
Submitted:
22 March 2024
Posted:
22 March 2024
You are already at the latest version
Abstract
Acetamiprid is a broad-spectrum neonicotinoid insecticide used in agriculture to control aphids. While recent studies have documented resistance to acetamiprid in several aphid species, the underlying mechanisms are still not fully understood. In this study, we analyzed the transcriptome and metatranscriptome of a laboratory strain of the pea aphid, Acyrthosiphon pisum (Harris, 1776), with reduced susceptibility to acetamiprid after nine generations of exposure to identify candidate genes and the microbiome involved in the adaptation process. Sequencing of the transcriptome of both selected (RS) and non-selected (SS) strains allowed the identification of 14858 genes and 4938 new transcripts. Most of the differentially expressed genes were associated with catalytic activities and metabolic pathways involving carbon and fatty acids. Specifically, the alcohol-forming fatty acyl-CoA reductase (FAR) and acyl-CoA synthetase (ACSF2), both involved in the synthesis of epidermal wax layer components, were significantly upregulated in RS, suggesting that adaptation to acetamiprid involves the synthesis of a thicker protective layer. Metatranscriptomic analysis revealed subtle shifts in the microbiome of RS. These results contribute to a deeper understanding of acetamiprid adaptation by the pea aphid and provide new insights for aphid control strategies.
Keywords:
omics
; insecticide resistance
; symbiotic bacteria
; gene expression analysis
; insect bioassays
1. Introduction
Aphids, small sap-sucking insects of the order Hemiptera, are notorious agricultural pests known for their ability to cause significant damage to crops through feeding activities and virus transmission [1,2]. As pests of economic importance, aphids pose formidable challenges to farmers worldwide, necessitating effective management strategies to mitigate their impact on crop health and yield. Acetamiprid is a neonicotinoid insecticide widely used in aphid management due to its systemic activity and broad spectrum of activity [3]. However, concerns have been raised regarding the overreliance on acetamiprid and its potential for inducing resistance in target pests [4].
At the center of the discussion on aphid control is the phenomenon of insecticide resistance, a widespread problem that has become a major obstacle in pest control [5,6]. Insecticide resistance arises from a complex interplay of genetic, physiological, and environmental factors, including the rapid evolution of resistance mechanisms within aphid populations. Resistance mechanisms include insensitivity of the target site, metabolic detoxification, and behavioral adaptations, which together reduce the efficacy of chemical insecticides [7,8]. Recent studies have documented resistance to acetamiprid in several aphid species [9,10,11,12,13]. For example, in one study, acetamiprid resistance in the melon aphid Aphis gossypii (Glover, 1877) was linked to overexpression of several P450 genes and suppressing their expression through RNA interference (RNAi) dramatically increased sensitivity to acetamiprid [13]. However, in a laboratory strain of Myzus persicae (Sulzer, 1776), resistance to acetamiprid was not accompanied by increased activity of P450 enzymes or any other known resistance mechanism [14], suggesting that different species may develop resistance to the same insecticide through divergent mechanisms.
In recent years, research has increasingly focused on the role of symbiotic bacteria in shaping aphid physiology and mediating responses to insecticides [15,16]. Symbiotic bacteria play a pivotal role in the growth and development of aphids [17]. The ubiquitous presence of the primary symbiont Buchnera aphidicola in aphids is ensured through vertical transmission via ovaries, maintaining its continuity from one generation to the next. Buchnera supplies essential amino acids and vitamin B, crucial nutrients often limited in plant phloem [18,19,20]. Another prevalent symbiont, Wolbachia, affects host reproduction to promote its proliferation and transmission [21,22,23,24]. Additionally, aphids can harbor secondary or facultative symbionts [25,26], transmitted both vertically and horizontally within and between species. Rickettsiella, a facultative endosymbiont, induces a change in aphid body color from red to green [27], potentially providing protection from natural enemies. Arsenophonus nasoniae influences the sex ratio of parasitic wasp offspring by selectively killing male embryos [28,29]. Hamiltonella strains, efficiently transmitted to aphid offspring, confer robust protection against dominant parasitoid species [30]. Li et al. observed that infection with H. defensa decreased the sensitivity of aphids to insecticides, particularly at low concentrations [31].
The pea aphid Acyrthosiphon pisum (Harris, 1776) is known worldwide as an important pest of legumes and pulses [32,33], where it causes significant economic losses. Despite the economic impact, there are currently no effective non-chemical methods to control this pest, so farmers rely heavily on chemical insecticides for control. Although resistance of the pea aphid to insecticides has not been extensively documented, a recent study found a high level of phenotypic resistance to several pyrethroids in field-collected strains compared to a susceptible reference strain [34]. Biochemical and molecular analyzes indicated the involvement of P450 and esterases in resistance: Increased P450 and esterase activities were observed in the resistant strains, and transcriptome profiling identified the P450 gene CYP6CY12 as highly overexpressed. These results highlight the occurrence of pyrethroid resistance in the pea aphid and the importance of understanding the molecular mechanisms underlying the development of resistance to other insecticides commonly used to control this species, including neonicotinoids.
In this study, we analyzed the transcriptome and metatranscriptome of pea aphids selected with acetamiprid for nine consecutive generations. Our aim was to investigate the constitutive changes in gene expression and microbiome of the acetamiprid-selected strain compared to the non-selected strain to identify possible target mechanisms involved in adaptation. Quantitative real-time PCR (qRT-PCR) was used to further validate the differentially expressed genes. Our results suggest a complex, multifaceted metabolic response to acetamiprid in the RS strain of pea aphid, with increased expression of enzymes involved in the synthesis of the epidermal wax layer components playing a central role in survival mechanisms. However, our analysis of the microbiome of RS suggests the involvement of two secondary symbionts in the adaptation process.
2. Materials and Methods
2.1. Aphids Rearing, Acetamiprid Exposure and Toxicity Assays
Aphids were reared according to the method described by Chang et al. [35]. A single apterous viviparous parthenogenetic A. pisum female was reared on broad bean (Vicia faba L.) seedlings in an incubator with a 16:8-hour light-dark cycle, 20°C temperature, and 60% relative humidity (RH).
For acetamiprid exposure, third-instar aphid nymphs were placed on leaves previously dipped in a 6.25 μg/ml acetamiprid solution in 0.01% dimethyl sulfoxide (DMSO) for 15~20 s, as described in Li et al. [31]. Surviving aphids were transferred to fresh leaves after 72 h. This procedure was replicated in three independent experiments with 10 individuals each. The acetamiprid exposure continued for nine consecutive generations to establish an acetamiprid-selected strain (RS). Aphids on leaves dipped in 0.01% DMSO served as negative control and were designated as non-selected strain (SS).
Toxicity bioassays were performed as per standard leaf dip toxicity bioassay, with minor modifications [36]. Briefly, serial dilutions of an acetamiprid stock in 0.01% DMSO were prepared with 0.1% Triton X-100 in water. Medium-sized broad bean leaves were dipped into acetamiprid dilutions (12.5 μg/ml, 6.25 μg/ml, 3.12 μg/ml, 1.56 μg/ml, 0.78 μg/ml) for 30 s each and then laid flat on a non-absorbent plastic to air dry for one hour. Control leaves were treated with 0.01% DMSO in 0.1% Triton X-100 alone. Thirty pea aphids were exposed to each concentration. Assay plates were incubated with a 16:8-hour light-dark cycle, 20°C temperature, and 60% RH, and mortality was recorded after three days (i.e., dead aphid failed to respond after gentle prodding). The bioassays were repeated three times.
