
Submitted:
19 March 2024
Posted:
19 March 2024
You are already at the latest version
Abstract
This study evaluates the efficacy of the salts, sodium metabisulfite (SMB), ammonium bicarbonate, sodium bicarbonate and potassium dihydrogen orthophosphate first in vitro against the main postharvest fruit rot fungi, Alternaria alternata, Botrytis cinerea, Penicillium italicum and Penicillium digitatum. Results showed that 0.2% SMB completely inhibited mycelium growth of the fungal species. Ammonium bicarbonate and sodium bicarbonate were less effective at 0.2% in inhibiting mycelial growth, ranging from 57.6% to 77.6%. The least effective was potassium dihydrogen or-thophosphate. Experiments were also performed in vivo on wounded apples inoculated with the most pathogenic fungus, B. cinerea, and treated with SMB at concentrations of 0.2, 0.5, 1, 2 and 3%, both preventively and curatively. Results based on the decay size showed that SMB, when used as a preventive treatment, had a reduced efficacy, even with the highest concentration. However, this salt proved to be very effective at 0.5% in curative treatment, since the decay was completely blocked. Our results suggest that the appropriate concentration of SMB for post-harvest treatment is 0.5% as a curative treatment. On the other hand, the 1% dose induced the onset of phytotoxicity around the wound. To assess the extent of phytotoxicity reaction, higher concentrations of 1-4% SMB were applied to wounded fruit. Apples and oranges were inoculated or not with B. cinerea and P. digitatum respectively. Doses of 1- 4% induced phytotoxicity in the form of a discoloured ring surrounding the wound on the epidermis of the fruit; this phytotoxicity enlarged as the concentration of SMB increased. The phytotoxic features were similar on apples and oranges. The methodological procedure made it possible to carry out a quantitative assess-ment of SMB phytotoxicity. This method is proposed as an easy-to-use technique for quantitatively estimating the phytotoxicity of antifungal compounds on post-harvest fruit.
Keywords:
control
; decay
; GRAS salts
; postharvest
; sodium metabisulfite
1. Introduction
Fresh fruit and vegetables still contain high amount of water after being harvested, leading to metabolic process, such as ripening and senescence. This relatively high water content is likely to play a key role in increasing the susceptibility of fruit to the fungi responsible for decomposition and the often-high levels of losses and waste [1]. Injuries on fruit during harvesting, postharvest handling, and commercialization are the primary cause of fungal infections [2]. Postharvest control of fruit rot has been relied on synthetic fungicides. However, resistant strains and increased residues in treated fruit are main factors limiting their use [3,4].
Alternatives are increasingly required to control postharvest diseases including the use of safe low-toxicity compounds, such as natural products with low environmental impact. Organic and inorganic salts, generally recognized as safe (GRAS) ingredients, are considered antifungal products possessing properties as postharvest fruit preservatives [5]. For that, GRAS salts were approved by the United States Food and Drug Administration (USFDA) and by the European Food Safety Authority (EFSA) allowing its use in food as conservatives [2,6].
GRAS salts such as carbonates, bicarbonates, sorbates, benzoates, parabens and metabisulfites, used as aqueous solutions or ingredients of edible coating are widely cited for controlling postharvest diseases [7,8,9,10,11,12]. Sulfites group, including sulfur dioxide (SO2) (E -numbers E 220-228), are currently approved for some uses on entire fresh fruit. Sulfur dioxide and sulfites are authorized as food additives in the EU [13]. Recent literature suggests that sulfur-containing salts, such as metabisulfites are effective in controlling important postharvest fruit diseases [14,15,16,17]. For instance, the GRAS salts sodium carbonate and sodium bicarbonate were reported fungistatic compounds able to reduce temporary postharvest decay without detectible phytotoxic effect on treated fruit [18]. These salts applied singly on fruit were effective against green mold on lemons and oranges but their activity was least on infected clementine and mandarins [12,19]. Moreover, application of hydrogen peroxide (20 g L−1) followed by potassium phosphite (20 g L−1) controlled both blue and green molds on ‘Eureka’ lemon fruit [20].
The objectives of the study were: (i) Evaluation of the in vitro antifungal activity of four GRAS salts against A. alternata, B. cinerea, P. digitatum and P. italicum, (ii) Assessment of the in vivo preventive and curative activity of the most promising salt to control decay on apple fruit, (iii) Identification of the signs of phytotoxicity in treated apples and oranges, for which a quantitative assessment was performed.
2. Materials and Methods
2.1. Fungal Species
Isolates of A. alternata, B. cinerea, P. digitatum and P. italicum were obtained from decayed fruit (mandarin and apple) from local markets ‘Le Kram’ and ‘Grombalia’ (Tunisia). Isolates were identified at the level of species and conserved in PDA medium at room temperature (around 20 °C for 7-14 d in obscurity) for further inoculation.
2.2. GRAS Salts Tested In Vitro
Sodium metabisulfite (SMB), ammonium bicarbonate (AMB), sodium bicarbonate (SB) and potassium dihydrogen orthophosphate (PDO) were firstly tested in vitro (PDA) against the main fungal species of postharvest fruit decay above cited. PDA medium was amended with the respective salt at 0.2 % before autoclaving at 120 °C during 20 min. PDA without salt was served as control. Mycelial plug (10 mm diameter) from 7 to 14 days-old cultures of each isolate was placed in the center of the medium in 90-mm plastic Petri dishes, then incubated at 20 ± 2 °C in obscurity.
Radial mycelial growth was determined in each Petri dish by measuring two perpendicular diameters of the fungal colony. These measurements were taken after 4, 7 and 9 days of incubation depending on the mycelial growth rate of the fungal species. Three replicates (3 Petri dishes) were used for each salt and fungal pathogen. The results were expressed as a percentage inhibition of mycelial growth using the following formula:
where dc = average diameter of the fungal colony on control plates (mm)and dt = average diameter of the fungal colony (mm) on Petri dishes amended with each salt.
Mycelium inhibition (%) = [(dc-dt)/dc] × 100,
2.3. GRAS Salt Tested on Fruit
The fruit used in the experiments were apples (Malus domestica. L) var. 'Golden' and oranges (Citrus sinesnsis. L) var. 'Maltaise' purchased from a supermarket in Ariana (Tunisia). The fruits were selected without wounds, with a healthy appearance, randomized, and surface disinfected by immersing them for 4 minutes in 0.72% sodium hypochlorite, rinsed twice with sterilized distilled water, and left to air dry under aseptic conditions.
Preliminary trial was undertaken in order to determine the appropriate kind of wound on fruit that allows rapid fungal progression. The fruit wounds were either 10 mm in diameter using a cork-borer, or five adjacent small punctures (within a 10 mm diameter circle) using a sterilized needle, with the control fruit intact. Figure 1 shows that a 10 mm diameter cork-borer wound was the most appropriate, as the larger the wound, the greater the diameter of the rot and its certainty of development. This type of wound was used in subsequent trials.
