
Submitted:
22 March 2024
Posted:
25 March 2024
You are already at the latest version
Abstract
Autoimmune kidney diseases arise from a loss of tolerance to antigens, often with unclear triggers. In this review we explore the role of the gut microbiome and how disease, diet and therapy can alter the gut microbiota consortium. Perturbations in the gut microbiota may induce the translocation of microbiota-derived inflammatory molecules such as liposaccharide (LPS) and other toxins systemically by penetrating the gut epithelial barrier. Once in the blood stream these pro-inflammatory mediators activate immune cells which release pro-inflammatory molecules many of which are antigens in autoimmune diseases. The ratio of gut bacteria Bacteroidetes:Firmicutes is associated with worse outcomes in multiple autoimmune kidney diseases including Lupus nephritis, MPO-ANCA vasculitis and Goodpasture’s syndrome. Therapies which enhance short chain fatty acid (SCFA)-producing bacteria in the gut have powerful therapeutic potential. Dietary fibre is fermented by gut bacteria which in turn release SCFAs which protect the gut barrier as well as modulate immune responses towards a tolerogenic anti-inflammatory state. Herein, we describe where the current field of research is and the strategies to harness the gut microbiome as potential therapy.
Keywords:
ANCA
; kidney disease
; SCFA
; glomerulonephritis
; vasculitis
; gut microbiota
; diet therapy
; autoimmunity
; immune responses
1. Introduction
There are several different types of inflammatory kidney diseases which are characterized by the development of glomerulonephritis whereby the glomeruli of the kidneys become inflamed and are no longer capable of filtering out metabolic waste. If left untreated these diseases can reach end stage renal disease where life-long dialysis or kidney transplant is required for survival. Although characterized as glomerulonephritis, each kidney disease can have very different etiologies and treatment strategies such as: Lupus nephritis, Anti-Neutrophil Cytoplasmic Antibody (ANCA)-vasculitis, IgA nephropathy, Acute Kidney Injury, Goodpasture’s syndrome and Focal Segmental Glomerulonephritis.
There is increasing evidence on the importance of gut microbiota in disease. Microbial perturbations have been implicated as a risk factor in several inflammatory diseases, such as type 1 diabetes, lupus nephritis and rheumatoid arthritis [1,2,3]. The gut microbiota, which is the community of microorganisms that reside in the human gut, plays a key role in the metabolism and fermentation of dietary fiber, including resistant starch (RS) [4,5,6]. RS is known to promote the growth and diversity of beneficial gut bacteria, which can have important effects on gut health and immune function. This review will evaluate the relationships between gut microbiota and autoimmune kidney diseases and explores its potential for new therapies that target the gut microbiota to improve outcomes of patients with autoimmune and inflammatory kidney diseases.
2. Gut Microbiota in Health and Disease
The gut microbiota is the community of microorganisms that inhabit the human gut. This includes bacteria, viruses, fungi, and other microorganisms, which collectively have a significant impact on human health. A key role of the gut microbiota is to metabolize and harvest nutrients from indigestible starches as it passes through the upper gastrointestinal tract (GI), which humans cannot enzymatically digest on their own [4,7] (Figure 1). RS is a type of dietary fiber that resists digestion in the small intestine and reaches the large intestine largely intact. In the large intestine, it can be fermented by gut microorganisms to produce SCFAs acting as a prebiotic to promote growth of certain beneficial bacteria such as Bifidobacteria and Lactobacilli which thrive on the benefits of SCFA production [4,5,6] (Figure 1). RS has been shown to have several health benefits, including improved glycaemic control, increased satiety, and reduced inflammation, which may be attributed in part to the effects of SCFAs produced by gut bacteria [8,9]. Recent studies have shown that gut microbiota imbalance plays a crucial role in the development and progression in kidney diseases like lupus nephritis, diabetic nephropathy, chronic kidney disorder and Kawasaki disease [2,5,10,11].
