
Submitted:
22 March 2024
Posted:
25 March 2024
You are already at the latest version
Abstract
ProAKAP4, a precursor of AKAP4 (A-kinase anchor protein) found in the flagellum of mammalian and non-mammalian spermatozoa, serves as a structural protein with established correlations to motility parameters across diverse species. This study aimed to determine the proAKAP4 levels evolution in thawed stallion semen over a 3-hour period, examining its correlation with motility descriptors and mitochondrial membrane potential. Utilizing sixteen ejaculates from four French warmblood stallions, the study involved maintaining thawed samples at 37°C for 3 hours, conducting proAKAP4 enzyme-linked immunosorbent assay (ELISA), computer-assisted sperm analysis (CASA), and mitochondrial membrane potential by JC-1 probe and flow cytometry at 0, 1, and 3 hours post-thawing. The findings indicate significant positive correlations (p≤0.05) between proAKAP4 levels and sperm total or progressive motility at all time points analyzed. Spermatozoa velocity descriptors (VAP, VCL, VSL) and spermatozoa lateral head displacement (ALH) display positive correlations (p≤0.05) with ProAKAP4 at the 0-hour post-thawing. ProAKAP4 concentration exhibits no discernible difference between batches with or without cryoprotectant. Notably, proAKAP4 consumption remains insignificant within the initial hour after thawing but becomes significant (p≤0.05) between 1 and 3 hours post-thawing. In summary, proAKAP4 demonstrates positive correlations with total and progressive motility in stallion semen for up to 3 hours after thawing, albeit showing a noticeable decrease starting from the first hour post-thawing, indicating a progressive consumption as a result of spermatozoa motile activity.
Keywords:
ProAKAP4
; AKAP4
; stallion
; spermatozoa
; motility
; mitochondrial activity.
1. Introduction
The cryopreservation of sperm over extended periods is pivotal for the survival, maintenance, and enhancement of genetic diversity and improvement within species and breeds worldwide [1]. In the equine field, semen cryopreservation has been important for different reasons like spreading good genetics worldwide, minimizing disease transmission, getting offspring after the death of the stallion [2] or keeping rare genetic lines from endangered breeds [1].
The most important point about frozen-thawed semen is to maintain the capacity of fertilization after thawing. One of the major impediments to the development of the horse frozen semen industry is the lower fertility with frozen semen versus to cooled semen [3]. However, Lomis (2001) [3] concluded that acceptable fertility with frozen semen is possible with good quality semen. A good quality semen once thawed will reach the fertilization site to penetrate the oocyte [1]. Thawed semen is considered good when its progressive motility is over 35%, [4] but this is not a warranty that the semen will be fertile [4,5]. The definition of semen fertility is not easy and precise [5], it is usually related to the seminal quality parameters [4,6]. The estimate of semen fertility has been always a challenge, frozen semen appears to have shorter longevity once thawed [2] and lower conception rates [7,8]. Insemination of the mare with frozen semen is a challenge due to the short lifetime of the spermatozoa once thawed. It is also difficult to predict how long the semen will be alive and motile after thawing to adapt to the ovarian scanning routine of the mare [8] for artificial insemination with frozen-thawed semen. The most commonly employed protocol to inseminate a mare with frozen semen to obtain a high pregnancy rate, the mare has to be inseminated within a 6-hour window around ovulation [2,9]. To reach this objective the mare has to be scanned regularly to catch the ovulation time.
The traditional semen analysis (sperm concentration, subjective motility and sperm morphoabnormalities) has a reduced predictive value in the evaluation of stallion fertility. An important number of sperm tests (e.g., sperm head morphometry, the hypoosmotic swelling test, acrosome integrity, membrane integrity, progesterone receptor exposure, DNA integrity or fragmentation, intracellular Calcium, intracellular ROS, mitochondrial activity…) have been developed in order to increase the sperm fertility prediction. However, most examine only a single or a narrow range of the attributes that a sperm must possess. Flow cytometry offers the possibility of quickly and simultaneously combining many of these analyzes [6]. On the other hand, the CASA-Mot (Computer Assisted Semen Analysis for Motility) can be used to rapidly assess total and progressive motility, as well as determine values for other kinematic variables [10]. Nowadays, the prediction of seminal quality has been usually related to motility parameters analyzed by CASA systems, flow cytometry analysis and formulas combining both [1]. However, semen quality is not always correlated with good fertility rates [11]. In a practical point of view, the search for new fertility predictors is an important field of research.
ProAKAP4 is a marker of sperm quality and male fertility in different species [12], also in stallions [13,14,15,16,17]. ProAKAP4 is the precursor of an A-kinase anchor protein 4 which is the main protein of the fibrous sheath of the principal piece of the flagellum [12,18]. As an important part of flagellum [19,20], proAKAP4 is a good marker of post-meiotic stage of spermatogenesis [12,21,22] as well as, a good marker of sperm progressive and total motility in stallion [13,14,15,16] and in other mammals like bulls [23], jackass [24], dogs [25], rabbits [26], rams [27] and dromedary [28].
This study aims to correlate motility descriptors and mitochondrial membrane potential with proAKAP4 concentration in stallion frozen-thawed semen and explore its evolution over three hours to assess the potential of proAKAP4 as a long-lasting motility indicator. Another aim of this study is to test how cryoprotectant affects proAKAP4 evolution.
2. Materials and Methods
2.1. Semen Samples
Sixteen ejaculates obtained from four French warmblood stallions, housed at Haras de Hus in Petit Mars, France, were utilized for this study, intended for both reproductive and sport (showjumping and dressage) purposes. The stallions ranged in age from 3 to 8 years at the time of semen collection. These animals were selected based on their proven fertility, excellent physical condition, and negative testing for Equine Arteritis, Infectious Anemia, and Contagious Metritis, conforming to CEE health requirements prior to semen production. Each stallion provided four ejaculates, from which three straws were extracted for further analysis.
Cryopreservation of all ejaculates was conducted between 2012 and 2016 at Haras de Hus, a collection center approved by European authorities. Consistent cryopreservation protocols were followed, employing INRAFreeze® as an extender and MiniDigitcool® (IMV Technologies; France) as the automatic freezer.
2.2. Experimental Groups
Three straws for each ejaculate were thawed at 37°C in a water bath for 50 seconds. Semen samples were deposited in an Eppendorf Tube containing 500µL of INRA96® previously warmed at 37°C. The resulting mix was gently shaken split into two aliquots. One aliquot was centrifuged at 600xg for 10 minutes to remove the cryoprotectant, the other one was not centrifuged. After centrifugation, the pellet was resuspended with 500µL of INRA96® pre-warmed at 37°C. Then both aliquots were kept inside an incubator at 37°C. The analyses were done at 0 hours, 1 hour and 3 hours after thawing.
