
Submitted:
21 March 2024
Posted:
22 March 2024
You are already at the latest version
Abstract
Maize is an important staple food crop in sub-Saharan Africa (SSA). However, the average grain yield on farmers’ fields falls considerably short of the demand of the region’s population. In addition, the normal endosperm maize widely consumed in SSA is deficient in vitamin A. Drought, a consequence of climate change, is a major constraint to maize production, resulting in seasonal low grain yield in farmers’ fields. The objectives of this study were to assess the performance of newly developed extra-early maturing provitamin A hybrids under managed drought and well-watered conditions, compare the outcomes of multiple trait base-index and multi-trait genotype-ideotype distance index selection procedures, and identify drought-tolerant hybrids with stable performance across contrasting environments for commercialization in SSA. One hundred and ninety provitamin A hybrids and six checks were evaluated under managed drought and well-watered conditions at Ikenne for two seasons between 2021 and 2023. A 14*14-lattice design was used for the field evaluations under both research conditions. Drought stress was achieved by the complete withdrawal of irrigation water 25 days after planting. Results revealed significant differences among the hybrids under drought and well-watered conditions. Grain yield, ears per plant, and plant aspect under managed drought were correlated to the same traits under well-watered conditions suggesting that the expression of these traits is governed by common genetic factors. Twenty-nine hybrids were identified as top-performing drought-tolerant hybrids by the multiple trait base index and multiple-trait genotype–ideotype distance index. Of the selected outstanding 29 hybrids, 34% were derived from crosses involving the tester TZEEIOR 197, demonstrating the outstanding genetic potential of this inbred line. Further analysis of the 29 selected hybrids revealed TZEEIOR 509 × TZEEIOR 197 as the hybrid that combined the most drought-tolerant adaptive traits. However, the hybrids TZEEIOR 526 × TZEEIOR 97, TZEEIOR 384 × TZEEIOR 30, TZEEIOR 515 × TZEEIOR 249, TZEEIOR 510 × TZEEIOR 197, TZEEIOR 479 × TZEEIOR 197, and TZEEIOR 458 × TZEEIOR 197 were identified as the most stable hybrids across drought and well-watered conditions. These hybrids should be extensively tested in multi-location trials for deployment and commercialization in SSA.
Keywords:
drought tolerance
; multiple trait base index
; multiple-trait genotype-ideotype distance index
; stability
; sub-Saharan Africa
1. Introduction
Maize is a major crop for human consumption and as animal feed in Sub-Saharan Africa (SSA). The crop has the potential to combat the food insecurity and malnutrition challenges presently facing the region. It currently provides 30% of dietary calories for millions of people in SSA [1]. Maize is cultivated on about 42.5 million hectares in Africa [2]. Most of the varieties cultivated by farmers are deficient in essential micronutrients, particularly vitamin A [3,4] which cannot be synthesized by the human body. The essentiality of this nutrient is demonstrated by the vulnerability of people who depend largely on maize for food to several major diseases, including river blindness (onchocerciasis), anemia, diarrhea, measles, malaria, and respiratory infections [5]. Vitamin A deficiency (VAD) is also known to impair the functionality of the immune system, increase susceptibility to diseases, cause night or complete blindness and death from severe illnesses [6]. Given its importance as a major staple cereal crop in African homes, it is imperative to develop and commercialize improved maize genotypes biofortified with essential nutrients such as provitamin A and with tolerance/resistance to major production constraints faced by SSA farmers. These constraints include drought, low soil nitrogen (low N), Striga hermonthica parasitism, fall armyworm infestation, and many diseases [7,8]. On farmers’ fields, these constraints usually occur jointly with devastating effects on maize grain yield. This, coupled with the low genetic potentials of varieties cultivated by most farmers have resulted in the low grain yields frequently observed on farmers field.
In the absence of other stress factors during the production cycle, drought is capable of causing grain yield reduction between 40 and 90% depending on the stage of plant growth and development when the drought occurs, and its severity [9,10,11]. The flowering (tassel and silk emergence) and post-flowering (grain filling) stages are the major determinants of photo-assimilate partitioning from the source to the sink and consequently the amount of grain yields obtained at harvest. These stages are widely considered as the most sensitive stages of maize development which, if affected by drought, can lead to considerable reduction in maize production and productivity [12,13,14]. Therefore, to address the dietary needs of people of SSA and high maize productivity in the sub-region, it is of utmost importance to develop new and improved maize hybrids that combine enhanced levels of provitamin A and high levels of tolerance to drought for commercialization in SSA. The first step to the realization of drought-tolerant PVA maize began in 2007 by the International Institute of Tropical Agriculture Maize Improvement Program (IITA-MIP). The program had as its aim the development of extra-early and early (80-85 and 90-95 days to maturity, respectively) varieties that combine high levels of drought tolerance and provitamin A for SSA farmers. Through this program, numerous extra-early inbred lines possessing drought tolerance genes and elevated levels of PVA were developed, commercialized, and are presently serving as invaluable germplasm resource for developing drought-tolerant PVA hybrids in several national and international breeding programs[8]. In SSA, hybrid development and commercialization are the major focus of many breeding programs owing to the superior performance of hybrids over open pollinated varieties (OPVs). When new hybrids are developed, it is important to understand their responses to drought in multiple locations to identify tolerant hybrids for commercialization. To achieve this goal, a good selection strategy is required.
Plant breeders usually develop new genotypes with superior performance by combining several desirable traits in these new genotypes [15]. The main difficulties that frequently prevent breeders from selecting optimal genotypes using this approach are a definition of the economic value of such traits and converting these into realistic economic weights [16]. These have been the major drawbacks of the commonly used multiple trait base index (MBI) method. To overcome these limitations, a multivariate selection index, the Multi-Trait Genotype–Ideotype Distance Index (MGIDI) has been developed [15]. This index takes multi-collinearity into account and identifies all variables that should be taken into account to lead to significant genetic gains from selection [15]. This study was conducted to (i) assess the performance of newly developed multiple stress-tolerant extra-early provitamin A hybrids under managed drought and well-watered research conditions, (ii) compare the selection outcomes resulting from the use of MBI and MGIDI, and (iii) identify outstanding drought-tolerant and stable hybrids across drought stress and well-watered environments.
2. Materials and Methods
2.1. Genetic Material
In an effort by the IITA Maize Improvement Program (MIP) to develop multiple-stress tolerant and/or resistant, extra-early maturing cultivars with elevated levels of PVA for farmers in SSA, the extra-early Striga-resistant cultivar, 2004 TZEE-Y STR C4 was crossed to Syn–Y-STR-34–1-1–1-1–2-1-B-B-B-B-B/NC354/SYN-Y-STR-34–1-1–1 (OR1), a source of high PVA, in 2007. This was aimed at transferring the genes for high β-carotene into the extra-early cultivar. The F1 was backcrossed to the extra-early cultivar and kernels of the BC1 with deep orange color were selected and advanced to the F2 and F3 stages. At the F3 stage, the lines with intense orange color were selected and recombined to obtain extra-early PVA cultivar 2009 TZEE-OR1 STR from which a new set of extra-early inbred lines were extracted, starting in 2011. By 2014, 224 S6 inbred lines, selected for the deep orange color, had been developed from the variety. This set of PVA inbred lines were assessed for tolerance to induced drought stress at Ikenne, Nigeria, during the 2014 and 2015 minor rainy seasons. Thereafter, the PVA inbred lines were advanced to the S7–S8 stages from which the kernels were sampled and subjected to chemical analyses at the Food and Nutrition Laboratory of IITA, Ibadan for the determination of their PVA contents[8]. After the generation of the PVA inbred lines from the extra-early PVA cultivar 2009 TZEE-OR1 STR, new set of PVA inbred lines were generated from the biparental populations of TZdEEI 12 x TZdEEI 95 and TZdEEI 7 x TZdEEI 12. The F1 hybrids from these crosses were advanced through several cycles of repeated inbreeding and selection based on deep orange kernel color. At the S5 stage, selected lines were evaluated under combined heat and drought stress at Kadawa (Nigeria) in 2021 (Badu-Apraku Unpublished). Based on the performance, 27 inbred lines (11 from TZdEEI 12 x TZdEEI 95 and 16 from TZdEEI 7 x TZdEEI 12) were selected. These inbred lines plus nine other inbred lines extracted from 2009 TZEE-OR1 STR (making a total of 36 inbred lines) and five drought-tolerant inbred testers, are the genetic materials used for the development of the new PVA hybrids evaluated in the present study. The pedigree information of the inbred lines is presented in Table S1.
2.2. Generation of Crosses
The 36 extra-early PVA inbred lines were crossed to the five drought-tolerant PVA testers using the line by tester (L×T) design to generate 180 testcrosses at IITA-Ikenne breeding nursery in 2021. In addition, the testers were crossed in a Diallel fashion to generate 10 hybrids, which were added to the testcross hybrids for the study. Six extra-early normal yellow endosperm hybrids were included as checks to make 196 hybrids.
