
Submitted:
20 March 2024
Posted:
21 March 2024
You are already at the latest version
Abstract
Black soldier fly larvae (BSFL) can use food by-products or residues as a growth substrate, and their use is considered a strategic solution to improve the sustainability and efficiency of animal production. BSFL need to be reared under controlled environmental conditions, as temperature can affect the welfare of the larvae and the efficiency of their bioconversion. The goal of this experiment was to test different diets: control, vegetable, carnivorous, omnivorous and to assess their effect on larval growth performance, chemical composition, and substrate temperature. BSFL (6-days-old) were randomly selected and allocated to one of the four diets for the entire experiment (8 days). The temperature in each container was measured twice daily using a thermal imaging camera. The vegetable diet led to significantly lower growth performance (p
Keywords:
Black soldier fly
; substrate
; bioconversion
; thermal imaging camera.
1. Introduction
Worldwide, the increasing population generates a huge volume of food waste (FW). Inadequate management and lack of proper handling of FW have serious adverse effects on the environment and human health [1]. Food losses reduce the amount of food available on the market. All the energy and resources, including water and land, used to grow or produce this food is also wasted. These massive food losses are terrible in a world where the population is expected to grow from seven to nine billion people by 2050 [2]. Furthermore, the high cost of a healthy diet and persistently high levels of poverty make food inaccessible to a large share of the world’s population [3,4,5]. With widespread food insecurity for hundreds of millions of people around the world, addressing FW is a critical factor in creating environmentally friendly, healthy and resilient food systems. The livelihoods of all people and animals depend on food security, which is achieved when all people have access to adequate nutritious food that is always accessible, satisfies their nutritional needs, and allows them to live active, healthy lives [6]. The UNEP Food Waste Index Report 2021 estimates that around 931 million tonnes of FW were generated in 2019 [7]. The UN has included Goal 12.3 (“ensure sustainable consumption and production patterns”) among the 17 Sustainable Development Goals in the 2030 Agenda and aims to halve FW. Management varies from country to country [8], and there are different recycling technologies of organic waste i.e., incineration, landfill anaerobic digestion, and compositing [9,10,11].
In the context of waste valorization, a promising strategy is the use of FW as a substrate for mass-insect rearing [12]. The use of insects in the bioconversion of FW is a new approach and an interesting example of a sustainable and circular economy. The use of Black soldier fly larvae (BSFL, Hermetia illucens L., Diptera: Stratiomyidae) in the FW treatment has emerged as a key innovation [13] due to the larvae high production rate, low cost, and shorter life cycle compared to other species. The extreme voracity and rapid development cycle of BSFL can also be exploited for the disposal of organic wastes with high environmental impact, such as manure from intensive livestock farming, agri-food industry waste, and urban solid waste [14,15]. Their rapid expansion is also due to the benefits associated with the utilization of waste. Most of the current research on BSF investigates the larval stage as it is the most suitable stage for recycling organic material, including FW, and produce biomass that can be used as feed [14,15,16,17].
Larvae can generate nutrient rich biomass consisting on average of 22.2-45.1% crude protein, 26.2-40.7% fat, and micronutrients [18,19,20,21]. The nutrient composition in terms of crude protein and lipid content of the biomass depends on the substrates provided and the rearing conditions [22]. Since nutrients affect the physiology, behaviour, and growth of larvae, studies on nutrient requirements, and specifically, protein and lipid requirements are being conducted [23,24]. The ability of BSFL to convert waste into high- value nutrient biomass has rapidly opened innovative economic perspectives for municipal solid waste management in different sectors. In addition, this biomass has been recommended as a suitable alternative to fishmeal or soybean meal in poultry, pig, and fish feed [20,22,25].
Insect rearing is complicated because insects require strict environmental conditions such as temperature and humidity, feeding and, stocking density. The rearing environment and diet must avoid adverse (or limiting) factors, both biotic (pathogens, parasitoids, and predators) and abiotic (temperature, humidity, air flow and light) [26,27]. Environmental conditions such as temperature, humidity, light (wavelength, intensity, polarization, and light colour temperature), and aeration are important, and determine successful copulation among individuals of the opposite sex, egg-laying activities, and mass production [28,29,30]. Maintaining appropriate ecological conditions is the most effective method to obtain a successful bioconversion from BSFL and manage the mass-rearing activities of BSF. Many researchers have examined the effect of different temperatures on the life cycle of BSF [31,32].
This research aims to analyses the effect of four different diets for BSFL on substrate temperature, growth performances, substrate reduction, and the chemical composition of the larval body. Four different diets were used: vegetable, carnivorous, omnivorous, and control. The EU forbids the use of animal products (except milk, eggs, and derivatives) and municipal trash, particularly the organic portions of municipal solid waste, for industrial insect farming systems [33,34]. Nonetheless, it is legal to utilize vegetable food and by-product residuals as an insects’ food source [34,35].
2. Materials and Methods
2.1. Colony
The study was conducted in the laboratories of the BEF Biosystems company (Turin, Italy). The BSF stock population was maintained in a breeding room from 2019. BSF adults were bred in a steel frame cage (100 x 63 x 110 cm) covered with a mosquito net. The light source was an LED panel, with a wavelength suitable for BSF, as described by Oonincx et al. [36], with a photoperiod of light: night (12:12). BSF were maintained in a climate-controlled room with a temperature of 27±1°C and a relative humidity of 65±5%. The adult flies were provided with water during their entire life and wooden sticks were placed in the cage as an oviposition substrate. The wooden sticks were checked every day and replaced every second day. Such practice ensured a synchronized development of the larvae during the experiment. The eggs were collected from the colony using the methods outlined by Dortmans et al. [37]. Using a paintbrush, the eggs originating from multiple females were placed in plastic cups from wooden sticks and then transferred to the plastic boxes in which the experiment took place (size: 60 x 40 x 12 cm). After hatching, the larvae were fed a mixture of chicken feed and water until the test starts.