Semen analysis encompassed the evaluation of kinematic parameters utilizing Computer-Assisted Semen Analysis (CASA) (section 2.3), determination of proAKAP4 concentrations (section 2.4), and assessment of mitochondrial membrane potential using JC-1 stain (section 2.5). The semen concentration of each sample was calculated employing CASA methodology.
2.3. Motility Analysis
Sperm motility assessment was conducted utilizing a Computer-Assisted Sperm Analysis (CASA) system (IVOS II ®; Hamilton Thorne; IMV Technologies; France). Post-thawing, all samples were maintained at 37 °C in a water bath for the initial minute, followed by transfer to a 37°C incubator for the subsequent 3 hours to sustain optimal temperature conditions for analysis. For examination, 3 μL of sperm samples were dispensed onto a prewarmed Leja® slide at 37 °C. Three distinct fields were examined to assess various motility descriptors including total motility (TM, %), progressive motility (PM, %), curvilinear velocity (VCL, μm/s), straight-line velocity (VSL, μm/s), average path velocity (VAP, μm/s), linearity (LIN, %), straightness (STR, %), amplitude of lateral head displacement (ALH, μm), and frequency of head displacement (BCF, Hz).
The CASA parameters were set as follows: magnification of x200 with 10 randomly selected fields; acquisition of 60 images per second; capture of 30 sequences in each analysis; particle area set to 3 pixels; connectivity threshold set at 6; minimum number of images required to calculate ALH set at 10. The cut-off value for motile spermatozoa was established as VAP ≥20 µm/s, while progressive motility was defined as STR ≥80% and VAP ≥30 µm/s.
2.4. ProAKAP4 ELISA
ProAKAP4 quantification was performed using a commercially available quantitative enzyme-linked immunosorbent assay (ELISA). The analysis was conducted at the ONIRIS Laboratory in Nantes, France, utilizing the Horse 4MID® kit (SPQI, Lille, France). This kit employs a quantitative ELISA sandwich assay designed to specifically detect and quantify proAKAP4 in equine semen samples. A reference curve was generated using a provided seven-point standard solution to determine the concentration of proAKAP4 present in each stallion's thawed semen sample.
Sample preparation followed the manufacturer's instructions for cryopreserved stallion semen. Forty microliters of diluted semen from each experimental group described in section 2.2 were mixed with 160 µl of specific lysis buffer included in the kit. After vigorous vortex mixing for one minute, 200 µL of dilution buffer was added and rapidly mixed before loading into the 96-well coated plate. The plate was then incubated for 2 hours under gentle agitation with an ELISA-plate horizontal shaker.
Following three successive washing steps, horseradish-conjugated detection antibody was added to each well and further incubated for 1 hour. Subsequently, TMB substrate was added, initiating a color reaction after 10 minutes of incubation. The intensity of the color reaction, measured spectrophotometrically with a 450 nm filter, is quantitatively proportional to the amount of proAKAP4 present in each sample. The color reaction was halted by adding a stop solution to each well, and color intensities were measured using a spectrophotometer.
Optical densities were then used to determine proAKAP4 concentration with the calculation sheet provided by the manufacturer (SPQI, Lille, France). Specifically, proAKAP4 concentration was expressed in ng per 10 million spermatozoa. This concentration was calculated using the formula: proAKAP4 ng/10 million spermatozoa = (proAKAP4 in ng/mL / spermatozoa M/mL) x 10 x 16 x (2/3). The 10x factor accounts for the final concentration in 10 million spermatozoa, while the 10x dilution factor corresponds to the post-thawed sample dilution during sample preparation. The 2/3 factor corresponds to the dilution volume loaded in each well of the 96-well plate in the Horse 4MID® assay.
2.5. Mitochondrial Membrane Potential
Mitochondrial membrane potential measurement was conducted utilizing the Guava® Easycyte II™ flow cytometer (Guava Technologies Inc., Hayward, CA, USA, distributed by IMV Technologies, France) and EasyKit™2 kit 37 of ready-to-use reagents (IMV Technologies, France). The EasyKit™2 comprises a 96-well plate containing the JC-1 fluorochrome, which is specific for mitochondrial membrane potential.
At each time point (0, 1, and 3 hours), 190 μL of EasyBuffer® (IMV Technologies, France), 10 μL of DMSO, and 1.75 μL of diluted semen were deposited in each well. Subsequently, the plate was incubated in darkness for 30 minutes at 37°C. Analysis of the wells was performed using the Guava® Easycyte II™ flow cytometer, calibrated beforehand with the Easy Check® kit (Guava Technologies Inc., MA, USA), and the Express Pro® software (IMV, France).
The EasyKit™2 fluorochrome emits maximum fluorescence at 525 nm (green) and 596 nm (red). For each well, 5,000 spermatozoa were analyzed. The JC-1 fluorescent stain serves as a marker of the intermediate piece of spermatozoa with high membrane potential and low membrane potential mitochondria. Polarized, active mitochondria are colored orange, while depolarized, inactive mitochondria are colored green, as per manufacturer-defined parameters.
Results are expressed as a percentage of "polarized" mitochondria (strongly or partially) and as a percentage of "depolarized" mitochondria. In our investigation, mitochondrial membrane potential is quantified by the percentage of "polarized" mitochondria, referred to as "active" mitochondria.
2.6. Statistical Analysis
The statistical analysis commenced with the assessment of data normality and homoscedasticity using the Shapiro-Wilk and Levene tests, respectively. In instances where data deviated from normal distribution, the arcsin √x transformation was employed to fulfill parametric assumptions. Subsequently, a mixed generalized linear model of repeated measures was applied to compare proAKAP4 concentration, total and progressive motility, and mitochondrial membrane potential of equine sperm across different incubation times (0 h, 1 h, and 3 h). Treatment (with cryoprotectant and without cryoprotectant) was considered as the fixed-effects factor, while individual horses were treated as random-effects factors. Furthermore, Pearson's correlation analysis was conducted to assess the correlation coefficients (r) between ProAKAP4 concentration and sperm motility parameters, as well as mitochondrial membrane potential, within each treatment and at each time point.