2.2. Transcriptomic Sequencing and Analysis
Total RNA of the 3rd-instar nymphs from RS and SS was extracted using the TRIzol (Thermo Fisher, Waltham, MA) method. Three independent experiments were conducted, and each sample had 30 individuals. Transcriptome libraries were constructed according to the method described by Wu et al. [37]. Raw reads were processed to remove 3’-adaptors and repeating reads. Clean reads underwent de novo assembly using the Trinity (version 2.0.6), TGICLL (version 2.1), and Phrap (Release 23.0) programs. The library was sequenced using the DNBSEQ (PE150, BGI, Beijing, China) according to the manufacturer’s instructions.
Clean reads were aligned to the NCBI non-redundant (NR) protein database, Swiss-Prot, Kyoto Encyclopedia of Genesa and Genomes (KEGG), and Cluster of Orthologous Group (COG) databases using Blastx (E-value ≤ 1E-05). Unigene sequences were aligned to protein databases (NR, Swiss-Prot, KEGG, and COG). Blast2GO (ver. 2.5.0) was used for gene ontology (GO) annotation of unigenes with the NR database [38]. WEGO software [39] was then used to perform functional classification of the GO terms for all unigenes. Pathway assignments followed the KEGG database. Unigene expression calculations used the FPKM (RPKM) method [40,41].
2.3. Metatranscriptomic Sequencing and Analysis
The total RNA of the 3rd-instar nymphs from RS and SS was extracted using the TRIzol method. Three independent experiments were conducted, and each sample had 30 individuals. After rRNA removing, fragments underwent end-repair and subsequent 3’ adenylation, followed by ligation of adaptors to the 3’ adenylated ends. The clean reads were obtained using SOAPnuke (version 1.5.0), Bowtie2 (version 2.2.5), and Samtools (version 1.2). High-quality reads were de novo assembled using MEGAHIT software [42]. The qualified library was sequenced using DNBSEQ (PE100, BGI, Beijing, China) according to the manufacturer’s instructions. The raw data from sequencing were used for subsequent bioinformatics analysis.
To generate annotation information, the protein sequences of genes were aligned against KEGG, COG, and Swiss-Prot databases using DIAMOND (E-value ≤ 1E-05) [43]. In contrast to the transcriptome, the taxonomic annotation relied on the Kraken LCA algorithm [44]. Based on the abundance profiles of species, the features (Genera, Phyla, and KOs) with significantly differential abundances between groups were determined using ANOVA. Differentially enriched KEGG pathways were identified [45,46].
2.4. Quantitative Real-Time PCR (qRT-PCR)
Total RNA of the 3rd-instar nymphs from RS and SS was extracted with the TRIzol method. The first strand of cDNA was synthesized with 2 μg total RNA by PrimeScript Ⅱ 1st Strand cDNA Synthesis Kit (TaKaRa, Dalian, China) using oligo d(T)15. qRT-PCR was performed according to the method described by Chang et al. [35]. The PCR program consisted of 40 cycles of denaturation at 95°C for 30s, annealing at 55°C for 30s, and extension at 72°C for 30s. The primers used are listed in Table 1. The 2-∆∆Ct method was used to calculate the relative expression of mRNAs from the Cts obtained in the PCR quantification (Ct is the cycle threshold, which indicates the number of cycles experienced when the fluorescent signal in each reaction tube reaches a set threshold). ΔCt represents the average Ct value of the sample minus the internal control. ΔCtΔCt represents the average Ct value of sample minus the control sample. Three independent experiments were conducted, and each sample was repeated three times. Three independent experiments were conducted (with 30 individuals for each), and each sample was repeated three times.
2.5. Data Analysis
The data were processed using SPSS Statistics 22.0 software. One-way analysis of variance (ANOVA) in conjunction with Tukey’s post-test and Student's t test, both at P < 0.05 as the significance level, were performed to determine differences between treatments. Three replicates were performed for each treatment and similar results were obtained. The standard error of the means was used to compare replicates.
Bioassays were analyzed using the open-source R environment [47]. Estimates of LC50, LC99 and LC10, 95% fiducial limits, and slopes were calculated by maximum likelihood logit regression analysis in a generalized linear model from individually fitted analyses of deviance as previously described [48]. Pairwise comparisons of LC50 values were significant (P < 0.05) when their respective 95 % fiducial limits did not overlap [48].
3. Results
3.1. Acetamiprid-Resistant Aphids
Insect bioassays showed that after nine consecutive generations of exposure to the insecticide, RS exhibited a significant 1.9-fold reduction in susceptibility to acetamiprid compared to SS (Table 2). Although RS displayed notably higher LC50 and LC99 values than SS, there were no apparent differences in LC5 values between the two strains (P > 0.05; overlapped 95% fiducial limits). This indicates similar susceptibility at lower, sublethal acetamiprid concentrations.
3.1. Transcriptomic Profiling Associated with Acetamiprid Resistance
RNA libraries were constructed and sequenced for RS and SS. This generated 45.57 and 46.16 million raw reads, respectively. After removal of low-quality reads and adaptor sequences, 43.13 and 42.42 million clean reads were obtained from RS and SS, respectively (Table S1). The analysis identified 14,858 expressed genes and 4938 new transcripts.
Expression analysis revealed 581 upregulated and 1220 downregulated genes in RS (Figure 1A). Among them, 15 genes showed significant upregulation, while 92 genes displayed significant downregulation in RS (P < 0.01) (Figure 1B). Upregulated genes included V-type H+-transporting ATPase subunit B (ATPeV1B), alanyl aminopeptidase (ANPEP), alcohol-forming fatty acyl-CoA reductase (FAR), while downregulated genes included alpha-N-acetylgalactosaminidase (NAGA), SRA stem-loop-interacting RNA-binding protein, mitochondrial (SLIRP), and polyhomeotic-like protein 1 (PHC1), among others (Table 3).
qRT-PCR experiments were performed to further validate the differentially expressed genes (Figure 1C). In general, the qRT-PCR results were consistent with the RNA-Seq results, except for ANPEP, which was significantly downregulated. The reason for this observation could be that the specific down-regulation of one ANPEP gene triggers a compensatory mechanism that induces the expression of another of the numerous ANPEP genes present in the pea aphid genome [49]. Genes that were significantly upregulated in RS (P < 0.01) include FAR and ACSF2. FAR catalyzes the formation of fatty alcohols from fatty acids and is a key enzyme involved in the synthesis of the epidermal wax layer. ACSF2 catalyzes the formation of fatty acyl-CoA from fatty acids and thus promotes the utilization of these fatty acids.
Gene Ontology (GO) analysis revealed that the majority of the differentially expressed genes (DEGs), totaling 628 genes (Figure 2A), were primarily associated with catalytic activity. These genes were mainly implicated in cofactor biosynthesis and the metabolism of carbon and fatty acids. Protein network analysis of these 628 genes identified three key nodes: triosephosphate isomerase (TPI), glucose-6-phosphate isomerase (PHI), and delta-1-pyrroline-5-carboxylate synthetase (P5CS). Notably, the expression of these key nodes decreased in the RS strain (Figure 2B). TPI and PHI serve as key enzymes in glycolysis, while P5CS plays a critical role in proline metabolism, suggesting metabolic reprogramming to cope with stress in the RS strain.
