
Submitted:
22 March 2024
Posted:
25 March 2024
You are already at the latest version
Abstract
A free-fall baited camera lander was launched for the first time on the Cosmonaut Sea shelf of East Antarctica at the depth of 694 m during the 38th Chinese National Antarctic Research Expedition (CHINARE) in 2022. We identified 31 unique taxa (23 of invertebrates and 8 of fishes) belonging to 8 phyla from 2403 pictures and 40 videos. The Antarctic jonasfish (Notolepis coatsi) was the most frequently observed fish taxa. Ten species of vulnerable marine ecosystem (VME) taxa were observed, accounting for 32% of all species. The maximum number (MaxN) of Natatolana meridionalis individuals per image frame was ten, and they were attracted to the bait. The macrobenthic community type was sessile suspension feeders with associated fauna (SSFA), which was shaped by the muddy substrata with scattered rocks. Rocks served as the best habitats for sessile fauna. The study reveals the megafauna community and their habitat by image survey in the Cosmonaut Sea for the first time. It helped us obtain Antarctic biodiversity baselines and monitoring data for future’s ecosystem health assessment and better protection.
Keywords:
Lander
; Cosmonaut Sea
; megafauna
; Antarctica
; Image survey
1. Introduction
The Southern Ocean is unique among the world's oceans in terms of its linkage with the other major ocean basins, its rich and unusual marine ecosystem, and its interaction with the physical climate system and the biogeochemistry of the region [1]. It comprises 15% of the world’s oceans and is home to thousands of endemic species [2,3,4]. The Southern Ocean has become a hot area for global research on climate change and ecological evolution due to its harsh natural environment, fragile ecosystem, and sensitivity to environmental variations. There is a growing need for marine biodiversity baselines and monitoring data to assess ocean ecosystem health, especially that around Antarctica where data are rare [5,6,7,8].
The Cosmonaut Sea (30°~60°E) is located to the west of Enderby Land in east Antarctica, and has been poorly explored [9]. Therefore, very few biological data have been recorded for the region [10,11,12,13], and existing ones mainly include the composition and distribution of phytoplankton, mesozooplankton, euphausiid larvae, krill, squid and Antarctic jonasfish [14,15,16]. But there is still no data about macrobenthos in the Cosmonaut Sea. Antarctic benthic ecosystems in the Cosmonaut Sea may be sentinels for monitoring the effects of climate change [17,18]. Macrobenthic communities in Antarctica differ in biodiversity and ecosystem functioning and are shaped by a variety of physical and biological drivers [19]. Some of these communities are unique in their occurrence and proportions of species and life forms. Some are typical for the entire Antarctic shelf but never occur with exactly the same proportions or compositions [18].
Trawls, sledges, and dredges are historically the common facilities for sampling the shelf benthic marine communities of Antarctica [12,20,21]. These traditional methods can help understanding the structure and function of the Antarctic benthic systems and are good for species identification of slow-moving macrobenthos. However, they cannot describe species behaviors and interactions, and have difficulty in capturing more mobile species [4]. Video surveys are the emerging methods to estimate the relative abundances of scavenging fishes and invertebrates [8,22,23]. Such studies can help us develop a better understanding of ecosystem patterns and processes in Antarctica, such as the Antarctic Peninsula, Prydz Bay and the Amundsen Sea, and the applicability has also been justified [4,24,25,26,27].
In this study, a free-fall baited camera lander was deployed for the first time on the Cosmonaut Sea shelf during the 38th Chinese National Antarctic Research Expedition (CHINARE) on 22 February 2022. The objective is to preliminarily survey fish and invertebrate communities, and basically understand the structure of the benthic community and the interactions among species on the Cosmonaut Sea shelf.
2. Method and Equipment
2.1. Location
2.2. Equipment
The lander system framework is made of titanium, consisting of batteries, an acoustic releaser, cameras, and traps (Figure 2). The batteries are rechargeable lithium battery combined with capacity greater than 96 Ah. The releaser is manufactured by iXblue. The load and communication distance of the iXblue releaser are not less than 2500 kg and 10 km, respectively. iXblue is powered by a No.1 alkaline battery or lithium battery. The Sea-Bird Scientific SBE 37 is equipped with high-accuracy temperature, conductivity and pressure sensors with an RS-232 interface, internal batteries, data storage and pump. The sampling interval was set to 120 seconds and the maximum observation depth was 7000 m. Two cameras were configured: one (CO01-016E, 3648×2736) was used to take photos, and the other (CO02-016HE, HD1080P) was used to record videos. The lander used a high efficiency LED lamp (CO04-50LI-6000AA), and the light was angled downward at a constant 45°from the horizontal plane. Two traps were used in the study: a large fish and invertebrate trap (82 cm × 47 cm × 47 cm), and invertebrate trap (φ10 cm × 50 cm).