Statistical analyses were conducted using the R statistical package (V 4.0.3, R Core Team; Vienna, Austria), while graphical representations were generated using GraphPad Prism software (V 8.4.0, GraphPad Software LLC; San Diego, CA, USA). The significance level was set at p < 0.05 for all analyses, and results are presented as mean ± standard error of the mean (SEM). Analyzes of the experimental data were performed with the statistical package R (V 4.0.3, R Core Team; Vienna, Austria), while the graphs were elaborated with the GraphPad Prism software (V 8.4.0, GraphPad Software LLC; San Diego, CA, USA). The minimum level of statistical significance was set at p < 0.05 for all analyses. Results are expressed as mean ± standard error of the mean (SEM).
3. Results
3.1. Correlation between proAKAP4, Motility Descriptors and Mitochondrial Membrane Potential
Total and progressive motility showed a positive correlation with proAKAP4 concentration at 0, 1 and 3 hours. However, considering the different descriptors of sperm motility, only those that refer to speed (VCL, VSL, VAP) and the sperm Lateral Head Displacement (ALH) evidenced a positive correlation with proAKAP4 levels at 0 hours (Table 1).
3.2. Evolution of proAKAP4, Motility Descriptors and Mitochondrial Membrane Potential over 3 Hours
No significant differences on proAKAP4 concentration were observed between the batch of samples with and without cryoprotectant at 0 and 1hours of incubation. However, regardless of cryoprotectant removal a significant decrease (P<0.05) of proAKAP4 levels was observed at 3 hours after thawing. The removal of the cryoprotectant had no effect on the concentration of proAKAP4 at 0 and 1h, but showed a significant decrease (P<0.05) at 3h when the cryoprotectant was eliminated (Figure 1).
The evolution of total motility (MT) and progressive motility (MP) in percentages is shown in Figure 2A,B. In terms of total motility (MT), there was no difference between samples with and without cryoprotectant and between incubation time. Regarding progressive motility (MP), no significant differences were observed between incubation time for booth type of samples, with or without cryoprotectant. Nevertheless, samples with cryoprotectant showed significantly lower progressive motility at all incubation times analyzed.
Analyzing the mitochondrial membrane potential, no significant difference was observed between samples (with or without cryoprotectant) at different incubation times. However, in both cases, removing or not the cryoprotectant, a significant (P<0.05) decrease between 0 and 1 hour of incubation was observed but not between 1 and 3 hours (Figure 3).
4. Discussion
The results of this experiment confirm a positive correlation between proAKAP4 concentration and total and progressive motility in a stallion frozen/thawed semen at any time of our study. This positive correlation has been found previously in fresh semen in stallions [13,14,16] and in other mammals like bulls [23,29,30], jackass [24], dromedaries [28] rams [27], bucks [31], pigs [32], dogs [25], mice [33,34], and even in humans [35,36]. In the present study, this correlation between proAKAP4 and motility parameters is sustained during the 3 hours post thaw in contrast with the same experiment in donkeys [24] where this correlation between motility descriptors and proAKAP4 concentration was only described at time 0 and 1 hour after thawing but not at 3 hours. Horse and donkey, although phylogenetically close, are different species with their own reproductive strategies. All kinematic parameters for donkeys are significantly higher than for horse. Donkey sperm is more rapid and linear than horse one and, possibly, the effect of proAKAP4 on the motility patterns is also different between booth species [37]. Nevertheless, obtained results confirm proAKAP4 as a good marker of sperm quality and longevity; kinematic parameters and longevity are both important for the fertilizing ability of frozen thawed spermatozoa [39,40].
However, analyzing the descriptors of the sperm trajectory only the velocity parameters (VCL, VCL and VAP) and the lateral head displacement (ALH), and only at 1 h of post thaw incubation but not later, correlate positively with proAKAP4 concentration. This correlation has been previously found in stallions [13] and in bulls [20,26,27,34]. In other mammals, like bulls, progressive motility [37] and velocity descriptors [38,39] have been described as sperm quality and even fertility markers. It seems evident that proAKAP4 has a relationship with the sperm motile ability but not an effect on the kind of movement, or trajectory description, in stallion. Further studies are needed in order to evaluate the effect on frozen/thawed semen fertility
ProAKAP4 as a reservoir of motility and energy is described in several articles and drawn as a "backpack" with the energy of spermatozoa [12,18,32]. ProAKAP4 does not significantly decrease in the first hour but diminishes between one and three hours; in this case, there is no significant difference between the groups containing or not cryoprotectants at 0 and 1h. However, at 3h the samples without cryoprotectant showed a significant decrease in the proAKAP4 concentration. On the other hand, removing the cryoprotectant higher levels of Progressive Motility were observed, indicating higher consumption of this energy “backpack”.
Total motility showed no significant difference between the centrifuged group (removing the cryoprotectant) and the non-centrifuged group (maintaining the cryoprotectant) and thus there is no non-significant difference between the 3 times studied. Centrifugation at 600G during 10 minutes has not affect significantly total motility and viability parameters [44]. However, removing the cryoprotectant the Progressive Motility evidenced higher values at all analyzed times. This may indicate a deleterious effect of the cryoprotectant on sperm motility and, as explained before, higher proAKAP4 consumption when the Progressive Motility is higher.
Comparing proAKAP4 with total and progressive motility, proAKAP4 holds well the first hour because it maintains motility and then suddenly decreases, but the motility has decreased slowly throughout the period. Comparing with data in donkeys [21], where our team carried out the same experiment, the results are very similar and comparable. It can be concluded that proAKAP4 is a structural protein that functions as the potential motility reserve of the equines spermatozoon [9] so by measuring the concentration it is probably possible to predict whether post-thaw motility will be constant over some time.
Longevity of post-thawed spermatozoa can be affected depending on the extender in different species. In deer, sperm longevity at four hours post-thawed is different depending on the cryoprotectant used [45]. In carp, supplement the extender with seminal plasma transferrin improves cryopreservation survival and longevity of the frozen-thawed semen [46]. In the present study, the used extender is an egg yolk-based extender (INRA Freeze ®). Egg yolk-based extenders are beneficial for semen preservation and post-thaw survival in tom cats [47] and significantly improve sperm thaw quality and longevity in African wild dog [48]. In stallion, cryoprotectants agents like phosphodiesterase inhibitors, caffeine, or taurine increase motility and longevity in post-thaw conditions [8].