2.3. Field Evaluation
The 196 hybrids were evaluated for agronomic performance at Ikenne (7°52′ N, 30°44′ E, 61 m a.s.l., 1200 mm mean annual precipitation) under managed drought during the dry seasons (November to February) of 2021-2022 and 2022-2023 and under well-watered conditions during the rainy seasons (June to September) of 2022 and 2023. The managed drought experiment was achieved by supplying 17 mm of sprinkler irrigation water per week up to 28 days after planting. The plants were allowed to depend on the available soil moisture to reach physiological maturity. Plots consisted of single rows, each 3 m long, with inter and intra-row spacing of 0.75 and 0.40 m, respectively. Three seeds were planted per hole and the seedlings were thinned to two plants per hill two weeks after planting to give a final plant population density of 66,666 plants/ha. A 14 × 14 lattice design with two replications was used for each experiment. Compound fertilizer, NPK 15:15:15, was applied at planting at the rate of 60 kg/ha of N, P, and K for managed-drought conditions while a similar rate of application was applied at 2 weeks after planting (WAP) for well-watered conditions. Top-dressing was done using urea (46% N) at the rate of 30 kg/ha N at 3 WAP for managed-drought conditions and 5 WAP for well-watered conditions. Weed control was achieved with the application of an herbicide formulation containing gramoxone and primextra at the rate of 5 l/ha as pre-emergence while subsequent weed control was by hand weeding.
2.4. Data Collection
Observations were made on grain yield (GY), anthesis-silking interval (ASI), plant height (PHT), ear height (EHT), ear per plant (EPP), plant aspect (PASP), ear aspect (EASP), stay green characteristic (STGC),and husk cover (HCV). Anthesis-silking interval (ASI) was determined as the difference between days to anthesis and days to silking. Plant height (PHT) and ear height (EHT) were measured as the distance in centimeters between the base of the plant and the first tassel branch and the top ear, respectively. Ears per plant (EPP) was obtained by dividing the number of ears harvested by the number of plants at harvest. Plants aspect (PASP) was rated on a scale of 1–9, where 1 = excellent and 9 = poor. Ear aspect (EASP) was recorded on a scale of 1–9, where 1 = clean, uniform, large, and well-filled ears and 9 = ears with undesirable features, such as diseases, small ears, ears rot, and ears with poorly filled grains. Stay green characteristic (STGC) or leaf death score (LD) was determined under drought-stress conditions at 70 days after planting (DAP) on a scale of 1 to 9, where 1 = almost all leaves green and 9 = virtually all leaves dead. Husk cover (HCV) was rated on a scale of 1 to 9, where 1 = husks tightly arranged and extended beyond the ear tip and 9 = ear tips exposed. Harvested ears from the managed-drought trial were shelled and grain yield (GY) (kg ha-1) was determined using the shelled grain weight. Grain yield, adjusted to 15 % moisture, was computed from the shelled grain weight. The grain yield (kg ha-1) of the rain-fed experiment was computed based on the field weight, assuming a shelling percentage of 80 at 15% moisture.
2.5. Data Analysis
Combined analysis of variance (ANOVA) was conducted for each of the research conditions using the mixed linear model (MLM) (eqn. 1) implemented in lmerTest package in R software 4.3.1 [17]. In the MLM, a combination of year and research condition (managed drought or well-watered) was considered as an environment. Thus, four environments namely, WW_22 (well-watered 2022), WW_23 (well-watered 2023), DS_22 (managed drought 2022), and DS_23 (managed drought 2023) were involved. Environment, block, and replications were considered as random effects while genotype was considered as fixed effect. The model for the analysis of variance is expressed as:
Y= µ + Rep + Rep (Blk) + G + E + G × E + e ……………
Where;
Y= phenotype; µ= mean of the trait; G= genotype; E= environment; Rep= replication; Blk= block; Rep (Blk)= replication nested in block; G × E = genotype by environment interaction; e= residual
The best linear unbiased predictions (BLUPs) under both research conditions were estimated from the ANOVA. The estimates were used to compute Pearson phenotypic correlation between grain yield and other traits under each research condition and between research conditions using the Metan package [18].The MGIDI proposed by Olivoto and Nardino [15] was used to select maize hybrids that combine drought-tolerant adaptive traits and high yield. The MGIDI is based on four principles namely(i) traits rescaling to ensure they have a 0–100 range, (ii) accounting for correlation structure and dimensionality reduction in the data, (iii) using desired traits value to plan Ideotype, and (iv) computing the distance between the planned Ideotype and the genotype.
Traits rescaling was done using the equation below
Where;
ϕoj and ηoj are the original minimum and maximum values for the trait j, respectively; ϕnj and ηnj are the new minimum and maximum values for trait j after rescaling, respectively; and θij is the original value for jth trait of the ith genotype.
The values for and were chosen in a manner that for traits in which negative gains are desired, = 0 and = 100 was used while traits in which positive gains are desired, = 100 and = 0. In the final rescaled two-way table (), the column has a 0–100 range.
Following traits rescaling, explanatory factor analysis was carried out. The rescaled values were first used to group correlated traits into factors before the factorial scores were estimated for the traits using Equation 3 below.
Where:
X is a p x 1 vector of the observations that were rescaled; is a p x 1 vector of the standardized means; L is a p x f matrix of the factorial loadings; f is a p x 1 vector of the common factors; and e is a p x 1 vector of error.
For p and f which represent the number of traits and common factors retained, respectively, the correlation matrix of the rescaled values were used to generate the eigen values and eigen vectors. For the initial loadings, factors with eigen values higher than one were considered before the varimax rotation criteria was implemented for analytic rotation and estimation of final loadings according to (Kaiser) [19]. The final scores were obtained using Equation 4 below.
Where;
F is a g × f matrix with the factorial scores; Z is a g × p matrix with the rescaled means; A is a p × f matrix of canonical loading, and R is a p × p correlation matrix between the traits. The g, f, and p represent the number of genotypes, the factor kept, and the measured traits, respectively.
Ideotype planning was done such that the Ideotype has the highest rescaled value of 100 for all the analyzed traits. The MGIDI was finally estimated using equation 5.
Where;
MGIDI is the multi-trait genotype–ideotype distance index for the ith genotype; Yij is the score of the ith genotype in the jth factor being g and f; and Yj is the jth score of the ideotype.
From the above equation, the lower the MGIDI score of a genotype, the closer the genotype is to the ideotype. Thereafter, the proportion of the MGIDI index of the genotypes explained by the correlated factor is used to display the strengths and weaknesses of the genotypes based on Equation 6.
Where;
is the proportion of the MGIDI index of the ith genotypes explained by the correlated jth factor; D2ij is the distance between the ith genotype and the ideotype for the jth factor.
Data processing and the computation of the index were carried out using the Metan package in R [18].
To ascertain the effectiveness of MGIDI, the results obtained from the latter were compared to those from a multiple traits base-index (MBI) proposed by Badu-Apraku et al. [20] that incorporates grain yield, plant and ear aspects, stay green characteristic, number of ears per plant, plant and ear height, husk cover ,and anthesis-silking interval. The MBI has been used for the selection of hybrids that combine drought tolerance with outstanding yield performance based on Equation 7 below.
MBI = (2 × GY) + EPP + PHT + EHT – PASP – EASP – ASI – STGC – HCV
Where;
MBI = multiple traits base index, GY =grain yield, EPP = ear per plant, PASP = plant aspect, EASP = ear aspect, ASI= anthesis-silking interval, STGC = stay green characteristic, HCV = husk cover, PHT = plant height, EHT= ear height.
The traits used in the MBI were standardized to reduce the effects of different scales of the traits. A positive MBI value indicates tolerance to drought while a negative value indicates susceptibility to drought. Yield reduction attributable to drought stress was estimated using Equation 8.
Forty hybrids from the MGIDI selection comprising 20 best, 10 average, and 10 worst performers were selected for GGE biplot analysis using Metan. The “mean vs. stability” view of the GGE biplot was used to determine hybrids with stable and high grain yields across managed drought and well-watered research conditions. The data were neither transformed nor standardized and were environment-centered. The model for the GGE biplot is shown in Equation 9 below.
Yij −Yj = λ1εi1ηj1 + λ2εi2ηj2 + εij
Where;
Yij is the average yield of genotype i in environment j, Yj is the average yield across genotypes in environment j, λ1 and λ2 are the singular values for PC1 and PC2 respectively, εi1 and εi2 are the PC1 and PC2 scores for genotype i, ηj1 and ηj2 are the PC1 and PC2 scores for genotype j, and εij is the error associated with the genotype i in environment j.
3. Results
3.1. Analysis of Variance for Grain Yield and Other Traits under Managed Drought and Well-Watered Conditions
The ANOVA results across well-watered conditions presented in Table 1 revealed significant (p < 0.05) genotype mean squares for all traits except EASP. Environment mean square was significant (p < 0.05) for GY, ASI, PASP, and EPP. Under managed drought, a significant (p < 0.001) genotype mean square was observed for all traits. The environment mean square was significant (p< 0.05) for ASI and HCV, while genotype x environment mean square was significant for ASI, PHT, EPP, and STGC (Table 2).
3.2. Selection of Outstanding PVA Hybrids with Drought Tolerance Using the MGIDI Selection Method
The MGIDI partitioned the measured traits into three factors. The first factor (FA1) was correlated with GY, EPP, PASP, EASP, EPP, ASI, and STGC. The second factor (FA2) was associated with HCV while the third factor (FA3) was correlated with PHT and EHT. All the measured traits showed desirable predicted gains from selection observed in the sense of selection and perfect goal. Generally, the MGIDI provided a total predicted gain from selection of 11.96 % for traits for which higher positive expression was desirable and -16.65 % for traits for which higher negative expression was desirable(Table 3). Using a selection intensity of 15%, 29 hybrids comprising 28 new hybrids and one commercial check (TZEEI 79 × TZEEI 82) were selected as drought-tolerant hybrids (Figure 1 and Table S2). Of the selected testcross hybrids, 10 hybrids (34%) were developed from crosses involving the tester TZEEIOR 197. The best five hybrids based on the MGIDI were TZEEIOR 510 × TZEEIOR 97, TZEEIOR 510 × TZEEIOR 249, TZEEIOR 321 × TZEEIOR 197, TZEEIOR 384 × TZEEIOR 30, and TZEEIOR 458 × TZEEIOR 197. Similarly, the worst five hybrids were TZEEIOR 523 × TZEEIOR 9A, TZEEIOR 221 × TZEEIOR 9A, TZEEIOR 24 × TZEEIOR 30, TZEEIOR 41 × TZEEIOR 30, and TZEEIOR 45 × TZEEIOR 30 (Table S2).
