
Submitted:
20 March 2024
Posted:
21 March 2024
You are already at the latest version
Abstract
Bacterial endophytes (120) were isolated from six halophytes (Distichlis spicata, Cynodon dactylon, Eragrostis obtusiflora, Suaeda torreyana, Kochia scoparia, and Baccharis salicifolia). These halophiles were molecularly identified and characterized with or without NaCl conditions. Characterization was based on tests such as indole acetic acid (IAA), exopolysaccharides (EPS), and siderophores (SID) production; solubilization of phosphate (P), potassium (K), zinc (Zn), and manganese (Mn); mineralization of phytate; enzymatic activity (acid and alkaline phosphatase, phytases, xylanases, and chitinases) and mineralization/solubilization mechanisms involved (organic acids and sugars). Eleven halophiles (tolerated NaCl 2.5 M) were identified as Oceanobacillus sp. (1), Bacillus sp. (2), Nesterenkonia sp. (3), Bacillus velezensis (4), Halobacillus sp. (5), Oceanobacillus sp. (6), B. velezensis (7) Halomonas sp. (8), B. pumilus (9), Pseudomonas sp. (10) and Oceanobacillus sp. (11). Biochemical responses of nutrient solubilization and enzymatic activity varied between bacteria and the presence of NaCl. Organic acids were involved in P-mineralization and nutrient solubilization. Tartaric acid was common in the solubilization of P, Zn, and K. Maleic and vanillic acid were only detected in Zn and K solubilization, respectively. Moreover, sugars appeared to be involved in the solubilization of nutrients; fructose was detected in solubilization tests. Therefore, these bacteria should be tested as a consortium to mitigate saline stress in glycophytes under a global climate change scheme that threatens to exacerbate soil salinity.
Keywords:
plant growth-promoting bacteria
; solubilization
; salinity
; enzyme activity
; organic acids
; food safety
; plant fitness.
1. Introduction
Salinity affects 20% of the world’s agricultural soils and is one of the leading causes of yield reduction in crops of economic interest, most of which are sensitive to salinity [1,2]. Soil salinity and sodicity will increase considerably due to climate change, so they have become a problem of global concern. Salts stress decreases the yield, nutrient content, and nutraceutical quality of food from crops of economic interest, which can cause deficiencies in humans and favor hidden hunger. Halophyte plants are essential for creating saline ecosystems due to their high tolerance and remediation potential [3]. These plants have evolved several strategies to survive in these environments, including increased cytoplasmic osmotic pressure, production of compatible solutes, exclusion of sodium ions from the cells, or their accumulation in the vacuole [4]. Another alternative used by halophyte plants to attenuate the effect of salt stress is related to their microbiome.
For decades, most research focused on rhizobacteria from saline soils and isolated from glycophytic crops (salinity-sensitive plants) [5,6]. However, more recently, research has focused on endophytic bacteria and their isolation from halophytes. Conventionally, endophytes are defined as bacteria or fungi that reside intercellularly in plant tissues and do not cause negative effects on plant growth [7]. Endophytes have advantages over rhizospheric ones because they are protected from biotic and abiotic environmental challenges. Therefore, endophytes are a potential tool for improving plant growth and yield under salt-stress conditions. Studies that have isolated endophytic bacteria from halophytes suggest increased adaptation and survival of these bacteria in saline. Moreover, inoculation with endophytic and high salinity-tolerant (halophilic) bacteria increases the tolerance of plants grown in saline soils [8].
Halophilic endophytic bacteria may have similar properties to rhizobacteria. For example, they solubilize nutrients [such as phosphorous (P), potassium ( K), zinc (Zn) or manganese (Mn)] to improve the efficiency of nutrient uptake, fix atmospheric nitrogen (N2), produce exopolysaccharides (EPS) and siderophores [iron (Fe) chelation], and modulate the level of phytohormones within plant tissues [9]. Endophytic bacteria can also synthesize plant hormones (auxins, cytokinins, abscisic acid, and gibberellins) that modify plant physiology to resist stress conditions [10].
Salinity is known to interfere with the availability of Fe, N, and other essential plant elements [11]. Endophytic bacteria can also secrete siderophores, which are high-affinity Fe-chelating compounds, so plants can easily access Fe-siderophore complexes for nutrition [9]. Some endophytic bacteria are N2-fixers and represent an important source of available N in saline-sodic soils. Phosphorus is an essential element for plants, but its availability is commonly low in saline soils. P-solubilizing and -mineralizing bacteria improve P nutrition in plants through chelation, ion exchange, acidification (by secretion of low molecular weight organic acids), and enzyme production (phosphatases and phytases) [12,13,14]. However, salinity is one of the main factors negatively influencing P solubilization by bacteria [15].
In saline-sodic soils, pH can decrease the availability of elements that play crucial metabolic roles in plants, such as micronutrients. These micronutrients acting as cofactors for several enzymes, are vital for plant development [16]. Their deficiencies can significantly impact crops of economic interest [17]. As mentioned, bacteria have the potential to solubilize other nutrients like K, Zn, or Mn. However, the processes and mechanisms involved in this solubilization remain poorly understood.
Enzyme production (hydrolytic proteases, cellulases, hemicellulases, xylanases, chitinases, pectinases, glucanases, and pectinases) is another characteristic of endophytic bacteria [9]. Some of these extracellular enzymes help plants establish systemic resistance against invasion by phytopathogenic bacteria or fungi. This information highlights the need to inoculate glycophytic crops with microorganisms from saline soils or halophytes. However, there is limited information on the effect of the presence or absence of NaCl on the plant growth-promoting properties of halophilic endophytic bacteria. There is also little information regarding compatible and functional bacterial consortia that can promote plant growth and mitigate salt stress.
Utilizing microbial consortia as an inoculum, particularly those derived from extreme environments, can be effective biotechnology tool [18,19]. Thses consortia by functionally complementing each other, have the potential to significantly enhance plant growth [20]. The research by Vaishnav et al. [21] underscores the importance of harnessing bacterial inoculants from saline environments for sustainable agriculture, addressing plant salt stress. Marghoob et al. [22] and Rajput et al. [23] have demostrated the effectiveness of consortiums of Aeromonas sp. and A. salmonicida, or Pantoea sp. and Erwinia rhapontici, in promoting wheat growth. Similarly, the bacterial consortium of Lysinibacillus sp. and Bacillus sp. encouraged tomato growth [24]. Therefore, The production of consortia-based bacterial inoculants from halophytes may be useful for salt-sensitive plants.
The objectives of the present research were: 1) To isolate endophytic bacteria from the roots of seven halophytes established in saline soils. 2) To biochemically characterize endophytic bacteria selected for their halotolerance under two salinity conditions (0 M and 2.5 M NaCl) for potential use in plant growth promotion. 3) To determine the production of organic acids and sugars in selected halophilic bacteria in P, Zn, K, and Mn solubilization. 4) To evaluate the activity of four extracellular enzymes in the halophilic endophytic bacteria. 5) To determine bacterial compatibility to form consortia for future use in salinity mitigation in glycophytic plants.
2. Results and Discussion
2.1. Isolation of Endophytic Bacteria by Halotolerance
A total of 120 endophytic bacterial isolates were obtained from halophyte roots. Fifty-eight endophyte isolates were tolerant to 0.1 M NaCl, 28 to 0.5 M NaCl, 22 to 1.5 M NaCl, and 11 to 2.5 M NaCl. The latter 11 isolates were considered halophilic based on the classification of salinity-tolerant microorganisms [25]; therefore, genomic analysis was performed on these isolates.
2.2. Molecular Identification
Phylogenetic analysis of the DNA sequences generated by 16S rRNA gene sequencing using NCBI databases revealed that each isolate corresponds to a single species, except isolates 4 and 7 (Figure 1). The BLAST algorithm for the sequences showed that isolate 1 corresponds to Oceanobacillus sp., isolate 2 to Bacillus sp., isolate 3 to Nesterenkonia sp., isolate 4 to Bacillus velezensis, isolate 5 to Halobacillus sp., isolate 6 to Oceanobacillus sp., isolate 7 to Bacillus velezensis, isolate 8 to Halomonas sp., isolate 9 to Bacillus pumilus, isolate 10 to Pseudomonas sp., and isolate 11 to Oceanobacillus sp., with degrees of proximity of 99% (Table 1).
Bacteria of the genus Oceanobacillus have been reported as halotolerant rhizobacteria [26] but not as an endophyte of halophytes. Mukhtar et al. [27] identified the rhizobacterium Halobacillus isolated from the halophyte Atriplex sp. The genus Nesterenkonia sp. was reported as a halotolerant endophyte isolated from the halophyte tree Populus euphratica. Moreover, species of the genus Halomonas have been reported as halophilic endophytes from the halophyte Arthrocnemum macrostachyum [28]. The bacteria B. velezensis and B. pumilus were previously reported as endophytes of non-halophyte plants . In addition, Szymanska et al. [31] described Pseudomonas stutzeri as a halophytic endophyte isolated from the halophyte Salicornia europea. This is the first report of B. velezensis and B. pumilus as endophytes isolated from halophytes.
2.3. Indolacetic Acid and Siderophore Production
All the tested bacterial isolates produced IAA. Bacteria growing in 2.5 M NaCl spiked media showed a significant increase in IAA production (p ≤ 0.001), except in isolates 10 (Pseudomonas sp.) and 11 (Oceanobacillus sp.) (Figure 2a). The maximum concentration of IAA was observed in Oceanobacillus sp. and B. velezensis (19.96 mg L-1 and 20.55 mg L-1, respectively). In contrast, the maximum concentration in the absence of NaCI was 12.81 mg L-1 in isolate 11 (Oceanobacillus sp.). These results are consistent with those reported by Mahgoub et al. [32], who evaluated the production of IAA in 18 endophytic bacteria isolated from halophytes. These authors observed that all bacteria produced more IAA when grown in a nutrient medium with 0.6 M NaCI than without NaCI. Other authors [33,34,35] observed that halotolerant endophytic bacteria (Marinilactibacillus kalidii, Stenotrophomonas pavanii, and B. licheniformis) isolated from halophytes produced 2.46 mg L-1, 20.5 mg L-1, and 27.0 mg L-1 of AIA, respectively in absence of NaCl. In the present investigation, isolate 7 (B. velezensis), had similar IAA production in the absence of NaCl to that of S. pavanii [34]. The isolation of halophilic endophytic bacteria is relevant because they are more adapted to salinity, and the presence of NaCl does not inhibit their capacity to produce auxins. IAA is a significant growth regulator for salt-stressed plants because it controls several growth processes, such as stem elongation, cell division, response to light and gravity, and cell differentiation [36].
Previous research has qualitatively shown that halotolerant endophytic bacteria produce siderophores [35,37,38], but few studies have tested this ability under saline conditions. Results from the present research show that isolates 1 (Oceanobacillus sp.), 2 (Bacillus sp.), 6 (Oceanobacillus sp.), 7 (B. velezensis), and 10 (Pseudomonas sp.) increased siderophore production in the saline medium compared to the control (Figure 2b). In the rest of the bacteria, the percentage of siderophores was similar with and without NaCl. Isolate 8 (Halomonas sp.) produced the highest percentage of siderophores (71% - 73%) in both salinity conditions. Such siderophore production was higher than that reported by Panwar et al. [39] for the halotolerant rhizobacterium Enterococcus faecium, which produced 60.33% in the absence of NaCl and 54.67% with 0.15 M NaCl; conversely, in Pantoea dispersa, the production was 32.11% and 39.23%, respectively. Endophytic bacteria produce siderophores, which improve Fe supply and enhance plant growth in saline soils with low Fe availability [40,41,42]. Therefore, endophytic bacteria could help solve this severe agricultural problem.
2.4. Nitrogenase Enzyme Activity and Exopolysaccharide Production
Isolate 3 (Nesterenkonia sp.), 4 (B. velezensis), 8 (Halomonas sp.), 7 (B. velezensis), and 9 (B. pumilus) had diazotrophic activity in the absence of NaCl (Table 2), which means that 45% of the bacteria presented this characteristic. In the presence of 2.5 M NaCl, isolates 3, 4, 8, 7, 9, and 10 showed diazotrophic activity. Shurigin et al. [43] found 5 of 20 bacteria isolated from the halophyte Haloxylon aphyllum fixed N2 in a semisolid medium without NaCl. Mahgoub et al. [32] observed that all 11 bacteria isolated from the halophyte A. macrostachyum fixed N2 in the absence of NaCl and in a saline medium (0.6 M NaCl). Enquahone et al. [44] observed that 81% of the halotolerant endophytic bacterial isolates from Sporobolus specatus (21 isolates in total) were positive for N2 fixation. Using endophytic bacterial isolates 3, 4, 8, 7, 9, and 10 with diazotrophic capacity could be a promising strategy to improve the growth of salt-sensitive plants.
Only 4 of 11 halophilic endophytic bacterial isolates were positive for EPS production (1, 4, 5, and 6) (Table 2). Remarkably, only isolate 4 (B. velezensis) was positive for both EPS production and N2 fixation. In recent years, halotolerant bacteria, particularly rhizobacteria, have been documented to improve plant salinity stress through EPS production [41]. EPS produced by halotolerant rhizobacteria can chelate free Na from the soil, restrict Na entry into plants, support biofilm formation, and improve soil stability. EPS production and biofilm formation are essential characteristics of endophytic bacteria under salinity stress conditions. Due to the ability to bind cations, bacterial EPS restricts the Na available for plant uptake and protects the root from high salt concentrations [45]. This study is among the pioneering works exploring EPS production in halophytic endophytes. Future research should address the production and functional characterization of EPS from endophytes, emphasizing the role of EPS in endophyte-plant interactions under saline conditions in halophyte and salinity-sensitive plants.
2.5. Nutrient Solubilization, Bacterial Growth, and pH Modification
2.5.1. Inorganic and Organic P
The ability of bacteria to solubilize P from Ca3(PO4)2 was decreased by the presence of NaCl (Figure 3a). The highest P solubilization in the absence of NaCl was observed for isolates 3 (Nesterenkonia sp.), 5 (Halobacillus sp.), 6 (Oceanobacillus sp.), and 10 (Pseudomonas sp.) with 15.46 mg P L-1, 15.88 mg L-1, 15.84 mg L-1, and 15.76 mg L-1, respectively. The maximum P solubilization was 11.05 mg L-1 in isolate 10 at 2.5 M NaCl.
Shahid et al. [46] reported that the non-halotolerant endophytic bacterium Priestia aryabhattai isolated from wheat enhanced P solubilization with increasing NaCl concentrations. The reported solubilization was 20 mg L-1 in the presence of 0.8 M NaCl and 40 mg L-1 in 2.5 M NaCl. Moreover, Serratia rubidaea, an endophyte isolated from the halophyte Chenopodium quinoa, solubilized 350.63 mg L-1 of P in the presence of 1.3 M NaCl [37]. Although the reported P concentration is higher than in the present study, the NaCl concentration tested by these authors was approximately half, and the analysis period was ten days instead of 48 h tested in the present study. Mahgoub et al. [32] observed that the endophytic bacterial strains BR1 (B. subtilis) and AR5 (B. thuringiensis), isolated from the halophytes A. macrostachyum and Spergularia marina, produced 211.58 mg L-1 and 182.48 mg L-1 of P, respectively in the presence of 0.6 M NaCl; however, the NaCl concentration used was four times lower than that of the present study.
In the solubilization P test, the bacteria acidified the culture medium. The average pH of the uninoculated NBRIP medium was 6.82 with and 6.86 without NaCl (Figure 3b). After 48 h of incubation, the average pH of the cultures was pH=5.4 without NaCl and pH=5.9 with NaCl. Therefore, P solubilization may be partially due to acidification; this was corroborated by positive correlations between soluble P concentration and pH with (r=0.75) and without NaCl (r=0.91). The lowest pH in the medium was observed with isolate 10 (Pseudomonas sp.) at both NaCl concentrations (pH=4.3 in the absence of NaCl and pH=4.6 with 2.4 M NaCl). This strain showed one of the highest P solubilization values (Figure 3a). Mahdi et al. [37] observed a decrease in pH (to 3.04 and 5.9) without NaCl and with 1.3 M NaCl. In the present study, no drastic pH changes were observed in the tests without salt, as observed by these authors.
In all solubilization tests, no differences in biomass were obtained among bacteria in the absence or presence of NaCl (Table S1). Moreover, the presence or absence of NaCl did not influence bacterial growth in all solubilization tests. Overall, bacterial biomass did not correlate with the solubilization capacity of the different studied elements.
This research shows the first report on the influence of NaCl on the production and concentration of organic acids of halophilic endophytic bacteria that solubilize inorganic P (Table 3). All bacteria secreted citric and succinic acids in the presence and absence of NaCl. Isolate 4 (B. velezensis) produced the highest concentration of citric acid, 15.5 mg L-1 with 2.5 M NaCl and 15.2 mg L-1 without NaCl. At the same time, no differences were observed among bacteria in succinic acid production in both salt conditions. In the absence of NaCl, all bacteria produced lactic acid, whereas in the presence of salt, no acid was produced by isolates 3 (Nesterenkonia sp.), 8 (Halomonas sp.), and 9 (B. pumilus). In the presence of NaCl, Oceanobacillus sp. isolates (1 and 10) produced the highest concentration of lactic acid (Table 3) compared to the other bacteria. NaCl increased lactic acid concentration only in isolate 6 (Oceanobacillus sp.), 7 (B. velezensis), 8 (Halomonas sp.), 9 (B. pumilus), and 10 (Pseudomonas sp.). Tartaric acid production was detected in isolates 3 (Nesterenkonia sp.) and 9 (B. pumilus), both in the absence and presence of NaCl, and no differences were observed in the tartaric acid production among these bacteria. Isolate 1 (Oceanobacillus sp.), 2 (Bacillus sp.), and 8 (Halomonas sp.) secreted higher amounts of citric acid in the presence of NaCl. However, NaCl did not increase the concentration of succinic acid, except in isolate 8 (Halomonas sp.) (Table 3). Paredes-Mendoza and Espinosa-Victoria [47] indicated that the predominant organic acids reported in the solubilization of inorganic P are succinic, citric, lactic, and tartaric. In this research, fumaric, oxalic, vanillic, malic, salicylic, and maleic acids were not detected in the extracts produced by the bacteria in the solubilization of nutrients.