Dietary intervention can change the composition of the human gut-microbiota [12]. A clinical trial comparing animal and plant-based diets confirmed that the human gut microbiota could be altered in a relatively brief period [13]. Animal-based diets expanded bile-tolerant microorganisms (Alistipes, Bilophila and Bacteroides), and decreased SCFA-producing bacteria (Roseburia, Eubacterium rectale, Ruminococcus bromii) [4,13,14]. A comparative study examining feces of children from a rural African village in Burkina Faso found that the children had a higher abundance of phylum Bacteriodetes (also known as Bacteroidota) and depleted Firmicutes from their high fiber diets, compared to European children who consumed a typical western diet (high in fat/salt and animal products) [15]. The gut microbiota is diverse and consists of many bacterial phyla such as Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria, Fusobacteria and Verrucomicrobia, with Bacteroidetes and Firmicutes being the two most dominant phyla, representing over 90% of the gut microbiota [16,17,18]. The balance and diversity of these microbial populations are believed to contribute to intestinal homeostasis and overall gut health. Hence, the ratio of Bacteroidetes:Firmicutes has been proposed as a marker for gut dysbiosis, with many studies reporting that patients with active disease often have a gut microbiota which are more dominated by the Firmicutes phylum [7,19]. The ratio is reported to be elevated in obesity [20] and inflammatory bowel disease [21] suggesting a potential link to gut dysbiosis. In type 2 diabetes, the ratio varies and is associated with different clinical parameters such as diet, lifestyle, genetics and environmental factors [22,23]. Alterations of the gut-microbiota brought upon by ‘western diets’ has been postulated to exacerbate inflammatory diseases [4]. Consumption of diet high in saturated fat often found in western diet contributes to the expansion of certain bacteria such as Bilophila wadsworthia, a pathobiont. Pathobionts are resident microorganisms within the host that are associated with chronic inflammatory conditions [24,25].
Increasing evidence of the role of gut microbiota in the exacerbation of autoimmune diseases have been widely reported in studies such as rheumatoid arthritis, systemic lupus erythematosus, and inflammatory bowel disease [26]. Bacterial populations with a predominantly pathobiont population namely, Bacilli and Enterobacteriaceae [27] are found to be involved in immune dysfunction in Kawasaki disease and IBD [28,29]. Treatment regimens of patients, disease severity and their diet and lifestyle may be a contributing factor in the populations of gut microbiota observed in patients. A more comprehensive investigation into the different factors involving the patients would elucidate the influence of treatment drugs, diet and the role of gut microbiota in disease.
3. Role of SCFAs in Modulating the Immune Response
SCFAs are by-products of gut microbial fermentation through metabolization of dietary fiber and glycosylated mucins [30]. The most abundantly produced SCFAs are acetate, butyrate and propionate, which have been shown to interact with G protein-coupled receptors (GPCRs) in the gut and other tissues [30,31]. GPCRs are a class of cell surface receptors that are involved in many important physiological processes, including the regulation of the immune system. In the gut, SCFAs have been shown to interact with several GPCRs, including GPR41, GPR43, GPR109a which are highly expressed on immune cells, including neutrophils, monocytes, and macrophages [5,31,32,33,34] (Figure 1). Activation of these receptors by SCFAs has been shown to have several beneficial effects on immune function, including the modulation of cytokine production, the promotion of regulatory T cells (Tregs), and the suppression of inflammation [30,31,32]. Following fermentation of dietary fiber, some SCFAs are able to interact with monocarboxylate transporters (MCTs) to be transported into and out of cells into the systemic circulation for use by other tissues [33].
SCFAs modulate the immune system by elucidating their effects on SCFA-sensing GPCRs on immune cell surfaces [35]. Butyrate was shown to induce extrathymic differentiation of Tregs and colonic FOXp3 Tregs through GPR43, as well as decrease pro-inflammatory cytokine gene expression of IL-12, IL-6 and NFκB protein RelB in dendritic cells (DCs), allowing DCs to facilitate Treg differentiation [35]. Butyrate could also reprogram macrophage polarization to an anti-inflammatory phenotype by activating metabolic pathways favoring the M2 phenotype [36]. Disturbances to butyrate-producing gut bacteria by broad spectrum antibiotics could perturb intestinal macrophages, shifting them to a pro-inflammatory state, highlighting how SCFA-dependent pathways are required for immune homeostasis [36].
4. Gut-Kidney Axis
Kidney disease is associated with increased intestinal permeability where endotoxin (a component of gut bacterial cell wall) translocates across the intestinal barrier into the blood stream, activating immune responses. Although data on the gut-kidney axis is sparse, evidence shows kidney injury leads to the accumulation of urea and other uremic toxins which can translocate into the intestinal lumen and perturbate the commensal bacteria [37]. Urea is converted to ammonia which damages tight junctions in the intestine, contributing to intestinal permeability. Conversely, the gut microbiota is also a source of metabolites that enhance the intestinal barrier, most notably anti-inflammatory SCFAs such as butyrate, propionate and acetate. A diet high in non-digestible carbohydrates, such as RS, promotes the growth of SCFA-producing bacteria, which are notably depleted in end stage renal disease (ESRD), lupus nephritis, and IgA nephropathy. There is a large body of evidence demonstrating that there is a significant difference in gut microbial diversity and metabolic pathways between healthy individuals and patients with inflammatory diseases. Given the role of SCFAs in maintaining immunological homeostasis both locally at the gut and systemically, it is critical to investigate the role of the gut-kidney axis in kidney diseases and test potential gut-targeted therapeutic interventions.