The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database assigned the differentially expressed genes to cofactor biosynthesis, carbon and fatty acid metabolism, and the ribosome pathways (Figure 2C). The expression of most genes assigned to metabolic pathways decreased in the RS strain (Figure 2D,E). In the ribosome pathway, however, gene expression increased in the RS strain, with the sole exception of the gene encoding the large subunit of ribosomal protein LP1 (RPLP1) (Figure 2F). GO and KEGG analysis showed that most DEGs in RS were concentrated in metabolic pathways and experienced partial repression. However, the induction of genes in the ribosome pathway suggests the activation of a potential adaptive response to the stress associated with acetamiprid exposure.
3.1. Metatranscriptomic Profiling Associated with Acetamiprid Resistance
Metatranscriptomic sequencing was used to analyze the differences in bacterial abundance between RS and SS. A substantial dataset was generated, yielding 240.40 and 240.39 million clean reads, with 24.04 billion and 24.01 billion clean bases from RS and SS, respectively (Table S2). Species annotation revealed a discrete increment in the abundance in four species, i.e. B. aphidicola, Herbasepirillum huttiense, Deelftia acidovorans, and Lactobacillus iners. Conversely, three species -Serratia symbiotica, Escherichia coli and Acinetobacter soli- exhibited decreased abundance (Figure 3A and Table 4).
Expression analysis unveiled 24 significantly upregulated genes and 29 significantly downregulated genes (P < 0.05) (Figure 3B). GO annotations indicated that most downregulated genes were related to actin filament organization, regulation of actin filament organization, and cell-substrate adhesion (Figure 3C). The actin cytoskeleton, an intracellular structure involved in the onset and control of cell shape and function, was found to regulate the ion channel activity (Figure 3D).
The sequencing results were further validated by qRT-PCR. The results showed significant (P < 0.05) downregulation of genes encoding MAP7, LIM domain protein (LIM3) and PRELI domain-containing protein 2 (PRELI) in RS (Figure 3E). MAP7 corresponds to the gene encoding the ensconsin. LIM3 encodes an RNA polymerase II transcription factor with a key role in neuron specification. PRELI, a protein-coding gene, may be involved in phosphatidic acid transfer activity and phospholipid transport and is located in the mitochondrial intermembrane space. The protection against acetamiprid may be attained through the downregulation of these genes.
4. Discussion
In the present study, we have obtained a strain RS of pea aphid that exhibits reduced susceptibility to acetamiprid and characterized it by transcriptomic and metatranscriptomic approaches. Most intriguingly, the RS strain, despite being exposed to acetamiprid for nine consecutive generations, exhibited only a twofold increase in LC50 compared to the unselected SS strain. In fact, documented cases of insecticide resistance in pea aphids are rare. Only one case has been reported so far, against pyrethroids [34,50]. This rarity suggests that resistance to most insecticides is low in this species. Furthermore, we found that key players in cellular metabolism and stress response, such as TPI, PHI and P5CS, were downregulated in the RS strain. This suggests that the insect has adopted strategies, such as metabolic reprogramming, that prioritize survival over growth to mitigate the negative effects of acetamiprid. While this adaptive response may improve short-term survival, it may impose a long-term fitness cost on the insect as resources are diverted from other essential biological functions, such as growth. Indeed, we observed differences in the growth cycle between the RS and SS strains: the RS strain required an additional 24 hours to reach the adult stage (data not shown), indicating a possible fitness cost in the absence of acetamiprid.
Both transcriptomic sequencing and qRT-PCR experiments have revealed the upregulation of ACSF2 and FAR, two important enzymes involved in lipid metabolism, in the RS strain. ACSF2 facilitates the conversion of fatty acids into fatty acyl-CoA, a crucial step in fatty acid metabolism. Meanwhile, FAR plays a key role in the conversion of fatty acyl-CoA into fatty alcohols, which are essential for the synthesis of cuticular hydrocarbons (CHCs). CHCs form the protective wax layer found on the surface of many aphids and mealybugs, shielding them from natural enemies and adverse environmental conditions [51,52]. The pea aphid is known to have a wax layer covering all parts of its body [53]. The constitutive upregulation of ACSF2 and FAR in RS due to prolonged exposure to acetamiprid may have increased the production of CHCs and, hence, the wax layer, ultimately reducing acetamiprid penetration. This explanation needs further investigation.
RNAi-mediated suppression of FAR expression in the cotton mealybug, Phenacoccus solenopsis Tinsley, resulted in reduced CHC levels in the wax layer [54]. Mealybugs with reduced CHC content exhibited increased mortality when exposed to desiccation and deltamethrin treatments, highlighting the importance of this enzyme for insect adaptation to water loss and insecticide stress. Therefore, we hypothesize that the potential mechanism responsible for reduced susceptibility to acetamiprid in the RS strain involves the thickening of the physical barrier through the wax layer that prevents the penetration of acetamiprid molecules into the aphid's body. This process may involve alterations in wax composition and layer deposition. However, while an increased wax layer may confer resistance to insecticides, it may also impose fitness costs on aphids, as the production of a thicker or more abundant wax layer requires resources and energy that could otherwise be allocated to essential biological processes such as growth.
Interestingly, genes encoding ribosomal proteins showed significant upregulation in RS transcriptome over SS transcriptome. While ribosomal proteins are primarily known for their involvement in protein synthesis within ribosomes, they have been found to have additional functions in various cellular processes. These proteins may interact with other molecules, such as proteins or nucleic acids, to exert these non-canonical functions [55]. Previously, Yu et al [56] linked the ribosomal protein S29 to deltamethrin resistance by binding to CYP6N3 -a member of the CYP6 class of cytochrome P450 enzymes involved in metabolic resistance to pyrethroids- and stimulating its degradation by the 26S proteasome. Overexpression of RPS29 reduced cell viability in the presence of deltamethrin. In addition, another study identified the ribosomal protein RpS2 as a potential receptor for the insecticidal protein Vip3Aa from Bacillus thuringiensis [57]. RNAi-mediated silencing of RpS2 gene expression in both transfected Sf21 cells and in larvae of Spodoptera litura (Fabricius, 1775) injected with double-stranded RNA resulted in reduced toxicity of the Vip3A protein. Further evidence for the specific upregulation of ribosomal proteins in response to a selective agent was provided by HT-SuperSAGE analysis of a Vip3Aa-selected population of Heliothis virescens (Fabricius, 1777), which confirmed this phenomenon [58]. Whether the constitutive overexpression of ribosomal proteins in acetamiprid-selected pea aphids responds to an increased demand for protein synthesis in response to stress or is involved in a specific adaptive mechanism remains to be elucidated.
In our study, we detected two secondary symbionts of pea aphids, namely H. huttiense and D. acidovorans, exclusively in RS, while L. iners was more prevalent in RS compared to SS. The precise impact of these secondary symbionts on the adaptive response of RS to acetamiprid remains uncertain. Some research suggests that symbiotic bacteria actively participate in detoxifying insecticides. For instance, Serratia oryzae has been implicated in insecticide resistance in Aedes albopictus (Skuse, 1894), contributing to resistance development by upregulating the expression and activity of metabolic detoxification enzymes in mosquitoes [59]. In A. gossypii, the composition of symbiotic bacteria undergoes significant changes after insecticide treatment [60]. Notably, antibiotic treatment has also been found to increase the sensitivity of A. gossypii to spirochetes [61]. However, the interaction between aphids and symbionts is intricate. While aphids may gain benefits from hosting symbionts, negative effects could lead to the reallocation of aphid energy resources [62]. For instance, infection of the corn leaf aphid, Rhopalosiphum maidis (Fitch, 1856), with H. defensa and Regiella insecticola could have a partially negative effect on aphid growth and development, despite endosymbionts were maintained in aphids over time [63]. Previously, these two symbionts were shown to protect aphids, with R. insecticola shielding A. pisum from the aphid-specific fungal entomopathogen Zoophthora occidentalis [64], and H. defensa reducing aphid susceptibility to insecticides [31].