No significant differences in pro AKAP4 concentration was observed in stallions between batches with or without cryoprotectant; however, in donkey [24], a significant difference in proAKAP4 concentration has been found between the samples with or without cryoprotectant. This difference can be explained because a different cryoprotectant was used in both experiments. Blommaert et al., [15] showed that proAKAP4 concentration differs significantly depending on the extender used in stallion thawed semen. Kowalczyk et al., [38] showed how the addition of Holothuroidea extract in the semen extender modulates positively the proAKAP4 concentration in the frozen/thawed bovine semen. In the present study, INRAFreeze ® was used to freeze all the ejaculates and in the case of donkey [24] BotuCrio ® was the election. Observed differences between donkey and horse can be explained because a different cryoprotectant has been used in both experiments; furthermore, other components could have an impact and there could be species-specific sensitivities, as explained before. Blommaert et al., [15] have shown that proAKAP4 concentration differs significantly depending on the extender used in stallion thawed semen. Further analysis comparing these 2 extenders (BotuCrio ® and INRAFreeze ®) is needed to elucidate if they exert the same effect on proAKAP4 or not. There are two hypotheses about these differences between extenders one is that a toxic effect might degrade proAKAP4 directly, and second, one extender might converse proAKAP4 more rapidly to AKAP4 than the other, dependent on the extender composition.
For mitochondrial membrane potential, using the JC1 stain, there is no significant difference between groups containing cryoprotectants or not, but the change over time is contrary to proAKAP4, decreasing rapidly between 0 and 1 hour only. The absence of a relationship between the mitochondrial membrane potential and proAKAP4 levels suggests independence between spermatozoa energy production and the transmission of motion. This can explain why in previous reports no direct relationship was observed between mitochondrial activity stimulation and spermatozoa motility increase [49,50].
5. Conclusions
In conclusion, proAKAP4 is a good marker of sperm quality correlating positively with total and progressive motility at any time in our study. It also correlates with velocity kinematic parameters at the moment of thawing. ProAKAP4 can be considered a long-lasting motility marker because it reduces at the time when motility parameters have decreased. Removing the cryoprotectant from frozen/thawed horse semen shows better maintenance of progressive sperm motility at 3h and lower proAKAP4 concentration. Greater progressive motility means more proAKAP4 consumption.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, JF.B. and L.B-A.; methodology, M.D-P., N.S, J.M. and L.B-A, .; software, M.D-P and I Y-O.; validation, M.D-P, I.Y-O, L.B-A and J.M.; formal analysis, M.D-P., I.Y-O and J.C.; investigation, M.D-P, V.M., N.S. and L.B-A.; resources, L.B-A and J.M.; data curation, M.D-P, N.S. and I.Y-O.; writing—original draft preparation, M.D..; writing—review and editing, J.C., J.M. and L.B-A.; visualization, M.D-P and J.M.; supervision, JF.B, J.M. and L.B-A.; project administration, L.B-A.; funding acquisition, J.M. and L.B-A. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by IFCE (Institut Français du Cheval et l’Equitation), grant number (CS-2021-014-PROAKAP-SPZ-qualité).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Obtained data are included in the article.
Acknowledgments
Thanks to “Haras de Hus” and especially Laure Normandin, for providing all the straws used in this research. Thanks to IMV technologies for the material provided. The present work was supported by Oniris, INSERM, and the French Ministry of Higher Education and Research. This research did not receive any commercial or not-for-profit sector support. Thanks to IFCE (Institut Français du Cheval et de l’Equitation) for the funding and supporting this project (CS-2021-014-PROAKAP-SPZ-qualité).
Conflicts of Interest
Nicolas Sergeant is cofounders of Spqi.
References
- Watson:, P.F. The causes of reduced fertility with cryopreserved semen. Animal Reproduction Science 2000, 60–61, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.D. Optimizing the use of frozen–thawed equine semen. Theriogenology 2008, 70, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Loomis PR, The equine frozen semen industry. Animal Reproduction Science 2001, 68, 191–200. [CrossRef] [PubMed]
- Barrier Battut, I.; Kempfer, A.; Lemasson, N.; Chevrier, L.; Camugli, S. Prediction of the fertility of stallion frozen-thawed semen using a combination of computer-assisted motility analysis microscopical observation and flow cytometry. Theriogenology 2017, 97, 186–200. [Google Scholar] [CrossRef] [PubMed]
- Kuisma, P.; Andersson, M.; Koskinen, E.; Katila, T. Fertility of frozen-thawed stallion semen cannot be predicted by the currently used laboratory methods. Acta Veterinaria Scandinavica 2006, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Colenbrander, B.; Gadella, B.; Stout, T. The Predictive Value of Semen Analysis in the Evaluation of Stallion Fertility. Reproduction in Domestic Animals 2003, 38, 305–311. [Google Scholar] [CrossRef]
- Vidament, M.; Dupere, A.M.; Julienne, P.; Evian, A.; Noue, P.; Palmer, E. Equine frozen semen: Freezability and fertility field results. Theriogenology 1997, 48, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Stephens, T.D.; Brooks, R.M.; Carrington, J.L.; Cheng, L.; Carrington, A.C.; Porr, C.A.; Splan, R.K. Effects of Pentoxifylline, Caffeine, and Taurine on Post-Thaw Motility and Longevity of Equine Frozen Semen. Journal of Equine Veterinary Science. 2013, 33, 615–621. [Google Scholar] [CrossRef]
- Sanchez, R.; Herrera, C.; Blanco, M.; Rosati, I.; Vlek, J.; Sieme, H. Effect of the dose using equine frozen semen and deep horn insemination in a large scale commercial equine program. Journal of Equine Veterinary Science 2016, 43, S78. [Google Scholar] [CrossRef]
- Giaretta, E.,; Munerato, M., Yeste, M., Galeati, G., Spinaci, M., Tamanini, C., Mari, G., Bucci, D. Implementing an open-access CASA software for the assessment of stallion sperm motility: Relationship with other sperm quality parameters. Anim. Reprod. Sci. 2017, 176, 11–19. [CrossRef]
- Kowalczyk, A.; Czerniawska-Piatkowska, E.; Kuczaj, M. Factors influencing the popularity of artificial insemination of mares in Europe. Animals 2019, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Carracedo, S.; Briand-Amirat, L.; Dordas-Perpinyà, M.; Ramos Escuredo, Y.; Delcombel, R.; Sergeant, N.; Delehedde, M. ProAKAP4 protein marker: Towards a functional approach to male fertility. Animal Reproduction Science 2022, 247, 107074. [Google Scholar] [CrossRef] [PubMed]