2.2. Diet Formulation
Four experimental diets were formulated: 1) control diet (D1) containing chicken feed, according to the scientific literature[14,38]; 2) vegetable diet (D2) with by-products such as carrots, potatoes, and brewer’s spent grain, these by-products being mixed in a weight ratio of 1:1:1; 3) omnivorous diet (D3) obtained by mixing a 1:1 ratio the vegetable diet and the carnivore diet; 4) carnivore diet (D4), obtained by mixing ground beef epiglottis and cod pulp in a ratio of 1:1 ratio. The vegetable raw materials for the preparation of the substrates (i.e., diets) were sourced locally and ground epiglottis and cod were purchased (Barf®, Tortona, AL, Italy).

Table 1.
The proximate composition of the four diets and young larvae (g kg-1 on a wet basis).
| Parameter | Initial Substrates | Young Larvae | |||
|---|---|---|---|---|---|
| D1i | D2 | D3 | D4 | ||
| Dry Matter | 278.0 | 304.1 | 255.8 | 245.2 | 302.6 |
| Organic matter | 239.2 | 284.6 | 238.5 | 230.2 | - |
| Crude Proteins | 40.1 | 20.1 | 81.5 | 121.8 | 112.9 |
| Crude Fat | 14.9 | 9.4 | 34.4 | 54.3 | 31.8 |
| Ash | 38.8 | 19.5 | 17.3 | 15.0 | 50.6 |
| Crude Fiber | 14.0 | 36.1 | 24.7 | - | 29.8 |
| Neutral detergent fiber | 48.1 | 68.8 | 85.1 | - | 43.8 |
| Acid detergent fiber | 22.9 | 59.0 | 35.5 | - | 37.6 |
| Acid detergent lignin | 7.60 | 18.7 | 12.5 | - | 7.2 |
| N-Free extractive ii | 612.53 | 720.39 | 382.94 | 220.41 | 77.4 |
| Starch | 94.1 | 135.3 | 51.9 | 6.1 | - |
i D1, Control; D2, Vegetable; D3, Omnivorous; D4, Carnivorous. ii Calculated as 100- [ crude protein+ fat +ash +fiber].
2.3. Experimental Design
300 larvae (6-days-old) were weighed individually from a pool of two kilograms of BSFL using an analytical balance (U.S. Solid, Cleveland, Ohio) to determine the average weight of larvae. Once the average weight of the larvae was determined, 32000 homogeneous larvae were selected. Sixteen groups of 2000 larvae (four replicates for each diet) were transferred to 16 plastic containers (32 x 23.5 x 11.5 cm). The containers were fitted with a mosquito net on top to prevent larval escape or possible contamination. The containers were chosen specifically to result in the same height of substrate across treatments (~8 cm).
The rearing facility-maintained larvae at the environmental conditions of 27±1°C, 65±5% relative humidity, and a photoperiod (12:12). Four replicates were conducted for each experimental diet and the amount of substrate in each replicate was calculated as 100mg larva/day as described by Diener et al. [14] throughout the trial period. A total amount of 2000g of substrate was allocated in each container to ensure a ten-days food supply. From the second day onwards, two ventilators provided constant ventilation to ensure that the substrate was thoroughly moisturized for the entire experiment duration (8 days). On the third day, the mosquito lids were removed to allow homogeneous drying of the substrates.
2.4. Larvae Growth
At the end of the experimental, the total biomass (larvae) and the residual rearing substrate were weighed (Kern, Balingen, Germany) for each container. The following parameters were then calculated Growth Rate (GR) and Substrate Reduction (SR):
GR= (larvae average final body weight (g) – larval initial body weight (g))/days of trial (d);
SR= [(distributed substrate (g) – residual substrate (g))/ distributed substrate (g)] x 100.
A sample of larvae was individually cleaned and photographed (Nippon Avionics CO., LTD, Shimamura-Building, Konobe-cho, Tsuzuki-ku, Yokohama-shi, Japan) orthogonally with a metric scale (mm). The ImageJ software package (v 1.50v, Bethesda, U.S.A.) was used to examine the photographs. The length of the larvae was recorded from the mouthparts to the lower part of the last abdominal segment.
2.5. Chemical Analysis and Utilization of Substrate Nutrients
Substrates and larvae were frozen at – 20° C and subsequently were analysed in the laboratories of Animal Production and Food Safety service (SPASA) of the Department of Veterinary Medical Sciences (DIMEVET) of the University of Bologna, Italy. Samples of larvae and substrates were freeze-dried (Olsa, Milano, Italy) to remove all moisture contained in the organic matter and were ground using a shredder (Broyeur mélangeur MB 950G KINEMATICA, Malters, Switzerland). All samples were analysed to measure: crude protein (CP) [39] using a Kiedahl nitrogen analyser (Gerhardt Vapodest50, Gerhardt Gmbh, Königswinter, Germany). For the nitrogen-to-protein conversion, the more precise N- factor of 4.67 proposed by Janssen et al. [40] was used instead of the standard conversion factor (6.25). Starch was determined according to AOAC method 996.11 and ether extract according to AOAC method 920.390020 [41]. Neutral detergent fiber (NDF), acid detergent fiber (ADF) acid detergent lignin (ADL) were analysed according to the method of Van Soestet al. [42] and ash was determined after 3 hours of combustion in a muffle furnace at 550°C (VULCAN 3-550, Dentsply Neytech Burlington, NJ, USA).
The formula to determine the percentage of degradation of each component (x) of the biomass was calculated based on the initial and final analytical composition of the substrates:
Degradationx = (Ax-Bx)/Ax × 100,
where A and B were the initial (A) and final (B) weight (g) of the x component of the diet in the substrate. Degradation was calculated for dry matter, crude protein, fats, ash, starch, and non-structural carbohydrates.
2.6. Substrate Temperature and Accumulated Degree Hours
The temperature of the substrate was recorded, and thermographic pictures were taken using a thermal imaging camera (Nippon Avionics CO., LTD, Shimamura-Building, Konobe-cho, Tsuzuki-ku, Yokohama-shi, Japan) to determine the highest and lowest temperatures of each box. The biomass of the larvae and the substrates were monitored twice a day (morning and afternoon) on days: 1, 2, 3, 5, 7, and 8. In this way, we obtained a temperature trend during the test. The thermal images were taken directly above each container and the hottest part of the surface was determined as the hot spot temperature. The software used to read the temperature was InfReC Analyzer NS9500 Standard (v. 5.0 C, Yokohama-shi, Japan).