This study is also the first to evaluate the mineralization of organic P from phytic acid in the absence and presence of NaCl. The results showed that, as in phosphate solubilization from Ca3(PO4)2, organic P mineralization was reduced in the presence of NaCl in all bacteria except isolates 1 (Oceanobacillus sp.) and 4 (B. velezensis). Bacterial isolates produced higher phosphate concentrations when phytic acid was used than with Ca3(PO4)2 (Figure 3c). Without NaCl, the average soluble phosphate concentration from phytic acid was 75% higher than with Ca3(PO4)2. Isolates 3 (Nesterenkonia sp.) and 9 (B. pumilus) showed the highest organic P mineralization in the presence of NaCl, but in the absence of NaCl, isolate 5 showed the highest (Halobacillus sp.) (Figure 3c). Insoluble organic P accounts for approximately 65% of total P in saline soils, while inorganic P accounts for 35%. Organic P is presented in different forms, including inositol phosphate, phosphomonoesters, phosphodiesters (phospholipids and nucleic acids), and phosphotriesters [48]. Therefore, the P mineralization process exhibited by some microorganisms, such as bacteria, is essential in saline soils. Bacteria mobilize insoluble organic P through mineralization and convert it into available P in the soil [49].
The results are relevant since most research has focused on evaluating solubilization and mineralization in halotolerant rhizobacteria, but limited information has been generated with halophilic endophytic bacteria. In this context, Liu et al. [50] reported that three halotolerant rhizobacteria (Pseudarthrobacter, Acinetobacter, and Pseudomonas) solubilized more inorganic P (Ca3(PO4)2) than organic P (phytic acid) under salt-free conditions. Other investigations highlight the mineralization of organic P in non-halotolerant rhizobacteria. For example, Rasul et al. [51] reported that Pantoea sp. solubilized (20 mg L-1) and mineralized P (100 mg L-1), while Ochrobactrum sp. was able to solubilize P (60 mg L-1) and mineralize P (150 mg L-1). In the present study, pH was not significantly modified during P mineralization with and without NaCl, contrary to what was observed with inorganic P. The average pH in the cultures without NaCl was 6.7, while in the presence of NaCl, the average pH was 6.8. The original pH of the Luria Bertani (LB) broth without NaCl was 7.0, and with NaCl, it was 7.3 (Figure 3d). P mineralization occurs by mechanisms other than the production of organic acids, which modify the pH of the culture medium [52]. For example, phytases, a subset of phosphatases, gradually dephosphorylate phytate to produce inositol and soluble P [53]. Alkaline and acid phosphatases also mineralize P [48]. The results of the present study suggest that halophilic endophytic bacteria have a high potential for application in cropping systems with organic fertilization and salinity problems.
2.5.2. Solubilization of K, Mn, and Zn
Mineral soils generally contain between 0.04% and 3% of K in the first 20 cm of soil. Ninety-eight per cent of the total K is mineral, unavailable to plants and microorganisms. Soil microorganisms use mechanisms to solubilize minerals (illite, micas, and feldspars) containing K, which plants can absorb [54]. Studies that use K-solubilizing bacteria to inoculate salinity-sensitive crops are important because this element can compete with high Na concentrations in the soil. The results of this research show that isolates 2 (Bacillus sp.), 4 (B. velezensis), 5 (Halobacillus sp.), 6 (Oceanobacillus sp.), and 8 (Halomonas sp.) increased their ability to solubilize K in the presence of NaCl. However, in the other isolates, solubilization was independent of NaCl. Isolate 4 was the most efficient in K solubilization (57.14 mg L-1) with salt, therefore, it could be used to improve the availability of this element in salinity-sensitive plants. In the non-saline medium, isolate 7 (B. velezensis) had the highest K solubilization (14.53 mg L-1; Figure 4a). Some studies have qualitatively evaluated K solubilization with endophytic bacteria isolated from halophytes [55,56]. However, the present research is the first one quantifying the K solubilization capacity of halophilic endophytic bacteria isolated from halophyte roots.
Recently, Ranawat et al. [57] observed that the halotolerant rhizobacterium E. hormaechei solubilized 97.5 mg de K L-1 in the absence of NaCl. Other investigations with halotolerant rhizobacteria have considered the presence of NaCl. Ashfaq et al. [58] found that K solubilization in the presence of NaCl decreased in 13 halotolerant rhizobacteria studied; the maximum K solubilization was 22.33 mg L-1 in the absence of NaCl and 16.58 with 0.7 M NaCl. The present investigation found a correlation between pH and K solubilization in both salinity conditions (r=0.74).
Our understanding of the mechanisms employed by halophilic endophytic bacteria e to solubilize K remains limited. However, the most well-known mechanisms include pH decrease through the production of organic acids and proton release. All bacterial isolates diminished the pH of the culture medium (Figure 4b), but B. velezensis (4) reduced it more significantly to 5.1. The organic acids produced to dissolve potassium minerals (illite and feldspar) are oxalic, gluconic, tartaric, 2-ketogluconic, citric, malic, succinic propionic, lactic, acetic, glycolic, malonic, and fumaric acids [59]. All bacteria evaluated in this investigation produced tartaric, malic, and citric acids, except isolate 8 (Pseudomonas sp.), which also produced vanillic acid. Vanillic acid production was previously reported in other Pseudomonas sp. strains but had not been related to the solubilization of K. Moreover, vanillic acid has been associated with resistance against phytopathogens [60]. Therefore, it is necessary to test whether isolate 8 (Pseudomonas sp.) can be used in biocontrol.
The isolates did not produce fumaric, oxalic, maleic, salicylic, succinic, lactic, and acetic acids (Table 4). Isolate 4 (B. velezensis) had the highest concentration of tartaric acid in saline condition (10.8 mg L-1), while in non-saline medium, isolate 7 did (B. velezensis), with 8.8 mg L-1. Isolate 9 (B. pumilus) produced the highest concentration of malic acid with and without NaCl (8.7 mg L-1 and 7.6 mg L-1, respectively). Also had the highest concentration of citric acid (Table 4). Salinity influenced the concentration of some organic acids, and this effect was different among isolates. Isolates 1 (Oceanobacillus sp.), 4 (B. velezensis), 7 (B. velezensis), and 10 (Pseudomonas sp.) produced more citric acid in the presence of salt. In addition, isolates 2 (Bacillus sp.), 3 (Nesterenkonia sp.), 9 (B. pumilus), and 10 (Pseudomonas sp.) produced more malic acid. Tartaric acid increased in the presence of salt in isolate 1 (Oceanobacillus sp.), 4 (B. velezensis), 5 (Halobacillus sp.), 6 (Oceanobacillus sp.), 7 (B. velezensis), 8(Halomonas sp.), 10 (Pseudomonas sp.), and 11 (Oceanobacillus sp.), while in isolate 3 (Nesterenkonia sp.) the concentration of tartaric acid decreased in the presence of salt.
Adding NaCl to the medium decreased Mn solubilization by Halomonas sp., Nesterenkonia sp., Pseudomonas sp., and Oceanobacillus sp., but not by the other bacteria (Figure 4c). Isolate 5 (Halobacillus sp.) solubilized the highest concentration of Mn with and without salt (1.36 and 1.30 mg L-1, respectively). The genus Halobacillus had been reported as growth promoters through P solubilization, but there were no previous reports of their role in Mn solubilization. Therefore, this is the first report indicating the role of Mn solubilization in halophilic endophytic bacteria. However, Yamaji et al. [61] and Dixit et al. [62] qualitatively analyzed the ability of non-halotolerant endophytic bacteria to solubilize Mn. Microorganisms reduce Mn4+ to Mn2+ through the production of protons and electron-transporting reducing agents that are oxidized [63], as shown below in Eq. 1. This equation represents the Mn2+ release reaction, in which there is no change in the oxidation state.
Only the research of Ijaz et al. [64] reported that the non-halotolerant rhizobacterium Bacillus sp. isolated from maize solubilized 10.73 mg Mn L-1 from MnO2 at 48 h. Soil salinity and sodicity influence Mn biogeochemistry [65,66].
All endophytic bacteria acidified the culture medium under both salinity conditions (Figure 4b, d, f). These results agree with other reports on Mn solubilization by pH decrease [63]. However, unlike P, K, and Zn solubilization, no correlation was observed between pH and Mn solubilization (r=0.21) in either salinity condition. Another solubilization mechanism may occur by utilizing MnO2 instead of oxygen as a final electron acceptor in the bacterial respiratory chain [67]; however, this mechanism remains to be tested in the bacteria evaluated in this study.
Ijaz et al. [64] reported that the following organic acids were involved in Mn solubilization when using MSB as a growth medium: formic, oxalic, salicylic, pyruvic, citric, and malic. In the present investigation, only citric and malic were detected (Table 4). In the present research, LB broth was used. Hence, differences in results may be due to different growth media used for this test. Similar to the solubilization of inorganic P and K, all bacteria secreted citric acid in the absence and presence of NaCl. Isolate 10 (Pseudomonas sp.) produced the highest concentration of citric acid with and without NaCl (15.8 and 16.0 mg L-1, respectively). None of the isolates produced fumaric, oxalic, lactic, acetic, succinic, tartaric, vanillic, or salicylic acids. Only isolates 1 (Oceanobacillus sp.), 2 (Bacillus sp.), 3 (Nesterenkonia sp.), and 9 (B. pumilus) produced malic acid in the absence and presence of NaCl. In the absence of NaCl, isolate 9 (B. pumilus) produced the highest concentration of malic acid (7.8 mg L-1), whereas, in the presence of NaCl, isolates 1 (Oceanobacillus sp.) and 2 (Bacillus sp.) produced the highest concentration of this acid (7.4 and 7.2 mg L-1, respectively). The presence of NaCl only increased citric acid concentrations in isolates 1 (Oceanobacillus sp.), 2 (Bacillus sp.), and 8 (Halomonas sp.), which showed similar values. In contrast, isolates 1, 2, and 9 increased their malic acid production in the presence of NaCl.
ZnO solubilization showed differences depending on the presence of salt and the bacterial strain (Figure 4e). Isolate 8 (Halomonas sp.) solubilized more Zn (13.33 mg L-1) in the presence of NaCl than the other isolates. In the absence of NaCl, isolate 4 (B. velezensis) had a higher solubilization capacity (107 mg L-1) than the other bacteria. Regarding the effect of NaCl, Isolate 2 (Bacillus sp.), 4 (B. velezensis), 5 (Halobacillus sp.), and 11 (Oceanobacillus sp.) solubilized more Zn in the absence of NaCl than in 2.5 M NaCl. These values are higher than those observed by Fatima et al. [68] in halotolerant rhizobacteria Alcaligenes AF7; this bacterium solubilized Zn to 2.79, 3.26, and 2.8 mg L-1 with 0, 0.3, and 0.7 M NaCl, respectively. According to Tewari and Arora [69] saline stress may be an important factor for nutrient solubilization in some halotolerant bacterial isolates.
The present investigation suggests that the ability of bacteria to solubilize Zn is relevant for agriculture since the availability of this element is fundamental for plant nutrition and the activity of several enzymes. Organic acid production may be one of the mechanisms involved in Zn solubilization [70], associated with decreased pH. The main acids involved in this process are oxalic, ferulic, caffeic, gallic, syringic, citric, 2-ketogluconic, gluconic, tartaric, maleic, and fumaric acids [71]. In this study, maleic y malic acids were produced by bacteria in both NaCl conditions. In contrast with P, K, and Mn solubilization, only bacterial strains 5 (Halobacillus sp.), 8 (Halomonas sp.), and 10 (Pseudomonas sp.) produced citric acid upon Zn solubilization (Cuadro 4). Isolates 1 (Oceanobacillus sp.), 2 (Bacillus sp.), 3 (Nesterenkonia sp.), 7 (B. velezensis), 8 (Halomonas sp.) and 11 (Oceanobacillus sp.) were the only ones that secreted tartaric acid regardless of NaCl concentrations.
No isolate produced fumaric, lactic, acetic, succinic, ferulic, or salicylic acid (Table 4). Isolates 4 (B. velezensis), 8 (Halomonas sp.), and 9 (B. pumilus) secreted the highest concentration of maleic acid in the presence of NaCl. In contrast, without NaCl, the highest concentration was secreted by isolate 4 (11.8 mg L-1). Isolate 8 produced the highest concentration of tartaric acid, 11.8 mg L-1 with salt and 11.6 mg L-1 without salt. The production of organic acids involved in Zn solubilization has been reported in non-halotolerant rhizobacteria. Upadhyay et al. [71] detected oxalic, maleic, tartaric, and fumaric acids in rhizobacterial ZnO solubilization. Li et al. [72] also identified oxalic, formic, tartaric, and acetic acids in the solubilization of ZnO by rhizobacteria, but the information is scarce for halotolerant endophytes. The present work is the first to identify malic acid production in Zn solubilization. In bacteria that secreted citric and tartaric acid, there was no difference in the concentration of these acids in the presence and absence of salt. In contrast, the presence of salt increased the concentration of malic acid in isolates 2 (Bacillus sp.), 3 (Nesterenkonia sp.), 4 (B. velezensis), 6 (Oceanobacillus sp.), and 7 (B. velezensis). The concentration of maleic acid change in the presence of salt in isolates 1 (Oceanobacillus sp.), 5 (Halobacillus sp.), 7, 8 (Halomonas sp.), 9 (B. pumilus), and 10 (Pseudomonas sp.) compared to the absence of salt.
2.5.3. Interaction between Organic Acids and the Solubilization Ion
Research to evaluate the solubilization capacity of bacteria generally focuses on a single ion, while few evaluate the production of organic acids involving two or more solubilizing ions. In the present work, some organic acids produced by the bacteria were common in the solubilization of different ions (Figure 5). In the solubilization of P, Mn, and K, citric acid was common and produced by all bacteria regardless of the presence of NaCl. In Zn solubilization, citric acid was detected only in four isolates. Succinic and lactic acids were only identified in the inorganic P solubilization. Tartaric acid was observed in the solubilization of P, Zn, and K, but not Mn. Malic acid was detected in the solubilization of Zn, Mn, and K, but not P. Maleic acid was only observed in the solubilization of Zn. Vanillic acid was only produced by isolate 8 (Halomonas sp.) during K solubilization. Future research should investigate the mechanisms involved in the solubilization of different nutrients.
2.5.4. Production of Sugars in Nutrient Solubilization Tests
Research on bacteria´s nutrient solubilization capacities has tested the effect of different carbon sources (glucose, fructose, and sucrose); however, no work has evaluated the exudation of sugars in the nutrient solubilization nor the presence or absence of NaCl. Thus, this research is the first to identify sugars present in the solubilization media of different nutrients (inorganic and organic P, K, Zn, and Mn) in the influence of NaCl. Zhang et al. [73] indicated that fructose is not only a carbon source but also a signalling molecule that triggers bacterially mediated mineralization processes of organic P. These authors observed that fructose stimulates the expression of phosphatase genes in bacteria and the rate of phosphatase release into the growth medium by regulating their protein secretory system. Importantly, in the present research, bacteria had both acid and alkaline phosphatase enzymatic activity. Although the highest fructose production was observed in the presence of inorganic P, some hypotheses exist regarding the role of fructose in this nutrient´s mineralization. In addition, in the present work, a positive correlation between fructose production and acid phosphatase activity was observed in 0 (r=0.62) and 2.5 M NaCl (r=0.64); however, this was not the case for alkaline phosphatase.
The presence or absence of salt impacted fructose concentrations depending on the solubilization ion and the bacterial strain (Table 5). Fructose production was common in all isolates except isolate five during Ca3(PO4)2 solubilization. In contrast, xylose production depended on the bacterial strain-none of the isolates produced arabinose, trehalose, maltose, sucrose, lactose, or xylose. Glucose was detected in the solubilization extracts but not reported since it was used in the culture broths as a carbon source. The highest average concentration of fructose was observed in K solubilization compared to the other solubilization ions, both in the absence and presence of NaCl. During the solubilization of inorganic P without NaCl, the average fructose concentration was 85 µg µL-1, and it doubled (164 µg µL-1) in the condition with NaCl. The two isolates of B. velezensis (isolates 4 and 7) produced the highest concentration of fructose both in the absence (458 and 463 µg µL-1, respectively) and presence of NaCl (459 and 456 µg µL-1, respectively). Except for isolate 4, fructose concentration increased under salt conditions (Table 5).
In organic P mineralization, fructose concentration was 13 µg µL-1 without salt and 16 µg µL-1 with salt. Fructose produced in the solubilization of inorganic P in the absence of salt was seven times higher than in organic P and ten times higher than in the presence of salt. No differences in fructose concentration were observed due to the effect of salt. Isolate 6 (Oceanobacillus sp.) produced high concentrations with and without salt (93 and 95 µg µL-1, respectively).
Similar to the solubilization of inorganic P, isolates 4 and 7 (B. velezensis) produced the most fructose in the solubilization of K, Mn, and Zn. The concentration of fructose in K solubilization was similar in both bacteria with (267 and 264 µg µL-1) and without salt (157 to 153 µg µL-1). Zn solubilization without salt (417 and 414 µg µL-1) was higher than when salt was added (349 and 347 µg µL-1) in the same two bacteria, respectively. Fructose was similarly produced in the absence or presence of NaCl (16.0 and 16.5 µg µL-1, respectively) during Mn solubilization. Moreover, isolates 4 and 7 secreted the highest concentration of fructose, both in the presence (33.4 and 32.0 µg µL-1, respectively) and absence of salt (32.0 and 35.0 µg µL-1, respectively). The concentration of fructose in the solubilization of inorganic P increased in the presence of salt in all bacteria. In contrast, the presence or absence of salt did not influence the concentration of fructuose coming from the extract of the mineralization of organic P.
Regarding K solubilization, all bacteria increased fructose secretion in the presence of salt, except isolates 3 (Nesterenkonia sp.) and 11 (Pseudomonas sp.). In Mn solubilization, only isolate 9 (B. pumilus) and 10 (Pseudomonas sp.) increased fructose concentration in the presence of salt, while the other bacteria had similar fructose concentrations. NaCl increased fructose concentration in the Zn solubilization extracts only in isolates 5 (Halobacillus sp.), 6 (Oceanobacillus sp.), 7 (B. velezensis), and 11 (Oceanobacillus sp.) (Table 5).