- Blommaert, D.; Sergeant, N.; Delehedde, M.; Jouy, N.; Mitchell, V.; Franck, T.; Donnay, I.; Lejeune, J.P.; Serteyn, D. Expression, localization, and concentration of A-kinase anchor protein 4 (AKAP4) and its precursor (proAKAP4) in equine semen: Promising marker correlated to the total and progressive motility in thawed spermatozoa. Theriogenology 2019, 131, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Griffin, R.A.; Swegen, A.; Baker, M.; Aitken, R.J.; Skerrett-Byrne, D.A.; Silva Rodriguez, A.; Martín-Cano, F.E.; Nixon, B.; Peña, F.J.; Delehedde, M.; Sergeant, N.; Gibb, Z. Mass spectrometry reveals distinct proteomic profiles in high- and low-quality stallion spermatozoa. Reproduction 2020, 160, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Blommaert, D.; Sergeant, N.; Delehedde, M.; Donnay, I.; Lejeune, J.P.; Franck, T.; Serteyn, D. First results about ProAKAP4 concentration in stallion semen after cryopreservation in two different freezing media. Cryobiology 2021, 102, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Dordas-Perpinyà, M.; Yanez-Ortiz, I.; Sergeant, N.; Mevel, V.; Bruyas, J.F.; Catalán, J.; Delehedde, M.; Briand-Amirat, L.; Miró, J. ProAKAP4 Concentration Is Related to Sperm Motility and Motile Sperm Subpopulations in Frozen–Thawed Horse Semen. Animals 2022, 12, 3417. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.Z.Y.; Dilger, A.; Bryan, E.; Bytnar, G.; Choi, S.; Cook, K.; Kline, K. Preliminary Study of Effects of Centrifugation of Stallion Semen on Motility and Pro-AKAP4 Concentration over 72 Hours Post-collection. Journal of Agricultural Science and Technology 2022, 12, 65–72. [Google Scholar]
- Delehedde, M.; Carracedo, S.; Selleslagh, M.; Eddarkaoui, S.; Amirat-Briand, L.; Sergeant, N. ProAKAP4 polypeptide as a biomarker of sperm functionality and male fertility disorders. Int J Gynecol and Reprod Sci 2019, 2, 13–19. [Google Scholar]
- Johnson, L.R.; Foster, J.A.; Haig-Ladewig, L.; Vanscoy, H.; Rubin, C.S.; Moss, S.B.; Gerton, G.L. Assembly of AKAP82, a protein kinase A anchor protein, into the fibrous sheath of mouse sperm. Developmental biology 1997, 192, 340–350. [Google Scholar] [CrossRef]
- Turner, R.M.; Johnson, L.R.; Haig-Ladewig, L.; Gerton, G.L.; Moss, S.B. An X-linked gene encodes a major human sperm fibrous sheath protein, hAKAP82: Genomic organization, protein kinase A-RII binding, and distribution of the precursor in the sperm tail. Journal of Biological Chemistry 1998, 273, 32135–32141. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, H.; Pask, A.J.; O'Brien, D.A.; Shaw, G.; Renfree, M.B. A-kinase anchoring protein 4 has a conserved role in mammalian spermatogenesis. Reproduction 2009, 137, 645. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Bernstein, I.R.; Cafe, S.L.; Delehedde, M.; Sergeant, N.; Anderson, A.L.; Bromfield, E.G. A kinase anchor protein 4 is vulnerable to oxidative adduction in male germ cells. Frontiers in Cell and Developmental Biology 2019, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Dordas-Perpinyà, M.; Sergeant, N.; Ruelle, I.; Bruyas, J.F.; Charreaux, F.; Michaud, S.; Carracedo, S.; Catalán, J.; Miró, J.; Delehedde, M.; Briand-Amirat, L. ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study. Vet. Sci 2022, 9, 224. [Google Scholar] [CrossRef] [PubMed]
- Dordas-Perpinyà, M.; Sergeant, N. , Yánez-Ortiz, I., Mevel, V., Catalán, J., Bruyas, J.F., Briand-Amirat, L., Miro, J. ProAKAP4 as a motility long-lasting marker in Catalan donkey spermatozoa. Anim Reprod Sci 2024, 262, 107427. [Google Scholar] [CrossRef] [PubMed]
- Le Couazer, D.; Delehedde, M.; Ruelle, I.; Sergeant, N.; Michaud, S.; Briand, L.; Bencharif, D. ProAKAP4 as a valuable marker to assess sperm quality in dogs. Reprod. Domest. Anim 2019, 54, 9192. [Google Scholar]
- Gardela, J.; Ruiz-Conca, M.; Palomares, A.; Olvera-Maneu, S.; García-Calvo, L.; López-Béjar, M.; Martínez-Pastor, F.; Álvarez-Rodríguez, M. Effect of Honey, Coenzyme Q10, and-Carotene-Tocopherol as Novel Additives in Rabbit-Sperm Cryopreservation Extender. Animals 2023, 13, 2392. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.; Anel-Lopez, L.; Neila-Montero, M.; Palacin-Martinez, C.; Montes-Garrido, R.; Alvarez, M.; de Paz, P.; Anel, L. ProAKAP4 as Novel Molecular Marker of Sperm Quality in Ram: An Integrative Study in Fresh, Cooled and Cryopreserved Sperm. Biomolecules 2020, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Malo, C.; Carracedo, S.; Delehedde, M.; Sergeant, N.; Skidmore, L. Identification of proAKAP4 in Dromedary sperm and their correlation with monthly semen parameters. Reprod. Fertil 2021, 2, 268–279. [Google Scholar] [CrossRef]
- Bastan, I.; Akcay, E. Quality assessment of frozen bull semen with the precursor A-kinase anchor protein 4 biomarker. Andrologia 2021, 53, e14164. [Google Scholar] [CrossRef]
- Marques de Almeida, A.B.; Hidalgo, M.M.T.; de Moraes, F.L.Z.; Trautwein, L.G.C.; de Fátima Schnitzer, J.; dos Santos Silva, L.A.; Rizzoto, G.; Pinheiro Ferreira, J.C.; Mello Martins, M.I. The proAKAP4 concentrations in Nelore bull sperm and their relation to FTAI conception rate results. Anim. Reprod. Sci. 2022, 247, 107156. [Google Scholar] [CrossRef]
- Fatet, A.; Sergeant, N.; Dordas-Perpinyà, M.; Drouet, B.; Ponthoreau, O.; Carracedo, S.; Bruyas, J.F.; Thorin, C.; Dlehedde, M.; Briand-Amirat, L. Sperm-specific protein proAKAP4 as a marker to evaluate sperm quality and fertility of Alpine and Saanen Bucks. Proceedings of the 25th Annual Conference of the European Society for Domestic Animal Reproduction (ESDAR). Reprod. Domest. Anim 2022, 57, 79–79. [Google Scholar]
- Sergeant, N.; Briand-Amirat, L.; Bencharif, D.; Delehedde, M. The Sperm Specific Protein Proakap4 as an Innovative Marker to Evaluate Sperm Quality and Fertility. J. Dairy Vet. Sci. 2019, 11, 555803. [Google Scholar] [CrossRef]
- Boersma, A.; Primus, J.; Wagner, B.; Broukal, V.; Andersen, L.; Pachner, B.; Dahlhoff, M.; Rülicke, T.; Auer, K.E. Influence of sperm cryopreservation on sperm motility and proAKAP4 concentration in mice. Reprod Med Biol 2022, 21, e12480. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.Y.; Chandanee, M.R.; Dissanayake, W.M.N.; Lee, S.M.; Yi, Y.J. Comparison of fertility competence in toll-like receptor 4 (TLR4)-knock out male mice fed a high-fat diet. J Biomed Transl Res 2023, 24, 41–52. [Google Scholar] [CrossRef]
- Sigala, J. Qualité du protéome du spermatozoïde humain et infertilité (Doctoral thesis) 2016, Université du Droit et de la Santé-Lille II, France. http://www.theses.fr/2016LIL2S039/document.