The result of the analysis to unmask the strengths and weaknesses of the selected extra-early maize hybrids is presented in Figure 2. In this figure, the proportion of each factor to the MGIDI index accounts for the strengths and weaknesses. The red, green, and blue lines represented the three factors, and the factor protruding the most to a genotype is indicated as the strength of the genotype. Based on this, entry 121 had strength for FA1 which is correlated to GY, EASP, PASP, EPP, and STGC. Entries 135, 86, 81, 70, 57, 47, and 164 had strength for FA2 which is correlated to PHT and EHT. Entries 22, 192, 19, 189, 173, 165, 159, 149, 144, 142, 120, 119, 118, 114, 110, 109, 106, 98, 93, 68, and 52 showed strength for FA3 which is correlated to ASI and HCV (Table 4 and Figure 2).
3.3. Selection of Drought-Tolerant Hybrids Using the Base Index derived from Multiple Traits
Based on the multiple traits base index (MBI) that integrated grain yield, plant aspect, ear aspect, anthesis-silking interval, stay green characteristic, ears per plant, plant height, ear height, and husk cover, 99 hybrids had positive selection base indices. The positive selection base index varied from 0.06 for Entry 150 to 15.93 for Entry 121. Three checks (TZEEI 79 × TZEEI 82, TZdEEI 1 × TZdEEI 12, and TZEEIY Pop STR C5 × TZEEI 58) were identified as drought-tolerant hybrids among the six checks used in the study. The best five top-performing hybrids under managed drought stress were Entries 121, 118, 179, 70, and 106. Three of these five hybrids, namely 70, 106, and 118, were also identified to be superior for drought tolerance using the MGIDI procedure. The five worst hybrids under managed drought were entries 168, 166, 26, 169, and 69 (Table S3). The selected hybrids from MGIDI were included in the list of the hybrids identified as drought–tolerant using MBI without consideration for selection intensity. However, when 15% selection intensity was applied as done for MGIDI, 21 of 29 hybrids (72%) selected were similar to the selection results from MGIDI (Figure 3).
3.4. Yield Performance and Grain Yield Reduction under Drought
Under drought stress condition, grain yield ranged from 198 kg/ha for hybrid TZEEIOR 41 × TZEEIOR 30 to 4044 kg/ha for TZEEIOR 509 × TZEEIOR 197 with an average of 2139 kg/ha. The best yielding hybrid (TZEEIOR 509 × TZEEIOR 197) had 34 % higher grain yield than the best check (TZEEI 79 × TZEEI 82) that had a grain yield of 3015 kg/ha. Under well-watered condition, grain yield varied from 1251kg/ha for TZEEIOR 28 × TZEEIOR 30 to 7455 kg/ha (TZEEIOR 509 × TZEEIOR 197) with a mean of 5568 kg/ha. The best check (TZEEIOR 12 × TZEEIOR 223) had a grain yield of 5732 kg/ha, which translates to 30 % grain yield reduction compared to the best hybrid (Table S4)
In general, grain yield reduction of the hybrids under managed drought stress compared with that of the well-watered conditions varied from 35 % for TZEEIOR 321 × TZEEIOR 249 to 89 % for TZEEIOR 41 × TZEEEIOR 30 with an average of 61 %. The grain yield reduction of the selected drought-tolerant hybrids from MGIDI under drought compared to that of the well-watered varied from 36 % for the check hybrid TZEEI 79 × TZEEI 82 to 69 % for TZEEIOR 202 × TZEEIOR 9A. In addition, 10 (~35%) and 26 (~90%) hybrids from the MGIDI selection had better performance than the selected commercial check under managed drought and well-watered conditions, respectively. The yield advantage of the hybrids with better performance than the selected check was 12 and 33 % under managed drought and well-watered conditions, respectively. For the selected hybrids from MBI, grain yield reduction varied from 35 % for TZEEIOR 321 × TZEEIOR 249 to 62.71 % for TZEEIOR 510 × TZEEIOR 197. Furthermore, 16 (~55%) and 27 (~93%) hybrids from the MBI selection had better performance than the selected commercial check under managed drought and well-watered conditions, respectively (Table 5).
Drought stress reduced PHT by 17 %, EHT by 9 %, EPP by 22 %, DP by 2 %, but increased ASI by 181 %, DS by 0.2%, HCV by 7 %, PASP by 25 %, and EASP by 11 % (Table S5).
3.5. Stability Analysis of Forty PVA Hybrids Comprising Best, Average, and Worst Performing Genotypes across all Environments
The GGE biplot view of the 40 extra-early provitamin A hybrids comprising 20 best, 10 average, and 10 worst performing hybrids from the MGIDI selection analyzed for performance stability is presented in Figure 4. The first and second principal components explained 94.8% of the total variation in grain yield of the extra-early PVA hybrids.
The best-yielding and most stable extra-early provitamin A hybrids were identified using the "mean vs. stability" biplot view across the test environments. The vertical line passing through the biplot origin is the average tester coordinate (ATC) which separates the hybrids based on yield performance. The farther the hybrid was to the right of the ATC, the better the yield performance of the hybrid and vice versa. The stability of the hybrids was determined by their projections from the ATC abscissa or the horizontal line. The shorter the hybrid projection from the horizontal line, the more stable the hybrid and vice versa. Based on these criteria, the five top-yielding hybrids across the test environments in descending order were entries 121, 70, 118, 142, and 159 while the most stable hybrids were entries 142, 149, 81, 68, 119, and 118. From this result, only entries 142 and 118 high high-yielding and stable across the test environment (Figure 4).
3.6. Phenotypic Correlations among Measured Traits under Managed Drought, Well-Watered, and across Research Conditions
The relationships among measured traits in each of the managed drought and well-watered conditions and across both research conditions are presented in Figure 5. Across managed drought environments, GY had significant (p < 0.001) positive correlation with EPP (0.76), EHT (0.29), and PHT (0.38) and significant (p < 0.001) negative correlations with EASP (-0.90), PASP (-0.69), STGC (-0.45), ASI (-0.43) (Figure 5A). Under well-watered conditions, significant (p < 0.001) positive correlations were obtained between GY and EHT (0.39), PHT (0.36), and EPP (0.59) while significant negative correlations were found between grain yield one hand and PASP (r = -0.71, p < 0.001,) and HCV (r = -0.15, p< 0.05) on the other (Figure 5B). Grain yield under managed drought and well-watered conditions had positive correlated (r = 0.49, p < 0.001,). EPP under managed drought had significant and positive correlation with EPP under well-watered conditions (r = 0.30, p < 0.001) and PASP under managed drought showed significant positive correlation with PASP under well-watered conditions (r = 0.2, p < 0.01) (Figure 5C).
4. Discussion
Genetic variability is crucial for the improvement of desired traits in crop improvement programs [21]. Significant genotype mean squares observed for grain yield and other traits under the contrasting research conditions was indicative of the presence of genetic variability among the newly developed extra-early provitamin A hybrids under drought and well-watered conditions. These results provide a basis for rapid genetic gains from selection and identification of hybrids with desirable traits for each research condition [22,23]. Significant environment mean squares observed for anthesis-silking interval and husk cover under managed drought indicated that the difference in environmental conditions that prevailed in the two years of evaluation affected these trait [24]. A similar trend of results was observed for grain yield, anthesis-silking interval, plant aspect, and ears per plant under well-watered conditions. The well-watered trials were carried out under rain-fed conditions. Considerable variation in rainfall occurs from year to year in Nigeria, even for the same site. Consequently, evaluation in several years is necessary, to expose materials being evaluated to the possible variation in rainfall, towards ensuring that materials that are selected are productive under wide rainfall regimes. Significant genotype x environment mean squares observed for anthesis-silking interval, plant height, ears per plant, and stay green characteristic under managed drought indicated that the genotypes differed in the response patterns to the two managed drought environments used in the present study. In contrast, the lack of significant environment and genotype x environment interaction mean squares for grain yield indicate that the relative performances of the genotypes were stable under the managed drought condition investigated. This is also true for the well-watered condition for which the mean squares for genotype by environment interactions were not significant. These results indicate that plant traits differ in their response to environmental conditions.