Accordingly, fruit were wounded twice in the equatorial diameter in opposite positions using a cork-borer (10 mm diameter, 2 mm depth) immediately before treatment or inoculation. The inoculation consists of a fungal mycelium 10 mm in diameter from the edge of a growing colony, which is placed upside down in each wound. The most effective salt in the in vitro experiment, sodium metabisulphite, was used in the following bioassays at different concentrations. For the curative treatment, apples inoculated with B. cinerea were incubated in room ambient (temperature around 16°C) for 24 h, then dipped for 1 min in SMB solutions at concentrations of 0.5, 1, 2 or 3%, and incubated at room temperature for 7 days. For the preventive treatment, the fruits were first treated with SMB at the concentrations mentioned above and kept at room temperature for 24 hours, then inoculated with B. cinerea and incubated for 7 days at the same room ambient. In all cases, the fruits were completely soaked for 1 min in a precise dose of SMB solution in recipients of 1 L each. Fruit soaked for 1 min in distilled water served as the positive control and the negative control was fruit soaked in a solution (4 mL/L) of the commercial fungicide 'Celest extra'.
Three replicates (3 fruit) per treatment were used and the decay diameter was measured after 7 days of incubation. Fruit rot diameter (mm) was measured twice perpendicularly (to maintain an average measurement per fruit) using a ruler. The percentage of mycelial inhibition was calculated as described below.
% inhibition = [(dc-dt)/dc] × 100, where dc = average diameter of the control fruit and dt = average diameter of the treated fruit.
2.4. GRAS Salt and Phytotoxicity
‘Golden' apples and 'Maltaise' oranges were wounded equatorially on two diametrically opposite sides. A first lot of fruit was inoculated by depositing a plug of mycelium (10 mm in diameter and 2 mm deep) of B. cinerea for the apples and of P. digitatum for the oranges, placed in the wound of the same size as the plug to completely cover the wound. After two hours, the inoculated fruit were immersed for 1 min in solutions each containing 0.5, 1, 2, 3 or 4% SMB. The second lot (non-inoculated) was wounded in the same way and then dipped for 1 minute in the SMB solutions mentioned above. The wounded fruits of the control were dipped in distilled water only. The fruits were incubated at an ambient temperature of 16 ± 2°C. Each treatment contained three fruits, one fruit being a replicate. The rot diameter of each fruit was measured (in mm using a ruler) twice perpendicularly to determine an average measurement per fruit.
The phytotoxicity caused by the application of SMB to the fruit was assessed at the end of the incubation period when rots reached the apical and/or the peduncle fruit surface. Oranges and apple were incubated for 6 days and 8 days respectively. The phytotoxicity of SMB on wounded fruit was estimated on the basis of the rot diameter. For that, we first calculated:
where ɸSMBI is the decay diameter of wounded, inoculated and treated fruit. ɸc+ is the diameter of wounded and inoculated but untreated fruit (positive control).
Decay inhibition (%) = [1- (ɸSMBI / ɸc+)] ×100
To distinguish between the fungal rot and that induced by the salt, the following approach was pursued. As the fruit rot diameter is the sum of the wound diameter and the rot size itself, it is more accurate to calculate only the rot size for wounded, inoculated and treated fruit (named: A) and the rot size for wounded and treated fruit only (named: B).
A (mm) = (ɸSMBI - ɸC-), where ɸSMBI is the decay diameter of wounded, inoculated and treated oranges or apples, ɸc- is the diameter of only wounded fruit (negative control).
B (mm) = (ɸSMB - ɸC-), where ɸSMB is the decay diameter of wounded and treated fruit.
The size of the rot caused by the fungus is therefore calculated as (A-B).
2.5. Statistical Analyses
Data from all experiments were subjected to analysis of variance (ANOVA). The main factors were GRAS salts and fungal species for the in vitro experiment, and treatments (preventive or curative) and salt concentrations for the in vivo experiment. For the phytotoxicity test, the main factors were SMB concentrations and inoculated or non-inoculated fruit.
3. Results
3.1. In Vitro Experiment
Analysis of variance shows a highly significant effect of GRAS salts, fungi and the interaction between these two factors on mycelial growth (Table 1). GRAS salts had a strong influence on mycelial growth, since the source of variation was high (F = 230.4). Figure 2 shows the inhibition of mycelial growth of B. cinerea, A. alternate, P. digitatum and P. italicum in Petri dishes (PDA) amended with each of the four salts after incubation at 20 °C. SMB at 0.2 % completely inhibited the mycelial growth of the four fungal species. For AMB, SB and PDO, significant differences were observed depending on the target pathogen (Figure 2). AMB differentially inhibited the mycelial growth of B. cinerea (77.6%), A. alternata (31.1%), P. digitatum (22.3%) and P.italicum (6.9%). However, SB was similarly effective in inhibiting mycelial growth in B. cinerea (56%) and P. digitatum (57.6%), followed by A. alternata (26.4%) and P. italicum (14.3%). The least effective salt was PDO, with a low inhibition of mycelial growth ranging from 26 % to -23 % (negative value means better growth stimulation than the control).
Furthermore, we carried out an additional test with SMB to check, by lowering this concentration to 0.1% SMB, whether there could be a mycelial growth by any of the fungal species at this lower concentration. It appeared at this concentration of 0.1% that A. alternata and P. italicum showed limited growth of 20 mm (80 mm control, inhibition rate of 75 %), 25 mm (74 mm control, inhibition rate of 66%) respectively, while B. cinerea, and P. digitatum were totally inhibited at this concentration. SMB therefore proved highly effective at a concentration of 0.2%, irrespective of the fungal species, and was used in subsequent in vivo trials to confirm this efficacy.
3.2. In Vivo Activity of Sodium Metabisulphite
SMB concentrations ranging from 0.2 to 3 % were evaluated in vivo as preventive and/or curative treatments. Significant differences between treatments (preventive and curative) and between SMB concentrations were recorded as indicated in the ANOVA (Figure 3, Legend). The preventive activity of SMB was less effective against B. cinerea on apples than the curative activity, due to a low reduction in rot compared with the curative treatment. Thus, the percentage of rot inhibition on apples inoculated and soaked preventively in 2% and 3% SMB was 37.6% and 58.8% respectively. However, the reduction in rot on apples inoculated and soaked curatively in 2% and 3% SMB solutions was 80% and 76.4% respectively, compared with the commercial fungicide, which was completely effective (100%) when used preventively or curatively.
Increasing SMB concentrations from 1% to 3% had no significant added effect on rot reduction, particularly in the curative treatment. However, a zone of necrosis similar to phytotoxicity was observed around the wound on the fruit skin, which increased with increasing SMB concentrations. As a result, other in vivo trials were conducted with this salt at low concentrations (0.2%, 0.5% and 1%). When applied at the 0.2% concentration, SMB reduced the size of rot on apples by up to 77.6% (Figure 4). However, the 0.5% concentration completely inhibited rot on inoculated apples (99.6%). Soaking the inoculated apples in a solution containing 1% SMB reduced decay by 95.2% with concomitant mild phytotoxicity on the fruit pericarp, such as whitening and collapse at the inoculation point. Thus, curative treatment with SMB at 0.5% or 1% was the most effective concentration, almost similar to the classical fungicide ('Celest Extra'). SMB was effective against rot caused by B. cinerea on apples at the non-apparent-phytotoxic dose of 0.5 %.
3.3. Phytotoxicity of Sodium Metabisulphite
Over time, fruit soaked in SMB solutions showed a translucent circular white rot (Figure 5) that extended in a circular pattern surrounding the wound that had been perforated by the cork-borer. According to the ANOVA in figure 5 (legend), the concentration of SMB was a significant factor in the induction of this rot zone, while the effect of the pathogen on this rot was not significant.