- Jumeau, F.; Sigala, J.; Dossou-Gbete, F.; Frimat, K.; Barbotin, A.L.; Buée, L.; Béhal, H.; Sergeant, N.; Mitchell, V. A-kinase anchor protein 4 precursors (pro-AKAP4) in human sperm. Andrology 2018, 6, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Gacem S, Valverde A, Catalan J, Yánez-Ortiz I, Soler C, Miró J. A new approach of sperm motility subpopulation structure in donkey and horse. Frontiers in Veterinary Science 2021, 8, 651477. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Gałeska, E.; Bubel, A. The concentration of proAKAP4 and other indicators of cryopotential of spermatozoa cryopreserved in extender with holothuroidea extract addition. Animals 2022, 12, 521. [Google Scholar] [CrossRef]
- Love, C.C. Relationship between sperm motility, morphology and the fertility of stallions. Theriogenology 2011, 76, 547–557. [Google Scholar] [CrossRef]
- Johannisson, A.; Cojkić, A.; Morrell, J.M. The relationship between sperm quality parameters and stallion fertility. Journal of Equine Veterinary Science 2023, 125, 10459. [Google Scholar] [CrossRef]
- Puglisi, R.; Pozzi, A.; Foglio, L.; Spanò, M.; Grollino, M.G.; Bongioni, G. The usefulness of combining traditional sperm assessments with in vitro heterospermic insemination to identify bulls of low fertility as estimated in vivo. Anim. Reprod. Sci 2012, 132, 17–28. [Google Scholar] [CrossRef]
- Farrell, P.B.; Presicce, G.A.; Brockett, C.C.; Foote, R.H. Quantification of bull sperm characteristics measured by computer-assisted sperm analysis (CASA) and the relationship to fertility. Theriogenology 1998, 49, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Á.; Polichronopoulos, T.; Gáspárdy, A.; Solti, L. Correlation between bull fertility and sperm cell velocity parameters generated by computer-assisted semen analysis. Acta Vet. Hung 2015, 63, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Len, J.A.; Jenkins, J.A.; Eilts, B.E.; Paccamonti, D.L.; Lyle, S.K.; Hosgood, G. Centrifugation has minimal effects on motility, viability, and acrosome integrity of equine sperm. Theriogenology 2008, 70, 582–583. [Google Scholar] [CrossRef]
- Cheng, F-P. ; Wu, J-T.; Chan, J-P.; Wang, J-S.; Fung, H-P.; Colenbrander, B.; Tung, K-C. The effect of different extenders on post-thaw sperm survival, acrosomal integrity and longevity in cryopreserved semen of Formosan Sika deer and Formosan Sambar deer. Theriogenology 2004, 61, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Shaliutina-Kolešová, A.; Dietrich, M.; Xian, M.; Nian, R. Seminal plasma transferrin effects on cryopreserved common carp Cyprinus carpio sperm and comparison with bovine serum albumin and antifreeze proteins. Animal Reproduction Science. 2019, 204, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, U.; Axnér, E. Epididymal and ejaculated cat spermatozoa are resistant to cold shock but egg yolk promotes sperm longevity during cold storage at 4°C. Theriogenology 2007, 67, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Van den Berghe, F.; Paris, M.; Brent Briggs, M.; Farstad, W.K.; Paris, D. A two-step dilution tris-egg yolk extender containing Equex STM significantly improves sperm cryopreservation in the African wild dog (Lycaon pictus). Cryobiology 2018, 80, 18–25. [Google Scholar] [CrossRef]
- Catalán, J.; Llavanera, M.; Bonilla-Correal, S.S.; Papas, M.; Gacem, S.; Rodríguez-Gil, J.E.; Yeste, M.; Miró, J. Irradiating frozen-thawed stallion sperm with red-light increases their resilience to withstand post-thaw incubation at 38ºC. Theriogenology 2020, 157, 85–05. [Google Scholar] [CrossRef]
- Catalán, J.; Papas, M.; Gacem, S.; Mateo-Orero, Y.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Red-light irradiation of horse spermatozoa increases mitochondrial activity and motility through changes in the motile sperm subpopulation structure. Biology 2020, 9, 254. [Google Scholar] [CrossRef]
Figure 1.
Change of proAKAP4 concentration after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) mean significant proAKAP4 concentration differences (p<0.05) between incubation times with or without cryoprotectant. Different letters (a,b) mean significant proAKAP4 concentration differences (p<0.05) at the same incubation time between treatment (With or without cryoprotectant).
Figure 1.
Change of proAKAP4 concentration after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) mean significant proAKAP4 concentration differences (p<0.05) between incubation times with or without cryoprotectant. Different letters (a,b) mean significant proAKAP4 concentration differences (p<0.05) at the same incubation time between treatment (With or without cryoprotectant).