5. Role of Microbiota in Specific Inflammatory Kidney Diseases
5.1. ANCA-Associated Vasculitis
ANCA-associated Vasculitis (AAV) is an autoimmune disease that affects small blood vessels, including those in the kidneys. AAV can be categorized into three clinical manifestations: microscopic polyangiitis (MPA), granulomatosis with polyangiitis (GPA) and eosinophilic granulomatosis with polyangiitis (EGPA) depending on the distribution of vascular inflammation, autoantigen specificity and presence/absence of granulomatosis with asthma. AAV can be identified through clinical pathological phenotypes, such as MPA when there is an absence of granulomatosis with asthma, GPA when there is granulomatosis but absence of asthma and EGPA when there is presence of granulomatosis with asthma [39]. Although target organs involved in AAV tend to be kidneys, skin, peripheral nerves, upper and lower respiratory tract, AAV can also be organ-specific depending on the AAV subtype [39]. A common clinical feature between the 3 clinical subtypes is the loss of tolerance to autoantigen specificity [39] and the generation of autoantibodies to cytoplasmic granules in neutrophils, which are myeloperoxidase (MPO), and proteinase 3 (PR3) that are present in the circulation of most patients [40,41]. These autoantibodies are known as ANCA [40,42]. ANCAs are thought to cause glomerular and vascular inflammation of AAV. The incidence of MPO-ANCA is predominantly found in the Asia Pacific region particularly the Chinese and Japanese populations whereas GPA-associated PR3-ANCA incidence decreases towards the equator, with higher incidence in Northern Europe, India, and the Middle East [43,44].
MPO-ANCA-associated glomerulonephritis (GN) is a rapidly progressive disease with a high mortality of 80% between 5 to 12 months if left untreated [45,46,47,48,49]. The disease often results in rapidly progressive crescentic GN which leads to renal failure and eventual death. Treatment often includes a combination of immunosuppression therapies with high-dose glucocorticoids and cyclophosphamide in active disease [48,49]. Current treatment regimens have greatly reduced the significantly high mortality rate by more than 90% [48]. However, because of the toxicity of immunosuppressant, the rate of drug-induced complications is high [50,51]. Furthermore, GN may relapse following withdrawal of treatment with corticosteroids and cyclophosphamide and often requires ongoing maintenance therapies [49,50,51]. Other therapies utilizing drugs such as Rituximab directed against B cells and plasma exchange have shown promising and higher recovery rates in clinical trials [51,52,53,54]. Novel and less invasive alternative therapies aimed at reducing inflammation in AAV or prolonging the period of remission will be a more focused and strategic approach at targeting MPO-ANCA-vasculitis. Recently, the association between gut microbiota and AAV was reported in a study conducted in a Chinese cohort [27]. Differences in the gut microbiota was observed in patients with AAV with a predominantly pathobiont population namely, Bacilli and Enterobacteriaceae [27].
Compelling evidence from preliminary studies in our laboratory on established murine model of MPO-AAV have shown that high fibre diet alters the ratio of Bacteroidetes:Firmicutes. HFD led to the enrichment of Bacteroidaceae, Muribaculaceae and Tannerellaceae which are families of SCFA-producing bacteria that come from the phylum Bacteroidetes. Prophylactic consumption of HFD ameliorated histological renal injury, which was associated with decreased glomerular immune cell (neutrophils, macrophages and CD4+ T cell) infiltration into the kidneys [55]. The study provides a proof of concept that in the context of MPO-AAV, a RS-diet can shift the balance of Firmicutes:Bacteroidetes. One method of overcoming the variable gut mirobiota population between mice and human is by using a fecal microbiota transplant (FMT) technique from a human donor into specific pathogen free (SPF) mice, which can mimic human microbiotas (humanised gnotobiotic mice) [56]. SPF mice have decreased gut diversity within their cecum compared to wild mice, possessing enterotypes correlating with low species richness (alpha diversity) and inflammation (Family Bacteroidaceae and Enterobacteriaceae) [57,58]. A study using SPF C57BL/6 mice found it can successfully engraft 85% of the donor’s gut microbiota upon FMT, encompassing all bacterial phyla, 11/12 bacterial class and 58/66 bacterial genus found in the donor’s gut microbiota [56].
