
Submitted:
18 March 2024
Posted:
21 March 2024
You are already at the latest version
Abstract
Alterations in the composition and diversity, metabolism, and products of the microbiome, and of the gut microbiome (GM), have been shown to be closely associated with the onset and progression of many human diseases, including cancer (i.e. hematological neoplasms). Acute lymphoblastic leukemia (ALL) is the most common form of childhood malignancy. Affected cases present typical alterations of GM, followed by inflammation, which contribute to its progression, response to therapy, as well as possible relapses. Recent evidence also reports that GM influences the development and functions of the newborn's hematopoietic system through the process of developmental programming during fetal life, as well as its susceptibility to the onset of onco-hematological pathologies, namely ALL. Furthermore, in children with ALL, GM has been found to vary in composition, variety, and functions during the clinical stages of ALL, and such variation may influence and predict the complications and prognosis of ALL after chemotherapy treatment or stem cell hematopoietic transplant. Here, we suggest some therapeutic strategies, which can be applied at two levels of intervention to recover the microbiome, and consequently prevent/delay ALL or arrest its progression.
Keywords:
Acute lymphoblastic leukemia (ALL)
; microbiome
; GM
; therapeutic strategies
1. Introduction
Leukemia represents a typical and heterogeneous group of hematological neoplasms, having the characteristic of clonal dominance of hematopoietic stem cells arrested in development and maturation multipotent progenitors [1]. This pathological condition is highly prevalent in children under 15 years of age (accounting for 30% of all cancers), where it represents the most common form of childhood malignant neoplasm. Precisely, the children leukemia forms mainly consist of acute lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML). Children within the age range of the 2-4 years are the commonest ALL cases and show an overall survival (OS) of above 90% [2]. ALL is characterized by a substantial proliferation of lymphoblasts accompanied by a decrease in the number of circulating mature cells, which cause bone marrow failure syndrome [3,4,5]. ALL may be of two forms: the first with a prevalence of 80% in children, and causing alterations in the B cell lineage, and the second, uncommon, characterized by affecting T cells. The major ALL complication, which can lead to mortality, is the involvement of the central nervous system (CNS), with the development of typical symptoms, including the loss of balance, headache, fainting, nausea or, more rarely, swallowing difficulties. The severity of such symptomatology depends on the leukemic infiltration in CNS, and consequently in its entity or better in the amount of CNS areas involved [6,7,8]. In addition, ALL appears underestimated, because underdiagnosed for being asymptomatic in most cases. Diagnosis of ALL consists in a complete cell blood count, a bone marrow puncture, and a biopsy. Disease confirmation is given by the presence for the 25% or more of lymphoblasts in the bone marrow, after the analysis of tissue sample, trough cytogenetic techniques or fluorescence in situ hybridization (FISH). Precisely, they consent to detect specific chromosomal abnormalities, and consequently of identifying specific subgroups associated with different disease outcomes [9]. Anemia, leukopenia, and a low number of platelets justify the typical symptoms of ALL. They determine the development of fatigue, weakness, shortness of breath during normal physical activities, light-headedness, dizziness or faintness, headaches, and a pale complexion, as well as recurrent fevers, frequent infections, easy bruising, red spots on the skin, called petechiae; frequent or severe nosebleeds; bleeding gums; and, in women, more frequent menstrual periods. Additional symptoms include weight loss or loss of appetite, swollen glands, bone and joint pain, difficulty breathing and an enlarged spleen or liver [10,11,12].
The ALL pathogenesis includes genetic alterations and environmental factors, i.e., alcohol, cigarette, and drugs, which, during pregnancy, can cause genetic mutations of the fetus [13]. Radiation exposure, chemicals and infections also represent other environmental factors, which can also contribute to ALL development. To date, however, the deep relationship between these factors and ALL investigations, and the exact mechanisms involved in ALL pathophysiology are still not clear [14]. Nevertheless, in the last years the interest has been focused on microbiome, since in adult ALL cases the role of microbiome has emerged in both development and progression, as well as in the response and side effects of chemotherapy drugs, infection during treatment, and therapeutic efficacy [15,16,17]. In addition, current evidence also reports the capacity of parents’s microbiome, and precisely of gut microbiome, to impact the newborn’s hematopoietic system (i.e., onset and function) via fetal developmental programming and its susceptibility after the birth to develop onco-hematological neoplasms [13]. Here, we concentrate our attention on describing and discussing on the literature evidence about the role of gut microbiome in the pediatric ALL, as well as in suggesting some therapeutic strategies aimed to recovery microbiome.
2. Microbiome and ALL
In the last decade, recent evidence has demonstrated that another crucial determinant (trigger) of the development of several diseases is the microbial dysfunction, or better the dysbiosis. Dysbiosis has been shown to impact the development and progression of many human diseases, ranging from immune diseases, cardiovascular diseases, neurodegenerative diseases, and cancer [18,19,20,21,22,23]. The very relevant role of microbiota and microbiome (its genome) has led to consider them as the second system of control of the functions of all the organs, tissues of human body after the brain. Indeed, it has been defined as the second brain, and regulates both the human health and disease. Among the human diseases, cancer onset and progression have been demonstrated to be significantly associated with microbial dysbiosis [24,25]. The contribution of the microbiome on cancer development has been under study since 2012 [26], and research explorations on adult ALL have started two years later [27]. In contrast, investigations on childhood ALL have been reported only after 2016 [28]. Probably, diverse issues have limited such studies, including contaminations during preparation via buffers or handling, or poor processing choices due to high sequencing costs. However, the recent progresses in the omics technologies and the reductions in costs in such investigations have facilitated the microbial research attention, and consequently of increasing the number of research studies on microbiome in both adult and children leukemia [29,30]. Of note, are the studies based on the Pax5/- and Sca1-ETV6-Runx1 mice model that first have demonstrated the key role of genetic predispositions in contributing to the alterations in the gut microbiome (GM), and in turn such variations can further impact the hematopoietic system (HS) and leukemia development. This evidence suggests of considering the analysis of GM as diagnostic leukemia tool in the future. In addition, the interesting results from epidemiological studies lead to suppose that a remarkable trigger of leukemia are the infections, impacting the most common ALL subtypes; infections during early childhood can determine the lack of microbial exposure in infancy, and consequently this can promote the emergence of this disease [31]. Remarkable also the results of a study conducted in 2018 on 42 pediatric ALL patients at different timepoints of therapy has revealed the presence of microbiome, which vary in function of the type of cancer. Such results, confirmed by another study performed in 2020, have suggested that the human microbiome might be a diagnostic tool for specific cancer types. Below, diverse concepts are cleared for better understanding the close relationship of microbiome and ALL in children [31,32,33].
3. Microbiota and Microbiome and Its Modulating Effects on Hematopoiesis in Health and Disease
Now, the research ‘s interest in exploring microbiota is rising in an exponential manner because of its key role in altering numerous mechanisms and processes related to human health and disease, such as cancer (e.g., ALL). Microbiota represents an important component of our body, constituted by many microorganisms (i.e., bacteria, viruses, and fungi). Precisely, it stims that microbial cells might be about 3.8 × 1013, with 3,000 species. The microbiota, organized in niches, is localized in many anatomical structures of our body: oral, vagina, skin, and gut. However, the prevalent region of microbiota is the gut, consequently here is defined as the gut microbiota (GM), with 500 species of bacteria, as well as yeasts, parasites and viruses. The major bacterial species found here include the phyla Firmicutes and Bacteroidetes. In adults, Firmicutes are the most abundant, followed by Bacteroidetes and Actinobacteria [34]. Firmicutes can be divided into Clostridium, Ruminococcus, and Eubacterium. The relationship between the bacterial species Bacteroidetes and Firmicutes has been shown to play an important role in health and disease [35]. The GM colonizes the newborn after birth and is composed of native flora and transient flora in food intake [36]. Accordingly, every person is characterized to have a specific and unique composition of GM, which evolves during the individual’s life, from childhood to adulthood [37]. Thus, GM shows dynamism and plasticity, which modulate its functions and the function of other tissue, organs, and systems, such HS [38]. The microbiota, and particularly GM, functions by mediating actions and effects, which are all protective, contributing to the body ‘s homeostasis. It protects via the body’s immune responses the host by pathogens, has metabolic activities, contributing to the synthesis of amino acids, vitamins, and enzymes, and indirectly controlling cellular proliferation, structure, development, and composition of the immune system, intestine through the development of intestinal villi and intestinal epithelial cells, contributing to the formation of the epithelial barrier.
Beyond these functions, GM also shows influences on other organs, for instance the CNS, in which metabolites released by the microbiota affect mood, stress regulation, and instinctive behaviors on cardiovascular and endocrine-metabolic systems; the cardiovascular system also results modulated by GM through disease risk factors causing hyperlipidemia, atherosclerosis and type 2 diabetes; and digestive system by the action of microbiota’s products absorbed from the gut and transferred to the liver [39] (see Figure 1). The GM also exerts a significant influence on individual’s physical and mental health. However, even if generally considered beneficial, some GM microbes can cause a potential danger to the human organism [38]. When the homeostasis of the microbiota is disturbed, most of it becomes pathogenic, remodeling occurs that alters the sophisticated balance of the microecosystem in the gastrointestinal tract, contributing to a variety of pathological conditions and negatively affecting the physiological processes of the host [39]. This imbalance, called “microbial dysbiosis”, shows a reduction of microbes favoring host health, and an excessive presence of pathogenic microbes with deleterious functions [40]. For instance, Bacteroides fragilis and Fusobacterium nucleatum, are known to induce a pro-inflammatory state in the colon, which could potentially increase oncogenic proliferation in the mucosal microenvironment [41,42].
Many risk factors can act on our microbiota, the first is immutable and is genetically hereditary, it cannot be modified; however, other factors can instead be modified, such as the environment and the way we are born and nourished during the first 1000 days of life, as well as the programming of the development of all organs, such as HS, during fetal life, depending on parents, mother and father [43]. Malnutrition or overnutrition in pregnancy is involved in influencing fetal programming, as well as other maternal and paternal factors modulate fetal programming. The components of gut flora are highly dependent on the influence of the environment in which we live such as rural, urban, industrialized, and non-industrialized areas [44]. The dysbiosis can inhibit the colonization of beneficial probiotic bacteria, promote harmful entero-pathogens proliferation, and alter innate receptor and cytokine signaling, thus affecting the immune system [45]. The dysbiosis can contribute and be associated with a myriad of pathological conditions, such as inflammatory bowel diseases, immune-mediated diseases, and neoplastic conditions, including hematological malignancies [40].
3.1. The GM Close Relationship with Hematopoiesis
Recent evidence reports that GM has a close relationship with hematopoiesis. Accordingly, the Balmer group have reported for the first time in 2014 that the different composition of the microbiota significantly influences both the configuration and quantity of myeloid cell lineages [46]. This could explain why the microbiota with fewer types of microorganisms constitutes an unfavorable factor for the success of an allogeneic stem cell transplant, as highlighted by Taur and collaborators. In fact, this unfavorable condition has been proven to be significantly related to a raise in mortality and a resulting lowered survival of transplanted patients [47]. Analogous findings have been found in mice treated with an antibiotic therapy [48]. In addition, it has been reported in mice that some aliments able to impact the microbiota, can indirectly modify HS function. For example, aliments rich in fibers specifically change the function of some microbiota ‘s bacteria, able to transform these aliments into short-chain fatty acids (SCFAs). Therefore, mice subjected to a diet rich in fiber’s aliments have been found to have higher bone marrow levels in myeloid precursors [49]. To the same manner, high systemic lipopolysaccharide levels related to GM dysbiosis have been showed to induce myelosuppressive effects via chronic activation of the immune innate Toll like receptor-4 expressed in hematopoietic stem cells (HSC) [49]. These remarks allow to suppose that the relationship between alteration of microbiota and the onset of several immune disorders, are linked with the development of age-related diseases.