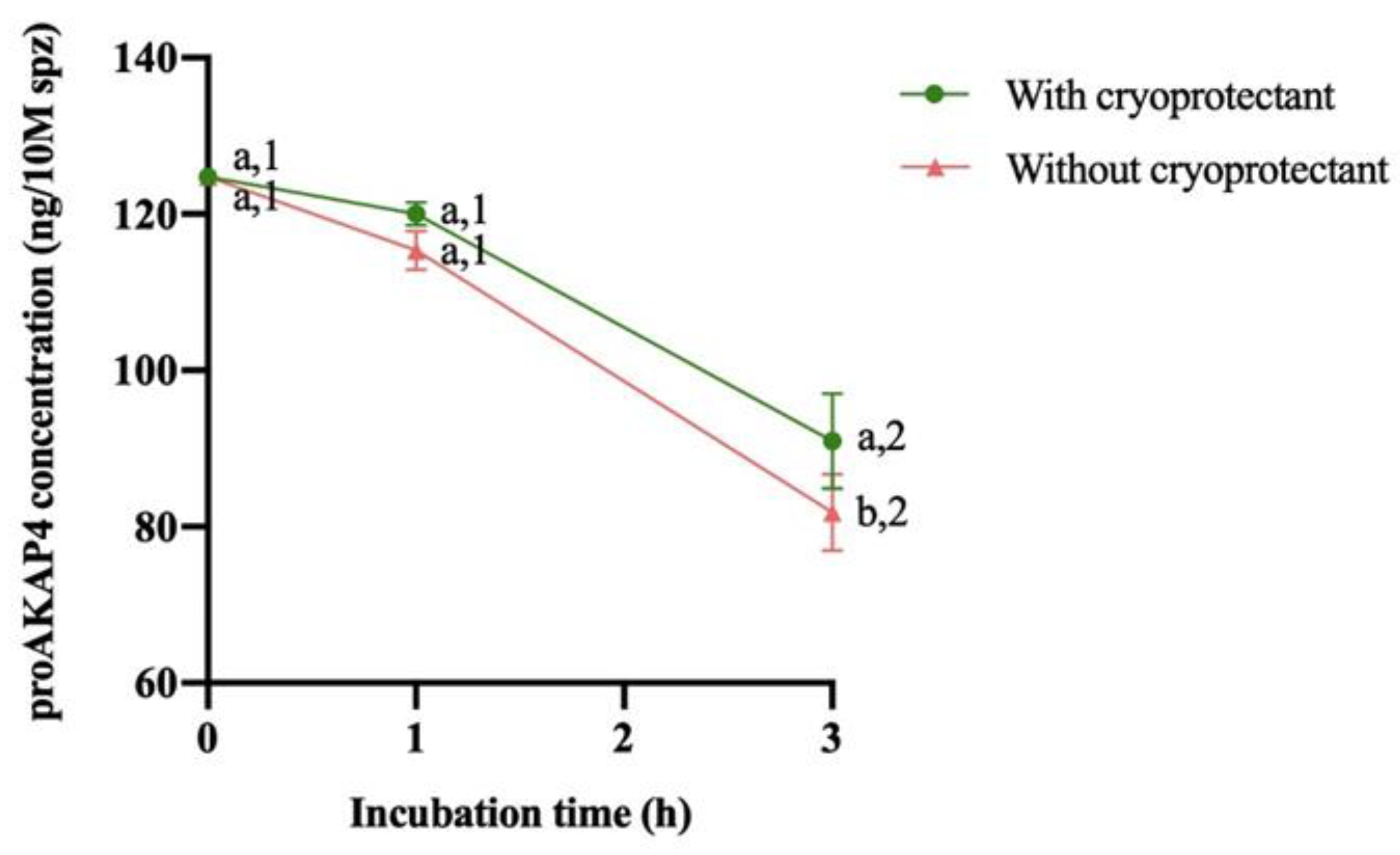
Figure 2.
Change of (A) total motility (%) and (B) progressive motility (%) after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) between mean significant differences (P<0.05) between incubation time with or without cryoprotectant. Different letters (a,b) show significant differences (P<0.05) between treatment (with or without cryoprotectant) at each incubation time.
Figure 2.
Change of (A) total motility (%) and (B) progressive motility (%) after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) between mean significant differences (P<0.05) between incubation time with or without cryoprotectant. Different letters (a,b) show significant differences (P<0.05) between treatment (with or without cryoprotectant) at each incubation time.
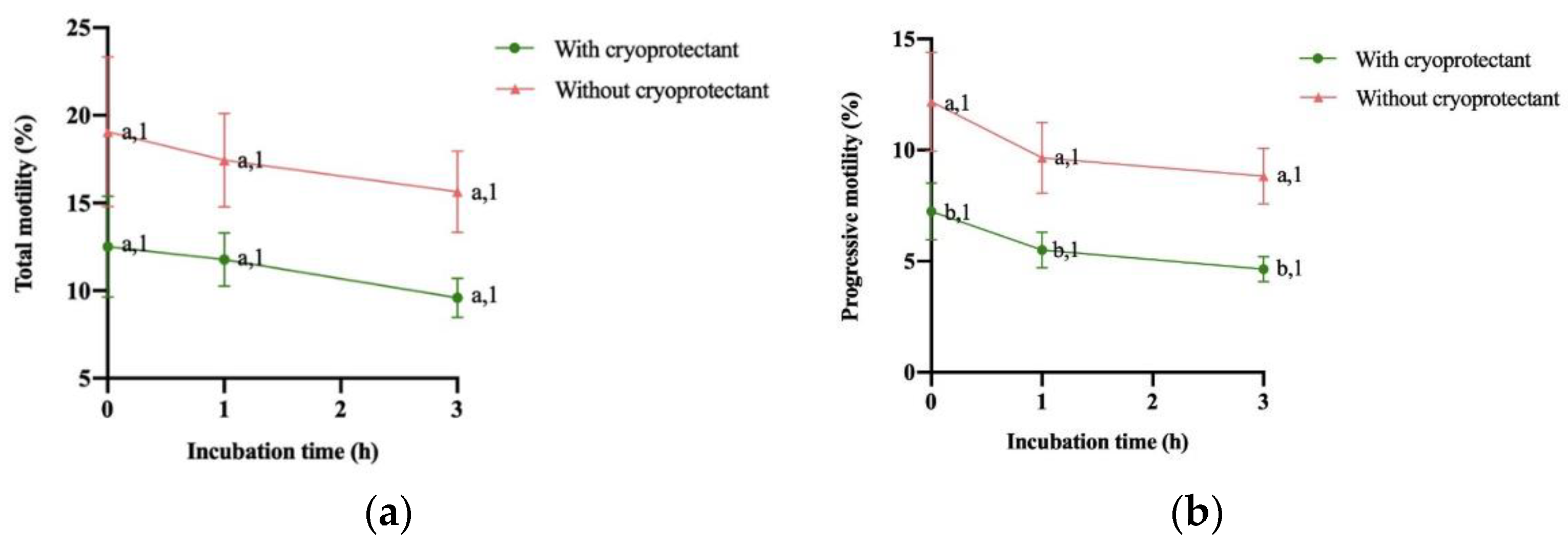
Figure 3.
Change of mitochondrial membrane potential (MMP, %) after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) mean significant differences (P<0,05) between incubation time for the same treatment (with or without cryoprotectant). No differences between treatment (with or without cryoprotectant) were observed at the same incubation time (a).
