
Submitted:
20 March 2024
Posted:
20 March 2024
You are already at the latest version
Abstract
According to previous studies the median raphe region (MRR) is known to contribute significantly to social behavior. Beside serotonin, there are reports of a small population of dopaminergic neu-rons in this region. Dopamine is linked to reward and locomotion, but very little has been known about its role in the MRR. To address that, we first confirmed the presence of dopaminergic cells in the MRR of mice (immunohistochemistry, RT-PCR) and humans (RT-PCR). Next, we used chemogenetic technology in mice containing Cre enzyme under the promoter of the dopamine transporter (DAT). With the help of an adeno-associated virus, artificial receptors were expressed in the dopaminergic cells of the MRR (DAT-MRR). Four weeks later, extensive behavioral charac-terization started 30 minutes after the injection of the artificial ligand (Clozapine-N-Oxide). Stimu-lation of DAT-MRR decreased social interest without influencing aggression and with an increase in social discrimination. Additionally, inhibition of the same cells increased the friendly social be-havior during social interaction test. No behavioral changes were detected in anxiety, memory and locomotion. All in all, dopaminergic cells were present both in the mice and human MRR, and manipulation of dopaminergic neurons of the MRR elicited a specific social response.
Keywords:
Dopamine
; Median Raphe Region
; Behavior
; DREADD
1. Introduction
It has been long known that mental illness can affect different behaviors resulting in social dysfunctions [1]. Psychopathologies such as schizophrenia, autism spectrum disorder and depression, are all good examples of how social relationships can be severely impacted [2,3,4]. According to current scientific evidence, environmental factors and multiple risk genes are the key features for the development of psychopathologies, causing subtle changes in the brain neurotransmission that leads ultimately to behavioral symptoms and emotional instability. One brain structure that is reportedly involved in many mental disorders is the serotoninergic median raphe nucleus.
The median raphe region (MRR, also known as superior central nucleus, B5+B8 [5]) is located in the midline of the brainstem, and it is constituted by the median and the paramedian raphe regions [6]. It is also part of the mesolimbic serotonergic pathway that projects to the septum and to the hippocampus [7,8]. This pathway has been the subject of a large number of studies, which demonstrate the involvement of MRR in anxiety [9,10,11,12,13] , social behavior [14,15,16,17], depression [18] and in the control of the circadian rhythms [19] by regulating the theta rhythms of the hippocampus [20,21].
Although the MRR is widely known as a serotoninergic nucleus, recent studies have shown that serotoninergic neurons are only a minority of the MRR (8.5% of all MRR neurons) and instead, the majority of MRR neurons are GABAergic (61%) or can be characterized by the presence of glutamatergic transporters (vesicular glutamate transporter 2 (VGluT2) and 3 (VGluT3)) [6,22]. The presence [23,24,25] and production [26,27,28,29] of dopamine (DA) in the MRR was repeatedly confirmed in rats, and its glial uptake – suggesting functional relevance- was also demonstrated [30]. It was even suggested that the hippocampus might get DAergic innervation from the MRR [31]. The presence of DA was also found in the MRR of deermice, Peromyscus maniculatus [32] and its production was shown also in chicken [33]. Nevertheless, most of the studies used tyrosine hydroxylase (TH) immunohistochemistry, which is the rate limiting enzyme of catecholamine synthesis including DA, noradrenaline and adrenaline. In rats considerable amount of noradrenaline was also detected in the MRR [23,24,25], therefore, confirmation of dopamine-β-hydroxylase (DBH, the key enzyme for noradrenaline synthesis) negativity of the TH positive cells seemed to be important [35]. Moreover, one study concentrating on the dorsal raphe (DR) did not find TH immunopositivity in the MRR of rats [34].
DA is present both in the central and peripheral nervous system, and it has been described as a key player in the regulation of a wide range of behaviors [36]. Imbalances in DA can contribute to the emergence of neurological and psychiatric disorders with disturbances in mood, locomotion and cognitive functions [37,38] such as Parkinson’s disease, schizophrenia, addiction and attention hyperactivity disorders [39,40,41,42]. Despite each condition having different etiology, most of these disorders are characterized by abnormal social behavior. Moreover, in some cases (especially in Parkinson’s disease and schizophrenia) the major target of presently available therapies is the dopaminergic system [44,45,46].
Our aim was (i) to confirm the presence of dopaminergic cells in the MRR of mice (by immunohistochemistry and RT-PCR) as well as in humans (by RT-PCR); (ii) investigate their role in social behavior by chemogenetics (virally introduced designer receptors exclusively active by designer drugs (DREADDs) activated by the synthetic ligand Clozapine-N-oxide (CNO) [47,48] in dopamine transporter-Cre (DAT-Cre) animals (Figure 1); (iii) summarize additional information on locomotion, anxiety and memory gained during the behavioral testing, as alterations of these behavior might influence the outcome of social tests.
2. Results
2.1. Presence of Dopaminergic Cells in the MRR
Double immunohistochemistry of the MRR showed the presence of TH positive cells, which were co-localized with red fluorescent protein (RFP) aimed to represent the dopaminergic cells in a DAT-Cre mice (Figure 2a–d). As described earlier [49], by using an antiserum against TH, we are able to visualize the cells which are involved in catecholamine synthesis, but we cannot discriminate dopaminergic, noradrenergic or adrenergic cells. However, DBH negativity of the TH positive cells suggest DA as end product. Indeed, the DBH-positive MRR cells of the DAT-Cre mice did not colocalize with the RFP-labelled cells (Figure 2e–h).
In a separate series we conducted triple immunohistochemistry visualizing RFP, TH and DBH on the same slides of control virus vector injected DAT-Cre animals (Figure 2i–m), which confirmed previously observed TH positivity and DBH negativity of RFP labelled cells.
Using double fluorescent immunohistochemistry against RFP and DAT we further supported the presence of DAT in these neurons (Figure 2n–q).
These results not only confirm the presence of TH positive and DBH negative dopaminergic cells in the MRR of mice but support that these cells (subsequently called DAT-MRR cells) can be effectively labelled and manipulated in the MRR region of DAT-Cre mice.
Additionally, we were able to confirm the presence of DAT positive cells both in the mouse and human brainstem by RT-PCR. In the mouse, not only DAT and TH, but also DBH mRNA expressions were found indicating that not only dopaminergic, but noradrenergic neurotransmission is also present (Figure 3a). Not only the MRR, but also the DR contained these markers. Moreover, in humans the MRR equivalent pontine raphe nucleus cells also express DAT (Figure 3b). However, the same expression was not detectable in cortical areas (Figure 3c).
2.2. Locomotion
In DAT-Cre mice manipulation of the DAT-MRR cells had no effect on locomotion in either studied parameters and tests (distance travelled in the OF, closed arm entries in EPM, total arm entries in Y-maze) (Figure 4).
2.3. Social Behavior
During the habituation phase of the sociability test (introduction of the animals to two empty boxes) there was a side preference in frequency (F (1,19) = 5.439, p = 0.030), but not in duration of sniffing (F(1,19) = 0.014, p = 0.906) and none of the treatment influenced the time spent with the objects (F(2,19) = 1.282, p = 0.300).
Introduction a juvenile mice into one of the small cages increased significantly the frequency (F(1,18)=35.155, p < 0.01) and time spent sniffing this cage (F(1,18) = 121.237, p < 0.01) (Figure 5a,b). However, the treatment had no effect on these parameters neither alone (frequency: F(2,18) = 0.287, p = 0.753; duration: F(2,18) = 1.458, p = 0.258), nor interacted with the presence of the stimulus mouse (frequency: F(2,18) = 0.356, p = 0.705; duration: F(2,18) = 0.925, p = 0.414). The index of social interest was above the 50% chance level in all studied groups, suggesting a social preference (single sample t-test: Control: t(8) = 18.006, p < 0.01; Excitatory: t(7) = 13.114, p < 0.01; Inhibitory: t(6) = 6.492, p < 0.01) (Figure 5c).
During the social discrimination test (SD; 24h after the CNO injection) an altered social interest (time spend sniffing both unfamiliar and familiar mice) was detectable (F(2,16) = 10.522, p < 0.01). More precisely, during this phase the excitatory group dealt with the stimulus animals significantly less than the control (p < 0.01) and inhibitory group (p < 0.01) (Figure 5d).
During the 10 min of the anxiogenic social interaction test (SIT) the animals initiated more friendly than aggressive encounter with each other (frequency: F(1,13) = 414.678, p < 0.01), without any influence of the treatment (F(2,13) = 1.423, p = 0.276). The main parameter, the time spent sniffing each other showed not only a significantly more time spending with friendly than aggressive behavior (F(1,13) = 61.476, p < 0.01), but also a significant interaction between the treatment and social behaviors (F(2,13) = 4.410, p = 0.034) (Figure 5e). The post hoc comparison revealed that the inhibitory group spent significantly more time with friendly social behavior than the control (p < 0.01) and excitatory group (p < 0.01) without significant differences in aggressive behavior.
During the 10 min of the resident-intruder test (RIT) the animals initiated more contact (F(1,15) = 190.106, p < 0.01) and spent significantly more time with friendly than with aggressive encounter (F(1,15) = 48.138, p < 0.01) (Figure 5f). In this case the treatment had no effect neither on social (frequency: F(2,15) = 1.134, p = 0.347; duration: F(2,15) = 0.020, p = 0.979), nor on aggressive behavior (frequency: F(2,15) = 1.042, p = 0.376; duration: F(2,15) = 0.727, p = 0.499).
2.4. Anxiety
As anxiety may influence the social behavior, especially during the social interaction test, we analyzed this parameter separately, in the elevated plus maze (EPM) test.
2.5. Memory
All animals showed intact short term memory exceeding the 50% chance level in the Y-maze test (Control: t(5)=9.313, p<0.01; Excitatory: t(7) = 5.161, p < 0.01; Inhibitory: t(6) = 5.208, p < 0.01) (Figure 6c). However, there was no significant difference between the groups (F(2,15) = 2.408, p = 0.123).
One day after the sociability test, the social recognition of a previously known mice did not show statistical significance (F(2,16) = 0.756, p = 0.485) (Figure 6d). However, DAT-MRR stimulated animals spent less time investigating the stimulus animals (F(2,16) = 10.522, p = 0.001) in compared to control and inhibitory group (p = 0.001; Figure 6e) with similar sniffing frequencies along the groups (F(2,16) = 0.429, p = 658; Figure 6f). Note that there was no direct manipulation of the DAT-MRR cells right before this test.
3. Discussion
Our results suggest that the MRR contains dopaminergic neurons (both in mice and in humans) and that these cells do not influence locomotion, anxiety and memory, however, their stimulation in mice decreased social behavior during the social discrimination test, whereas their inhibition increased the friendly social behavior in the social interaction test.
The presence of DAT in the human pontin raphe nucleus confirmed the translational value of our results. Despite the adequate RNA content (see Supplementary Table S1.) only very faint expression was observed in sample 6. It is possible that this person was in agony for a longer time than others (see Supplementary Table S2.) and his oxygen supply may have been permanently reduced due to previous pneumonia as well, which may negatively affect RNA integrity. However, it cannot be ruled out that the sampling does not always succeed with the same accuracy, which is also why there may be differences in the DAT expression of the samples. The absence of DAT mRNA in cortical areas was in line with a previous rat study [52] and can be explained by its expression in the cell bodies rather than on axon terminals.
Table 1 summarizes all the observed changes during behavioral experiments performed on DAT-Cre mice in this study.
In the tests used to measure locomotor activity of the mice (OF, closed arm entries in EPM and total arm entries in Y-maze) no difference was found between excitatory, inhibitory and control groups. According to the literature [53,54,55], DA is directly responsible for the locomotor activity. However, mainly the nigrostriatal pathway was implicated in this behavior. Additionally, a previous study has shown that injecting different drugs (GABA agonists, opioid agonists) into the MRR may lead to hyperlocomotion [56]. Not all effect was antagonized by haloperidol (D2 antagonist) injection, suggesting a DA-dependent as well as an independent MRR-related influence on locomotion [56,57]. We have to add, due to its vast projection [58], it is not easy to determine which pathway the DAT-MRR is involved. Nevertheless, in light of the current findings, we might conclude that dopaminergic cells from the MRR might have a different role than the nigrostriatal pathway.