In inorganic P solubilization, xylose was only produced by isolates 10 (Pseudomonas sp., with 229.5 µg µL-1) and 5 (Halobacillus sp. with 29.0 µg µL-1 ). During P mineralization with and without salt, isolates 3 (Nesterenkonia sp.), 4 (B. velezensis), 5 (Halobacillus sp.), and 7 (B. velezensis) produced xylose. Nesterenkonia sp. produced the highest concentration of xylose with 2.5 (8.0 µg µL-1) and 0 M NaCl (8.5 µg µL-1). In K solubilization, the highest xylose concentration was observed in isolates 3 (Nesterenkonia sp.), 4 (B. velezensis), 7 (B. velezensis), and 9 (B. pumilus); in Mn solubilization, isolates 4 and 7 had the highest; and in Zn solubilization, isolates 3 and 9 had more xylose (Table 5).
In K solubilization, NaCl did not influence xylose concentration in isolates 3 (Nesterenkonia sp.), 4 (B. velezensis), 7 (B. velezensis), and 9 (B. pumilus); the same result was observed for Mn in isolates 4 and 7 (B. velezensis), and for Zn in isolates 3 (Nesterenkonia sp.) and 9 (B. pumilus). Contrary to what was observed for fructose concentration, adding NaCl only influenced the xylose concentration in the broth during the solubilization of inorganic P in isolates 5 (Halobacillus sp.) and 10 (Pseudomonas sp.). Sharma et al. [74] reported the bacterial production of xylose and its subsequent conversion to aldonic acid, following the transformation of glucose by the enzyme glucose dehydrogenase. This acid can participate in the efficient solubilization of inorganic P. Thus, the present investigation further supports the participation of xylose as an intermediate to aldonic acid in the solubilization of other nutrient sources such as Zn and Mn, but especially K (Table 5). Future studies should further analyze the participation of xylose in the solubilization of other nutrients.
2.6. Enzyme Activity in Different Salinity Conditions
The extracellular activity of alkaline and acid phosphatase enzymes in halotolerant rhizobacteria has been reported [75]. Shabaan et al. [76] qualitatively informed the extracellular enzyme activity of alkaline and acid phosphatase in 25 isolates of halotolerant rhizobacteria. The present study also provides information on the activity of these enzymes in halophilic endophytic bacteria and their response to different NaCl concentrations. All 11 bacteria showed alkaline and acid phosphatase enzyme activity (Table 6), but NaCl decreased alkaline phosphatase enzyme activity, whereas acid phosphatase activity increased. The maximum alkaline phosphatase activity was observed without NaCl in all bacteria except isolate 8 (Halomonas sp.), which only showed activity in the presence of salt (Table 6). Cummings and Gilmour [77] also observed that bacteria of the genus Halomonas stimulated their metabolism only in the presence of NaCl. The highest alkaline phosphatase activity (34 of µg mL-1 h-1) in 0 M NaCl was observed in isolate 2 (Bacillus sp.). This activity is lower than that reported by Shabaan et al. [76] in the halotolerant rhizobacterium SMH-7 in 0 M NaCl (164 of µg mL-1 h-1). However, the activity was higher than that observed by Barrera et al. [78] in the non-halotolerant rhizobacterium Kosakonia radicincitans in 0.1 and 2.2 M NaCl (1 µg mL-1 h-1 at both salt concentrations). Contrary to what was observed for alkaline phosphatase activity, the maximum acid phosphatase activity in all bacteria was observed in 1.5 M NaCl; this was ten times higher than in 0 M NaCl. The acid phosphatase activity in 0 and 1.5 M NaCl was, on average, 15 and 20 times higher than alkaline phosphatase at the same NaCl concentrations, respectively. In 0 M NaCl, the highest enzymatic activity for acid phosphatase (51 µg mL-1 h-1) was found in isolate 10 (Pseudomonas sp.), while isolates 1 (Oceanobacillus sp.), 2 (Bacillus sp.), 3 (Nesterenkonia sp.), and 8 (Halomonas sp.) showed no activity.
Interestingly, isolate 8 had no acid or alkaline phosphatase activity in 0 M NaCl but did so in the presence of salt. At 1.5 M NaCl, the highest acid phosphatase activity was found in isolates 4 (B. velezensis), 5 (Halobacillus sp.), 6 (Oceanobacillus sp.), 7 (B. velezensis), and 10 (Pseudomonas sp.). In contrast, in 2.5 M NaCl, the highest activity was in isolate 5 (Halobacillus sp.). These concentrations were lower than those reported by Shabaan et al. [76] in a halotolerant rhizobacteria (356.93 µg mL-1 h-1) but higher (2 µg mL-1 h-1) in two NaCl concentrations (0.1 y 2.2 M) than those produced by the halotolerant bacteria K. radicincitans [78].
In this study, alkaline phosphatase activity had a low correlation (r=0.32) with inorganic P solubilization, while acid phosphatase had a medium correlation (r=0.62). These results suggest that for P solubilization, halophilic endophytic bacteria release acid phosphatase in addition to producing organic acids. At pH < 7, the acid phosphatase enzyme shows higher activity [79].
Phytase transforms P from phytic acid to soluble P [53]. In the present work, all 11 halophilic endophytic bacteria produced phytase (Table 6) and released soluble P to use phytate as the sole source of organic P. Phytase activity correlated positively with the soluble concentration of P (r=0.72). Phytase activity increased in the presence of NaCl; it was two and five times higher at 1.5 and 2.5 M NaCl, respectively, compared to 0 M. Isolates 3 (Nesterenkonia sp.), 5 (Halobacillus sp.), and 9 (B. pumilus) had the highest phytase activity at 0 M of P (90.12, 90.38, and 90.01 µg mL-1 h-1, respectively). In comparison, isolate 9 (B. pumilus) had 120.85 at 1.5 M, and isolate 7 (B. velezensis) had 164.72 µg mL-1 h-1 at 2.5 M NaCl. As observed in alkaline and acid phosphatase activity, isolate 8 (Halomonas sp.) had no phytase activity in 0 M NaCl. These results are important because they show that enzymes related to P solubilization and mineralization in isolate 8 are only activated in the presence of NaCl. Importantly, phytase activity in these bacteria is higher than that observed in the halotolerant rhizobacterium P. azotoformas (10.08 µg P mL-1 h-1) in 0 M NaCl [80].
Halophilic endophytic microorganisms associated with halophytes are a potential source of several hydrolytic enzymes that are functional in saline environments, such as xylanases and chitinases. Wejse et al. [81] referred to these enzymes as multi-extremophiles because they are functional in high salt concentrations (>1 M NaCl), which is not the case for most other proteins. In the present research, enzyme profile analysis revealed the potential for xylanase production, both in the presence and absence of NaCl, in all bacterial isolates except for Oceanobacillus sp. isolates 1 and 2 and isolate 10 (Pseudomonas sp.) (Table 7). Isolate 8 (Halomonas sp.) showed no xylanase activity in 0 M NaCl. The presence of NaCl increased xylanase activity in all the producing bacteria, especially at 2.5 M NaCl compared to 1.5 and 0 M. Xylanase activity in 0 M NaCl was not different among the bacteria; however, isolate 11 (Oceanobacillus sp.) presented the highest xylanase activity in 1.5 and 2.5 M (1.89 and 5.32 µM mL-1 h-1, respectively), different from the rest of the isolates. Xylanase activity in the genus Oceanobacillus was qualitatively observed by Rohban et al., [82].
Khan et al. [83] showed that the halotolerant endophyte bacterium Bacillus sp. TKE4 showed xylanase activity in the absence of NaCl (9.4 µM mL-1 h-1), which is higher than the average activity found in the present research in isolate 11 (1.89 mL-1 h-1). Giridhar et al. [84] reported that the halotolerant endophyte Gracilibacillus sp. displayed xylanase activity in 0 and 2 M NaCl (0.9 and 2.8 µM mL-1 h-1, respectively). The halotolerant bacterium Marinimicrobium sp. maintained the same xylanase activity (60 µM mL-1 h-1) at two concentrations of NaCl (1.7 and 3.5 M). Xylanase production by endophytic bacteria allows them to degrade plant cell walls and leads to colonization. Xylanases are also related to the biological control of phytopathogenic fungi [85]. Therefore, bacteria with xylanase activity could be used as microbial inoculants for biological control. Future research should test this potential bacterial trait under in vitro and field conditions.
The present study also found that all halophilic endophytic bacteria, with exception of isolates 10 (Pseudomonas sp.) and 11 (Oceanobacillus sp.), exhibites chitinace activity. In particular, isolate 10 showed no xylanase or chitinase activity. Similarly, isolate 8 (Halomonas sp.) in 0 M NaCl showed no activity of both enzymes; however, these two enzymes were active in the presence of NaCl. As occurred with xylanases, the presence of NaCl increased chitinase activity. Chitinase activity in 1.5 and 2.5 M NaCl was similar among bacteria but differed in the absence of NaCl. These results evidence the highly tolerant nature of chitinase to salt. Finding enzymes that show optimal activity at various salt concentrations is relevant because salt does not limit their stability [86]. The rhizobacterium B. subtilis had chitinase activity of 120 µM mL-1 h-1 at 120 h in 0 M NaCl [87]. This activity was much higher than that exhibited by the bacteria in the present research analyzed in an incubation time of 48 h (between 1.24 and 1.90 µM mL-1 h-1). The halotolerant rhizobacterium Planococcus rifitoensis had chitinase activity of 10 µM mL-1 h-1 in 0 M NaCl, while in 1.7 M NaCl, it decreased to 6 µM mL-1 h-1. These findings indicate that similar to bacteria with xylanase activity, bacteria with chitinase activity could be used as a biological control of phytopathogenic fungi [87].
2.7. In vitro Compatibility between Bacterial Isolates
Bacterial compatibility is critical for formulating inoculants that promote plant growth [88]. Table S2 shows that there was compatibility between most of the bacteria. However, isolate 6 (Oceanobacillus sp.) was incompatible with isolates 5 (Halobacillus sp.) and 10 (Pseudomonas sp.). Isolate 3 (Nesterenkonia sp.) was incompatible with 4 and 7 (B. velezensis) and 10 (Pseudomonas sp.). Based on the multivariate analyses (reported in the next section), five bacterial isolates (2, Bacillus sp., 4, B. velezensis, 5, Halobacillus sp., 8, Halomonas sp. and, 9, B. pumilus) had a remarkably better biochemical performance than the rest of the halophilic endophytic bacteria and were compatible with each other (Figure 6). These promising results support the elaboration of a bacterial consortium that could potentiate these bacteria´s beneficial functions. Designing bacterial consortia has gained interest as a suitable strategy for sustainable agricultural production. A bacterial consortium generally consists of two or more compatible bacteria of different species in a synergistic interaction. In this context, the endophytic bacteria isolated from halophyte roots showed functional complementarity in plant-growth-promoting properties that promote phytostimulation, which may contribute to host plant productivity under salt stress, as suggested by Gaiero et al., [89]. Therefore, crop inoculation with this consortium may be a viable strategy for sustainable crop production in salinity-based agriculture, which includes crop production in arid and semi-arid environments.
2.8. Principal Component Analysis
Based on the PCA eigenvalues, only 21 variables (of the 42 analyzed) are relevant for selecting halophilic endophytic bacteria in this study (Table S3). The first three components of the PCA explained 67.4% of the total accumulated variance of the 21 analyzed variables (Figure 7). PC1 accumulated 31.3% of the variance and included the following bacterial traits as the ones with the greatest influence: nitrogenase and phytase enzyme activity, AIA production, P mineralization, Mn and K solubilization, siderophores production, and fructose production (especially in 2.5 M NaCl). PC2 explained 19.6% of the variance and was influenced by AIA production in the absence of NaCl and inorganic P solubilization, acid phosphatase, and citric acid production (both in the absence and presence of salt). PC3 accumulated 16.4% of the variance, and the related variables were nitrogenase enzyme activity in the absence of salt and EPS in both salt conditions. The comparative analysis showed that plant-growth-promoting properties such as P, K solubilization, phytic acid mineralization, and the production of siderophores, citric acid, and EPS were positively associated with both NaCl conditions; i.e., the presence of NaCl did not negatively affect these biochemical properties. Endophytic isolates 2 (Bacillus sp.), 4 (B. velezensis), 5 (Halobacillus sp.), 8 (Halomonas sp.), and 9 (B. pumilus) were of most significant interest due to their favorable response to 2.5 M NaCl. These strains; which are compatible each other, could be inoculated in glycophyte cultures to mitigate salt stress.
3. Materials and Methods
3.1. Selection of Halophytes, Sampling, and Root Disinfection
Fine roots of six dominant halophytes (Distichlis spicata, Cynodon dactylon, Eragrostis obtusiflora, Suaeda torreyana, Kochia scoparia, and Baccharis salicifolia) were sampled in an area in the east of the State of Mexico in the former Lake Texcoco (19.27°N, 98.54°O and 2,236 masl; Figure S1). The site is characterized by its salt content [90]. Samples were placed in separate plastic bags inside a cooler (4 °C) and immediately transported to the laboratory for analysis.
Roots were disinfected according to the methodology of Albdaiwi et al. [26]. Briefly, the roots were washed under running water for 10 min to remove adhering soil particles, placed in 70% ethanol for 1 min, and rinsed three times with sterile distilled water. A 3% sodium hypochlorite solution was added and left for 5 min, followed by six rinses with sterile distilled water. After the sixth rinse, an aliquot (100 μL) of wash was seeded onto Luria Bertani (LB) culture medium and incubated at 28 °C for 15 days. Surface disinfection was considered successful when no growth was observed on the medium.
3.2. Isolation of Endophytic Bacteria by Halotolerance
The isolation of bacteria from the roots of each halophyte was performed by halotolerance in LB media with increasing concentrations of NaCl (0.1, 0.5, 1.5, and 2.5 M). Initially, 0.5 g of the root tissue was macerated in a mortar with 10 mL of sterile distilled water. Subsequently, serial decimal dilutions were prepared, and 100 µL of the last dilution (10-3) was placed in Petri dishes prepared with LB medium in the different NaCl concentrations. The Petri dishes were incubated for 15 days at 28 °C [2]. Bacteria isolated at the 2.5 M NaCl concentration were purified, molecularly identified, and biochemically characterized. Bacteria grown at the other NaCl concentrations were cryopreserved in 20% glycerol.
3.3. Molecular Identification of Halophilic Endophytic Bacteria
DNA isolation was performed according to Töpper et al. [91]. Bacteria were placed in LB medium and incubated at 28 °C for 48 h at 160 rpm. Then, 1 mL of each bacterial suspension was placed in an Eppendorf tube and centrifuged at 14,000 rpm for 3 min. Subsequently, DNA extraction was performed with the commercial DNeasy Blood & Tissue Kit from Qiagen according to the manufacturer’s instructions. The 16S rDNA gene fragment was amplified by polymerase chain reaction (PCR) with the conditions shown in Table S4. The universal primers were 16SF direct: 5′-GCCTAACACATGCAAGTC-3′ and 16SR reverse: 5′-AAGGAGGTGATCCAGCCGCA-3′. A reaction mixture was prepared for each oligonucleotide pair, as described in Table S4. The PCR product was approximately 1500 bp, which was verified on 1% agarose gel. Subsequently, the 16S rRNA sequences were compared with the GenBank database using the NCBI BLAST nucleotide search. A multiple sequence alignment was constructed with the ClustalX 1.8 software package (http://www.clustal.org/clustal2), and a phylogenetic tree was prepared using the neighbor-joining method in MEGA v6.1 software (www.megasoftware.net), with confidence tested by bootstrap analysis (1000 replicates).
3.4. Biochemical Characterization of Halophilic Endophytic Bacteria
Halophilic bacteria were characterized according to the production of indole acetic acid (IAA), siderophores, and exopolysaccharides (EPS); nutrient solubilization (P, K, Zn, and Mn); phytic acid mineralization; nitrogenase enzyme activity; production of organic acids and sugars in the solubilization broth of all nutrient solubilization tests; and enzyme activity (acid and alkaline phosphatase, phytases, xylanases, and chitinases).
For all evaluations, bacteria were propagated in LB medium supplemented with 2.5 M NaCl and incubated for 48 h at 28 °C. Bacterial cultures were adjusted to 0.5 absorbance at 600 nm (equivalent to 1x106 bacteria mL-1). From each bacterial suspension, 1 mL was taken and placed in 10 mL of prepared LB broth to measure the solubilization of each nutrient, siderophore production, and IAA separately. These tests were performed in the presence (2.5 M) and absence of NaCl.
3.4.1. Indole Acetic Acid and Siderophore Production
IAA production was measured using the procedure described by Bano and Musarrat [92] in LB broth supplemented with 1 mg L-1 of L-tryptophan. Samples were incubated at 35 °C for 48 h and then centrifuged at 9,000 rpm for 10 min. One mL of the supernatant was placed in a test tube, and two mL of Salkowski reagent was added. The samples were incubated for 30 min until a pink color was obtained, which is characteristic of IAA production. The absorbance of the sample was measured in a spectrophotometer at a wavelength of 530 nm [93], while the concentration was determined based on a standard curve with concentrations between 12-60 mg L-1 of IAA.
The quantification of siderophores was performed using the methodology of Arora and Verma [94]. Bacterial cultures were grown in LB broth and centrifuged at 10,000 rpm for 10 min. The supernatant (0.5 mL) bacterial was mixed with 0.5 mL CAS reagent, and after 20 min, the absorbance was measured at 630 nm. Siderophore production was calculated using the following formula and reported as a percentage [95].
Where:
Ar= reference absorbance (CAS solution and uninoculated culture medium), and As= absorbance of the sample (CAS solution and uninoculated supernatant).
3.4.2. Exopolysaccharide and Nitrogenase Enzyme Production
EPS production was evaluated using the method modified by Chaudhari et al. [96]. The LB medium was prepared with 0.8 g L-1 of Congo red dye supplemented with 3% sucrose. Bacterial isolates were seeded in Petri dishes with the prepared medium and incubated at 37 °C for 48 h. According to Arciola et al. [97], bacterial colonies that were very dark to almost black were designated as positive for EPS production.
The ability to fix atmospheric nitrogen was evaluated according to the methodology of Xie et al. [98] in a semisolid sucrose malate medium with bromothymol blue. After one week, the change in color from green to blue was considered positive for nitrogenase activity.