Similar, to previous work by Harnden and Tomberlin [32], we calculated the thermal summation model to obtain the accumulated degree hours (ADH) or days (ADD) for each of the diets used in the present experimental. The ADH was calculated using the minimum, maximum and means temperature, the critical temperature of the larvae, and the duration of the experiment. The formula was calculated using a low development threshold (LDT) of 12°C [43]:
ADH= temperature of substrate (°C)- LDT (°C) × time (h),
where the substrate temperature was the minimum, maximum and mean recorded by the thermal imaging camera.
2.7. Statistical Analysis
Statistical analysis was performed using Statistica (StatSoft Inc., release 12, 2013). The Shapiro-Wilk test was used to verify normal distribution of the dependent variables for each combination of groups within-subject and between-subject factors. Levene’s test was used to verify the homogeneity of variances for each combination of groups of within- and between-subject factors. All data except substrate temperature were subjected to one-way analysis of variance (ANOVA) with diet as a fixed factor. Substrate temperature was analysed using ANOVA with diet and day of experiment as fixed factors. Tukey’s test was applied for pairs comparisons. The statistical significance threshold was set at p<0.05).
3. Results
3.1. Larvae Gowth
The effect of the rearing substrate on the development of BSF larvae over time is reported in Table 2. Overall, D1 (control) showed the best results, while D2(vegetable) performed the worst in terms of larval growth. All variables presented in Table 2 significantly differ with a p-value <0.01. In particular, the larval biomass of D2 was significantly lower than that of D1, resulting in a significantly higher residual biomass of the substrate in D2 compared to D3 (omnivorous) and D4 (carnivorous) (p<0.01). Larval weight and length were significantly higher in the D1 compared to the other groups (p<0.01). Moreover, the growth rate of the control group was significantly higher compared to the other groups, while the substrate reduction was significantly lower for group D2 compared to all the other groups. As expected, the larvae meal yield from the D2 was the lowest. In summary, the D2 diet yielded the lowest biomass of larvae meal.
3.2. Larvae Chemical Composition
As shown in Table 3, the proximate composition of the larvae varied between the different diets, and in particular the protein, ash, and fiber content were highly variable. Larvae fed a vegetable diet had significantly lower protein content than those fed a carnivorous diet (p<0.05). The fat content of the larvae was statistically lower in D2 compared to the other groups (p<0.01), and the ash content was similar only in groups D3 and D4 and significantly lower compared to D1 and D2. Crude fiber was statistically different (p<0.01) between all groups except D1, with D4 having the lowest and D2 the highest fiber content, while fiber fractions (NDF, ADF, ADL) did not show any significant difference between diets. Non-structural carbohydrates were lowest in D1 and highest in D2 (p<0.01), with an intermediate value of D3 and D4.
3.3. Substrates Chemical Composition
The results of substrate degradation are summarized in Table 4. All variables showed statistical differences between the groups in degradation. DM degradation was lower in D1 than other groups(p<0.01). In addition, OM degradation was lower in D1 than in all other groups (p<0.01). Crude protein degradation was higher in D3 than in D1 and D4 (p<0.01) and lower in D1 than in D2 and D3 (p<0.05). Fat degradation was lower in D4 than in all other groups (p<0.01), ash degradation was lower in D1 than the other groups (p<0.01). Starch degradation was higher in D4 compared to the other groups (p<0.01), and the degradation of non-structural carbohydrates was higher in D2 than in D1 and D3 (p<0.01) and also inD4 (p<0.05).
3.4. Substrate Temperature
The maximum and minimum temperatures recorded in the different substrates of the BSFL are shown in Figure 1. The temperature trend over the course of the experiment was different for the different diets. An effect of the diet was observed: the D1 group exhibited a statistically significant difference, being lower compared to the other groups. A time effect was observed, with mimimum temperatures being significantly higher on day 7 (p<0.01) than on all other days (p<0.01). There were significant differences in both maximum and minimum temperature, with the lowest temperature measured on the last day compared to the two measurements on day 5 and day 7 (p<0.01).
3.5. Accumulated Degree Hours
The maximum, minimum and mean ADH values of BSFL reared on different diets are shown in Table 5. It was found that the diets had not significant effect on minimum, maximum and mean ADH values during the experiment period. The maximum ADH value was recorder in D3 while the minimum value was observed in D1. The mean ADH values ranged between 2964 and 3072.
4. Discussion
One of the proposed strategies to reduce food waste and loss is insect-based bioconversion [15,44]. By converting organic waste into useful nutrients, BSFL can play a significant role in recovering these lost nutrients, as BSF is characterized by a low (i.e., very favorable) food conversion ratio. All diets used in this trial allowed larvae to grow and develop, although at different extents. BSFL reduced all four substrates and provided a different larval composition depending on the composition of the diet. Considering the control group (D1) as the most successful, it is clear that D3 achieved the best values among the tested diets in terms of larval growth, as this group showed a high larval biomass and a low residual substrate. From a nutritional point of view, the content of crude protein and crude fat in D3 larvae is also similar to that of D1 larvae (Table 3); also, DM value is similar to the control diet (Table 2). Our results are in agreement with those present by Tschirner et al. [45] who showed that the standard substrate group had the best results in terms of total larvae yield, individual larvae weight, and substrate consumption compared to the protein and fiber groups. Due to the differences in minerals and microelements, indeed the D1 presented the highest content in ashes and showed better growth performances compared to the other groups in terms of individual larval weights. As reported by Rebora et al. [46], BSFL deposit the carbonate calcium as calcite on the exoskeleton in the last stages of the life cycle, while in the young larval stages it is deposited mainly in the form of amorphous calcium. The granules of calcium carbonate are stored in organs, the Malpighian tubules. So, the effect of minerals, carbonate calcium is one the most important, should impact the cycle of BSFL, no research evaluated this point until now. Despite the D2 diet showing similar substrate utilization coefficients to the control diets, crude protein and fat contents were lower (approximately one half compared to D1) resulting in the worst performances. This underlines that protein and fat were limiting factors in this diet. These findings are consistent with the trial conducted by Bellezza-Oddone et al. [23] where better growth performance was obtained in diets using a fat content of 4.5%. The chemical composition of D2 larvae mirrors the chemical composition of the diet. Conversely, D3 and D4 diets, despite being higher in protein and fat, as also found by Nguyen et al. [47] did not lead to improved growth performances compared to D1 diet. This result may suggest sugar content as a limiting factor in these diets. However, D4 larvae, due to both their higher DM content compared to other diets and the high levels of crude protein and fat, allowed the production of quantities similar to D1 in terms of larvae yield, protein and fat. In particular, larvae fed on a carnivorous diet had significantly higher DM content than those fed other diets, resulting in a higher larvae meal yield. As suggested by Gold et al. [48] a well-balanced combination of substrates enhances growth and lowers variability, which is likely why larvae fed an omnivorous diet performed better in the present study.