The size of the rot was measured on oranges and apples inoculated and treated with increasing concentrations of SMB from 0.5 to 4 % (Table 2). In this case, rot size ranged from 2.5 to 9.5 mm for apples (8 dpi) and from 0.9 to 14.9 mm for oranges (6 dpi). When the fruits were treated only, rot size ranged from 2.8 to 11.7 mm for apples and from 1.3 to 9.2 mm for oranges. As a result, the percentage of rot inhibition compared to the control (inoculated untreated fruit) decreased with increasing SMB concentration, ranging from 98.7 (0.5 % SMB) to 78.2 (4 % SMB) for oranges and from 93.2 (0.5 % SMB) to 71.2 % (4 % SMB) for apples.
These results indicate that SMB concentrations appear to be fully effective from 0.5% and that the necrosis produced on fruit treated with concentrations exceeding 0.5% is not due to fungal infection but rather linked to SMB phytotoxicity, which is also confirmed by the ANOVA. To exploit this point, we took, for each concentration, the values for inoculated-treated and non-inoculation-treated fruit as replicates of SMB treatment only in order to determine the average size of the necrosis for each concentration. Such adjustment of the data made it possible to determine a linear regression linking the concentration of SMB (x) to the corresponding average size of the necrosis (y). This gives the equations y= 3.76.x-0.2 with an R2 = 0.99 for oranges and y=2.47.x with an R2 = 0.96 for apples. Figure 6 shows these two straight lines, which fit the measured values perfectly.
To validate the accuracy of this linear relationship between SMB concentration and necrosis diameter, we performed another trial in which apples and oranges were only wounded and treated with higher concentrations of SMB (7 % and 10 %), in addition to the control (Figure 7).
The diameter of the necrosis on the fruit for each treatment after 6 days for oranges and 8 days for apples is shown in the Table 3 illustrating the values expected according to the two linear formulae mentioned above and those measured. It appears from Chi-square test that the hypothesis of equality between the observed and expected values is well-validated (null hypothesis accepted).
4. Discussion
Our results from in vitro trials using GRAS salts showed that 0.2% AMB induced 77.6% inhibition against B. cinerea, but showed limited activity against other fungal species. SB was less effective than AMB, with inhibition of 56 and 57.6% against B. cinerea and P. digitatum respectively. The PDO was not only ineffective but also induced mycelial growth that was greater than that of the control, particularly in the case of P. digitatum and P. italicum. Previous results indicated that at a concentration of 0.16 - 0.50 %, ammonium, potassium and sodium bicarbonates inhibited approximately 95% of the in vitro growth of B. cinerea colonies [21]. This inhibition is explained by the buffering capacity of the carbonate, which alkalinises the environment of pathogens such as B. cinerea, which needs an acidic medium to grow properly [22]. Zhao et al. [23] evaluated in vitro the antifungal activity of seventeen GRAS salts against four postharvest fungal diseases of citrus fruit. Their results, based on inhibition rates, pointed to differential reactions of fungi to the salts, in agreement with our results. Thus, sodium silicate (1%) was 100% inhibitory against all tested fungi. In contrast, sodium carbonate (1%) was 100% effective against P. digitatum and P. italicum, but had no activity against G. citri-aurantii or G. gloeosporioides. In this respect, Lyousfi et al. [24] evaluated in vitro some organic and inorganic salts (food additives) as antifungal agents against Monilinia fructigena. Most additives showed significant inhibition of mycelial growth, but the extent of the inhibition varied according to the additive and its concentration (sodium bicarbonate, sodium carbonate, copper sulfate being the most effective, while ammonium carbonate and citric acid were the least effective).
Sulphur-containing food additives act effectively in vitro against various postharvest fungal diseases. This was observed in our in vitro tests with SMB at 0.2%, which was the most effective, irrespective of the fungal species, as mycelial growth was completely inhibited at this concentration. Kim et al. [25] highlighted that sodium metabisulfite was one of the most effective salts for controlling microbial growth. Tian et al. [26] demonstrated the strong in vitro antifungal activity of potassium metabisulfite and sodium hyposulfite (0.1%) against four kiwifruit soft rot fungi, inhibiting mycelial growth in a concentration-dependent manner. SMB completely inhibited mycelial growth of A. alternata and B. cinerea when used at a concentration of 0.2 M (3.8 %) [6]. The sulphur salt completely inhibited the mycelial growth of P. italicum at 0.4 % [9] and of P. digitatum at 2 % [27]. Inhibition of P. digitatum and P. italicum by potassium and sodium metabisulfite in Petri Dish at concentrations of 0.19-1.9 % has also been reported [5]. Sodium metabisulfite at a concentration of 0.19 % completely inhibited mycelial growth of A. solani [6]. In another study, 0.01% SMB caused total inhibition of conidial germination and germ-tube elongation in Venturia inaequalis [28], such concentration, according to our results on mycelial growth, seems to be a very low dose to ensure total inhibition in vitro. Our in vitro results showed that 0.2% SMB is effective in ensuring total inhibition of fungi in vitro, but that 0.1% is not sufficient to achieve the same result and is therefore considered a limiting concentration for in vitro efficacy.
According to Talibi et al. [29], the ability of GRAS salts to control postharvest diseases can be estimated by in vitro experiments, but in vivo bioassays are still needed to confirm the in vitro results. In the first part of our in vivo study, inoculated apples were dipped in different concentrations of SMB (1 %, 2 % and 3 %) as preventive and curative treatments. Results showed that SMB was ineffective when used preventively at all concentrations. However, the 1 % curative dip showed high efficacy (92.4%) compared with the commercial fungicide (100 %). In the second part of the study, we tested SMB curatively on apples at concentrations of 0.2 %, 0.5 % and 1%. The results showed that SMB, when used curatively at 0.5%, was effective in controlling rot on apples inoculated with B. cinerea.
Sulphite salts dissolved in aqueous solutions are released as sulphur dioxide (SO2) (67.4%) [13,30]. It has been reported that the mode of action of sulphite salts depends on pH [31]. Contact between the salt solution and the fruit pericarp can modify the pH of the wound in which the fungus develops [5]. The antifungal activity of sodium metabisulphite may increase because the pH at the point of inoculation is lower than that of the surrounding fruit tissue. The sulphur dioxide released by metabisulphites alters the permeability of the fungal membrane [31] and accumulates in the cytoplasm, inhibiting mycelial growth through intercellular reactions [13,30]. Moreover, other factors may influence the mode of action of sulfur-containing fungistatic salts. Accordingly, it has been reported that pathogenicity of some fungal pathogens increases, and are able to alkalinize the medium in the peel infection wound, such as Alternaria. Whereas other species, such as Penicillium acidify the ambient [32,33,34].
The ineffectiveness of SMB when applied as a preventive measure could be attributed to the gradual dissipation of the sulphur component, so that the subsequent fungal infection lost its full activity. However, there are reports on other GRAS salts without a sulphur component, such as potassium sorbate, showing that they are also ineffective as a preventative measure. For example, Olmedo et al. [35] reported that potassium sorbate was tested against green and blue mould on lemon fruit inoculated and treated preventively and curatively. They pointed out that no efficacy was observed as a preventive measure, whereas curative treatment was effective. The same observation was underlined when thermal treatments were used. In fact, Lydakis and Aked [36] reported that ‘Sultanina' table grape berries inoculated with B. cinerea after vapour heat treatment (preventive treatment) were more susceptible to the fungal disease than the control. Nevertheless, curative heat treatments applied after inoculation significantly reduced infection compared to control. In a review, Palou et al. [37] considered that the lack of preventive activity is one of the main handicaps of GRAS salts. Our results on the ineffectiveness of preventive treatment of fruit with SMB are in accordance with the previous findings using other means of treatments.