As reported above, malnutrition or overnutrition in pregnancy is involved in influencing fetal developmental programming. Besides, raising evidence shows the role of environmental experiences to influence intestinal microbiota, in early life, emphasizing a “microbial programming phenomenon” (widely quoted in [51]). In this context, obesity has been displayed to be a significant challenge for the health of both mothers and children. In children, obesity impacts the development of both microbiota and immune responses. Despite its importance, it is unclear whether weight during pregnancy, as well as the composition and functional quality of the maternal microbiota, can mediate adverse effects on offspring. However, Kozyrskyj and colleagues have reviewed literature from human studies and established that maternal obesity can modulate both the composition and function of gut microbiota in new-born [52]. Vertical transport of microbiota and the release or inhibition of release of their metabolic products have been hypothesized as possible mechanisms [53]. In the next paragraphs, the GM effects on hematopoiesis are better described, as well as the role of developmental programming of HS.
3.2. Experimental Evidence for the Effects of Microbiota on Hematopoiesis
Research on the GM topic is still in its primary development; however, some promoting results concerning the relationship between GM and hematopoiesis are existing. In this context, it has been demonstrated that several GM products impact hematopoiesis, and bone marrow cellular pool composition correlates significantly with GM structure and heterogeneity [54]. Of substantial help are the mouse studies, because they significantly contribute to identifying GM effects on HS. Particularly, studies in germ-free mice have evidenced reduced immunological functions. Such altered phenotype has been demonstrated to be associated with an increased susceptibility towards intestinal and systemic infections [55]. These results emphasize the value of GM functions. As above reported, GM can execute numerous, specific biochemical and molecular reactions, and consequently to synthesize a wide spectrum of molecules and metabolites that human cells are not able to produce. Some GM compounds exercise an effect on the hematopoietic functions. For example, SCFAs interact with the immune system, through the G-protein-coupled receptors (GPCR), which are expressed in many immunological cells. In diverse cases, SCFAs mediate an anti-inflammatory effect through the inactivation of the NF-κB pathway. From a hematopoietic point of view, SCFAs promote the generation of specific subsets of T cells [56,57]. In another study in mice, the impacts of using fermentable fibers as dietary supplementation on the immune system have been analyzed. The results have reported the increase in fiber bioavailability leads to the expansion of members from the phylum Bacteroidetes. In turn, this results in a significant local (intestinal) and systemic increment of SCFA. Boosted SCFA concentration improves the proliferation of dendritic cell precursors in the bone marrow through the activation of GPR41. An interesting datum is that GM variations are accompanied by alterations in the lung microbiota, even though they do not cause an increase of SCFA levels, possibly due to the lack of suitable substrates [58]. As mentioned earlier, antibiotic treatments often cause severe hematopoietic damages through suppression of its activities. Such a condition is not caused to the direct effect of antibiotic drugs. Accordingly, significant data have been obtained by Josefdottir and co-workers [59]. They have explored the harmful effect of antibiotics by depleting mouse GM via antibiotic treatment. GM depletion in mice determines a substantial damage of the hematopoietic function, including reduced cell count in the blood corresponding with decreased cellularity in bone marrow. In the study, they have established that the GM depletion and not due the antibiotic treatment caused this acquired hematopoietic phenotype. GM transplant demonstrated, indeed, restoring physiological hematopoiesis. They have remarked that the immunological impairment was linked to the GM alterations induced by antibiotics, causing an immune-phenotype identical to the one of Stat-1 deficient mice. This has led to suppose that the suppression of the Stat-1 signaling induced by GM suppression is systemic and not limited to the intestinal area [60].
The global abovementioned results are extremely far-reaching if it considers the GM plasticity and variability. GM is extremely vulnerable to environmental factors and nutritional factors, as showed in twin studies [61,62]. Moreover, GM alterations correlate with age, even if some specific features of the centenarians ‘s GM have been correlated to the longevity phenotype [62,63,64,65,66]. Taken together, the great connection between microbiota and hematopoiesis constitutes a new significant factor, able to modulate the fate of the individual immune-biography traits [63], in health and disease [66].
3.3. Developmental Programming of HS and the Susceptibility of Leukemia
Well established evidence has recognized that the body human systems, such as nervous, endocrine, immune and cardiovascular systems have been programmed during the fetal development [67,68,69,70,71,72,73,74,75,76]. The fetal period (and particularly the embryonal phases; “critical windows”) is highly susceptible to many environmental stressors, that can impact the life after the born, the adult period and its health status [77]. Thus, it characterizes a essential period of development, distinguished by a high rate of cellular proliferation and plasticity in the developing systems. These remarks are significantly associated with the new concepts on the high vulnerability of our systems to maternal, environmental and intrauterine stressors, during their embryonic development [51,67,68,69,70,71,72,73,74,75,76].
Harmful developmental conditions can influence the (epi)genetic and physiological processes of fetal development, thereby permanently changing the construction and functionality of hypothalamic-pituitary-adrenal (HPA) axis and systems of the progeny, predisposing them to disease development during the life after the born [51,67,68,69,70,71,72,73,74,75,76]. All the illustrated concepts imply that HS function, destiny, and disease in the life after the born are clearly dependent on the developmental programming processes. Endogenous and gestational risk factors have been demonstrated to influence this process, and the related mechanisms and effects induced on hypothalamic-pituitary-adrenal (HPA) axis, immune, endocrine, and metabolic systems, inducing HS developmental programming, and the outcomes in life after the born [13].
Overall, these concepts have led to interesting evidence on the critical relevance of the close relationship between the maternal and fetal micro-environment, and its significant contribution in HS developmental programming and susceptibility to diseases, such as leukemia [67,68,69,70,71,72,73,74,75,76,77,78]. Accordingly, some studies have reported that the prematurity at birth and intrauterine growth restriction (IUGR) may be the major risk factors for alterations in HS, contributing to HS increased susceptibility to HS diseases, such as leukemia [67,68,69,70,71,72,73,74,75,76,77,78].
4. Close Relationship between ALL and Microbiota
Recent evidence reports microbiome alterations in children with ALL. They have been observed in all the steps of the ALL management: at both the time of diagnosis, during treatment, and in contributing to the onset of complications and differences observed after the completion of therapy. At the onset of disease, a small variation in the oral and gut microbiota/microbiome of ALL children patients is already detected [79]. During the ALL treatment, the administration of chemotherapeutics and antibiotics causes ulterior alterations in the microbiome, by determining an additional reduction in the diversity of composition and types of the microbiota’s microorganisms. Such results in the dominance of Enteroccocaceae being predictive of infections. Furthermore, microbiota deterioration, including Faecalibacterium depletion, may extend even for several years after completion of ALL treatment, inducing potential long-term health effects. Nevertheless, it would not be clear whether the observed alterations represent a fundamental trigger for the development of ALL or are caused by immunological variations that precede the onset of ALL. Monitoring large pediatric cohorts could be useful to provide direct evidence whether genotype already determines microbial composition, even without disease onset. Taking all of this together, there is always a clear need for precise characterization and modulation of patients’ microbiomes during therapy, to better understand the microbial influence on leukemogenesis, minimize side effects, and improve the efficacy of treatment [79].
In the next paragraphs, we stress these concepts, for better understanding the message of such review.
4.1. Gut Microbiota and Children Susceptibility for ALL Onset
The GM role is manifest from birth of newborns, and its composition and delivery from mother to children can significantly influence the onset and type of cancer. For example, children born via caesarean partition show a higher ALL risk, because they are not influenced by the maternal vaginal microbiota, which can facilitate them to form the later gut microbiota [79]. However, two recent studies have reported a non-association between the type of delivery and the occurrence of brain cancer or lymphomas [1,80]. Another study, and precisely a meta-analysis conducted on 25 studies, 18 of which met all inclusion criteria, has evaluated the association between breastfeeding and the ALL onset. All meta-analyses of subgroups of the 18 studies have significantly shown that from 14% to 19% of all childhood leukemia cases may be prevented by breastfeeding for 6 months or more. The reason is linked to the immunomodulating effect of the mother’s milk, by which specific nutrients, antibodies, or anti-inflammatory factors are delivered to the baby [81]. Based on such interesting results, it suggests that the early GM in newborns is significantly dependent on and affected by the type of delivery, the form of feeding, hospitalization of the newborn and the use of antibiotics. Altogether, these mentioned factors appear to favor or not the development of an advantageous or helpful microbiome in newborn, characterized by relatively higher richness in Bacteroides and Bifidobacterium and lower numbers of Clostridium difficile or Escherichia coli [82].
4.2. Alterations in Microbiome at the Children ALL Diagnosis
As above reported, the alterations in the microbiome may interest both oral and GUT microbiome. The oral mucosa constitutes the first natural protective barrier, and its changes can favor the onset of ALL complications and consequent diseases. Accordingly, it is imperative to know the ALL influence on the oral mucosa at the time of diagnosis and during chemotherapy. However, very limited number of studies has investigated this aspect. Precisely, one study has focused our interest on such issue, reporting that some ALL cases are affected by tooth and show jaw pain, gingival swelling, and loose teeth, which might be attributed at an impaired microbiome [31,83]. The analysis of oral samples from newly diagnosed ALL patients, when compared with healthy control children has demonstrated that Firmicutes and Fusobacteria were significantly different. Precisely, ALL patients were characterized by significantly higher levels of Firmicutes, and the reduced presence of Fusobacteria. Among Firmicutes, Granulicatella and Veillonella, were more abundant in ALL patients [83]. Taken together, ALL patient samples showed a diminished microbial variety and smaller abundance than controls, demonstrating a dysbiosis able to increase the susceptibility of ALL cases to infection risks. Certainly, further studies need to confirm this aspect.
Investigations on GUT microbiome from ALL cases at diagnosis have reported an increased abundance of Faecalibacterium, Bacterioides, or Parabacterioides and Firmicutes [84,85]. However, the diarrhea-causing Clostridium has been observed to be less abundant in ALL children, where the Lachnospiraceae, including Roseburia and Blautia, produce SCFAs with an anti-inflammatory effect [86]. Despite this, some ALL patients during the initial disease manifestation have subjected at antibiotic treatment to limit their increased susceptibility to infection. Consequently, such treatment determines a significant decrease in ALL patients treated of microbiome diversity [85]. Based on these results, Bai and coworkers have suggested that Bacteriodales and Enterococcaceae of the phylum Firmicutes might be used as a promising biomarker for ALL, but only in children without antibiotic treatment [85]. In 2020, Liu and coworkers on the largest cohort of cases examined until now, and precisely on 70 newly diagnosed ALL patients, enrolled at time of diagnosis [84], have observed no difference in the alpha diversity of microbiome, and confirmed the increase in Bacteroides species, hence, in the beta diversity. Specifically, Bacterioides uniformis and Bacteroides fragiles were significantly augmented in cases of ALL in pre-chemotherapy step.