Substrate degradation finding indicate that DM degradation, particularly OM and CP, was lower in D1 compared to the other groups. Considering that D1 showed the best growth rates, it can be supposed that the quantity of substrate supplied may be excessive in relation to the number of larvae present. The degradation of starch was higher in D4 compared to the other groups, primarily because this diet had the lowest starch content. This result aligns with Barragan-Fonseca et al.[49] who observed that larvae yield was greater on diets with a high non-structural carbohydrate content. However, according to the growth performance of D1, it is clear that the larvae’s ability to grow is influenced by factors beyond the amount of proteins and carbohydrates.
It is commonly known that environmental factors influence the growth and survival of insects. BSF is sensitive to several environmental factors, the most important abiotic factor being temperature [49,50,51] Numerous studies examined the effects of using organic side streams as feeding substrates or laboratory-reared meals at constant temperature on the life-history characteristics of BSFL. [31,32,49,52]. As demonstrated also by Shumo et al. [53], both environmental temperature and substrate type significantly influence BSF larval development. In the study of Ribeiro et al. [54] under continuous feeding conditions, seven different isolated vegetables (wheat bran, pumpkin, apple, grape pomace, red onion, red cabbage, and spinach) were used as substrates. The conditions for the substrates were varied and included natural and 70% substrate moisture at three different temperatures: 20°, 25°, and 30° C. The authors showed that higher temperatures contributed to the rapid development of BSFL. In the present study, contrasting differences in minimum substrate temperatures were observed in the D1 group, indicating the best performance. Therefore, this aspect was not a factor affecting larval development. Harnden and Tomberlin [32] found that the minimum ADH (degree hours required for a species to complete each developmental stage or to achieve a specified larval length or weight) required to complete larval development differed significantly for each diet and temperature. The ADH for completion of larval growth was lower for larvae reared on cereal meal and higher for larvae reared on pork. Overall, these results confirm that both temperature and diet significantly affect the development of BSF during the egg, larval, and prepupal stages. In our study, in which we applied a similar model to the ADH, it was found that the type of diet had no effect on the temperature of the larvae during their development. This seems clear for the experimental design in this study, which aimed to investigate whether the different diet composition of the diet has an effect on the temperature produced by the secondary fermentation triggered by larval activity. Indeed, all replicates were kept at the same room temperature (27°C), instead in Harden and Tomberlin [32] used high different temperatures such 24.9°C, 27.6°C and 32.2°C in the climate chamber. Another important aspect to consider is that as the larvae grow larger, they produce more heat, but as the days go by, they feed less and the heat decreases. Such a general shift in substrate temperature indicates changes in digestive activity and metabolism during larval ontogeny [55]. According to Georgescu et al. [24], during the growth of BSF larvae from the third to the last instar, the mass-specific metabolic rate decreases from about 12 to 2 μW/mg, while weight of the larva increases from about 1 to 90 mg. According to McEachern [55], the age and weight of the larvae have a major influence on the amount of heat produced by BSF larvae. The maximum amount of heat is produced by aggregates when the larvae are larger and older. The study also found that the larvae release less heat per gram of body weight as they grow. In fact, our study agrees with the cited study as it shows that substrate temperatures (minimum and maximum) increased with the age of the larvae, together with the increase in the amount of heat produced, but that the temperature began to decrease with increasing weight. Furthermore, even at a constant population density, the larvae do not consistently produce the same amount of heat per individual or per gram of body weight. Instead, the heat production of BSFL varies greatly depending on the size of the group in which they live. Temperature fluctuations within a group can be advantageous as they allow the larvae to thermoregulate through their behaviour. A potential disadvantage is that the larvae are likely to be exposed to inappropriate temperatures during their movement. As reported by Li et al. [56] the present study confirmed a similar trend in substrate temperature throughout the experiment. In their study, the maximum substrate temperature increased as the experiment progressed, peaked around the fourth day of the experiment, and then began to decrease.
The temperature measurements taken in this study reflect the temperature at the surface of the biomass. This identification could be even more interesting as the larvae prefer to remain on the surface in darkness rather than just in the inner layers. This assessment could be even more interesting and intriguing than measuring the temperature inside the substrate (e.g. with a probe). Therefore, it can be assumed that the observed differences in growth performance are a consequence of the different composition of the substrate, and are not due to the substrate temperatures themselves.
5. Conclusions
The aim of this study was to evaluate the effects of different food waste and residues (ranging from a vegetable to a carnivorous diet) as rearing substrates for BSFL on their development, particularly on degradation/bioconversion efficiency and variations in surface temperatures. Several factors are probably responsible for the different larvae development in the different diets. The most influential factor was the different nutrient content of the animal- and plant-based diets. Larvae fed on diets containing meat exhibited favorable growth performance. It is important to note that, according to regulations, feeding larvae with animal-derived products is not permitted. However, further research may be warranted to explore the potential utilization of this type of waste and to provide a more precise scientific framework for quantitative and qualitative nutrient requirements and for measuring larval digestibility and metabolic heat production.