Phytotoxicity of sodium metabisulfite was assessed at concentrations from 0.5 to 4 % on apples and oranges. Necrosis was developed on fruit peel around the point of injury even though on un-inoculated fruit. This confirms that necrosis is not only produced by pathogenic fungi, but also by SMB at concentrations above 0.5 %. An increase of the necrosis area was registered even in healthy fruit (fruit wounded and treated but not inoculated). In literature, salts inducing phytotoxicity on fruit pericarp was described. For example, the application of copper sulfate has shown phytotoxic effects on fruit rinds, leading to visible darkening, and sinking at the inoculation point [24]. Martínez-Blay et al. [5] pointed out the phytotoxicity of sodium and potassium metabisulfite as well as aluminum and potassium sulfate on citrus fruit.
We can assume that dipping healthy fruit in high concentrations of SMB would affect the cellular cohesion of the fruit pericarp, which would then allow the fruit to be infected more rapidly. In addition, since the natural antagonists of the microbiome, which are normally latent on the pericarp, would be disrupted after treatment of the fruit, the pathogen would be able to multiply again without mechanical/antagonistic barriers. For this reason, using a higher level of sodium metabisulphite will not improve the inhibition of rot size, but will increase the sensitivity of the fruit skin to the side effect of this salt. Our analyses highlighted that the appropriate concentration was 0.5% in the curative treatment without showing any noticeable phytotoxicity on the fruit. Thus, Martinez-Blay et al. [5] reported that the application of SMB at a concentration of 100 mM was phytotoxic on 'Valencia' oranges, but that 50 mM caused no apparent damage to the rind of the treated fruit and was very effective against mould. The 50 mM concentration (0.85 %) is very close to 0.50 %, showing in our trials efficacy with no apparent phytotoxic reaction in treated fruit.
To our knowledge, the evaluation of salt phytotoxicity was mainly based on the qualitative visual description of the symptoms appearing after treatment of the fruit. For example, in dip treatments of stone fruit to control fungal pathogens after harvest using food additives, phytotoxicity on the fruit skin caused by the salts was assessed visually on a scale of 1 to 4, depending on the surface area covered by fungal lesions [38]. Delisle-Houde et al. [39] used also visual scale of 0-4 to assess the severity of phytotoxicity of different salts on foliar surface of lettuce infected with the bacteria ‘Pseudomonas cichorii’ causing lettuce varnish spot. Our analyses on severity of phytotoxicity used a quantitative procedure by relating the size of necrosis measured on treated fruit to the corresponding concentration of salt. We suggest using the protocol/methodology applied here as a model for a quantitative analysis of the phytotoxicity of compounds used to treat fruit. This suggestion may become valid after other tests including different compounds on various fruit (apples are the best candidates thanks to the obvious necrosis margin).
Author Contributions
M.B.A; Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, and Writing – original draft, Writing – review & editing, M.A; Formal analysis, Data curation, Software, Visualization, and Writing, Writing – original draft, Writing – review & editing. All authors have read and agreed to the published version of the manuscript.
Funding
Ministry of higher Education and Scientific Research of Tunisia funded this research in the framework of PRIMA, project acronym StopMedWaste ‘Innovative Sustainable technologies TO extend the shelf-life of Perishable MEDiterranean fresh fruit, vegetables and aromatic plants and to reduce WASTE’, Project Number 1556.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Sample Availability
Samples of the compounds are available from the authors.
References
- Romanazzi, G.; Moumni, M. Chitosan and other edible coatings to extend shelf life, manage postharvest decay, and reduce loss and waste of fresh fruits and vegetables. Current Opinion in Biotechnol 2022, 78, 102834. [Google Scholar] [CrossRef] [PubMed]
- Palou, L. Postharvest treatments with GRAS salts to control fresh fruit decay. Hortic 2018. 4, 46. [CrossRef]
- Brent, K.J.; Hollomon, D.W. Fungicide resistance in crop pathogens: how can it be managed?, 3rd ed.; Brussels GIFAP,1995; pp 48-56.
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant-and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharv Biol Technol 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Martínez-Blay,V. ; Taberner, V.; Pérez-Gago, M.B.; Palou, L. Control of major citrus postharvest diseases by sulfur-containing food additives. Int J Food Microbio 2020, 330, 108713. [Google Scholar]
- Mills, A.A.S.; Platt, H.W. , Hurta, R.A. Effect of salt compounds on mycelial growth, sporulation and spore germination of various potato pathogens. Postharv Biol Technol 2004, 34, 341–350. [Google Scholar] [CrossRef]
- Guimaraes, J. E.; de la Fuente, B.; Pérez-Gago, M. B.; Andradas, C.; Carbó, R.; Mattiuz, B. H.; Palou, L. Antifungal activity of GRAS salts against Lasiodiplodia theobromae in vitro and as ingredients of hydroxypropyl methylcellulose-lipid composite edible coatings to control Diplodia stem-end rot and maintain postharvest quality of citrus fruit. Int J Food Microbiol 2019, 301, 9–18. [Google Scholar] [CrossRef]
- Montesinos -Herrero, C.; Moscoso -Ramírez, P.A.; Palou, L. Evaluation of sodium benzoate and other food additives for the control of citrus postharvest green and blue molds. Postharv Biol Technol 2016, 115, 72–80. [Google Scholar] [CrossRef]
- Askarne, L.; Boubaker, H.; Boudyach, E. H.; Aoumar, A.A.B. Use of food additives to control postharvest citrus blue mold disease. Atlas J Biol 2013, 2, 147–153. [Google Scholar] [CrossRef]
- Valencia-Chamorro, S.A.; Perez-Gago, M.B.; Del Rio, M. A.; Palou, L. Curative and preventive activity of hydroxypropyl methylcellulose-lipid edible composite coatings containing antifungal food additives to control citrus postharvest green and blue molds. J Agric Food Chem 2009, 57, 2770–2777. [Google Scholar] [CrossRef]
- Smilanick, J. L.; Mansour, M. F.; Gabler, F.M.; Sorenson, D. Control of citrus postharvest green mold and sour rot by potassium sorbate combined with heat and fungicides. Postharv Biol Technol 2008, 47, 226–238. [Google Scholar] [CrossRef]
- Palou, L.; Usall, J.; Muñoz, J.A.; Smilanick, J.L.; Viñas, I. Hot water, sodium carbonate, and sodium bicarbonate for the control of postharvest green and blue molds of clementine mandarins. Postharv Biol Technol 2002, 24, 93–96. [Google Scholar] [CrossRef]
- EFSA Panel on Food additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of sulfur dioxide (E 220), sodium sulfite (E 221), sodium bisulfite (E 222), sodium metabisulfite (E 223), potassium metabisulfite (E 224), calcium sulfite (E 226), calcium bisulfite (E 227) and potassium bisulfite (E 228) as food additives. EFSA Journal 2016, 14, 4438. [Google Scholar]