3.4.3. Quantitative Determination of Nutrients Solubilization and Mechanisms Involved
The methodology described by Battini et al. [99] was used to determine P solubilization and P mineralization. Solubilization was determined from an inorganic source (Ca3(PO4)2), and mineralization was determined from an organic one (phytic acid: C6H18O24P6). In addition, NBRIP and PSM (Phytate Screening Medium) culture media were prepared for inorganic P solubilization and organic P mineralization, respectively. K solubilization was measured with the methodology of Parmar and Sindhu [100] using a mineral medium with 3% potassium feldspar. Feldspars are among the most important minerals in the soil, and their chemical formula is KAlSi3O8. Mn and Zn solubilization was evaluated using 50 mM MnO2 and 0.1% Zn, respectively, according to the methodology described by Sanket et al. [67].
After inoculation with the bacteria, all the solubilization tests were incubated at 28 °C for 48 h at 120 rpm. Absorbance was determined at 600 nm in a UV-visible spectrometer as an indirect measure of bacterial growth (Cary 50, Varian). Then, the samples were filtered with Whatman No. 40 filter paper and centrifuged at 9,000 rpm for 10 min. The cell-free supernatants were measured for pH (potentiometrically) and soluble nutrient concentration (P, K, Mn, and Zn). All tests had three replicates for evaluating each solubilization ion, and a treatment without inoculation (control) was included.
A 0.5 mL aliquot of the bacterial cell-free supernatant was placed in a test tube, and 1 mL of vanadate-molybdate solution and 3.5 mL of deionized water were added to quantify soluble P. The samples were allowed to stand for 10 min until they developed a yellow color. Then, absorbance was measured with a spectrophotometer (Perkin Elmer 3110) at a wavelength of 400 nm [101]. The concentration of soluble P was determined by comparing the absorbance of the sample with a standard curve with concentrations ranging from 1-6 mg L-1. Soluble K was measured by flame emission photometry (Jenway PFP7). Soluble Mn and Zn concentrations were quantified with an atomic absorption spectrometer (Perkin Elmer 3110). Certified standards were used to prepare the respective calibration curve standards with deionized water (0.2 Mohms).
3.4.3.1. Production of Organic Acids and Sugars
Bacterial cell-free supernatants from inorganic P, K, Mn, and Zn solubilization were frozen until organic acid analysis [47]. Samples were thawed, sonicated for 20 min (at 40kHz; Branson 1510), and filtered with a 0.22 µm nylon mesh. The samples were then analyzed by HPLC-IR (LDC Analytical IR Detector Varian ProStar) with a Phenomenex Rezex column (ROA, Organic acid H+ 300X 7.8 mm). The following 12 acids (Sigma) were used as standards: fumaric, citric, oxalic, lactic, acetic, acetic, malic, tartaric, salicylic, maleic, vanillic, and succinic.
Sugars were determined according to the methodology of Murkovic and Derler [102] in bacteria-free solubilization extracts. The standards used were D-glucose, D-fructose, D-sucrucose, D-lactose, D-maltose, D-xylose, D-arabinose, and trehalose. Calibration curves were prepared with a concentration of 1 mg mL-1 of these standards. The HPLC system configuration was 25 water: 75 acetonitrile. In addition, separation of the compounds was performed with a CarboPac PA1 anion exchange column (250 x 4 mm; Dionex Corp.) and a CarboPac PA1 guard column (50 x 4 mm; Dionex) with a volume of 1 µL and a constant flow rate of 1 mL min-1.
3.4.4. Extracellular Enzyme Activity at Different Salinity Conditions
To measure the extracellular enzyme activity of acid and alkaline phosphatase, phytase, xylanase, and chitinase, each bacterium separately was cultured in LB broth without NaCl and adjusted bacterial density to 0.5 absorbance at 600 nm. Enzyme activity was evaluated at three NaCl concentrations (0, 1.5, and 2.5 M). All enzyme activity assays were performed in triplicate, and a control was included.
Phosphatase activity was determined using disodium p-nitrophenylphosphate (PNPP 0.025 M) as a colorimetric substrate [99]. Each bacterial isolate was cultured at 28 °C for 48 h in NBRIP medium at pH = 7 supplemented with Ca3(PO4)2 (5 g L-1), and 1 mL of adjusted culture medium was added. Cell-free supernatant samples were obtained by centrifuging 2 mL of culture of each bacterial isolate at 10,000 rpm for 10 min. In a test tube, 0.5 mL of NBRIP supernatant, 0.5 mL of substrate (NBRIP medium), and 2 mL of 0.5 modified universal buffer (MUB) were added. The MUB was adjusted to pH = 6.5 for acid phosphatase and pH = 11 for alkaline phosphatase. The reactions were carried out at 37 ± 1 °C for 60 min and stopped by adding 0.5 mL of 0.5 M CaCl2 and 2 mL of 0.5 M NaOH. The samples were filtered using Whatman No. 42 filter paper. The p-nitrophenol (PNP) formed was measured spectrophotometrically at 400 nm. Three independent replicates per treatment were performed. A unit of enzyme activity (U) was expressed as micrograms of PNP released per milliliter per hour.
An NBRIP medium supplemented with phytic acid (2 g L-1) was prepared for quantitative analysis of extracellular phytase production, and 1 mL of bacterial suspension adjusted to 0.5 absorbance was added. After incubation at 28 °C for 48 h, the cultures were centrifuged at 10,000 rpm for 10 min, and the supernatant was used for extracellular phytase estimation. Phytase activity determines the amount of inorganic P released into the medium [103]; therefore, two mixtures were prepared to assess the rate of released P. In Mixture 1, the supernatant (0.2 mL) was mixed with 0.5 g of phytic acid dissolved in 100 mL of sodium acetate buffer (0.2 M, pH = 5.5). After 30 min of incubation at 28 °C, the reaction was stopped by adding 10 mL of 15% trichloroacetic acid. In Mixture 2, 100 mL sulfuric acid (1 M) was mixed with 100 mL ascorbic acid (10%) and 4 mL ammonium molybdate (3:1:0.1) 2.5%. For phytase estimation, 100 µL of Mixture 1 was mixed with 900 µL of Mixture 2 and incubated at 28 °C for 1 h. The absorbance of the final mixtures was measured at 400 nm [99]. A unit of enzyme activity (U) was expressed as micrograms of P released per milliliter per hour.
For quantitative analysis of extracellular xylanase production, LB broth was prepared with 1% xylan and 1 mL of bacterial culture medium adjusted to 0.5 absorbance. After incubation at 28 °C for 48 h, the cultures were centrifuged at 10,000 rpm for 10 min to obtain the supernatant, which was used for extracellular xylanase estimation. Xylanase activity was estimated by measuring the yield of reducing sugars with modification of the 3,5-dinitrosalicylic acid (DNS) method [104]. The supernatant (50 µL) was mixed with 50 µL of 0.1 M citrate buffer (pH = 4.8) and 1% xylan and incubated at 55 °C for 60 min. Then, 200 µL of DNS solution was added to the reaction, which was maintained in a boiling water bath for 5 min. The released reducing sugars were estimated using a standard curve of 1 mg mL-1 glucose.
Chitinase activity was determined by the method of Ramírez et al. [105]. The LB broth medium was supplemented with colloidal chitin (10%), and the bacterial suspension (1 mL), adjusted to 0.5 absorbance, was incubated at 28 °C for 48 h. The bacterial cultures were centrifuged at 10,000 rpm for 10 min to obtain the supernatant. Then, 10% colloidal chitin and 1 mL of supernatant were mixed and incubated at 30 °C for 60 min. The reaction was stopped by adding 1 mL of 1% NaOH. The product was determined by 3,5-dinitrosalicylic acid (DNS) assay, and the absorbance was measured at 535 nm. Chitinase activity was defined as the enzyme required to produce 1 μM of N- acetylglucosamine per hour per mL of supernatant [106].
3.5. Compatibility Between Bacterial Isolates
For the possible association of bacteria (bacterial consortia), in vitro compatibility between bacterial isolates was tested in triplicate, as described by Sundaramoorthy et al. [107]. Bacterial isolates were inoculated in Petri dishes with LB medium, two by two, in an extended manner. The isolates were distributed in perpendicular lines. The Petri dishes were incubated at 28 °C for 72 h. Compatible bacterial isolates grew on top of each other, while non-compatible isolates formed inhibition areas between combinations.
3.6. Statistical Analysis
The Shapiro-Wilk normality test was performed for all quantitative variables, while homogeneity of variances was corroborated with Bartlett’s test (α=0.05). All variables presented normal distribution, so data transformation was not necessary. ANOVA (p ≤ 0.05) and Tukey’s mean comparison (p ≤ 0.05) tests were performed with R software version 4.0.5. Principal component analysis (PCA) was performed with the R package Factoextra version 4.0.5 [108].
4. Conclusions
Of the 120 root endophytic bacteria isolated from halophytes, 11 were halophilic and exhibited promising characteristics to improve plant growth, both in the presence and absence of NaCl. Therefore, these bacteria can be used as microbial inoculants to promote plant growth, biostimulate nutrient solubilization, and tested for their contribution to sustainable agriculture under salinity conditions due to their adaptation to these conditions. The isolated bacteria had a higher affinity for organic P mineralization compared to inorganic P. Salt concentration increased nutrient solubilization depending on bacterial strain and solubilization ion. The nitrogenase activity found in some bacteria in the presence of salt is relevant since, under saline conditions, nitrogen limits plant growth and food production. Results showed a relationship between the production of organic acids and the solubilization of nutrients that have low availability in saline soils, such as K, Zn, and Mn. An exclusive production of maleic acid in Zn solubilization and fructose and xylose in nutrient solubilization was observed. The presence of salt influenced fructose but not xylose secretion in some bacteria. The activity of acid phosphatase, phytase, xylanase, and chitinase enzymes increased in the presence of salt, showing their stability at the salt concentrations tested. Halophilic endophytic bacteria displayed xylanase and chitinase activity, and these enzymes should be tested in biological control agents to protect crops from phytopathogenic fungi. Finally, the fact that these bacteria are mostly compatible provides a solid basis for the future formulation of consortia with desirable biochemical characteristics to help mitigate salt stress.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. This research includes four supplementary tables and one supplementary figure. Table S1: 16S gen amplification process by PCR to molecular identification of halophilic endophytic bacteria, Table S2: Growth of 11 bacteria in the nutrient solubilization tests, Table S3: Compatibility among halophilic endophytic bacteria isolated from halophytes, Table S4: Eigenvalues, variance, and stronger variables from principal component analysis, Figure S1: Location of sampling site to the isolation of endophytic bacteria from roots of six halophytes.
Author Contributions
JABG contributed to the research conceptualization, methodology, validation, formal analysis, investigation, data curation, writing of original draft preparation, writing review & editing. YEKR participated in molecular analysis, writing of original draft preparation, writing review & editing, and visualization. RCG was involved in the soil and plant chemical analysis, methodology, resources, writing of original draft preparation, and writing review & editing. MCGC contributed to research conceptualization, methodology, resources, data curation, writing original draft preparation, writing review and editing, supervision, project administration, and funding acquisition. MEHL participated in the enzyme and molecular analyses and resources. RMSH contributed to the organic acids and sugar analyses and resources. BEHC was involved in the statistical analysis and interpretation of principal component analysis. All authors read and accept the final document version.
Funding
This study was supported by Colegio de Postgraduados.
Data Availability Statement
All information generated in this research is presented.
Acknowledgments
The authors thank Dr. Heike Vibrans Lindemann from the Posgrado de Botánica, Colegio de Postgraduados, Campus Montecillo, for the taxonomical identification of halophytes. We also appreciate the technical support from MC. Jesús Eulises Corona Sánchez for elemental analysis. JABG thanks the Mexican Council of Humanities, Science, and Technology (CONAHCYT) for his PhD scholarship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siddikee, A.; Chauhan, P.S.; Anandham, R.; Han, G.-H.; Sa, T. Isolation, Characterization, and Use for Plant Growth Promotion Under Salt Stress, of ACC Deaminase-Producing Halotolerant Bacteria Derived from Coastal Soil. J. Microbiol. Biotechnol. 2010, 20, 1577–1584. [Google Scholar] [CrossRef]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. SpringerPlus 2013, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zandi, E. , Jafari, A. A., & Mirakhorli, R. Studying the effects of growth stages on forage quality of two halophytes in Garmsar. Iranian Journal of Range and Desert Research, 2017, 24, 467-473. [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Naz, I.; Bano, A. Biochemical, molecular characterization and growth promoting effects of phosphate solubilizing Pseudomonas sp. isolated from weeds grown in salt range of Pakistan. Plant Soil 2010, 334, 199–207. [Google Scholar] [CrossRef]
- Shin, W.; Siddikee, A.; Joe, M.M.; Benson, A.; Kim, K.; Selvakumar, G.; Kang, Y.; Jeon, S.; Samaddar, S.; Chatterjee, P.; et al. Halotolerant Plant Growth Promoting Bacteria Mediated Salinity Stress Amelioration in Plants. Korean J. Soil Sci. Fertil. 2016, 49, 355–367. [Google Scholar] [CrossRef]
- Coombs, J.T.; Franco, C.M.M. Isolation and Identification of Actinobacteria from Surface-Sterilized Wheat Roots. Appl. Environ. Microbiol. 2003, 69, 5603–5608. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, S.; Malik, K.; Mehnaz, S. Microbiome of Halophytes: Diversity and Importance for Plant Health and Productivity. Microbiol. Biotechnol. Lett. 2019, 47, 1–10. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Bacterial endophyte mediated plant tolerance to salinity: growth responses and mechanisms of action. World J. Microbiol. Biotechnol. 2020, 36, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial mediation of plant hormone status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Abbas, G.; Saqib, M.; Akhtar, J.; Haq, M.A.U. Interactive effects of salinity and iron deficiency on different rice genotypes. J. Plant Nutr. Soil Sci. 2015, 178, 306–311. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A. Bacillus species as the most promising bacterial biocontrol agents in rhizosphere and endorhiza of plants grown in rotation with each other. Eur. J. Plant Pathol. 2017, 150, 497–506. [Google Scholar] [CrossRef]
- Puente, M.E.; Bashan, Y.; Li, C.Y.; Lebsky, V.K. Microbial Populations and Activities in the Rhizoplane of Rock-Weathering Desert Plants. I. Root Colonization and Weathering of Igneous Rocks. Plant Biol. 2004, 6, 629–642. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresour. Technol. 2017, 247, 190–199. [Google Scholar] [CrossRef]
- Seshachala, U.; Tallapragada, P. Phosphate Solubilizers from the Rhizospher of Piper nigrum L. in Karnataka, India. Chil. J. Agric. Res. 2012, 72, 397–403. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Kachare, S.; Tiwari, S. Molecular diversity of 1-aminocyclopropane-1-carboxylate (ACC) deaminase producing PGPR from wheat (Triticum aestivum L.) rhizosphere. Plant Soil 2016, 414, 213–227. [Google Scholar] [CrossRef]
- Alloway, B.J. Soil factors associated with zinc deficiency in crops and humans. Environ. Geochem. Health 2009, 31, 537–548. [Google Scholar] [CrossRef]
- Bradáčová, K.; Florea, A.S.; Bar-Tal, A.; Minz, D.; Yermiyahu, U.; Shawahna, R.; Kraut-Cohen, J.; Zolti, A.; Erel, R.; Dietel, K.; et al. Microbial Consortia versus Single-Strain Inoculants: An Advantage in PGPM-Assisted Tomato Production? Agronomy 2019, 9, 105. [Google Scholar] [CrossRef]
- Liu, X.; Mei, S.; Salles, J.F. Inoculated microbial consortia perform better than single strains in living soil: A meta-analysis. Appl. Soil Ecol. 2023, 190. [Google Scholar] [CrossRef]
- Shenoy, V. V. , & Kalagudi, G. M. Meta-bug and near-isogenic strain consortia concepts for plant growth promoting rhizobacteria. In 6th International PGPR workshop, India, section VII—mechanism of biocontrol. 2003, p (Vol. 108).