Author Contributions
Conceptualization Sardi L. and Meneguz M.; methodology Meneguz M.; software Sardi L. and Belperio S.; validation Belperio S. and Cattaneo A.; formal analysis Belperio S.; resources Meneguz M. and Sardi L..; data curation Belperio S. and Cattaneo A.; writing—original draft preparation Belperio S.; writing—review and editing Nannoni E and Meneguz M.; visualization Belperio S.; supervision Martelli G. and Dabbou S.; project administration Sardi L.; funding acquisition Sardi L and Meneguz M.
Funding
This research received no external funding.
Institutional Review Board Statement
Larvae were raised in standard farming conditions and were not subjected to any invasive procedure; therefore, no review was needed. No increased mortality or signs larvae suffering were observed during the trial.
Acknowledgments
This project received technical support from T. Barber.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ambaye, T.G.; Rene, E.R.; Nizami, A.-S.; Dupont, C.; Vaccari, M.; Van Hullebusch, E.D. Beneficial Role of Biochar Addition on the Anaerobic Digestion of Food Waste: A Systematic and Critical Review of the Operational Parameters and Mechanisms. Journal of Environmental Management 2021, 290, 112537. [Google Scholar] [CrossRef] [PubMed]
- FAO, IFAD, UNICEF, WFP and WHO. 2023 In Brief to The State of Food Security and Nutrition in the World 2023Urbanization, Agrifood Systems Transformation and Healthy Diets across the Rural–Urban Continuum.; FAO; IFAD; UNICEF; WFP; WHO; Rome, FAO., 2023; ISBN 978-92-5-137952-3.
- Food and Agriculture Organization of the United Nations,The State of Food Security and Nutriion in the World。 Trasforming Food Systems for Food Security, Improved Nutrition and Affordable Healthy Diets for All “ FAO Rome, 2021. [CrossRef]
- FAO, IFAD, UNICEF, WFP and WHO. 2022. The State of Food Security and Nutrition in the World 2022. Repurposing Food and Agricultural Policies to Make Healthy Diets More Affordable.; FAO: Rome, FAO, 2022. [Google Scholar] [CrossRef]
- Food and Agriculture Organisation Water for Sustainable Food and Agriculture Water for Sustainable Food and Agriculture; 2017; ISBN 9789251099773.
- Clapp, J.; Moseley, W.G.; Burlingame, B.; Termine, P. Viewpoint: The Case for a Six-Dimensional Food Security Framework. Food Policy 2022, 106, 102164. [Google Scholar] [CrossRef]
- United Nations Environment Programme (2021). Food Waste Index Report 2021. Nairobi ISBN 978928073 8513.
- OECD; Food and Agriculture Organization of the United Nations OECD-FAO Agricultural Outlook 2023-2032; OECD-FAO Agricultural Outlook; OECD, 2023; ISBN 978-92-64-61933-3.
- Awasthi, S.K.; Sarsaiya, S.; Awasthi, M.K.; Liu, T.; Zhao, J.; Kumar, S.; Zhang, Z. Changes in Global Trends in Food Waste Composting: Research Challenges and Opportunities. Bioresource Technology 2020, 299, 122555. [Google Scholar] [CrossRef] [PubMed]
- Tiwary, A.; Williams, I.D.; Pant, D.C.; Kishore, V.V.N. Assessment and Mitigation of the Environmental Burdens to Air from Land Applied Food-Based Digestate. Environmental Pollution 2015, 203, 262–270. [Google Scholar] [CrossRef]
- Pham, T.P.T.; Kaushik, R.; Parshetti, G.K.; Mahmood, R.; Balasubramanian, R. Food Waste-to-Energy Conversion Technologies: Current Status and Future Directions. Waste Management 2015, 38, 399–408. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental Impact of Food Waste Bioconversion by Insects: Application of Life Cycle Assessment to Process Using Hermetia Illucens. Journal of Cleaner Production 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ristow, B.; Rahayu, T.; Putra, N.S.; Widya Yuwono, N.; Nisa’, K.; Mategeko, B.; Smetana, S.; Saki, M.; Nawaz, A.; et al. Black Soldier Fly Larvae (BSFL) and Their Affinity for Organic Waste Processing. Waste Management 2022, 140, 1–13. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of Organic Material by Black Soldier Fly Larvae: Establishing Optimal Feeding Rates. Waste Manag Res 2009, 27, 603–610. [Google Scholar] [CrossRef]
- Surendra, K.C.; Tomberlin, J.K.; Van Huis, A.; Cammack, J.A.; Heckmann, L.-H.L.; Khanal, S.K. Rethinking Organic Wastes Bioconversion: Evaluating the Potential of the Black Soldier Fly (Hermetia Illucens (L.)) (Diptera: Stratiomyidae) (BSF). Waste Management 2020, 117, 58–80. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Van Huis, A. Black Soldier Fly from Pest to ‘Crown Jewel’ of the Insects as Feed Industry: An Historical Perspective. JIFF 2020, 6, 1–4. [Google Scholar] [CrossRef]
- Lalander, C.; Ermolaev, E.; Wiklicky, V.; Vinnerås, B. Process Efficiency and Ventilation Requirement in Black Soldier Fly Larvae Composting of Substrates with High Water Content. Science of The Total Environment 2020, 729, 138968. [Google Scholar] [CrossRef] [PubMed]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional Composition of Black Soldier Fly ( Hermetia Illucens ) Prepupae Reared on Different Organic Waste Substrates. J Sci Food Agric 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Schlüter, O.K. Nutritional Composition and Safety Aspects of Edible Insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-Art on Use of Insects as Animal Feed. Animal Feed Science and Technology 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinnerås, B. Effects of Feedstock on Larval Development and Process Efficiency in Waste Treatment with Black Soldier Fly (Hermetia Illucens). Journal of Cleaner Production 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing Substrate Impacts Growth and Macronutrient Composition of Hermetia Illucens (L.) (Diptera: Stratiomyidae) Larvae Produced at an Industrial Scale. Sci Rep 2020, 10, 19448. [Google Scholar] [CrossRef]
- Bellezza Oddon, S.; Biasato, I.; Resconi, A.; Gasco, L. Determination of Lipid Requirements in Black Soldier Fly through Semi-Purified Diets. Sci Rep 2022, 12, 10922. [Google Scholar] [CrossRef]
- Georgescu, B.; Boaru, A.M.; Muntean, L.; Sima, N.; Struți, D.I.; Păpuc, T.A.; Georgescu, C. Modulating the Fatty Acid Profiles of Hermetia Illucens Larvae Fats by Dietary Enrichment with Different Oilseeds: A Sustainable Way for Future Use in Feed and Food. Insects 2022, 13, 801. [Google Scholar] [CrossRef] [PubMed]
- Barragán-Fonseca, K.Y.; Barragán-Fonseca, K.B.; Verschoor, G.; Van Loon, J.J.; Dicke, M. Insects for Peace. Current Opinion in Insect Science 2020, 40, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Leppla, N.C. Rearing of Insects Second Edi.; Elsevier Inc., 2009; ISBN 9780123741448.