4.3. Alterations in Microbiome during Children ALL Treatment with Chemotherapy
The ALL treatment with chemotherapy starts immediately after the diagnosis and is characterized by 3 phases over 2–3 years. Such treatment is well recognized to modify the bacterial composition and cause a shift in reduction of the white blood cell counts. Such determines the development of typical symptoms in childhood ALL, including anemia-derived fatigue, fever, infection, and even easy bleeding. Therapy starts directly after diagnosis, with chemotherapy given in 3 phases over 2–3 years. In addition, infections are characteristic. Accordingly, a study conducted in 409 newly diagnosed ALL patients, has demonstrated the presence of 1313 microbiologically derived infections during therapy [87]. This has led to hypothesize the role of impaired intestinal microbiome in the significant increase susceptibility of the documented infections. On the other hand, a study has demonstrated that the composition of microbiome changes comparing it before and after the treatment, with a specific reduction of Lachnospiraceae and Roseburia in patients [88,89]. A larger study from Hakim and cowokers on 199 ALL patients conducted during the three different times of chemotherapy has evidenced not significant differences in the mean variety of microbiome, which showed baseline features; however, after chemotherapy the microbial diversity significantly decreased, with a different bacterial composition: Bacterioidetes, Faecalibacterium, Ruminococcaceae, Actinobacteria, and Verrucomicrobia significantly reduced, while other taxa, Clostridiaceae, Streptococcaceae, Lactobacillaceae, Enterococcaceae, and Firmicutes, augmented [88].
However, a longitudinal observation study from Chua and coworkers has analyzed the temporal differences in the gut microbiome from seven ALL patients before, during and after chemotherapy and from controls [89]. They have observed that antibiotic treatment before chemotherapy was not the cause of the differences observed in large inter-individual variability of ALL patients compared to healthy children in microbiome composition. Nevertheless, it has been detected that Bacteroidetes was significantly enhanced before chemotherapy, although its large quantity declined after therapy. In contrast, Firmicutes and Actinobacteria, increased following chemotherapy to a similar level of healthy controls. Precisely, five genera resulted to have a lower abundance after therapy: Bacteroides and Prevotella, belonging to the phylum Bacteroidetes; Fusobacterium; and Atopobium and Corynebacterium from Fusobacteria and Actinobacteria, respectively. Furthermore, only Bifidobacterium (Actinobacteria) was significantly higher in the post-chemotherapy samples. This commensal occurs immediately following birth and results to be able to use human milk oligosaccharides, as well as other carbon sources. Furthermore, it has a protective role in preventing intestinal inflammation in infants since it prevents an increase of Proteobacteria which are associated with dysbiosis and negative health outcome [87,90].
Finally, long-term adult survivors of pediatric ALL who completed therapy at least 5 years have exhibited decreased microbial variety than healthy controls, with a remarkable enhancement of Actinobacteria and depletion of Faecalibacterium [89]. Furthermore, the researchers in these individuals, have detected enhanced T-cell activation and chronic inflammation, suggesting a correlation between dysregulated microbial taxa and immune dysregulation [89]. The increased infection risk in survivors, a high prevalence of chronic health conditions and an elevated risk of mortality and morbidity have been also investigated [91]. Thus, microbial dysregulation determined by the influences of chemotherapy and antibiotics during ALL treatment may have long-term effects on the development of other diseases, such as obesity or diabetes, in adult survivors of pediatric ALL [92,93].
5. Diverse Levels of Strategies to Recovery Microbiome and Prevent All Onset
5.1. The First Level of Health-Promoting Strategies
Microbiome is crucial for the life of an individual, and comparably to other systems, it shows an interindividual variation, particularly in ALL cases. Today, the growing evidence suggests that these differences originate not in later stages of life cycle of an individual, but rather in the early periods. Such last constitute critical windows of rapid growth under a strong epigenetic remodeling, and, showing, thereby, high phenotypical dynamicity and plasticity during which an organism results predominantly susceptible to environmental conditions, harmfully impacting tissue, organ and system development and disease tendency later in life or after some years from the birth. A clear example are the ALL cases (widely quoted in [51,94]).
We, here, have reported, how HS is developmentally programmed, as other systems, and, thereby, in adulthood, or after some years at the birth, it results the representation of the developmental programming, and particularly the consequence of numerous developmental programming events. Among these, the endothelium (re) programming is first included, being the hub of the HS origin, homeostasis, and function, as well as of other human systems [95,96,97,98]. In turn, endothelium (re)programming influences HS and immune programming with the cooperation of hormonal and metabolic alterations (altered HPA axis and increased release of cortisol, but also of other crucial hormones). After, they induce epigenetic and microbial programming in progeny. The combination of all these programming conditions makes the progeny ‘s HS susceptible to assume long-term structural and functional alterations, that permanently modulate its functions, raise disease risk and velocity to get old. However, their prevention, as well as the prevention of consequent pathologies at long or short (i.e., ALL) onset, may be feasible, and consequently represent a certainty and not an illusion. Accordingly, evidence supports the positive effects of a healthful diet (i.e., Mediterranean) physical activity, low stress, non-usage of smoking, alcohol, and drugs during pregnancy [99,100,101,102,103].
Here, we first suggest of recommending such strategies fat both parents who want to have kids, before and during pregnancy. McGowan and Matthews 2018 have supported such strategies, stressing the role of both parents and their lifestyle in developmental programming. Precisely, they affirm that parental danger (not only maternal, but also paternal), related to stress and/or to their changed clinical status related to be affected to eventual diseases (i.e., hypertension, type 2 diabetes) or to have an insalubrious lifestyle (linked to consumption of alcohol, use of drugs, diet, or to be smoker and sedentary), causes deep biological effects on both fetal development and subsequent functionality of HPA axis and specific systems. Furthermore, these effects show of being species-, gender-, and age-specific, and vary in function of timing and duration of exposure, as stressed by McGowan and Matthews (2018) [75].
5.2. Some Therapeutic Approaches of Second Level
In addition to the first level of health-promoting strategies, we also recommend of utilizing some therapeutic approaches of second level to apply during prenatal and neonatal periods or in adult life. Among these, importance has been given to the pharmacological targets/targeting of pathways involved in the crosstalk of endothelium-HS, cellular/tissue reprogramming, use of miRNAs, and modulation of microbiota of both parents and new-borns through the innovative method of fecal microbiota transplantation (FMT) [104]. Precisely, FMT is a promising treatment for diseases related to gut dysbiosis, as it can help to rebalance the composition and function of the gut microbiota by transferring fecal preparations from healthy donors. Each gram of human feces contains ≈1011 bacterial cells, 108-109 virus-like particles (most are bacteriophages), ≈107 colonocytes, ≈108 archaea, ≈106 fungi, protists, and metabolites, [105]. The efficacy of the treatment can be explained by considering, for example, metabolites derived from the phylum Firmicutes, particularly the SCFAs and secondary bile acids, which with their beneficial roles such as fortifying the intestinal barrier and alleviating inflammation, promoting host homeostasis [106]. FMT can directly change the recipient’s GM for normalizing the composition and to provide a therapeutic benefit. FMT has initially been applied for the treatment of recurrent and refractory Clostridium difficile infection, thanks to the decisive consensus of United States Food and Drug Administration, occurred in 2013. Today, its application is not only limited to gastrointestinal disorders, but to other diseases [104]. Accordingly, a recent systematic review highlighted that FMT can be adopted for the treatment of 85 specific diseases in clinical settings globally from 2011 to 2021 [107]. Furthermore, a study conducted at the University of Minnesota, enrolling patients suffering from acute myeloid leukemia and patients undergoing hematopoietic cell transplantation, experimented with microbiota transplantation. The subjects enrolled were fragile and could contract a high number of infections, also linked to alterations in the microbiota (dysbiosis) following the treatment. Although microbiota transplantation does not have a significant effect against infections, it can still normalize the composition of the microbiota, obtaining therapeutic benefits by improving the diversity of intestinal microorganisms, increasing the levels of some anaerobic commensal bacteria, and reducing the concentration of other species that could be the cause of some disorders [108].
Although FMT appears to be a generally safe therapeutic method with few adverse effects, it is, however, necessary to monitor clinical efficacy and long-term adverse events [104]. Consequently, it is imperative to fix regular follow-up for both identifying the periodicity and length of FMT treatment and monitoring the clinical efficacy and long-term adverse events. In addition, further studies are needed for developing personalized FMT treatments for everyone and his clinical conditions according to diverse features of hosts and diseases/conditions to treat, such as adverse HS programming, and leukemia.
Another strategy might be the use of serotherapeutic therapy, that includes three therapeutic approaches: (i) the treatment with molecules able to selectively kill senescent cells (SCs), that is, senolytics; (ii) the use of compounds having the capacity to reduce the proinflammatory program of SCs, or that modify the senescent phenotype, that is, senomorphics; and (iii) prevention of the accumulation of senescent cells [109,110,111,112]. All these measures may consent a) to reduce the effects of developmental adversity, b) to favor a well-matched HS development programming principally acting on endothelium, and 3) to delay/retard the onset of leukemia in children and adulthood. Furthermore, the development of other optional treatments is increasing. These last have the aim to recover the disturbed epigenetic profiles linked to altered programming. However, the detrimental epigenetic alterations are understood to be potentially reversible; therefore, they may likely be corrected by certain lifestyle factors such as diet, physical activity, etc., as well as by pharmacological interventions specifically targeted at epigenome [13,14,15,16]. If these therapeutic strategies will be established, then such an approach would provide a way to slow down the epigenetic clock and to modify the dynamics of epigenetic age during the life course and, thus, to slowing down and/or delaying age-related processes [94,117,118]. The lack of specific biomarkers for monitoring developmental programming makes difficult to test and verify the biological effects of eventual interventions and treatments. The group of Goswami has suggested the telomere length as optimal biomarker for developmental programming [119]. The epigenetic age indices such as DNA methylation-based biomarkers are also considered now as another promising option [120]. These obstacles, as well as the need to identify the unknown longterm outcomes of the described interventions and therapeutic approaches, reflect several gaps and the need of performing further studies. Multidisciplinary investigations are particularly suggested, being everyone the result of the sophisticated interplay of environmental factors with its genome, trascriptome, proteome, metabolome, microbiome, epigenome, exposome, as evidenced in describing and discussing on HS programming. Further studies are hence imperative, in different types of patients and with different conditions and diseases [104].
Diet plays a key role from the earliest days of life in human health in cell metabolism, regulation of the GM and immunological processes via epigenomic factors, as above mentioned, [121]. Many studies have reported a correlation between habits such as smoking and drinking alcohol during pregnancy and increased risk of leukemia, but other dietary factors also have an important influence [122,123]. Intake of fruit and vegetables provides the necessary folic acid intake to avoid the risk of leukemia in children, and it has been reported that maternal fruit and vegetable consumption is inversely related to childhood ALL [124]. In California, in accordance with this premise, a study has been conducted for evaluating the link between maternal diet quality prior to pregnancy, considering a diet quality index, and the risk of childhood ALL. On the other hand, maternal malnutrition and low micronutrient levels could cause elevated maternal cortisol concentrations, affecting the development of the fetal immune system, and interfering with normal immune cell proliferation and organogenesis [125]. Studies have reported a correlation between diet and risk of ALL. It has been observed by some researchers that a diet that includes fish, seafood, beans, and beef correlates with a low risk of ALL [126]. Instead, the risk of ALL may increases when mothers consume various foods such as sugars or syrups [127]. Regarding the risk of ALL in children, there is a positive association between the risk of this disease with consumption of more coffee and/or caffeinated beverages [128,129].
Dietary habits can influence the diversity of the GM and dietary components can influence both the microbial population and its metabolic activity. The Mediterranean diet has been proposed and it includes foods such as vegetables mainly, fiber and omega-3 fatty acids and animal protein and saturated fats but in smaller quantities. Adherence to the Mediterranean diet leads to an increase in certain bacteria such as Bifidobacteria, Lactobacilli, Prevotella, Eubacteria, Bacteroides, in contrast, a high-fat diet leads to an increase in Bacteroides and Enterobacteria and a decrease in Bifidobacteria, Lactobacilli, Prevotella and Eubacteria. Different studies affirm this diet-dependent change in the microbiota [130].