- Vaishnav, A. , Varma, A., Tuteja, N., Choudhary, D.K. PGPR-mediated amelioration of crops under salt stress. In: Choudhary, D., Varma, A., Tuteja, N. (eds) Plant-Microbe Interaction: An Approach to Sustainable Agriculture. Springer, Singapore, 2016. [CrossRef]
- Marghoob, M.U.; Nawaz, A.; Ahmad, M.; Waheed, M.Q.; Khan, M.H.; Imtiaz, M.; Islam, E.U.; Imran, A.; Mubeen, F. Assessment of halotolerant bacterial and fungal consortia for augmentation of wheat in saline soils. Front. Microbiol. 2023, 14, 1207784. [Google Scholar] [CrossRef]
- Rajput, L., Imran, A., Mubeen, F., & Hafeez, F. Y. Wheat (Triticum aestivum L.) growth promotion by halo-tolerant PGPR-consortium. Soil Environment 2018, 37, 178–189. [CrossRef]
- Damodaran, T.; Jha, S.K.; Kumari, S.; Gupta, G.; Mishra, V.K.; Sharma, P.C.; Gopal, R.; Singh, A.; Jat, H.S. Development of Halotolerant Microbial Consortia for Salt Stress Mitigation and Sustainable Tomato Production in Sodic Soils: An Enzyme Mechanism Approach. Sustainability 2023, 15, 5186. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y.; Alananbeh, K.M.; Al-Sayaydeh, R. Isolation and Characterization of Halotolerant Plant Growth Promoting Rhizobacteria From Durum Wheat (Triticum turgidum subsp. durum) Cultivated in Saline Areas of the Dead Sea Region. Front. Microbiol. 2019, 10, 1639. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, S.; Mehnaz, S.; Mirza, M.S.; Malik, K.A. Isolation and characterization of bacteria associated with the rhizosphere of halophytes (Salsola stocksii and Atriplex amnicola) for production of hydrolytic enzymes. Braz. J. Microbiol. 2019, 50, 85–97. [Google Scholar] [CrossRef]
- Mora-Ruiz, M.d.R.; Font-Verdera, F.; Orfila, A.; Rita, J.; Rosselló-Móra, R. Endophytic microbial diversity of the halophyteArthrocnemum macrostachyumacross plant compartments. FEMS Microbiol. Ecol. 2016, 92, fiw145. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. The Endophytic Bacteria Bacillus velezensis Lle-9, Isolated from Lilium leucanthum, Harbors Antifungal Activity and Plant Growth-Promoting Effects. J. Microbiol. Biotechnol. 2020, 30, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Brahim, A.H.; Ben Ali, M.; Daoud, L.; Jlidi, M.; Akremi, I.; Hmani, H.; Feto, N.A.; Ben Ali, M. Biopriming of Durum Wheat Seeds with Endophytic Diazotrophic Bacteria Enhances Tolerance to Fusarium Head Blight and Salinity. Microorganisms 2022, 10, 970. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, S.; Dąbrowska, G.B.; Tyburski, J.; Niedojadło, K.; Piernik, A.; Hrynkiewicz, K. Boosting the Brassica napus L. tolerance to salinity by the halotolerant strain Pseudomonas stutzeri ISE12. Environ. Exp. Bot. 2019, 163, 55–68. [Google Scholar] [CrossRef]
- Mahgoub, H.A.M.; Fouda, A.; Eid, A.M.; Ewais, E.E.-D.; Hassan, S.E.-D. Biotechnological application of plant growth-promoting endophytic bacteria isolated from halophytic plants to ameliorate salinity tolerance of Vicia faba L. Plant Biotechnol. Rep. 2021, 15, 819–843. [Google Scholar] [CrossRef]
- Shurigin, V.; Egamberdieva, D.; Li, L.; Davranov, K.; Panosyan, H.; Birkeland, N.-K.; Wirth, S.; Bellingrath-Kimura, S.D. Endophytic bacteria associated with halophyte Seidlitzia rosmarinus Ehrenb. ex Boiss. from saline soil of Uzbekistan and their plant beneficial traits. J. Arid. Land 2020, 12, 730–740. [Google Scholar] [CrossRef]
- Huang, Q.-S.; Yan, Z.-F.; Chen, X.-Q.; Du, Y.-Y.; Li, J.; Liu, Z.-Z.; Xia, W.; Chen, S.; Wu, J. Accelerated biodegradation of polyethylene terephthalate by Thermobifida fusca cutinase mediated by Stenotrophomonas pavanii. Sci. Total. Environ. 2022, 808, 152107. [Google Scholar] [CrossRef]
- Bhutani, N.; Maheshwari, R.; Sharma, N.; Kumar, P.; Dang, A.S.; Suneja, P. Characterization of halo-tolerant plant growth promoting endophytic Bacillus licheniformis MHN 12. J. Genet. Eng. Biotechnol. 2022, 20, 113–12. [Google Scholar] [CrossRef]
- Dhungana, S.A.; Itoh, K. Effects of Co-Inoculation of Indole-3-Acetic Acid-Producing and -Degrading Bacterial Endophytes on Plant Growth. Horticulturae 2019, 5, 17. [Google Scholar] [CrossRef]
- Mahdi, I.; Hafidi, M.; Allaoui, A.; Biskri, L. Halotolerant Endophytic Bacterium Serratia rubidaea ED1 Enhances Phosphate Solubilization and Promotes Seed Germination. Agriculture 2021, 11, 224. [Google Scholar] [CrossRef]
- Patel, R.; Parekh, V.B. Culturable endophytic bacteria from halotolerant Salicornia brachata L. : Isolation and plant growth promoting traits. Indian J. Appl. Microbiol. 2018, 21, 10–21. [Google Scholar] [CrossRef]
- Panwar, M.; Tewari, R.; Nayyar, H. Native halo-tolerant plant growth promoting rhizobacteria Enterococcus and Pantoea sp. improve seed yield of Mungbean (Vigna radiata L.) under soil salinity by reducing sodium uptake and stress injury. Physiol. Mol. Biol. Plants 2016, 22, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Kearl, J.; McNary, C.; Lowman, J.S.; Mei, C.; Aanderud, Z.T.; Smith, S.T.; West, J.; Colton, E.; Hamson, M.; Nielsen, B.L. Salt-Tolerant Halophyte Rhizosphere Bacteria Stimulate Growth of Alfalfa in Salty Soil. Front. Microbiol. 2019, 10, 1849. [Google Scholar] [CrossRef] [PubMed]
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A hope for cultivation of saline soils. J. King Saud Univ. - Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Vyacheslav, S.; Begali, A.; Kakhramon, D.; Zafar, I. Bacterial endophytes from halophyte black saxaul (Haloxylon aphyllum Minkw.) and their plant growth-promoting properties. J. Appl. Biol. Biotechnol. 2022, 10, 45–53. [Google Scholar] [CrossRef]
- Enquahone, S.; van Marle, G.; Simachew, A. Plant growth-promoting characteristics of halotolerant endophytic bacteria isolated from Sporobolus specatus (Vahr) Kunth and Cyperus laevigatus L. of Ethiopian rift valley lakes. Arch. Microbiol. 2022, 204, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ruppel, S.; Franken, P.; Witzel, K. Properties of the halophyte microbiome and their implications for plant salt tolerance. Funct. Plant Biol. 2013, 40, 940–951. [Google Scholar] [CrossRef]
- Shahid, M.; Zeyad, M.T.; Syed, A.; Singh, U.B.; Mohamed, A.; Bahkali, A.H.; Elgorban, A.M.; Pichtel, J. Stress-Tolerant Endophytic Isolate Priestia aryabhattai BPR-9 Modulates Physio-Biochemical Mechanisms in Wheat (Triticum aestivum L.) for Enhanced Salt Tolerance. Int. J. Environ. Res. Public Heal. 2022, 19, 10883. [Google Scholar] [CrossRef]
- Paredes-Mendoza, M. , & Espinosa-Victoria, D. Ácidos orgánicos producidos por rizobacterias que solubilizan fosfato: una revisión crítica. Terra Latinoamericana, 2010, 28, 61–70. [Google Scholar]
- Rodrı́guez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef] [PubMed]
- Dey, G.; Banerjee, P.; Sharma, R.K.; Maity, J.P.; Etesami, H.; Shaw, A.K.; Huang, Y.-H.; Huang, H.-B.; Chen, C.-Y. Management of Phosphorus in Salinity-Stressed Agriculture for Sustainable Crop Production by Salt-Tolerant Phosphate-Solubilizing Bacteria—A Review. Agronomy 2021, 11, 1552. [Google Scholar] [CrossRef]
- Li, W.C.; Ye, Z.H.; Wong, M.H. Metal mobilization and production of short-chain organic acids by rhizosphere bacteria associated with a Cd/Zn hyperaccumulating plant, Sedum alfredii. Plant Soil 2009, 326, 453–467. [Google Scholar] [CrossRef]
- Rasul, M.; Yasmin, S.; Yahya, M.; Breitkreuz, C.; Tarkka, M.; Reitz, T. The wheat growth-promoting traits of Ochrobactrum and Pantoea species, responsible for solubilization of different P sources, are ensured by genes encoding enzymes of multiple P-releasing pathways. Microbiol. Res. 2021, 246, 126703. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Rosales, E., Pasoz, M., Sanroman, M. A. Solid-state fermentation for food applications. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2018, 319–355. [CrossRef]
- Etesami, H.; Beattie, G.A. Plant-Microbe Interactions in Adaptation of Agricultural Crops to Abiotic Stress Conditions. In Probiotics and Plant Health; Springer Singapore: Singapore, Singapore, 2017; pp. 163–200. [Google Scholar] [CrossRef]
- Patel, P. K. , Jadav, N. J., & Gohil, V. K. N. Direct effects of potassium, sulphur and KSB on growth, yield and quality of direct seeded rice under direct seeded rice-chickpea cropping system. The Pharma Innovation Journal 2023, SP-12, 846-851.
- Bakelli, A., Amrani, S., Nacer, A., Bouri, M., & Sahin, F. Klebsiella sp. S6. A halotolerant rhizosphere bacterium of Phragmites communis L. with potential plant-growth promotion of pepper. Analele Universitatii din Oradea, Fascicula Biologie, 2022, 29, 92-100.
- Ranawat, B.; Bachani, P.; Singh, A.; Mishra, S. Enterobacter hormaechei as Plant Growth-Promoting Bacteria for Improvement in Lycopersicum esculentum. Curr. Microbiol. 2021, 78, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.; Hassan, H.M.; Ghazali, A.H.A.; Ahmad, M. Halotolerant potassium solubilizing plant growth promoting rhizobacteria may improve potassium availability under saline conditions. Environ. Monit. Assess. 2020, 192, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zarjani, J.K.; Aliasgharzad, N.; Oustan, S.; Emadi, M.; Ahmadi, A. Isolation and characterization of potassium solubilizing bacteria in some Iranian soils. Arch. Agron. Soil Sci. 2013, 59, 1713–1723. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, P.; Tiwari, R.; Meena, K.K.; Yandigeri, M.; Singh, D.P.; Arora, D.K. Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biol. Fertil. Soils 2011, 47, 907–916. [Google Scholar] [CrossRef]
- Yamaji, K.; Nagata, S.; Haruma, T.; Ohnuki, T.; Kozaki, T.; Watanabe, N.; Nanba, K. Root endophytic bacteria of a 137Cs and Mn accumulator plant, Eleutherococcus sciadophylloides, increase 137Cs and Mn desorption in the soil. J. Environ. Radioact. 2016, 153, 112–119. [Google Scholar] [CrossRef]
- Dixit, S. J. , Kuttan, A. K., & Shrivastava, R. M. Genetic characterization of sulphur and iron oxidizing bacteria in manganese mining area of Balaghat and Chhindwara, Madhya Pradesh, 2018, India.
- Uren, N. Chemical reduction of an insoluble higher oxide of manganese by plant roots. J. Plant Nutr. 1981, 4, 65–71. [Google Scholar] [CrossRef]
- Ijaz, A.; Mumtaz, M.Z.; Wang, X.; Ahmad, M.; Saqib, M.; Maqbool, H.; Zaheer, A.; Wang, W.; Mustafa, A. Insights Into Manganese Solubilizing Bacillus spp. for Improving Plant Growth and Manganese Uptake in Maize. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.B.; Jensen, P.E.; Husted, S. Manganese Deficiency in Plants: The Impact on Photosystem II. Trends Plant Sci. 2016, 21, 622–632. [Google Scholar] [CrossRef]
- Schmidt, S.B.; Husted, S. The Biochemical Properties of Manganese in Plants. Plants 2019, 8, 381. [Google Scholar] [CrossRef]
- Sanket, A.S.; Ghosh, S.; Sahoo, R.; Nayak, S.; Das, A.P. Molecular Identification of Acidophilic Manganese (Mn)-Solubilizing Bacteria from Mining Effluents and Their Application in Mineral Beneficiation. Geomicrobiol. J. 2016, 34, 71–80. [Google Scholar] [CrossRef]
- Fatima, T.; Mishra, I.; Verma, R.; Arora, N.K. Mechanisms of halotolerant plant growth promoting Alcaligenes sp. involved in salt tolerance and enhancement of the growth of rice under salinity stress. 3 Biotech 2020, 10, 361. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Fluorescent Pseudomonas sp. PF17 as an efficient plant growth regulator and biocontrol agent for sunflower crop under saline conditions. Symbiosis 2016, 68, 99–108. [Google Scholar] [CrossRef]
- Ramesh, A.; Sharma, S.K.; Sharma, M.P.; Yadav, N.; Joshi, O.P. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in Vertisols of central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Upadhyay, H.; Gangola, S.; Sharma, A.; Singh, A.; Maithani, D.; Joshi, S. Contribution of zinc solubilizing bacterial isolates on enhanced zinc uptake and growth promotion of maize (Zea mays L.). Folia Microbiol. 2021, 66, 1–11. [Google Scholar] [CrossRef]
- Li, W.C.; Ye, Z.H.; Wong, M.H. Metal mobilization and production of short-chain organic acids by rhizosphere bacteria associated with a Cd/Zn hyperaccumulating plant, Sedum alfredii. Plant Soil 2009, 326, 453–467. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, N.; Fan, J.; Wang, F.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi stimulate organic phosphate mobilization associated with changing bacterial community structure under field conditions. Environ. Microbiol. 2018, 20, 2639–2651. [Google Scholar] [CrossRef]
- Sharma, V.; Kumar, V.; Archana, G.; Kumar, G.N. Substrate specificity of glucose dehydrogenase (GDH) ofEnterobacter asburiaePSI3 and rock phosphate solubilization with GDH substrates as C sources. Can. J. Microbiol. 2005, 51, 477–482. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 178, 104124. [Google Scholar] [CrossRef]
- Shabaan, M. Preliminary evaluation of halotolerant PGPRs for promoting maize growth under salt stressed gnotobiotic conditions. Pak. J. Agric. Sci. 2021, 58, 1491–1500. [Google Scholar] [CrossRef]
- Cummings, S.P.; Gilmour, D.J. The effect of NaCl on the growth of a Halomonas species: accumulation and utilization of compatible solutes. Microbiology 1995, 141, 1413–1418. [Google Scholar] [CrossRef]
- Barrera, M.C.; Jakobs-Schoenwandt, D.; Gómez, M.I.; Becker, M.; Patel, A.V.; Ruppel, S. Salt stress and hydroxyectoine enhance phosphate solubilisation and plant colonisation capacity of Kosakonia radicincitans. J. Adv. Res. 2019, 19, 91–97. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springer Plus 2013, 2, 587. [Google Scholar] [CrossRef]
- Belkebla, N.; Bessai, S.A.; Melo, J.; Caeiro, M.F.; Cruz, C.; Nabti, E.-H. Restoration of Triticum aestivum Growth under Salt Stress by Phosphate-Solubilizing Bacterium Isolated from Southern Algeria. Agronomy 2022, 12, 2050. [Google Scholar] [CrossRef]
- Wejse, P.L.; Ingvorsen, K.; Mortensen, K.K. Purification and characterisation of two extremely halotolerant xylanases from a novel halophilic bacterium. Extremophiles 2003, 7, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Rohban, R.; Amoozegar, M.A.; Ventosa, A. Screening and isolation of halophilic bacteria producing extracellular hydrolyses from Howz Soltan Lake, Iran. J. Ind. Microbiol. Biotechnol. 2008, 36, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Sohail, M.; Ali, S.T. Arthrocnemum macrostachyum biomass: a novel substrate for multienzyme preparation by endophytic Bacillus species. Biomass- Convers. Biorefinery, 2023. [Google Scholar] [CrossRef]
- Giridhar, P.V.; Chandra, T. Production of novel halo-alkali-thermo-stable xylanase by a newly isolated moderately halophilic and alkali-tolerant Gracilibacillus sp. TSCPVG. Process. Biochem. 2010, 45, 1730–1737. [Google Scholar] [CrossRef]
- Siah, A.; Deweer, C.; Duyme, F.; Sanssené, J.; Durand, R.; Halama, P.; Reignault, P. Correlation of in planta endo-beta-1,4-xylanase activity with the necrotrophic phase of the hemibiotrophic fungus Mycosphaerella graminicola. Plant Pathol. 2010, 59, 661–670. [Google Scholar] [CrossRef]
- Moreno, M.D.L.; Pérez, D.; García, M.T.; Mellado, E. Halophilic Bacteria as a Source of Novel Hydrolytic Enzymes. Life 2013, 3, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Badiaa, E.; Abdeljabbar, H.; Mohamed, R.H.; Abdellatif, B.; Najla, S.-Z. In vivo and in vitro evaluation of antifungal activities from a halotolerant Bacillus subtilis strain J9. Afr. J. Microbiol. Res. 2012, 6, 4073–4083. [Google Scholar] [CrossRef]
- Santiago, C.D.; Yagi, S.; Ijima, M.; Nashimoto, T.; Sawada, M.; Ikeda, S.; Asano, K.; Orikasa, Y.; Ohwada, T. Bacterial Compatibility in Combined Inoculations Enhances the Growth of Potato Seedlings. Microbes Environ. 2017, 32, 14–23. [Google Scholar] [CrossRef]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef]
- Valenzuela-Encinas, C.; Neria-González, I.; Alcántara-Hernández, R.J.; Enríquez-Aragón, J.A.; Estrada-Alvarado, I.; Hernández-Rodríguez, C.; Dendooven, L.; Marsch, R. Phylogenetic analysis of the archaeal community in an alkaline-saline soil of the former lake Texcoco (Mexico). Extremophiles 2007, 12, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Töpper, B.; Larsen, A.; Thingstad, T.F.; Thyrhaug, R.; Sandaa, R.-A. Bacterial community composition in an Arctic phytoplankton mesocosm bloom: the impact of silicate and glucose. Polar Biol. 2010, 33, 1557–1565. [Google Scholar] [CrossRef]
- Bano, N.; Musarrat, J. Characterization of a New Pseudomonas aeruginosa Strain NJ-15 as a Potential Biocontrol Agent. Curr. Microbiol. 2003, 46, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Hussein, K.A.; Joo, J.H. Isolation and characterization of rhizomicrobial isolates for phosphate solubilization and indole acetic acid production. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 847–855. [Google Scholar] [CrossRef]
- Arora, N.K.; Verma, M. Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. 3 Biotech 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.M. Iron acquisition in microbial pathogenesis. Trends Microbiol. 1993, 1, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, R., Hajoori, M., Suthar, M., Desai, S., & Bharthana-Vesu, S. Isolation, screening and characterization of marine bacteria for exopolysaccharide production. Bioscience Discovery 2017, 8, 643–9.