- Palma, L.; Ceballos, S.J.; Johnson, P.C.; Niemeier, D.; Pitesky, M.; VanderGheynst, J.S. Cultivation of Black Soldier Fly Larvae on Almond Byproducts: Impacts of Aeration and Moisture on Larvae Growth and Composition. J Sci Food Agric 2018, 98, 5893–5900. [Google Scholar] [CrossRef]
- Singh, A.; Kumari, K. An Inclusive Approach for Organic Waste Treatment and Valorisation Using Black Soldier Fly Larvae: A Review. Journal of Environmental Management 2019, 251, 109569. [Google Scholar] [CrossRef]
- Lemke, N.B.; Dickerson, A.J.; Tomberlin, J.K. No Neonates without Adults: A Review of Adult Black Soldier Fly Biology, Hermetia Illucens (Diptera: Stratiomyidae). BioEssays 2023, 45, 2200162. [Google Scholar] [CrossRef]
- Meneguz, M. , Miranda C.D., Cammack J.A. and J.K. Tomberlin Adult Behaviour as the next Frontier Fo Optimising Industrial Production of Clack Soldie Fly Hermetia Illucens (L.) (Diptera: Stratiomydae).
- Chia, S.Y.; Tanga, C.M.; Khamis, F.M.; Mohamed, S.A.; Salifu, D.; Sevgan, S.; Fiaboe, K.K.M.; Niassy, S.; Van Loon, J.J.A.; Dicke, M.; et al. Threshold Temperatures and Thermal Requirements of Black Soldier Fly Hermetia Illucens: Implications for Mass Production. PLoS ONE 2018, 13, e0206097. [Google Scholar] [CrossRef]
- Harnden, L.M.; Tomberlin, J.K. Effects of Temperature and Diet on Black Soldier Fly, Hermetia Illucens (L.) (Diptera: Stratiomyidae), Development. Forensic Science International 2016, 266, 109–116. [Google Scholar] [CrossRef]
- European Commission, E. (2001). REGULATION (EC) No 999/2001 OF THE EUROPEAN 608 PARLIAMENT AND OF THE COUNCIL of Laying down Rules for the Prevention, Control and Eradication of Certain Transmissible Spongiform Encephalopathies Https://Eur-Lex.Europa.Eu/Legalcontent/EN/ALL/?uri=CELEX%3A32001R09991. 22 May.
- Regulation (EC) No 1069/2009 of the European Parliament and of the Council of Laying down Health Rules as Regards Animal by-Products and Derived Products Not Intended for Human Consumption and Repealing Regulation (EC) No 1774/2002 (Animal by-Products Regulation). 21 October.
- Commission Regulation (EU) No 142/2011 of Implementing Regulation (EC) No 1069/2009 of the European Parliament and of the Council Laying down Health Rules as Regards Animal by-Products and Derived Products Not Intended for Human Consumption and Implementing Council Directive 97/78/EC as Regards Certain Samples and Items Exempt from Veterinary Checks at the Border under That DirectiveText with EEA Relevance. 25 February.
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Dortmans, B.; Diener, S.; Verstappen, B.; Zurbrügg, C. Black Soldier Fly Biowaste Processing.
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Influence of Resources on <I>Hermetia Illucens</I> (Diptera: Stratiomyidae) Larval Development. jnl. med. entom. 2013, 50, 898–906. [Google Scholar] [CrossRef]
- AOAC Official Method 981.10 Crude Protein in Meat.
- Janssen, R.H.; Vincken, J.-P.; Van Den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio Molitor, Alphitobius Diaperinus, and Hermetia Illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- AOAC: Official Methods of Analysis (Volume 1) (15th ed.), Assoc. Off. Anal. Chem., Arlington, VA (1990).
- Standardization of Analytical Methodology for Feeds: Proceedings of a Workshop Held in Ottawa, Canada, 12 - 14 March 1979; Pigden, W.J., Ed.; International Development Research Centre: Ottawa, 1980; ISBN 978-0-88936-217-8.
- Holmes, L. Role of Abiotic Factors on the Development and Life History of the Black Soldier Fly, Hermetia Illucens (L.) (Diptera: Stratiomyidae).
- Van Huis, A.; Oonincx, D.G.A.B. The Environmental Sustainability of Insects as Food and Feed. A Review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of Different Growing Substrates and Processing on the Nutrient Composition of Black Soldier Fly Larvae Destined for Animal Feed. JIFF 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S.; Saitta, V.; Morelli Venturi, D.; Li, C.; Gorb, S. The Armoured Cuticle of the Black Soldier Fly Hermetia Illucens. Sci Rep 2023, 13, 22101. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of Black Soldier Fly (Diptera: Stratiomyidae) Larvae to Recycle Food Waste. Environmental Entomology 2015, 44, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.; Egger, J.; Scheidegger, A.; Zurbrügg, C.; Bruno, D.; Bonelli, M.; Tettamanti, G.; Casartelli, M.; Schmitt, E.; Kerkaert, B.; et al. Estimating Black Soldier Fly Larvae Biowaste Conversion Performance by Simulation of Midgut Digestion. Waste Management 2020, 112, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; Van Loon, J.J.A. Influence of Larval Density and Dietary Nutrient Concentration on Performance, Body Protein, and Fat Contents of Black Soldier Fly Larvae ( Hermetia Illucens ). Entomologia Exp Applicata 2018, 166, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Park,H.BlackSoldierFlyLarvaeManual2016https://scholarworks.umass.edu/sustainableumass_studentshowcase/14.
