
Submitted:
18 March 2024
Posted:
19 March 2024
You are already at the latest version
Abstract
To investigate the effect of prohexadione-calcium (Pro-Ca) primed on seed germination and seedling growth of oilseed rape under salt stress, to alleviate the inhibition of soil salinization on the growth of oilseed rape, and to improve the sustainability of agricultural production. Using Huayouza 158R and Huayouza 62 as test materials, the regulation of seed germination and seedling growth under the stresses of 0, 100, and 150 mM NaCl (S0, S100, and S150) was investigated after the primed of Pro-Ca. We conducted six treatments, S0: non-primed + 0 mM NaCl, PCS0: Pro-Ca primed + 0 mM NaCl, S100: non-primed + 100 mM NaCl, PCS100: Pro-Ca primed + 100 mM NaCl, S150: non-primed + 150mM NaCl, Pro-Ca+S150: Pro-Ca primed + 150 mM NaCl. The results showed that salt stress inhibited seed germination and seedling growth, and Pro-Ca primed significantly increased the germination index and vigor index of oilseed rape seeds. The average germination time and malondialdehyde (MDA) content were significantly reduced, which resulted in the optimal initiation fiddling concentration of 8mg·L-1. The morphological parameters of oilseed rape decreased significantly under salt stress, with leaf area decreasing by 70.69% and 75.03% and total chlorophyll content decreasing by 18.55% and 18.68% at S150. With the rise of salt stress, the changes of each photosynthetic parameter were somewhat similar, and Pn, Gs, Ci, and Tr continued to decline, indicating that the effects of salt stress on photosynthesis in oilseed rape might be mainly attributed to stomatal restriction and inhibition of photosynthetic pigment synthesis. While Na+ and MDA contents increased significantly, reaching a peak at S150, indicating that Na+ content increased significantly under salt stress, which resulted in the slow growth of oilseed rape and weak plants under the influence of Na+ salt damage. After Pro-Ca primed, leaf area and photosynthetic pigment synthesis increased, and the initiation treatment promoted the rise of Pn, Gs, Ci, and Tr under salt stress. The Na+ content decreased by 7.29% and 6.96% in PCS100 compared with S100, decreased by 9.23% and 25.02% in PCS150 compared with S150, and increased in K+ and Mg2+ after Pro-Ca primed. At this time, the antioxidant enzyme activities increased, and the MDA content and relative conductivity decreased. It was concluded that Pro-Ca primed improved the photosynthetic capacity of oilseed rape by promoting leaf growth, stomatal opening, and photosynthetic pigment synthesis. At the same time, reducing Na+ content and promoting K+ and Mg2+ absorption and antioxidant enzyme activities together alleviate salt damage and improve salt tolerance of oilseed rape.
Keywords:
Oilseed rape
; salt stress
; prohexadione-calcium
; photosynthetic capacity
; ion metabolism
1. Introduction
As the global population continues to increase, so does the human demand for food. Therefore, finding ways to improve crop yield and quality is a top priority for food security [1]. The growth of crops, however, has always been under the stress of an unfavorable environment. However, the growth of crops has been facing adverse environmental stresses such as drought, salt stress, insect pests, and so on. These biotic or abiotic stresses are serious constraints to crop yield and quality [2,3]. With the development of society, environmental pollution caused by improper agricultural management, sea level rise due to global warming, and a series of problems have gradually increased soil salinization, and salt stress has become a major adverse factor in agricultural production [4]. The total area of saline soil in the world is more than 8.3 × 108 hm2, which is about 25% of the total land area, spreading over all regions of the world. According to statistics, more than half of the irrigated land in the world is jeopardized by salinity and waterlogging, which seriously restricts the growth and development of crops [5]. Salt stress inhibits seed germination and plant growth and development, resulting in reduced yields. Salt stress injury to plants is mainly caused by Na+, high concentrations of Na+ accumulation, but also inhibits the absorption of K+, the accumulation of excessive Na+ will make the biofilm function impaired, affecting the osmotic balance, and causing physiological and metabolic disorders [6]. A large number of studies have shown that chlorophyll synthesis in leaves is inhibited under salt stress, and the photosynthetic rate of plants decreases, resulting in slow growth [7]. The plant’s photosynthetic rate decreases, resulting in slow growth.
Rapeseed is a vital oilseed, vegetable, and forage crop, with the planting area accounting for about 30% of the world and China’s production accounting for about 20% of the world’s production [8]. It is also an essential protein and edible oil source and can be used for ornamental purposes, oil extraction, wintergreen fodder, and fresh grass feeding [9]. As an essential green manure crop, winter oilseed rape proliferates, has a significant nutrient body, good mulching effect, low planting cost, and is essential for farmland soil protection and soil fertilization [10]. It has become vital to meet the increasing food demand by developing salt-tolerant varieties suitable for growing in saline soils.
Prohexadione-calcium (Pro-Ca) is a cyclohexane carboxylic acid plant growth retardant, which can regulate flowering time, improve plant fruiting rate, control plant growth, improve fruit yield and quality, and improve resistance to stress, etc., and has no residual toxicity and no pollution [11]. Because of its low toxicity and no residue, it is increasingly attracting widespread attention. In recent years, the positive effects of Pro-Ca on plant growth and development under adversity have been gradually discovered. Foliar spraying of Pro-Ca can alleviate the reduction of superoxide dismutase (SOD), peroxidase activity (POD), and catalase (CAT) activities in tobacco leaves under low-temperature stress, promote proline biosynthesis, and regulate the relative balance of reactive oxygen species (ROS) [12]. Improved root vigor, induced chlorophyll biosynthesis, increased leaf soluble protein content, and increased fruit weight and yield in peanuts [13]. Improve photosynthetic characteristics and promote soluble substance synthesis in soybeans under salt stress [14]. More and more researchers hope to improve crop salinity tolerance by developing and applying Pro-Ca.
In contrast, seed initiation technology can improve seed germination, seedling emergence, and seedling quality, which is conducive to the cultivation of solid seedlings and even improve plant resistance [15]. Therefore, in this experiment, Pro-Ca was used as an initiator to investigate the regulatory effects on seed germination and seedling growth of oilseed rape under salt stress. This study aimed to reveal the regulatory mechanism of Pro-Ca primed to alleviate salt stress in oilseed rape seedlings, to investigate the effect on salt tolerance of oilseed rape and its physiological metabolism mechanism changes after Pro-Ca primed, to solve the inhibition of soil salinity on oilseed rape growth, and to improve the sustainability of agricultural production. The results showed that prohexadione-calcium priming improved the photosynthetic capacity of oilseed rape by promoting leaf growth, stomatal opening, and photosynthetic pigment synthesis. Meanwhile, it reduced Na+ content and promoted K+ and Mg2+ absorption and antioxidant enzyme activities, which jointly alleviated salt damage and improved salt tolerance of oilseed rape.
2. Materials and Methods
2.1. Materials for Testing
The test oilseed rape varieties were Huayouza 158R (moderately salt-tolerant) and Huayouza 62 (salt-tolerant), and the test regulator was prohexadione-calcium (Pro-Ca), both of which were supplied by the Salt and Alkali Tolerance and Chemistry Control Group of Coastal Agricultural College, Guangdong Ocean University.
2.2. Experimental Design
2.2.1. Screening Test for Optimal Concentration of Prohexadione-Calcium
Seeds of full and uniform size were selected and sterilized by soaking in 3 % hydrogen peroxide (H2O2) for 10 min and rinsed three to five times with distilled water. The sterilized seeds were evenly placed in Petri dishes (9 cm in diameter) lined with three layers of filter paper, and the filter paper was moistened with 10 mL of Pro-Ca at concentrations of 0 mg-L-1; 2 mg-L-1; 4 mg-L-1; 8 mg-L-1; 16 mg-L-1; and 32 mg-L-1, respectively, and sealed with sealing film. The petri dishes were placed in the incubator and initiated in the dark at 20 °C for 8 h. After initiation, the seeds were cleaned from the surface solution with distilled water, the seeds were drained with absorbent paper, and then placed in a cool and ventilated place to naturally air-dry for 48 h ( 20-25°C, 40%-45% relative humidity), and then the seeds were determined to be used for germination test by weighing to determine that the seeds could be used for germination test after reaching the initial water content to screen out the optimal The optimum concentration was selected. The germination test was carried out in Petri dishes with 10 ml of salt solution (0, 100, and 150 mM, corresponding to S0, S100, and S150, respectively) at different concentrations, moistening the germination paper, and placing 25 seeds uniformly in Petri dishes and incubating them for 7 days to observe seed germination; the cotyledons were sampled on the 7th day to determine the malondialdehyde content.
(1) EA0: 0 mg-L-1 Pro-Ca primed.
(2) EA2: 2 mg-L-1 Pro-Ca primed.
(3) EA4: 4 mg-L-1 Pro-Ca primed.
(4) EA8: 8 mg-L-1 Pro-Ca primed.
(5) EA16: 16 mg-L-1 Pro-Ca primed.
(6) EA32: 32 mg-L-1 Pro-Ca primed.
2.2.2. Effect of Pro-Ca Primed on the Growth of Oilseed Rape Seedlings under Salt Stress
Concentration screening tests were conducted to derive the optimum initiating concentration, and soil cultivation was carried out in 19 cm × 14 cm × 17 cm pots. The soil was 2.5 kg per pot, and 1 L of salt solution of different concentrations (0, 100, and 150 mM) was mixed with the soil to simulate different salt stress environments. The study was conducted from 2022-2023 in a daylight-linked greenhouse (natural light conditions, day/night temperature difference of 25/20 ± 2 °C, relative humidity of 60%) at the Binhai College of Agriculture, Guangdong Ocean University (latitude: 21°8′56″ N, longitude: 110°17′58″ E, altitude: 20 m). Six holes were made in the soil of each pot and three seeds were sown in each hole, totaling 18 seeds. When the plant developed three true leaves, the plants were thinned leaving one plant in each hole for a total of six plants. Six treatments were set up for each variety. Samples were taken on the 36th day after sowing, and the relevant indexes were measured.