Figure 3.
Change of mitochondrial membrane potential (MMP, %) after thawing and during the following 3 hours of storage at 37°C (mean ± SEM). Different numbers (1,2) mean significant differences (P<0,05) between incubation time for the same treatment (with or without cryoprotectant). No differences between treatment (with or without cryoprotectant) were observed at the same incubation time (a).
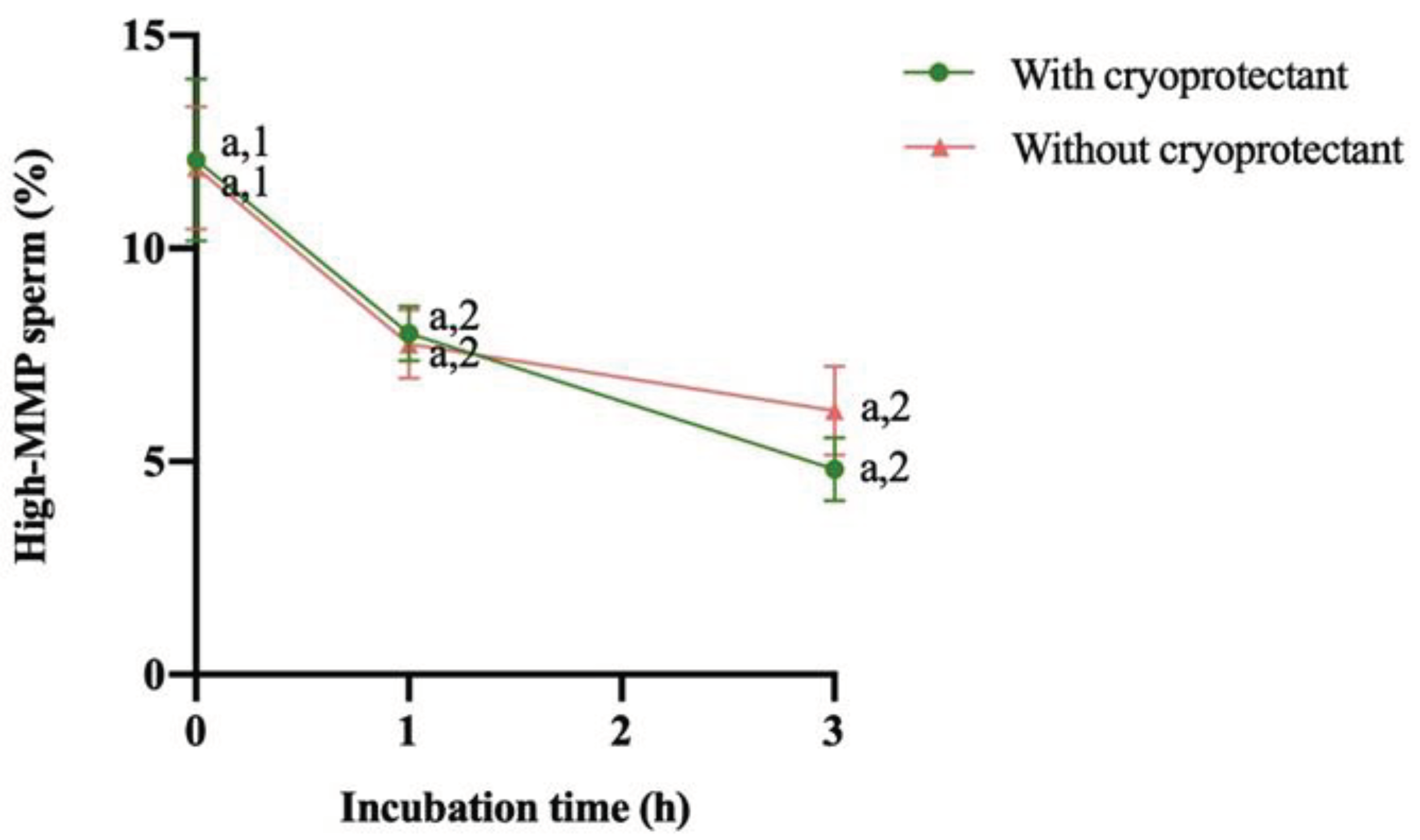

Table 1.
Pearson’s correlation between proAKAP4 concentration in ng/10 M (millions) spermatozoa and motility descriptors and mitochondrial membrane potential at 0, 1 and 3 hours after thawing. Total motility (TM, %), progressive motility (PM, %), curvilinear velocity (VCL, μm/s), straight-line velocity (VSL, μm/s), average path velocity (VAP, μm/s), linearity (LIN, %), straightness (STR, %), amplitude of lateral head displacement (ALH, μm) and frequency of head displacement (BCF, Hz), mitochondrial membrane potential (JC1). *mean significant correlation (p<0.05).
Table 1.
Pearson’s correlation between proAKAP4 concentration in ng/10 M (millions) spermatozoa and motility descriptors and mitochondrial membrane potential at 0, 1 and 3 hours after thawing. Total motility (TM, %), progressive motility (PM, %), curvilinear velocity (VCL, μm/s), straight-line velocity (VSL, μm/s), average path velocity (VAP, μm/s), linearity (LIN, %), straightness (STR, %), amplitude of lateral head displacement (ALH, μm) and frequency of head displacement (BCF, Hz), mitochondrial membrane potential (JC1). *mean significant correlation (p<0.05).
| 0 hours | 1 hour | 3 hours | ||||
| r | p | r | p | r | p | |
| TM | 0.421 | 0.018* | 0.435 | 0.027* | 0.496 | 0.024* |
| PM | 0.352 | 0.041* | 0.367 | 0.039* | 0.402 | 0.027* |
| VCL | 0.666 | 0.000* | 0.211 | 0.247 | 0.222 | 0.223 |
| VSL | 0.440 | 0.012* | 0.139 | 0.449 | 0.120 | 0.511 |
| VAP | 0.527 | 0.002* | 0.160 | 0.382 | 0.105 | 0.567 |
| LIN | -0.273 | 0.131 | -0.069 | 0.707 | -0.243 | 0.181 |
| STR | -0.151 | 0.408 | -0.168 | 0.359 | -0.020 | 0.914 |
| ALH | 0.570 | 0.001* | 0.288 | 0.110 | 0.213 | 0.241 |
| BCF | 0.060 | 0.744 | -0.205 | 0.260 | 0.122 | 0.505 |
| JC1 | 0.190 | 0.298 | 0.135 | 0.461 | 0.338 | 0.058 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