5.2. Goodpasture’s Syndrome
Goodpasture’s syndrome is also known as Type I Crescentic GN with Linear Immunoglobulin Deposition or Autoimmune Anti-glomerular Basement Membrane (anti-GBM)-associated GN. The disease accounts for around 10% of cases of crescentic GN [59]. The disease is associated with humoral autoimmune responses to the non-collagenous domain of α3 type IV collagen, which is a constituent of the GBM [60]. It is recognized by the linear deposition of the anti-GBM antibody in the GBM [60]. Antibody autoreactivity results in complement activation by the Fc receptor that can cause glomerular injury by recruitment of macrophages and neutrophils to the glomerular capillaries causing diminished renal function observed by proteinuria and hematuria [60]. Goodpasture’s syndrome can be a serious and potentially life-threatening condition, and prompt and aggressive treatment is essential for the best possible outcome. Treatment typically involves plasma exchange and medications that suppress the immune system, such as corticosteroids and immunosuppressants [61].
There is currently no evidence linking Goodpasture’s syndrome to the gut microbiota. There has been one study in experimental anti-GBM in rats that suggests that SCFA treatment can attenuate pathological endpoints in anti-GBM disease. In particular, butyrate was able to reduce urine protein, serum creatinine and glomerular crescent formation [62]. However, certain medications (cyclophosphamide and prednisolone, see Section 6) used to treat Goodpasture’s syndrome, such as immunosuppressants, can affect the gut microbiota. This can lead to an increased risk of developing infections and other complications. Thus, alterations in gut microbiota composition and function may play a role in the development of Goodpasture’s syndrome.
5.3. IgA Vasculitis
IgA vasculitis, also known as Henoch-Schonlein purpura, is a type of vasculitis that primarily affects small blood vessels, including those in the skin, kidneys, and gastrointestinal tract. It is caused by the deposition of IgA immune complexes in the blood vessel walls, leading to inflammation and damage to the blood vessels. The exact cause of this condition is unknown, but it is thought to be related to an abnormal immune response triggered by an infection or other environmental factors. Symptoms of IgA vasculitis can include a rash on the skin, joint pain and swelling, abdominal pain, and blood in the urine. Diagnosis is typically based on a combination of clinical symptoms and laboratory tests, including blood tests and a kidney biopsy. Neutrophils are the predominant cells found in cutaneous and gastrointestinal biopsies of patients with IgA vasculitis [63]. Studies have shown that NETs play a role in the pathogenesis of IgA vasculitis [64]. In particular, NETs have been found to be involved in the formation of immune complexes and in the activation of the complement system, which can lead to tissue damage [65,66]. It is possible that NETs are activated by bacterial products that escape through tight junctional proteins between the gut epithelial cells. Both LPS from gram negative bacteria and CpG can activate NETs through the pattern recognition receptors TLR4 and TLR9 respectively [67,68,69,70]. Treatment for IgA vasculitis may include medications to reduce inflammation and control symptoms, such as non-steroidal anti-inflammatory drugs (NSAIDs), corticosteroids, and immunosuppressive drugs, further causing pertubations of gut bacteria. In most cases, the condition resolves on its own within a few weeks to months, but some individuals may experience long-term kidney damage or other complications.
The relative abundance of Actinobacteria in patients with IgA vasculitis is lowered compared with healthy controls [71]. As IgA vasculitis can be heterogenous in presentation, this study categorized patients according to organ types affected. Interestingly species diversity of gut bacteria changed according to the different categories. Those patients that mainly had skin and manifestations with no gut involvement had a dominance of the Actinobacteria family. Whereas overall diversity was decreased in all patients with IgA vasculitis compared to healthy subjects. A study of inherited microbiota from mother to children in IgA Vasculitis has demonstrated that the gut microbiome is strongly associated with their mothers [72]. In particular the abundance of both Magamonas and Lacotbacillus was positively correlated with their mother’s abundance of the same gut bacteria. A positive correlation between clinical factors and the bacterial species from children with IgA vasculitis was found. Alisitipes putredinis was associated with fibrinogen, and Bifidobacetrium longum was associated with partial thromboplastin time (PPT, is an indication of how long it takes your blood to clot). Dialister abundance was decreased in children with IgA vasculitis. This finding agrees with other studies that have shown that Dialister abundance is also decreased in other inflammatory diseases such as eczema in children. A reduction in the abundance of Ruminococcus was also noted in children with IgA vasculitis. This is an important finding as Ruminoccous is responsible for the production of the SCFAs butyrate and propionate which are anti-inflammatory in terms of T cell activation. In agreement with these findings, another study in children found Ruminococcus was the most abundant bacteria in convalescent children whereas Veillonella was associated with acute IgA Vasculitis [73]. This suggests that Ruminoccous is associated with gut health, and methods which would enrich for Ruminoccous either through diet or prebiotics may be beneficial in IgA vasculitis.