(1) S0: non-primed + 0 mM NaCl. Non-primed followed by growth in a 0 mM NaCl salt stress environment.
(2) PCS0: Pro-Ca primed + 0 mM NaCl. Pro-Ca primed followed by growth in a 0 mM NaCl salt stress environment.
(3) S100: non-primed + 100 mM NaCl. Non-primed followed by growth in a 100 mM NaCl salt stress environment.
(4) PCS100: Pro-Ca primed + 100 mM NaCl. Pro-Ca primed followed by growth in 100 mM NaCl salt stress environment.
(5) S150: non-primed + 150 mM NaCl. Non-primed followed by growth in a 150 mM NaCl salt stress environment.
(6) PCS150: Pro-Ca primed + 150 mM NaCl. Pro-Ca primed followed by growth in 150 mM NaCl salt stress environment.
2.3. Measurement Items and Methods
2.3.1. Prohexadione-Calcium Concentration Screening Test
After sowing, the number of germinated seeds was recorded daily. After 7 d of incubation, the cotyledons were frozen in liquid nitrogen and stored in a refrigerator at -80°C. The MDA content was determined by the thiobarbituric acid method [16]. The MDA content was determined by the thiobarbituric acid method. After that, the remaining oilseed rape was rinsed with water, and the fresh weight of the young shoots was determined after drying the surface water with absorbent paper. The formula for calculating the relevant indexes is as follows:
Germination index (GI) = ∑Gt/Dt
Vitality Index (VI) = GI × S
Mean germination time (MGT)=∑(Gt×Dt)/∑Gt
Dt represents the number of days to germination; Gt represents the number of germinated seeds per day corresponding to Dt; S is the fresh weight of normal seedlings [17].
2.3.2. Effect of Pro-Ca Primed on the Growth of Oilseed Rape Seedlings under Salt Stress
Determination of Morphological Indicators
Samples were taken after 36 d of growth under salt stress to determine morphology. Root height and root length were measured with a scale; stem thickness was measured with an electronic vernier caliper (the base of the stem was 1 cm away from the root system); leaf area was recorded by a leaf area meter; the root system and the above-ground part of the seedlings were separated, and the surface water was sucked up by blotting paper and put in a kraft paper bag, and then placed in an oven at 80 °C for 48 h, which was used to determine the dry weight of the root system and the above-ground dry weight and to calculate the seedling strength index.
Seedling strength index = (root dry mass/above-ground dry mass + stem thickness/plant height) / (root dry mass + above-ground dry mass) [18].
Measurement of Photosynthetic Pigments
The inverted bilobed leaves of oilseed rape were perforated with a perforator, and 0.1 g of leaves were weighed into test tubes. Photosynthetic pigments were extracted by adding 10 ml of anhydrous ethanol to the test tubes, and the absorption values at 665 nm, 649 nm, and 470 nm were determined spectrophotometrically using Eq [19]. Chlorophyll a and Chlorophyll b, carotenoids, and total chlorophyll contents were calculated.
Chlorophyll a = 13.95 A665 - 6.88 A649
Chlorophyll b = 24.96 A649 - 7.32 A665
Total Chlorophyll = Chlorophyll a + Chlorophyll b
Crotenoids = (1000 A470 - 2.05 Chlorophyll a-114.8 Chlorophyll b) / 245
Measurement of Photosynthetic Properties
The net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide (Ci), and transpiration rate (Tr) of oilseed rape seedlings were recorded after 36 d of salt stress at 14:00~16:00 pm on a sunny day with a portable photosynthesis measurement system, LI-6800 (LI-COR, Inc., USA). The CO2 concentration in the leaf chamber was 400 μmol mol−1, the light intensity was set at 1000 μmol m−2 s−1, the leaf temperature was 25 ± 1 °C, and the relative air humidity ranged from 65% to 70%.
Determination of Membrane Lipid Peroxidation Products
MDA content was determined using the thiobarbituric acid (TBA) method [16]. Seedling leaf tissue 0.5 g was weighed and ground in liquid nitrogen to powdered form, added to 10 ml of phosphate buffer (pH 7.8), and extracted with 0.6% TBA prepared with 10 % trichloroacetic acid (TCA). It was placed under boiling water at 100 °C for 15 min and then rapidly cooled on ice. After centrifugation at 7000 g for 20 min, the absorbance of the supernatant was measured at 450 nm, 532 nm, and 600 nm, respectively (UV-Vis spectrophotometer, GENESYS 180, below).
Weigh 0.1 g of leaves in a sealed test tube and add 10 ml of deionized water. R1 was measured by immersion in a dark environment for 12 hours; after that, R2 was measured by cooling in boiling water in a water bath for 30 minutes.
relative conductivity (EL): EL= R1 / R2 ×100% [20].
Determination of Antioxidant Enzymes
Determination of antioxidant enzymes: 0.5 g of frozen leaf samples were accurately weighed and ground in 10 mL of 0.05 mol·L−1 pre-cooled phosphate buffer (pH 7.8) to form a homogenate, centrifuged at 12,000 g for 20 min at four °C, and the supernatant was used for the determination of the activities of SOD, POD, CAT, and ascorbic acid peroxidase (APX). SOD activity was determined by measuring the change in absorbance at 560 nm according to the method of Giannopolitis, and one unit of SOD activity was defined as the amount of enzyme that inhibited 50% of NBT photoreduction [21]. POD activity was determined by the oxidation rate of guaiacol at 470 nm by Zhang and Liping [22]. CAT activity was determined by the decrease in absorbance per minute at 240 nm caused by the depletion of H2O2 by the Maehly method [23]. APX activity was determined by the amount of change in absorbance per minute at 290 nm by Ahanger [24].
Determination of Ionic Content
After 36 d of salt stress, dry samples were taken to determine the content of each ion. A 150 mg dry mass of leaves was digested with 6 ml HNO3 /HClO4 (4:1, v/v), and the supernatant was analyzed after the solution was clarified. Inductively coupled plasma emission spectroscopy (Prodigy XP, LEEMAN, Inc., USA) was used to determine the Na+, Mg2+, and Ca2+ contents, and a flame photometer (Sherwood M410, Inc., UK) was used to determine the K+ content in the dry samples [25].
Statistical Analysis
All data were analyzed by one-way ANOVA using SPSS 25.0. Duncan’s test was used to evaluate the difference among different treatments; P < 0.05 indicated that there was a statistical difference between treatments. Origin 2021 software was used for plotting.
3. Results
3.1. Effect of Pro-Ca Primed on Seed Germination under Salt Stress
As shown in Figure 1, there were some differences after priming different concentrations of Pro-Ca, but the inhibition of seed germination by salt stress was effectively alleviated after priming. As the degree of salt stress increased, the germination and vigor indexes decreased, and the average germination time increased. The germination index of Huayouza 158R increased significantly by 29.17% to 56.71% (Figure 1A), the vigor index increased significantly by 21.42% to 81.23% (Figure 1C), and the average germination time was shortened by 3.41% to 7.77% (Figure 1E) after primed by Pro-Ca. The germination index of Huayouza 62 increased significantly by 21.82% ~ 61.31% (Figure 1B), the vigor index increased significantly by 20.09% ~ 107.62% (Figure 1D), and the average germination time was shortened by 3.51% ~ 7.51% (Figure 1F). A comparison between the two varieties under salt stress revealed that although the average germination time was not significantly different, the germination index and seed vigor of Huayouza 62 were higher under salt stress. The inhibitory effect of salt stress on seed germination could be alleviated by the priming of Pro-Ca, and the germination index and seed vigor first increased and then decreased after treatment with different concentrations and then reached the peak value in the EA8 treatment:
3.2. Effect of Pro-Ca Primed on Fresh Weight and Malondialdehyde of Young Shoots
Under unstressed (S0) conditions, fresh weight increased (Figure 2A,B), and malondialdehyde content decreased (Figure 2C,D) after Pro-Ca primed, suggesting that Pro-Ca had a growth-promoting effect on oilseed rape. Salt stress (S100 and S150) inhibited the growth of young shoots, which resulted in a significant decrease in fresh weight and an increase in malondialdehyde content, and the inhibitory effect of salt stress was significantly alleviated by Pro-Ca primed. Compared with EA0, only EA8 treatment significantly increased the fresh weight of Huayouza 158R under salt stress, and EA4, EA8, and EA16 treatments significantly increased the fresh weight of Huayouza 62. The malondialdehyde content of Huayouza 158R was significantly decreased in all treatments, and the malondialdehyde content of Huayouza 62 was significantly decreased in EA8 and EA32. Huayouza 158R was more sensitive to Pro-Ca primed, and the optimal initiation concentration was EA8.
3.3. Effect of Pro-Ca Primed on the Morphology of Oilseed Rape under Salt Stress
The salt stress treatments (S100 and S150) sustained significant decreases in plant height, root length, leaf area, and stoutness index compared to the unstressed treatment (S0). The salt stress treatment S150 decreased 49.45%, 24.83%, 70.69%, and 80.05% in Huayouza 158R and 50.41%, 26.40%, 75.03%, and 59.27% in Huayouza 62, respectively (Figure 3A–D). After Pro-Ca-tuned initiation of Huayouza 158R, PCS0 treatment significantly promoted the growth of oilseed rape compared with S0 (Figure 3A–D), PCS100 treatment significantly increased plant height, leaf area, and stoutness index compared with S100 (Figure 3A,C,D), and PCS150 treatment significantly increased only root length compared with S150 (Figure 3B). After Pro-Ca primed, plant height, leaf area and seedling index increased significantly in the PCS0 treatment compared with S0 (Figure 3A,C,D), only leaf area increased significantly in the PCS100 treatment compared with S100 (Figure 3C), and only root length increased significantly in the PCS150 treatment compared with S150 (Figure 3B). The experiments showed that Pro-Ca priming could alleviate the inhibitory effect of salt stress on the growth of oilseed rape and induce robust development of oilseed rape.