In a study by De Filippis and colleagues, in 153 individuals habitually following omnivore, vegetarian or vegan diets, adherence to this diet has been observed and shown to be associated with increased levels of fiber-degrading SCFA (short chain fatty acids), Prevotella and Firmicutes [131].
In subjects eating the Mediterranean diet, the Prevotella-Bacteroides ratio was higher, indicating that a diet rich in natural fibre and resistant starch has a positive effect on the bacterial composition of human subjects [132] A study conducted by Garcia’s group has focused on the eating habits of healthy subjects to test the variability of the microbiota adhering to the Mediterranean diet. What is observed following a questionnaire to which the volunteer subjects are submitted is that adherence to the Mediterranean diet allows a decrease in the Firmicutes-Bacteroidetes ratio and greater presence of Bacteroidetes was associated with lower animal protein intake. High consumption of animal protein, saturated fats, and sugars affected gut microbiota diversity [133]. To confirm the observations of the above studies a preclinical study conducted on mice by Nagpal et al. analyzed the gut microbiome after adhering to either the typical Western diet or the Mediterranean diet [134] They found that the microbiome of the study participants consuming the typical Western diet was significantly more diverse than the microbiome of the participants consuming the typical Mediterranean diet. They found that the microbiota of study participants consuming the Mediterranean diet was significantly more diverse than the microbiota of participants consuming the Western diet, characterized by in lard, beef tallow, butter, eggs, cholesterol, casein, lactoalbumin, dextrin, high-fructose corn syrup and sucrose. They also had a higher abundance of Lactobacillus, Clostridium, Faecalibacterium and Oscillospira and a lower abundance of Ruminococcus and Coprococcus [134] Another study states that the diet can alter the composition of the microbiota very rapidly, in less than a week, as shown by 31 in his study in which the consumption of particular types of foods is attested to that produces predictable changes in existing host bacterial genera. This influences host immune and metabolic parameters, with broad implications for human health [130]. Microbes in the distal intestine, where they are abundant, contribute to host health through the biosynthesis of essential vitamins and amino acids and the generation of important metabolic byproducts from food components not digested by the small intestine [135].
Several studies have been able to comprehensively investigate the impact of diet component on gut microbial composition. The effects of dietary protein on the gut microbiota were first described in 1977. Protein consumption is positively correlated with overall microbial diversity [136]. The consumption of whey and pea protein extract results in the increase of Bifdobacterium and intestinal Lactobacillus, while whey also reduces the pathogens Bacteroides fragilis and Clostridium perfringens [137]. Pea protein also has been associated with increased levels of intestinal SCFAs, which are considered anti-inflammatory and important for maintaining the mucosal barrier [138]. In contrast, consumption of animal protein causes an increase in the number of bile-tolerant anaerobes, such as Bacteroides, Alistipes, and Bilophila [136]. One study found that subjects on a high-protein/low-carbohydrate diet carbohydrate had a reduced presence of Roseburia and Eubacterium rectale in the gut microbiota and lower butyrate in the feces [139]. In their study, De Filippo et al.similarly observed lower fecal SCFAs in Italian subjects who consumed a protein-rich diet [140].
Adherence to the Mediterranean diet has also been shown to have positive effects on health, with a reduction in inflammatory molecules and thus a protective role against oncological diseases.
Regarding the panel of inflammation-related markers, a study was conducted examining the effects of the Mediterranean diet on the inflammatory profile. 612 non-fragile or pre-fragile subjects in five European countries (UK, France, Netherlands, Italy and Poland) were analysed before and after a 12-month Mediterranean diet intervention. A negative correlation in the inflammatory markers CRP, IL-17 and IL-2 with positive levels of the anti-inflammatory cytokine IL-10 was observed following adherence to the diet. This confirms how diet and in particular adherence to the Mediterranean diet can positively influence health by reducing the risk of chronic inflammatory [141].
In addition to their effects on the composition of the microbiota, prebiotics also produce significant changes in metabolic and immune markers.
Several different studies have observed reductions in the proinflammatory cytokine IL-6, associated with the intake of non-digestible carbohydrates present in whole grains [142].
West et al. noted an increase in plasma levels of the anti-inflammatory cytokine IL-10 with consumption of high-amylose corn starch butyrate [143].
Therefore, prebiotics are thought to have a beneficial effect on the immune and metabolic function of the gut, and this is believed to be due to increased production of SCFAs and strengthening of gastrointestinal tract-associated lymphoid tissue (GALT) resulting from fibre fermentation [144].
New evidence has reported that specific nutrients exert different actions on metabolic outcomes, depending on individual microbial patterns subject to specific individuals or conditions, suggesting the important role of personalized human nutritional treatment [145].
Another recent therapy is based on the use of butyrate-producing bacteria, which as previously mentioned has a protective role against ALL disease. These strains, such as those of Clostridium butyricum and Butyricicoccus pullicaecorum are thought to be specific next-generation niche probiotics and have good bile tolerance, viability, and metabolism and can be genetically manipulated to increase their ability to produce butyrate [146,147]. For example, heterologous genes required for butyrate production from butyrate from acetyl-CoA can be introduced by inactivating the gene that codes for conversion of acetyl-CoA to acetate and the gene encoding aldehyde/alcohol dehydrogenase for ethanol production or simply disrupt a CoA transferase gene, which could be an alternative pathway for acetate production. In addition, to obtain higher levels of butyrate and thus greater abundance of butyrate producers in the gut, co-culture can be carried out to obtain an interactive microbial population composed of more than microbes.
6. Conclusions
Although studies regarding GM are growing in number, the correlation between pediatric cancers, such as ALL, and GM is not yet well recognized. It is complicated to study due to the relatively small number of patients (cohorts) and difficulties in sample collection. Additionally, children with ALL receive chemotherapy and radiation therapy, as well as intensive antibiotic prophylaxis, to inhibit potential infections. These treatments have a great effect on the entire body, including the bone marrow, liver, and gastrointestinal tract, and directly and indirectly affect the GM. Furthermore, no prospective study has been designed for ALL to date.
The relevance of the GM is confirmed by several studies regarding disease development, the efficacy of therapy, staging, or side effect manifestations. It requires more investigation, but in the future, it might be feasible to identify the microbiome profile of an individual before starting antitumor therapy to predict efficacy or to choose appropriate and personalized therapy. GM could also be used as a biomarker. It remains unanswered whether dysbiosis is a consequence or cause of neoplasms. Microbiome studies in the pediatric oncology population are limited, and associations are still unclear. More studies with larger cohorts are needed to be done to help develop more personalized and successful therapy in pediatric oncology. Finally, additional studies are imperative to test the long-term effects of the therapeutical strategies suggested to be applied at two levels of intervention (see Figure 2A,B).
Author Contributions
CRB was involved in design, wrote, and edited the paper. MC was involved in the figures and data literature curation. CRB, MC and TSA revised the manuscript. All authors participated in the study, and they read and approved the final version of the paper.
References
- Greaves M. (2018). A causal mechanism for childhood acute lymphoblastic leukaemia. Nature reviews. Cancer, 18(8), 471–484. [CrossRef]
- Rajagopala, S. V., Yooseph, S., Harkins, D. M., Moncera, K. J., Zabokrtsky, K. B., Torralba, M. G., Tovchigrechko, A., Highlander, S. K., Pieper, R., Sender, L., & Nelson, K. E. (2016). Gastrointestinal microbial populations can distinguish pediatric and adolescent Acute Lymphoblastic Leukemia (ALL) at the time of disease diagnosis. BMC genomics, 17(1), 635. [CrossRef]
- Ekpa, Q. L., Akahara, P. C., Anderson, A. M., Adekoya, O. O., Ajayi, O. O., Alabi, P. O., Okobi, O. E., Jaiyeola, O., & Ekanem, M. S. (2023). A Review of Acute Lymphocytic Leukemia (ALL) in the Pediatric Population: Evaluating Current Trends and Changes in Guidelines in the Past Decade. Cureus, 15(12), e49930. [CrossRef]
- Seth, R., & Singh, A. (2015). Leukemias in Children. Indian journal of pediatrics, 82(9), 817–824. [CrossRef]
- Tarlock, K.; Dahl, G.; Lacayo, N. Acute myeloid leukemia in children. In Wintrobe’s Clinical Hematology, 14th ed.; Lippincott Williams &Wilkins: Philadelphia, PA, USA, 2018; pp. 5016–5095.
- de Ville de Goyet, M., Kicinski, M., Suciu, S., Vandecruys, E., Uyttebroeck, A., Ferster, A., Freycon, C., Plat, G., Thomas, C., Barbati, M., Dresse, M. F., Paillard, C., Pluchart, C., Simon, P., Chantrain, C., Minckes, O., van der Werff Ten Bosch, J., Bertrand, Y., Rohrlich, P., Millot, F., European Organisation for Research, Treatment of Cancer (EORTC) Children’s Leukemia Group (CLG) (2024). Long-term neurotoxicity among childhood acute lymphoblastic leukaemia survivors enrolled between 1971 and 1998 in EORTC Children Leukemia Group studies. Discover. Oncology, 15(1), 20. [CrossRef]
- Del Principe, M. I., Maurillo, L., Buccisano, F., Sconocchia, G., Cefalo, M., De Santis, G., Di Veroli, A., Ditto, C., Nasso, D., Postorino, M., Refrigeri, M., Attrotto, C., Del Poeta, G., Lo-Coco, F., Amadori, S., & Venditti, A. (2014). Central nervous system involvement in adult acute lymphoblastic leukemia: diagnostic tools, prophylaxis, and therapy. Mediterranean journal of hematology and infectious diseases, 6(1), e2014075. [CrossRef]
- Johnston, D. L., Alonzo, T. A., Gerbing, R. B., Aplenc, R., Woods, W. G., Meshinchi, S., & Gamis, A. S. (2017). Central nervous system disease in pediatric acute myeloid leukemia: A report from the Children’s Oncology Group. Pediatric blood & cancer, 64(12), 10.1002/pbc.26612. [CrossRef]
- Van Delft, F. W., & Saha, V. (2004). Molecular techniques to improve outcome in childhood ALL. Methods in molecular medicine, 91, 111–122. [CrossRef]
- Cho, S. Y., Cheng, A. C., & Cheng, M. C. (2000). Oral care for children with leukaemia. Hong Kong medical journal = Xianggang yi xue za zhi, 6(2), 203–208.