5.4. IgA Nephropathy
IgA nephropathy, also known as Berger’s disease, is a kidney disease characterized by the deposition of pathogenic immunoglobulin A1 immune complexes in the glomeruli of the kidney. The disease involves mesangial proliferation and the deposition of inflammatory responses by the infiltration of macrophages, monocytes and T cells, leading to damage and scarring of the kidney tissue. The exact cause of IgA nephropathy is not fully understood, but it is believed to be related to bacterial infection and autoantibodies specific for IgA1 that cause the formation of IgA1 immune complexes in the circulation, some of which deposit in the kidneys resulting in glomerular injury [74,75]. Recent research has also suggested a possible link between IgA nephropathy and the gut microbiota.
Several studies have found alterations in the gut microbiome of individuals with IgA nephropathy, including a decrease in beneficial bacteria and an increase in pathogenic bacteria. The imbalance of the gut microbiota populations is often observed in populations of Bacteroidetes and Firmicutes between IgA nephropathy patients and healthy controls [76,77,78]. The gut mucosal system serves as a barrier between the external environment and the body’s internal tissues. IgA is predominantly found in mucosal secretions, including those in the gastrointestinal tract. Thus, IgA is crucial in the regulation of immune defence. The dysfunction of intestinal barrier can lead to increased intestinal permeability, which may allow for the translocation of bacteria products and antigens into the bloodstream, triggering an immune response and influx of IgA1 deposition in the kidney. Overall, the link between gut microbiome and IgA nephropathy is an area of active research, and further studies are needed to fully understand the mechanisms involved and to identify potential therapeutic targets.
5.5. Lupus Nephritis
Lupus Nephritis is one of the most common and serious complications of Systemic Lupus Erythematosus (SLE). It is estimated that around 30% to 60% of individuals with SLE develop lupus nephritis and is one of the leading causes of morbidity and death among patients with SLE [79]. The relationship between lupus nephritis and the gut microbiome is an area of ongoing research, and the exact mechanisms and interactions are not yet fully understood. While there is limited research specifically examining the Bacteroidetes/Firmicutes ratio in the context of lupus nephritis, some studies have explored the gut microbiota composition in SLE. Thus, the alteration of gut microbiota by Firmicutes/Bacteroidetes ratio is commonly used as a marker for pathological symptoms and severity. Increased intestinal permeability is normally observed in individuals with SLE, which is often correlated with changes in the gut microbiota, including Firmicutes/Bacteroidetes ratio. However, the Firmicutes/Bacteroidetes ratio in SLE and lupus nephritis are not consistent across all studies [80]. Additionally, alterations in the gut microbiota composition, including changes in the Bacteroidetes/Firmicutes ratio, may be influenced by various factors such as diet, medication, disease activity, and individual variations. Therefore, it is challenging to draw definitive conclusions about the role of the Bacteroidetes/Firmicutes ratio specifically in SLE.
A dominance of Ruminococcus in patients with SLE is associated with an increased number or circulating Tregs, however an abundance of Ruminococcus gnavus is 5 times the abundance seen in healthy patients [81,82]. SLE patients with active disease according to the Systemic Lupus Erythematosus Disease Activity Index (SLEDAI) have an increased titre of anti-Ruminococcus gnavus antibodies in their serum. Studies using mice models of lupus have demonstrated that probiotics containing both Lactobacillus rhamnosus and Lactobacillus delbruekii are able to down regulate microRNAs (miRNAs) associated with biomarkers in SLE. miR-181a which can modulate T cell receptor signaling and control B and T cell differentiation is increased in the serum of patients with active SLE [83]. Peripheral blood mononuclear cells from patients with SLE incubated with both Lactobacillus rhamnosus and Lactobacillus delbruekii had significant downregulation of miR-181a and miR-155, another miRNA associated with disease in SLE patients. This suggests that prebiotics containing both Lactobacillus rhamnosus and Lactobacillus delbruekii may have the potential to reduce inflammation in SLE.