2.3. Deployment Process
The lander was deployed and recovered during the 38th CHINARE on the R/V Xuelong2 icebreaker, to a depth of 694 m. The equipment was deployed at 20:09 and landed on the sea floor at 20:26 on 22 February 2022 (UTC).
The lander deployment process had four stages: setup, deployment, recovery and data exporting. Prior to deployment, the cameras were activated through underwater systems by a computer. The camera was preprogrammed to take 1 picture every 30 sec, and the video camera was preprogrammed to take 10 min of video every 30 min. The survey time was set according to the battery charging time. Then, baits (chicken legs and Silver sillago) were loaded into the traps. The last step was connecting the acoustic releaser to the cement block. Deployment was completed, and the lander was dropped into the sea. The dropping rate was approximately 1 m/s. Location and bottom depths were based on triangulation of the acoustic releases after the deployment [28]. The lander was recovered in the daytime when the sea was calm. The ship travelled to the deployment area for lander recovery. To recover the lander, the deck unit was used to search for the signal from the acoustic releaser, and the distance between the equipment and the ship was measured. Then, the deck unit released the signal to recover the lander. The rising rate was approximately 1 m/s. The crew searched for the lander, which was attached to orange balls from the bridge. Finally, the equipment was found and brought up onto the deck.
The lander was washed using fresh water after being placed on the deck. Then, the samples were collected from the trap cage. Following retrieval, an external hard disk was used to store the images and videos from the cameras, and the data from SBE 37 for further analysis.
2.4. Metrics and Biodiversity Analyses
Annotations were made from the video footages to identify species to the highest taxonomic resolution [8]. A relative abundance metric-the maximum number (MaxN) was used in the video survey. Counts of MaxN for individuals of each species in a video frame were performed rather than the total tally per deployment to avoid double-counting [29]. Other variables were also recorded, such as the time for a taxon’s first arrival of, and the time when the maximum number of a taxon was observed upon the landing of the device on the seafloor [8,23,30].
3. Results
A total of 2403 photos and 40 videos were recorded underwater by the lander during approximately 20 hours.
3.1. Substrate Type
The images were used to determine substrate characteristics. The transect was dominated by muddy substrate containing some rocks. The rocks were scattered on the sea floor, and served as substrata for different species, such as sponges, bryozoans and corals. These species, in turn, served as living substrata for ophiuroids, asteroids, holothurians and others (Figure 3).
3.2. Observation of the Hydrological Environment
The SBE 37 started to record data on salinity, temperature and pressure at the bottom at 21:00. The pressure was relatively stable with the average of 701.081 dbar and increased slowly during the whole observation period. The highest pressure was 701.545 dbar at 16:06, and the lowest one was 700.542 at 23:42. The average value of salinity and temperature was 34.6096 psu and 0.1091 ℃, respectively. The highest salinity was 34.6446 psu at 5:42, and the highest value of temperature was 0.1584 ℃ at 8:20. Salinity and temperature varied strongly (Figure 4) with a sharp fall at 13:00. The salinity reached its lowest value (34.5511 psu) at 13:42. The temperature reached its lowest value (-0.0478 ℃) at 13:32. The lower temperature and salinity lasted approximately one hour, and they quickly increased at 2:20 p.m. Data here may imply that there was a low-temperature and low-salinity water mass passing over the bottom.
3.3. Recorded Taxa
Thirty-one species were identified, representing 8 phyla, 20 orders and 30 families (Table 1). Invertebrates accounted for 23 species, and fishes accounted for 8 species. Bryozoa dominated in the images (Figure 5).
Three fish were the most frequently occurring fish taxa. Notolepis coatsi appeared in 37 videos with even up to 4 individuals in one frame, in 29 of 2403 photos with up to 3 individuals in one photo. Trematomus lepidorhinus was found in 16 of 40 videos, and 809 of 2403 photos. Melanostigma gelatinosum appeared in 117 of 2403 photos. Dissostichus mawsoni was recorded only once by the cameras at 3:35, 23 February 2022.
Ten species of VME (vulnerable marine ecosystem) taxa were observed, accounting for 32% of all species. VME taxa include sponges, sea mats and cold-water corals, which can provide important habitat for a diversity of marine organisms [31]. Sea pens, sea anemones and sea squirts were the VME taxa commonly observed (Table 1).
Nothoadmete sp. was firstly observed at 21:09, and the MaxN per image frame was four. Natatolana meridionalis was observed at 9:17, and the MaxN per image frame was ten. These two species were the main ones influencing the MaxN of individuals per image frame each hour.
The species caught in the traps were Isopoda (N. meridionalis), Amphipoda (Abyssorchomene sp.) and fishes (N. coatsi) with 9, 13 and 3 individuals, respectively.