In summary, we have shown that acetamiprid-selected pea aphid RS strain exhibits changes in gene expression, the most interesting of which was the constitutive overexpression of genes related to the synthesis of the components of the epidermal wax layer. Overall, the development of an increased waxy layer represents a mechanism by which the pea aphid can mitigate the effects of acetamiprid. This emphasizes the importance of understanding wax biosynthesis and its role in acetamiprid resistance for effective pest management strategies. We have also shown the differential infection of the acetamiprid-selected RS strain with two specific secondary symbionts. Understanding the trade-offs that aphids must make to survive acetamiprid exposure while harboring these secondary symbionts is crucial for developing sustainable pest management strategies that consider both immediate efficacy and long-term ecological impact.
Supplementary Materials
Table S1: Summary of transcriptomic sequencing data; Table S2: Summary of metatranscriptomic sequencing data.
Author Contributions
Conceptualization, D.L. and Y.W.; methodology, Z.C.; software, S.G.; validation, Z.C., X.Z. and Y.Q.; formal analysis, Z.C.; investigation, Z.C.; resources, Y.K.; data curation, Z.C. and C.A.-P; writing—original draft preparation, Z.C.; writing—review and editing, C.A. -P and D.L.; visualization, Z.C. and K.Z.; supervision, D.L., Y.W and C.A. -P; project administration, D.L., Y.W. and C.A. -P; funding acquisition, D.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work and the APC were funded by the National Natural Science Foundation of China (No. 31970480), the Natural Science Foundation of Henan province (No. 212300410063) and the Key Research Project of Henan Province (No. 231111111000).
Data Availability Statement
The Raw transcriptomic and metatranscriptomic sequencing data presented in this study are openly available in NCBI with SRA number SRR26347092 and SRR25998188, respectively.
Acknowledgments
We thank Dr. Feng Cui from the Institute of Zoology, Chinese Academy of Sciences for kindly providing the parthenogenetic pea aphids of Acyrthosiphon pisum.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analysis, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Huang, X.L.; Qiao, G.X. Research status and trends in Aphidology. Acta Entomol. Sin. 2006, 1017–1026. [Google Scholar] [CrossRef]
- Guerrieria, E.; Digilio, M.C. Aphid-plant interactions: a review. J. Plant Inter. 2008, 3(4), 223–232. [Google Scholar] [CrossRef]
- Phogat, A.; Singh, J.; Kumar, V.; Malik, V. Toxicity of the acetamiprid insecticide for mammals: a review. Environ. Chem. Lett. 2022, 20, 1453–1478. [Google Scholar] [CrossRef]
- Ijaz, M.; Afzal, M.B.S.; Shad, S.A. Resistance risk analysis to acetamiprid and other insecticides in Acetamiprid-Selected population of Phenacoccus solenopsis. Phytoparasitica 2016, 44, 177–186. [Google Scholar] [CrossRef]
- Simon, J.-C.; Peccoud, J. Rapid evolution of aphid pests in agricultural environments. Curr. Opin. Insect Sci. 2018, 26, 17–24. [Google Scholar] [CrossRef]
- Li, R.; Cheng, S.; Chen, Z.; Guo, T.; Liang, P.; Zhen, C.; Wang, J.; Zhang, L.; Liang, P.; Gao, X. Establishment of toxicity and susceptibility baseline of broflanilide for aphis Gossypii glove. Insects 2022, 13, 1033. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Nauen, R. The molecular mechanisms of insecticide resistance in aphid crop pests. Insect Biochem. Mol. Biol. 2023, 156, 103937. [Google Scholar] [CrossRef] [PubMed]
- Kaleem Ullah, R.M.; Gao, F.; Sikandar, A.; Wu, H. Insights into the effects of insecticides on aphids (Hemiptera: Aphididae): resistance mechanisms and molecular basis. Inter. J Mol. Sci. 2023, 24(7), 6750. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Denholm, I. Resistance of insect pests to neonicotinoid insecticides: Current status and future prospects. Arch. Insect Biochem. Physiol. 2005, 58, 200–215. [Google Scholar] [CrossRef]
- Hirata, K.; Kiyota, R.; Matsuura, A.; Toda, S.; Yamamoto, A.; Iwasa, T. Association between the R81T mutation in the nicotinic acetylcholine receptor β1 subunit of Aphis gossypii and the differential resistance to acetamiprid and imidacloprid. J. Pestic. Sci. 2015, 40, 25–31. [Google Scholar] [CrossRef]
- Shi, X.; Jiang, L.; Wang, H.; Qiao, K.; Wang, D.; Wang, K. Toxicities and sublethal effects of seven neonicotinoid insecticides on survival, growth and reproduction of imidacloprid-resistant cotton aphid, Aphis gossypii. Pest Manag. Sci. 2011, 67, 1528–1533. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Functional analysis of cytochrome P450 genes linked with acetamiprid resistance in melon aphid, Aphis gossypii. Pesticide Biochem. Physiol. 2020, 170, 104687. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Acetamiprid resistance and fitness costs of melon aphid, Aphis gossypii: An age-stage, two-sex life table study. Pesticide Biochem. Physiol. 2021, 171, 104729. [Google Scholar] [CrossRef]
- Berber, G.; Demirci, B.; Toprak, U.; Inak, E.; Yorulmaz, S. Acetamiprid resistance in the green peach aphid Myzus persicae (Sulzer) (Hemiptera: Aphididae): selection, cross-resistance, biochemical and molecular resistance mechanisms. JAFAG 2022, 39(3), 136–142. [Google Scholar] [CrossRef]