5.6. Acute Kidney Injury
The impact of gut microbiota on the development of acute kidney injury (AKI) have been explored in germ-free mice with ischemia-reperfusion injury (IRI). Where the transplantation of gut microbiota from stool of normal mice resulted in renoprotection quantified by the degree of histological renal damage and function [84], suggesting the role of gut microbiota in the development or severity of AKI. Interestingly, another study demonstrated that the depletion of gut microbiota with antibiotics exacerbated renal damage and function in an IRI AKI murine model [85]. Furthermore, the study reported that resident F4/80 macrophages associated with the initiation of renal inflammation following IRI and chemokines CX3CR1 and CCR2 expressions on F4/80+ macrophages were reduced in gut microbiota-depleted mice. Re-inoculation of gut microbiota by fecal transplantation from normal mice into antibiotic-treated mice did not alleviate renal injury. Mice with induced bilateral IRI were protected from gut inflammation but not intestinal leakage with probiotic supplementation with Bifidobacterium bifidum BGN4 and Bifidobacterium longum [86]. This was accompanied with a decreased infiltration of both innate cells (macrophages and neutrophils) and Th17 cells. The antibiotic amoxicillin has been shown to accelerate recovery in mice after IRI. The protective mechanism was not found in germ-free mice suggesting that gut bacteria have a direct impact on regulating immune responses. IL-17a CD4+ T cells were decreased in the kidneys of mice treated with amoxicillin, but this effect was not seen in germ-free mice [87]. This gives further impetus to investigate the use of probiotics as adjunct therapy in AKI.
5.7. Diabetic Nephropathy
The state of diabetes is associated with an alteration of microbial composition [88] and furthermore the gastrointestinal microbiome of patients with diabetic kidney disease (DKD) is differential compared to the state of diabetes without kidney injury. The genus Escherichia-Shigella has been shown to discriminate those with DKD from either those with T2DM and no kidney injury [89,90] and healthy controls [89,91]. Escherichia-Shigella belongs to the phyla Pseudomonadota (previously known as Proteobacteria), a phyla of gram-negative bacteria, and this phyla is enriched in DKD participants compared with T2DM with no renal injury [90]. Furthermore, in comparison to healthy controls, Pseudomonadota has been observed to be enriched in patients with T2DM and CKD stage 4-5 [92], CKD stage 4-5 [93] ESRD requiring dialysis [94,95] and IgA Nephropathy [76]. Patients with CKD have been shown to have higher serum LPS compared to healthy controls [92] indicative of decreased integrity of the intestinal barrier and increased presence of Pseudomonadota [96]. The presence of gram-negative Pseudomonadota in the blood would lead to macrophage activation via TLR4 recognition of LPS which signals through either MYD88 or TRIF to activate NFқB and cause the subsequent production of proinflammatory cytokines such as TNFα, IL-6 and Type I interferon.
It has been posited that Enterobacteriaceae, a family of bacteria in the Pseudomonadota phyla, are particular contributors to inflammatory sequalae in a number of conditions affecting the gastrointestinal [97,98], renal [99] and hepatic [100] systems. This has led to the concept of Enterobacteriaceae blooms [97], whereby an increase in this taxa drives a proinflammatory response, ultimately contributing to morbidity and mortality across a number of disease conditions [101]. From a therapeutic standpoint, dietary intervention has been demonstrated to be successful in reducing Enterobacteraciae in mouse models of DKD [19] as well as human CKD patients [102]. Treatment approaches that modulate the gut microbiota and prevent overexpansion of Enterobacteraciae may represent potential treatment modalities in DKD and other renal inflammatory conditions.
6. Gut Microbiota and Interactions with Standard of Care Immunosuppressants
The critical role of gut microbiota in maintaining bodily functions is well-established. How the microbiota interacts with therapeutics commonly used in standard of care for inflammatory diseases (cyclophosphamide, prednisolone, rituximab and cyclosporine) is not well-defined. Emerging evidence suggests that microbiota and therapeutic treatments interact with each other. In particular, the gut microbiota can influence drug pharmokinetics through altercation of metabolic processing, and either increase or decrease bioavailability of drugs. An example of this is cyclosporine A, a calcineurin inhibitor which is commonly used in autoimmune diseases and in transplants to prevent rejection [103].