3.4. Effect of Pro-Ca Primed on Photosynthetic Pigments in Oilseed Rape under Salt Stress
As shown in Figure 4, the salt stress treatment (S150) significantly decreased the content of each photosynthetic pigment by 17.07%, 19.22%, 17.66%, and 18.55% compared with S0. The chlorophyll a and total chlorophyll contents of Huayouza 158R continued to decrease significantly in the S150 treatment compared with the S100 treatment, while Huayouza 62 tended to flatten out with insignificant changes. PCS0 showed a significant increase in photosynthetic pigment content compared with S0 (Figure 4A–D). Comparing PCS100 with S100, the indexes of Huayouza 158R increased significantly by 17.73%, 30.07%, 20.05%, and 20.69%, respectively (Figure 4A–D); Huayouza 62 only showed a significant increase in chlorophyll a, which increased by 9.68% (Figure 4A). Comparing PCS150 with S150, chlorophyll a and total chlorophyll content of Huayouza 158R increased significantly by 11.08% and 11.32% (Figure 4A,D), and chlorophyll a and carotenoid content of Huayouza 62 increased significantly (Figure 4A,C) by 10.00% and 18.53%. From comparing PCS100 with S100, it can be concluded that Pro-Ca primed significantly promoted the synthesis of photosynthetic pigments in Huayouza 158R (Figure 4A–D). Comparison of PCS150 with S150 revealed that after Pro-Ca primed, Huayouza 158R mainly synthesized chlorophyll a (Figure 4A), and Huayouza 62 mainly synthesized carotenoids (Figure 4C). The two varieties were identical in that the changes in chlorophyll content tended to be the same (Figure 4A).
3.5. Effect of Pro-Ca Primed on Photosynthetic Properties of Oilseed Rape under Salt Stress
In the S100 treatment compared with S0, Tr of Huayouza 62 decreased significantly, and the changes of the remaining indicators were not significant; in the S150 treatment compared with S0, the indicators decreased significantly (Figure 5A–D). Comparing PCS0 with S0, the Tr of Huayouza 62 significantly increased by 28.23% (Figure 5D), and the remaining indicators’ changes were insignificant. PCS100 compared with S100, Pn and Ci significantly increased by 18.11% and 9.32% in Huayouza 158R (Figure 5A,C), and Pn and Tr significantly increased by 19.99% and 38.85% in Huayouza 62 (Figure 5A,D). Comparing PCS150 with S150, the indexes of Huayouza 158R increased significantly, Pn, Gs, and Ci of Huayouza 62 increased significantly, and the change of Tr was not significant (Figure 5A–D). Pro-Ca primed promoted a significant increase in Pn in oilseed rape under salt stress (Figure 5A).
3.6. Effect of Pro-Ca Primed on Membrane Lipid Peroxidation in Oilseed Rape under Salt Stress
The relative conductivity and MDA content of Huayouza 158R leave significantly and consistently increased under salt stress (S100 and S150) compared with unstressed (S0). The relative conductivity of S100-treated leaves of Huayouza 62 increased significantly, and the MDA content changed insignificantly, while the relative conductivity and MDA content of S150-treated leaves increased significantly (Figure 6A,B). PCS0 showed a downward trend in relative conductivity and a significant decrease of 12.44% in MDA content in Huayouza 158R leaves when compared with S0 (Figure 6A). PCS100 showed a significant decrease of 23.87% in the relative conductivity of Huayouza 158R leaves when compared with S100. Comparing PCS150 with S150, the relative conductivity and MDA content of Huayouza 158R leaves decreased significantly by 31.57% and 7.32%, and the MDA content of Huayouza 62 decreased significantly by 17.39%. The results indicated that the relative conductivity and MDA content of leaves of oilseed rape seedlings changed significantly under salt stress; these treatments primed by Pro-Ca could reduce the relative conductivity and MDA content of leaves.
3.7. Effect of Pro-Ca Primed on Antioxidant Enzymes in Oilseed Rape under Salt Stress
As shown in Figure 7, POD and APX activities increased significantly under salt stress (S100 and S150) compared with unstressed (S0) (Figure 7B,D), and CAT activities changed insignificantly (Figure 7C), with a sustained and significant increase in SOD activity in Huayouza 158R, and a significant increase in Huayouza 62 in the S150 treatment (Figure 7A). Comparing PCS0 with S0, SOD and POD activities of Huayouza 158R increased significantly by 18.28% and 30.44%, and CAT and APX activities changed insignificantly. SOD, POD, and APX activities of Huayouza 62 increased significantly by 11.21%, 33.53%, and 10.90%, and CAT changed insignificantly. SOD and POD of Huayouza 158R increased significantly by 11.37% and 19.53% when PCS100 was compared with S100. The activities of’s antioxidant enzymes of Huayouza 62 increased significantly by 20.22%, 13.72%, 16.23%, and 6.96%, respectively. Comparing PCS150 with S150, all the indexes increased significantly. Huayouza 158R increased by 18.61%, 22.59%, 18.58%, and 9.20%, and Huayouza 62 increased by 8.92%, 6.76%, 14.12% and 16.70%.
3.8. Effect of Pro-Ca Primed on Ion Content of Oilseed Rape under Salt Stress
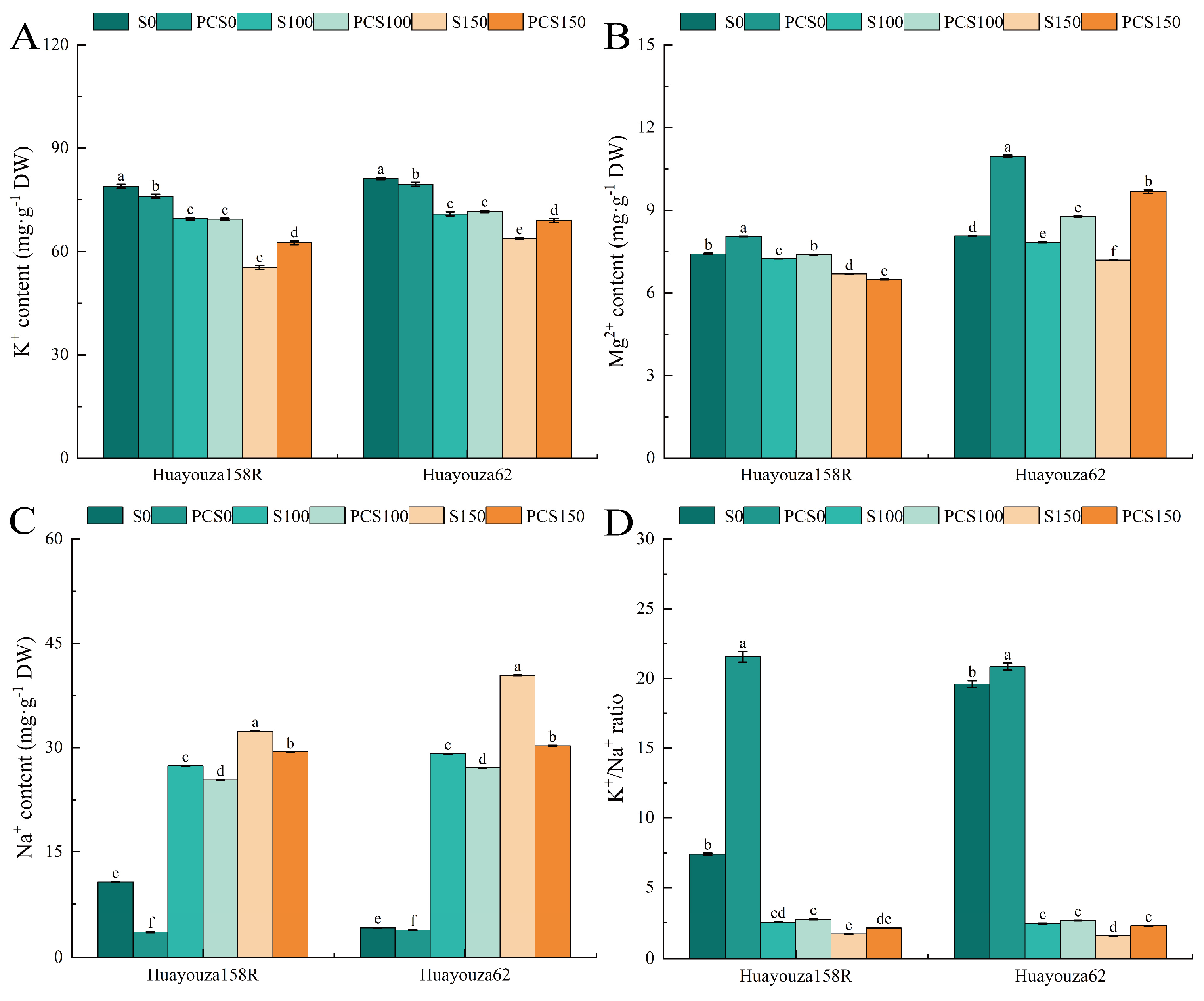
As shown in Figure 8, salt stress (S100 and S150) showed significant decreases in K+, Mg2+, and K+/Na+ and a significant increase in Na+ content compared to unstressed (S0) (Figure 8A–D). PCS0 showed a significant decrease in K+ and Na+ content and a significant increase in Mg2+ and K+/Na+ compared to S0. PCS100 showed insignificant changes in K+ and K+/Na+ compared with S100; Mg2+ increased significantly by 2.16% and 11.91% in the two varieties, respectively (Figure 8B,D). Na+ content decreased significantly, by 7.29% and 6.96%, respectively (Figure 8C). Comparing PCS150 with S150, the K+ content of Huayouza 158R increased significantly by 12.92%, and the Mg2+ and Na+ contents decreased significantly by 3.09% and 9.23%. The K+, Mg2+, and K+/Na+ of Huayouza 62 increased significantly by 8.36%, 34.62%, and 44.51%, respectively, and the Na+ content decreased significantly by 25.02% (Figure 8A–D).