- Jaffar, S.; Ahmad, S.; Lu, Y. Contribution of insect gut microbiota and their associated enzymes in insect physiology and biodegradation of pesticides. Front. Microbiol. 2022, 13, 979383. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Khan, M.M.; Bamisile, B.S.; Hafeez, M.; Qasim, M.; Rasheed, M.T.; Rasheed, M.A.; Ahmad, S.; Shahid, M.I.; Xu, Y. Role of insect gut microbiota in pesticide degradation: a review. Front. Microbiol. 2022, 13, 870462. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Guo, J.Y.; Long, C.Y.; Liu, H.; Wan, F.H. Advances in endosymbionts and their functions in insects. Acta Entomol. Sin. 2014, 57, 111–122. [Google Scholar] [CrossRef]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: aphids and their symbiotic bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huang, X.L.; Qiao, G.X. Trends in research on the primary endosymbiont of aphids, Buchnera aphidicola. Chin. J. Appl. Entomol. 2013, 50, 1419–1427. [Google Scholar] [CrossRef]
- Tian, P.-P.; Zhang, Y.-L.; Huang, J.-L.; Li, W.-Y.; Liu, X.-D. Arsenophonus interacts with Buchnera to improve growth performance of aphids under amino acid stress. Microbiol. Spectr. 2023, 11, e01792–23. [Google Scholar] [CrossRef] [PubMed]
- Sl, D.; K, B.; Hr, B.; Bf, J.; W, Z.; F, R.; Sl, O. Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem. Mol. Biol. 1999, 29. [Google Scholar] [CrossRef]
- Gong, P.; Shen, Z.R.; Li, Z.H. Wolbachia endosymbionts and their manipulation of reproduction of arthropod hosts. Acta Entomol. Sin. 2002, 241–252. [Google Scholar] [CrossRef]
- Jiang, X.F.; Wang, L.; Zhang, L.; Luo, L.Z. Molecular detection of Wolbachia in three species of vegetable aphids collected from Beijing suburb. Plant Prot. 2009, 35, 63–65. [Google Scholar] [CrossRef]
- Bi, J.; Wang, Y.-F. The effect of the endosymbiont Wolbachia on the behavior of insect hosts. Insect Sci. 2020, 27, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Fukatsu, T.; Nikoh, N.; Kawai, R.; Koga, R. The secondary endosymbiotic bacterium of the pea aphid Acyrthosiphon pisum (Insecta: Homoptera). Appl. Environ. Microbiol. 2000, 66, 2748–2758. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.B.; Zhang, J.; Guo, J.Y.; Guo, J.Y.; Liu, H.; Wan, F.H. Advances of the secondary endosymbionts in sap-feeding insects. J. Biosaf 2016, 25, 92-98+122. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T.; Matsumoto, S.; Simon, J.-C.; Fukatsu, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef]
- Bressan, A.; Terlizzi, F.; Credi, R. Independent origins of vectored plant pathogenic bacteria from arthropod-associated Arsenophonus endosymbionts. Microb. Ecol. 2012, 63, 628–638. [Google Scholar] [CrossRef]
- Xu, C.; Wang, L.S.; Zhu, X.Z.; Wang, L.; Li, D.Y.; Zhang, K.X.; Ji, J.C.; Luo, J.Y. The relationship between symbiotic bacteria and wing dymorphism in Rhopalosiphum padi. Chin. J. Biol. Control. 2022, 38, 205–214. [Google Scholar] [CrossRef]
- Wu, T.; Monnin, D.; Lee, R.A.R.; Henry, L.M. Local adaptation to hosts and parasitoids shape Hamiltonella defensa genotypes across aphid species. Proc. R. Soc. B Biol. Sci. 2022, 289, 20221269. [Google Scholar] [CrossRef]
- Li, Q.; Sun, J.; Qin, Y.; Fan, J.; Zhang, Y.; Tan, X.; Hou, M.; Chen, J. Reduced insecticide susceptibility of the wheat aphid Sitobion miscanthi after infection by the secondary bacterial symbiont Hamiltonella defensa. Pest Manag. Sci. 2021, 77, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yuan, M.L. Biological and ecological characteristics of Acyrthosiphon pisum. Pratacultural Sci. 2017, 34, 1727–1740. [Google Scholar] [CrossRef]
- Sandhi, R.K.; Reddy, G.V.P. Biology, ecology, and management strategies for pea aphid (Hemiptera: Aphididae) in pulse crops. J. Integr. Pest Manag. 2020, 11(1), 18. [Google Scholar] [CrossRef]
- Müller, V.; Benjamin, B.; Lueke, B.; Mazzoni, E.; Pym, A.; Bass, C.; Nauen, R. Molecular characterization of pyrethroid resistance in field-collected populations of the pea aphid, Acyrthosiphon pisum. Entomol. Gen. 2023. [Google Scholar] [CrossRef]
- Chang, M.; Cheng, H.; Cai, Z.; Qian, Y.; Zhang, K.; Yang, L.; Ma, N.; Li, D. miR-92a-1-P5 modulated expression of the flightin gene regulates flight muscle formation and wing extension in the Pea Aphid, Acyrthosiphon pisum (Hemiptera: Aphidoidea). J. Insect Sci. 2022, 22, 14. [Google Scholar] [CrossRef] [PubMed]
- Moores, G.D.; Gao, X.; Denholm, I.; Devonshire, A.L. Characterisation of insensitive acetylcholinesterase in insecticide-resistant cotton aphids, Aphis gossypii Glover (Homoptera: Aphididae). Pestic. Biochem. Physiol. 1996, 56, 102–110. [Google Scholar] [CrossRef]
- Wu, B.; Huang, L.; Qiu, W.J.; Liu, X.; Shen, Y.W.; Liu, Y.P.; Yang, Z.L.; Li, X.M.; Cui, B.; Xu, S.D.; et al. Small nucleolar RNA Sf-15 regulates proliferation and apoptosis of Spodoptera frugiperda Sf9 cells. BMC Mol. Biol. 2019, 20. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef]
- Audic, S.; Claverie, J.-M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.R.; Nielsen, J. Uncovering Transcriptional Regulation of Metabolism by Using Metabolic Network Topology. Proc. Natl. Acad. Sci. USA 2005, 102, 2685–2689. [Google Scholar] [CrossRef] [PubMed]
- Matsouaka, R.A.; Singhal, A.B.; Betensky, R.A. An Optimal Wilcoxon–Mann–Whitney Test of Mortality and a Continuous Outcome. Stat. Methods Med. Res. 2018, 27, 2384–2400. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2024. Available online: http://www.R-project.org/ (accessed on 12 March 2024).
- Yang, L.; Sun, Y.; Chang, M.; Zhang, Y.; Qiao, H.; Huang, S.; Kan, Y.; Yao, L.; Li, D.; Ayra-Pardo, C. RNA interference-mediated knockdown of Bombyx mori haemocyte-specific cathepsin l (Cat l)-like cysteine protease gene increases Bacillus thuringiensis kurstaki toxicity and reproduction in insect cadavers. Toxins 2022, 14, 394. [Google Scholar] [CrossRef] [PubMed]
- Crawley, M.J. The R Book, John Wiley and Sons Ltd: Chichester, West Sussex, UK, 2007; pp 1-877.
- The International Aphid Genomics Consortium. Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 2010, 8(2), e1000313. [Google Scholar] [CrossRef]
- Arthropod Pesticide Resistance Database. Available online: https://www.pesticideresistance.org/ (accessed on 11 March 2024).