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Detection of biofilm-forming strains of Staphylococcus epidermidis and S. aureus. Expert Rev. Mol. Diagn. 2002, 2, 478–484. [Google Scholar] [CrossRef]
- Xie, G.H.; Cui, Z.; Yu, J.; Yan, J.; Hai, W.; Steinberger, Y. Identification of nif genes in N2-fixing bacterial strains isolated from rice fields along the Yangtze River Plain. J. Basic Microbiol. 2006, 46, 56–63. [Google Scholar] [CrossRef]
- Battini, F.; Cristani, C.; Giovannetti, M.; Agnolucci, M. Multifunctionality and diversity of culturable bacterial communities strictly associated with spores of the plant beneficial symbiont Rhizophagus intraradices. Microbiol. Res. 2016, 183, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Parmar, P.; Sindhu, S.S. The novel and efficient method for isolating potassium solubilizing bacteria from rhizosphere soil. Geomicrobiol. J. 2018, 36, 130–136. [Google Scholar] [CrossRef]
- de Freitas, J.R.; Banerjee, M.R.; Germida, J.J. Phosphate-solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica napus L.). Biol. Fertil. Soils 1997, 24, 358–364. [Google Scholar] [CrossRef]
- Murkovic, M.; Derler, K. Analysis of amino acids and carbohydrates in green coffee. J. Biochem. Biophys. Methods 2006, 69, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, P.; Jorquera, M.A.; Sangwan, P.; Kumar, P.; Verma, A.K.; Agrawal, S. Isolation of phytase-producing bacteria from Himalayan soils and their effect on growth and phosphorus uptake of Indian mustard (Brassica juncea). World J. Microbiol. Biotechnol. 2013, 29, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Ramı́rez, M.G.; Avelizapa, L.R.; Avelizapa, N.R.; Camarillo, R.C. Colloidal chitin stained with Remazol Brilliant Blue R®, a useful substrate to select chitinolytic microorganisms and to evaluate chitinases. J. Microbiol. Methods 2004, 56, 213–219. [Google Scholar] [CrossRef]
- Rojas-Avelizapa, L.; Cruz-Camarillo, R.; Guerrero, M.; Rodríguez-Vázquez, R.; Ibarra, J. Selection and characterization of a proteo-chitinolytic strain of Bacillus thuringiensis, able to grow in shrimp waste media. World J. Microbiol. Biotechnol. 1999, 15, 299–308. [Google Scholar] [CrossRef]
- Sundaramoorthy, S.; Raguchander, T.; Ragupathi, N.; Samiyappan, R. Combinatorial effect of endophytic and plant growth promoting rhizobacteria against wilt disease of Capsicum annum L. caused by Fusarium solani. Biol. Control. 2012. [Google Scholar] [CrossRef]
- Kassambara, A., Mundt, F., Package ‘factoextra’. Extract and visualize the results of multivariate data analyses. 2017, Available online: http://www.sthda.com/english/rpkgs/factoextra.
Figure 1.
Phylogenetic tree established by the neighbor-joining method based on 16S rRNA sequences and closely related sequences.
Figure 1.
Phylogenetic tree established by the neighbor-joining method based on 16S rRNA sequences and closely related sequences.
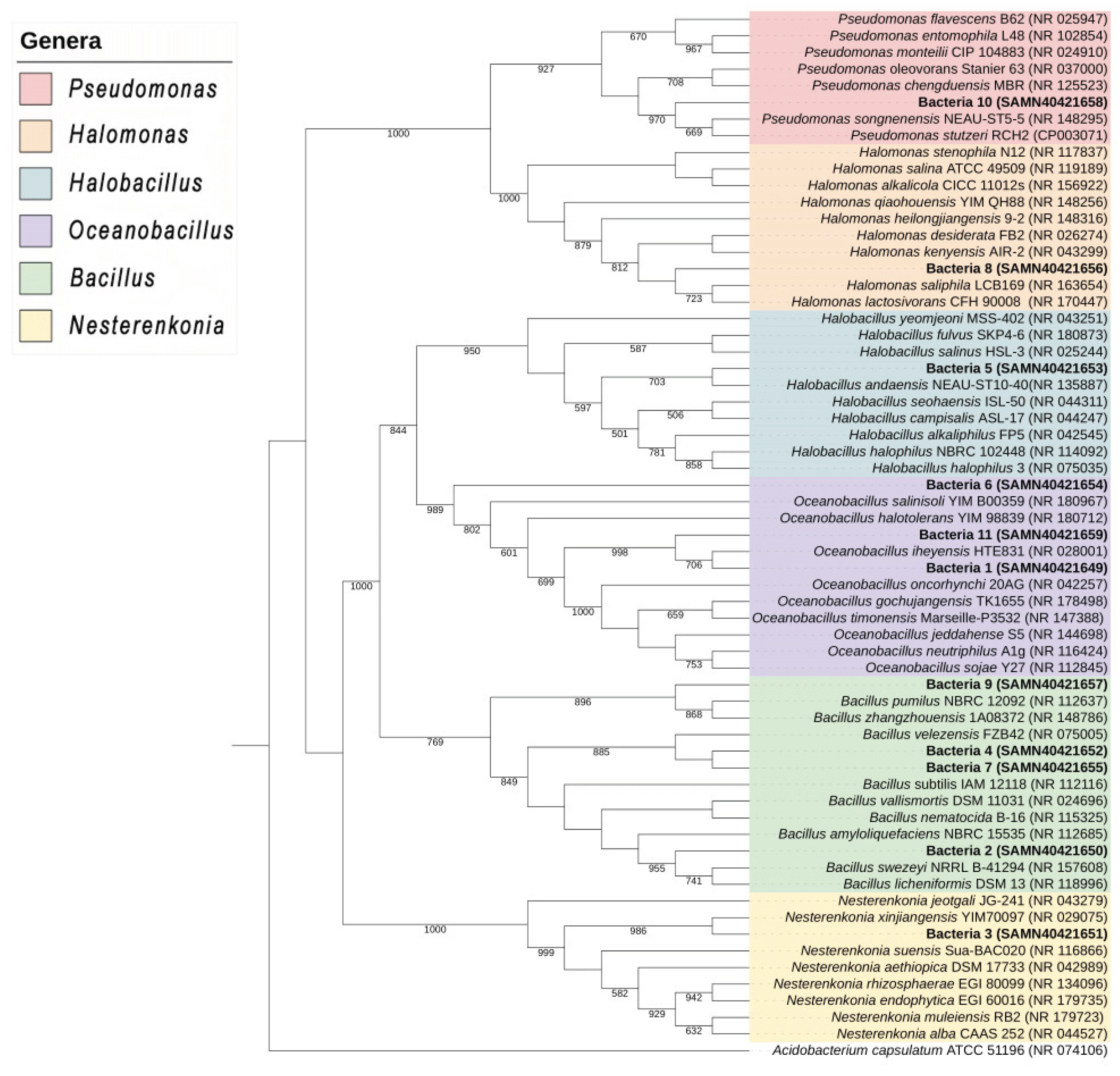
Figure 2.
Effect of NaCl on indol acetic acid (a) and siderophores (b) production by endophytic halophytic bacteria isolated from halophytes. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the production of IAA and siderophores when comparing endophytic bacteria in 2.5 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 0 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05).
Figure 2.
Effect of NaCl on indol acetic acid (a) and siderophores (b) production by endophytic halophytic bacteria isolated from halophytes. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the production of IAA and siderophores when comparing endophytic bacteria in 2.5 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 0 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05).
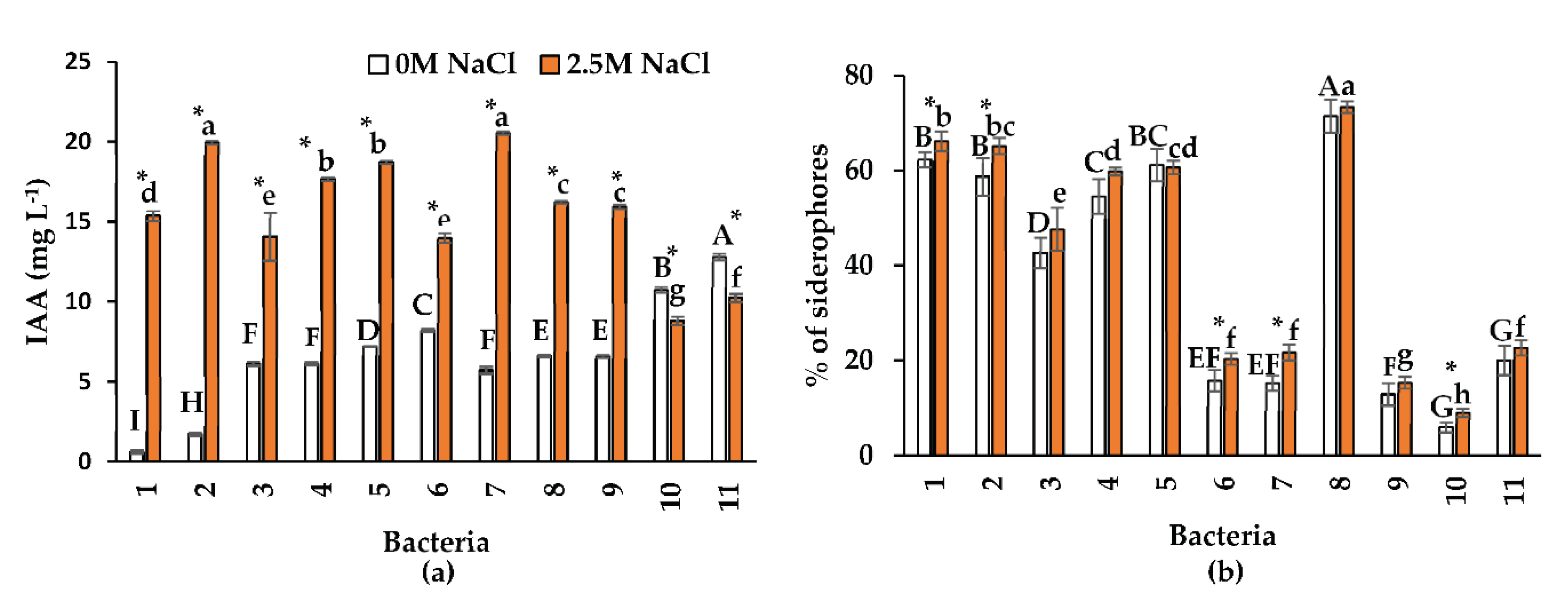
Figure 3.
Soluble P concentration and pH in the free-bacteria extract after solubilization test (48 h) by halophilic endophytic bacteria in two NaCl concentrations. (a,b) when using Ca3(PO4)2, or (c,d) phytic acid. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the soluble P concentration and pH when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05).
Figure 3.
Soluble P concentration and pH in the free-bacteria extract after solubilization test (48 h) by halophilic endophytic bacteria in two NaCl concentrations. (a,b) when using Ca3(PO4)2, or (c,d) phytic acid. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the soluble P concentration and pH when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05).
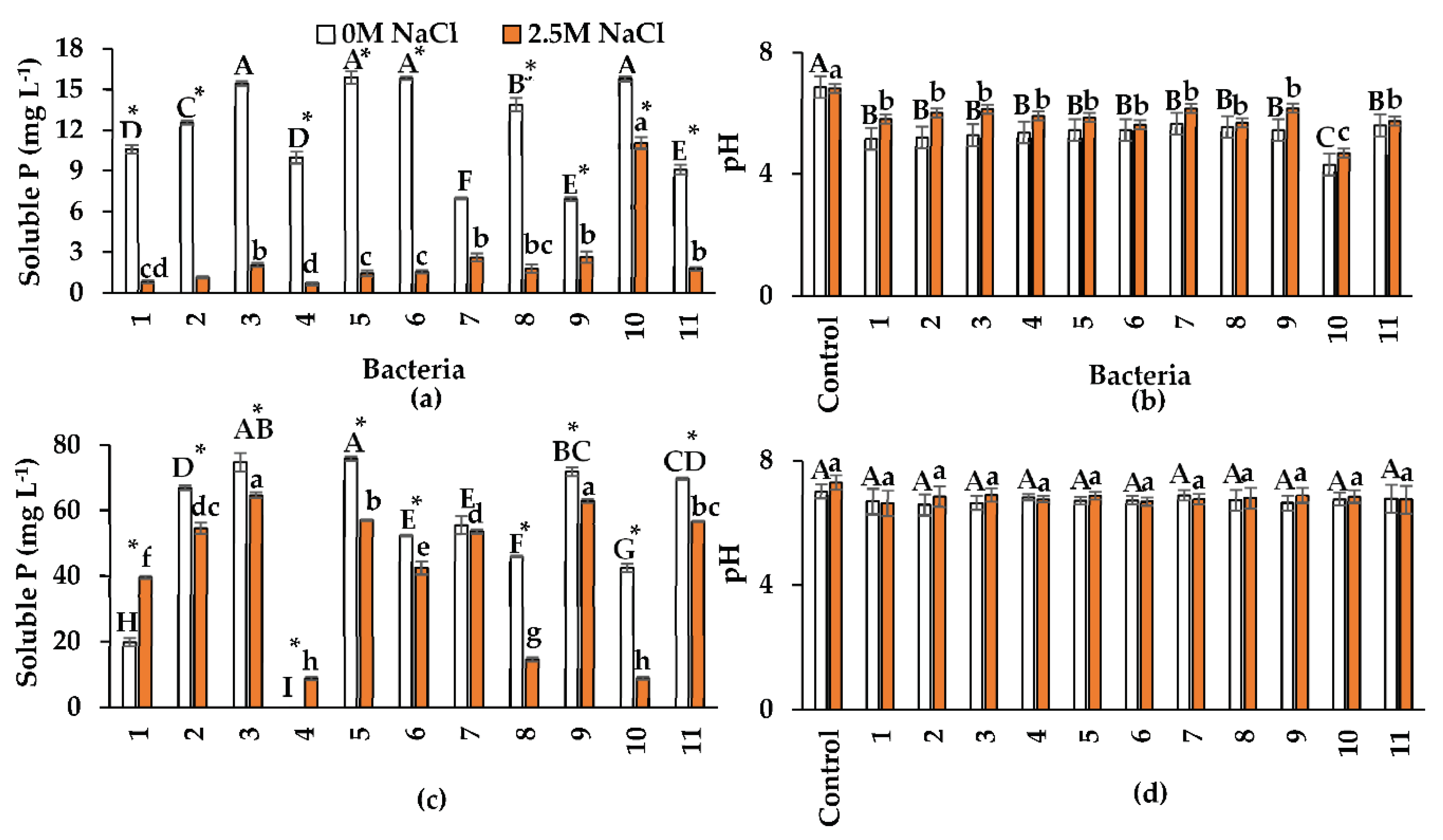
Figure 4.
Soluble concentration of K (a, b), Mn (c, d) and Zn (e, f), and pH in the free-bacteria extract after solubilization test (48 h) by halophilic endophytic bacteria in two NaCl concentrations. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the soluble P concentration and pH when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *Shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05). e) A better visualization of results obtained with NaCl is shown.
Figure 4.
Soluble concentration of K (a, b), Mn (c, d) and Zn (e, f), and pH in the free-bacteria extract after solubilization test (48 h) by halophilic endophytic bacteria in two NaCl concentrations. Values correspond to average ± standard deviation, n=3. Different capital letters show differences in the soluble P concentration and pH when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *Shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05). e) A better visualization of results obtained with NaCl is shown.
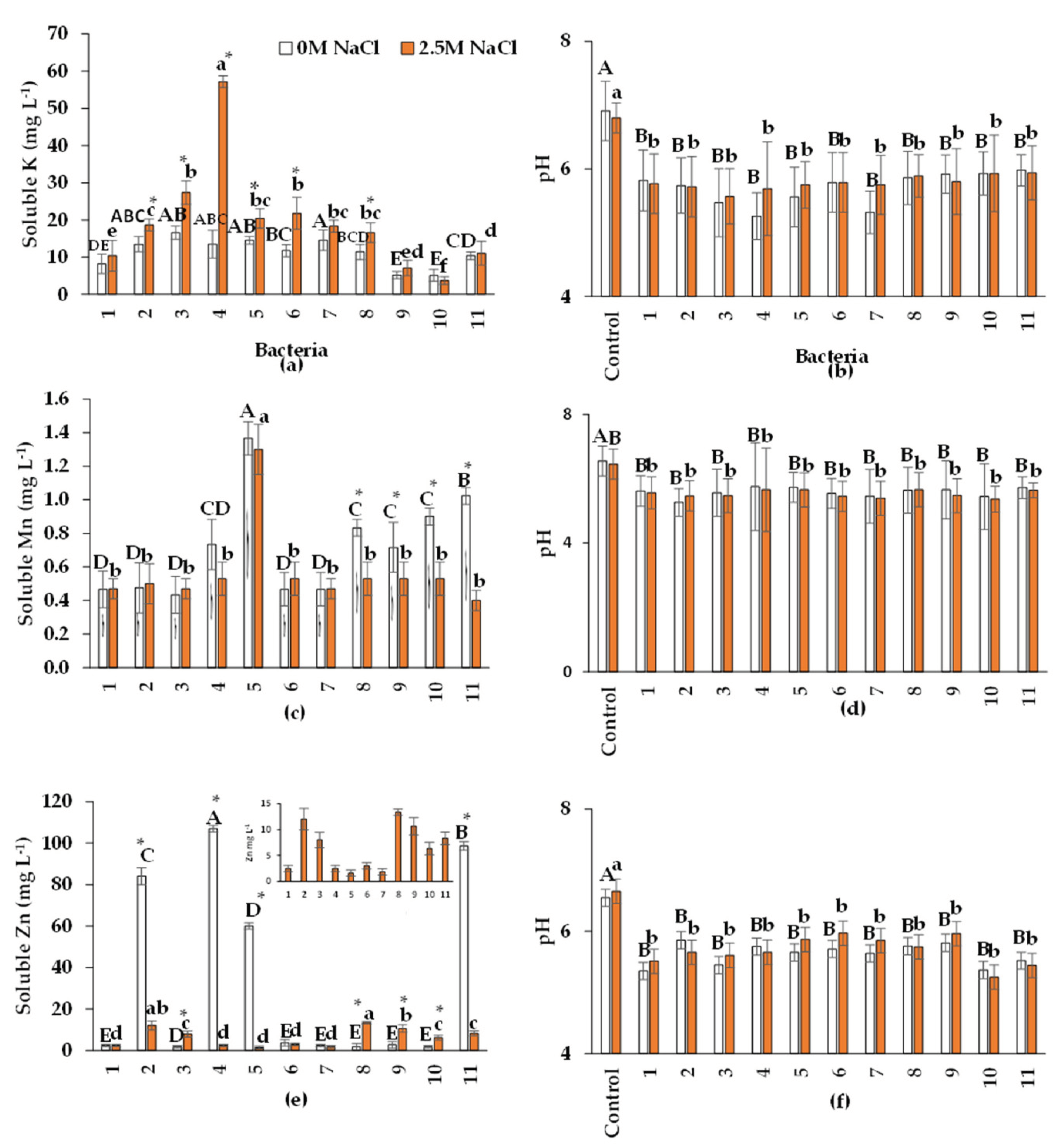
Figure 5.
Ven diagram presenting the organic acids secreted in each solubilization ion and common among solubilization tests.
Figure 5.
Ven diagram presenting the organic acids secreted in each solubilization ion and common among solubilization tests.