4. Discussion
Seed germination is the beginning of the life course of oilseed rape, and the germination and seedling stages are more sensitive to salt stress during the growth and development of oilseed rape. Studies have concluded that germination index, vigor index, and seedling fresh weight can be used as effective indicators for screening salt tolerance in oilseed rape at the germination stage [26]. Some scholars have found that sorghum seed shoot length, root length, germination potential, and germination rate showed a sequential decrease with increasing salt concentration and different degrees of delayed germination [27]. The results of the present study were similar to it; with increasing salt concentration germination index and vigor index continued to decline, and the average germination time increased, inhibiting the vigor of oilseed rape seeds and weakening the germination ability (Figure 1). With the increase in salt concentration, the germination rate, germination index, and vigor index decreased more, and the seed vigor index was more sensitive than the germination rate and germination index under salt stress [28]. The experiment concluded that the vigor index decreased the most with increasing salt stress (Figure 1), similar to its results, and the more sensitive change of seed vigor index under salt stress was verified. After being primed with different concentrations of Pro-Ca, the seed germination index and vigor index increased significantly, and the average germination time decreased significantly, indicating that primed could alleviate the inhibitory effect of salt stress on seed germination and improve germination neatness [29]. Comparison among the concentrations revealed that the seed germination and vigor indexes tended to increase and decrease. The average germination time decreased and then increased (Figure 1), so EA8 (8 mg·L-1) initiation had the best effect in promoting the germination of oilseed rape. Seeds affected by salt stress would inhibit seed germination growth with weak young shoots and yellowing cotyledons. Injury by salt stress deepens cell membrane lipid peroxidation, resulting in elevated MDA content, so the degree of injury by salt stress was assessed by measuring the MDA content of young shoots [30]. As a result of salt stress, the fresh weight of young oilseed rape shoots decreased, and the MDA content tended to increase. Pro-Ca primed an increase in fresh weight and a significant decrease in MDA content under salt stress (Figure 2), indicating that Pro-Ca promotes the growth of young oilseed rape shoots under salt stress and reduces cellular lipid peroxidation. It was concluded that the optimal initiation concentration of Pro-Ca primed was EA8 (8 mg·L-1).
Salt stress leads to slow growth and development and dwarf plants [31]. Our study showed that salt stress inhibited plant height, root length, and leaf area in oilseed rape, with a significant decrease in the stoutness index (Figure 3), typical of developmental retardation. It was found that under salt stress (S150), plant height and leaf area decreased by 49.45% and 70.69% in Huayouza 158R, 50.41% and 75.03% in Huayouza 62, and 24.83% and 26.40% in root length, respectively. The inhibition of plant height and leaf area was higher than that of root length, which was inconsistent with the trend of changes in previous studies [32], probably due to the limited space in pots, which restricts the growth of the root system. Pro-Ca is primed to promote the growth and development of oilseed rape and leaf growth (Figure 3C), which provides the basis for photosynthesis in oilseed rape. The strong seedling index can reflect whether the plant thrives [17], and after Pro-Ca was primed, the strong seedling index was significantly increased under unstressed conditions, which promoted plant robustness (Figure 3D). It showed an increasing trend under salt stress, and the PCS100 treatment of Huayouza 158R was significantly higher than S100, indicating that Pro-Ca primed could significantly improve the growth inhibition of Huayouza 158R by salt stress.
Most reports indicated a decrease in photosynthetic capacity and significant inhibition of photosynthetic pigment synthesis under salt stress. Prolonged salt stress reduces the biosynthesis of the chlorophyll protein-lipid complex, which affects the photosynthetic system and results in stunted plant growth [33,34,35]. However, no uniform conclusion has emerged on the changes in photosynthetic pigments under salt stress, with some studies showing that chlorophyll content increases in salt-tolerant plants under salt stress [36]. In contrast, others suggest that the elevation of chlorophyll content is temporary, increasing and then decreasing with the continued effects of salt stress [37]. In this study, it was shown that the sprouting growth of oilseed rape under salt stress inhibits the synthesis of photosynthetic pigments, and the salt-tolerant variety Huayouza 62 has a higher level of photosynthetic pigment content (Figure 4). It was found that although the level of photosynthetic pigment content of Huayouza 158R was lower, the photosynthetic pigment content was significantly increased after initiation, and the effect was better than that of Huayouza 62. Maintaining a certain level of photosynthetic pigment can improve light energy conversion efficiency and maintain photosynthetic activity. It has been pointed out that photosynthetic parameters were significantly reduced, and growth and development were inhibited under salt stress [38]. In this study, Pn, Gs, Ci, and Tr were significantly decreased at S150 (Figure 5), which indicated that the decrease in Pn under salt stress was due to stomatal restriction, reduced stomatal opening to reduce transpiration, and decreased inter-cellular CO2 content, resulting in a significant decrease in Pn. Leaf stunting and photosynthetic pigment content decreased under salt stress, while photosynthetic parameters such as Pn only decreased significantly at S150, suggesting that oilseed rape had some resistance at S100. The study also showed that Pro-Ca priming promoted Pn by promoting stomatal opening and intercellular CO2 content, and the promotion became more significant with the elevation of salt stress. This suggests that Pro-Ca priming seems to play a more significant role in oilseed rape when it is more severely inhibited, and the exact mechanism of its effect needs to be further investigated.
The ionic damage of salt stress on oilseed rape is mainly caused by Na+; excessive Na+ will disturb the plant metabolism, cause damage to the cell membrane system, increase the permeability of the cell membrane, and electrolyte extravasation, resulting in cellular metabolism disorders and the accumulation of reactive oxygen species, which will lead to the inhibition of plant growth or even death [39]. When the concentration of Na+ and Cl- in the plant is too high, it also interferes with the uptake of nitrogen and nutrients such as K+ and Mg2+ by the root system, which leads to plant stunting [40]. The present study showed a significant increase in Na+ content and a significant decrease in K+ and Mg2+ content under salt stress (Figure 8). Excessive accumulation of Na+, increased cellular plasma peroxidation, and damage to membrane lipids or membrane proteins caused a significant increase in relative conductivity and MDA content of oilseed rape leaves (Figure 6), which is consistent with the results of previous studies. The central metal element for chlorophyll synthesis is Mg [41]. The significant decrease in Mg2+ content may further affect chlorophyll synthesis, and the reduced photosynthetic capacity leads to slow development. It was found that antioxidant enzyme activities were elevated at this time, and oilseed rape activated the self-defense mechanism (Figure 7). The antioxidant defense system is a crucial mechanism for plants to regulate the balance of reactive oxygen species and resist environmental stress [42]. The changes in antioxidant enzyme activities under salt stress were variable, and it was noted that the activities of SOD, CAT, and APX were significantly enhanced in tomato seedlings under salt stress [43]. Our results were similar to theirs; SOD, POD, and APX activities increased significantly under salt stress except for CAT activity (Figure 7), which may be related to the degree of salt stress and cultivar salt tolerance, and the antioxidant defense system of oilseed rape was enhanced under salt stress. After Pro-Ca primed, Na+ content decreased significantly, and K+ and Mg2+ showed an increasing trend (Figure 8). A large amount of Na+ accumulation inhibits K+ uptake, and researchers have suggested that the K+/ Na+ ratio can reflect the degree of salt stress in crops [44]. The K+ content (Figure 8A), which decreased after Pro-Ca primed under unstressed conditions, is worth noting. Under salt stress conditions, it increased after Pro-Ca primed; however, both K+/ Na+ increased (Figure 8D). This suggests that the mode of regulation is inconsistent, with the unstressed condition primarily contributing to a decrease in Na+ content and the salt stress condition contributing to a decrease in Na+ while also promoting the uptake of potassium ions. The study also showed that Pro-Ca primed a decrease in relative conductivity and MDA content and increased antioxidant enzyme activities. Therefore, our results strongly suggest that Pro-Ca primed alleviates salt stress in oilseed rape.
5. Conclusions
In this experiment, salt stress inhibited seed vigor, and seed vigor increased significantly after Pro-Ca was primed, and malondialdehyde content decreased significantly, alleviating salt stress inhibition and promoting seed germination. Salt stress inhibited stomatal opening and photosynthetic pigment synthesis, and the Pn decreased; meanwhile, the Na+ content increased significantly, and the relative conductivity and MDA content of leaves increased significantly under the influence of salt damage, and the growth of oilseed rape was slow, and the plants were weak. Pro-Ca primed promoted leaf growth, stomatal opening, and photosynthetic pigment synthesis, improved leaf photosynthetic capacity, and promoted the growth of oilseed rape seedlings, with a rise in seedling strength index; it also inhibited Na+ uptake, promoted K+ and Mg2+ uptake, and increased the activity of antioxidant enzymes while the K+/Na+ increased, which effectively mitigated ionic toxicity. Therefore, Pro-Ca primed can improve the photosynthetic capacity and antioxidant enzyme activity of oilseed rape by promoting leaf growth, reducing Na+ content, and promoting K+ and Mg2+ uptake to jointly alleviate the salt damage and improve the salt tolerance of oilseed rape.
Author Contributions
Conceptualization, Peng Deng, Naijie Feng and Dianfeng Zheng; methodology, Naijie Feng and Peng Deng; software, Peng Deng; validation, Peng Deng; formal analysis, Peng Deng and Naijie Feng; investigation, Peng Deng; resources, Naijie Feng and Dianfeng Zheng; data curation, Peng Deng and Naijie Feng.; writing—original draft preparation, X.X.; writing—review and editing, X.X.; visualization, X.X.; supervision, X.X.; project administration, X.X.; funding acquisition, Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
Funding Source National 13th Five-Year Plan Key R&D Program Project (2019YFD1002205); Guangdong Provincial Department of Education General College Innovation Team Project (2021KCXTD011); Guangdong Provincial Department of Education Graduate Student Innovation Forum (2022XSLT036).
Data Availability Statement
We accept raw data uploads.