4. Discussion
4.1. Substrate Type and Assemblages
Seabed imagery was used to determine substrate characteristics. Substrate type and water depth have frequently been associated with the distribution of benthic biota on the Antarctic shelf [32,33]. Muddy substrate with scattered rocks was found in this study. Many rocks may be dropstones from iceberg scouring [33]. Rocks were found to serve as substrata for sessile taxa, such as sponges, bryozoans and corals. These species, in turn, served as living substrata for ophiuroids, asteroids, holothurians and others in this survey. Rocks are important for building benthic communities, and they provide a hard attachment site and an elevated position off the bottom enhancing the food supply for sessile suspension feeders, such as sponges, bryozoans, pennatulaceun, and actiniae [32]. Deposit feeders are often associated with soft substrates where they are able to feed on fine particles [34]. Amphipods were associated with muddy substrates from the images recorded, and the phytodetritus in muddy substrates may be the food source for them.
4.2. Hydrological Environment and Assemblages
Meijers et al. [9] found seasonal Antarctic Bottom Water (AABW) produced in the region (60°E), which moved down the slope and was deflected westward due to the Coriolis force. The mass water mixed with Antarctic Circumpolar Current (ACC) and Weddell Gyre waters above it as it moved westward across the regions 30°E、40°E and 50°E,eroding the strong characteristics observed in the region (60°E). Lowered acoustic Doppler current profiler (LADCP) data showed that the bottom water had large velocities near the deployment area [9]. The different postures of U. carpenteri showed high-strength and disordered current at the bottom. The current at the bottom brought a large number of organic debris, which provided the food source for sessile suspension feeders, such as sponges, colder-water corals and sea pens. This was important for structuring the benthic community.
4.3. Comments on Fishes and Invertebrates in Images
Notolepis coatsi was the most frequently encountered fish taxon and the second-largest fish biomass in the Cosmonaut Sea of this study like that in Prydz Bay [15,16]. It is a midsize bathypelagic fish widely distributed around the Antarctic continental shelf [16]. N. coatsi may feed only on krill [35] and account for 50% of the food consumed by Antarctic fur seals (Arctocephalus gazella) [36]. Therefore, N. coatsi plays an important role in the marine food web, which directly links krill and marine mammals in the Southern Ocean. N. coatsi is regarded as an ideal organism to evaluate the role of the Cosmonaut Sea, because of its large population size, position in the food web and specialized diet [16]. Additionally, we not only explored N. coatsi behavior, but also trapped the individuals using the lander.
Dissostichus mawsoni was observed by photographic survey for the first time in the Cosmonaut Sea and is a large nototheniid species endemic to the Antarctic continental shelf. The Antarctic toothfish is an important fishery resource in the Southern Ocean and plays an important role in Antarctic ecosystems [11]. Sufficient data and stock information are needed to establish fisheries, but such information is rare in the Cosmonaut Sea. The lander can help us assess the relative abundance and population size structure of the Antarctic toothfish in the Cosmonaut Sea.
In total, 10 VME taxonomic categories were observed from the lander imagery, accounting for 32% of all species. The VME taxa distributed sparsely, and most of them lived on rocks. The presence of benthic invertebrates contributes to the creation of complex three-dimensional structures and provides substrata for other organisms called sessile suspension feeders with associated fauna (SSFA) [32,33]. The main sessile suspension feeders recorded in this survey were Bryozoa. Associated fauna included Ophiuroidea, Asteroidea, Gastropoda, Isopoda and others. Isopoda and Amphipoda were the main taxa in the trap, so the bait is possibly attractive for them. Natatolana meridionalis firstly arrived at the bait fifty-one minutes after the lander arrived at the seafloor. The bait was consumed for approximately eight hours until it was fully exhausted. Data from the lander can help assess changes in benthic community diversity and their associated habitat structure.
4.4. Observation Advantages and Limitations of the Lander
A major advantage of the lander is that it can explore fauna behavior, including the interactions between species. It is a powerful tool for the detection of rare predatory fish species [37], which are difficultly found through physical sampling. The structure of the community should be visually determined to better understand vulnerable marine ecosystems. The lander thus allows us to clarify anthropogenic impacts, such as fishing and climate change. Also, the density of data for macrobenthos is low, especially in the Cosmonaut Sea. Although traditional trawling is advisable for the collection of all faunal types for higher resolution taxonomic investigations, the lander adds an existing monitoring program to extend biological observing capacity into the Antarctic Ocean [8]. Moreover, the lander is an unnamed vehicle that falls to the seafloor unattached to any cable, subsequently operating autonomously at the bottom, and it can carry diverse instruments for environmental parameter collections [38].