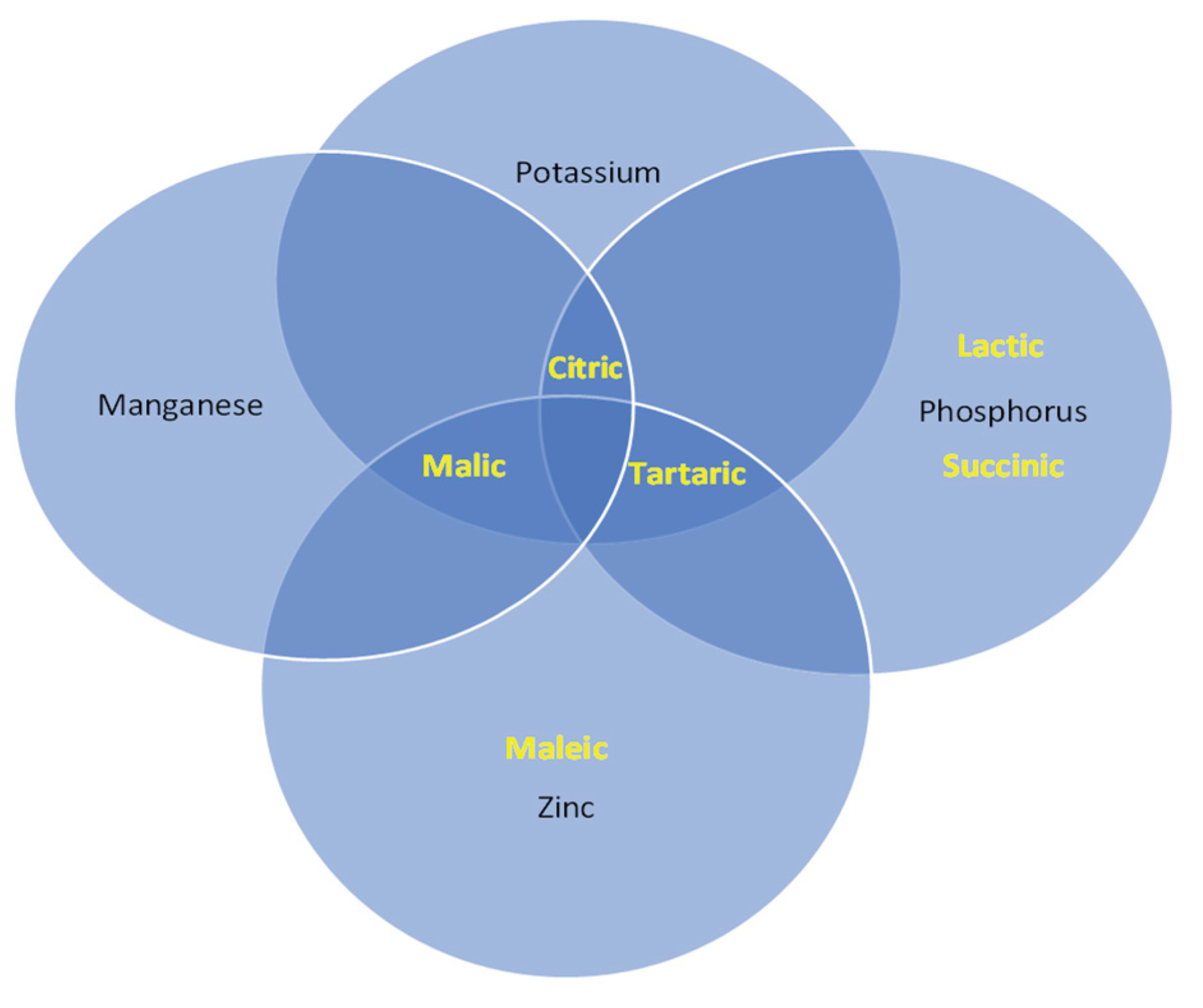
Figure 6.
Bacterial compatibility of one selected consortium with beneficial properties such as plant growth promotion, nutrient solubilization, and enzyme production.
Figure 6.
Bacterial compatibility of one selected consortium with beneficial properties such as plant growth promotion, nutrient solubilization, and enzyme production.
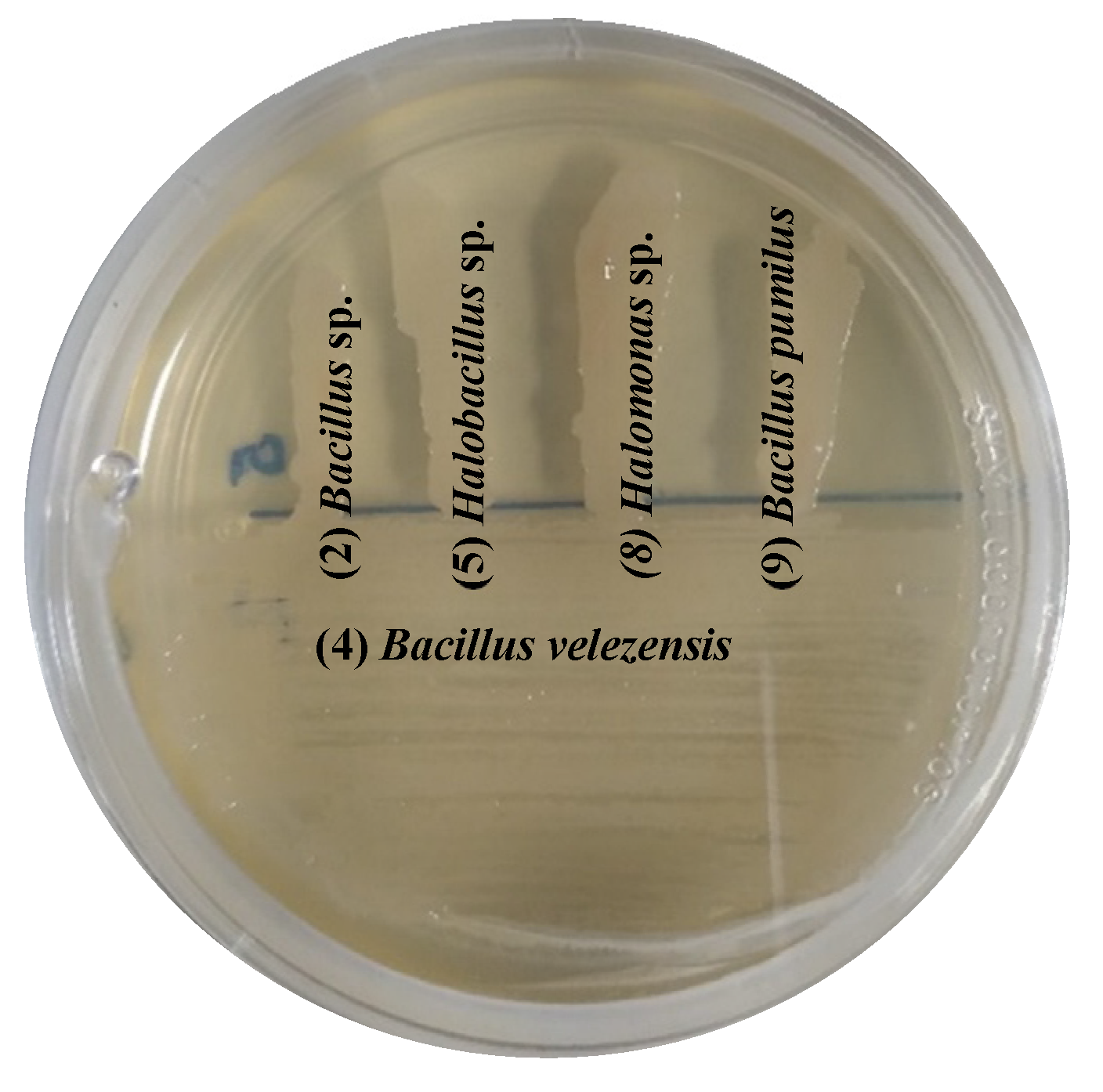
Figure 7.
Principal component analysis (PCA) from the biochemical characterization of halophilic endophytic bacteria isolated from six halophytes. (a) Biochemical variables analyzed in different NaCl concentrations. Circles show similar response in the both NaCl concentrations tested. (b) Distribution of bacteria in the PCA. Variables showed in black for 0 M NaCl and in red for 2.5 M NaCl. Indoleacetic acid (IAA), exopolysaccharides (EPS), soluble manganese (Mn), soluble potassium (K), soluble phosphate (Pi) from solubilization with Ca3(PO4)2, soluble phosphate (P) from phytic acid solubilization test, soluble zinc (Zn). The numbers represent the bacterial isolates.
Figure 7.
Principal component analysis (PCA) from the biochemical characterization of halophilic endophytic bacteria isolated from six halophytes. (a) Biochemical variables analyzed in different NaCl concentrations. Circles show similar response in the both NaCl concentrations tested. (b) Distribution of bacteria in the PCA. Variables showed in black for 0 M NaCl and in red for 2.5 M NaCl. Indoleacetic acid (IAA), exopolysaccharides (EPS), soluble manganese (Mn), soluble potassium (K), soluble phosphate (Pi) from solubilization with Ca3(PO4)2, soluble phosphate (P) from phytic acid solubilization test, soluble zinc (Zn). The numbers represent the bacterial isolates.
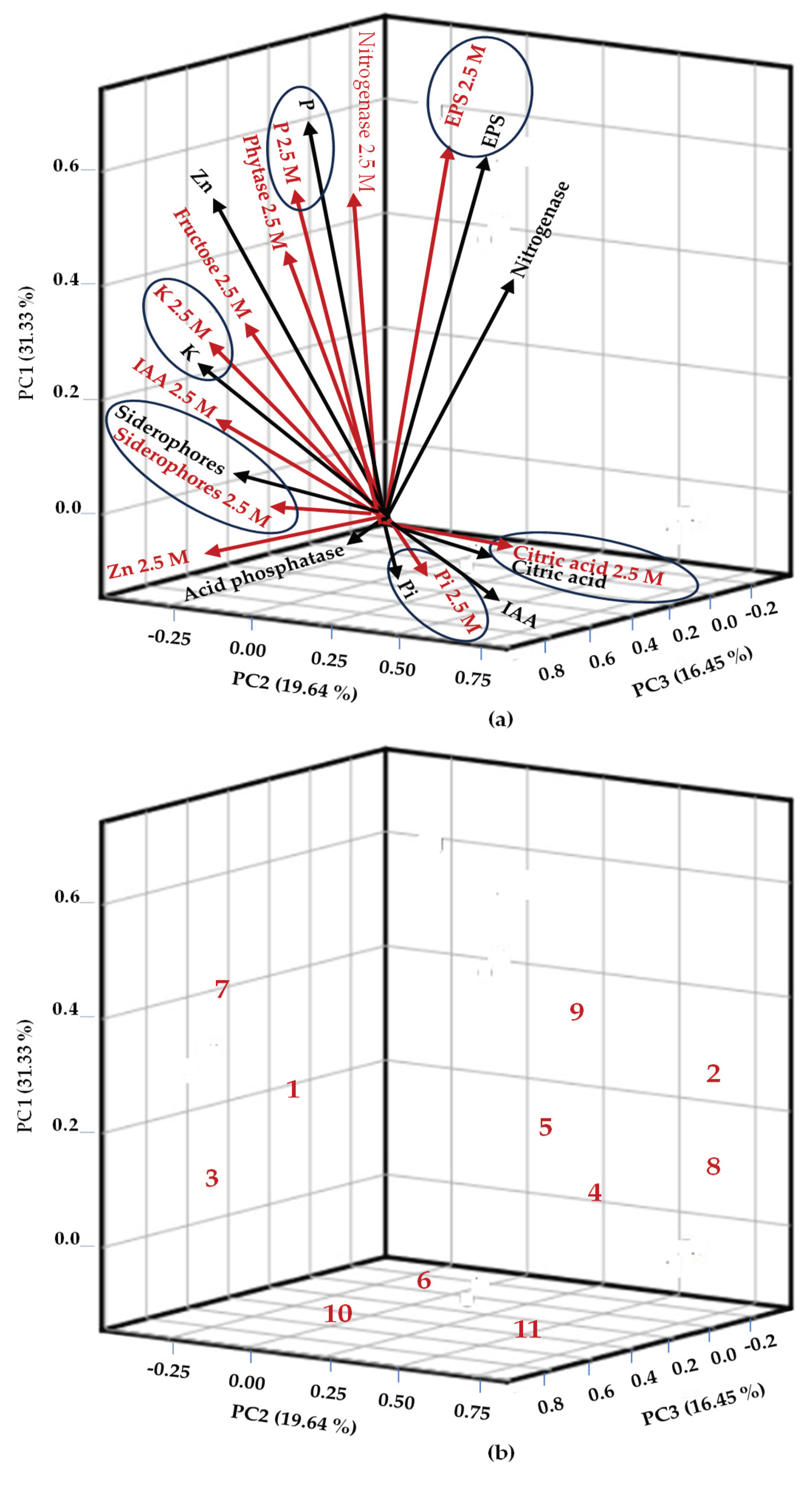

Table 1.
Electrical conductivity (EC) of rhizosphere soil and molecular identification of endophyte bacteria isolated from six halophytes.
Table 1.
Electrical conductivity (EC) of rhizosphere soil and molecular identification of endophyte bacteria isolated from six halophytes.
| Halophyte | EC (dS m-1) | Bacterial code | Molecular identification | |
| Sequence homology (% identity) | NCBI accession number | |||
| D. spicata | 16 | 1 | 99 | SAMN40421649 |
| D. spicata | 12 | 2 | 99 | SAMN40421650 |
| K. scoparia | 7 | 3 | 99 | SAMN40421651 |
| S. torreyana | 12 | 4 | 99 | SAMN40421652 |
| C. dactylon | 6 | 5 | 99 | SAMN40421653 |
| D. spicata | 8 | 6 | 99 | SAMN40421654 |
| K. scoparia | 5 | 7 | 99 | SAMN40421655 |
| K. scoparia | 8 | 8 | 99 | SAMN40421656 |
| E. obtusiflora | 9 | 9 | 99 | SAMN40421657 |
| S. torreyana | 7 | 10 | 99 | SAMN40421658 |
| B. salicifolia | 6 | 11 | 99 | SAMN40421659 |
Table 2.
Qualitative nitrogenase determination and exopolysaccharides produced by halophilic endophytic bateria isolated from halophytes.
Table 2.
Qualitative nitrogenase determination and exopolysaccharides produced by halophilic endophytic bateria isolated from halophytes.
| Bacteria | Nitrogenase enzyme | Exopolysaccharide | ||
| NaCl 0M | NaCl 2.5M |
NaCl 0M | NaCl 2.5M | |
| 1 | - | - | + | + |
| 2 | - | - | - | - |
| 3 | + | + | - | - |
| 4 | + | +++ | + | + |
| 5 | - | - | + | + |
| 6 | - | - | + | + |
| 7 | + | + | - | - |
| 8 | +++ | ++ | - | - |
| 9 | ++ | +++ | - | - |
| 10 | - | +++ | - | - |
| 11 | - | - | - | - |
-,+,++,+++ denotes: none, low, moderate and high nitrogenase enzyme response, respectively in 11 endophytic bacteria. Exopolysaccharide production: (+) with or (-) without production. n=3. .
Table 3.
Organic acids (mg L-1) produced by halophilic endophytic bacteria in NBRIP- Ca3(PO4)2 broth in the presence or not of NaCl.
Table 3.
Organic acids (mg L-1) produced by halophilic endophytic bacteria in NBRIP- Ca3(PO4)2 broth in the presence or not of NaCl.
| 0 M NaCl | 2.5 M NaCl | |||||||
| Bacteria | Citric | Succinic | Lactic | Tartaric | Citric | Succinic | Lactic | Tartaric |
| 1 | 6.3±0.4B* | 1.8±0.2A | 4.7±0.3A | nd | 7.2±0.1b* | 1.3±0.1a | 4.20±0.2a | nd |
| 2 | 6.2±0.4B* | 1.4±0.4A | 5.2±0.3A* | nd | 4.6±0.1b* | 1.0±0.2a | 4.0±0.2a* | nd |
| 3 | 1.4±0.2C | 1.2±0.1A | 0.4±0.2B* | 1.8±0.1A | 1.9±0.2b | 1.2±0.1a | nd* | 1.4±0.1a |
| 4 | 15.5±0.5A | 0.4±0.1A | 0.8±0.2B | nd | 15.2±0.3a | 0.8±0.3a | 1.2±0.2b | nd |
| 5 | 5.6±0.3B | 0.6±0.3A | 1.0±0.1B | nd | 5.2±0.1b | 0.4±0.3a | 1.0±0.2b | nd |
| 6 | 5.6±0.6B | 0.8±0.3A | 0.6±0.1B* | nd | 6.5±0.3b | 1.0±0.4a | 1.8±0.2b* | nd |
| 7 | 2.2±0.1C | 1.4±0.2A | 0.6±0.2B* | nd | 2.8±0.3b | 1.2±0.4a | 1.4±0.3b* | nd |
| 8 | 1.4±0.1C* | 0.4±0.2A* | 1.4±0.2B* | nd | 2.1±0.5b* | 1.0±0.1a* | nd* | nd |
| 9 | 2.6±0.1C | 1.0±0.1A | 1.0±0.3B* | 1.4±0.1A | 2.4±0.2b | 1.4±0.1a | nd* | 1.0±0.2a |
| 10 | 3.2±0.3C | 1.2±0.1A | 3.2±0.5A* | nd | 3.8±0.2c | 1.0±0.2a | 4.0±0.1a* | nd |
| 11 | 3.1±0.1C | 1.0±0.2A | 1.8±0.2B | nd | 3.6±0.1c | 1.0±0.2a | 2.0±0.1b | nd |
Average ± and standard deviation, n=3. Different capital letters show differences when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences in organic acid production when comparing endophytic bacteria in 2.5 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05). Nd=no detected.
Table 4.
Organic acids (mg L-1) produced by endophytic bacteria when solubilizing K, Mn, and Zn are influenced by 2.5 M NaCl concentration.
Table 4.