- Tomberlin, J.K.; Adler, P.H.; Myers, H.M. Development of the Black Soldier Fly (Diptera: Stratiomyidae) in Relation to Temperature. ENVIRONMENTAL ENTOMOLOGY 2009, 38. [Google Scholar] [CrossRef] [PubMed]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of Rearing Substrate on Growth Performance, Waste Reduction Efficiency and Chemical Composition of Black Soldier Fly ( Hermetia illucens ) Larvae. J Sci Food Agric 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Shumo, M.; Khamis, F.; Tanga, C.; Fiaboe, K.; Subramanian, S.; Ekesi, S.; Van Huis, A.; Borgemeister, C. Influence of Temperature on Selected Life-History Traits of Black Soldier Fly (Hermetia Illucens) Reared on Two Common Urban Organic Waste Streams in Kenya. Animals 2019, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, N.; Costa, R.; Ameixa, O.M.C.C. The Influence of Non-Optimal Rearing Conditions and Substrates on the Performance of the Black Soldier Fly (Hermetia Illucens). Insects 2022, 13, 639. [Google Scholar] [CrossRef]
- McEachern, T. Determining Heat Production of Black Solderi Fly Larvae, Hermitia Illucens, to Design Rearing Structures at Livestock Facilities. 2018. [CrossRef]
- Li, C.; Addeo, N.F.; Rusch, T.W.; Tarone, A.M.; Tomberlin, J.K. Black Soldier Fly (Diptera: Stratiomyidae) Larval Heat Generation and Management. Insect Science 2023, 30, 964–974. [Google Scholar] [CrossRef]
Figure 1.
Temperatures maximum and minimum (°C) of rearing substrates in the days of the trial report.
Figure 1.
Temperatures maximum and minimum (°C) of rearing substrates in the days of the trial report.
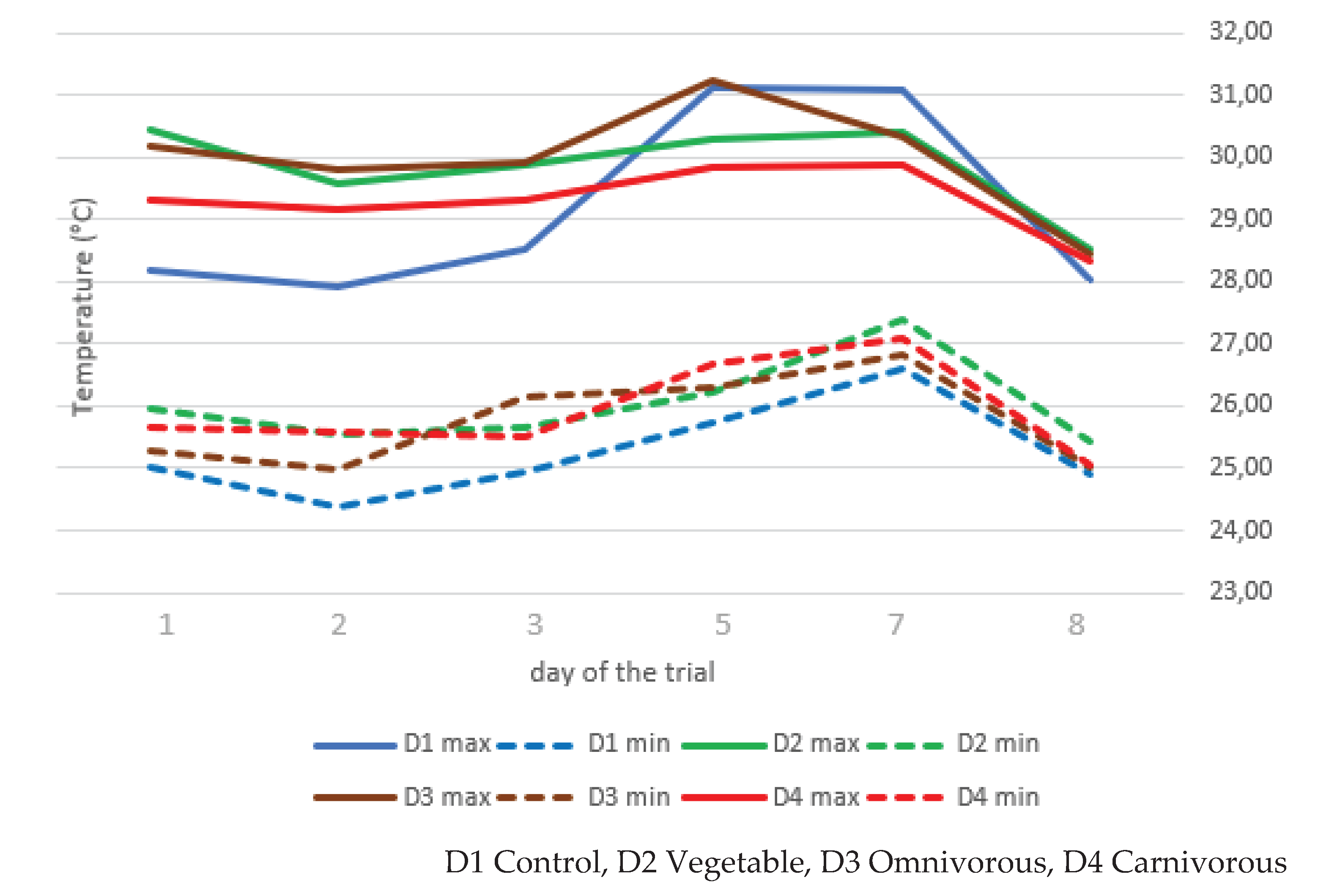
Table 2.