- Fathi, A., Mirzarahimi, M., & Farajkhah, H. (2021). Réponse à un schéma chimiothérapeutique administré à des enfants atteints de LAL à cellules pré-B à risque élevé selon le protocole COG. Canadian oncology nursing journal = Revue canadienne de nursing oncologique, 31(3), 334–338. [CrossRef]
- Lim, H. C., & Kim, C. S. (2014). Oral signs of acute leukemia for early detection. Journal of periodontal & implant science, 44(6), 293–299. [CrossRef]
- Balistreri, C. R., Garagnani, P., Madonna, R., Vaiserman, A., & Melino, G. (2019). Developmental programming of adult haematopoiesis system. Ageing research reviews, 54, 100918. [CrossRef]
- Tebbi C. K. (2021). Etiology of Acute Leukemia: A Review. Cancers, 13(9), 2256. [CrossRef]
- Pagani, I. S., Poudel, G., & Wardill, H. R. (2022). A Gut Instinct on Leukaemia: A New Mechanistic Hypothesis for Microbiota-Immune Crosstalk in Disease Progression and Relapse. Microorganisms, 10(4), 713. [CrossRef]
- Oldenburg, M., Rüchel, N., Janssen, S., Borkhardt, A., & Gössling, K. L. (2021). The Microbiome in Childhood Acute Lymphoblastic Leukemia. Cancers, 13(19), 4947. [CrossRef]
- Soares, S. C., Roux, L. J. D., Castro, A. R., Silva, C. C., Rodrigues, R., Macho, V. M. P., Silva, F., & Costa, C. (2023). Oral Manifestations: A Warning-Sign in Children with Hematological Disease Acute Lymphocytic Leukemia. Hematology reports, 15(3), 491–502. [CrossRef]
- Balistreri, C. R., & Monastero, R. (2023). Neuroinflammation and Neurodegenerative Diseases: How Much Do We Still Not Know?. Brain sciences, 14(1), 19. [CrossRef]
- Schirò, G., Iacono, S., & Balistreri, C. R. (2023). The Role of Human Microbiota in Myasthenia Gravis: A Narrative Review. Neurology international, 15(1), 392–404. [CrossRef]
- Scola, L., Giarratana, R. M., Torre, S., Argano, V., Lio, D., & Balistreri, C. R. (2019). On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches. International journal of molecular sciences, 20(23), 6015. [CrossRef]
- Forte, A., Balistreri, C. R., De Feo, M., Della Corte, A., Hellstrand, P., Persson, L., & Nilsson, B. O. (2019). Polyamines and microbiota in bicuspid and tricuspid aortic valve aortopathy. Journal of molecular and cellular cardiology, 129, 179–187. [CrossRef]
- Maleki, S., Razavi, S. H., Yadav, H., & Letizia Manca, M. (2024). New horizon to the world of gut microbiome: seeds germination. Critical reviews in food science and nutrition, 1–19. Advance online publication. [CrossRef]
- Balistreri C. R. (2022). Promising Strategies for Preserving Adult Endothelium Health and Reversing Its Dysfunction: From Liquid Biopsy to New Omics Technologies and Noninvasive Circulating Biomarkers. International journal of molecular sciences, 23(14), 7548. [CrossRef]
- Zheng, D., Liwinski, T., & Elinav, E. (2020). Interaction between microbiota and immunity in health and disease. Cell research, 30(6), 492–506. [CrossRef]
- Wen, Y., Jin, R., & Chen, H. (2019). Interactions Between Gut Microbiota and Acute Childhood Leukemia. Frontiers in microbiology, 10, 1300. [CrossRef]
- Schwabe, R. F., & Jobin, C. (2013). The microbiome and cancer. Nature reviews. Cancer, 13(11), 800–812. [CrossRef]
- Holler, E., Butzhammer, P., Schmid, K., Hundsrucker, C., Koestler, J., Peter, K., Zhu, W., Sporrer, D., Hehlgans, T., Kreutz, M., Holler, B., Wolff, D., Edinger, M., Andreesen, R., Levine, J. E., Ferrara, J. L., Gessner, A., Spang, R., & Oefner, P. J. (2014). Metagenomic analysis of the stool microbiome in patients receiving allogeneic stem cell transplantation: loss of diversity is associated with use of systemic antibiotics and more pronounced in gastrointestinal graft-versus-host disease. Biology of blood and marrow transplantation : journal of the American Society for Blood and Marrow Transplantation, 20(5), 640–645. [CrossRef]
- Rajagopala, S. V., Yooseph, S., Harkins, D. M., Moncera, K. J., Zabokrtsky, K. B., Torralba, M. G., Tovchigrechko, A., Highlander, S. K., Pieper, R., Sender, L., & Nelson, K. E. (2016). Gastrointestinal microbial populations can distinguish pediatric and adolescent Acute Lymphoblastic Leukemia (ALL) at the time of disease diagnosis. BMC genomics, 17(1), 635. [CrossRef]
- Scola, L., Giarratana, R. M., Torre, S., Argano, V., Lio, D., & Balistreri, C. R. (2019). On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches. International journal of molecular sciences, 20(23), 6015. [CrossRef]
- Balistreri C. R. (2022). Promising Strategies for Preserving Adult Endothelium Health and Reversing Its Dysfunction: From Liquid Biopsy to New Omics Technologies and Noninvasive Circulating Biomarkers. International journal of molecular sciences, 23(14), 7548. [CrossRef]
- Greaves, M., Cazzaniga, V., & Ford, A. (2021). Can we prevent childhood Leukaemia?. Leukemia, 35(5), 1258–1264. [CrossRef]
- Nycz, B. T., Dominguez, S. R., Friedman, D., Hilden, J. M., Ir, D., Robertson, C. E., & Frank, D. N. (2018). Evaluation of bloodstream infections, Clostridium difficile infections, and gut microbiota in pediatric oncology patients. PloS one, 13(1), e0191232. [CrossRef]
- Poore, G. D., Kopylova, E., Zhu, Q., Carpenter, C., Fraraccio, S., Wandro, S., Kosciolek, T., Janssen, S., Metcalf, J., Song, S. J., Kanbar, J., Miller-Montgomery, S., Heaton, R., Mckay, R., Patel, S. P., Swafford, A. D., & Knight, R. (2020). Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature, 579(7800), 567–574. [CrossRef]
- Turpin, W., Espin-Garcia, O., Xu, W., Silverberg, M. S., Kevans, D., Smith, M. I., Guttman, D. S., Griffiths, A., Panaccione, R., Otley, A., Xu, L., Shestopaloff, K., Moreno-Hagelsieb, G., GEM Project Research Consortium, Paterson, A. D., & Croitoru, K. (2016). Association of host genome with intestinal microbial composition in a large healthy cohort. Nature genetics, 48(11), 1413–1417. [CrossRef]
- Beam, A., Clinger, E., & Hao, L. (2021). Effect of Diet and Dietary Components on the Composition of the Gut Microbiota. Nutrients, 13(8), 2795. [CrossRef]
- Ma, T., Chen, Y., Li, L. J., & Zhang, L. S. (2021). Opportunities and Challenges for Gut Microbiota in Acute Leukemia. Frontiers in oncology, 11, 692951. [CrossRef]
- https://www.poliambulatoriomodoetia.it/salute/microbiota-intestinale/.
- Adak, A., & Khan, M. R. (2019). An insight into gut microbiota and its functionalities. Cellular and molecular life sciences : CMLS, 76(3), 473–493. [CrossRef]
- https://www.santagostino.it/it/santagostinopedia/microbiota-intestinale.
- Upadhyay Banskota, S., Skupa, S. A., El-Gamal, D., & D’Angelo, C. R. (2023). Defining the Role of the Gut Microbiome in the Pathogenesis and Treatment of Lymphoid Malignancies. International journal of molecular sciences, 24(3), 2309. [CrossRef]
- Wu, S., Rhee, K. J., Albesiano, E., Rabizadeh, S., Wu, X., Yen, H. R., Huso, D. L., Brancati, F. L., Wick, E., McAllister, F., Housseau, F., Pardoll, D. M., & Sears, C. L. (2009). A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nature medicine, 15(9), 1016–1022. [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215.
- Balistreri, C. R., Garagnani, P., Madonna, R., Vaiserman, A., & Melino, G. (2019). Developmental programming of adult haematopoiesis system. Ageing research reviews, 54, 100918. [CrossRef]
- Groussin, M., Poyet, M., Sistiaga, A., Kearney, S. M., Moniz, K., Noel, M., Hooker, J., Gibbons, S. M., Segurel, L., Froment, A., Mohamed, R. S., Fezeu, A., Juimo, V. A., Lafosse, S., Tabe, F. E., Girard, C., Iqaluk, D., Nguyen, L. T. T., Shapiro, B. J., Lehtimäki, J., … Alm, E. J. (2021). Elevated rates of horizontal gene transfer in the industrialized human microbiome. Cell, 184(8), 2053–2067.e18. [CrossRef]
- Alexander, J. L., Wilson, I. D., Teare, J., Marchesi, J. R., Nicholson, J. K., and Kinross, J. M. (2017). Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 14, 356–365. doi:10.1038/nrgastro.2017.20.