However, the lander cannot acquire images of small sessile fauna (meiofauna, infauna). Quantitative analysis like density per unit cannot be achieved, and only the relative measures of abundance (MaxN) can be applied at this moment. As the lander could help to better understand the current spatial variability on the Cosmonaut Sea shelf fauna, data here would serve as a supplement of the baseline for future comparisons [4].
Acknowledgement
We thank Leader Ning Xu, Captain Zhimin Xiao, Chief Engineer Xiaodong Chen, Chief Officer Hao Xing, Second Officer Tieyuan Li, Third Officer Xian Su, Boatswain Yaokui Fu and all crew aboard the XUE LONG 2 Icebreaker for the security and logistical support during the 38th Chinese National Antarctic Research Expedition. We extend our heartfelt thanks to Dr. Pei-long Ju, Dr. Hai-bo Li, Dr. Ji-chang Zhang, Dr. Cai Zhang and Dr. Li-wei Kou for help in the sampling and data analysis processes.
Author Contribution Statement
Jianfeng Mou: Investigation, Data curation, Writing-original draft, Validation. Xuebao He: Conceptualization, Software, Writing-review & editing. Kun Liu: Data curation, Software. Yaqin Huang: Data curation, Writing-review & editing. Shuyi Zhang: Data curation. Yongcan Zu: Data curation, Writing-review & editing. Yanan Liu: Data curation, Writing-review & editing. Shunan Cao: Investigation, Supervision. Musheng Lan: Investigation, Supervision. Xing Miao: Investigation. Heshan Lin: Supervision, Conceptualization. Wenhua Liu: Supervision, Conceptualization, Resources.
Funding
This work was supported by The National Natural Science Foundation of China under contract (No. 42076237, No. 42106227, No. 42106065) and Impact and Response of Antarctic Seas to Climate Change (IRASCC2020-2025-NO.01-02-02 & 02-02).
Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Availability of data and material
Not applicable.
Ethics approval
Although our research was about benthos in the Antarctica, all the data were from the images of the lander. Therefore, we believe an ethical review process was not required for our study.
Code availability
Not applicable.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Consent for publication
Not applicable.
References
- Busalacchi, A.J. The role of the Southern Ocean in global processes: an earth system science approach. Antarct. Sci. 2004, 16, 363–368. [Google Scholar] [CrossRef]
- Convey, P.; Stevens, M.I. Antarctic Biodiversity. Science 2007, 317, 1877–1878. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.J. Antarctic Marine Biodiversity – What Do We Know About the Distribution of Life in the Southern Ocean? PLOS ONE 2010, 5, e11683. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, A.M.; Goodell, W.; Salinas-De-León, P.; Ballesteros, E.; Berkenpas, E.; Capurro, A.P.; Cárdenas, C.A.; Hüne, M.; Lagger, C.; Landaeta, M.F.; et al. Spatial patterns of continental shelf faunal community structure along the Western Antarctic Peninsula. PLOS ONE 2020, 15, e0239895. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J.; Coll, M.; Danovaro, R.; Halpin, P.; Ojaveer, H.; Miloslavich, P. A Census of Marine Biodiversity Knowledge, Resources, and Future Challenges. PLOS ONE 2010, 5, e12110. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.A.; Bett, B.J.; Gates, A.R.; Heimbach, P.; Howe, B.M.; Janssen, F.; McCurdy, A.; Ruhl, H.A.; Snelgrove, P.; Stocks, K.I.; et al. Global Observing Needs in the Deep Ocean. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Rogers, A.; Aburto-Oropeza, O.; Appeltans, W.; Assis, J. Critical Habitats and Biodiversity: Inventory, Thresholds and Governance. Washington, DC: World Resources Institute, 2020.