During the habituation phase of the sociability test, there was no sign of fear from the object with any group tested. Although DA is implicated in fear, it is more connected to the extinction than the realization of the fear response [59,60] and the ventral tegmental area (VTA) is suggested as a main source [59]. However, D1 receptors of the prefrontal cortex has a role in the acquisition of contextual fear conditioning [61]. We cannot entirely close out that dopaminergic cells of the MRR may project to this area, as DA may colocalize with VGluT3 [62], and previous studies showed VGluT3 innervation from the MRR to the prefrontal region [63]. Additionally, our recent study suggested that MRR VGluT2 neurons regulate the acquisition of negative experience in mice [22]. However, it is still not yet known if there is a DA-VGluT2 interaction in the MRR. Further studies are required to address the detailed role of DA cells of the MRR in the context of fear.
In the second phase of the sociability test, the social interested was investigated, but no changes were observed among the groups. All animals displayed more interest toward the conspecific stimulus mice rather than the empty cage. In contrast to our results, Bariselli, et al. [64] described that inhibition of DA neurons decreased the sociability among conspecifics, however, they investigated the DA neurons of the VTA. In our hands during the social discrimination phase, excitation of the DAT-MRR cells resulted in a decrease in social interest toward both conspecific stimulus mice. This result corroborates with the finding of the social interaction test, where inhibition of the DAT-MRR cells increased the friendly social behavior. In contrast, Liu, et al. [65] found that the dendritic cell fact 1 (Dcf1) knockout mice displayed social interaction deficit and it could have been reverted by DA or D1 receptor agonist, suggesting that lower rather than enhanced dopaminergic tone will lead to reduced social interest. This difference might be due to the divergent role of DA on different brain areas and also the usage of different genetic techniques. Nevertheless, our results suggest that the dopaminergic neurons of the MRR might be also involved in the regulation of social behavior, an important manifestation of normal as well as pathological behavior.
In the resident intruder test, no statistically significant difference was observed between the studied groups. This might be due to the fact that social behavior and ethological aggression are regulated by slightly different mechanisms [66]. Another possible explanation is that the later experiment was carried out in dark, thus, was less distressing for the nocturnal animal. Indeed, previously it was shown that the outcome of the test is highly dependent on the anxiogenic nature of the environment. Although both tests (SIT and RIT) represent a social challenge and induce similar behaviors, but involve different contexts, thereby different levels of anxiety. For example, Haller, et al. [67] showed that the cannabinoid receptor 1 (CB1) KO mice was more aggressive during RIT, while less aggressive during SIT compared to wild-type, which was explained by the different stressfulness of the tests (e.g. familiar vs non-familiar environment; light vs dark during the test).
Although we were not able to find any anxiety-related effect in the EPM test, this test was done also in dark, where the CB1 KO animals also behaved normally [67]. However, our EPM results were in contrast to the findings of Bahi and Dreyer [68], presenting a decreased anxiety both in the OF and EPM after silencing the DAT in the nucleus accumbens. Once again, this variation is probably due to the different brain region targeted, and the environmental conditions may also influence the outcome (the mentioned study was done during the light, inactive phase of the animals). Nonetheless, given the successful history of evaluating the anxiety-like behavior in mice with the EPM test [69,70,71], our results might indicate that manipulation of DAT-MRR cells does not have a strong effect on anxiety control.
Previous studies have shown that acquisition and consolidation of memory involve dopaminergic activity [72,73]. In addition, a recent study found that a decrease in dopaminergic cells might be a neurocognitive signature of Alzheimer’s disease [74]. All these studies suggest a close relationship between DA and memory. We assessed working memory by Y-Maze and social memory by SD without treatment effects. In the SD test, the test mice acted as if they have never met the familiar mice. This result corroborates with the notion that social memory only lasts a few hours under laboratory conditions [75,76], although it has been reported that vasopressin release as well as group housing could prolong this effect [76,77,78]. These results might indicate that DAT-MRR is not involved in either working or social memory. However, it might have a role in spatial memory. Indeed, injection of a D1/D5 antagonist into the CA1 region of the hippocampus decreased the SD abilities at 24h [79]. As VGluT3-MRR cells projecting to the hippocampus [63] may be co-localized with DA, they might provide the source of DA in the hippocampus. Although, these assumptions need further confirmation.
Our experiments have certain limitations that need to be addressed in future studies. (i) We tried to use the most optimal tests to measure each behavior, but in order to have a general view we used a long test battery and repeated CNO injections. However, our previous optogenetic manipulation showed that a single 5 min stimulation of the MRR may induce long lasting, plastic changes in the animal’s behavior [80]. (ii) Although one might expect that the excitatory and inhibitory groups would behave in opposite ways, our results did not confirm this assumption. Nevertheless, it is not that surprising as the two types of DREADD sequences affect different signaling pathways (Gq and Gi). Moreover, stimulation seems to be a more active process, while inhibition mostly diminishes the effect of other stimulatory signals.
Our results further extend our understanding regarding the role of dopaminergic cells in our brain and raise some questions that could be addressed in future research. This is utmost important as many people worldwide take drugs influencing their dopaminergic system [81] such as: L-DOPA for treatment of Parkinson’s disease, [82], Aripiprazole for the treatment of schizophrenia and bipolar disorder [83,84], Tetrabenazine for the treatment of Huntington’s Disease [85] and Pramipexole for the treatment of Restless legs syndrome [86] and essential tremor [87], which might have unwanted behavioral side effects leading to discontinuation of the pharmacotherapy. We focused on social behavior as it is a fundamental property of every day interactions and serves as the basis for survival and reproduction [88]. Understanding the mechanisms behind this behavior could help scientists to provide a better treatment for those who suffer from psychological and psychiatric disorders.
4. Materials and Methods
4.1. Animals
Adult male mice on C57BL/6J background containing Cre enzyme under DAT [43] promoter were obtained from The Jackson Laboratory (USA; Stock No.: 006660), and local colonies were maintained at the Institute of Experimental Medicine, Budapest, Hungary. The experimental animals were all heterozygous for the Cre enzyme (see reduced DAT expression in homozygous mice [89]) and were from C57BL/6J mothers to avoid any maternal impact. The mice were group housed (2–3 mice/cage) till the beginning of the behavioral examination, when individual housing began to enhance social interest. Male juvenile (30–45-day-old) C57BL/6J mice were used as stimulus animals for the sociability and resident intruder test as previously suggested [90,91]. For RT-PCR C57BL/6J adult male mice were used from the same local colony. The animals were kept in standard environment (21 ± 1 °C, 12 h light/dark cycle with light on at 7pm) and had access to food (standard laboratory chow; Charles River, Hungary) and water ad libitum. The behavioral examinations were at the beginning of dark, active phase of the animals under red light, as it is known that rodents are nocturnal animals [92].
The experiments were approved by the Workplace Animal Welfare Committee of Institute of Experimental Medicine and National Scientific Ethical Committee on Animal Experimentation of Hungary (PEI/001/33-4/2013, PE/EA/254-7/2019) and were performed according to the European Communities Council Directive recommendations for the care and use of laboratory animals (2010/63/EU).
4.2. Surgery
The mice were anesthetized with intraperitoneal (i.p.) injection of 0.2ml anesthetics (0.5 ml of ketamine, 0.1 ml of xylazine and 2.4 ml of physiological saline) (Medicus Partner, Hungary). Then, buprenorphine (Bupaq; 0.1 mg/kg) was given to them subcutaneously as a painkiller.
Three different virus constructs were used containing the fluorophore mCherry (referred later as red fluorescent protein, RFP) and/or DREADD sequence between two loxP loci and under the control of a neuron-specific Syn promoter. In DAT-Cre mice these vectors limit the expression of RFP and DREADD to dopaminergic cells (Figure 2) [43]. The rAAV8 / hSyn-DIO-hM3D(Gq)-mCherry (5.9 × 1012 gc/ml, Addgene #44361) construct activates, while the rAAV8 / hSyn-DIO-HM4D(Gi)-mCherry (1.9 × 1013 gc/ml, Addgene #44362) construct inhibits the cells. Controls were injected with rAAV8 / hSyn-DIO-mCherry (4.1 × 1012 gc/ml, Addgene #50459) virus vector. Twenty nl of the virus was injected via glass pipettes (tip diameter 20–30 μm) connected to a MicroSyringe Pump Controller (World Precision Instruments, Sarasota, FL, USA) into the MRR of mice fixed in a stereotaxic system (David Kopf Instruments). The coordinates were the following: AP: −4.1mm of the bregma, DV: 4.6 mm and 0 mm lateral to midline.
4.3. Behavioral Analysis
Tests were carried out between 9–13 h in a separate room of the animal facility and were recorded by digital camera (Samsung SNB 7000). The order of tests was the following: sociability, social interaction (SIT), resident intruder (RIT), elevated plus maze (EPM) and y-maze (Figure 1). In order to activate the DREADD, CNO (Tocris; 1 mg/kg dissolved in 10ml saline, injected in 0.1 ml/10 g volume) was injected 30 minutes before each behavioral test. To balance for any possible CNO-induced effects all the groups (including controls) got CNO. Data were analyzed later by computer-based event recorder H77, Noldus EthoVision or Solomon Coder by an experimenter blind to the treatment groups. Each test apparatus was cleaned with 20% ethanol, and dried prior the next animal was introduced.
4.3.1. Open Field Test (OF)
The animals were placed in a non-transparent white plastic box (40 cm × 36 cm × 19 cm) for 5 minutes under infrared light. EthoVision software was used to measure the distance traveled and the time spent in the peripheral and central zones (see [93], for further details about the protocol). The locomotor activity and anxiety-like behavior were measured.
4.3.2. Sociability Test
The sociability test directly followed the open field test without further CNO injection. It consisted of three phases: habituation, sociability, and social discrimination. In the habituation phase, two small cages were put inside the plastic box, placed on opposite sides and the test animal had 5 minutes to explore and get familiar with the cages. During the sociability phase one stimulus mouse (juvenile male C57Bl6 mice [16,94]) was put inside one of the cages. The test mice then had another 5 minutes to explore the test cage. Next social discrimination was performed 24 hours after the sociability phase without further CNO injection. During this phase we put the same stimulus mice on the opposite edge of the cage (in order to avoid side preference), and we put a new stimulus mouse under the other cage. Here a comparison was made between the time that the test mice spent with the new and old stimuli mouse. This phase also lasted 5 minutes. This test examined the lasting memory effect of the stimulation/inhibition of MRR.
The time and frequency sniffing the small cages (either empty or containing a conspecific, familiar or unfamiliar) were measured by H77 event recorder.
The sociability index (SI) and social discrimination (SD) were calculated using the Equation (1):
where tmouse: time spent sniffing the box containing a juvenile mouse; tcage: time spent sniffing the empty cage; (SD): Social discrimination; tnew: time spent with a new mouse; told: time spent with the old, familiar mouse.
4.3.3. Social Interaction Test (SIT)
The test took place in two phases, habituation and social interaction [95]. The first phase happened in the day before the social interaction phase, where the mice were put alone in a test box (new, unknow environment) for 10 minutes without CNO injection. The same box was used on the next day for the second phase. The habituation phase was done to make the animals familiar with the new environment, and to enhance social interaction. Both the unfamiliarity of the environment and the lighting during the test enhance anxiety, thereby the results of this test might be also interpreted in relation to anxiety [95].
On the second day, two test mice (same treatment) were put together for 10 minutes 30 min after an intraperitoneal injection of CNO (1 mg/10 ml/kg). During these 10 minutes the animal’s behavior were recorded. The following parameters were taken into consideration: Aggressive, Defensive, Social and Other. The aggressive behavior included any type of aggressive and abnormal behavior from the mice, e.g. biting, chasing, mating. Defensive behavior was categorized whenever the mouse was fleeing from attacks, and when they were hitting back after being attacked. The animal behaviors were labelled as social whenever one animal was sniffing the other. The “other” criteria were used when the animals were not interacting with each other. The time and frequency of these behaviors were analyzed by Solomon Coder program.