In the case of cyclophosphamide (CYC) it has been shown that it can alter the composition of the microbiota of the small intestine and aids in the translocation of gram-positive bacteria into secondary lymphoid organs. It is thought that CYC injures the gut mucosal layer, damaging tight junctions and allowing the escape of bacteria [104]. Rituximab (RTX) the B cell-depleting antibody, induces mucosal damage in a similar manner to CYC. This results in increased inflammatory cells, and gut permeability in RTX-treated mice. In particular, Lactobacillus reuteri (L.reuteri) is reduced in the gut. In vitro studies demonstrated that L.reuteri inhibits inflammation in LPS-stimulated cultured mesenteric lymph node cells.
Inflammation in mice can be reduced via reintroduction of L.reuteri into RTX-treated mice. Prednisolone commonly prescribed for inflammation in autoimmune kidney diseases [105] alters the gut microbiota enriching for the Anaerobacterium species in mice studies. This was accompanied with a decrease in immunomodulatory SCFAs propionate and isobutyrate. Given patients are often on long term doses of prednisolone, probiotics or diets that can encourage the growth of the bacteria that are lost are worth pursuing.
7. The Role of SCFAs and Innate Cells
SCFAs play a significant role in modulating the function of innate immune cells. Innate immune cells are a crucial component of the immune system that provides immediate defense against pathogens. SCFAs, which are produced through the fermentation of dietary fiber by gut microbiota, have been shown to influence the function of various innate immune cells by regulating various protein molecules such as NLRPR3 inflammasome and toll-like receptors (TLRs) within the context of the innate immune system. The inflammasome is a multiprotein complex involved in the activation of inflammatory responses. Its regulation by SCFAs, particularly butyrate has been shown to inhibit the NLRP3 inflammasome thus contributing to the control of inflammation [106,107]. Additionally, SCFAs act as energy substances to preserve the intestinal barrier and homeostasis and inhibit autophagy, and as histone deacetylase inhibitors to suppress NLRP3 inflammasome [107]. SCFAs also modulate the activity of TLRs, a family of proteins that play a crucial role in recognizing microbial components, hence regulating the innate immune response to pathogens.
The influence of SCFAs on TLR signaling have been demonstrated in studies where TLR3 and TLR4 activation were downregulated, particularly by butyrate and propionate [108,109]. Butyrate and propionate have been shown to reduce the production of proinflammatory cytokines such as tumor necrosis factor-alpha, interleukin-6 and interleukin 1 beta, which are mediated through the inhibition of nuclear factor kappaB (NF-κB) induced by TLR activation [109,110]. Additionally, butyrate has been found to modulate the expression of TLR4 and the phosphorylation of mitogen-activated protein kinases and NF-κB in colon cancer cells [108]. These findings suggest that butyrate and propionate is involved in the modulation of immune responses and inflammation through the regulation of TLR3 and TLR4 activation. Microbial triggers or dysregulation of the immune response mediated by TLR ligation can contribute to the development and exacerbation of kidney disease. TLR4 is well-known for its role in recognizing LPS, a component of the gram-negative bacteria cell wall. Studies have shown that there is an upregulation of TLR4 in CKD [111] and an activation of TLR4 by LPS triggers a pro-inflammatory response which contributes to the development and progression of kidney pathologies [112]. Activation of TLR4 and TLR9 have been demonstrated to contribute to neutrophil recruitment and subsequently the exacerbation of autoimmune ANCA-associated GN [113,114]. Furthermore, TLR2, TLR4 and TLR9 have been shown to be involved in both glomerular and tubulointerstitial compartments of the kidneys in patients with AAV, with TLR4 being the most prominent suggestive of its central role in the inflammatory processes associated with AAV in the kidneys [115]. Understanding the roles of TLRs in kidney diseases is crucial for the development of targeted therapeutic strategies. Modulating TLR signaling via SCFAs interactions may represent a potential approach to mitigating inflammation and alleviating kidney injury. However, it is important to note that the effects of SCFAs may vary depending on the cell type and context, as they have been shown to increase TNFα-induced inflammation in lung mesenchymal cells [116]. Further research is needed to fully understand the mechanisms and potential therapeutic applications of SCFAs in modulating inflammatory responses. Nonetheless, the contribution of SCFAs on the promotion of an anti-inflammatory environment, influencing downstream signaling pathway associated with TLR activation is promising. By regulating TLR signalling, SCFAs contribute to the fine-tuning of immune responses, helping to maintain a balance between effective defence against pathogens and prevention of excessive inflammation or inappropriate immune reactions.