- Pope, R.D. Some aphid waxes, their form and function (Homoptera: Aphididae). J. Nat. Hist. 1983, 17, 489–506. [Google Scholar] [CrossRef]
- Ammar, E.D.; Alessandro, R.T.; Hall, D.G. Ultrastructural and chemical studies on waxy secretions and wax-producing structures on the integument of the woolly oak aphid Stegophylla brevirostris Quednau (Hemiptera: Aphididae). J. Microsc. Ultrastruct. 2013, 1, 43–50. [Google Scholar] [CrossRef]
- Kanturski, M.; Świątek, P.; Trela, J.; Borowiak-Sobkowiak, B.; Wieczorek, K. Micromorphology of the model species pea aphid Acyrthosiphon pisum (Hemiptera, Aphididae) with special emphasis on the sensilla structure. Eur. Zoo. J. 2020, 87(1), 336–356. [Google Scholar] [CrossRef]
- Tong, H.; Wang, Y.; Wang, S.; Omar, M.A.A.; Li, Z.; Li, Z.; Ding, S.; Ao, Y.; Wang, Y.; Li, F.; Jiang, M. Fatty acyl-CoA reductase influences wax biosynthesis in the cotton mealybug, Phenacoccus solenopsis Tinsley. Commun. Biol. 2022, 5(1), 1108. [Google Scholar] [CrossRef]
- Zhou, X.; Liao, W.J.; Liao, J.M.; Liao, P.; Lu, H. Ribosomal proteins: functions beyond the ribosome. J. Mol. Cell. Biol. 2015, 7(2), 92–104. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Ma, K.; Sun, L.; Hu, H.; Zou, F.; Guo, Q.; Lei, Z.; Zhou, D.; Sun, Y.; et al. Ribosomal protein S29 regulates metabolic insecticide resistance through binding and degradation of CYP6N3. PLOS ONE 2014, 9, e94611. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sachdev, B.; Sharma, N.; Seth, R.; Bhatnagar, R.K. Interaction of Bacillus thuringiensis vegetative insecticidal protein with ribosomal S2 protein triggers larvicidal activity in Spodoptera frugiperda. Appl. Environ. Microbiol. 2010, 76(21), 7202–7209. [Google Scholar] [CrossRef] [PubMed]
- Ayra-Pardo, C.; Ochagavía, M.E.; Raymond, B.; Gulzar, A.; Rodríguez-Cabrera, L.; Rodríguez de la Noval, C.; Morán Bertot, I.; Terauchi, R.; Yoshida, K.; Matsumura, H.; Téllez Rodríguez, P.; Hernández Hernández, D.; Borrás-Hidalgo, O.; Wright, D.J. HT-SuperSAGE of the gut tissue of a Vip3Aa-resistant Heliothis virescens (Lepidoptera: Noctuidae) strain provides insights into the basis of resistance. Insect Sci. 2019, 26(3), 479–498. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Guo, X.X.; Lou, Z.W.; Wang, H.Y.; Cheng, P. Correlation research between Serratia oryzae and insecticide-resistance of Aedes albopictus. J. Med. Pest Control 2023, 39, 205–210. [Google Scholar] [CrossRef]
- Zhang, S.; Luo, J.; Wang, L.; Zhang, L.; Zhu, X.; Jiang, W.; Cui, J. Bacterial communities in natural versus pesticide-treated Aphis gossypii populations in North China. MicrobiologyOpen 2019, 8, e00652. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, Y.; Zheng, C.; Gao, X.; Wei, X.; Xi, J.; Peng, T.; Shang, Q. Rapid evolution of symbiotic bacteria populations in spirotetramat-resistant aphis Gossypii glover revealed by pyrosequencing. Comp. Biochem. Physiol. Part D Genomics Proteomics 2016, 20, 151–158. [Google Scholar] [CrossRef]
- Zytynska, S.E.; Tighiouart, K.; Frago, E. Benefits and costs of hosting facultative symbionts in plant-sucking insects: A meta-analysis. Mol. Ecol. 2021, 30, 2483–2494. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Zhang, T.; Bai, S.; He, K.; Zhang, Y.; Francis, F.; Wang, Z. Secondary symbionts affect aphid fitness and the titer of primary symbiont. Front. Plant Sci. 2023, 14, 1096750. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.J.; Spragg, C.J.; Altincicek, B.; Gerardo, N.M. Symbiont-mediated protection against fungal pathogens in pea aphids: a role for pathogen specificity? Appl. Environ. Microbiol. 2013, 79, 2455–2458. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Analysis of differentially expressed genes (DEGs) between RS and SS aphid strains in the transcriptome. (A) Bar chart of the number of DEGs. Red represents the number of upregulated DEGs in RS, while blue represents the number of downregulated DEGs. (B) Volcano plot of significant DEGs. Dots represent individual genes. Blue dots represent significantly downregulated DEGs in RS, and red dots represent significantly upregulated DEGs. Grey dots indicate DEGs that are not significant between RS and SS. The data were analyzed with ANOVA (P < 0.05). (C) Quantitative real-time PCR (qRT-PCR) verification of ATP6V1, DLGAP1, FAR, and other genes after acetamiprid treatment. Each sample with different genes had three replicates. Significance analysis was conducted with ANOVA. Values are means ± SEM of three experiments. ** P < 0.01.
Figure 1.
Analysis of differentially expressed genes (DEGs) between RS and SS aphid strains in the transcriptome. (A) Bar chart of the number of DEGs. Red represents the number of upregulated DEGs in RS, while blue represents the number of downregulated DEGs. (B) Volcano plot of significant DEGs. Dots represent individual genes. Blue dots represent significantly downregulated DEGs in RS, and red dots represent significantly upregulated DEGs. Grey dots indicate DEGs that are not significant between RS and SS. The data were analyzed with ANOVA (P < 0.05). (C) Quantitative real-time PCR (qRT-PCR) verification of ATP6V1, DLGAP1, FAR, and other genes after acetamiprid treatment. Each sample with different genes had three replicates. Significance analysis was conducted with ANOVA. Values are means ± SEM of three experiments. ** P < 0.01.
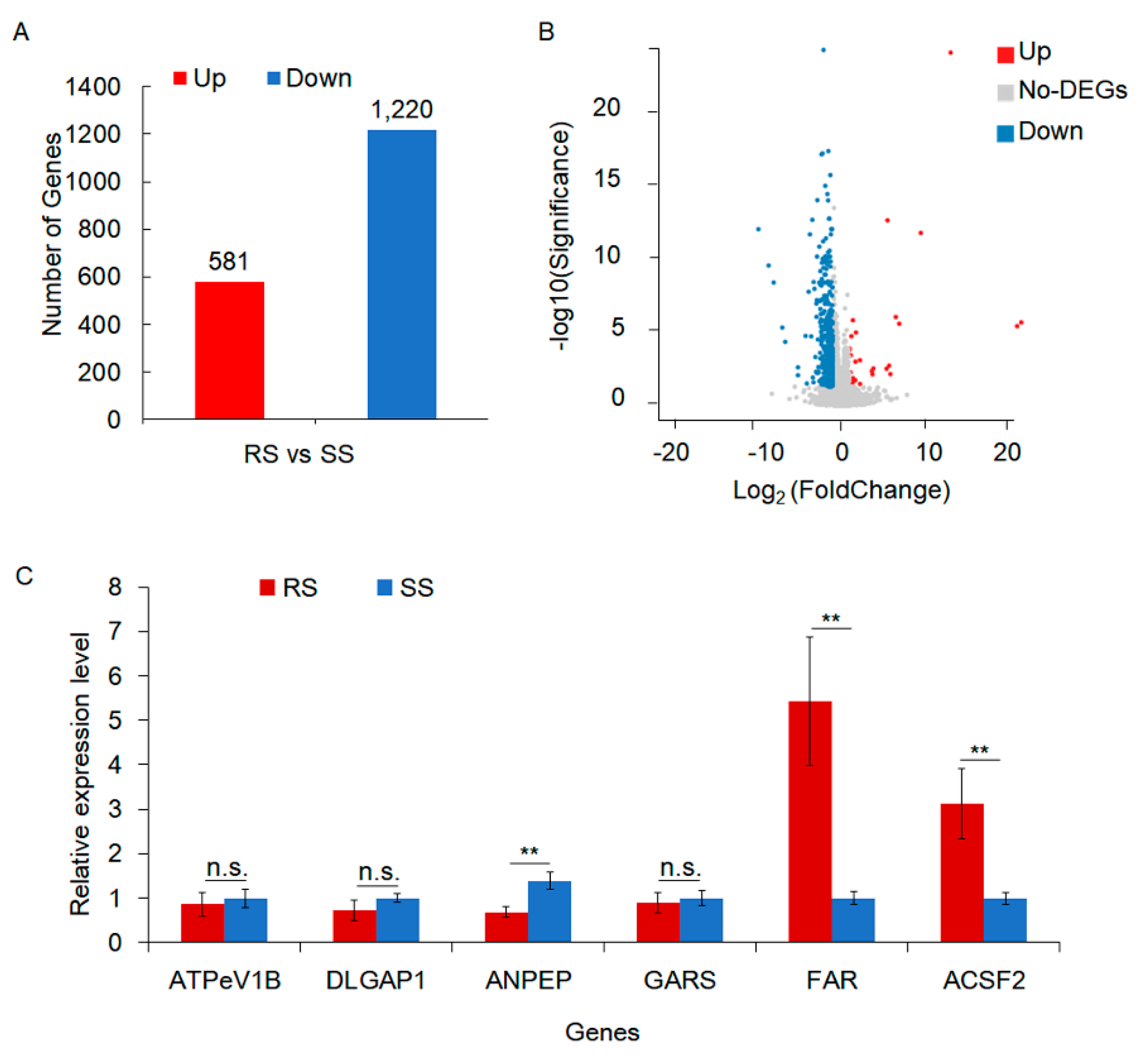
Figure 2.