- Bhatta, U.K. . Alternative management approaches of citrus diseases caused by Penicillium digitatum (green mold) and Penicillium italicum (blue mold). Front Plant Sci 2022, 12, 833328. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Zou, Y.; Li, W.; He, F.; Huang, X.; Wu, X. Pre-and postharvest measures used to control decay and mycotoxigenic fungi in potato (Solanum tuberosum L.) during storage. Critic Rev Food Sci Nutri 2021, 62, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Edelenbos, M. Postharvest environmentally and human-friendly pre-treatments to minimize carrot waste in the supply chain caused by physiological disorders and fungi. Trends Food Sci Technol 2021, 112, 88–98. [Google Scholar] [CrossRef]
- Nilmini, R.K.; Kodituwakku, T.D.; Abeywickrama, K.; Kuruppu, M. In vitro and in vivo Application of Eco-friendly Treatments to Control Postharvest Stem-end Rot of Naturally Infected Avocado (cv. Pollock). J Agric Sci 2021, 16. [Google Scholar] [CrossRef]
- Usall, J.; Smilanick, J.; Palou, L.; Denis-Arrue, N.; Teixidó, N.; Torres, R.; Vinas, I. Preventive and curative activity of combined treatments of sodium carbonates and Pantoea agglomerans CPA-2 to control postharvest green mold of citrus fruit. Postharv Biol Technol 2008, 50, 1–7. [Google Scholar] [CrossRef]
- Smilanick, J.L.; Mackey, B.E.; Reese, R.; Usall, J.; Margosan, D.A. Influence of concentration of soda ash, temperature, and immersion period on the control of postharvest green mold of oranges. Plant Dis 1997, 81, 379–382. [Google Scholar] [CrossRef]
- Cerioni, L.; Sepulveda, M.; Rubio-Ames, Z.; Volentini, S.I.; Rodríguez-Montelongo, L.; Smilanick, J.L.; Ramallo, J.; Rapisarda, V.A. Control of lemon postharvest diseases by low-toxicity salts combined with hydrogen peroxide and heat. Postharvest Biol Technol 2013, 83, 17–21. [Google Scholar] [CrossRef]
- Palou, L.; Usall, J. Smilanick, J.L. Aguilar, M.J. Viñas, I. Evaluation of food additives and low-toxicity compounds as alternative chemicals for the control of Penicillium digitatum and Penicillium italicum on citrus fruit. Pest Manag Sci 2001, 58, 459–466. [Google Scholar] [CrossRef]
- Alaoui, F.T.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Ait Ben Aoumar, A. Control of gray mold disease of tomato by postharvest application of organic acids and salts. Plant Pathol J 2017, 16, 62–72. [Google Scholar]
- Zhao, J.; Wang, Y.; Liu, Q.; Liu, S.; Pan, H.; Cheng, Y. , & Long, C. The GRAS Salts of Na2SiO3 and EDTA-Na2 Control Citrus Postharvest Pathogens by Disrupting the Cell Membrane. Foods 2023, 12, 2368. [Google Scholar] [PubMed]
- Lyousfi, N.; Legrifi, I.; Ennahli, N.; Blenzar, A.; Amiri, S.; Laasli, S. E.; Lahlali, R. Evaluating Food Additives Based on Organic and Inorganic Salts as Antifungal Agents against Monilinia fructigena and Maintaining Postharvest Quality of Apple Fruit. J Fungi 2023, 9, 762. [Google Scholar] [CrossRef]
- Kim, Y. H.; Kim, G. H.; Yoon, K. S.; Shankar, S.; Rhim, J.W. Comparative antibacterial and antifungal activities of sulfur nanoparticles capped with chitosan. Microbial Pathogen 2020, 144, 104178. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, L.; Wang, R.; Ji, N.; Ma, C.; Lei, J.; Zhang, X. Pullulan-Based Active Coating Incorporating Potassium Metabisulfite Maintains Postharvest Quality and Induces Disease Resistance to Soft Rot in Kiwifruit. Foods 2023, 12, 3197. [Google Scholar] [CrossRef] [PubMed]
- Enginsu, S.; Erper, I.; Yildirim, E.; Avci, S.; Turkkan, M. Evaluation of antifungal activity of organic and inorganic salts against Penicillium digitatum, the causal agent of citrus green mould. Abstract Book 4th Int. Agric. Congress. 2018, Temmuz, Turkey. [Google Scholar]
- Arslan, U. Efficacy of boric acid, monopotassium phosphate and sodium metabisulfite on the control of apple scab. J Phytopathol 2016, 164, 678–685. [Google Scholar] [CrossRef]
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E. H.; Aoumar, A. A. B. In vitro and in vivo antifungal activities of organic and inorganic salts against citrus sour rot agent Geotrichum candidum. Plant Pathol. J, 2011, 10, 138–145. [Google Scholar] [CrossRef]
- Davidson, P.M.; Junja, V.K.; Branen, A.L. Antimicrobial agents. In Food Additives; Branen, A.L., Davidson, P.M., Salminen, S., Thorngate, J., Eds.; Marcel Dekker Inc.: New York, USA, 2003; pp. 563–620. [Google Scholar]
- Divol, B.; Du Toit, M.; Duckitt, E. Surviving in the presence of sulphur dioxide: Strategies developed by wine yeasts. Appl. Microbiol. Biotechnol. 2012, 95, 601–613. [Google Scholar] [CrossRef]
- Prusky, D. , Yakoby, N. Pathogenic fungi: leading or led by ambient pH? Mol Plant Pathol 2003, 4, 509–516. [Google Scholar] [CrossRef]
- Prusky, D.; Lichter, A. Mechanisms modulating fungal attack in postharvest pathogen interactions and their control. Eur J Plant Pathol 2008, 121, 281–289. [Google Scholar] [CrossRef]
- Zhang, Z.; Dvir, O.; Pesis, E.; Pick, U.; Lichter, A. Weak organic acids and inhibitors of pH homeostasis suppress growth of Penicillium infesting litchi fruits. J phytopathol 2005, 153, 667–673. [Google Scholar] [CrossRef]
- Olmedo, G.M.; Debes, M.A.; Sepúlveda, M.; Ramallo, J.; Rapisarda, V.A.; Cerioni, L.; Volentini, S.I. Overcoming lemon postharvest molds caused by Penicillium spp. multiresistant isolates by the application of potassium sorbate in aqueous and wax treatments. J Food Sci 2023. [Google Scholar] [CrossRef] [PubMed]
- Lydakis, D.; Aked, J. Vapour heat treatment of Sultanina table grapes. I: control of Botrytis cinerea. Postharv Biol Technol 2003, 27, 109–116. [Google Scholar] [CrossRef]
- Palou, L.; Smilanick, J. L.; Droby, S. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharv Rev 2008, 2, 1–16. [Google Scholar]
- Martínez-Blay, V.; Taberner, V.; Pérez-Gago, M.B.; Palou, L. Postharvest Treatments with Sulfur-Containing Food Additives to Control Major Fungal Pathogens of Stone Fruits. Foods 2021, 10, 2115. [Google Scholar] [CrossRef] [PubMed]
- Delisle-Houde, M.; Affia, H.; R.J., Tweddell. Evaluation of different salts for the control of lettuce varnish spot: when phytotoxicity rules. J Plant Sci 2018, 753–761. [Google Scholar] [CrossRef]
Figure 1.
Importance of wound type allowing a better growth of B. cinerea on apple fruit.

Figure 2.
Percentage inhibition of mycelial growth of B. cinerea, A. alternata, P. digitatum and P. italicum on PDA amended with one of the four 0.2% GRAS salts, incubated at 20 °C. Bars with standard error are averages of three replicates. SMB: sodium metabisulphite, AMB: ammonium bicarbonate, SB: sodium bicarbonate PDO: potassium dihydrogen orthophosphate. For B. cinerea, mycelial growth was determined after 4 days, for P. digitatum after 7 days and for A. alternata and P.italicum after 9 days.