- Balmer, M. L., Schürch, C. M., Saito, Y., Geuking, M. B., Li, H., Cuenca, M., Kovtonyuk, L. V., McCoy, K. D., Hapfelmeier, S., Ochsenbein, A. F., Manz, M. G., Slack, E., & Macpherson, A. J. (2014). Microbiota-derived compounds drive steady-state granulopoiesis via MyD88/TICAM signaling. Journal of immunology (Baltimore, Md. : 1950), 193(10), 5273–5283. [CrossRef]
- Taur, Y., Jenq, R. R., Perales, M. A., Littmann, E. R., Morjaria, S., Ling, L., No, D., Gobourne, A., Viale, A., Dahi, P. B., Ponce, D. M., Barker, J. N., Giralt, S., van den Brink, M., & Pamer, E. G. (2014). The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood, 124(7), 1174–1182. [CrossRef]
- Espinoza, J. L., Elbadry, M. I., & Nakao, S. (2016). An altered gut microbiota may trigger autoimmune-mediated acquired bone marrow failure syndromes. Clinical immunology (Orlando, Fla.), 171, 62–64. [CrossRef]
- Trompette, A., Gollwitzer, E. S., Yadava, K., Sichelstiel, A. K., Sprenger, N., Ngom-Bru, C., Blanchard, C., Junt, T., Nicod, L. P., Harris, N. L., & Marsland, B. J. (2014). Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nature medicine, 20(2), 159–166. [CrossRef]
- Zapata, H. J., & Quagliarello, V. J. (2015). The microbiota and microbiome in aging: potential implications in health and age-related diseases. Journal of the American Geriatrics Society, 63(4), 776–781. [CrossRef]
- Vaiserman, A., Koliada, A., & Lushchak, O. (2018). Developmental programming of aging trajectory. Ageing research reviews, 47, 105–122. [CrossRef]
- Kozyrskyj, A. L., Kalu, R., Koleva, P. T., & Bridgman, S. L. (2016). Fetal programming of overweight through the microbiome: boys are disproportionately affected. Journal of developmental origins of health and disease, 7(1), 25–34. [CrossRef]
- Weissman I. L. (1994). Developmental switches in the immune system. Cell, 76(2), 207–218. [CrossRef]
- Khosravi, A., Yáñez, A., Price, J. G., Chow, A., Merad, M., Goodridge, H. S., & Mazmanian, S. K. (2014). Gut microbiota promote hematopoiesis to control bacterial infection. Cell host & microbe, 15(3), 374–381. [CrossRef]
- Smith, S. E., Li, J., Garbett, K., Mirnics, K., & Patterson, P. H. (2007). Maternal immune activation alters fetal brain development through interleukin-6. The Journal of neuroscience : the official journal of the Society for Neuroscience, 27(40), 10695–10702. [CrossRef]
- Singh, N., Thangaraju, M., Prasad, P. D., Martin, P. M., Lambert, N. A., Boettger, T., Offermanns, S., & Ganapathy, V. (2010). Blockade of dendritic cell development by bacterial fermentation products butyrate and propionate through a transporter (Slc5a8)-dependent inhibition of histone deacetylases. The Journal of biological chemistry, 285(36), 27601–27608. [CrossRef]
- Kurita-Ochiai, T., Fukushima, K., & Ochiai, K. (1995). Volatile fatty acids, metabolic by-products of periodontopathic bacteria, inhibit lymphocyte proliferation and cytokine production. Journal of dental research, 74(7), 1367–1373. [CrossRef]
- Zimmerman, M. A., Singh, N., Martin, P. M., Thangaraju, M., Ganapathy, V., Waller, J. L., Shi, H., Robertson, K. D., Munn, D. H., & Liu, K. (2012). Butyrate suppresses colonic inflammation through HDAC1-dependent Fas upregulation and Fas-mediated apoptosis of T cells. American journal of physiology. Gastrointestinal and liver physiology, 302(12), G1405–G1415. [CrossRef]
- Josefsdottir, K. S., Baldridge, M. T., Kadmon, C. S., & King, K. Y. (2017). Antibiotics impair murine hematopoiesis by depleting the intestinal microbiota. Blood, 129(6), 729–739. [CrossRef]
- Baldridge, M. T., Nice, T. J., McCune, B. T., Yokoyama, C. C., Kambal, A., Wheadon, M., Diamond, M. S., Ivanova, Y., Artyomov, M., & Virgin, H. W. (2015). Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science (New York, N.Y.), 347(6219), 266–269. [CrossRef]
- Chow, J., Lee, S. M., Shen, Y., Khosravi, A., & Mazmanian, S. K. (2010). Host-bacterial symbiosis in health and disease. Advances in immunology, 107, 243–274. [CrossRef]
- Biagi, E., Candela, M., Franceschi, C., & Brigidi, P. (2011). The aging gut microbiota: new perspectives. Ageing research reviews, 10(4), 428–429. [CrossRef]
- Franceschi, C., Bonafè, M., Valensin, S., Olivieri, F., De Luca, M., Ottaviani, E., & De Benedictis, G. (2000). Inflamm-aging. An evolutionary perspective on immunosenescence. Annals of the New York Academy of Sciences, 908, 244–254. [CrossRef]
- Rampelli, S., Candela, M., Turroni, S., Biagi, E., Collino, S., Franceschi, C., O’Toole, P. W., & Brigidi, P. (2013). Functional metagenomic profiling of intestinal microbiome in extreme ageing. Aging, 5(12), 902–912. [CrossRef]
- Biagi, E., Franceschi, C., Rampelli, S., Severgnini, M., Ostan, R., Turroni, S., Consolandi, C., Quercia, S., Scurti, M., Monti, D., Capri, M., Brigidi, P., & Candela, M. (2016). Gut Microbiota and Extreme Longevity. Current biology : CB, 26(11), 1480–1485. [CrossRef]
- Grignolio, A., Mishto, M., Faria, A. M., Garagnani, P., Franceschi, C., & Tieri, P. (2014). Towards a liquid self: how time, geography, and life experiences reshape the biological identity. Frontiers in immunology, 5, 153. [CrossRef]
- Fisher, R. E., Steele, M., & Karrow, N. A. (2012). Fetal programming of the neuroendocrine-immune system and metabolic disease. Journal of pregnancy, 2012, 792934. [CrossRef]
- Bateson, P., Gluckman, P., & Hanson, M. (2014). The biology of developmental plasticity and the Predictive Adaptive Response hypothesis. The Journal of physiology, 592(11), 2357–2368. [CrossRef]
- Faa, G., Marcialis, M. A., Ravarino, A., Piras, M., Pintus, M. C., & Fanos, V. (2014). Fetal programming of the human brain: is there a link with insurgence of neurodegenerative disorders in adulthood?. Current medicinal chemistry, 21(33), 3854–3876. [CrossRef]
- Alexander, B. T., Dasinger, J. H., & Intapad, S. (2015). Fetal programming and cardiovascular pathology. Comprehensive Physiology, 5(2), 997–1025. [CrossRef]
- Bateson P. (2015). Why are individuals so different from each other?. Heredity, 115(4), 285–292. [CrossRef]
- Projecto-Garcia, J., Biddle, J. F., & Ragsdale, E. J. (2017). Decoding the architecture and origins of mechanisms for developmental polyphenism. Current opinion in genetics & development, 47, 1–8. [CrossRef]
- Kwon, E. J., & Kim, Y. J. (2017). What is fetal programming?: a lifetime health is under the control of in utero health. Obstetrics & gynecology science, 60(6), 506–519. [CrossRef]
- Miranda, J. O., Ramalho, C., Henriques-Coelho, T., & Areias, J. C. (2017). Fetal programming as a predictor of adult health or disease: the need to reevaluate fetal heart function. Heart failure reviews, 22(6), 861–877. [CrossRef]
- McGowan, P. O., & Matthews, S. G. (2018). Prenatal Stress, Glucocorticoids, and Developmental Programming of the Stress Response. Endocrinology, 159(1), 69–82. [CrossRef]
- Walker, D. J., & Spencer, K. A. (2018). Glucocorticoid programming of neuroimmune function. General and comparative endocrinology, 256, 80–88. [CrossRef]
- Wells J. C. (2017). Worldwide variability in growth and its association with health: Incorporating body composition, developmental plasticity, and intergenerational effects. American journal of human biology : the official journal of the Human Biology Council, 29(2), 10.1002/ajhb.22954. [CrossRef]
- Zohdi, V., Lim, K., Pearson, J. T., & Black, M. J. (2014). Developmental programming of cardiovascular disease following intrauterine growth restriction: findings utilising a rat model of maternal protein restriction. Nutrients, 7(1), 119–152. [CrossRef]
- Wang, R., Wiemels, J. L., Metayer, C., Morimoto, L., Francis, S. S., Kadan-Lottick, N., DeWan, A. T., Zhang, Y., & Ma, X. (2017). Cesarean Section and Risk of Childhood Acute Lymphoblastic Leukemia in a Population-Based, Record-Linkage Study in California. American journal of epidemiology, 185(2), 96–105. [CrossRef]
- Greenbaum, S., Sheiner, E., Wainstock, T., Segal, I., Ben-Harush, M., Sergienko, R., & Walfisch, A. (2018). Cesarean Delivery and Childhood Malignancies: A Single-Center, Population-Based Cohort Study. The Journal of pediatrics, 197, 292–296.e3. [CrossRef]
- Amitay, E. L., & Keinan-Boker, L. (2015). Breastfeeding and Childhood Leukemia Incidence: A Meta-analysis and Systematic Review. JAMA pediatrics, 169(6), e151025. [CrossRef]
- Penders, J., Thijs, C., Vink, C., Stelma, F. F., Snijders, B., Kummeling, I., van den Brandt, P. A., & Stobberingh, E. E. (2006). Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics, 118(2), 511–521. [CrossRef]
- Wang, Y., Xue, J., Zhou, X., You, M., Du, Q., Yang, X., He, J., Zou, J., Cheng, L., Li, M., Li, Y., Zhu, Y., Li, J., Shi, W., & Xu, X. (2014). Oral microbiota distinguishes acute lymphoblastic leukemia pediatric hosts from healthy populations. PloS one, 9(7), e102116. [CrossRef]
- Liu, X., Zou, Y., Ruan, M., Chang, L., Chen, X., Wang, S., Yang, W., Zhang, L., Guo, Y., Chen, Y., Zhang, Y., He, H., Gan, Y., Wang, K., & Zhu, X. (2020). Pediatric Acute Lymphoblastic Leukemia Patients Exhibit Distinctive Alterations in the Gut Microbiota. Frontiers in cellular and infection microbiology, 10, 558799. [CrossRef]
- Bai, L., Zhou, P., Li, D., & Ju, X. (2017). Changes in the gastrointestinal microbiota of children with acute lymphoblastic leukaemia and its association with antibiotics in the short term. Journal of medical microbiology, 66(9), 1297–1307. [CrossRef]
- Vacca, M., Celano, G., Calabrese, F. M., Portincasa, P., Gobbetti, M., & De Angelis, M. (2020). The Controversial Role of Human Gut Lachnospiraceae. Microorganisms, 8(4), 573. [CrossRef]
- Milani, C., Duranti, S., Bottacini, F., Casey, E., Turroni, F., Mahony, J., Belzer, C., Delgado Palacio, S., Arboleya Montes, S., Mancabelli, L., Lugli, G. A., Rodriguez, J. M., Bode, L., de Vos, W., Gueimonde, M., Margolles, A., van Sinderen, D., & Ventura, M. (2017). The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiology and molecular biology reviews : MMBR, 81(4), e00036-17. [CrossRef]
- Hakim, H., Dallas, R., Wolf, J., Tang, L., Schultz-Cherry, S., Darling, V., Johnson, C., Karlsson, E. A., Chang, T. C., Jeha, S., Pui, C. H., Sun, Y., Pounds, S., Hayden, R. T., Tuomanen, E., & Rosch, J. W. (2018). Gut Microbiome Composition Predicts Infection Risk During Chemotherapy in Children With Acute Lymphoblastic Leukemia. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America, 67(4), 541–548. [CrossRef]
- Chua, L. L., Rajasuriar, R., Azanan, M. S., Abdullah, N. K., Tang, M. S., Lee, S. C., Woo, Y. L., Lim, Y. A., Ariffin, H., & Loke, P. (2017). Reduced microbial diversity in adult survivors of childhood acute lymphoblastic leukemia and microbial associations with increased immune activation. Microbiome, 5(1), 35. [CrossRef]
- Henrick, B. M., Chew, S., Casaburi, G., Brown, H. K., Frese, S. A., Zhou, Y., Underwood, M. A., & Smilowitz, J. T. (2019). Colonization by B. infantis EVC001 modulates enteric inflammation in exclusively breastfed infants. Pediatric research, 86(6), 749–757. [CrossRef]
- Thomas, R., Wong, W. S. W., Saadon, R., Vilboux, T., Deeken, J., Niederhuber, J., Hourigan, S. K., & Yang, E. (2020). Gut microbial composition difference between pediatric ALL survivors and siblings. Pediatric hematology and oncology, 37(6), 475–488. [CrossRef]
- Hudson, M. M., Ness, K. K., Gurney, J. G., Mulrooney, D. A., Chemaitilly, W., Krull, K. R., Green, D. M., Armstrong, G. T., Nottage, K. A., Jones, K. E., Sklar, C. A., Srivastava, D. K., & Robison, L. L. (2013). Clinical ascertainment of health outcomes among adults treated for childhood cancer. JAMA, 309(22), 2371–2381. [CrossRef]
- Armstrong, G. T., Kawashima, T., Leisenring, W., Stratton, K., Stovall, M., Hudson, M. M., Sklar, C. A., Robison, L. L., & Oeffinger, K. C. (2014). Aging and risk of severe, disabling, life-threatening, and fatal events in the childhood cancer survivor study. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 32(12), 1218–1227. [CrossRef]
- Vaiserman A. (2018). Developmental Tuning of Epigenetic Clock. Frontiers in genetics, 9, 584. [CrossRef]
- Pinho, S., & Frenette, P. S. (2019). Haematopoietic stem cell activity and interactions with the niche. Nature reviews. Molecular cell biology, 20(5), 303–320. [CrossRef]
- Balistreri, C.R. (Ed.), 2016. Endothelial Progenitor Cells (EPCs) in Ageing and Age-Related Diseases: from Their Physiological and Pathological Implications toTranslation in Personalized Medicine [Special Issue]. Mechanisms Ageing Devolopment, pp. 1–80 159.
- Balistrieri, C.R., 2017a. Endothelial Progenitor Cells: A New Real Hope or Only an Unrealizable Dream? Springer International Publishing, Dordrecht, pp. 1–80.