- Giddens, J.; Turchik, A.; Goodell, W.; Rodriguez, M.; Delaney, D. The National Geographic Society Deep-Sea Camera System: A Low-Cost Remote Video Survey Instrument to Advance Biodiversity Observation in the Deep Ocean. Front. Mar. Sci. 2021, 7. [Google Scholar] [CrossRef]
- Meijers, A.; Klocker, A.; Bindoff, N.; Williams, G.; Marsland, S. The circulation and water masses of the Antarctic shelf and continental slope between 30 and 80∘E. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2010, 57, 723–737. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Peck, L.S. Vulnerability of Antarctic shelf biodiversity to predicted regional warming. Clim. Res. 2008, 37, 149–163. [Google Scholar] [CrossRef]
- Barnes, D.K.; Clarke, A. Antarctic marine biology. Curr. Biol. 2011, 21, R451–R457. [Google Scholar] [CrossRef] [PubMed]
- Linse, K.; Griffiths, H.J.; Barnes, D.K.; Brandt, A.; Davey, N.; David, B.; De Grave, S.; D′Acoz, C.D.; Eléaume, M.; Glover, A.G.; et al. The macro- and megabenthic fauna on the continental shelf of the eastern Amundsen Sea, Antarctica. Cont. Shelf Res. 2013, 68, 80–90. [Google Scholar] [CrossRef]
- Mou, J.; Liu, K.; Huang, Y.; Lin, J.; He, X.; Zhang, S.; Li, D.; Zu, Y.; Chen, Z.; Fu, S.; et al. Species Diversity and Community Structure of Macrobenthos in the Cosmonaut Sea, East Antarctica. Diversity 2023, 15, 1197. [Google Scholar] [CrossRef]
- Hunt, B.; Pakhomov, E.; Trotsenko, B. The macrozooplankton of the Cosmonaut Sea, east Antarctica (30°E–60°E), 1987–1990. Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2007, 54, 1042–1069. [Google Scholar] [CrossRef]
- Van de Putte, A.P.; Jackson, G.D.; Pakhomov, E.; Flores, H.; Volckaert, F.A. Distribution of squid and fish in the pelagic zone of the Cosmonaut Sea and Prydz Bay region during the BROKE-West campaign. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2010, 57, 956–967. [Google Scholar] [CrossRef]
- Ran, Q.; Duan, M.; Wang, P.; Ye, Z.; Mou, J.; Wang, X.; Tian, Y.; Zhang, C.; Qiao, H.; Zhang, J. Predicting the current habitat suitability and future habitat changes of Antarctic jonasfish Notolepis coatsorum in the Southern Ocean. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2022, 199. [Google Scholar] [CrossRef]
- Sahade, R.; Lagger, C.; Torre, L.; Momo, F.; Monien, P.; Schloss, I.; Barnes, D.K.A.; Servetto, N.; Tarantelli, S.; Tatián, M.; et al. Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Sci. Adv. 2015, 1, e1500050. [Google Scholar] [CrossRef] [PubMed]
- Mou, J.; Liu, K.; Huang, Y.; He, X.; Zhang, S.; Wang, J.; Lin, J.; Lin, H.; Liu, W. The macro-and megabenthic fauna on the continental shelf of Prydz Bay, east Antarctica. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2022, 198. [Google Scholar] [CrossRef]
- Gutt, J. Some “driving forces” structuring communities of the sublittoral Antarctic macrobenthos. Antarct. Sci. 2000, 12, 297–313. [Google Scholar] [CrossRef]
- Arnaud, P.M.; López, C.M.; Olaso, I.; Ramil, F.; Ramos-Esplá, A.A.; Ramos, A. Semi-quantitative study of macrobenthic fauna in the region of the South Shetland Islands and the Antarctic Peninsula. Polar Biol. 1998, 19, 160–166. [Google Scholar] [CrossRef]
- Malyutina, M. Russian deep-sea investigations of Antarctic fauna. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2004, 51, 1551–1570. [Google Scholar] [CrossRef]
- Linley, T.D.; Gerringer, M.E.; Yancey, P.H.; Drazen, J.C.; Weinstock, C.L.; Jamieson, A.J. Fishes of the hadal zone including new species, in situ observations and depth records of Liparidae. Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2016, 114, 99–110. [Google Scholar] [CrossRef]
- Leitner, A.B.; Neuheimer, A.B.; Donlon, E.; Smith, C.R.; Drazen, J.C. Environmental and bathymetric influences on abyssal bait-attending communities of the Clarion Clipperton Zone. Deep. Sea Res. Part I: Oceanogr. Res. Pap. 2017, 125, 65–80. [Google Scholar] [CrossRef]
- Riddle, M.J.; Craven, M.; Goldsworthy, P.M.; Carsey, F. A diverse benthic assemblage 100 km from open water under the Amery Ice Shelf, Antarctica. Paleoceanogr. 2007, 22. [Google Scholar] [CrossRef]
- Sumida P.Y.G., Bernardino A.F., Stedall V.P., Glover, A.G.; Smith, C.R. Temporal changes in benthic mega-faunal abundance and composition across the West Antarctic Peninsula shelf: Results from video surveys. Deep. Res Part II Top Stud. Oceanogr. 2008, 55, 2465–2477. [CrossRef]
- Eastman, J.T.; Amsler, M.O.; Aronson, R.B.; Thatje, S.; McClintock, J.B.; Vos, S.C.; Kaeli, J.W.; Singh, H.; La Mesa, M. Photographic survey of benthos provides insights into the Antarctic fish fauna from the Marguerite Bay slope and the Amundsen Sea. Antarct. Sci. 2012, 25, 31–43. [Google Scholar] [CrossRef]
- Ambroso, S.; Salazar, J.; Zapata-Guardiola, R.; Federwisch, L.; Richter, C.; Gili, J.M.; Teixidó, N. Pristine populations of habitat-forming gorgonian species on the Antarctic continental shelf. Sci. Rep. 2017, 7, 12251–12251. [Google Scholar] [CrossRef]
- Alberty, M.; Sprintall, J.; MacKinnon, J.; Germineaud, C.; Cravatte, S.; Ganachaud, A. Moored Observations of Transport in the Solomon Sea. J. Geophys. Res. Oceans 2019, 124, 8166–8192. [Google Scholar] [CrossRef]
- Langlois, T. , Williams, J., Monk, J., Bouchet, P.; Currey, L.; Goetze, J.; Harasti, D.; Huveneers, C.; Lerodiaconou, D.; Malcolm, H.; Whitmarsh, S. Marine sampling field manual for benthic stereo BRUVS (Baited Remote Underwater Videos). In Field Manuals for Marine Sampling to Monitor Australian Waters, Przeslawski R, Foster S (Eds). National Environmental Science Programme (NESP). 2018, 82–104.