In the present study, both MGIDI and MBI were each used to identify 29 genotypes combining drought tolerance with high yield. Of the selected genotypes from both selection procedures, 72% similarity was observed. As reported by several authors [15,18,25,26], multiple trait base index is usually affected by multicollinearity of traits which in turn affects the efficiency of selection. MGIDI is, however, not affected by traits multicollinearity making it a better selection procedure when ideotype design is of major interest. In addition, MGIDI provides the knowledge of the strengths and weaknesses of selected ideotype, which could guide plant breeders in identifying traits for further improvement. MGIDI has been used to develop climate-resilient maize hybrids through the development of better selection approaches and optimization of breeding strategies [27]. Improvement in crop performance, the goal of all plant breeding programs, is usually pursued through the assembly in new varieties of genes for various desirable plant traits associated with high grain productivity [28]. The selected 29 drought-tolerant PVA hybrids from MGIDI comprised of 28 new hybrids and one commercial check. The 10 and 26 hybrids ranked above this commercial check under managed drought and well-watered conditions have an average yield advantage of 12% and 33%, respectively. These hybrids show promise for commercialization in SSA as they are not only able to escape drought being extra-early hybrids, but also have the inherent ability to withstand drought occurring during the growing season. These hybrids could also be useful for developing open-pollinated varieties, three-way and double cross hybrids for drought-prone areas in SSA. Ten hybrids from those selected by MGIDI had the tester TZEEIOR 197 as a parent. In a previous study [29], this tester was reported as an outstanding inbred line with positive and significant general combining ability effects for grain yield under drought. These results indicate TZEEIOR 197 to possess genes for drought tolerance as well as show it as an excellent combiner that could be used for the development of more drought-tolerant extra-early PVA hybrids. In the present study, the superior combining ability was further demonstrated by the fact that hybrid combinations involving this tester were more in number than those from any other inbred tester among the selected hybrids combining drought tolerance with high yield.
Examination of the strengths and weaknesses of the MGIDI selected hybrids revealed hybrid 121 (TZEEIOR 509 × TZEEIOR 197) as the only genotype associated with the first factor. This factor correlates with high grain yield, increased ears per plant, and improved ear and plant aspects under drought conditions, an indication that this hybrid has desirable genes for these four traits under managed drought conditions. This provides a basis for its high grain yield under drought stress as well as under well-watered environments. Hybrids associated with the second factor could be selected and used as parent for plant and ear height improvement while hybrids associated to the third factor could be used as parent in a breeding program for the improvement of anthesis-silking interval and husk cover improvement. The initiation of three-way and double-cross hybridization program among these promising hybrids could further increase the number of hybrids with desired performance under drought stress.
In this study, the extended anthesis-silking interval combined with reduction in ears per plant, plant height, ear height and increased plant and ear aspects, and husk cover observed under drought stress contributed to the reduction in grain yield under drought. The imposed drought stress covered the flowering and grain filling periods. Increased anthesis-silking interval is known to increase the incidence of barrenness [30]. The grain yield loss obtained in the present study ranged from 35 to 89 % with an average of 61% under managed drought conditions. These yield losses fall within the range reported by other authors [14,20,31,32]. The magnitude of grain yield reduction as a result of drought is known to depend on the duration and intensity of the drought stress [33,34].
Given that the occurrence of drought is unpredictable and crop varieties must optimize productivity under diverse environmental conditions, improved varieties must show outstanding and stable performance under both drought and well-watered conditions. Hybrids that showed these attributes in the present study and whose superior performances were stably expressed viz. TZEEIOR 509 × TZEEIOR 197, TZEEIOR 458 × TZEEIOR 197 and TZEEIOR 510 × TZEEIOR 97, have great potential for commercialization for cultivation in drought-prone environments and qualify for more extensive testing under diverse conditions.
Grain yield, ears per plant, and plant aspect under managed drought were correlated to the same traits under well-watered conditions suggesting that the expression of these traits is governed by common genetic factors. Similar results have been reported by several authors [31,35,36,37]. The presence of strong associations between grain yield on one hand and ears per plant, plant and ear heights, anthesis-silking interval, plant and ear aspect, and the stay green characteristic under managed drought are indicative of potential of the latter for use as secondary traits for indirect selection in grain yield improvement. These traits have been used to develop selection index for simultaneous improvement of grain yield and drought tolerance with considerable success [38,39]. In the present study, the relationship between grain yield and anthesis-silking interval under well-watered conditions was not significant but significant under managed drought conditions. These results are consistent with the findings of Bolanos and Edmeades [40] who reported that grain yield was weakly correlated to anthesis-silking interval under optimum condition but both are strongly correlated under moisture stress. Knowledge of trait correlations and how the relationships that they define change under contrasting research conditions are useful in guiding plant breeders in the implementation of a breeding program targeting improvements in one trait through improvement in another that is easier to measure [27].
5. Conclusions
The results of this study revealed significant genetic variability among the newly developed extra-early maturing provitamin A hybrids. Some of the hybrids have considerable potential for mitigating food insecurity and reducing malnutrition in SSA. The inbred line TZEEIOR 197 possesses outstanding attributes for the development of extra-early provitamin A hybrids for drought prone environments in SSA. Both multiple trait base index and Multi-traits Genotype-Ideotype Distance Index selection outcome showed 72% similarity. The hybrids TZEEIOR 509 × TZEEIOR 197, TZEEIOR 458 × TZEEIOR 197 and TZEEIOR 510 × TZEEIOR 97 were identified as the most outstanding drought-tolerant hybrids based on the per se performance, yield stability analysis, and Multiple-traits genotype-ideotype distance index. These hybrids should be further tested in multiple stress environments towards efforts to commercialize them in SSA. Additionally, the hybrids could be used as parents for population development, extraction of new and improved inbred lines for further development of new maize ideotypes for farmers in SSA.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Pedigree information of the genetic materials used for the study; Table S2: : Ranking of 196 hybrids using the Multi-Trait Genotype-Ideotype Distance Index (MGIDI); Table S3: Ranking of 196 hybrids using multiple trait base index (MBI), Table S4: Grain yield performance of 196 hybrids evaluated under managed drought and well-watered conditions at Ikenne during 2021-2022 and 2022-2023 dry season and 2022 and 2023 main season, Table S5: traits reduction under managed drought compared to that under well-watered conditions
Author Contributions
Conceptualization, T.O.B., B.B.-A and V.O.A.; methodology, T.O.B.,B.B.-A.,I.I.A; supervisions, B.B.-A.; V.O.A., formal analysis, T.O.B., and I.I.A.; writing—original draft preparation, T.O.B.; writing—review and editing, B.B-A,V.O.A,I.I.A, and K.R.N . All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by African Union through Pan African University and the Bill and Melinda Gates Foundation [OPP1134248].
Data Availability Statement
The datasets used in the present study have been deposited at the IITA CKAN repository.
Acknowledgments
The authors are grateful to the staff of the Maize Improvement Unit of the International Institute of Tropical Agriculture, Ibadan, Nigeria, the African Union Commission, and the African Development Bank through the Pan African University Life and Earth Science Institute (PAULESI) for providing funds for the research. The authors are also grateful for the funding support of the Bill & Melinda Gates Foundation under the Accelerated Genetic Gain in Maize and Wheat (AGG) Projects.