4.3.4. Resident Intruder Test (RIT)
For this experiment, the test mice (resident) were isolated in a cage for a week started on day 1 of testing, during the CNO injection. During this period the cages were not cleaned as olfactory cues help to determine the territoriality of the animal [96].
The test started when one stimulus mouse (intruder) was introduced to the cage of the test mice. For this experiment, we prioritized younger and smaller stimulus mice, as they tend to be submissive to older/bigger mice. Our intent with this was to observe only the aggression coming from the dominance of the test mice. The test was 10 minutes, and the behavior was classified in the same way as it was done during the SIT using Solomon Coder program.
4.3.5. Elevated Plus Maze Test (EPM)
The test is based on the natural explorative behavior of the rodents in new environments and widely used to assess anxiety in laboratory animals [97]. Due to the natural aversion of the mice from elevated and open places, they are expected to spend more time in the closed arm [98].
The EPM apparatus consisted of two open arms and two closed arms (with walls, open from the top; 67 cm × 7 cm × 30 cm) crossed in the middle perpendicularly with each other. We put the mice in the centrum of the EPM and let them freely explore the apparatus for 5 minutes. The number of entries into the arms and the time spent in them were measured by H77 event recorder. The number of closed arm entries reflect locomotion, while time spent in open arm is the widely accepted measure of anxiety. The number of entries into the open/(open+closed) arm * 100 is considered as a locomotor independent measure of anxiety [17,51,99,100,101].
4.3.6. Y-Maze Test
It detects the ability of short-term learning and memory of the rodent. The apparatus consists of three arms (A, B and C) at 120 degrees, connected by a central zone (CZ). On this experiment, a test mouse was put into arm, and while it explored the maze, it was expected to enter the other arms consecutively. An animal with a good short memory is able to remember the arms where it has been just before and will demonstrate a propensity to visit the one less recently visited arm (e.g. “good” alteration: ABC, BCA, CAB, ACB, CBA, BAC). On the other hand, a mouse with poor short memory, will keep entering the last visited arm [102].
The animals were tested for 5 minutes and the order of visited arms were observed on the video recordings. In order to calculate the percent (%) alternation the formula below was used (Equation (2)).
where (% Alt): Percent alternation; n”good” alterations: number of “good” alterations; narm entries: total number of arm entries.
4.4. Immunohistochemistry
Upon termination of the experiments (i.e. 6 weeks after AAV injection) mice were deeply anesthetized and transcardially perfused with 0.1M phosphate buffered saline (PBS) for 1 min, then with 4% (w/v) paraformaldehyde in PBS for 20 min. Brains were taken out, and post-fixed for 24 h in fixative at +4 °C than cryoprotected by 20% glucose-PBS solution for 24 h at +4 °C. Thirty μm-thick coronal sections were prepared on a sliding microtome.
4.4.1. Confirmation of Dopaminergic Cells in the MRR
On selected brain slices of control virus injected mice two double fluorescent immunohistochemistry was conducted. DA was visualized by positive TH and negative DBH immunohistochemistry together with the RFP immunohistochemistry visualizing the dopaminergic cells in DAT-Cre mice after control virus injection.
Primary antibodies were diluted in PBS (a-TH (host: rabbit), 1:1000, DiaSorin, Stillwater, USA or a-DBH (host: rabbit), 1:1000, PA1-18314, Pierce) and brain samples were incubated for 3 nights. After washing, sections were incubated in secondary antibody solution for 1 h (Biotin SP conj. goat a-rabbit 1:1000, Vector, CatNo: BA 1000; diluted in PBS). Slices were incubated in Extravidin-peroxidase (Lot#053M4811V, Sigma) for 1 h. After multiple washes, Fictiramin (488 nm) fluorophore was used for 10 min. Then, anti-RFP primary antibody (incubation time: 2 nights) and Alexa fluor 488 fluorescent secondary antibody (incubation time: 2 h) was used for DREADD immunolabeling. Sections were mounted with Mowiol.
In a separate set of animal 2 weeks after control virus injection TH and DBH co-positivity was studied on the same slides together with RFP expression in a triple fluorescent immunohistochemistry. On parallel sections RFP and DAT co-expression was also studied. For these staining the following antibodies were used: Primaries: 1:100 000 a-RFP, sheep, acquired from Csaba Fekete at the Institute of Experimental Medicine, Budapest, Hungary; 1:1000 a-DAT, rat, MAB369, Sigma-Aldrich; 1:300 a-TH, mouse, #45648, Cell Signalling Technology; 1:500 a-mouse Fab fragment, donkey, 715-007-003, Jackson ImmunoResearch; 1:2000 a-DBH, rabbit, EPR20385, Abcam. Secondaries: 1:500 a-sheep A594, donkey, A-11016, Invitrogen; 1:500 a-rat A488, donkey, A-21208, Invitrogen; 1:1000 biotin conjugated a-mouse, donkey, 715-065-150, Jackson ImmunoResearch; 1:500 a-rabbit A488, A-21206, Invitrogen; and 1:500 streptavidin conjugated Pacific Blue fluorescent dye, S11222, ThermoFisher Scientific. Slices were mounted in PBS and covered with glycerol.
Sections were evaluated with fluorescent C2 confocal laser-scanning microscope (Nikon Europe, 100× and 20× objective).
4.4.2. Verification of the Virus Injection
The RFP signal was enhanced by a nickel-3,3′-diaminobenzidine (Ni-DAB) immunohistochemistry carried out with a rabbit anti-RFP primary antibody and a biotinylated secondary anti-rabbit antibody coupled to an avidin-biotin complex (ABC). The RFP was labeled with a rabbit polyclonal antibody. The primary antibody (1:4000) was detected by biotinylated anti-rabbit goat serum (1:1000) and avidin–biotin complex diluted in Tris buffer (1:1000, Vectastain ABC Kit, Vector Laboratories, USA). The peroxidase reaction was developed in the presence of diaminobenzidine tetrahydrochloride (0.2mg/ml), nickel–ammonium sulphate (0.1%) and hydrogen peroxide (0.003%) dissolved in Tris buffer. The sections were mounted on glass slides and covered by a DPX mounting medium. The virus infected area was localized on micrographs by using an overlay of the stereotaxic atlas images on the series of images of the MRR [50].
If there was no staining, or it was unilateral, or other brain regions (e.g. DR) were also stained the animal and the data belonging to it were excluded from the statistical analysis (Figure 7).
4.5. Verification of the Presence of DA in the Raphe by RT-PCR
For measurements in mice, wild-type C57BL6/J animals were sacrificed, and their DR and MRR region were dissected by punch needles, then fresh froze on dry ice. Samples were kept on −80°C until further preparations for one-step PCR.
In the case of human samples, the study was approved by the Hungarian Medical Research Council—Scientific and Research Ethical Committee (Egészségügyi Tudományos Tanács—Tudományos és Kutatásetikai Bizottság, #40197-2/2019/EKU), in accordance with the Ethical Rules for Using Human Tissues for Medical Research in Hungary (HM 34/1999) and the Code of Ethics of the World Medical Association (Declaration of Helsinki). Post-mortem brain samples were obtained from the Human Brain Tissue Bank, Semmelweis University (Budapest, Hungary). The brain of twelve individuals who died of natural causes were used in the measurement. The details of the samples can be found in Supplementary Table S1 and S2.
4.5.1. RNA Extraction and cDNA Synthesis
For mouse samples, frozen samples were thawed and total RNA was processed according to RNeasy® Mini Kit instructions (QIAGEN, #74106). Reverse transcription (High-Capacity RNA-to-cDNA™ Kit, Thermo Fisher #4387406) and one-step PCR (DreamTaqTM Green PCR Master Mix, Thermo Fisher, K1085) were done by Biometra Tone (Analytik Jena). The following primers were designed using Primer-BLAST (NCBI) and were acquired from Integrated DNA Technologies: glyceraldehyde 3-phosphate dehydrogenase (GAPDH, housekeeping gene), DAT, TH and DBH (see Supplementary Table S3.).
For human samples, total RNA was isolated by using the RNeasy® Mini Kit according to the manufacturer’s instructions. RNA was diluted into RNase-free water. The quality and quantity of extracted RNA was determined using NanoDrop ND-1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and only those with A260/A280 ratio between 1.8 and 2.1 were used in subsequent experiments (Supplemetary Table S1). The isolated RNA concentration was calculated and normalized with RNase-free water and reverse transcribed into cDNA using SuperScript II reverse transcriptase kit (Invitrogen, Carlsbad, CA, USA) (see Supplementary Table S2 for a list of all samples.). After 10-fold dilution, 2.5 μL of the resulting cDNA was used as template in PCR. The PCR reactions were performed on CFX-96 C1000 Touch Real-Time System (Bio-Rad Laboratories) with iTaq DNA polymerase (Bio-Rad Laboratories, Hercules, CA, USA) in total volumes of 12.5 μL under the following conditions: 95 °C for 3 min, followed by 35 cycles of 95°C for 0.5 min, 60°C for 0.5 min, and 72 °C for 1 min. All the determinations were conducted in duplicate. The primers used for RT-PCR were synthesized by Integrated DNA Technologies, Inc. (IDT, Coralville, IA, USA) and used at 300 nM final concentration. Sequences of primers are listed in Supplementary Table S3.
4.5.2. Agarose Gel Electrophoresis
One third of the amplified PCR product was loaded in an agarose gel (1.2%) containing Eco Safe nucleic acid staining solution (1:20,000, Pacific Image Electronics) and the electrophoresis was conducted in 1× TAE (40 mM Tris-Acetate, 1 mM EDTA, pH 8.0) buffer. The applied separation voltage was 100 V. After the electrophoresis the DNA bands were visualized by UV transillumination.
4.6. Statistics
Data were analyzed with StatSoft Statistica 13.0 (Tulsa, USA) utilizing single sample t-test (Sociability Index, spontaneous alteration, Discrimination Index, in comparison to 50%), one-way analysis of variance ANOVA (parameter: treatment) or repeated measure ANOVA (Sociability, social interaction, resident-intruder and Social Discrimination) followed by Fisher LSD post hoc comparison. Data are expressed as mean ± SEM and p < 0.05 was considered statistically significant. All marks on figures represent the results of post hoc comparison, while main ANOVA effects are described in the text.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, D.Z; methodology, E.S. Z.E.T. F. D. A.D.; formal analysis, T.C. C.L.F. P.C. D.V. F.D.; investigation, T.C. B.T. C.L.F. P.C. D.V. Z.E.T. F.D. Á.D; resources, D.Z. Á.D.; writing—original draft preparation, T.C:, D.Z..; writing—review and editing, B.T. C.L.F. P.C. E.S. D.V. Z.E.T. F.D. A.D.; visualization, T.C. C.L.F. F.D.; supervision, D.Z. Z.E.T. A.D.; project administration, D.Z..; funding acquisition, D. Z.. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Brain Research Program (NAP 3.0) of the Hungarian Academy of Sciences; the National Research Development and Innovation Office of Hungary (grant numbers K141934, K138763, K134221 and K120311), as well as the Thematic Excellence Program 2021 Health Sub-program of the Ministry for Innovation and Technology in Hungary (within the framework of the TKP2021-EGA-16 project of the Pécs University as well as TKP2021-EGA-25 project of the Semmelweis University).