- Linley, T.D.; Craig, J.; Jamieson, A.J.; Priede, I.G. Bathyal and abyssal demersal bait-attending fauna of the Eastern Mediterranean Sea. Mar. Biol. 2018, 165, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Brasier, M.J.; Grant, S.M.; Trathan, P.N.; Allcock, L.; Ashford, O.; Blagbrough, H.; Brandt, A.; Danis, B.; Downey, R.; Eléaume, M.P.; et al. Benthic biodiversity in the South Orkney Islands Southern Shelf Marine Protected Area. Biodiversity 2018, 1–15. [Google Scholar] [CrossRef]
- Smith, J.; O'Brien, P.E.; Stark, J.S.; Johnstone, G.J.; Riddle, M.J. Integrating multibeam sonar and underwater video data to map benthic habitats in an East Antarctic nearshore environment. Estuarine, Coast. Shelf Sci. 2015, 164, 520–536. [Google Scholar] [CrossRef]
- Post, A.L.; Lavoie, C.; Domack, E.W.; Leventer, A.; Shevenell, A.; Fraser, A.D. ; the NBP 14-02 SCIENCE TEAM Environmental drivers of benthic communities and habitat heterogeneity on an East Antarctic shelf. Antarct. Sci. 2016, 29, 17–32. [Google Scholar] [CrossRef]
- Jones, D.O.B. , Bett, B.J. and Tyler, P.A. Depth-related changes to 860 density, diversity and structure of benthic megafaunal assemblages 861 in the Fimbul ice shelf region, Weddell Sea, Antarctica. Polar Biol. 2007, 862 (30), 1579–1592.
- Gon, O. and Heemstra, P.C. Fishes of the Southern Ocean. J.L.B. Smith Institute of Ichthyology Grahamstown. 1990.
- Ciaputa, P. and Siciński, J. Seasonal and annual changes in Antarctic Fur seal (Arctocephalus gazella) diet in the area of Admiralty Bay, King George Island, South Shetland Islands. Pol. Polar Res. 2006, 171–184.
- Fujiwara, Y.; Tsuchida, S.; Kawato, M.; Masuda, K.; Sakaguchi, S.O.; Sado, T.; Miya, M.; Yoshida, T. Detection of the Largest Deep-Sea-Endemic Teleost Fish at Depths of Over 2,000 m Through a Combination of eDNA Metabarcoding and Baited Camera Observations. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Brandt, A.; Gutt, J.; Hildebrandt, M.; Pawlowski, J.; Schwendner, J.; Soltwedel, T.; Thomsen, L. Cutting the Umbilical: New Technological Perspectives in Benthic Deep-Sea Research. J. Mar. Sci. Eng. 2016, 4, 36. [Google Scholar] [CrossRef]
Figure 1.
Sampling station of the lander.