Organic acids (mg L-1) produced by endophytic bacteria when solubilizing K, Mn, and Zn are influenced by 2.5 M NaCl concentration.
| 0 M NaCl | 2.5 M NaCl | |||||||||
| Bacteria | Citric | Malic | Tartaric | Maleic | Vanillic | Citric | Malic | Tartaric | Maleic | Vanillic |
| K | ||||||||||
| 1 | 5.2±0.1B* | 0.4±0.1C | 1.7±0.2B* | Nd | Nd | 4.0±0.2b* | 0.8±0.1b | 2.8±0.1b* | Nd | Nd |
| 2 | 4.8±0.1B | 0.8±0.1C* | 1.3±0.2B | Nd | Nd | 4.7±0.3b | 1.6±0.2b* | 2.0±0.1b | Nd | Nd |
| 3 | 1.9±0.3C | 1.0±0.1C* | 8.9±0.1A* | Nd | Nd | 2.2±0.1b | 1.8±0.2b* | 2.2±0.1b* | Nd | Nd |
| 4 | 0.5±0.5C* | 3.2±0.2B | 2.0±0.1B* | Nd | Nd | 1.2±0.1c* | 2.8±0.1b | 10.8±0.2a* | Nd | Nd |
| 5 | 4.2±0.2B | 2.0±0.3B | 2.0±0.3B* | Nd | Nd | 3.9±0.2b | 1.6±0.3b | 3.7±0.2b* | Nd | Nd |
| 6 | 1.2±0.2C | 2.0±0.2B | 1.8±0.5B* | Nd | Nd | 0.7±0.2c | 1.8±0.3b | 4.2±0.3b* | Nd | Nd |
| 7 | 4.3±0.1C* | 1.2±0.2C | 8.8±0.5A* | Nd | Nd | 1.0±0.3c* | 1.7±0.4b | 4.7±0.4b* | Nd | Nd |
| 8 | 4.7±0.1B | 1.5±0.1C | 1.0±0.2B* | Nd | 6.0±0.1 | 4.6±0.5b | 1.6±0.4b | 7.5±0.6a* | Nd | 6.2±0.2 |
| 9 | 11.1±0.2A | 7.6±0.1A* | 1.7±0.1B | Nd | Nd | 10.5±0.5a | 8.7±0.1a* | 2.4±0.4b | Nd | Nd |
| 10 | 14.0±0.2* | 0.5±0.3C* | 1.3±0.2B* | Nd | Nd | 4.7±0.2b* | 1.2±0.2b* | 2.8±0.3b* | Nd | Nd |
| 11 | 4.9±0.1B | 1.0±0.2C | 1.0±0.3B* | Nd | Nd | 5.0±0.1b | 1.5±0.1b | 2.2±0.1b* | Nd | Nd |
| Mn | ||||||||||
| 1 | 8.9±0.3B* | 4.8±0.2B* | Nd | Nd | Nd | 10.5±0.3b* | 7.4±0.1a* | Nd | Nd | Nd |
| 2 | 9.0±0.4B* | 4.9±0.2B* | Nd | Nd | Nd | 10.2±0.4b* | 7.2±0.2a* | Nd | Nd | Nd |
| 3 | 3.8±0.1C | 0.4±0.1C | Nd | Nd | Nd | 3.4±0.2c | 0.6±0.2b | Nd | Nd | Nd |
| 4 | 5.0±0.1C | Nd | Nd | Nd | Nd | 5.4±0.1c | Nd | Nd | Nd | Nd |
| 5 | 4.7±0.2C | Nd | Nd | Nd | Nd | 5.0±0.1c | Nd | Nd | Nd | Nd |
| 6 | 4.0±0.1C | Nd | Nd | Nd | Nd | 4.8±0.2c | Nd | Nd | Nd | Nd |
| 7 | 4.2±0.2C | Nd | Nd | Nd | Nd | 4.0±0.4c | Nd | Nd | Nd | Nd |
| 8 | 3.0±0.2C* | Nd | Nd | Nd | Nd | 4.6±0.2c* | Nd | Nd | Nd | Nd |
| 9 | 4.0±0.1C | 7.8±0.2A* | Nd | Nd | Nd | 4.6±0.3c | 6.8±0.2a* | Nd | Nd | Nd |
| 10 | 15.8±0.1A | Nd | Nd | Nd | Nd | 16.0±0.3a | Nd | Nd | Nd | Nd |
| 11 | 9.2±0.2B | Nd | Nd | Nd | Nd | 10.0±0.1b | Nd | Nd | Nd | Nd |
| Zn | ||||||||||
| 1 | Nd | 3.0±0.1A | 4.5±0.2B | 3.4±0.1B* | Nd | Nd | 3.2±0.1a | 4.0±0.1b | 2.8±0.1b* | Nd |
| 2 | Nd | 3.5±0.2A* | 3.6±0.2B | 3.0±0.2B | Nd | Nd | 2.0±0.2a* | 3.2±0.2b | 2.6±0.2b | Nd |
| 3 | Nd | 2.6±0.1A* | 2.8±0.1B | 1.2±0.4C | Nd | Nd | 4.7±0.1a* | 3.0±0.1b | 1.6±0.5c | Nd |
| 4 | Nd | 2.8±0.2A* | Nd | 11.8±0.1A | Nd | Nd | 3.4±0.2a* | Nd | 10.4±0.4a | Nd |
| 5 | 8.0±0.3A | 4.3±0.3A | Nd | 5.0±0.2B* | Nd | 7.8±0.3a | 4.7±0.3a | Nd | 3.6±0.3b* | Nd |
| 6 | Nd | 1.2±0.3B* | Nd | 4.8±0.2B | Nd | Nd | 4.0±0.3a* | Nd | 5.6±0.2b | Nd |
| 7 | Nd | 1.0±0.4B* | 3.4±0.1B | 1.0±0.3C* | Nd | Nd | 3.7±0.5a* | 4.0 | 2.8±0.3b* | Nd |
| 8 | 9.2±0.2A | 3.4±0.1A | 11.8±0.1A | 5.4±0.4B* | Nd | 9.3±0.1a | 3.2±0.4a | 11.6±0.3a | 6.4±0.1a* | Nd |
| 9 | Nd | 3.0±0.2A | Nd | 1.4±0.1C* | Nd | Nd | 3.8±0.5a | Nd | 10.5±0.2a* | Nd |
| 10 | 9.2±0.1A | 1.3±0.1B | Nd | 2.9±0.4C* | Nd | 8.6±0.2a | 2.0±0.3a | Nd | 4.7±0.1a* | Nd |
| 11 | Nd | 2.8±0.1A | 5.0±0.2B | 3.0±0.2B | Nd | Nd | 2.2±0.1a | 4.6±0.2b | 3.8±0.2b | Nd |
Average ± and standard deviation, n=3. Different capital letters show differences in organic acid production when comparing endophytic bacteria in 0 M NaCl. Lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05). Nd=not detected.
Table 5.
Sugar production by halophilic endophyte bacteria in different ion-solubilization tests under NaCl effect.
Table 5.
Sugar production by halophilic endophyte bacteria in different ion-solubilization tests under NaCl effect.
| 0 M NaCl | 2.5 M NaCl | |||||||||
| Bacteria | Ca3(PO4)2 | Phytate | K | Mn | Zn | Ca3(PO4)2 | Phytate | K | Mn | Zn |
| Fructose | ||||||||||
| 1 | 55.3±0.4D* | 20.4±1.8B | 129.7±0.1B* | 3.6±0.1C | 3.7±0.4G | 61.3±1.6d* | 21.6±0.8b | 160.5±1.9b* | 3.4±0.1d | 4.0±0.4h |
| 2 | 57.5±0.5D* | 2.0±0.6D | 122.8±0.7B* | 4.7±0.2C | 4.7±0.5G | 65.7±2.5d* | 1.8±0.5c | 151.0±2.5b* | 5.0±0.5d | 4.7±0.3h |
| 3 | 295.4±1.2B* | 1.7±0.1D | 16.6±0.5E | 16.5±0.4B | 111.1±0.2D | 333.4±1.2b* | 2.8±0.6c | 16.8±0.1e | 18.2±0.2c | 114.8±05c |
| 4 | 458.8±0.3A | 2.3±0.5D | 157.5±1.5A* | 32.0±0.2A | 414.0±0.3A* | 459.4±3.0a | 4.7±1.0c | 267.6±0.2a* | 33.4±0.4a | 349.4±0.5a* |
| 5 | Nd | 1.7±0.1D | 3.5±1.0F* | 24.7±0.2B | 343.5±0.2B* | Nd | 2.2±0.3c | 8.4±1.0g* | 23.9±0.3b | 83.3±1.0d* |
| 6 | 153.1±0.2C* | 93.1±1.0A | 113.3±1.4B* | 18.7±0.5B | 275.3±0.4C* | 308.6±2.0b* | 95.1±0.5a | 128.3±0.1c* | 20.6±0.3b | 282.3±0.5b* |
| 7 | 463.8±0.4A* | 2.2±0.3D | 153.1±0.7A* | 35.0±0.3A* | 417.9±0.4A* | 456.2±0.5a | 2.4±0.3c | 264.8±0.1a* | 32.0±0.5a* | 347.4±0.5a* |
| 8 | 2.3±0.5F* | 1.9±0.3D | 35.3±0.2D* | 16.4±0.2B | 16.0±0.5F | 6.7±0.6e | 1.5±0.4c | 43.8±0.1e* | 17.4±0.3c | 16.1±0.5g |
| 9 | 151.7±0.6C* | 16.2±1.4B | 70.8±0.1C* | 1.9±0.6D* | 3.5±0.6G | 322.8±1.0b* | 18.5±0.9b | 113.1±0.2d* | 3.6±0.2d* | 2.9±0.3h |
| 10 | 184.5±0.2C* | 7.0±0.3C | 78.1±0.4C* | 13.1±0.1B* | 38.1±0.2E | 226.7±0.4c* | 7.3±0.1c | 104.0±0.2d* | 16.3±0.5c* | 39.1±0.1e |
| 11 | 8.7±0.1E* | 1.8±0.4D | 16.7±0.4E | 9.5±0.3C | 18.8±0.1F* | 19.6±0.6e* | 2.5±0.2c | 18.7±0.1f* | 8.7±0.1d | 25.8±0.1f* |
| Xylose | ||||||||||
| 1 | Nd | Nd | 6.4±0.3B | Nd | Nd | Nd | Nd | 7.4±0.1b | Nd | Nd |
| 2 | Nd | Nd | 8.6±0.4B | Nd | Nd | Nd | Nd | 9.6±0.2b | Nd | Nd |
| 3 | Nd | 8.5±0.2A | 67.0±0.4A | Nd | 23.2±0.4A | Nd | 8.0±0.4a | 68.0±0.4a | Nd | 24.6±0.6a |
| 4 | Nd | 6.4±0.1AB | 65.7±0.1A | 3.6±0.1A | Nd | Nd | 7.8±0.4ab | 67.7±0.5a | 4.8±0.1a | Nd |
| 5 | 29.0±1.5B* | 5.70.1B | 5.4±0.2B | Nd | Nd | 228.1±2.8a* | 6.40.2b | 6.2±0.6b | Nd | Nd |
| 6 | Nd | Nd | 8.2±0.1B | Nd | Nd | Nd | Nd | 9.4±0.1b | Nd | Nd |
| 7 | Nd | 3.4±0.2C | 70.1±0.3A | 3.8±0.2A | Nd | Nd | 4.2±0.1b | 73.1±0.1a | 4.6±0.3a | Nd |
| 8 | Nd* | Nd | Nd | Nd | Nd | 4.4±0.4b* | Nd | Nd | Nd | Nd |
| 9 | Nd | Nd | 66.6±0.4A | Nd | 24.6±0.3A | Nd | Nd | 68.6±0.2a | Nd | 25.7±1.0a |
| 10 | 229.5±1.8A* | Nd | Nd | Nd | Nd | Nd* | Nd | Nd | Nd | Nd |
| 11 | Nd | Nd | 6.4±0.6B | Nd | Nd | Nd | Nd | 7.8±0.3b | Nd | Nd |
Average ± and standard deviation, n=3. Different capital letters show differences in sugar production when comparing endophytic bacteria in 0 M NaCl. Different lowercase letters show differences when comparing endophytic bacteria in 2.5 M NaCl. *shows a significant difference when comparing the same bacteria in 0 or 2.5 M NaCl. In all cases Tukey (α=0.05). Nd=not detected.
Table 6.
Solublization-phosphate related enzymatic activity in halophilic endophytic bacteria in three NaCl concentrations.
Table 6.
Solublization-phosphate related enzymatic activity in halophilic endophytic bacteria in three NaCl concentrations.
|
Alkaline phosphatase (Nitrophenylphosphate µg mL-1 h-1) |
Acid phosphatase (Nitrophenylphosphate µg mL-1 h-1) |
|||||
| NaCl (M) | NaCl (M) | |||||
| Bacteria | 0 | 1.5 | 2.5 | 0 | 1.5 | 2.5 |
| 1 | 27.62±2.3bA | 8.38±2.8aB | 3.1±0.9cC | Nd | 52.56±2.7bA | 42.36±1.8cB |
| 2 | 34.45±2.7aA | 7.75±2.8aB | 3.1±0.9cC | Nd | 59.67±3.2aA | 48.85±2.4bB |
| 3 | 18.00±2.3cA | 8.62±2.3aB | 7.75±1.2aB | Nd | 49.78±2.7bA | 49.27±3.8bA |
| 4 | 28.24±2.8bA | 6.20±2.4aB | 4.64±1.9bB | 47.00±1.4bB | 60.29±4.1aA | 57.82±2.9aA |
| 5 | 16.44±1.9cA | 9.86±1.9aB | 4.34±1.4bC | 44.94±1.4bB | 62.87±2.8aA | 62.43±2.3aA |
| 6 | 27.00±2.4bA | 6.44±1.8aB | 6.82±1.8bB | 27.11±2.3dC | 60.29±4.1aA | 50.71±2.4bB |
| 7 | 23.27±1.8bA | 3.58±1.4bB | 3.79±1.9cB | 23.81±3.2dC | 60.29±4.1aA | 42.67±1.9bB |
| 8 | Nd | 8.51±1.9aA | 9.24±1.9aA | Nd | 62.87±2.8aA | 42.05±1.4bB |
| 9 | 29.79±2.3bA | 9.31±1.9aB | 8.69±1.9aB | 33.19±1.7cC | 63.80±2.8aA | 51.94±2.3bB |
| 10 | 25.14±1.8bA | 5.51±1.9aB | 9.55±1.9aB | 51.32±1.2aC | 62.15±2.9aA | 51.33±1.4bB |
| 11 | 17.69±2.4cA | 9.55±1.9aB | 7.44±1.4abB | 39.27±5.2cC | 51.33±1.7bA | 44.21±1.8cB |
| Interval | 16.44-34.45 | 3.58-9.86 | 3.1-9.25 | 23.81-51.32 | 49.78-63.80 | 42.05-62.43 |
|
Phytase (Phosphate µg mL-1 h-1) |
||||||
| NaCl (M) | ||||||
| Bacteria | 0 | 1.5 | 2.5 | |||
| 1 | 80.21±1.8bC | 107.12±2.1bB | 124.16±2.8cA | |||
| 2 | 70.01±1.2cC | 100.15±2.8bB | 122.18±2.4cA | |||
| 3 | 90.12±2.8aA | 87.56±3.2cA | 94.75±2.5dA | |||
| 4 | 4.24±2.4fC | 67.24±3.6dB | 117.74±2.8cA | |||
| 5 | 90.38±2.2aC | 100.08±3.8bB | 144.53±94bA | |||
| 6 | 70.45±2.6cC | 87.34±3.3cB | 110.22±2.9cA | |||
| 7 | 70.56±2.1cC | 100.75±3.1bB | 164.72±3.1aA | |||
| 8 | Nd | 84.74±3.4cB | 144.94±3.5bA | |||
| 9 | 90.01±2.6aC | 120.85±3.6aB | 154.42±3.4aA | |||
| 10 | 50.83±2.8dC | 104.16±3.1bB | 140.74±3.1bA | |||
| 11 | 40.12±2.6eC | 97.12±3.5bB | 147.48±3.2bA | |||
| Interval | 4.24-90.38 | 67.24-107.12 | 94.75-164.72 | |||
Lowercase letters show differences when comparing enzymatic activity between bacteria at their respective NaCl concentration. Different capital letters show differences when comparing different concentrations of NaCl in the same bacteria. In all cases Tukey (α=0.05). Nd=not detected.
Table 7.
Hydrolytic enzymes activity in halophilic endophytic bacteria at three NaCl concentrations.
Table 7.
Hydrolytic enzymes activity in halophilic endophytic bacteria at three NaCl concentrations.
|
Xylanase (µg mL-1 h-1) |
Chitinase (µg mL-1 h-1) |
|||||
| NaCl (M) | NaCl (M) | |||||
| Bacteria | 0 | 1.5 | 2.5 | 0 | 1.5 | 2.5 |
| 1 | Nd | Nd | Nd | 1.62±0.931aB | 6.85±1.021aA | 6.70±0.923aA |
| 2 | Nd | Nd | Nd | 1.90±0.161aB | 6.43±0.212aA | 5.48±0.546aA |
| 3 | 0.09±0.007aB | 0.76±0.004bB | 3.14±0.005bA | 1.67±0.112aB | 6.92±0.283aA | 6.54±0.283aA |
| 4 | 0.14±0.012aB | 0.68±0.005bB | 3.01±0.010bA | 1.24±0.932aB | 6.48±0.827aA | 5.45±1.093aA |
| 5 | 0.06±0.008aC | 0.62±0.004bB | 2.14±0.003bA | 1.45±0.836aB | 6.87±0.927aA | 6.30±0.864aA |
| 6 | 0.06±0.008aC | 0.67±0.006bB | 2.92±0.005bA | 1.86±0.732aB | 6.56±1.052aA | 6.28±0.834aA |
| 7 | 0.02±0.034aC | 0.76±0.005bB | 2.74±0.008bA | 1.90±0.623aB | 5.68±1.024aA | 6.34±0.941aA |
| 8 | Nd | 0.79±0.004bB | 2.80±0.005bA | Nd | 6.14±0.962aA | 5.85±0.841aA |
| 9 | 0.09±0.005aC | 0.83±0.004bB | 3.22±0.007bA | 1.78±0.924aB | 6.24±0.863aA | 6.35±0.974aA |
| 10 | Nd | Nd | Nd | Nd | Nd | Nd |
| 11 | 0.08±0.015aC | 1.89±0.010aB | 5.32±0.008aA | Nd | Nd | Nd |
| Interval | 0.02-0.14 | 0.62-1.89 | 2.14-5.32 | 1.24-1.90 | 5.68-6.92 | 5.45-6.70 |
Lowercase letters show differences when comparing enzymatic activity between bacteria at their respective NaCl concentration. Different capital letters show differences when comparing different concentrations of NaCl in the same bacteria. In all cases Tukey (α=0.05). Nd=not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