Institutional Review Board Statement
All tests were approved by the local committee of animal health and care (PEI/001/33-4/2013, PE/EA/254-7/2019) and performed according to the European Communities Council Directive recommendations for the care and use of laboratory animals (2010/63/EU).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available upon request.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Thompson, A.H.; Bland, R.C. Social dysfunction and mental illness in a community sample. Can J Psychiatry 1995, 40, 15–20. [Google Scholar] [CrossRef]
- Klin, A.; Volkmar, F.R.; Sparrow, S.S. Autistic social dysfunction: some limitations of the theory of mind hypothesis. J Child Psychol Psychiatry 1992, 33, 861–876. [Google Scholar] [CrossRef]
- Seivewright, H.; Tyrer, P.; Johnson, T. Persistent social dysfunction in anxious and depressed patients with personality disorder. Acta Psychiatr Scand 2004, 109, 104–109. [Google Scholar] [CrossRef]
- Stanghellini, G.; Ballerini, M. Dis-sociality: the phenomenological approach to social dysfunction in schizophrenia. World Psychiatry 2002, 1, 102–106. [Google Scholar] [PubMed]
- JP, H. The Neuronatomy of the Serotonergic System. Handbook of Behavioral Neuroscience 2010, 21, 51–64. [Google Scholar]
- Sos, K.E.; Mayer, M.I.; Cserep, C.; Takacs, F.S.; Szonyi, A.; Freund, T.F.; Nyiri, G. Cellular architecture and transmitter phenotypes of neurons of the mouse median raphe region. Brain Struct Funct 2017, 222, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G. Ascending serotonin neuron diversity under two umbrellas. Brain Struct Funct 2016, 221, 3347–3360. [Google Scholar] [CrossRef] [PubMed]
- Geyer, M.A.; Braff, D.L. Startle habituation and sensorimotor gating in schizophrenia and related animal models. Schizophr Bull 1987, 13, 643–668. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, Y.; Tanaka, K.F.; Tsunematsu, T.; Yamanaka, A.; Yoshioka, M. Optogenetic activation of serotonergic neurons enhances anxiety-like behaviour in mice. Int J Neuropsychopharmacol 2014, 17, 1777–1783. [Google Scholar] [CrossRef]
- Graeff, F.G.; Guimaraes, F.S.; De Andrade, T.G.; Deakin, J.F. Role of 5-HT in stress, anxiety, and depression. Pharmacol Biochem Behav 1996, 54, 129–141. [Google Scholar] [CrossRef]
- Dos Santos, L.; de Andrade, T.G.; Zangrossi, H. Jr. Serotonergic neurons in the median raphe nucleus regulate inhibitory avoidance but not escape behavior in the rat elevated T-maze test of anxiety. Psychopharmacology (Berl) 2005, 179, 733–741. [Google Scholar] [CrossRef]
- Andrews, N.; File, S.E.; Fernandes, C.; Gonzalez, L.E.; Barnes, N.M. Evidence that the median raphe nucleus--dorsal hippocampal pathway mediates diazepam withdrawal-induced anxiety. Psychopharmacology (Berl) 1997, 130, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.G.; Zangrossi, H. Jr.; Graeff, F.G. The median raphe nucleus in anxiety revisited. J Psychopharmacol 2013, 27, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, Y.; Tsutsui-Kimura, I.; Sasamori, H.; Nebuka, M.; Nishitani, N.; Tanaka, K.F.; Yamanaka, A.; Yoshioka, M. Different roles of distinct serotonergic pathways in anxiety-like behavior, antidepressant-like, and anti-impulsive effects. Neuropharmacology 2020, 167, 107703. [Google Scholar] [CrossRef] [PubMed]
- Teissier, A.; Chemiakine, A.; Inbar, B.; Bagchi, S.; Ray, R.S.; Palmiter, R.D.; Dymecki, S.M.; Moore, H.; Ansorge, M.S. Activity of Raphe Serotonergic Neurons Controls Emotional Behaviors. Cell Rep 2015, 13, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, C.L.; Bellardie, M.; Torok, B.; Sipos, E.; Toth, B.; Baranyi, M.; Sperlagh, B.; Dobos-Kovacs, M.; Chaillou, E.; Zelena, D. Pharmacogenetic excitation of the median raphe region affects social and depressive-like behavior and core body temperature in male mice. Life Sci 2021, 286, 120037. [Google Scholar] [CrossRef] [PubMed]
- Chaves, T.; Torok, B.; Fazekas, C.L.; Correia, P.; Sipos, E.; Varkonyi, D.; Hellinger, A.; Erk, D.; Zelena, D. Median raphe region GABAergic neurons contribute to social interest in mouse. Life Sci 2022, 289, 120223. [Google Scholar] [CrossRef]
- Numasawa, Y.; Hattori, T.; Ishiai, S.; Kobayashi, Z.; Kamata, T.; Kotera, M.; Ishibashi, S.; Sanjo, N.; Mizusawa, H.; Yokota, T. Depressive disorder may be associated with raphe nuclei lesions in patients with brainstem infarction. J Affect Disord 2017, 213, 191–198. [Google Scholar] [CrossRef]
- Morin, L.P. Neuroanatomy of the extended circadian rhythm system. Exp Neurol 2013, 243, 4–20. [Google Scholar] [CrossRef]
- Sorman, E.; Wang, D.; Hajos, M.; Kocsis, B. Control of hippocampal theta rhythm by serotonin: role of 5-HT2c receptors. Neuropharmacology 2011, 61, 489–494. [Google Scholar] [CrossRef]
- Jackson, J.; Dickson, C.T.; Bland, B.H. Median raphe stimulation disrupts hippocampal theta via rapid inhibition and state-dependent phase reset of theta-related neural circuitry. J Neurophysiol 2008, 99, 3009–3026. [Google Scholar] [CrossRef]
- Szonyi, A.; Zicho, K.; Barth, A.M.; Gonczi, R.T.; Schlingloff, D.; Torok, B.; Sipos, E.; Major, A.; Bardoczi, Z.; Sos, K.E.; Gulyas, A.I.; Varga, V.; Zelena, D.; Freund, T.F.; Nyiri, G. Median raphe controls acquisition of negative experience in the mouse. Science 2019, 366, 6469. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, J.M.; Grobecker, H.; Zivin, J. Catecholamines in the raphe nuclei of the rat. Brain Res 1976, 114, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Loullis, C.C.; Felten, D.L.; Shea, P.A. HPLC determination of biogenic amines in discrete brain areas in food deprived rats. Pharmacol Biochem Behav 1979, 11, 89–93. [Google Scholar] [CrossRef]
- Zangen, A.; Overstreet, D.H.; Yadid, G. Increased catecholamine levels in specific brain regions of a rat model of depression: normalization by chronic antidepressant treatment. Brain Res 1999, 824, 243–250. [Google Scholar] [CrossRef]
- Trulson, M.E.; Cannon, M.S.; Raese, J.D. Identification of dopamine-containing cell bodies in the dorsal and median raphe nuclei of the rat brain using tyrosine hydroxylase immunochemistry. Brain Res Bull 1985, 15, 229–234. [Google Scholar] [CrossRef]
- Ochi, J.; Shimizu, K. Occurrence of dopamine-containing neurons in the midbrain raphe nuclei of the rat. Neurosci Lett 1978, 8, 317–320. [Google Scholar] [CrossRef]
- Jahanshahi, A.; Steinbusch, H.W.; Temel, Y. Distribution of dopaminergic cell bodies in the median raphe nucleus of the rat brain. J Chem Neuroanat 2013, 53, 60–63. [Google Scholar] [CrossRef]
- Kocabicak, E.; Jahanshahi, A.; Schonfeld, L.; Hescham, S.A.; Temel, Y.; Tan, S. Deep Brain Stimulation of the Rat Subthalamic Nucleus Induced Inhibition of Median Raphe Serotonergic and Dopaminergic Neurotransmission. Turk Neurosurg 2015, 25, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Liesi, P.; Paetau, A.; Rechardt, L.; Dahl, D. Glial uptake of monoamines in primary cultures of rat median raphe nucleus and cerebellum. A combined monoamine fluorescence and glial fibrillary acidic protein immunofluorescence study. Histochemistry 1981, 73, 239–250. [Google Scholar] [CrossRef]
- Reymann, K.; Pohle, W.; Muller-Welde, P.; Ott, T. Dopaminergic innervation of the hippocampus: evidence for midbrain raphe neurons as the site of origin. Biomed Biochim Acta 1983, 42, 1247–1255. [Google Scholar]
- Lin, L.H.; Pivorun, E.B. Analysis of serotonin, dopamine and their metabolites in the caudate putamen, the suprachiasmatic nucleus and the median raphe nucleus of euthermic and torpid deermice, Peromyscus maniculatus. Pharmacol Biochem Behav 1989, 33, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Aoki, N.; Mori, C.; Serizawa, S.; Kihara-Negishi, F.; Homma, K.J.; Yamaguchi, S. Dopaminergic nuclei in the chick midbrain express serotonin receptor subfamily genes. Front Physiol 2022, 13, 1030621. [Google Scholar] [CrossRef]
- Stratford, T.R.; Wirtshafter, D. Ascending dopaminergic projections from the dorsal raphe nucleus in the rat. Brain Res 1990, 511, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Nevue, A.A.; Elde, C.J.; Perkel, D.J.; Portfors, C.V. Dopaminergic Input to the Inferior Colliculus in Mice. Front Neuroanat 2015, 9, 168. [Google Scholar] [CrossRef]
- Le Moal, M.; Simon, H. Mesocorticolimbic dopaminergic network: functional and regulatory roles. Physiol Rev 1991, 71, 155–234. [Google Scholar] [CrossRef]
- Creese, I.; Iversen, S.D. The role of forebrain dopamine systems in amphetamine induced stereotyped behavior in the rat. Psychopharmacologia 1974, 39, 345–357. [Google Scholar] [CrossRef]
- Ryan, J.M. Pharmacologic approach to aggression in neuropsychiatric disorders. Semin Clin Neuropsychiatry 2000, 5, 238–249. [Google Scholar] [CrossRef]
- Association, A.P. Diagnostic and Statistical Manual of Mental Disorders. 5th ed.; Washington, DC, 2013.