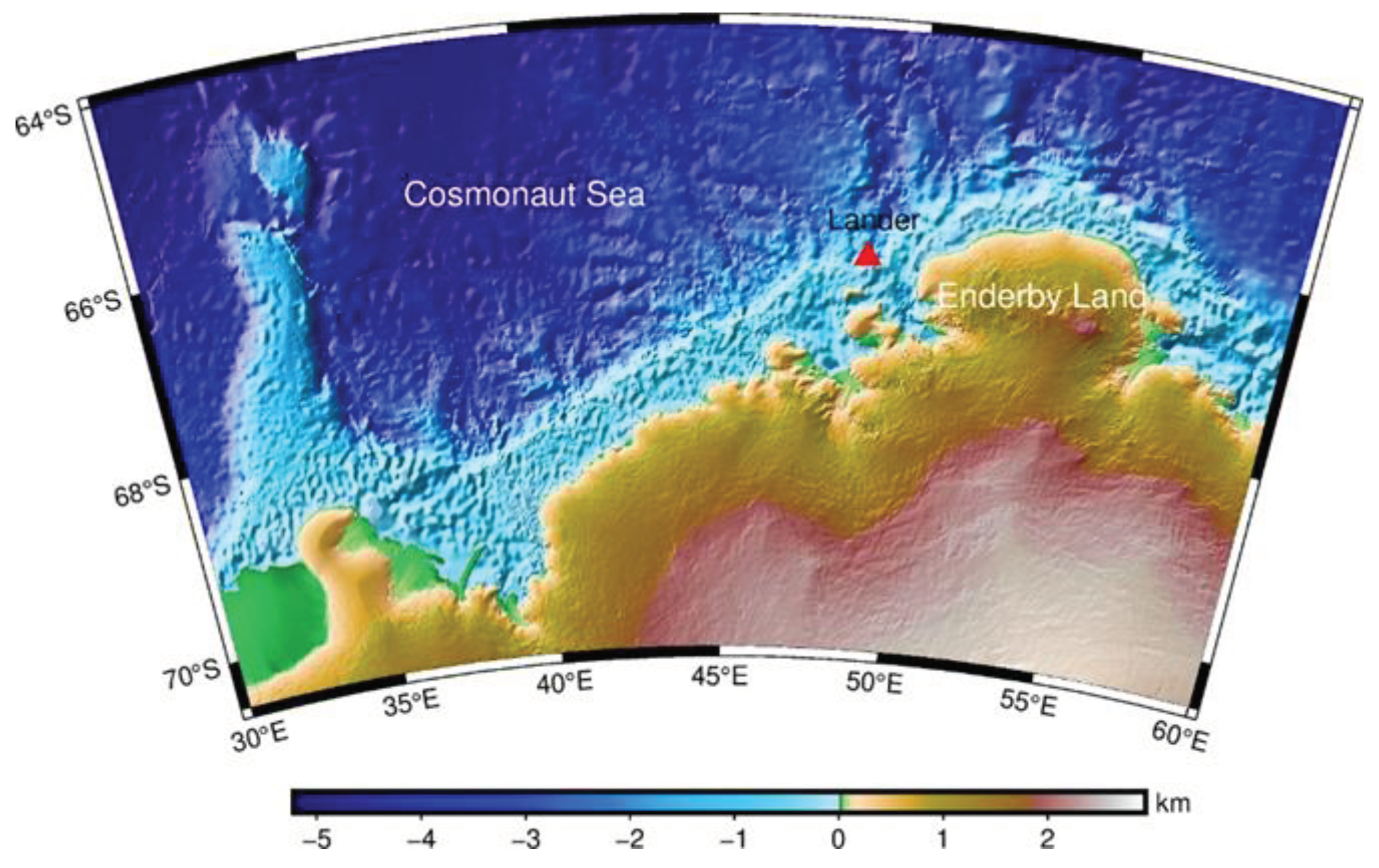
Figure 2.
Structure of the free-fall baited camera lander.

Figure 3.
Substrate type of the Cosmonaut sea shelf.

Figure 4.
Data collected by the SBE 37 recorder.
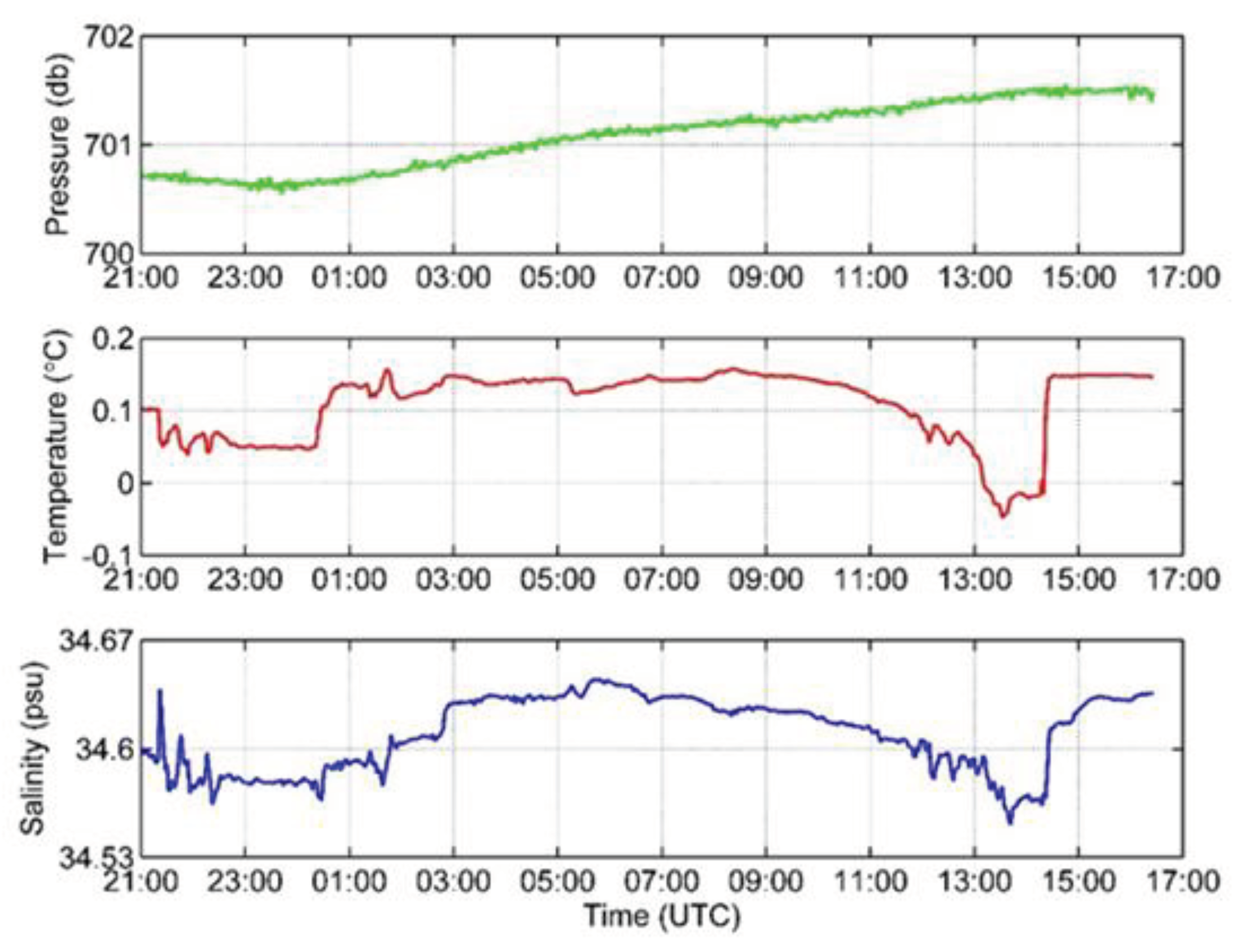
Figure 5.
Fish images from the shelf of Cosmonaut Sea.


Table 1.
Maximum number (MaxN) of individuals of each species.
| Phylum | Order | Family | Genus species | MaxN | VME |
|---|---|---|---|---|---|
| Porifera | Demospongiae | Demospongiae und. | 1 | ✔ | |
| Cnidaria | Scleralcyonacea | Umbellulidae | Umbellula carpenteri | 1 | ✔ |
| Mopseidae | Primnoisis sp. | 1 | ✔ | ||
| Actiniaria | Actinostolidae | Stomphia sp. | 2 | ✔ | |
| Scleralcyonacea | Scleralcyonacea und. | 3 | ✔ | ||
| Mollusca | Neogastropoda | Cancellariidae | Nothoadmete sp. | 4 | |
| Cochlespiridae | Aforia sp. | 1 | |||
| Elasipodida | Psychropotidae | Psychroteuthis glacialis | 1 | ||
| Arthropoda | Euphausiacea | Euphausiidae | Euphausia superba | 5 | |
| Decapoda | Thoridae | Lebbeus sp. | 1 | ||
| Nematocarcinidae | Nematocarcinus lanceopes | 1 | |||
| Isopoda | Cirolanidae | Natatolanameridionalis | 10 | ||