Acknowledgments
We thank Prof. Naijie Feng and Prof. Dianfeng Zheng for their support. Help in materials, funding, and revising the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jat Baloch, M.Y.; Zhang, W.; Sultana, T.; Akram, M.; Shoumik, B.A.A.; Khan, M.Z.; Farooq, M.A. Utilization of sewage sludge to manage saline–alkali soil and increase crop production: Is it safe or not? Environ. Technol. Innov. 2023, 32. [Google Scholar] [CrossRef]
- Radha, B.; Sunitha, N.C.; Sah, R.P.; TP, M.A.; Krishna, G.K.; Umesh, D.K.; Thomas, S.; Anilkumar, C.; Upadhyay, S.; Kumar, A.; et al. Physiological and molecular implications of multiple abiotic stresses on yield and quality of rice. Frontiers in Plant Science 2023, 13. [Google Scholar] [CrossRef]
- Kalaji; Hazem, M. Emerging technologies and management of crop stress tolerance || The use of chlorophyll fluorescence kinetics analysis to study the performance of photosynthetic machinery in plants; Elsevier Inc.: 2014; pp. 347–384.
- Kim, J.B.; So, J.M.; Bae, D.H. Global warming impacts on severe drought characteristics in Asia monsoon region. Water 2020, 12, 1360. [Google Scholar] [CrossRef]
- Zhang, K.; Chang, L.; Li, G.; Li, Y. Advances and future research in ecological stoichiometry under saline-alkali stress. Environmental Science and Pollution Research 2022, 30, 5475–5486. [Google Scholar] [CrossRef]
- Etesami, H.; Fatemi, H.; Rizwan, M. Interactions of nanoparticles and salinity stress at physiological, biochemical and molecular levels in plants: A review. Ecotoxicology and Environmental Safety 2021, 225. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.N.; Imran, S.; Hannan, A.; Paul, N.C.; Mahamud, M.A.; Chakrobortty, J.; Sarker, P.; Irin, I.J.; Brestic, M.; Rhaman, M.S. Organic amendments for mitigation of salinity stress in plants: A review. Life 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Xian, M.; Li, J. Current situation and development trends of rapeseed variety improvement in China. Journal of China Agricultural University 2024, 50–62. [Google Scholar] [CrossRef]
- Feng, H.; Wang, H. Security strategy for the nation’s edible vegetable oil supplies under the new circumstances. Chinese Journal of Oil Crop Sciences 2024, 1–7. [Google Scholar] [CrossRef]
- Jiao, J.; Sun, W.; Fang, Y.; Chen, Q.; Cui, X.; Mu, C.; Bai, J.; An, L.; Luo, X.; Wan, H.; et al. Effects of different winter rape mulchings on soil physical and chemical properties and microorganisms. Agricultural Research in the Arid Areas 2023, 41, 128–139. [Google Scholar]
- Nakayama, I.; Miyazawa, T.; Kobayashi, M.; Kamiya, Y.; Abe, H.; Sakurai, A. Effects of a new plant growth regulator Prohexadione Calcium (BX-112) on shoot elongation caused by exogenously applied gibberellins in rice (Oryza sativa L.) seedlings. Plant and Cell Physiology 1990, 31, 195–200. [Google Scholar] [CrossRef]
- Pan, M.; Yin, Y.; Shen, F.; Luo, B.; Tang, X.; He, H.; Chen, D. Effects of Prohexadione Calcium on physiological index of tobacco seedlings during cold stress. Southwest China Journal of Agricultural Sciences 2016, 29, 6. [Google Scholar] [CrossRef]
- Du, L.; Fan, T.; Wang, C.; Wan, G.; Jiang, T.; Zhang, J.; Wang, T.; Chen, K. Effect of Prohexadione Calcium on senescence, yield and quality of summer-planting peanut. Journal of Peanut Science 2008, 37, 32–36. [Google Scholar]
- Feng, N.; Yu, M.; Li, Y.; Jin, D.; Zheng, D. Prohexadione-calcium alleviates saline-alkali stress in soybean seedlings by improving the photosynthesis and up-regulating antioxidant defense. Ecotoxicology Environmental Safety 2021, 220, 112369. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. Journal of Plant Physiology 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Anastasiou, E.; Lorentz, K.O.; Stein, G.J.; Mitchell, P.D. Prehistoric schistosomiasis parasite found in the Middle East. The Lancet Infectious Diseases 2014, 14, 553–554. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous melatonin improves seedling health index and drought tolerance in tomato. Plant Growth Regulation 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzyme in isolated chloroplasts. polyphenoloxidase in beta vulgaris. Plant Physiol 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.; Du, X.; Tang, H.; Shen, C.; Wu, J. Salicylic acid alleviates the adverse effects of salt stress in Torreya grandis cv. merrillii seedlings by activating photosynthesis and enhancing antioxidant systems. PLoS ONE 2014, 9, 0109492. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants 1 2. Plant Physiol. 1977. [Google Scholar] [CrossRef]
- Zhang, L.; Pei, Y.; Wang, H.; Jin, Z.; Liu, Z.; Qiao, Z.; Fang, H.; Zhang, Y. Hydrogen sulfide alleviates cadmium-induced cell death through restraining ROS accumulation in roots of Brassica rapa L. ssp. pekinensis. Oxidative Medicine Cellular Longevity 2015, 2015, 1–11. [Google Scholar] [CrossRef]
- Wei, M.Y.; Liu, J.Y.; Li, H.; Hu, W.J.; Shen, Z.J.; Qiao, F.; Zhu, C.Q.; Chen, J.; Liu, X.; Zheng, H.L. Proteomic analysis reveals the protective role of exogenous hydrogen sulfide against salt stress in rice seedlings. Nitric Oxide 2021, 111–112, 14–30. [Google Scholar] [CrossRef]
- Abass, A.M.; Nasser, A.M.; Leonard, W.; Alamri, S.A.; Pravej, A.; Muhammad, A.; Parvaiz, A.; Yuan, H. Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. International Journal of Genomics 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Yadav, S.P.S.; Adhikari, R.; Paudel, P.; Shah, B.; Pokhrel, S.; Puri, S.; Adhikari, R.; Bhujel, S. Effect of different chemical priming agents on physiological and morphological characteristics of rice (Oryza sativa L.). Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Chen, X.; Zhang, R.; Xing, Y.; Jiang, B.; Li, B.; Xu, X.; Zhou, Y. The efficacy of different seed priming agents for promoting sorghum germination under salt stress. PLoS ONE 2021, 16. [Google Scholar] [CrossRef]
- Monjezi, N.; Yaghoubian, I.; Smith, D.L. Cell-free supernatant of Devosia sp. (strain SL43) mitigates the adverse effects of salt stress on soybean (Glycine max L.) seed vigor index. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef]
- Pereira, A. Plant abiotic stress challenges from the changing environment. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef]
- Ibrahimova, U.; Zivcak, M.; Gasparovic, K.; Rastogi, A.; Allakhverdiev, S.I.; Yang, X.; Brestic, M. Electron and proton transport in wheat exposed to salt stress: Is the increase of the thylakoid membrane proton conductivity responsible for decreasing the photosynthetic activity in sensitive genotypes? Photosynth. Res. 2021, 150, 195–211. [Google Scholar] [CrossRef]
- Sezer, İ.; Kiremit, M.S.; Öztürk, E.; Subrata, B.A.G.; Osman, H.M.; Akay, H.; Arslan, H. Role of melatonin in improving leaf mineral content and growth of sweet corn seedlings under different soil salinity levels. Scientia Horticulturae 2021, 288. [Google Scholar] [CrossRef]
- Khan, M.N.; Li, Y.; Fu, C.; Hu, J.; Chen, L.; Yan, J.; Khan, Z.; Wu, H.; Li, Z. CeO2 nanoparticles seed priming increases salicylic acid level and ROS scavenging ability to improve rapeseed salt tolerance. Global Challenges 2022, 6. [Google Scholar] [CrossRef]
- Guo, H.; Hu, Z.; Zhang, H.; Min, W.; Hou, Z. Comparative effects of salt and alkali stress on antioxidant system in cotton (Gossypium Hirsutum L.) leaves. Open Chemistry 2019, 17, 1352–1360. [Google Scholar] [CrossRef]
- Yin, J.; Jia, J.; Lian, Z.; Hu, Y.; Guo, J.; Huo, H.; Zhu, Y.; Gong, H. Silicon enhances the salt tolerance of cucumber through increasing polyamine accumulation and decreasing oxidative damage. Ecotoxicology environmental safety 2019, 169, 8. [Google Scholar] [CrossRef]
- Xue, F.; Liu, W.; Cao, H.; Song, L.; Ji, S.; Tong, L.; Ding, R. Stomatal conductance of tomato leaves is regulated by both abscisic acid and leaf water potential under combined water and salt stress. Physiologia Plantarum 2021, 172, 2070–2078. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Hou, H. Effect of salt stress on growth, physiological parameters, and ionic concentration of water dropwort (Oenanthe javanica) cultivars. Frontiers in Plant Science 2021, 12, 660409. [Google Scholar] [CrossRef]
- Gadelha, C.G.; Cotta Coutinho, I.A.; Kennedy, D.P.P.; Sergimar; Miguel, E.D.C.; De Carvalho, H.H.; Lopes, L.D.S.; Gomes-Filho, E. Sodium uptake and transport regulation, and photosynthetic efficiency maintenance as the basis of differential salt tolerance in rice cultivars. Environmental experimental botany 2021, 192. [Google Scholar] [CrossRef]
- Tian, L.; Tan, L.; Liu, F.; Cai, H.; Sun, C. Identification of quantitative trait loci associated with salt tolerance at seedling stage from Oryza rufipogon. Journal of Genetics and Genomics 2011, 38, 593–601. [Google Scholar] [CrossRef]
- Kong, X.-H. Toxcity of heavy metal ions to plants and their toxicological mechanism. Journal of Longyan Teachers Colleg 2005. [Google Scholar] [CrossRef]
- Wang, G.; Cao, F.; You, Q.; Qi, W. Effects of salt stress on concentrations of Na+,K+ and Na+/K+ in the leaves of four tree species and evaluation of salt tolerance. Journal of Plant Resources and Environment 2001, 10, 5. [Google Scholar] [CrossRef]
- Moynier, F.; Fujii, T. Theoretical isotopic fractionation of magnesium between chlorophylls. Scientific Reports 2017, 7. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.O.N.; Miller, G.A.D. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2011, 35, 259–270. [Google Scholar] [CrossRef]
- Hernández, J.A.; Ferrer, M.A.; Jiménez, A.; Barceló, A.R.; Sevilla, F. Antioxidant systems and O2-/H2O2 production in the apoplast of pea leaves. Its relation with salt-induced necrotic lesions in minor veins. American Society of Plant Biologists 2001, 127, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Chuamnakthong, S.; Nampei, M.; Ueda, A. Characterization of Na+ exclusion mechanism in rice under saline-alkaline stress conditions. Plant Science 2019, 287. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of Pro-Ca primed on the germination of oilseed rape under salt stress (day 7). Germination index (A and B), vigor index (C and D), and average germination time (E and F). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 1.