- Davis, K.L.; Kahn, R.S.; Ko, G.; Davidson, M. Dopamine in schizophrenia: a review and reconceptualization. Am J Psychiatry 1991, 148, 1474–1486. [Google Scholar] [PubMed]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Tomasi, D.; Telang, F. Addiction: beyond dopamine reward circuitry. Proc Natl Acad Sci U S A 2011, 108, 15037–15042. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Kollins, S.H.; Wigal, T.L.; Newcorn, J.H.; Telang, F.; Fowler, J.S.; Zhu, W.; Logan, J.; Ma, Y.; Pradhan, K.; Wong, C.; Swanson, J.M. Evaluating dopamine reward pathway in ADHD: clinical implications. JAMA 2009, 302, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.N.; Pei, Z.J.; Zhang, J.; Vlasov, Y.K.; Davis, T.; Taylor, E.; Weng, F.J.; Van Dort, J.C.; Solt, K.; Brown, N.E. The Role of Glutamatergic and Dopaminergic Neurons in the Periaqueductal Gray/Dorsal Raphe: Separating Analgesia and Anxiety. eNeuro 2019, 6, ENEURO–0018. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.T.; Zhang, Q.; Wen, S.Y.; Chen, F.F.; Zhou, C.Q. Efficacy and safety of non-ergot dopamine-receptor agonists as an adjunct to levodopa in advanced Parkinson’s disease: A network meta-analysis. Eur J Neurol 2022. [Google Scholar] [CrossRef]
- Juza, R.; Musilek, K.; Mezeiova, E.; Soukup, O.; Korabecny, J. Recent advances in dopamine D(2) receptor ligands in the treatment of neuropsychiatric disorders. Med Res Rev 2023, 43, 55–211. [Google Scholar] [CrossRef]
- Novak, G.; Seeman, M.V. Dopamine, Psychosis, and Symptom Fluctuation: A Narrative Review. Healthcare (Basel) 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci U S A 2007, 104, 5163–5168. [Google Scholar] [CrossRef]
- Roth, B.L. DREADDs for Neuroscientists. Neuron 2016, 89, 683–694. [Google Scholar] [CrossRef]
- Lewis, D.A.; Campbell, M.J.; Foote, S.L.; Goldstein, M.; Morrison, J.H. The distribution of tyrosine hydroxylase-immunoreactive fibers in primate neocortex is widespread but regionally specific. J Neurosci 1987, 7, 279–290. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J.; Franklin, K.B.J. The mouse brain in stereotaxic coordinates., 2nd ed.; Academic Press: San Diego, 2001. [Google Scholar]
- Mlynarik, M.; Zelena, D.; Bagdy, G.; Makara, G.B.; Jezova, D. Signs of attenuated depression-like behavior in vasopressin deficient Brattleboro rats. Horm Behav 2007, 51, 395–405. [Google Scholar] [CrossRef]
- Shimada, S.; Kitayama, S.; Walther, D.; Uhl, G. Dopamine transporter mRNA: dense expression in ventral midbrain neurons. Brain Res Mol Brain Res 1992, 13, 359–362. [Google Scholar] [CrossRef]
- Tran, A.H.; Tamura, R.; Uwano, T.; Kobayashi, T.; Katsuki, M.; Ono, T. Dopamine D1 receptors involved in locomotor activity and accumbens neural responses to prediction of reward associated with place. Proc Natl Acad Sci U S A 2005, 102, 2117–2122. [Google Scholar] [CrossRef] [PubMed]
- Ryczko, D.; Dubuc, R. Dopamine and the Brainstem Locomotor Networks: From Lamprey to Human. Front Neurosci 2017, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Beninger, R.J. The role of dopamine in locomotor activity and learning. Brain Res 1983, 287, 173–196. [Google Scholar] [CrossRef]
- Shim, I.; Stratford, T.R.; Wirtshafter, D. Dopamine is differentially involved in the locomotor hyperactivity produced by manipulations of opioid, GABA and glutamate receptors in the median raphe nucleus. Behav Brain Res 2014, 261, 65–70. [Google Scholar] [CrossRef]
- Wirtshafter, D.; Klitenick, M.A.; Asin, K.E. Is dopamine involved in the hyperactivity produced by injections of muscimol into the median raphe nucleus? Pharmacol Biochem Behav 1988, 30, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Vertes, R.P.; Martin, G.F. Autoradiographic analysis of ascending projections from the pontine and mesencephalic reticular formation and the median raphe nucleus in the rat. J Comp Neurol 1988, 275, 511–541. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Hernandez, X.I.; Vogel, P.; Betz, S.; Kalisch, R.; Sigurdsson, T.; Duvarci, S. Dopamine neurons drive fear extinction learning by signaling the omission of expected aversive outcomes. Elife 2018, 7. [Google Scholar] [CrossRef]
- Kalisch, R.; Gerlicher, A.M.V.; Duvarci, S. A Dopaminergic Basis for Fear Extinction. Trends Cogn Sci 2019, 23, 274–277. [Google Scholar] [CrossRef]
- Stubbendorff, C.; Hale, E.; Cassaday, H.J.; Bast, T.; Stevenson, C.W. Dopamine D1-like receptors in the dorsomedial prefrontal cortex regulate contextual fear conditioning. Psychopharmacology (Berl) 2019, 236, 1771–1782. [Google Scholar] [CrossRef]
- Fyk-Kolodziej, B.E.; Shimano, T.; Gafoor, D.; Mirza, N.; Griffith, R.D.; Gong, T.W.; Holt, A.G. Dopamine in the auditory brainstem and midbrain: co-localization with amino acid neurotransmitters and gene expression following cochlear trauma. Front Neuroanat 2015, 9, 88. [Google Scholar] [CrossRef]
- Szonyi, A.; Mayer, M.I.; Cserep, C.; Takacs, V.T.; Watanabe, M.; Freund, T.F.; Nyiri, G. The ascending median raphe projections are mainly glutamatergic in the mouse forebrain. Brain Struct Funct 2016, 221, 735–751. [Google Scholar] [CrossRef]
- Bariselli, S.; Hornberg, H.; Prevost-Solie, C.; Musardo, S.; Hatstatt-Burkle, L.; Scheiffele, P.; Bellone, C. Role of VTA dopamine neurons and neuroligin 3 in sociability traits related to nonfamiliar conspecific interaction. Nat Commun 2018, 9, 3173. [Google Scholar] [CrossRef]
- Liu, Q.; Shi, J.; Lin, R.; Wen, T. Dopamine and dopamine receptor D1 associated with decreased social interaction. Behav Brain Res 2017, 324, 51–57. [Google Scholar] [CrossRef]
- de Boer, S.F.; Olivier, B.; Veening, J.; Koolhaas, J.M. The neurobiology of offensive aggression: Revealing a modular view. Physiol Behav 2015, 146, 111–127. [Google Scholar] [CrossRef]
- Haller, J.; Varga, B.; Ledent, C.; Barna, I.; Freund, T.F. Context-dependent effects of CB1 cannabinoid gene disruption on anxiety-like and social behaviour in mice. Eur J Neurosci 2004, 19, 1906–1912. [Google Scholar] [CrossRef]
- Bahi, A.; Dreyer, J.L. Dopamine transporter (DAT) knockdown in the nucleus accumbens improves anxiety- and depression-related behaviors in adult mice. Behav Brain Res 2019, 359, 104–115. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Johnson, N.J.; Carr, J.; Hodgson, T.P. Resistance of experientially-induced changes in murine plus-maze behaviour to altered retest conditions. Behav Brain Res 1997, 86, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.J.; Dalvi, A. Anxiety, defence and the elevated plus-maze. Neurosci Biobehav Rev 1997, 21, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N. What’s wrong with my mouse? : behavioral phenotyping of transgenic and knockout mice., 2nd ed.; Wiley-Interscience: Hoboken, N.J. 2007. [Google Scholar]
- Grogan, J.; Bogacz, R.; Tsivos, D.; Whone, A.; Coulthard, E. Dopamine and Consolidation of Episodic Memory: Timing is Everything. J Cogn Neurosci 2015, 27, 2035–2050. [Google Scholar] [CrossRef]
- Guzman-Ramos, K.; Moreno-Castilla, P.; Castro-Cruz, M.; McGaugh, J.L.; Martinez-Coria, H.; LaFerla, F.M.; Bermudez-Rattoni, F. Restoration of dopamine release deficits during object recognition memory acquisition attenuates cognitive impairment in a triple transgenic mice model of Alzheimer’s disease. Learn Mem 2012, 19, 453–460. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.; Venneri, A. Volume and Connectivity of the Ventral Tegmental Area are Linked to Neurocognitive Signatures of Alzheimer’s Disease in Humans. J Alzheimers Dis 2018, 63, 167–180. [Google Scholar] [CrossRef]
- Thor, D.H.; Holloway, W.R. Social Memory of the Male Laboratory Rat. Journal of Comparative and Physiological Psychology 1982, 96, 1000–1006. [Google Scholar] [CrossRef]
- Bluthe, R.M.; Gheusi, G.; Dantzer, R. Gonadal steroids influence the involvement of arginine vasopressin in social recognition in mice. Psychoneuroendocrinology 1993, 18, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.S.; Williams Avram, S.K.; Cymerblit-Sabba, A.; Song, J.; Young, W.S. Targeted activation of the hippocampal CA2 area strongly enhances social memory. Mol Psychiatry 2016, 21, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Kogan, J.H.; Frankland, P.W.; Silva, A.J. Long-term memory underlying hippocampus-dependent social recognition in mice. Hippocampus 2000, 10, 47–56. [Google Scholar] [CrossRef]
- Garrido Zinn, C.; Clairis, N.; Silva Cavalcante, L.E.; Furini, C.R.; de Carvalho Myskiw, J.; Izquierdo, I. Major neurotransmitter systems in dorsal hippocampus and basolateral amygdala control social recognition memory. Proc Natl Acad Sci U S A 2016, 113, E4914–E4919. [Google Scholar] [CrossRef] [PubMed]
- Balazsfi, D.G.; Zelena, D.; Farkas, L.; Demeter, K.; Barna, I.; Cserep, C.; Takacs, V.T.; Nyiri, G.; Goloncser, F.; Sperlagh, B.; Freund, T.F.; Haller, J. Median raphe region stimulation alone generates remote, but not recent fear memory traces. PLoS One 2017, 12, e0181264. [Google Scholar] [CrossRef]
- Foley, P.B. Dopamine in psychiatry: a historical perspective. J Neural Transm (Vienna) 2019, 126, 473–479. [Google Scholar] [CrossRef]
- Calne, D.B.; Sandler, M. L-Dopa and Parkinsonism. Nature 1970, 226, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Stip, E.; Tourjman, V. Aripiprazole in schizophrenia and schizoaffective disorder: A review. Clin Ther 2010, 32 (Suppl. 1), S3–20. [Google Scholar] [CrossRef]
- Muneer, A. The Treatment of Adult Bipolar Disorder with Aripiprazole: A Systematic Review. Cureus 2016, 8, e562. [Google Scholar] [CrossRef] [PubMed]
- Ondo, W.G.; Tintner, R.; Thomas, M.; Jankovic, J. Tetrabenazine treatment for Huntington’s disease-associated chorea. Clin Neuropharmacol 2002, 25, 300–302. [Google Scholar] [CrossRef] [PubMed]
- Winlow, W. Pramipexole in restless legs syndrome: an evidence-based review of its effectiveness on clinical outcomes. Core Evid 2005, 1, 35–42. [Google Scholar]
- Herceg, M.; Nagy, F.; Pal, E.; Janszky, J.; Kesmarky, I.; Komoly, S.; Kovacs, N. Pramipexole may be an effective treatment option in essential tremor. Clin Neuropharmacol 2012, 35, 73–76. [Google Scholar] [CrossRef]
- Sandi, C.; Haller, J. Stress and the social brain: behavioural effects and neurobiological mechanisms. Nat Rev Neurosci 2015, 16, 290–304. [Google Scholar] [CrossRef]
- Backman, C.M.; Malik, N.; Zhang, Y.; Shan, L.; Grinberg, A.; Hoffer, B.J.; Westphal, H.; Tomac, A.C. Characterization of a mouse strain expressing Cre recombinase from the 3’ untranslated region of the dopamine transporter locus. Genesis 2006, 44, 383–390. [Google Scholar] [CrossRef]
- Perna, J.C.; Wotjak, C.T.; Stork, O.; Engelmann, M. Timing of presentation and nature of stimuli determine retroactive interference with social recognition memory in mice. Physiol Behav 2015, 143, 10–14. [Google Scholar] [CrossRef]
- Hadicke, J.; Engelmann, M. Social investigation and long-term recognition memory performance in 129S1/SvImJ and C57BL/6JOlaHsd mice and their hybrids. PLoS One 2013, 8, e54427. [Google Scholar] [CrossRef] [PubMed]
- Hut, R.A.; Van der Zee, E.A. The cholinergic system, circadian rhythmicity, and time memory. Behav Brain Res 2011, 221, 466–480. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J Vis Exp 2015, 96, e52434. [Google Scholar]
- Camats Perna, J.; Engelmann, M. Recognizing Others: Rodent’s Social Memories. Curr Top Behav Neurosci 2017, 30, 25–45. [Google Scholar] [PubMed]
- Haller, J.; Halasz, J.; Makara, G.B. Housing conditions and the anxiolytic efficacy of buspirone: the relationship between main and side effects. Behav Pharmacol 2000, 11, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Coppens, C.M.; de Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J. The resident-intruder paradigm: a standardized test for aggression, violence and social stress. J Vis Exp 2013, 77, e4367. [Google Scholar]
- Komada, M.; Takao, K.; Miyakawa, T. Elevated plus maze for mice. J Vis Exp 2008, 22. [Google Scholar]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Varkonyi, D.; Torok, B.; Sipos, E.; Fazekas, C.L.; Banrevi, K.; Correia, P.; Chaves, T.; Farkas, S.; Szabo, A.; Martinez-Bellver, S.; Hangya, B.; Zelena, D. Investigation of Anxiety- and Depressive-like Symptoms in 4- and 8-Month-Old Male Triple Transgenic Mouse Models of Alzheimer’s Disease. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Knoll, J.; Zelena, D.; Timar, J.; Baghy, K.; Mervai, Z.; Miklya, I. Synthetic enhancer compounds, besides acting on biogenic amine system, influence the glutamate transmission and stress response. Behav Brain Res 2020, 378, 112290. [Google Scholar] [CrossRef]
- Balazsfi, D.; Fodor, A.; Torok, B.; Ferenczi, S.; Kovacs, K.J.; Haller, J.; Zelena, D. Enhanced innate fear and altered stress axis regulation in VGluT3 knockout mice. Stress 2018, 21, 151–161. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-Maze for Assessment of Spatial Working and Reference Memory in Mice. Methods Mol Biol 2019, 1916, 105–111. [Google Scholar]
Figure 1.