| Amphipoda | Uristidae | Abyssorchomene sp. | 13 (trap) | ||
| Echinodermata | Velatida | Pterasteridae | Pteraster sp. | 1 | |
| Paxillosida | Astropectinidae | Psilaster charcoti | 1 | ||
| Ophiurida | Ophiopyrgidae | Ophioplinthus sp. | 5 | ||
| Ophiacanthidae | Ophiacanthidae und. | 1 | |||
| Crinoidea | Crinoidea und. | 1 | ✔ | ||
| Bryozoa | Cyclostomatida | Horneridae | Horner sp. | 10 | ✔ |
| Cheilostomatida | Austroflustra vulgaris | 6 | ✔ | ||
| Cellariidae | Melicerita obliqua | 1 | ✔ | ||
| Brachiopoda | Terebratulida | Terebratulinae | Terebratulinae und. | 1 | |
| Chordata | Aplousobranchia | Polyclinidae | Polyclinidae und. | 2 | ✔ |
| Perciformes | Nototheniidae | Dissostichus mawsoni | 1 | ||
| Trematomus lepidorhinus | 1 | ||||
| Zoarcidae | Melanostigma gelatinosum | 2 | |||
| Artedidraconidae | Pogonophryne sp. | 1 | |||
| Channichthyidae | Chionobathyscus dewitti | 1 | |||
| Perciformes und. | 1 | ||||
| Myctophiformes | Myctophidae | Gymnoscopelus sp. | 1 | ||
| Aulopiformes | Paralepididae | Notolepis coatsi | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