Conflicts of Interest
The authors declare no conflict of interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- FAOSTAT. 2023. Available online: https://www.fao.org/faostat/en/#data/QCL.
- Kurilich, A.C.; Juvik, J.A. Quantification of carotenoid and tocopherol antioxidants in Zea mays. J Agric Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef]
- Tien, L.D.; Duc, C.H.; Quynh, L.N. Improving nutritional quality of plant proteins through genetic engineering. Curr Genomics. 2016, 17, 220–229. [Google Scholar] [CrossRef]
- Villamor, E.; Fawzi, W.W. Vitamin A supplementation: Implications for morbidity and mortality in children. Journal of Infectious Diseases. 2000, 182, S122–S133. [Google Scholar] [CrossRef]
- Sommer, A. Vitamin A deficiency and clinical disease: An historical overview. J Nutr. 2008, 138, 1835–1839. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Beyene, Y.; Babu, R.; Nair, S.; Gowda, M.; Das, B.; et al. Quantitative trait loci mapping and molecular breeding for developing stress resilient maize for Sub-Saharan Africa. Crop Sci. 2015, 55, 1449–1459. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B. Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Springer, 2017; 230p. [Google Scholar]
- NeSmith, D.S.; Ritchie, J.T. Effects of soil water-deficits during tassel emergence on development and yield component of maize (Zea mays). F Crop Res. 1992, 28, 251–256. [Google Scholar] [CrossRef]
- Menkir, A.; Akintunde, A.O. Evaluation of the performance of maize hybrids, improved open-pollinated and farmers’ local varieties under well-watered and drought stress conditions. Maydica 2001, 46, 227–238. [Google Scholar]
- Badu-Apraku, B.; Fontem, L.A.; Akinwale, R.O.; Oyekunle, M. Biplot analysis of diallel crosses of early maturing tropical yellow maize inbreds in stress and nonstress environments. Crop Sci. 2011, 51, 173–188. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M. Genetic analysis of grain yield and other traits of extra-early yellow maize inbreds and hybrid performance under contrasting environments. F Crop Res. 2012, 129, 99–110. [Google Scholar] [CrossRef]
- Jazayeri, S.M.; Torres, R.V. Genomic and transcriptomic approaches toward plant selection. J Sci Res Rev Cienc e Investig. 2017, 2, 54. [Google Scholar] [CrossRef]
- Campos, H.; Cooper, M.; Edmeades, G.O.; Löffler, C.; Schussler, J.R.; Ibañez, M. Changes in drought tolerance in maize associated with fifty years of breeding for yield in the U.S. corn belt. Maydica. 2006, 51, 369–381. [Google Scholar]
- Olivoto, T.; Nardino, M. MGIDI: Toward an effective multivariate selection in biological experiments. Bioinformatics. 2021, 37, 1383–1389. [Google Scholar] [CrossRef]
- Bizari, E.H.; Val, B.H.P.; Pereira, E.D.M.; Mauro, A.O.D.; Unêda-Trevisoli, S.H. Selection indices for agronomic traits in segregating populations of soybean. Rev Cienc Agron. 2017, 48, 110–117. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J Stat Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C. metan: An R package for multi-environment trial analysis. Methods Ecol Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Kaiser, H.F. The varimax criterion for analytic rotation in factor analysis. Psychometrika 1958, 23, 187–200. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M.; Akinwale, R.O. Selection of extra-early maize inbreds under low N and drought at flowering and grain-filling for hybrid production. Maydica. 2011, 56, 29–42. [Google Scholar]
- Yue, H.; Wei, J.; Xie, J.; Chen, S.; Peng, H.; Cao, H.; et al. A study on genotype-by-environment interaction analysis for agronomic traits of maize genotypes across huang-huai-hai region in china. Phyton-International J Exp Bot. 2022, 91, 57–81. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Betrán, F.J.; Rooney, L.W. Combining abilities of quality protein maize inbreds. Crop Sci. 2004, 44, 1997–2005. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Annor, B.; Oyekunle, M.; Akinwale, R.O.; Fakorede, M.A.B.; Talabi, A.O.; et al. Grouping of early maturing quality protein maize inbreds based on SNP markers and combining ability under multiple environments. F Crop Res. 2015, 183, 169–183. [Google Scholar] [CrossRef]
- Bhadmus, O.A.; Badu-apraku, B.; Adeyemo, O.A.; Ogunkanmi, A.L. Genetic analysis of early white quality protein maize inbreds and derived hybrids under low-nitrogen and combined drought and heat stress environments. Plants. 2021, 10, 2596. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Olivoto, T. Multicollinearity in Path Analysis: A Simple Method to Reduce Its Effects. Agron. J. 2017, 109, 131–142. [Google Scholar] [CrossRef]
- Singamsetti, A.; Zaidi, P.H.; Seetharam, K.; Vinayan, M.T.; Olivoto, T.; Mahato, A.; et al. Genetic gains in tropical maize hybrids across moisture regimes with multi-trait-based index selection. Front Plant Sci. 2023, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pour-Aboughadareh, A.; Sanjani, S.; Nikkhah-Chamanabad, H.; Mehrvar, M.R.; Asadi, A.; Amini, A. Identification of salt-tolerant barley genotypes using multiple-traits index and yield performance at the early growth and maturity stages. Bull Natl Res Cent. 2021, 45, 117. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Aderounmu, M.; Lum, A.F.; et al. Genetic studies of extra-early provitamin-A maize inbred lines and their hybrids in multiple environments. Crop Sci. 2020, 60, 1325–1345. [Google Scholar] [CrossRef] [PubMed]
- Edmeades, G.O.; Bolanos, J.; Hernandez, M.; Bello, S. Causes for silk delay in a lowland tropical maize population. Crop Sci. 1993, 33, 1029–1035. [Google Scholar] [CrossRef]
- Oyekunle, M. Genetic analysis and molecular characterisation of early maturing maize (Zea mays L.) inbred lines for drought tolerance; University of Ibadan, 2014. [Google Scholar]
- Badu-Apraku, B.; Menkir, A.; Ajala, S.O.; Akinwale, R.O.; Oyekunle, M.; Obeng-Antwi, K. Performance of tropical early-maturing maize cultivars in multiple stress environments. Can J Plant Sci. 2010, 90, 831–852. [Google Scholar] [CrossRef]
- Széles, A.; Horváth, É.; Simon, K.; Zagyi, P.; Huzsvai, L. Maize production under drought stress: Nutrient supply, yield prediction. Plants. 2023, 12, 3301. [Google Scholar] [CrossRef]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; et al. Impact of water deficit stress in maize: Phenology and yield components. Sci Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Meseka, S.; Menkir, A.; Bossey, B.; Mengesha, W. Performance assessment of drought tolerant maize hybrids under combined drought and heat stress. Agronomy. 2018, 8, 274. [Google Scholar] [CrossRef] [PubMed]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; N’guetta, A.S.P. Assessment of genetic diversity for drought, heat and combined drought and heat stress tolerance in early maturing maize landraces. Plants. 2019, 8, 1–19. [Google Scholar] [CrossRef]
- Cairns, J.E.; Crossa, J.; Zaidi, P.H.; Grudloyma, P.; Sanchez, C.; Luis, A.J.; et al. Identification of drought, heat, and combined drought and heat tolerant donors in maize. Crop Sci. 2013, 53, 1335–1346. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Akaogu, I.C.; Akinwale, R.O.; et al. Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci. 2016, 56, 183–199. [Google Scholar] [CrossRef]
- Nasser, L.M.; Badu-Apraku, B.; Gracen, V.E.; Mafouasson, H.N.A. Combining ability of early-maturing yellow maize inbreds under combined drought and heat stress and well-watered environments. Agronomy. 2020, 10, 19–22. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. F Crop Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
Figure 1.
View of the selected (red dots) and non-selected (grey dots) hybrids based on multiple-traits genotype-ideotype distance index. The lower the MGIDI index of a genotype on the left side of the graph, the closer the genotype is to an ideotype.
Figure 1.
View of the selected (red dots) and non-selected (grey dots) hybrids based on multiple-traits genotype-ideotype distance index. The lower the MGIDI index of a genotype on the left side of the graph, the closer the genotype is to an ideotype.
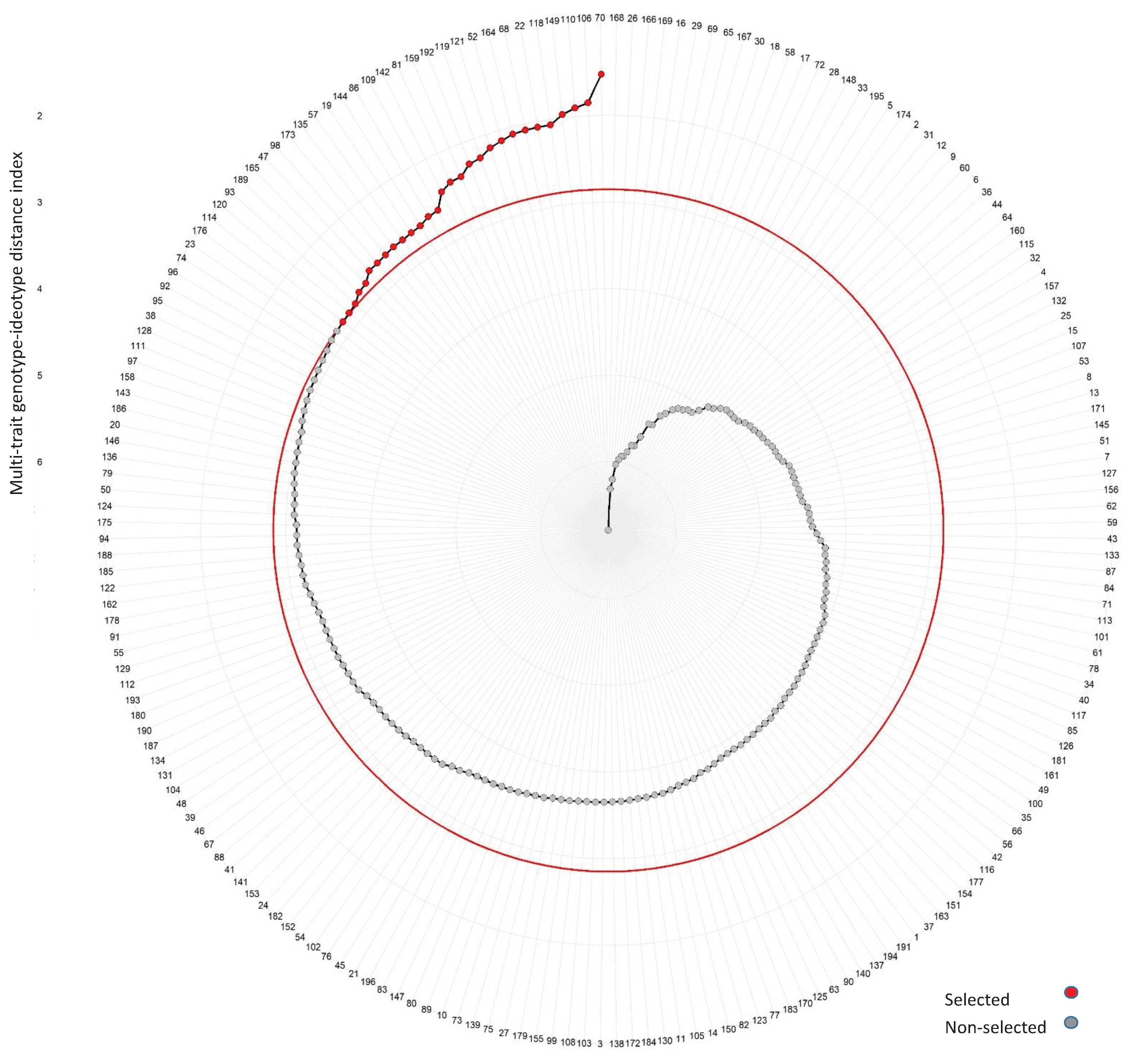
Figure 2.
The strengths and weaknesses plot of the selected 29 hybrids shown as the proportion of each factor based on the computed MGIDI. The red, green and blue lines represent the three factors and the factor protruding the most to a genotype is indicated as the strength of the genotype. The black broken circle shows the theoretical value if all the factors contributed equally.
Figure 2.
The strengths and weaknesses plot of the selected 29 hybrids shown as the proportion of each factor based on the computed MGIDI. The red, green and blue lines represent the three factors and the factor protruding the most to a genotype is indicated as the strength of the genotype. The black broken circle shows the theoretical value if all the factors contributed equally.