Figure 2.
Percentage inhibition of mycelial growth of B. cinerea, A. alternata, P. digitatum and P. italicum on PDA amended with one of the four 0.2% GRAS salts, incubated at 20 °C. Bars with standard error are averages of three replicates. SMB: sodium metabisulphite, AMB: ammonium bicarbonate, SB: sodium bicarbonate PDO: potassium dihydrogen orthophosphate. For B. cinerea, mycelial growth was determined after 4 days, for P. digitatum after 7 days and for A. alternata and P.italicum after 9 days.
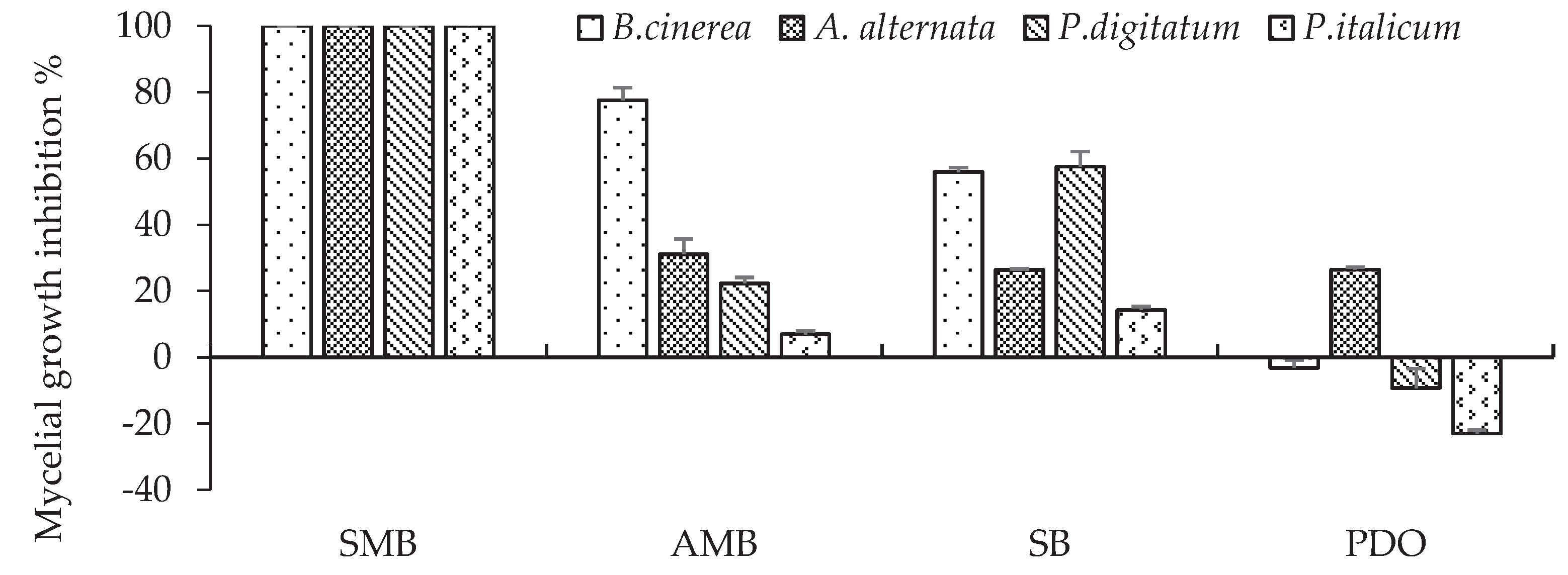
Figure 3.
Percentage of rot inhibition by sodium metabisulfite treatment (1-3 %) of 'Golden' apples inoculated with Botrytis cinerea. Fruits were dipped preventively or curatively in different concentrations of sodium metabisulphite and then incubated at room temperature for 7 days. The bars represent the standard error on the mean of three replicates. Means with different letters are significantly different at p ≤ 0.05. ANOVA summary: F ratios: Treatment (152.9**), SMB concentration (5.6*) and interaction Treatment × SMB concentration (22.4**).
Figure 3.
Percentage of rot inhibition by sodium metabisulfite treatment (1-3 %) of 'Golden' apples inoculated with Botrytis cinerea. Fruits were dipped preventively or curatively in different concentrations of sodium metabisulphite and then incubated at room temperature for 7 days. The bars represent the standard error on the mean of three replicates. Means with different letters are significantly different at p ≤ 0.05. ANOVA summary: F ratios: Treatment (152.9**), SMB concentration (5.6*) and interaction Treatment × SMB concentration (22.4**).
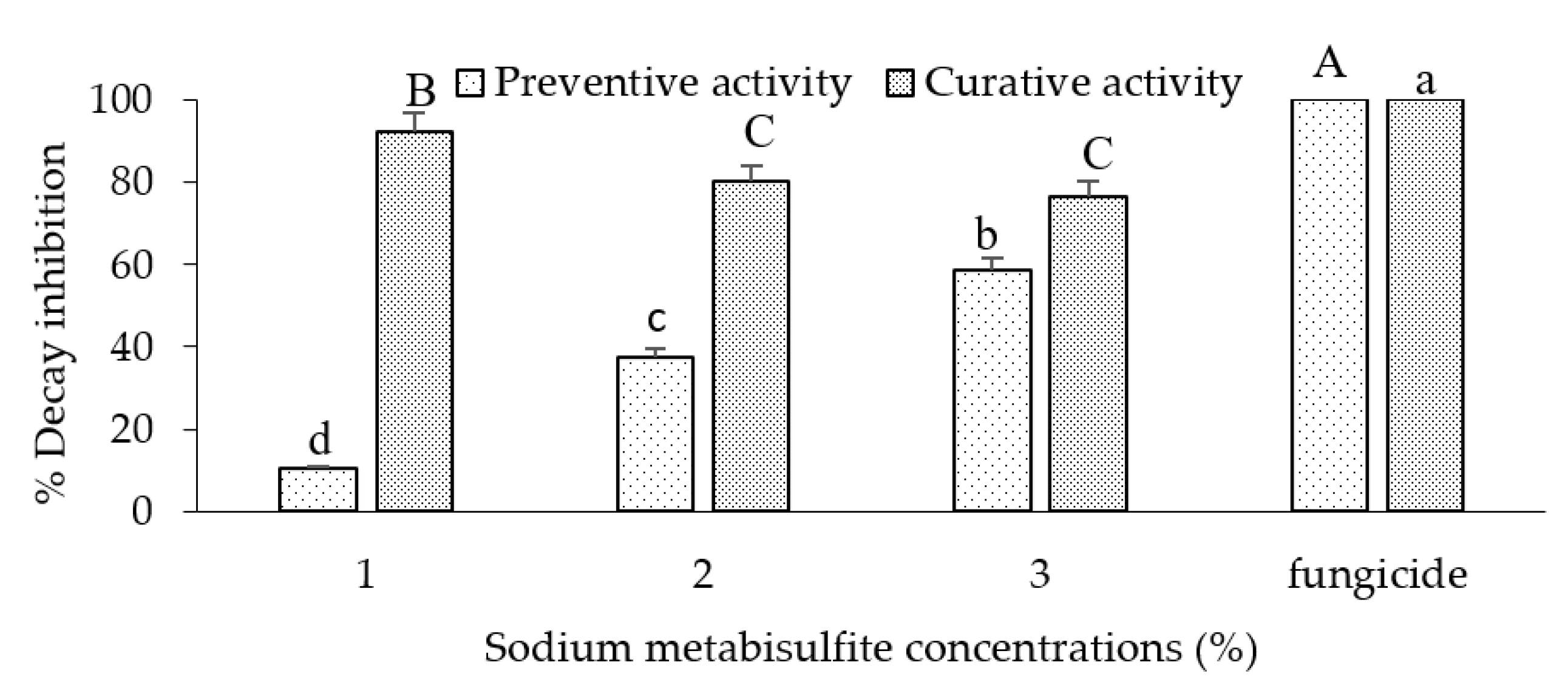
Figure 4.