Gene ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of differentially expressed genes (DEGs) between RS and SS strains in the transcriptome. (A) GO classification of DEGs, showing the number of DEGs in different categories. Most of the DEGs were genes related to the biosynthesis of cellular anatomical entities, catalytic activity, and metabolic processes. (B) 628 genes of catalytic activity network with three genes at key nodes, including triosephosphate isomerase (TPI), glucose-6-phosphate isomerase (PHI) and delta-1-pyrroline-5-carboxylate synthetase (P5CS). (C) KEGG enrichment of DEGs. (D) Heatmap analysis of hierarchical clustering of DEGs in fatty acid metabolism. (E) Heatmap analysis of hierarchical clustering of DEGs in carbon metabolism. (F) Heatmap analysis of hierarchical clustering of DEGs in the ribosome. Red and blue indicate high and low expression in RS, respectively.
Figure 2.
Gene ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of differentially expressed genes (DEGs) between RS and SS strains in the transcriptome. (A) GO classification of DEGs, showing the number of DEGs in different categories. Most of the DEGs were genes related to the biosynthesis of cellular anatomical entities, catalytic activity, and metabolic processes. (B) 628 genes of catalytic activity network with three genes at key nodes, including triosephosphate isomerase (TPI), glucose-6-phosphate isomerase (PHI) and delta-1-pyrroline-5-carboxylate synthetase (P5CS). (C) KEGG enrichment of DEGs. (D) Heatmap analysis of hierarchical clustering of DEGs in fatty acid metabolism. (E) Heatmap analysis of hierarchical clustering of DEGs in carbon metabolism. (F) Heatmap analysis of hierarchical clustering of DEGs in the ribosome. Red and blue indicate high and low expression in RS, respectively.
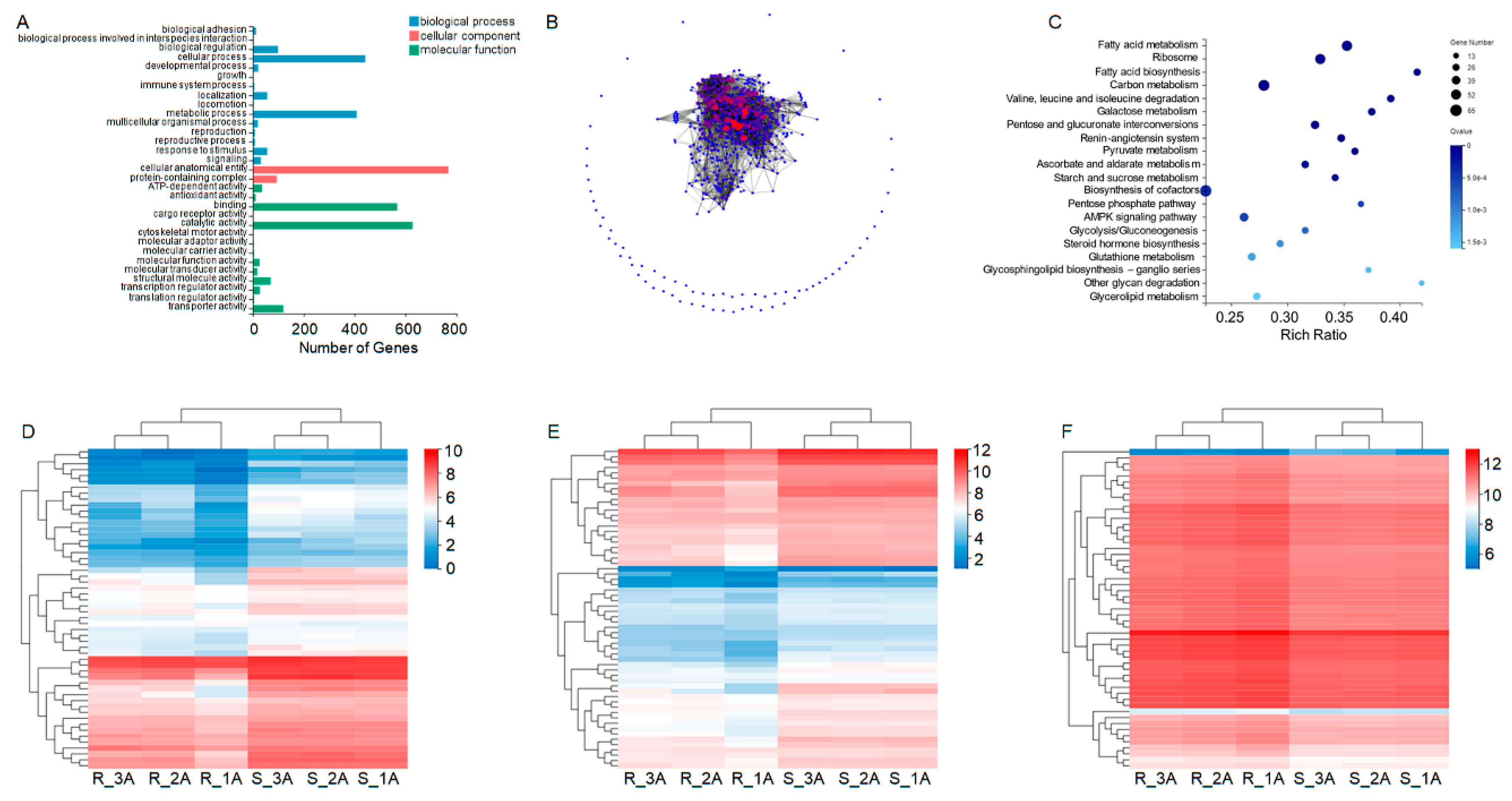
Figure 3.
Analysis of differentially expressed genes (DEGs) between RS and SS strains in the metatranscriptome. (A) Bar chart of the relative abundance of bacteria. Different colors represent different species, according to the legend on the right. The color filling of the intervals and lines can visually reflect the changes in the relative abundance of the species. (B) Volcano plot of significant DEGs in the metatranscriptome. Dots represent individual genes. Blue dots represent significantly downregulated DEGs in RS, and red dots represent significantly upregulated DEGs. Grey dots indicate DEGs that are not significant between RS and SS. The data were analyzed using ANOVA (P < 0.05). (C) Gene ontology (GO) classification of downregulated DEGs in the metatranscriptome. (D) EuKaryotic Ortholog Groups (KOG) of downregulated DEGs. The horizontal axis represents the gene ratio. (E) Quantitative real-time PCR (qRT-PCR) expression levels of TCF4/12, LIM3, MAP7 and PREL. qRT-PCR of different genes was replicated three times. Significance analysis was conducted with ANOVA. Values are means ± SEM of three experiments. * P < 0.05.