Timeline of the behavioral test battery. Thirty minutes before all experiments (except social discrimination /SD/ test) animals were injected intraperitoneally by clozapine-N-oxide (CNO, 1 mg/10 ml/kg diluted in saline). (a) Adeno-associated viral (AAV) vectors (20 nl) containing control, stimulatory or inhibitory designer receptor exclusively activated by designer drug ( DREADD) sequences were microinjected into the median raphe region (MRR; anteroposterior: −4.1 mm; mediolateral: 0 mm; dorsoventral: −4.6 mm from Bregma) of dopamine transporter-Cre mice. For details see methods section. (b) Animals had 14 days to recover after surgery and an additional 14 days to habituate to reversed light-dark cycle. This 28-day incubation time is also enough for DREADD expression. (c) On the first test day, 5 min open field (OF), 5 min object habituation, then 5 min sociability test was conducted. (d) On Day 2, SD test was performed for 5 min to measure social memory. (e) On Day 5, social behavior was measured by social interaction test (SIT) for 10 min. (f) On Day 8 Resident-intruder test (RIT) was performed for 10 min to investigate aggressive behavior. (g) On Day 10, to study anxiety-like behavior, elevated plus maze (EPM) test was used for 5 min. (h) On Day 13, Y-maze test was conducted for 5 min to measure working memory. (i) On the 16th experimental day animals were transcardially perfused 2 h after CNO injection.
Figure 1.
Timeline of the behavioral test battery. Thirty minutes before all experiments (except social discrimination /SD/ test) animals were injected intraperitoneally by clozapine-N-oxide (CNO, 1 mg/10 ml/kg diluted in saline). (a) Adeno-associated viral (AAV) vectors (20 nl) containing control, stimulatory or inhibitory designer receptor exclusively activated by designer drug ( DREADD) sequences were microinjected into the median raphe region (MRR; anteroposterior: −4.1 mm; mediolateral: 0 mm; dorsoventral: −4.6 mm from Bregma) of dopamine transporter-Cre mice. For details see methods section. (b) Animals had 14 days to recover after surgery and an additional 14 days to habituate to reversed light-dark cycle. This 28-day incubation time is also enough for DREADD expression. (c) On the first test day, 5 min open field (OF), 5 min object habituation, then 5 min sociability test was conducted. (d) On Day 2, SD test was performed for 5 min to measure social memory. (e) On Day 5, social behavior was measured by social interaction test (SIT) for 10 min. (f) On Day 8 Resident-intruder test (RIT) was performed for 10 min to investigate aggressive behavior. (g) On Day 10, to study anxiety-like behavior, elevated plus maze (EPM) test was used for 5 min. (h) On Day 13, Y-maze test was conducted for 5 min to measure working memory. (i) On the 16th experimental day animals were transcardially perfused 2 h after CNO injection.
Figure 2.
Confirmation of the chemogenetic technique by immunohistochemistry 6 weeks after injection of rAAV8 / hSyn-DIO-mCherry (Addgene #50459) into the median raphe region (MRR) of dopamine transporter Cre (DAT-Cre) mice. Subregions (MRR and PMR /paramedian raphe/) are defined based on the Mouse Brain Atlas [50]. Position of the coronal section is indicated in each image, relative to the Bregma. (a) Fluorescent micrographs show representative MRR sections with red fluorescent protein (RFP; mCherry, the fluorophore suggesting the presence of the designer receptor exclusively activated by designer drug (DREADD); red)) and tyrosine hydroxylase (TH, rate-limiting enzyme of catecholamine synthesis; green) labelling. (b–d): Confocal laser scanning images show high colocalization between the labelling of RFP and TH in representative MRR neurons (arrows). (e) Fluorescent micrographs show representative MRR sections with RFP (red) and dopamine beta-hydroxylase (DBH, the enzyme which catalyses the conversion of dopamine to norepinephrine; green) labelling. (f–h) Confocal laser scanning images show no colocalization between the labelling of RFP and DBH. (i–m) Confocal laser scanning images of triple immunohistochemical staining of DREADD marker RPF positive, TH positive but DBH negative neurons in the MRR, indicating dopaminergic phenotype. (n–q) Confocal laser scanning images show colocalization of RFP and DAT.
Figure 2.
Confirmation of the chemogenetic technique by immunohistochemistry 6 weeks after injection of rAAV8 / hSyn-DIO-mCherry (Addgene #50459) into the median raphe region (MRR) of dopamine transporter Cre (DAT-Cre) mice. Subregions (MRR and PMR /paramedian raphe/) are defined based on the Mouse Brain Atlas [50]. Position of the coronal section is indicated in each image, relative to the Bregma. (a) Fluorescent micrographs show representative MRR sections with red fluorescent protein (RFP; mCherry, the fluorophore suggesting the presence of the designer receptor exclusively activated by designer drug (DREADD); red)) and tyrosine hydroxylase (TH, rate-limiting enzyme of catecholamine synthesis; green) labelling. (b–d): Confocal laser scanning images show high colocalization between the labelling of RFP and TH in representative MRR neurons (arrows). (e) Fluorescent micrographs show representative MRR sections with RFP (red) and dopamine beta-hydroxylase (DBH, the enzyme which catalyses the conversion of dopamine to norepinephrine; green) labelling. (f–h) Confocal laser scanning images show no colocalization between the labelling of RFP and DBH. (i–m) Confocal laser scanning images of triple immunohistochemical staining of DREADD marker RPF positive, TH positive but DBH negative neurons in the MRR, indicating dopaminergic phenotype. (n–q) Confocal laser scanning images show colocalization of RFP and DAT.
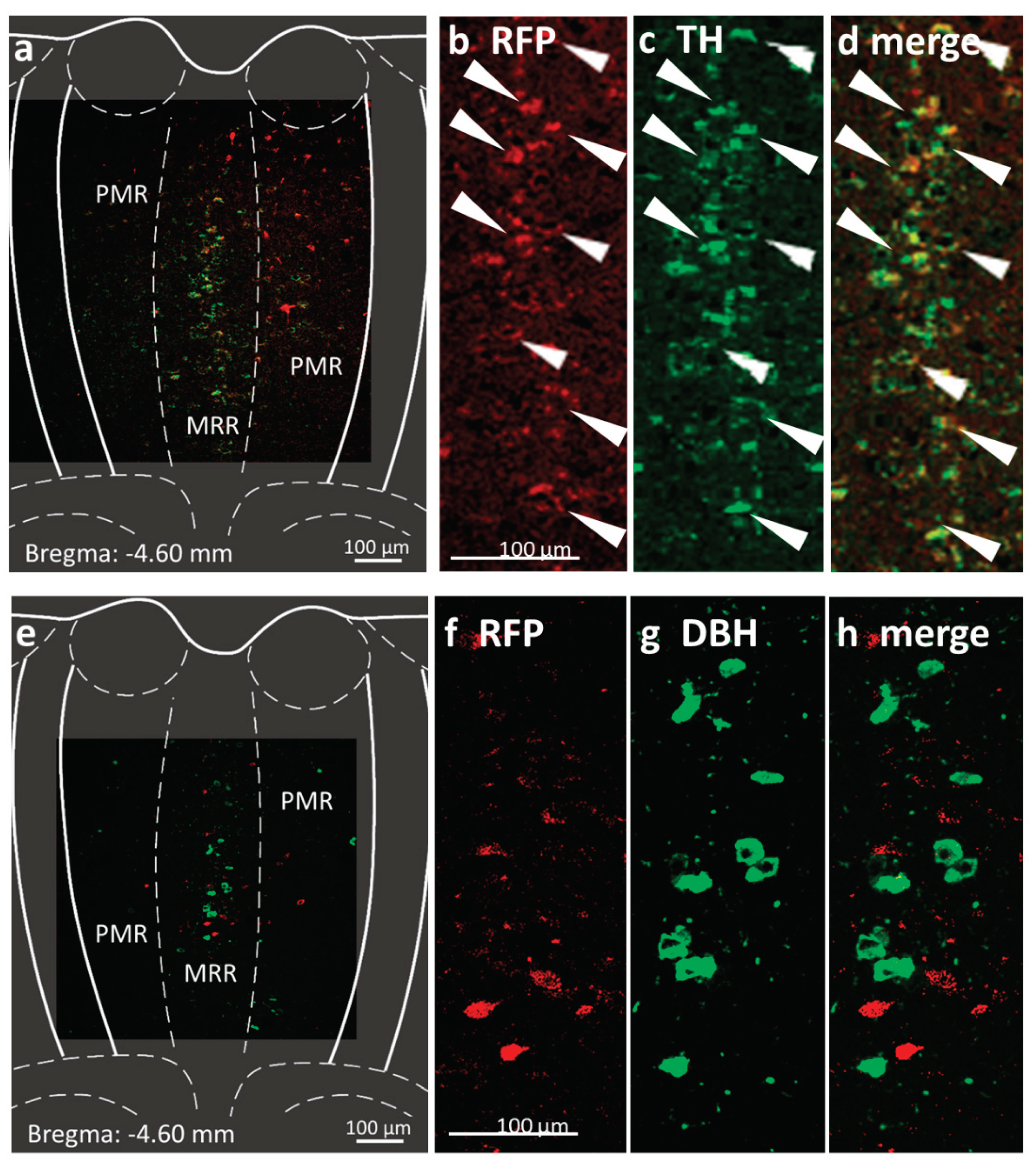
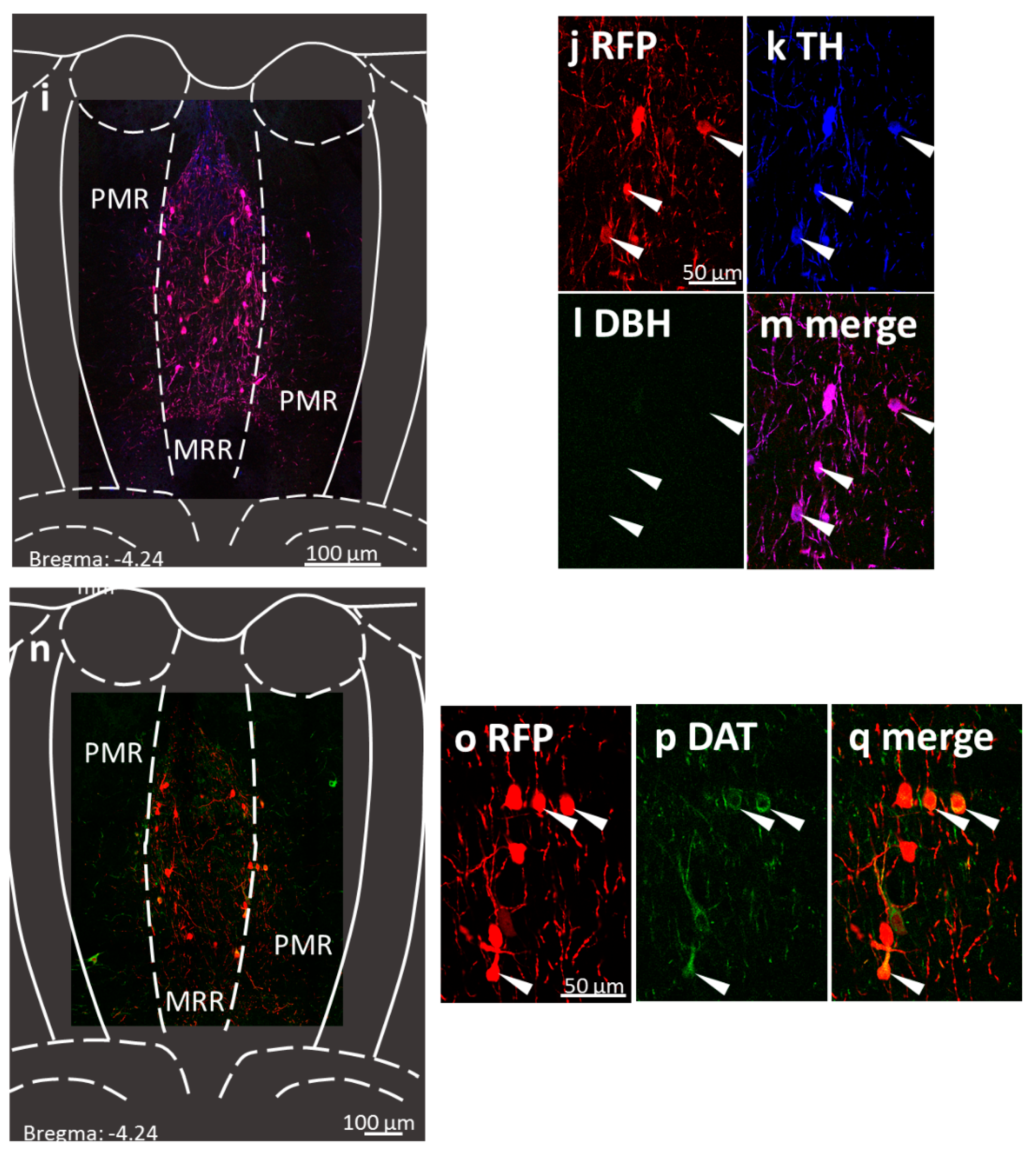
Figure 3.