- Balistreri, C.R., 2017b. The current obsession of age-related diseases and the race in identifying effective treatments and preventive interventions. J. Pathobiol. Physiol. 1, 2.
- Hillier, S. E., & Olander, E. K. (2017). Women’s dietary changes before and during pregnancy: A systematic review. Midwifery, 49, 19–31. [CrossRef]
- Biagi, C., Nunzio, M. D., Bordoni, A., Gori, D., & Lanari, M. (2019). Effect of Adherence to Mediterranean Diet during Pregnancy on Children’s Health: A Systematic Review. Nutrients, 11(5), 997. [CrossRef]
- Thompson, E. L., Vamos, C. A., & Daley, E. M. (2017). Physical activity during pregnancy and the role of theory in promoting positive behavior change: A systematic review. Journal of sport and health science, 6(2), 198–206. [CrossRef]
- Ng, Q. X., Venkatanarayanan, N., Loke, W., Yeo, W. S., Lim, D. Y., Chan, H. W., & Sim, W. S. (2019). A meta-analysis of the effectiveness of yoga-based interventions for maternal depression during pregnancy. Complementary therapies in clinical practice, 34, 8–12. [CrossRef]
- Hill, B., Kothe, E. J., Currie, S., Danby, M., Lang, A. Y., Bailey, C., Moran, L. J., Teede, H., North, M., Bruce, L. J., & Skouteris, H. (2019). A systematic mapping review of the associations between pregnancy intentions and health-related lifestyle behaviours or psychological wellbeing. Preventive medicine reports, 14, 100869. [CrossRef]
- Wang, J. W., Kuo, C. H., Kuo, F. C., Wang, Y. K., Hsu, W. H., Yu, F. J., Hu, H. M., Hsu, P. I., Wang, J. Y., & Wu, D. C. (2019). Fecal microbiota transplantation: Review and update. Journal of the Formosan Medical Association = Taiwan yi zhi, 118 Suppl 1, S23–S31. [CrossRef]
- De Paepe, M., Leclerc, M., Tinsley, C. R., & Petit, M. A. (2014). Bacteriophages: an underestimated role in human and animal health?. Frontiers in cellular and infection microbiology, 4, 39. [CrossRef]
- Morrison, D. J., & Preston, T. (2016). Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut microbes, 7(3), 189–200. [CrossRef]
- Yu, Y., Wang, W., & Zhang, F. (2023). The Next Generation Fecal Microbiota Transplantation: To Transplant Bacteria or Virome. Advanced science (Weinheim, Baden-Wurttemberg, Germany), 10(35), e2301097. [CrossRef]
- Rashidi, A., Ebadi, M., Rehman, T. U., Elhusseini, H., Kazadi, D., Halaweish, H., Khan, M. H., Hoeschen, A., Cao, Q., Luo, X., Kabage, A. J., Lopez, S., Holtan, S. G., Weisdorf, D. J., Khoruts, A., & Staley, C. (2023). Randomized Double-Blind Phase II Trial of Fecal Microbiota Transplantation Versus Placebo in Allogeneic Hematopoietic Cell Transplantation and AML. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 41(34), 5306–5319. [CrossRef]
- Baumann K. (2018). Rejuvenating senolytics. Nature reviews. Molecular cell biology, 19(9), 543. [CrossRef]
- Olivieri, F., Prattichizzo, F., Grillari, J., & Balistreri, C. R. (2018). Cellular Senescence and Inflammaging in Age-Related Diseases. Mediators of inflammation, 2018, 9076485. [CrossRef]
- Gurău, F., Baldoni, S., Prattichizzo, F., Espinosa, E., Amenta, F., Procopio, A. D., Albertini, M. C., Bonafè, M., & Olivieri, F. (2018). Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing research reviews, 46, 14–31. [CrossRef]
- Xu, M., Pirtskhalava, T., Farr, J. N., Weigand, B. M., Palmer, A. K., Weivoda, M. M., Inman, C. L., Ogrodnik, M. B., Hachfeld, C. M., Fraser, D. G., Onken, J. L., Johnson, K. O., Verzosa, G. C., Langhi, L. G. P., Weigl, M., Giorgadze, N., LeBrasseur, N. K., Miller, J. D., Jurk, D., Singh, R. J., … Kirkland, J. L. (2018). Senolytics improve physical function and increase lifespan in old age. Nature medicine, 24(8), 1246–1256. [CrossRef]
- Abdul, Q. A., Yu, B. P., Chung, H. Y., Jung, H. A., & Choi, J. S. (2017). Epigenetic modifications of gene expression by lifestyle and environment. Archives of pharmacal research, 40(11), 1219–1237. [CrossRef]
- Wallace, R. G., Twomey, L. C., Custaud, M. A., Turner, J. D., Moyna, N., Cummins, P. M., & Murphy, R. P. (2018). The role of epigenetics in cardiovascular health and ageing: A focus on physical activity and nutrition. Mechanisms of ageing and development, 174, 76–85. [CrossRef]
- Ferioli, M., Zauli, G., Maiorano, P., Milani, D., Mirandola, P., & Neri, L. M. (2019). Role of physical exercise in the regulation of epigenetic mechanisms in inflammation, cancer, neurodegenerative diseases, and aging process. Journal of cellular physiology, 234(9), 14852–14864. [CrossRef]
- Pasyukova, E. G., & Vaiserman, A. M. (2017). HDAC inhibitors: A new promising drug class in anti-aging research. Mechanisms of ageing and development, 166, 6–15. [CrossRef]
- Fransquet, P. D., Wrigglesworth, J., Woods, R. L., Ernst, M. E., & Ryan, J. (2019). The epigenetic clock as a predictor of disease and mortality risk: a systematic review and meta-analysis. Clinical epigenetics, 11(1), 62. [CrossRef]
- Xiao, F. H., Wang, H. T., & Kong, Q. P. (2019). Dynamic DNA Methylation During Aging: A “Prophet” of Age-Related Outcomes. Frontiers in genetics, 10, 107. [CrossRef]
- Whiteman, V. E., Goswami, A., & Salihu, H. M. (2017). Telomere length and fetal programming: A review of recent scientific advances. American journal of reproductive immunology (New York, N.Y. : 1989), 77(5), 10.1111/aji.12661. [CrossRef]
- Horvath, S., & Raj, K. (2018). DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nature reviews. Genetics, 19(6), 371–384. [CrossRef]
- Janakiraman, M., Salei, N., & Krishnamoorthy, G. (2023). High salt diet does not impact the development of acute myeloid leukemia in mice. Cancer immunology, immunotherapy : CII, 72(1), 265–273. [CrossRef]
- Karalexi, M. A., Dessypris, N., Thomopoulos, T. P., Ntouvelis, E., Kantzanou, M., Diamantaras, A. A., Moschovi, M., Baka, M., Hatzipantelis, E., Kourti, M., Polychronopoulou, S., Stiakaki, E., Mora, A. M., Wunsch-Filho, V., Infante-Rivard, C., Loutradis, D., & Petridou, E. T. (2017). Parental alcohol consumption and risk of leukemia in the offspring: a systematic review and meta-analysis. European journal of cancer prevention : the official journal of the European Cancer Prevention Organisation (ECP), 26(5), 433–441. [CrossRef]
- Fenech M. (2001). The role of folic acid and Vitamin B12 in genomic stability of human cells. Mutation research, 475(1-2), 57–67. [CrossRef]
- Kwan, M. L., Jensen, C. D., Block, G., Hudes, M. L., Chu, L. W., & Buffler, P. A. (2009). Maternal diet and risk of childhood acute lymphoblastic leukemia. Public health reports (Washington, D.C. : 1974), 124(4), 503–514. [CrossRef]
- Palmer A. C. (2011). Nutritionally mediated programming of the developing immune system. Advances in nutrition (Bethesda, Md.), 2(5), 377–395. [CrossRef]
- Devine, S. M., & Larson, R. A. (1994). Acute leukemia in adults: recent developments in diagnosis and treatment. CA: a cancer journal for clinicians, 44(6), 326–352. [CrossRef]
- Bonaventure, A., Rudant, J., Goujon-Bellec, S., Orsi, L., Leverger, G., Baruchel, A., Bertrand, Y., Nelken, B., Pasquet, M., Michel, G., Sirvent, N., Bordigoni, P., Ducassou, S., Rialland, X., Zelenika, D., Hémon, D., & Clavel, J. (2013). Childhood acute leukemia, maternal beverage intake during pregnancy, and metabolic polymorphisms. Cancer causes & control : CCC, 24(4), 783–793. [CrossRef]
- Blanco-Lopez, J., Iguacel, I., Pisanu, S., Almeida, C. C. B., Steliarova-Foucher, E., Sierens, C., Gunter, M. J., Ladas, E. J., Barr, R. D., Van Herck, K., & Huybrechts, I. (2023). Role of Maternal Diet in the Risk of Childhood Acute Leukemia: A Systematic Review and Meta-Analysis. International journal of environmental research and public health, 20(7), 5428. [CrossRef]
- Ayub, A., Ahmad, Q. M., Javed, T., Hayat, M. Z., Farooq, M. A., Anwar, H. M. Z., & Khan, M. A. (2020). Evaluation of diet as a risk factor in the development of childhood leukaemia: a case control study. JPMA. The Journal of the Pakistan Medical Association, 70(3), 404–409. [CrossRef]
- Singh, R. K., Chang, H. W., Yan, D., Lee, K. M., Ucmak, D., Wong, K., Abrouk, M., Farahnik, B., Nakamura, M., Zhu, T. H., Bhutani, T., & Liao, W. (2017). Influence of diet on the gut microbiome and implications for human health. Journal of translational medicine, 15(1), 73. [CrossRef]
- Aman, P., Pettersson, D., Zhang, J. X., Tidehag, P., & Hallmans, G. (1995). Starch and dietary fiber components are excreted and degraded to variable extents in ileostomy subjects consuming mixed diets with wheat- or oat-bran bread. The Journal of nutrition, 125(9), 2341–2347. [CrossRef]
- De Filippis, F., Pellegrini, N., Vannini, L., Jeffery, I. B., La Storia, A., Laghi, L., Serrazanetti, D. I., Di Cagno, R., Ferrocino, I., Lazzi, C., Turroni, S., Cocolin, L., Brigidi, P., Neviani, E., Gobbetti, M., O’Toole, P. W., & Ercolini, D. (2016). High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut, 65(11), 1812–1821. [CrossRef]
- Garcia-Mantrana, I., Selma-Royo, M., Alcantara, C., & Collado, M. C. (2018). Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Frontiers in microbiology, 9, 890. [CrossRef]
- Nagpal, R., Shively, C. A., Appt, S. A., Register, T. C., Michalson, K. T., Vitolins, M. Z., & Yadav, H. (2018). Gut Microbiome Composition in Non-human Primates Consuming a Western or Mediterranean Diet. Frontiers in nutrition, 5, 28. [CrossRef]
- Bäckhed, F., Ley, R. E., Sonnenburg, J. L., Peterson, D. A., & Gordon, J. I. (2005). Host-bacterial mutualism in the human intestine. Science (New York, N.Y.), 307(5717), 1915–1920. [CrossRef]
- Cotillard, A., Kennedy, S. P., Kong, L. C., Prifti, E., Pons, N., Le Chatelier, E., Almeida, M., Quinquis, B., Levenez, F., Galleron, N., Gougis, S., Rizkalla, S., Batto, J. M., Renault, P., ANR MicroObes consortium, Doré, J., Zucker, J. D., Clément, K., & Ehrlich, S. D. (2013). Dietary intervention impact on gut microbial gene richness. Nature, 500(7464), 585–588. [CrossRef]
- Romond, M. B., Ais, A., Guillemot, F., Bounouader, R., Cortot, A., & Romond, C. (1998). Cell-free whey from milk fermented with Bifidobacterium breve C50 used to modify the colonic microflora of healthy subjects. Journal of dairy science, 81(5), 1229–1235. [CrossRef]
- Kim, C. H., Park, J., & Kim, M. (2014). Gut microbiota-derived short-chain Fatty acids, T cells, and inflammation. Immune network, 14(6), 277–288. [CrossRef]
- Russell, W. R., Gratz, S. W., Duncan, S. H., Holtrop, G., Ince, J., Scobbie, L., Duncan, G., Johnstone, A. M., Lobley, G. E., Wallace, R. J., Duthie, G. G., & Flint, H. J. (2011). High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. The American journal of clinical nutrition, 93(5), 1062–1072. [CrossRef]
- De Filippo, C., Cavalieri, D., Di Paola, M., Ramazzotti, M., Poullet, J. B., Massart, S., Collini, S., Pieraccini, G., & Lionetti, P. (2010). Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proceedings of the National Academy of Sciences of the United States of America, 107(33), 14691–14696. [CrossRef]
- Ghosh, T. S., Rampelli, S., Jeffery, I. B., Santoro, A., Neto, M., Capri, M., Giampieri, E., Jennings, A., Candela, M., Turroni, S., Zoetendal, E. G., Hermes, G. D. A., Elodie, C., Meunier, N., Brugere, C. M., Pujos-Guillot, E., Berendsen, A. M., De Groot, L. C. P. G. M., Feskins, E. J. M., Kaluza, J., … O’Toole, P. W. (2020). Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: the NU-AGE 1-year dietary intervention across five European countries. Gut, 69(7), 1218–1228. [CrossRef]
- Keim, N. L., & Martin, R. J. (2014). Dietary whole grain–microbiota interactions: insights into mechanisms for human health. Advances in nutrition (Bethesda, Md.), 5(5), 556–557. [CrossRef]
- West, N. P., Christophersen, C. T., Pyne, D. B., Cripps, A. W., Conlon, M. A., Topping, D. L., Kang, S., McSweeney, C. S., Fricker, P. A., Aguirre, D., & Clarke, J. M. (2013). Butyrylated starch increases colonic butyrate concentration but has limited effects on immunity in healthy physically active individuals. Exercise immunology review, 19, 102–119.