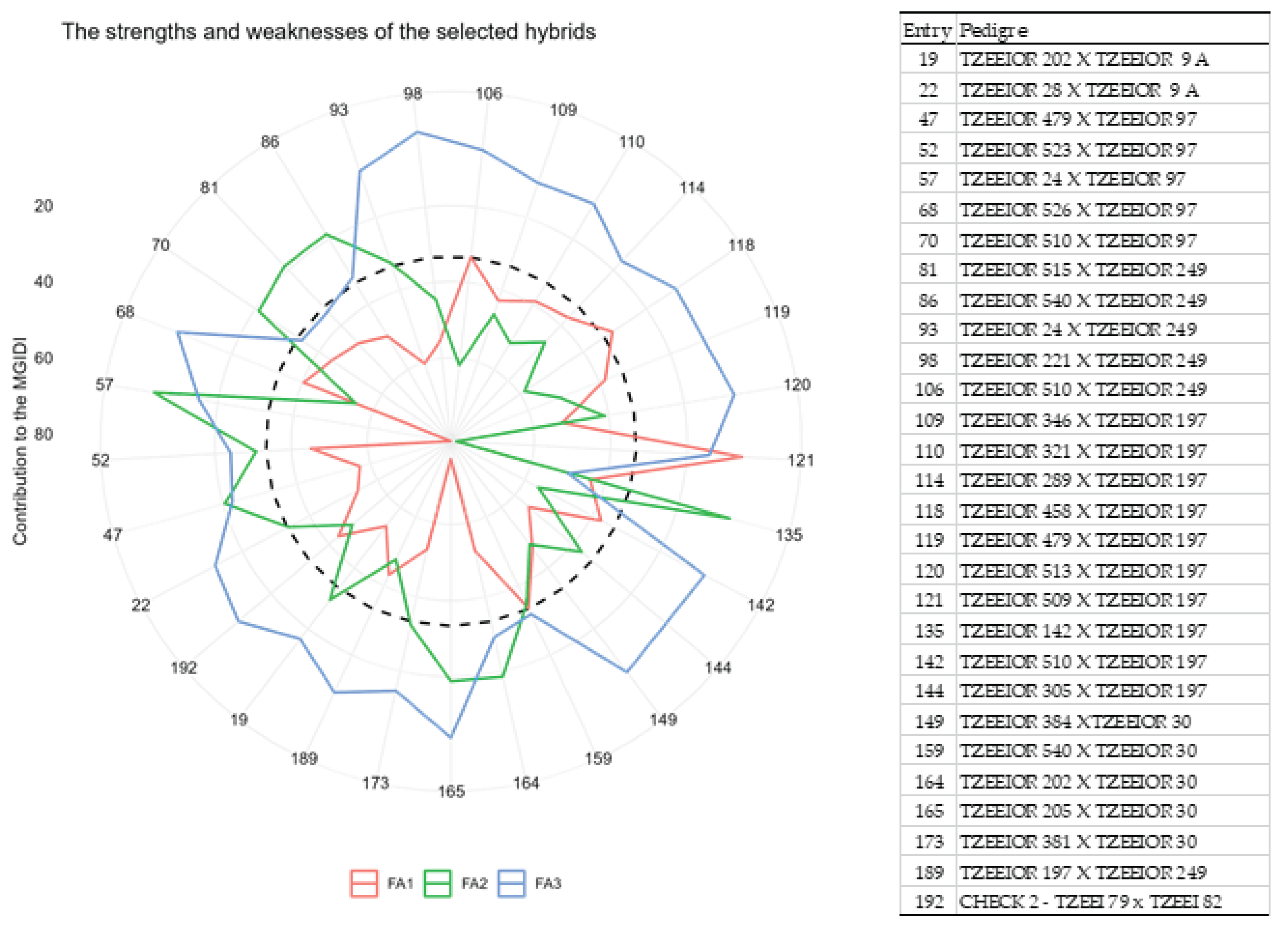
Figure 3.
Venn diagram showing the similarity level (as the intersection) between the multiple base index (MBI) and multi-trait genotype-ideotype distance index (MGIDI) methods.
Figure 3.
Venn diagram showing the similarity level (as the intersection) between the multiple base index (MBI) and multi-trait genotype-ideotype distance index (MGIDI) methods.
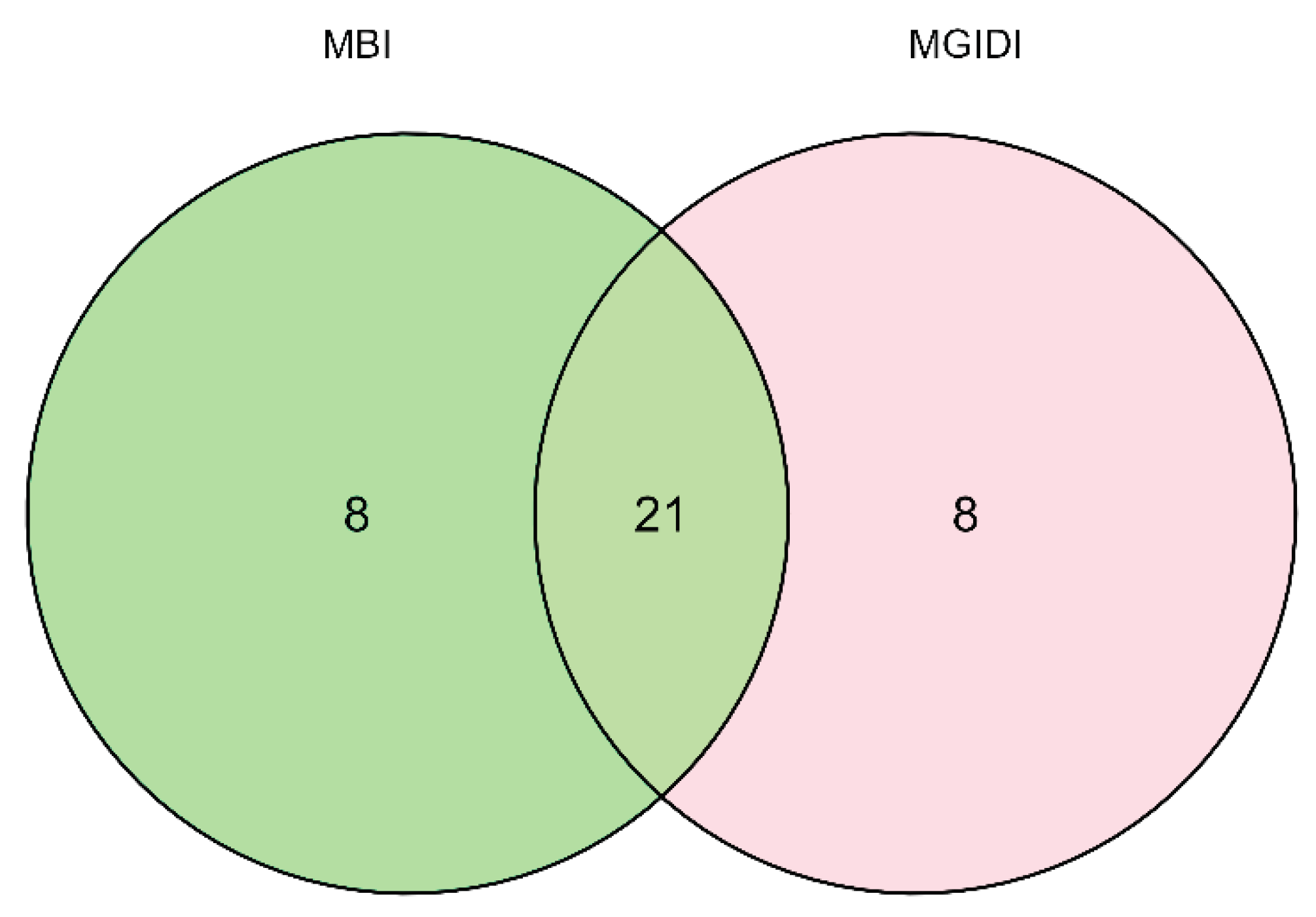
Figure 4.
Mean performance and stability of 40 extra-early provitamin A maize hybrids based on grain yield as measured by the principal components across four environments at Ikenne.
Figure 4.
Mean performance and stability of 40 extra-early provitamin A maize hybrids based on grain yield as measured by the principal components across four environments at Ikenne.