Amplification products obtained by RT-PCR. (a) In the wild-type mouse (C57BL/6J, the background strain for the Cre mice used in this study), both the dorsal raphe (DR) and median raphe region (MRR) samples showed measurable mRNA expression of dopamine transporter (DAT) and tyrosine hydroxylase (TH). Interestingly, as suggested by literature, the noradrenergic marker dopamine-β-hydroxylase (DBH) was also detectable. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. (b) Samples of control human pontine raphe nucleus (which contain the human equivalent of mouse median raphe nucleus) were obtained from the Human Brain Bank (Semmelweis University, Budapest, Hungary) and ran in gel electrophoresis, for detection of DAT expression. The product displayed matched the length of DAT. (c) Negative controls from 6 cortical samples (2 from temporal cortex and 4 from frontopolar cortex). No DAT expression were observed in these two cortical areas.
Figure 3.
Amplification products obtained by RT-PCR. (a) In the wild-type mouse (C57BL/6J, the background strain for the Cre mice used in this study), both the dorsal raphe (DR) and median raphe region (MRR) samples showed measurable mRNA expression of dopamine transporter (DAT) and tyrosine hydroxylase (TH). Interestingly, as suggested by literature, the noradrenergic marker dopamine-β-hydroxylase (DBH) was also detectable. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. (b) Samples of control human pontine raphe nucleus (which contain the human equivalent of mouse median raphe nucleus) were obtained from the Human Brain Bank (Semmelweis University, Budapest, Hungary) and ran in gel electrophoresis, for detection of DAT expression. The product displayed matched the length of DAT. (c) Negative controls from 6 cortical samples (2 from temporal cortex and 4 from frontopolar cortex). No DAT expression were observed in these two cortical areas.
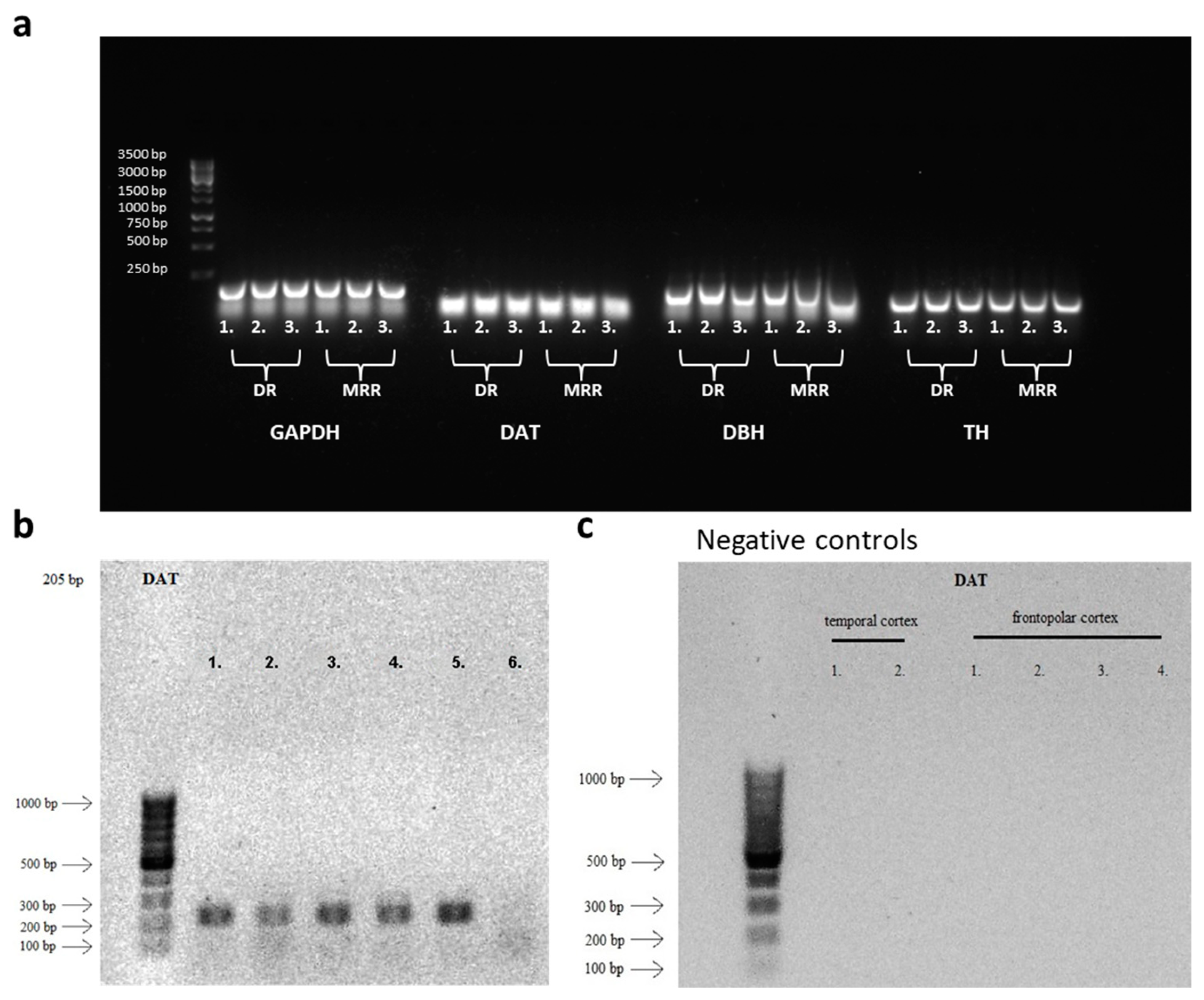
Figure 4.
Locomotion after chemogenetic manipulation of median raphe region dopaminergic cells (DAT-MRR) in male dopamine transporter Cre mice. No difference was observed between treatment groups. (a) Distance travelled in cm in the Open Field test (OF). (b) Number of closed arm entries during the 5 min elevated plus maze (EPM) test. (c) Number of entries in the arms of the Y-maze. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values.
Figure 4.
Locomotion after chemogenetic manipulation of median raphe region dopaminergic cells (DAT-MRR) in male dopamine transporter Cre mice. No difference was observed between treatment groups. (a) Distance travelled in cm in the Open Field test (OF). (b) Number of closed arm entries during the 5 min elevated plus maze (EPM) test. (c) Number of entries in the arms of the Y-maze. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values.
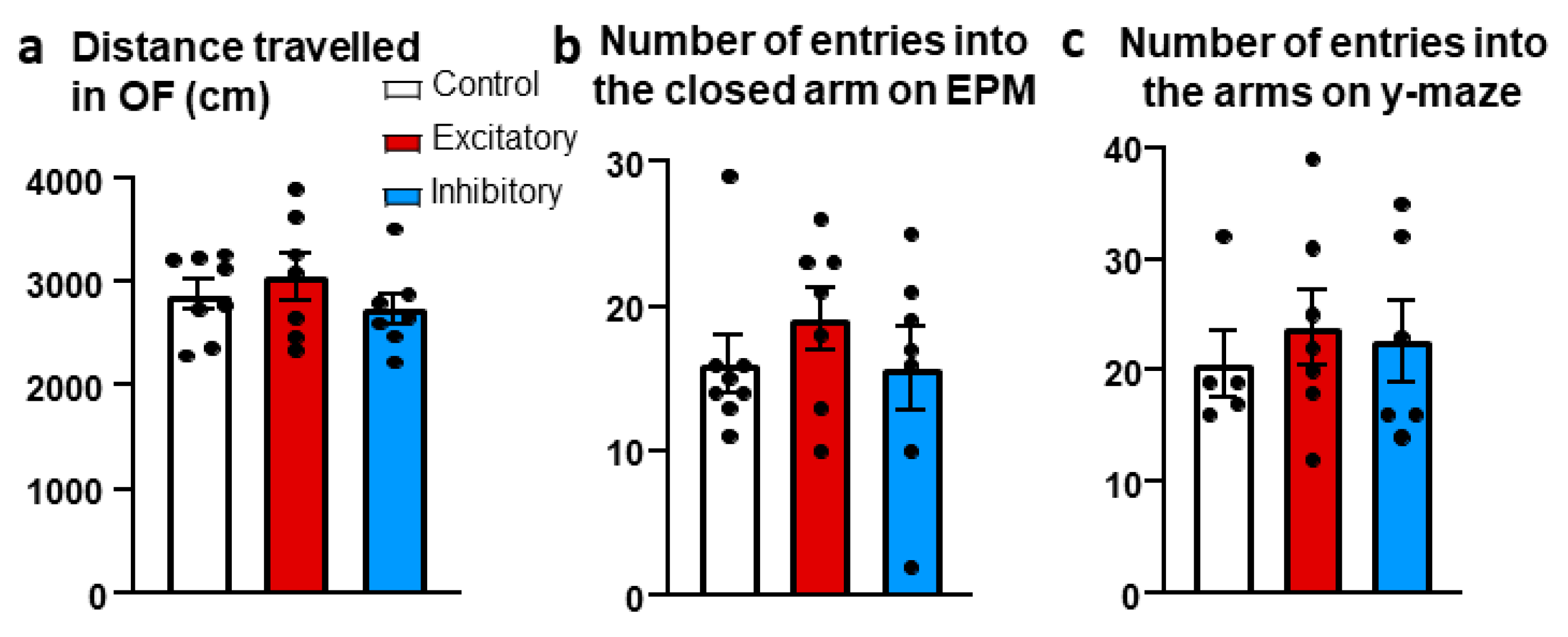
Figure 5.
Results of Social Behavior tests. (a) Frequency of the interaction (sniffing) between the test mice and the empty cage (box) or caged mice (mouse). (b) Time of this interaction. (c) Social Index displayed the social preference toward the stimulus mice. For more details, see methods section. All groups performed above the chance level 50. (d) Time spent interacting with mice (caged familiar and caged unfamiliar mice) during social discrimination. Chemogenetic stimulation of the dopaminergic cells of the median raphe region (DAT-MRR) decreased social interest. (e) Time spent interacting with conspecific animal during social interaction test. Chemogenetic inhibition of DAT-MRR cells increased social interest. (f) Time spent interacting with conspecific animals during resident intruder test. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values. * p < 0.01 vs control.
Figure 5.