Percentage of rot inhibition in apple fruit var. 'Golden' inoculated with B. cinerea and curatively dipped in different concentrations (0.2-1%) of sodium metabisulphite then incubated at room temperature for 7 d. Bars represent the standard error on the mean of three replicates. Means with different letters are significantly different at p ≤ 0.05. ANOVA summary: F-ratio of SMB concentrations (14.23*).
Figure 4.
Percentage of rot inhibition in apple fruit var. 'Golden' inoculated with B. cinerea and curatively dipped in different concentrations (0.2-1%) of sodium metabisulphite then incubated at room temperature for 7 d. Bars represent the standard error on the mean of three replicates. Means with different letters are significantly different at p ≤ 0.05. ANOVA summary: F-ratio of SMB concentrations (14.23*).
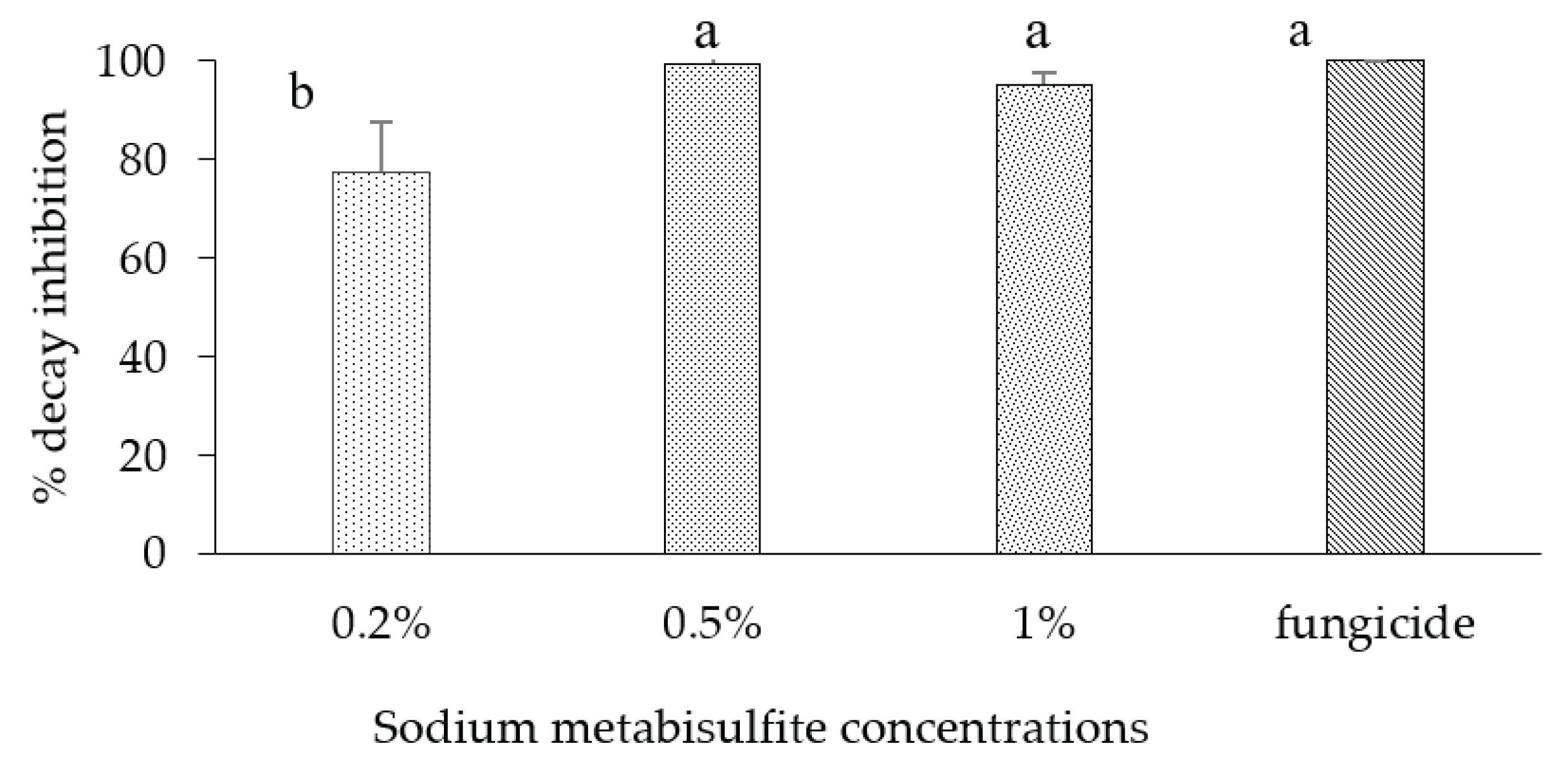
Figure 5.
Phytotoxicity (as rot size) of sodium metabisulphite in relation to increasing concentrations from 0.5 to 4% on apples inoculated/un-inoculated with B. cinerea (left) and on oranges inoculated/un-inoculated with P. digitatum (right). ANOVA Sumary of the phtytoxicity in apple fruit: Factor SMB concentration: F value = (11.6*), Factor pathogen: F value = (2.6 ns), Interaction SMB concentration×pathogen: F value = (2.6)*. ANOVA Sumary of the phtytoxicity in orange fruit: Factor SMB concentration: F value = (22.4*), Factor pathogen: F value = (0.2 ns), Interaction SMB concentration×pathogen: F value = (0.6 ns). * Significance at p ≤ 0.05, ns not significant.
Figure 5.
Phytotoxicity (as rot size) of sodium metabisulphite in relation to increasing concentrations from 0.5 to 4% on apples inoculated/un-inoculated with B. cinerea (left) and on oranges inoculated/un-inoculated with P. digitatum (right). ANOVA Sumary of the phtytoxicity in apple fruit: Factor SMB concentration: F value = (11.6*), Factor pathogen: F value = (2.6 ns), Interaction SMB concentration×pathogen: F value = (2.6)*. ANOVA Sumary of the phtytoxicity in orange fruit: Factor SMB concentration: F value = (22.4*), Factor pathogen: F value = (0.2 ns), Interaction SMB concentration×pathogen: F value = (0.6 ns). * Significance at p ≤ 0.05, ns not significant.

Figure 6.
Linear regression between sodium metabisulphite concentration (x) and rot diameter size (y) measured on oranges (6 dpi) and apples (8 dpi). Each point is the mean diameter of eight replicates (fruit). Vertical bars represent standard deviations.
Figure 6.
Linear regression between sodium metabisulphite concentration (x) and rot diameter size (y) measured on oranges (6 dpi) and apples (8 dpi). Each point is the mean diameter of eight replicates (fruit). Vertical bars represent standard deviations.