Effect of Pro-Ca primed on the germination of oilseed rape under salt stress (day 7). Germination index (A and B), vigor index (C and D), and average germination time (E and F). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
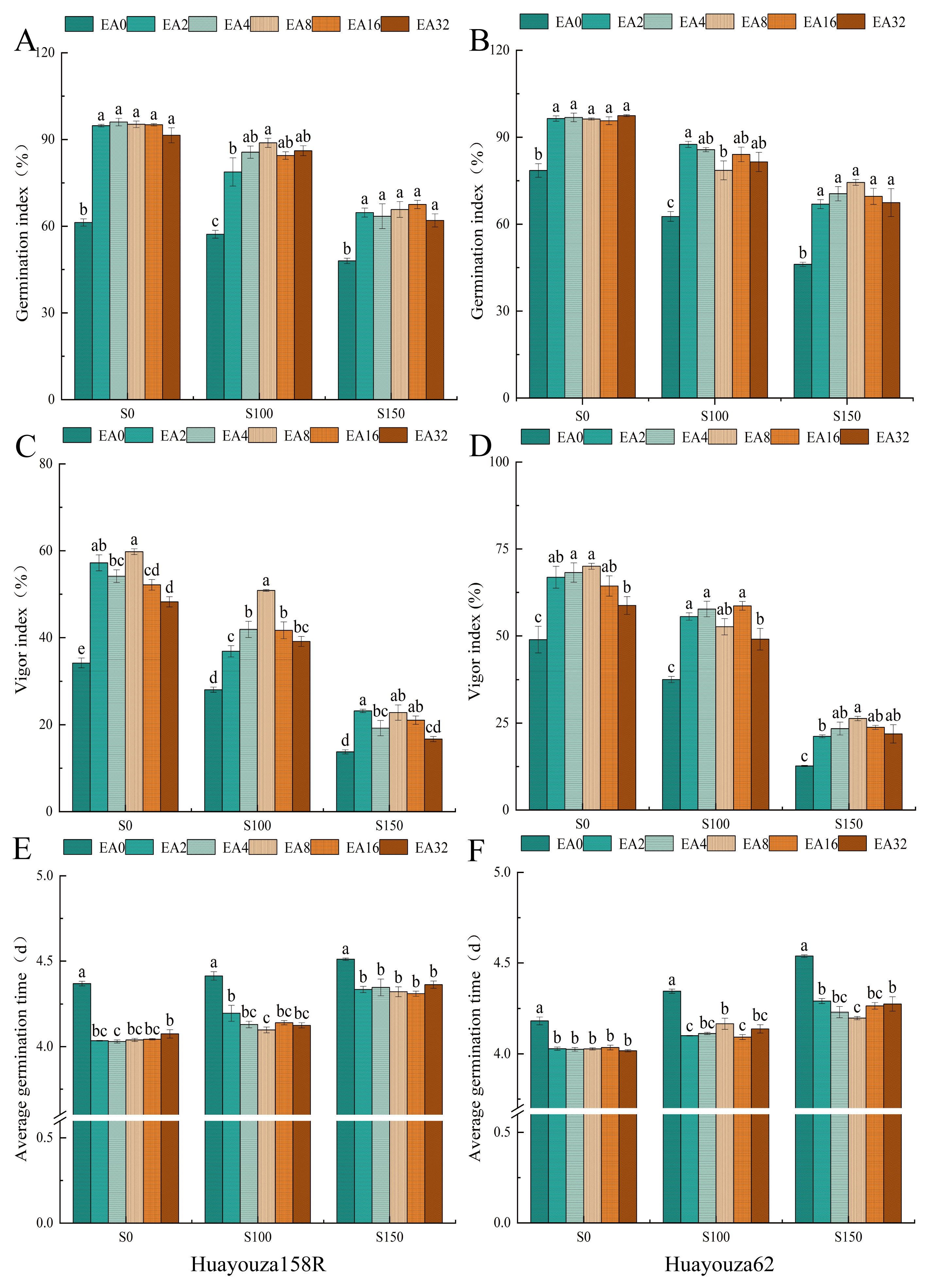
Figure 2.
Effect of Pro-Ca primed on fresh weight and malondialdehyde in oilseed rape under salt stress (day 7). Fresh weight (A and B), MDA: malondialdehyde (C and D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 2.
Effect of Pro-Ca primed on fresh weight and malondialdehyde in oilseed rape under salt stress (day 7). Fresh weight (A and B), MDA: malondialdehyde (C and D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
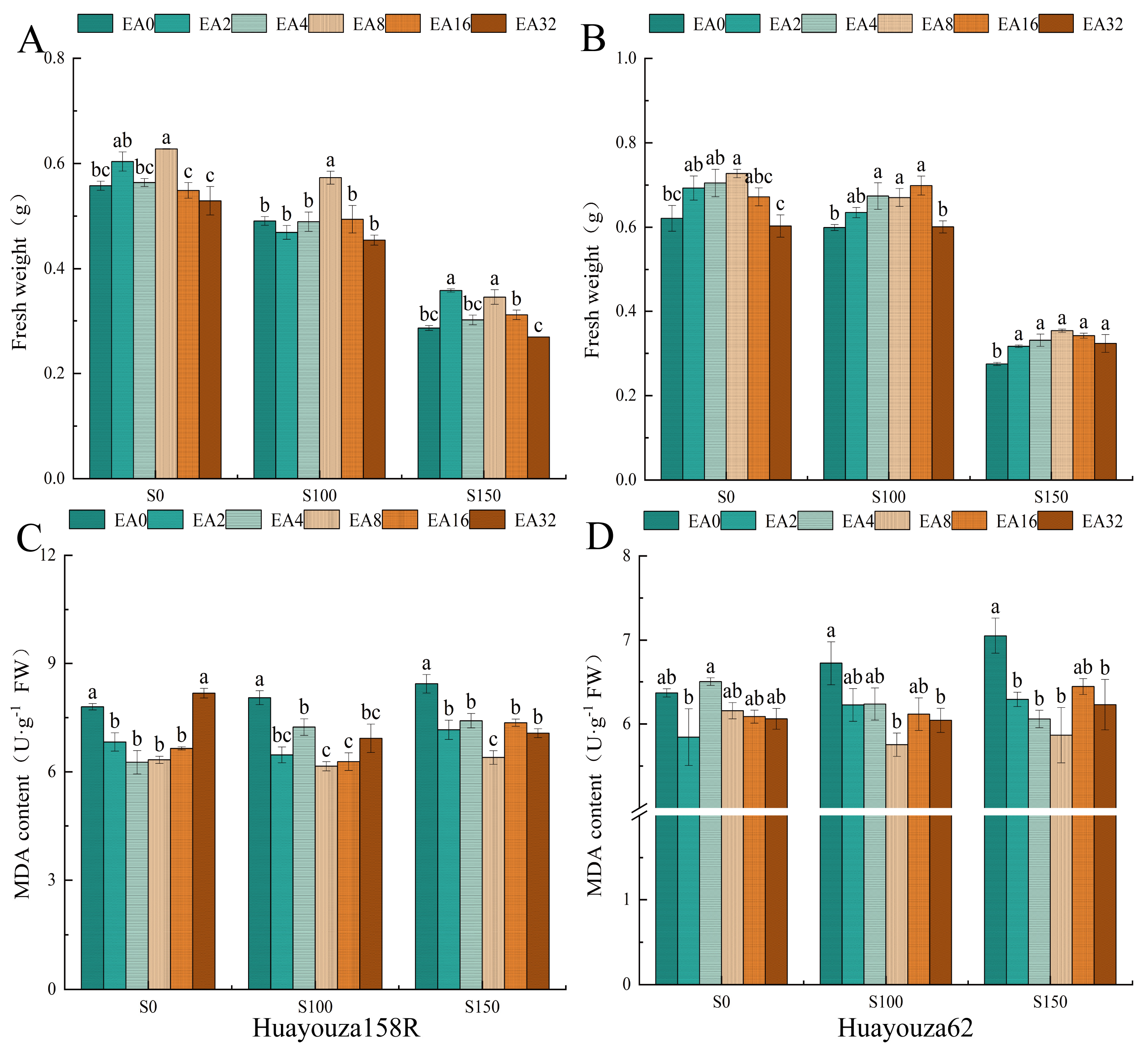
Figure 3.
Effect of Pro-Ca primed on morphological development of oilseed rape under salt stress. Plant height (A), root length (B), leaf area (C), and seedling strength index (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 3.