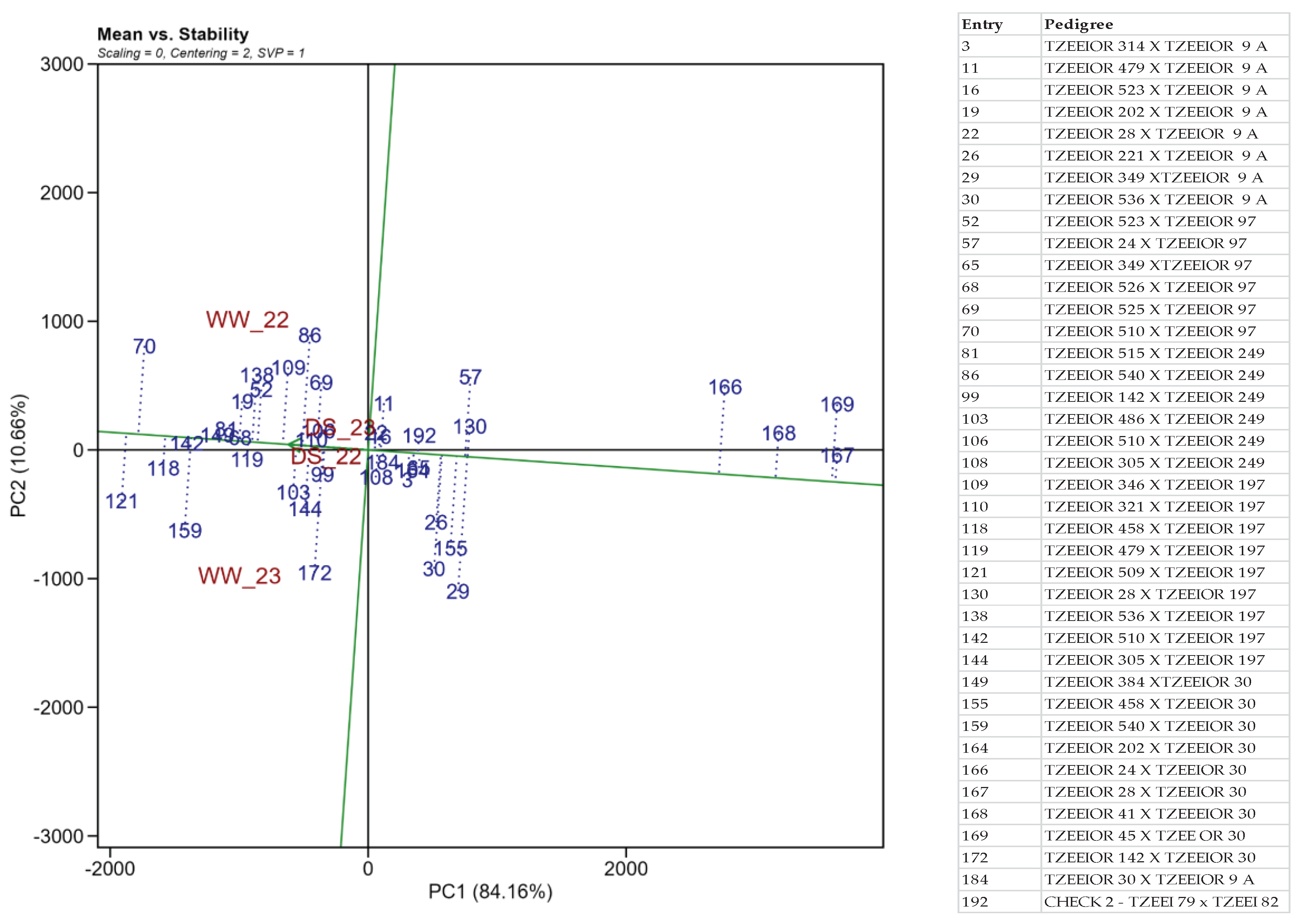
Figure 5.
Phenotypic correlations among measured traits under (A) managed drought, (B) well-watered, and (C) between contrasting research conditions.
Figure 5.
Phenotypic correlations among measured traits under (A) managed drought, (B) well-watered, and (C) between contrasting research conditions.
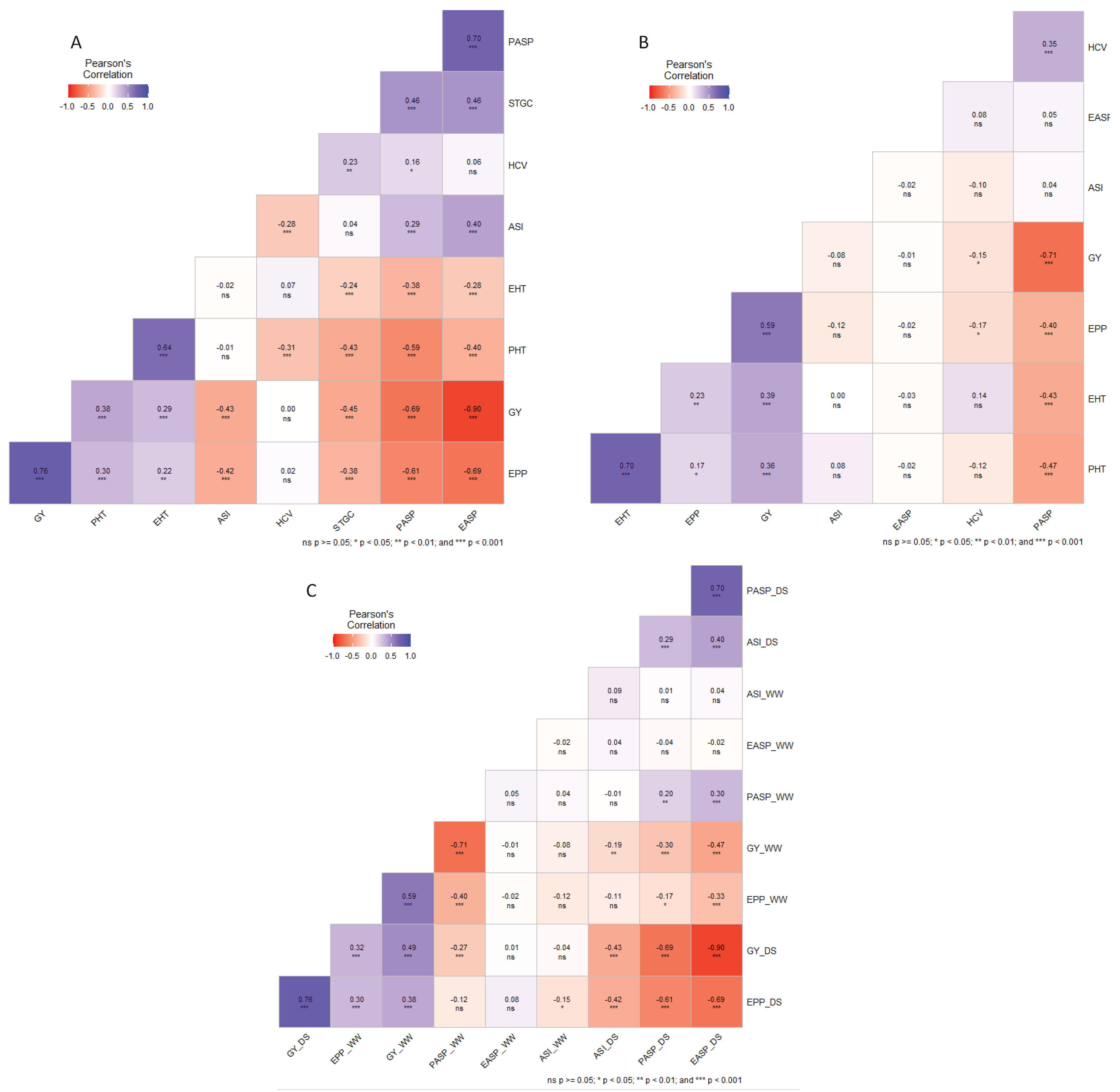

Table 1.
Mean squares of grain yield and other measured traits across well-watered environments.
| Source of variation | DF | GY | ASI | PHT | EHT | PASP | EASP | EPP | HCV |
| Environment (Env) | 1 | 37593481* | 46.76* | 6.65ns | 50.26ns | 37.12* | 3.46ns | 1.25* | 3.71ns |
| Genotype (Gen) | 195 | 4330349*** | 0.71** | 422.55*** | 192.61*** | 2.07*** | 0.85ns | 0.02*** | 7.19*** |
| Env × Gen | 195 | 1174838ns | 0.42ns | 67.95ns | 54.75ns | 0.65ns | 1.00ns | 0.01ns | 0.82ns |
| Residual | 1108155 | 0.40 | 69.28 | 52.12 | 0.89 | 0.75 | 0.01 | 0.84 | |
| Min | 590.5 | 0.00 | 129.2 | 57 | 2 | 2 | 0.38 | 1 | |
| Max | 10064 | 4.00 | 213.8 | 180 | 8 | 8 | 1.27 | 8 | |
| Mean | 5568 | 0.62 | 179.3 | 83.56 | 4.44 | 4.68 | 0.92 | 3.92 |
Env: environment; GY: grain yield; ASI: anthesis-silking interval; PHT: plant height; EHT: ear height; PASP: plant aspect; EASP: ear aspect; EPP: ear per plant; HCV: husk cover; *,**,*** : significant at 0.05, 0.01 and 0.001 probability levels, respectively. ns : non-significant.
Table 2.
Mean squares of grain yield and others agronomic traits across drought stress environments.
Table 2.
Mean squares of grain yield and others agronomic traits across drought stress environments.
| Source of variation | DF | GY | ASI | PHT | EHT | PASP | EASP | EPP | HCV | STGC |
| Environment (Env) | 1 | 1560132ns | 35.57* | 673.19ns | 2.05ns | 8.51ns | 2.45ns | 0.19ns | 81.67*** | 0.10ns |
| Genotype (Gen) | 195 | 1464750*** | 3.085*** | 485.84*** | 193.81*** | 1.55*** | 1.61*** | 0.05*** | 3.90*** | 1.93*** |
| Env × Gen | 195 | 979359.6ns | 2.16* | 258.2** | 70.25ns | 1.08ns | 1.15ns | 0.04** | 0.95ns | 1.07*** |
| Residual | 835346 | 1.57ns | 191.5 | 65.2 | 0.98 | 1 | 0.03 | 0.78 | 0.99 | |
| Min | 0 | 0 | 65 | 34 | 2 | 1 | 0 | 1 | 2 | |
| Max | 8407 | 9 | 205.8 | 128.4 | 9 | 9 | 1.17 | 8 | 9 | |
| Mean | 2138.74 | 1.5 | 147.9 | 76.27 | 5.42 | 5.13 | 0.71 | 3.97 | 5.02 |
Env: environment; GY: grain yield; ASI: anthesis-silking interval; PHT: plant height; EHT: ear height; PASP: plant aspect; EASP: ear aspect; EPP: ear per plant; HCV: husk cover; STGC: stay green characteristic; *,**,***: significant at 0.05, 0.01 and 0.001 probability levels, respectively; ns: non-significant.