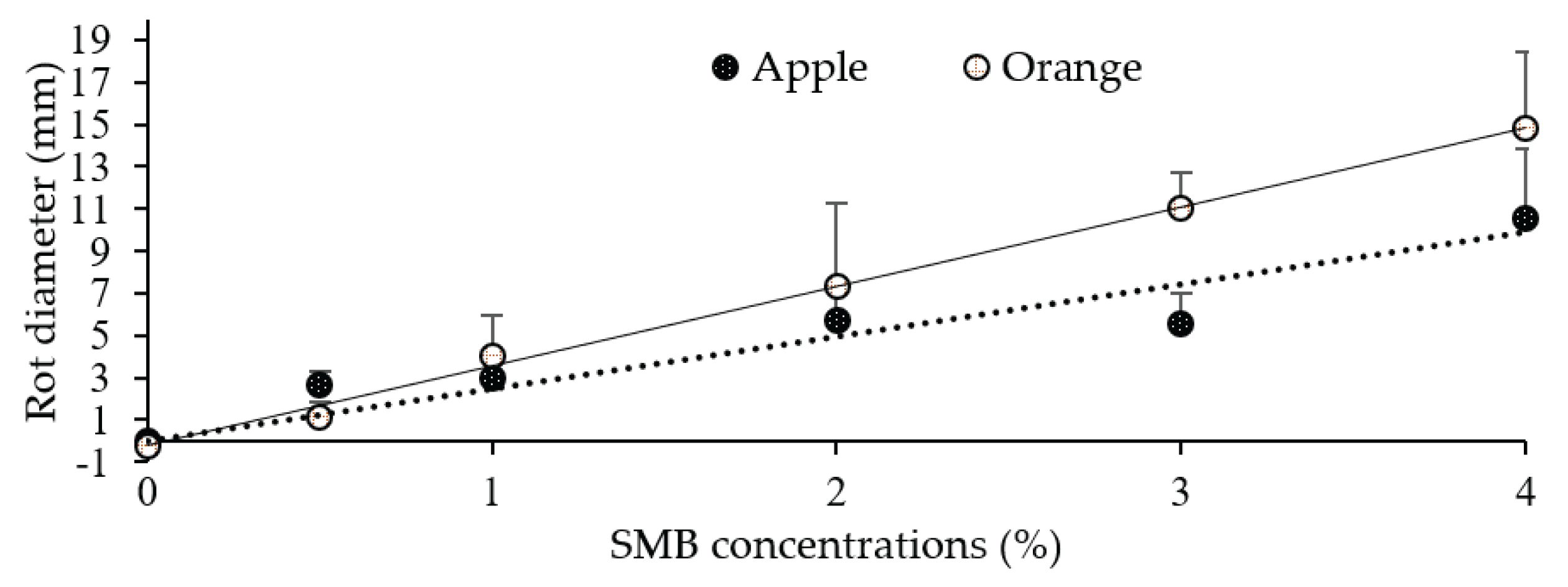
Figure 7.
Evolution of the phytotoxic reaction to SMB in the form of a circular necrosis around the wound on 'Golden' apples and 'Thompson' citrus fruit treated with 7 % and 10 % SMB and incubated at 18 °C for 6 days.
Figure 7.
Evolution of the phytotoxic reaction to SMB in the form of a circular necrosis around the wound on 'Golden' apples and 'Thompson' citrus fruit treated with 7 % and 10 % SMB and incubated at 18 °C for 6 days.


Table 1.
Analysis of variance in the effect of GRAS salts on mycelial inhibition of various fungi.
| Source of variation | DF | F-value |
| Factor A (GRAS salts) | 3 | 230.4** |
| Factor B (fungi) | 3 | 18.2* |
| Interaction A×B | 9 | 10.6* |
*Significant at p ≤ 0.05, GRAS salts were sodium metabisulfite, ammonium bicarbonate, sodium bicarbonate and potassium dihydrogen orthophosphate. The pathogens were A. alternata, B. cinerea, P. digitatum and P. italicum.
Table 2.
Evaluation of the phytotoxicity of SMB by the size of rots on oranges and apples treated with increasing SMB concentrations.
Table 2.
Evaluation of the phytotoxicity of SMB by the size of rots on oranges and apples treated with increasing SMB concentrations.
| SMB concentrations % (x) | 0.5 | 1 | 2 | 3 | 4 |
| Oranges (cv. ‘Maltaise’) | |||||
| Decay inhibition (%)1 | 98.7 | 94.1 | 88.0 | 84.8 | 78.2 |
| A (mm)= ɸSMBI - ɸC- | 0.9 ± 0. 1 | 4 ± 1.4 | 8.2 ± 5.6 | 10.4 ± 1.1 | 14.9 ± 3.5 |
| B (mm) = ɸSMB - ɸC- | 1.3 ± 0.3 | 4 ± 2.1 | 6.4 ± 2.3 | 11.7 ± 1.5 | 9.2 ± 2.9 |
| Fungal effect (A-B) (mm) | -0.4 | 0 | 1.8 | -1.3 | 5.7 |
| (A+B) /2, [y observed] (mm) | 1.1 | 4 | 7.3 | 11.05 | 12.05 |
| y = a. x +b, [y calculated] | y = 3.76.x - 0.2 ; R2 = 0.99 | ||||
| Apples (cv. ‘Golden’) | |||||
| Decay inhibition (%)1 | 93.2 | 91.6 | 84.7 | 87.1 | 71.7 |
| A (mm)= ɸSMBI - ɸC- | 2.5 ± 0.3 | 2.5 ± 0.6 | 5.1 ± 1.6 | 5.8 ± 1.3 | 9.5± 2.3 |
| B (mm) = ɸSMB - ɸC- | 2.8 ± 0.9 | 3.5 ± 1.7 | 6.3 ± 1.4 | 5.3 ± 1.6 | 11.7 ± 4.2 |
| Fungal effect (A-B) (mm) | 0.3 | 1.0 | 1.3 | -0.5 | 2.2 |
| (A+B) /2, [y observed] (mm) | 2.7 | 3.0 | 5.7 | 5.6 | 10.6 |
| y = a. x +b, [y calculated] | y = 2.47.x ; R2 = 0.96 | ||||
Decay inhibition (%) = [1- (ɸSMBI / ɸc+)] ×100; ɸSMBI: Decay diameter of inoculated treated oranges or apples; ɸSMB: Decay diameter of un-inoculated treated fruit;ɸc+: Positive control (diameter of untreated, inoculated wounded fruitdiameter); ɸc-:Negative control (diameter of untreated, un-inoculated wounded fruit). .
Table 3.
Mean values of rot diameter (mm) measured and those expected from the respective linear regressions of apples and oranges in relation to SMB concentrations and chi-square test comparing the critical and calculated chi-square values.
Table 3.
Mean values of rot diameter (mm) measured and those expected from the respective linear regressions of apples and oranges in relation to SMB concentrations and chi-square test comparing the critical and calculated chi-square values.
| SMB (%) | Observed values | Expected values | Chi-square test | |
| Oranges cv. ‘Thompson’ |
7 | 26.02 | 26.12 | χ2c = 0.809 χ2 (0.05, 1) = 3.84 |
| 10 | 31.9 | 37.4 | ||
| Apples cv. ‘Golden’ | 7 | 16.7 | 17.3 | χ2 c = 0.23 χ2 (0.05, 1) = 3.84 |
| 10 | 22.4 | 24.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