Effect of Pro-Ca primed on morphological development of oilseed rape under salt stress. Plant height (A), root length (B), leaf area (C), and seedling strength index (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
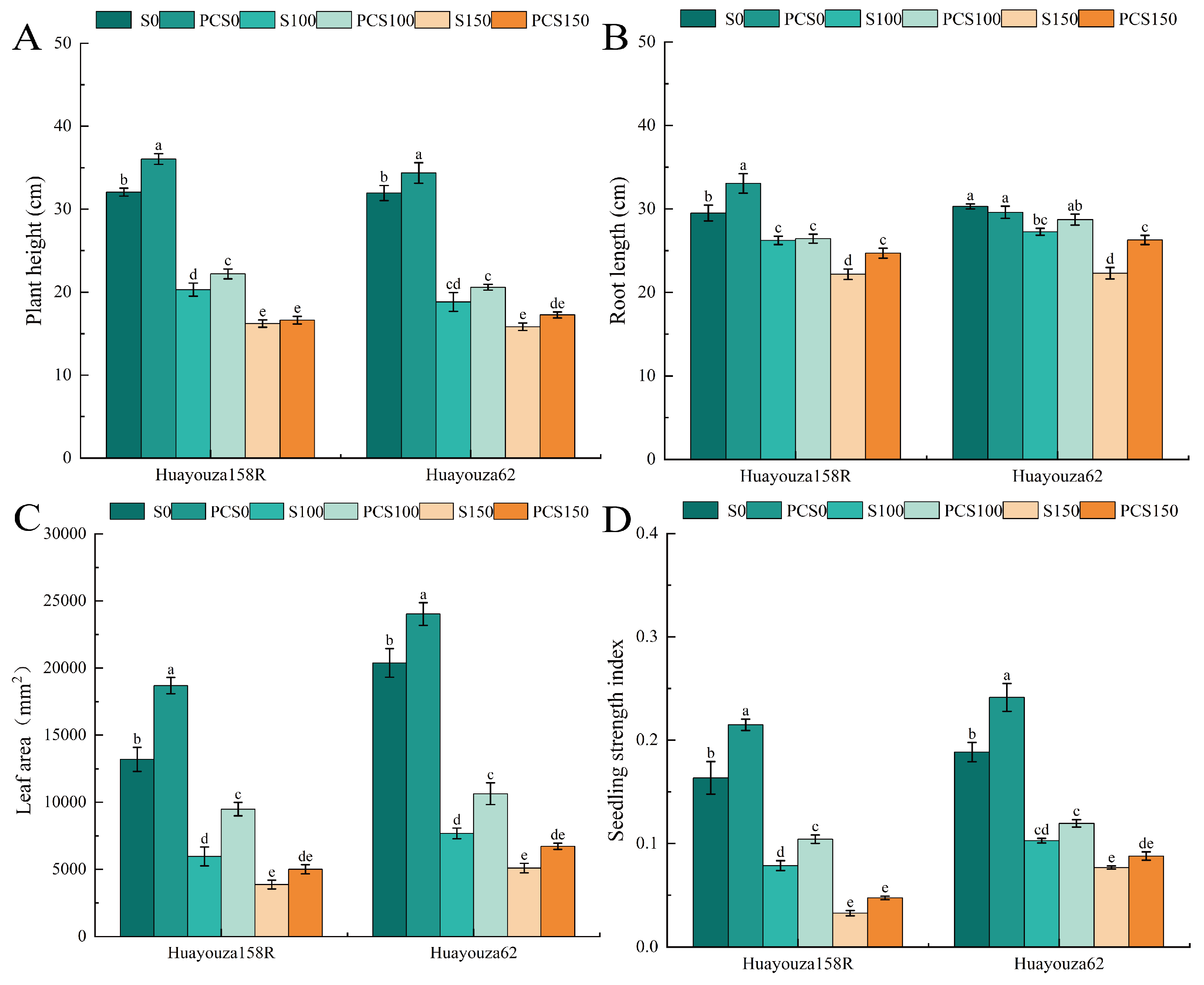
Figure 4.
Effect of Pro-Ca primed on photosynthetic pigments in oilseed rape under salt stress. Chlorophyll a (A), chlorophyll b (B), carotenoids (C), total chlorophyll (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 4.
Effect of Pro-Ca primed on photosynthetic pigments in oilseed rape under salt stress. Chlorophyll a (A), chlorophyll b (B), carotenoids (C), total chlorophyll (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
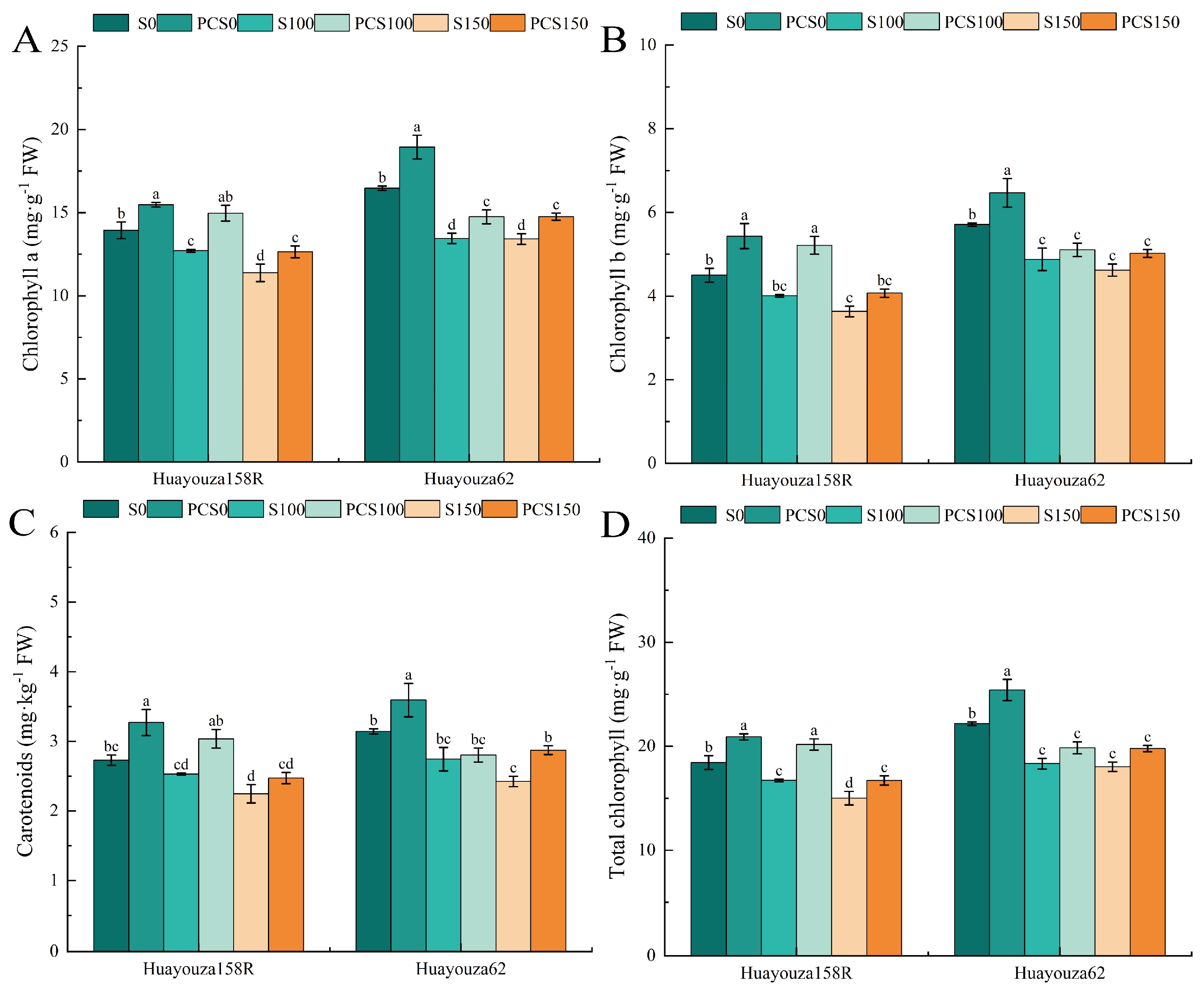
Figure 5.
Effect of Pro-Ca primed on photosynthetic properties of oilseed rape under salt stress. Pn: net photosynthetic rate (A), Gs: stomatal conductance (B), Ci: inter-cellular CO2 concentration (C), Tr: transpiration rate (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 5.
Effect of Pro-Ca primed on photosynthetic properties of oilseed rape under salt stress. Pn: net photosynthetic rate (A), Gs: stomatal conductance (B), Ci: inter-cellular CO2 concentration (C), Tr: transpiration rate (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
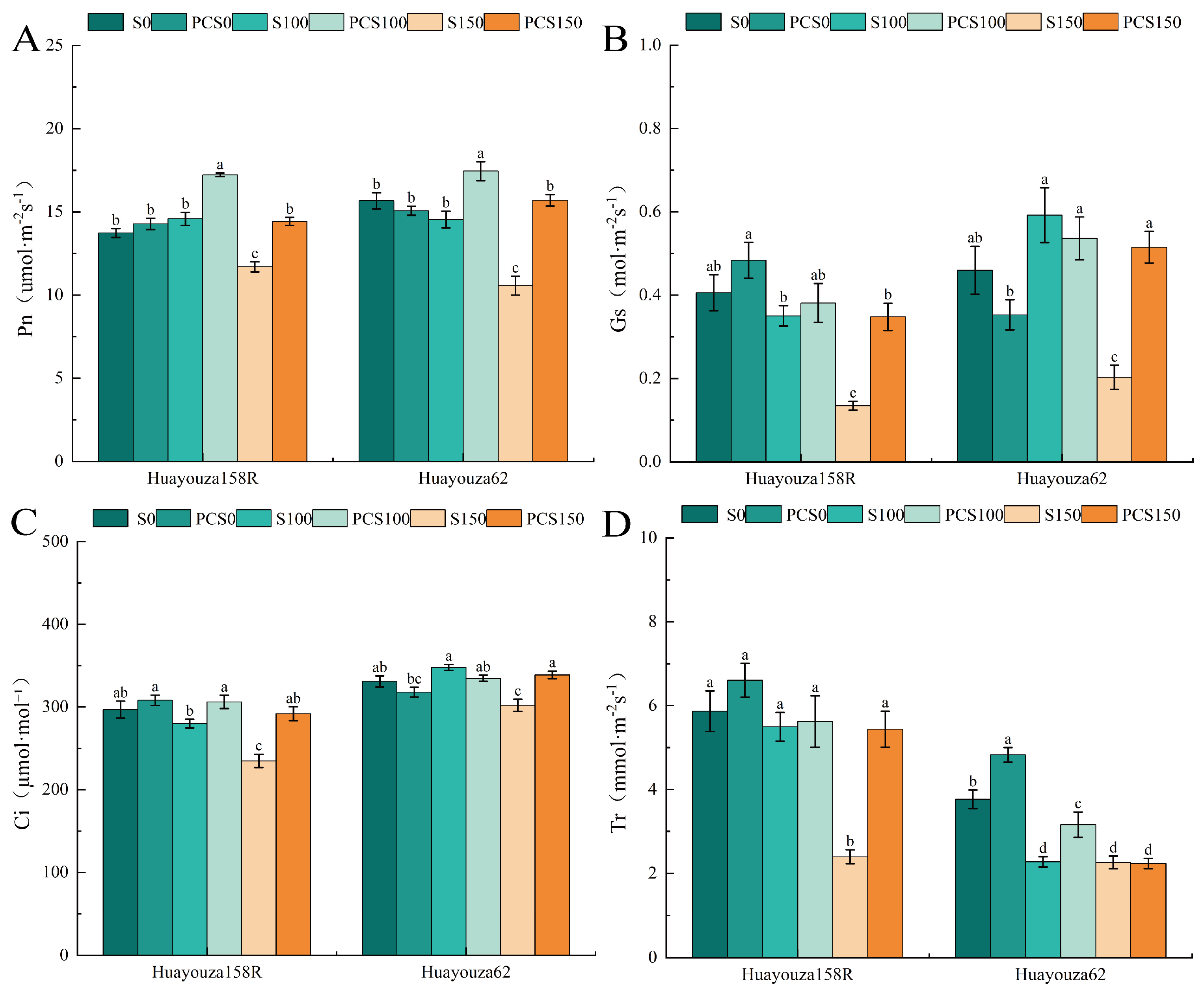
Figure 6.