Figure 3.
Analysis of differentially expressed genes (DEGs) between RS and SS strains in the metatranscriptome. (A) Bar chart of the relative abundance of bacteria. Different colors represent different species, according to the legend on the right. The color filling of the intervals and lines can visually reflect the changes in the relative abundance of the species. (B) Volcano plot of significant DEGs in the metatranscriptome. Dots represent individual genes. Blue dots represent significantly downregulated DEGs in RS, and red dots represent significantly upregulated DEGs. Grey dots indicate DEGs that are not significant between RS and SS. The data were analyzed using ANOVA (P < 0.05). (C) Gene ontology (GO) classification of downregulated DEGs in the metatranscriptome. (D) EuKaryotic Ortholog Groups (KOG) of downregulated DEGs. The horizontal axis represents the gene ratio. (E) Quantitative real-time PCR (qRT-PCR) expression levels of TCF4/12, LIM3, MAP7 and PREL. qRT-PCR of different genes was replicated three times. Significance analysis was conducted with ANOVA. Values are means ± SEM of three experiments. * P < 0.05.

Table 1.
List of primers used in this study.
| Genes | Forward primer (5'-3') | Reverse primer (5'-3') |
|---|---|---|
| ATP6V1 (NM_001293544) | TCGTCAAATCTATCCACCAA | AATGCCTCTTCTCCCACAAC |
| DLGAP1 (XM_029490798) | AATTCCTCGGTTTATGTGAG | ATTGCCTTGCGTTGTTCTTC |
| ANPEP (XM_001950011) | TTGGATGGGCATTGTTTCTA | ATAGTCCATATCACCGACCT |
| GARS (XM_003245009) | TCATTGCCTCCATTAGTAGC | ATTTGTTCCATTGAATCCCT |
| FAR (XM_003242260) | ACTACGAGTCACCACCTTTG | TTTCTGCTTTCGCATACATT |
| ACSF2 (XM_016806965) | CGCCAACTCTACAAGACAAC | CATGACAAGATACCCACGAA |
| TCF4/12 (XM_016808432) | TCGCCCGATGATGATAGTGT | GTGCCGTCCAAGTAATAAGA |
| LIM3 (XM_008188468) | GAACGCAGAACAGTAAAGAA | CTGGTATAATAACGGAGGAA |
| MAP7 (XM_008190631) | AGAGTTGCGGTTGCAGTTGG | TGTTGCTCGGCAGATTCAGT |
| PRELI (XM_029488204) | GAAGAATGTTGGTATGACGA | CATGTTGGATTTGGTGTAAT |
Table 2.
Acetamiprid toxicity to non-selected SS and selected RS pea aphid strains expressed as LC50, LC99, and LC5. These values represent the concentration that causes death in 50%, 99% and 5% of the population, respectively, expressed in µg acetamiprid per ml (ppm). In all cases, the 95% fiducial limits are given in parentheses.
Table 2.
Acetamiprid toxicity to non-selected SS and selected RS pea aphid strains expressed as LC50, LC99, and LC5. These values represent the concentration that causes death in 50%, 99% and 5% of the population, respectively, expressed in µg acetamiprid per ml (ppm). In all cases, the 95% fiducial limits are given in parentheses.
| Aphid strain | LC50 | LC99 | LC5 | Slope (± S.E.) | na | RRb |
|---|---|---|---|---|---|---|
| SS | 2.68 (2.46-2.92) | 32.10 (31.34-32.87) | 0.55 (0.01-1.09) | 6.38 ± 0.37 | 150 | – |
| RS | 5.09 (4.88-5.31) | 44.75 (44.02-45.48) | 1.26 (0.81-1.72) | 8.28 ± 0.56 | 150 | 1.9 |
aNumber of larvae used in the bioassays, including control. bRR, resistance ratio, is the LC50 for RS strain divided by the LC50 for SS strain.
Table 3.
Top 10 downregulated and upregulated DEGs in the RS transcriptome.
| Gene ID | log2(R/S)* | Annotation |
|---|---|---|
| LOC115034459 | -10.09 | alpha-N-acetylgalactosaminidase |
| LOC100573163 | -8.84 | glycerol-3-phosphate dehydrogenase (NAD+) |
| LOC103308887 | -8.23 | SRA stem-loop-interacting RNA-binding protein, mitochondrial |
| LOC100168848 | -7.17 | NULL |
| LOC100572322 | -6.82 | polyhomeotic-like protein 1 |
| LOC100570759 | -5.26 | MFS transporter, PAT family, solute carrier family 33 (acetyl-CoA transportor), member 1 |
| LOC100570250 | -5.26 | lysophospholipid acyltransferase 7 |
| LOC100159801 | -4.33 | gamma-glutamyltranspeptidase / glutathione hydrolase / leukotriene-C4 hydrolase |
| LOC103310139 | -4.18 | leucine-rich repeat and immunoglobulin-like domain-containing nogo receptor-interacting protein |
| LOC100162019 | -3.96 | lactase-phlorizin hydrolase |
| LOC100169462 | 13.39 | V-type H+-transporting ATPase subunit B |
| LOC100575793 | 9.74 | glycyl-tRNA synthetase |
| LOC100159545 | 7.11 | discs, large-associated protein 1 |
| LOC107882136 | 6.69 | Alanyl aminopeptidase |
| LOC100569077 | 6.02 | NULL |
| LOC100571352 | 5.87 | lysosomal acid phosphatase |
| LOC100570391 | 5.67 | alcohol-forming fatty acyl-CoA reductase |
| LOC103310260 | 5.56 | medium-chain acyl-CoA ligase, mitochondrial |
| LOC100570903 | 3.95 | SWI/SNF related-matrix-associated actin-dependent regulator of chromatin subfamily C |
| LOC115034301 | 3.84 | glutamate receptor, ionotropic, invertebrate |
* log2(R/S) < 0: Downregulated DEG; log2(R/S) > 0: Upregulated DEG.
Table 4.
Abundance of bacteria.
| Sample | B. aphidicola (%) | S. symbiotica (%) | E. coli (%) | H. huttiense (%) | A. soli (%) | D. acidovorans (%) | L. iners (%) |
|---|---|---|---|---|---|---|---|
| RS_1 | 87.97 | 11.09 | 0.44 | 0.32 | 0.18 | 0.0 | 0.0 |
| RS_2 | 88.78 | 10.74 | 0.28 | 0.0 | 0.04 | 0.13 | 0.04 |
| RS_3 | 90.93 | 8.07 | 0.18 | 0.02 | 0.01 | 0.0 | 0.79 |
| SS_1 | 90.70 | 8.49 | 0.46 | 0.0 | 0.05 | 0.0 | 0.29 |
| SS_2 | 86.42 | 13.34 | 0.24 | 0.0 | 0.0 | 0.0 | 0.0 |
| SS_3 | 86.23 | 13.04 | 0.50 | 0.0 | 0.23 | 0.0 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