- Schley, P. D., & Field, C. J. (2002). The immune-enhancing effects of dietary fibres and prebiotics. The British journal of nutrition, 87 Suppl 2, S221–S230. [CrossRef]
- Ponzo, V., Fedele, D., Goitre, I., Leone, F., Lezo, A., Monzeglio, C., Finocchiaro, C., Ghigo, E., & Bo, S. (2019). Diet-Gut Microbiota Interactions and Gestational Diabetes Mellitus (GDM). Nutrients, 11(2), 330. [CrossRef]
- Stoeva, M. K., Garcia-So, J., Justice, N., Myers, J., Tyagi, S., Nemchek, M., McMurdie, P. J., Kolterman, O., & Eid, J. (2021). Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut microbes, 13(1), 1–28. [CrossRef]
- Geng, H. W., Yin, F. Y., Zhang, Z. F., Gong, X., & Yang, Y. (2021). Butyrate Suppresses Glucose Metabolism of Colorectal Cancer Cells via GPR109a-AKT Signaling Pathway and Enhances Chemotherapy. Frontiers in molecular biosciences, 8, 634874. [CrossRef]
Figure 1.
Presence of microbiota in the anatomical structures of our body and influence on different districts of our body.
Figure 1.
Presence of microbiota in the anatomical structures of our body and influence on different districts of our body.
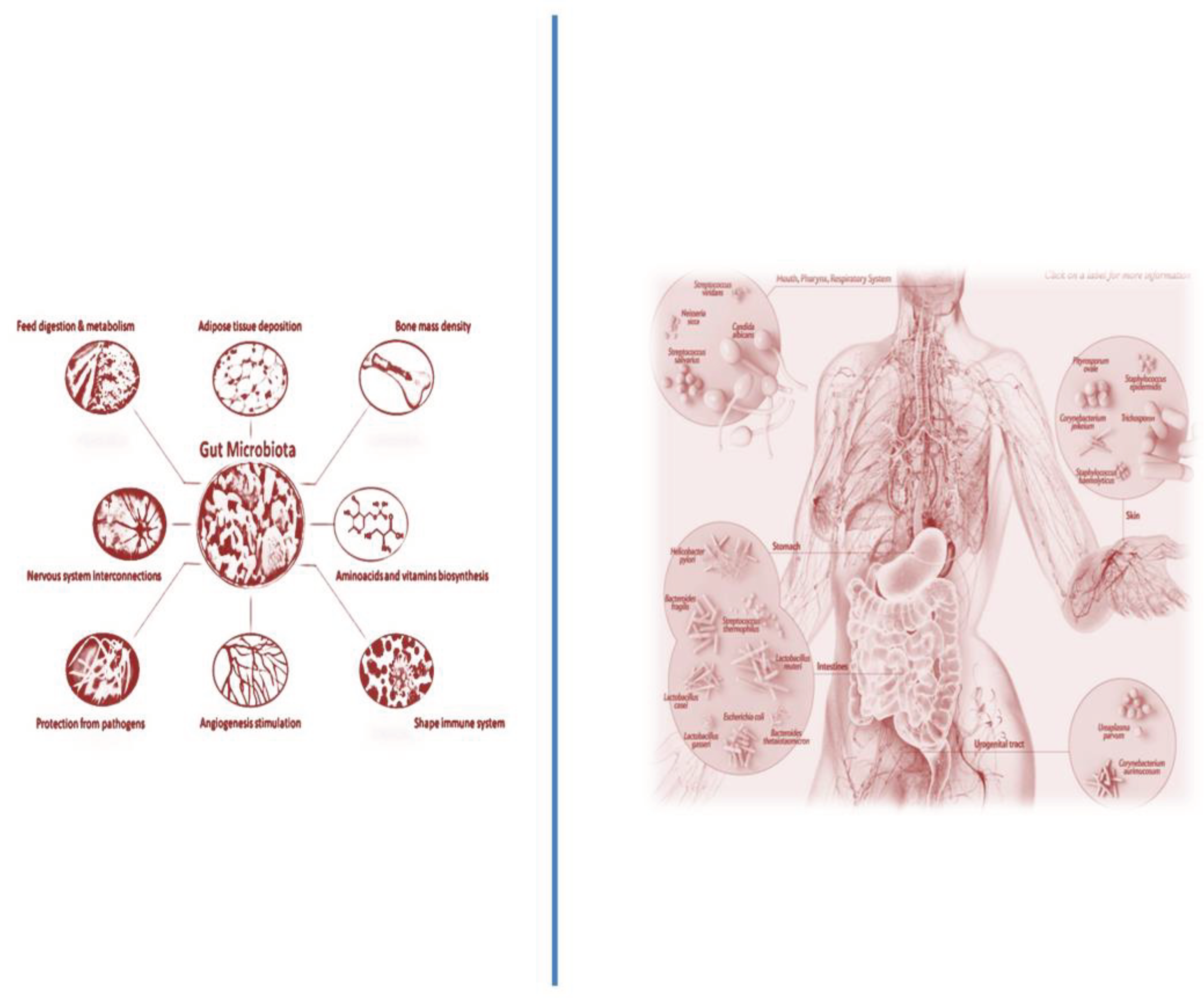
Figure 2.
Two levels of interventions, recommendations, and therapeutical strategies in parents (A) and in children after the birth (B).
Figure 2.
Two levels of interventions, recommendations, and therapeutical strategies in parents (A) and in children after the birth (B).
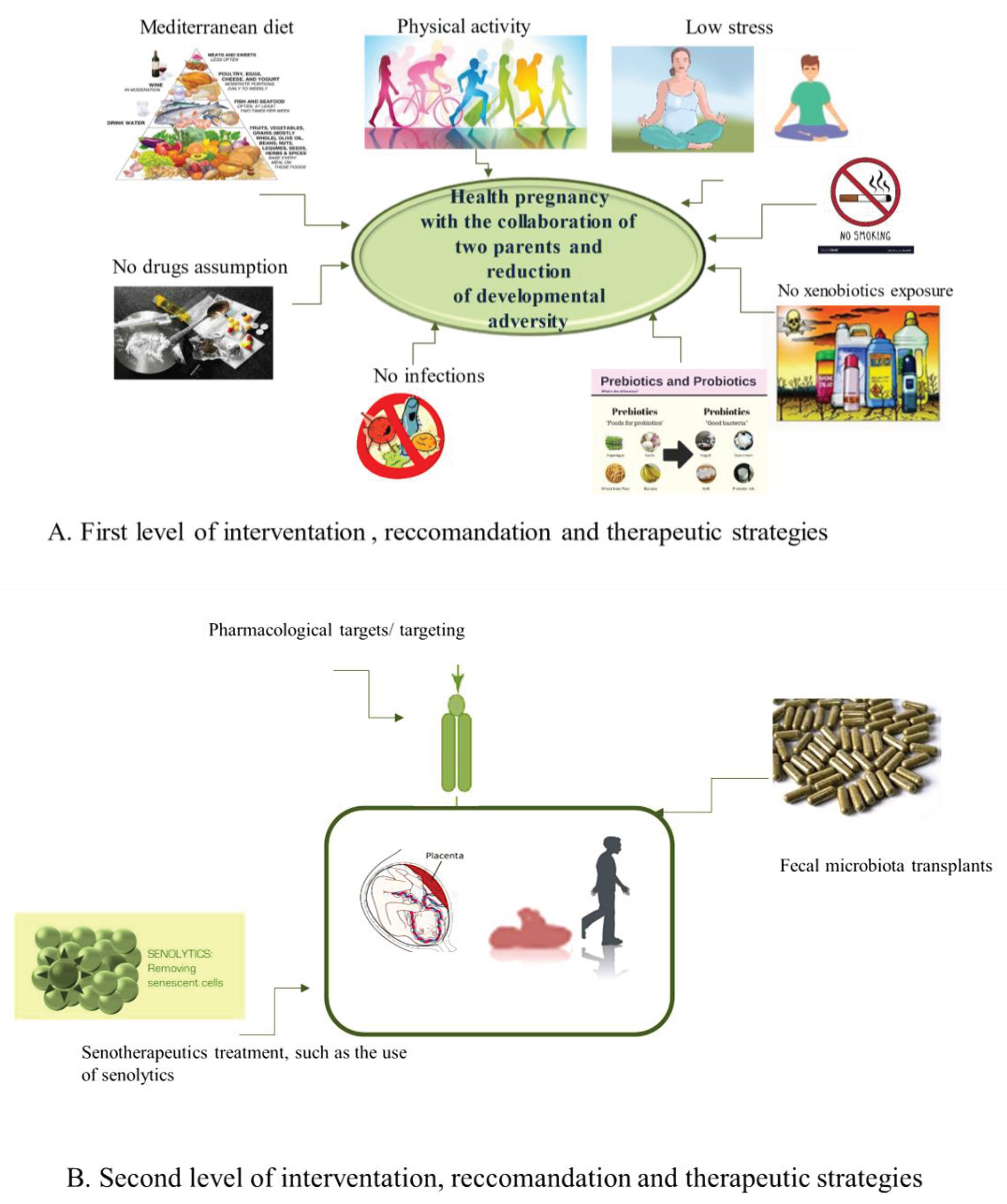
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