Effect of Pro-Ca primed on membrane lipid peroxidation in oilseed rape under salt stress. Leaf relative conductivity (A), MDA: malondialdehyde (B). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 6.
Effect of Pro-Ca primed on membrane lipid peroxidation in oilseed rape under salt stress. Leaf relative conductivity (A), MDA: malondialdehyde (B). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
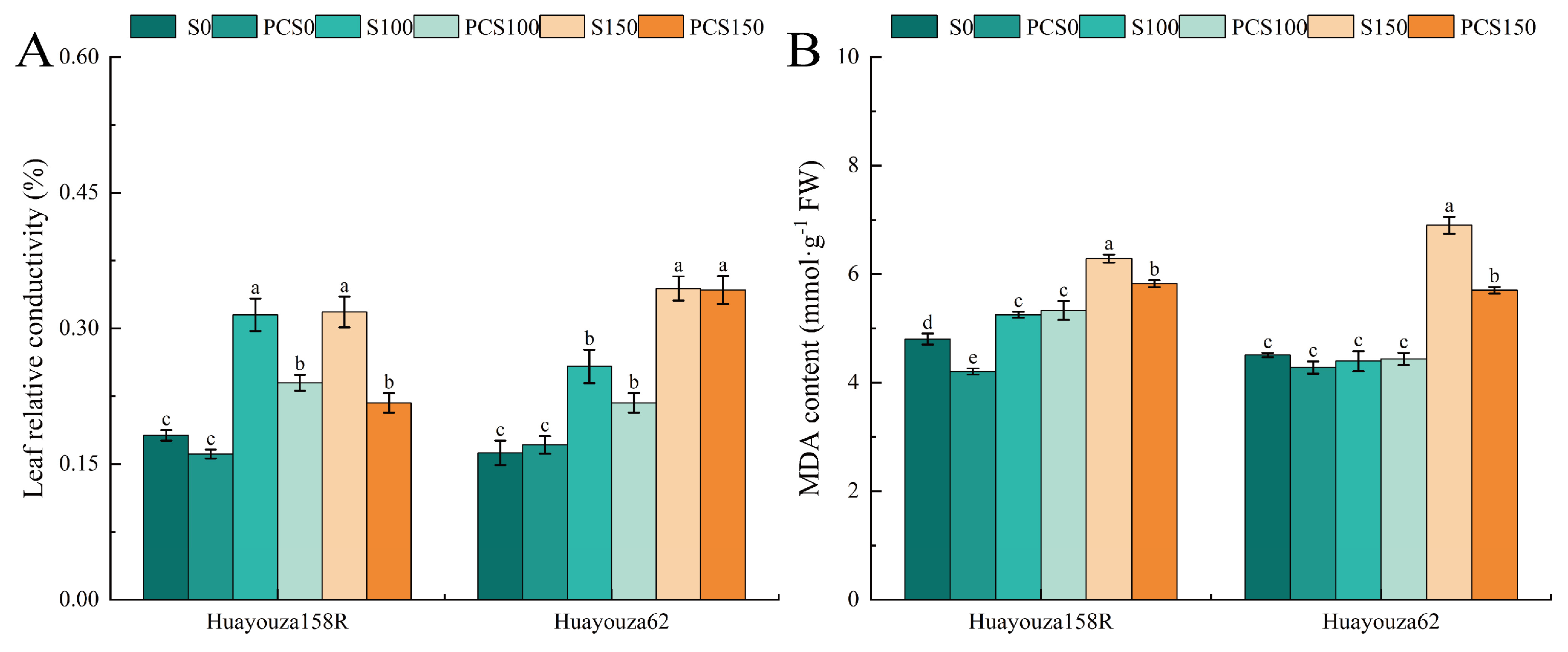
Figure 7.
Effect of Pro-Ca primed on antioxidant enzymes in oilseed rape under salt stress. SOD: superoxide dismutase (A), POD: peroxidase (B), CAT: catalase (C), APX: ascorbate peroxidase (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 7.
Effect of Pro-Ca primed on antioxidant enzymes in oilseed rape under salt stress. SOD: superoxide dismutase (A), POD: peroxidase (B), CAT: catalase (C), APX: ascorbate peroxidase (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
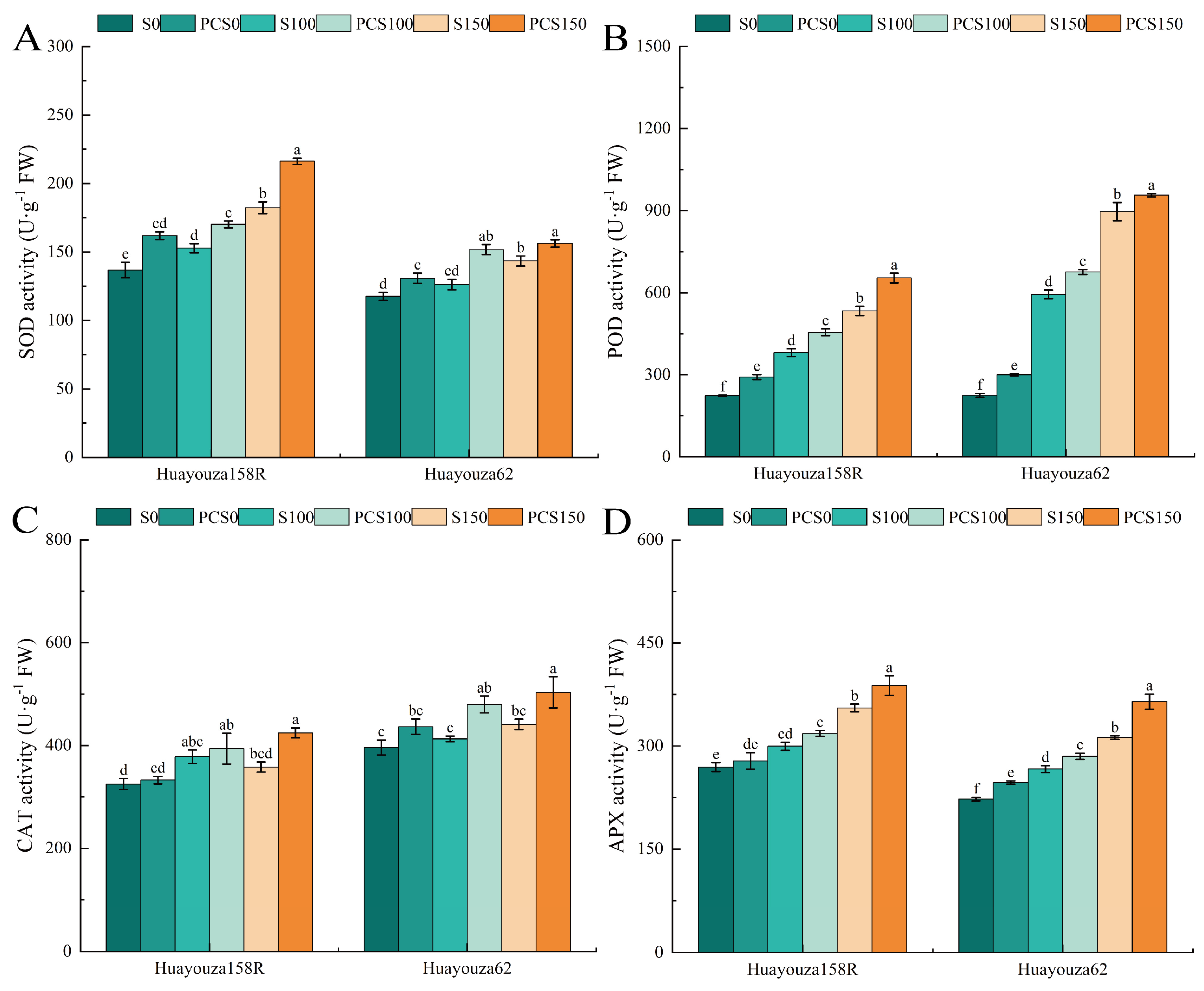
Figure 8.
Effect of Pro-Ca primed on ion content of oilseed rape under salt stress. K+: potassium ions (A), Mg2+: magnesium ions (B), Na+: sodium ions (C), K+ /Na+: potassium to sodium ion ratio (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Figure 8.
Effect of Pro-Ca primed on ion content of oilseed rape under salt stress. K+: potassium ions (A), Mg2+: magnesium ions (B), Na+: sodium ions (C), K+ /Na+: potassium to sodium ion ratio (D). Data are expressed as mean ± standard error and are not significantly different with the same letter by LSD test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