Larval and residual biomass, larvae weight and length, grow rate; substrate reduction, dry matter larvae and larvae meal yield of black soldier fly larvae reared on different substrates.
Table 2.
Larval and residual biomass, larvae weight and length, grow rate; substrate reduction, dry matter larvae and larvae meal yield of black soldier fly larvae reared on different substrates.
| Parameter | D1i | D2 | D3 | D4 | p-Value |
|---|---|---|---|---|---|
| Larval biomass (g)ii | 376.9±34.603A | 278.0±49.558B | 335.1±19.274AB | 305.9±24.212AB | p<0.01 |
| Residual biomass (g) | 380.1±22.533AB | 476.8±±53.485B | 313.5±14.925A | 304.7±26.268A | p<0.01 |
| larvae weight (g) | 0.16±0.024A | 0.10±0.011B | 0.12±0.006B | 0.11±0.014B | p<0.01 |
| Larvae lenght (cm) | 1.75±0.182Aa | 1.40±0.097Bb | 1.46±0.081ABb | 1.43±0.111ABb | p<0.01 |
| Growth rate (%) | 1.60±0.389A | 0.82±0.139B | 1.03±0.074B | 0.94±0.176B | p<0.01 |
| Substrate reduction (%) | 82.72±1.024B | 77.41±2.847A | 84.32±0.746B | 84.77±1.313B | p<0.01 |
| Dry Matter larvae (%) | 37.05±7.132AB | 26.82±5.273B | 35.11±5.504AB | 44.43±3.952A | p<0.01 |
| Larvae meal yield (g DM) | 138.56±21.924B | 76.31±29.348A | 117.52±18.292AB | 135.40±8.242B | p<0.01 |
i D1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous. ii g kg-1 on a wet basis. A, B p<0.01; a, b p<0.05.
Table 3.
Proximate composition (g kg-1 on dry matter) of black soldier fly larvae reared on control (D1), vegetable (D2), omnivorous (D3), and carnivorous (D4) diets.
Table 3.
Proximate composition (g kg-1 on dry matter) of black soldier fly larvae reared on control (D1), vegetable (D2), omnivorous (D3), and carnivorous (D4) diets.
| Black soldier fly larvae | |||||
|---|---|---|---|---|---|
| Parameter | D1i | D2 | D3 | D4 | p-Value |
| Crude Protein | 28.1±0.494ab | 27.3±3.475a | 28.8±3.467ab | 33.0±0.679b | p<0.05 |
| Crude Fat | 26.1±1.184b | 9.7±1.998a | 30.29±2.530b | 25.59±6.976b | p<0.01 |
| Ash | 15.3±0.284c | 10.8±1.385b | 6.1±0.787a | 4.6±0.211a | p<0.01 |
| Crude Fiber | 6.2±0.335ab | 9.1±0.378c | 6.5±0.177b | 5.8±0.297a | p<0.01 |
| Neutral detergent fiber | 12.8±4.427 | 16.3±3.433 | 12.7±2.398 | 11.5±1.254 | p<0.21 |
| Acid detergent fiber | 8.9±0.666 | 15.1±2.638 | 12.9±5.333 | 13.9±5.133 | p<0.18 |
| Acid detergent lignin | 1.1±0.076 | 2.34±0.268 | 2.5±1.087 | 2.1±1.037 | p<0.10 |
| Non-Free extractive ii | 24.3±0.704a | 43.1±4.786c | 28.2±3.492ab | 31.0±7.043b | p<0.01 |
iD1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous. ii Calculated as 100- [ crude protein+ fat+ash+fiber] A, B p<0.01; a, b p<0.05.
Table 4.
Chemical composition (g kg-1 on a wet basis) of final substrate and degradation (%) of the substrate: D1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous diet.
Table 4.
Chemical composition (g kg-1 on a wet basis) of final substrate and degradation (%) of the substrate: D1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous diet.
| Degradation (%) | |||||
|---|---|---|---|---|---|
| Parameter | D1 | D2 | D3 | D4 | p-Value |
| DM | 45.90±4.52A | 58.36±4.29B | 60.51±2.02B | 57.72±2.40B | p<0.01 |
| OM | 50.81±4.31A | 58.99±4.18B | 61.33±2.27B | 59.18±2.64B | p<0.01 |
| CP | 54.38±5.78Aa | 63.69±5.76ABb | 71.78±3.71Bb | 59.91±2.32Aab | p<0.01 |
| CF | 96.41±1.89B | 89.90±3.38B | 91.05±3.24B | 64.01±5.70A | p<0.01 |
| Ash | 15.84±9.71A | 49.16±5.82B | 49.32±4.86B | 34.69±3.82B | p<0.01 |
| Starch | 42.45±4.36B | 34.59±12.62B | 40.35±5.45B | 98.06±0.37A | p<0.01 |
| NFE | 49.59±4.55A | 61.44±4.60B | 43.61±3.79A | 52.68±3.67Ab | p<0.01 |
D1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous; DM: dry matter; OM= organic matter; CP=crude protein; CF=crude fat; NFE= N-free extractive; A, B p<0.01.
Table 5.
Maximum, minimum and mean accumulated degree hours (ADH, LDT= 12°C) required by BSFL.
| Temperature | D1 | D2 | D3 | D4 | p-Value |
|---|---|---|---|---|---|
| MAX | 3359.5±177.686 | 3430.24±187.64 | 3455.98±139.40 | 3314.7±104.66 | p=0.27 |
| MIN | 2568.4±101.98 | 2713.1±121.02 | 2646.6±64.06 | 2690.1±69.47 | p=0.10 |
| MEANS | 2964±136.32 | 3071.7±149.78 | 3051.3±98.14 | 3002.4±82.94 | p=0..29 |
MAX=ADG in relation to the maximum substrate temperature; MIN= ADG at the minimum substrate temperature; MEANS= ADG in relation to the average of the minimum and maximum substrate temperatures; Temperature max, min and means are reported in degrees (°C); D1 Control, D2 Vegetable, D3 Omnivorous, D4 Carnivorous.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