Results of Social Behavior tests. (a) Frequency of the interaction (sniffing) between the test mice and the empty cage (box) or caged mice (mouse). (b) Time of this interaction. (c) Social Index displayed the social preference toward the stimulus mice. For more details, see methods section. All groups performed above the chance level 50. (d) Time spent interacting with mice (caged familiar and caged unfamiliar mice) during social discrimination. Chemogenetic stimulation of the dopaminergic cells of the median raphe region (DAT-MRR) decreased social interest. (e) Time spent interacting with conspecific animal during social interaction test. Chemogenetic inhibition of DAT-MRR cells increased social interest. (f) Time spent interacting with conspecific animals during resident intruder test. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values. * p < 0.01 vs control.
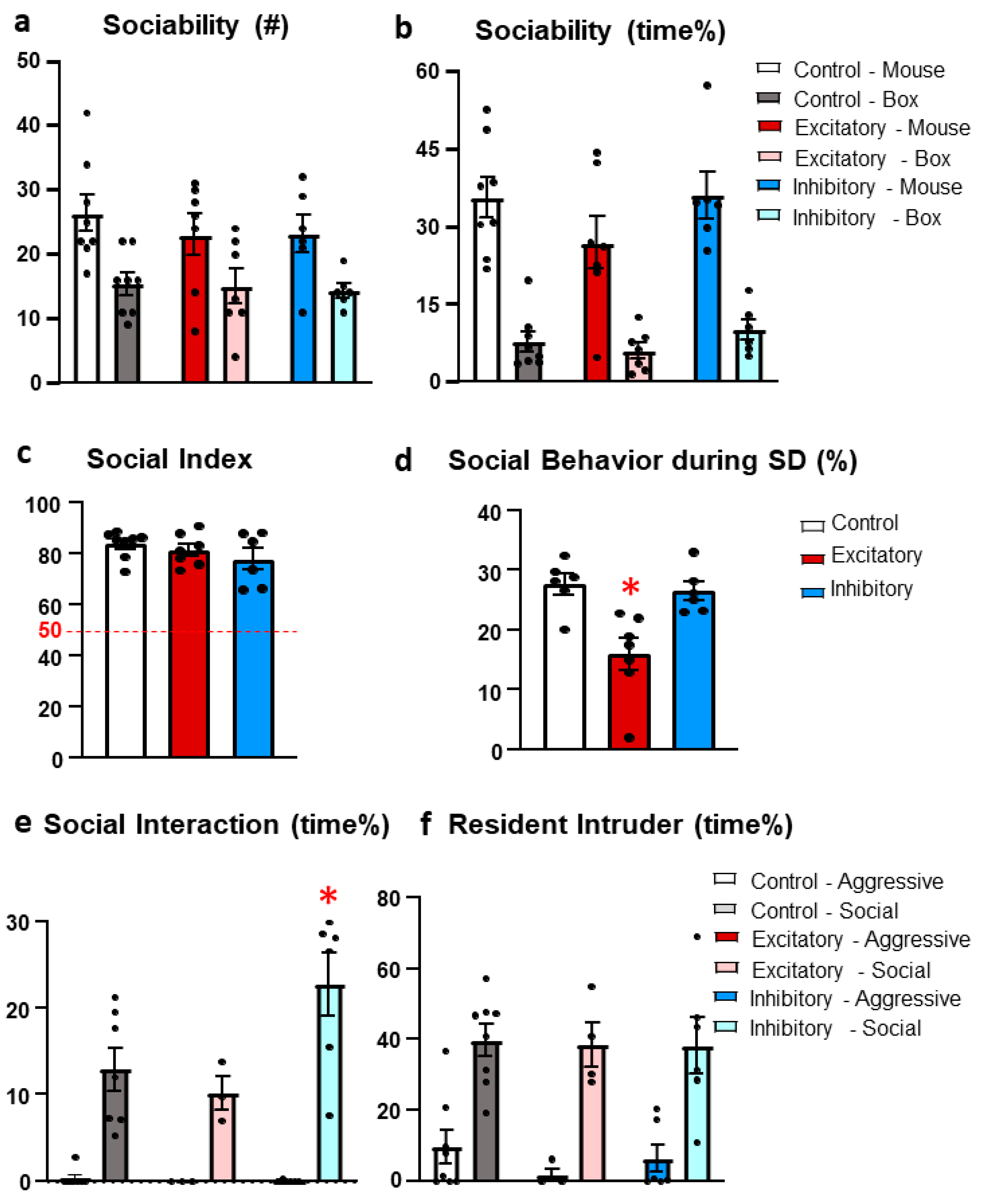
Figure 6.
Results of Anxiety and Memory tests. (a) The time spent in the open arm in the elevated plus maze during the 5 min observation was without group difference. (b) The open/total arm entries shows the index of anxiety [51] without group difference. (c) Percentage of “good” alternations in the Y-maze (see Section 4.3.6.). All animals performed above the chance level (marked red, 50). (d) Discrimination index during the social discrimination test. Animals were not able to distinguish between familiar and unfamiliar mice 24h after “sampling” (no significant difference from the 50% chance level). (e) The time the test animal spent investigating old and new stimulus animal during the social discrimination test. Chemogenetic stimulation of the dopaminergic cells of the median raphe region (DAT-MRR) decreased overall social interest. (f) Frequency in which the animals spent with the caged conspecific animals during the social discrimination test revealed no difference between groups. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values. * p < 0.01 vs control and inhibitory groups.
Figure 6.
Results of Anxiety and Memory tests. (a) The time spent in the open arm in the elevated plus maze during the 5 min observation was without group difference. (b) The open/total arm entries shows the index of anxiety [51] without group difference. (c) Percentage of “good” alternations in the Y-maze (see Section 4.3.6.). All animals performed above the chance level (marked red, 50). (d) Discrimination index during the social discrimination test. Animals were not able to distinguish between familiar and unfamiliar mice 24h after “sampling” (no significant difference from the 50% chance level). (e) The time the test animal spent investigating old and new stimulus animal during the social discrimination test. Chemogenetic stimulation of the dopaminergic cells of the median raphe region (DAT-MRR) decreased overall social interest. (f) Frequency in which the animals spent with the caged conspecific animals during the social discrimination test revealed no difference between groups. Data (n = 7–8) are expressed as average ± SEM. Dots represent individual values. * p < 0.01 vs control and inhibitory groups.
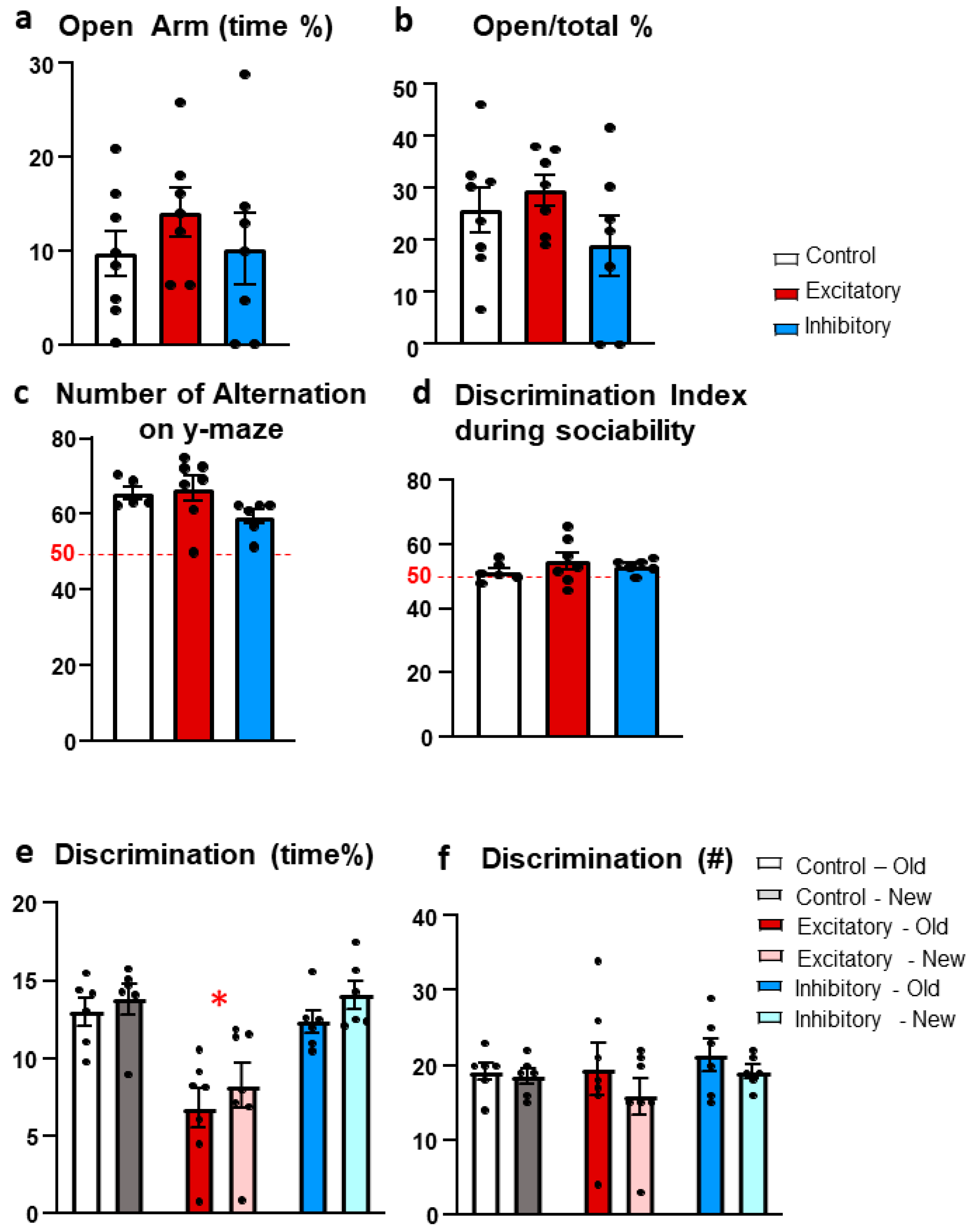
Figure 7.
Virus injection details. (a) Schematic illustration of the targeted mouse median raphe region (MRR) in coronal section according to the Mouse Brain in Stereotaxic Coordinates [50]. (b) Representative image of an appropriate virus expression (visualized by nickel-3,3′-diaminobenzidine staining for the reporter protein red fluorescent protein [RFP]) in the MRR of a dopamine-transporter-Cre mice 6 weeks after 10nl rAAV8/hSyn-DIO-HM4D(Gi)-mCherry (Addgene #44362) injection. (c) Representative figure of a rejected virus expression with staining both in the dorsal raphe (DR) and MRR. Aq: cerebral aqueduct. Coordinates: anterior-posterior: −4.1 mm; lateral: 0.0 mm; dorso-ventral: −4.6 mm from Bregma.
Figure 7.
Virus injection details. (a) Schematic illustration of the targeted mouse median raphe region (MRR) in coronal section according to the Mouse Brain in Stereotaxic Coordinates [50]. (b) Representative image of an appropriate virus expression (visualized by nickel-3,3′-diaminobenzidine staining for the reporter protein red fluorescent protein [RFP]) in the MRR of a dopamine-transporter-Cre mice 6 weeks after 10nl rAAV8/hSyn-DIO-HM4D(Gi)-mCherry (Addgene #44362) injection. (c) Representative figure of a rejected virus expression with staining both in the dorsal raphe (DR) and MRR. Aq: cerebral aqueduct. Coordinates: anterior-posterior: −4.1 mm; lateral: 0.0 mm; dorso-ventral: −4.6 mm from Bregma.

Table 1.
Summary of results.
| Function | Test | Excitatory | Inhibitory |
|---|---|---|---|
| Locomotion | OF | ∅ | ∅ |
| EPM | ∅ | ∅ | |
| Y Maze | ∅ | ∅ | |
| Social Behavior |
Sociability | Decreased social interest | ∅ |
| Social Interaction | ∅ | Increased social behavior | |
| Resident Intruder | ∅ | ∅ | |
| Anxiety | EPM | ∅ | ∅ |
| Memory | Y Maze | ∅ | ∅ |
| Social discrimination at 24 h | ∅ | ∅ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