Table 3.
Factorial loadings and predicted genetic gains of 196 PVA hybrids based on MGIDI.
| traits | FA1 | FA2 | FA3 | Xo | Xs | Predicted gain | sense | goal |
| ASI | -0.62 | 0.49 | -0.13 | 1.51 | 1.49 | -1.03 | decrease | 100 |
| PHT | -0.28 | -0.33 | 0.80 | 147.87 | 148.24 | 0.25 | increase | 100 |
| EHT | -0.11 | 0.13 | 0.93 | 76.36 | 78.69 | 3.04 | increase | 100 |
| HCV | 0.00 | -0.90 | -0.02 | 3.97 | 3.57 | -10.13 | decrease | 100 |
| PASP | -0.73 | -0.20 | 0.41 | 5.42 | 5.41 | -0.17 | decrease | 100 |
| EASP | -0.89 | -0.08 | 0.19 | 5.13 | 5.06 | -1.24 | decrease | 100 |
| EPP | -0.85 | 0.02 | 0.11 | 0.71 | 0.73 | 2.63 | increase | 100 |
| GY | -0.91 | -0.02 | 0.19 | 2138.74 | 2267.90 | 6.04 | increase | 100 |
| STGC | -0.48 | -0.47 | 0.27 | 5.02 | 4.82 | -4.08 | decrease | 100 |
NB: (Bold values represent traits with high contribution to each component). ASI: anthesis-silking interval, PHT: plant height, EHT: ear height, HCV: husk cover, PASP: plant aspect, EASP: ear aspect, EPP: ear per plant, GY: grain yield, STGC: stay green characteristic
Table 4.
Factorial loadings, communalities and uniqueness of 29 selected PVA hybrids based on the Multi-Trait Genotype–Ideotype Distance Index.
Table 4.
Factorial loadings, communalities and uniqueness of 29 selected PVA hybrids based on the Multi-Trait Genotype–Ideotype Distance Index.
| Traits | FA1 | FA2 | FA3 | Communality | Uniqueness |
| ASI | 0.13 | 0.26 | 0.87 | 0.84 | 0.16 |
| PHT | 0.15 | -0.84 | -0.05 | 0.73 | 0.27 |
| EHT | -0.27 | -0.71 | 0.10 | 0.59 | 0.41 |
| HCV | -0.02 | 0.36 | -0.74 | 0.69 | 0.31 |
| PASP | 0.72 | 0.21 | 0.01 | 0.57 | 0.43 |
| EASP | 0.91 | -0.06 | 0.03 | 0.82 | 0.18 |
| EPP | 0.74 | 0.00 | 0.14 | 0.57 | 0.43 |
| GY | 0.81 | -0.08 | 0.19 | 0.69 | 0.31 |
| STGC | 0.38 | 0.09 | -0.18 | 0.18 | 0.82 |
ASI: anthesis-silking interval, PHT: plant height, EHT: ear height, HCV: husk cover, PASP: plant aspect, EASP: ear aspect, EPP: ear per plant, GY: grain yield, STGC: stay green characteristic.
Table 5.
Grain yield (kg/ha) of selected drought-tolerant extra-early maturing hybrids identified from both MGIDI and multiple traits base-index with 72 % level of similarity.
Table 5.
Grain yield (kg/ha) of selected drought-tolerant extra-early maturing hybrids identified from both MGIDI and multiple traits base-index with 72 % level of similarity.
| Entries | Genotypes | GY (kg/ha) under managed drought conditions | GY (kg/ha) under well-watered conditions | YR% | MBI | MGIDI |
| 19 | TZEEIOR 202 × TZEEIOR 9 A | 2098.17 | 6714.28 | 68.75 | 5.39* | 2.65 |
| 22 | TZEEIOR 28 × TZEEIOR 9 A | 2501.02 | 5341.02 | 53.17 | 7.79 | 2.06 |
| 39 | TZEEIOR 314 × TZEEIOR 97 | 3604.17 | 6993.1 | 48.46 | 8.38 | 3.37* |
| 47 | TZEEIOR 479 × TZEEIOR 97 | 2857.75 | 5247.55 | 45.54 | 7.70 | 2.69 |
| 52 | TZEEIOR 523 × TZEEIOR 97 | 3054.03 | 6385.56 | 52.17 | 10.5 | 2.12 |
| 57 | TZEEIOR 24 × TZEEIOR 97 | 2502.56 | 4392.24 | 43.02 | 6.41 | 2.66 |
| 68 | TZEEIOR 526 × TZEEIOR 97 | 3318.75 | 6435.87 | 48.43 | 9.69 | 2.07 |
| 70 | TZEEIOR 510 × TZEEIOR 97 | 3568.91 | 7379.62 | 51.64 | 15.07 | 1.53 |
| 74 | TZEEIOR 321 × TZEEIOR 249 | 3207.81 | 4918.39 | 34.78 | 9.31 | 2.89* |
| 81 | TZEEIOR 515 × TZEEIOR 249 | 2917.19 | 6722.3 | 56.6 | 9.81 | 2.36 |
| 86 | TZEEIOR 540 × TZEEIOR 249 | 2922.97 | 5818.24 | 49.76 | 9.02 | 2.60 |
| 92 | TZEEIOR 205 × TZEEIOR 249 | 2558.98 | 5240.41 | 51.17 | 8.46 | 2.91* |
| 93 | TZEEIOR 24 × TZEEIOR 249 | 1988.74 | 4367.42 | 54.46 | 2.29* | 2.83 |
| 98 | TZEEIOR 221 × TZEEIOR 249 | 2156.96 | 5186.18 | 58.41 | 4.19* | 2.68 |
| 106 | TZEEIOR 510 × TZEEIOR 249 | 3294.78 | 5845.61 | 43.64 | 12.15 | 1.85 |
| 109 | TZEEIOR 346 × TZEEIOR 197 | 2438.1 | 6397.55 | 61.89 | 6.58 | 2.59 |
| 110 | TZEEIOR 321 × TZEEIOR 197 | 3116.3 | 5714.04 | 45.46 | 11.24 | 1.90 |
| 114 | TZEEIOR 289 × TZEEIOR 197 | 2873.28 | 7341.28 | 60.86 | 7.61 | 2.85 |
| 118 | TZEEIOR 458 × TZEEIOR 197 | 3642.91 | 7062.23 | 48.42 | 15.91 | 2.06 |
| 119 | TZEEIOR 479 × TZEEIOR 197 | 3073.8 | 6428.36 | 52.18 | 10.41 | 2.24 |
| 120 | TZEEIOR 513 × TZEEIOR 197 | 2799.11 | 6953.46 | 59.75 | 6.03* | 2.85 |
| 121 | TZEEIOR 509 × TZEEIOR 197 | 4044.13 | 7454.93 | 45.75 | 15.94 | 2.17 |
| 128 | TZEEIOR 205 × TZEEIOR 197 | 2308.17 | 5989.75 | 61.46 | 6.71 | 2.95* |
| 135 | TZEEIOR 142 × TZEEIOR 197 | 2578.45 | 5953.68 | 56.69 | 6.03* | 2.66 |
| 142 | TZEEIOR 510 × TZEEIOR 197 | 2628.79 | 7050.98 | 62.72 | 8.90 | 2.42 |
| 144 | TZEEIOR 305 × TZEEIOR 197 | 2648.3 | 5885.05 | 55 | 6.08* | 2.64 |
| 149 | TZEEIOR 384 ×TZEEIOR 30 | 3414.92 | 6752.58 | 49.43 | 10.71 | 1.96 |
| 158 | TZEEIOR 509 × TZEEIOR 30 | 3096.97 | 5469.03 | 43.37 | 6.84 | 3.01* |
| 159 | TZEEIOR 540 × TZEEIOR 30 | 3261.14 | 7086.18 | 53.98 | 10.69 | 2.36 |
| 164 | TZEEIOR 202 × TZEEIOR 30 | 2297.08 | 4917.55 | 53.29 | 8.97 | 2.08 |
| 165 | TZEEIOR 205 × TZEEIOR 30 | 2359.71 | 6279.68 | 62.42 | 4.80* | 2.76 |
| 173 | TZEEIOR 381 × TZEEIOR 30 | 2999.96 | 5897.69 | 49.13 | 6.97 | 2.67 |
| 175 | TZEEIOR 536 × TZEEIOR 30 | 3498.09 | 7390.22 | 52.67 | 6.34 | 3.13* |
| 178 | TZEEIOR 525 × TZEEIOR 30 | 3217.1 | 5617.52 | 42.73 | 8.85 | 3.17* |
| 179 | TZEEIOR 510 × TZEEIOR 30 | 3805.47 | 6392.13 | 40.47 | 15.18 | 3.63* |
| 189 | TZEEIOR 197 × TZEEIOR 249 | 2299.81 | 5464.1 | 57.91 | 4.53 | 2.78 |
| 192 | CHECK 2 - TZEEI 79 × TZEEI 82 | 3015.16 | 4720.41 | 36.13 | 7.91 | 2.26 |
GY: grain yield; YR: yield reduction; MBI: multiple base index; MGIDI: multi-traits genotype-ideotype distance index; *: hybrids that were not selected as drought-tolerant using the alternative selection method.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