Neutrophils are the key mediator of injury in AAV. Autoimmunity to the major neutrophil enzyme MPO or PR3 results in the generation of ANCA which binds to activated neutrophils and triggers a unique form of pathological cell death termed “neutrophil extracellular traps” (NETs) or NETosis. NETs release webs of DNA containing injurious enzymes that cause inflammation of the blood vessels depositing the autoantigen MPO or PR3. The relationship between neutrophils and the gut microbiota in the generation of autoimmunity in AAV remain to be explored. Neutrophils are highly influenced by microbial metabolites particularly butyrate which have been found to inhibit NETosis [117]. It is well established that administration of SCFAs, or promoting the growth of SCFA-producing gut bacteria skews the inflammatory response towards tolerance. The gut microbiota is capable of regulating neutrophil function including controlling the magnitude of inflammatory responses, and influences neutrophil activation and recruitment [118]. Targeting neutrophil-butyrate signaling pathways which inhibit its activation highlights the potential for new therapies to improve outcomes in patients with AAV.
In the model of experimental autoimmune ANCA-induced GN, neutrophils are known to be the primary immune cells to traffic to the glomeruli and subsequently cause glomerular MPO deposition and injury in the kidneys [119]. Neutrophils are key players in the innate immune system. They are the most abundant type of leukocytes and play a crucial role in the immediate response to infections. SCFAs have been shown to inhibit neutrophil activation through various mechanisms. SCFAs, particularly propionate and butyrate were demonstrated to downregulate the production and release of proinflammatory mediators by neutrophils such as nitric oxide and pro-inflammatory cytokines TNFα and cytokine-induced chemoattractant 2αβ [120]. This effect is mediated by the inhibition of HDAC activity and NF-κB activation [120]. HDAC play a role in the formation of NETs which are released by activated neutrophils [121] and involves the citrullination of histones by peptidyl arginine deiminase 4 [122]. Studies have shown that butyrate-mediated inhibition of HDAC can enhance the differentiation and function of Tregs, thereby contributing to immune tolerance and regulation of inflammation (Figure 1). In the context of ANCA-vasculitis, butyrate-mediated HDAC inhibition could affect the expression of genes involved in neutrophil activation and NET formation, thereby impacting NETosis.
Furthermore, activation of the GPR43 receptor by SCFAs is found to induce a chemotactic response in neutrophils [123,124]. The GPR43 receptor has been shown to be expressed in renal tissues [125] and is identified as a key player in the activation of neutrophils [126]. SCFAs, especially acetate and propionate are able to activate the GPR43 on the surface of neutrophils and modulate the chemotaxis of neutrophils to and from sites of infection or inflammation [124] through the expression of chemokine receptors including CXCL1 and CXCL2 on the neutrophils [124]. By modulating the production and responsiveness to chemokines, SCFAs may contribute to the regulation of kidney inflammation, potentially limiting excessive neutrophil recruitment and activation and consequently dampening inflammatory responses and disease. More recently, studies on anti-GBM GN in rats demonstrated that treatment of SCFAs, particularly butyrate ameliorated disease severity with a decrease in T cell activation and an increase Treg cell differentiation [127]. While mechanisms by which SCFAs dampen immune responses and GN were not investigated, it is plausible that the interaction between butyrate and GPR43 expressed on renal cells cause the increase the Treg population and play a significant role in the development of GN.
8. SCFAs Modulate T Regulatory Cells
SCFAs have been shown to play a significant role in the regulation of Tregs differentiation and function. SCFAs, particularly butyrate, have been found to promote the differentiation of naive T cells into effector T cells or Tregs, depending on the immunological milieu. This process occurs through the inhibition of HDACs, leading to increased histone acetylation and gene transcription associated with Treg differentiation and regulation of the mTOR-S6K pathway [128]. As Tregs play a pivotal role in regulating the immune system and maintaining immune tolerance, the involvement of SCFAs is crucial in the promotion of immune homeostasis and regulation of immune responses, particularly in ANCA-associated vasculitis, where disease is the result of autoimmunity to the autoantigen. Research has consistently shown that, SCFAs, particularly butyrate enhances the suppressive function of Tregs by increasing the production of anti-inflammatory cytokine IL-10 and transforming growth factor-beta (TGF-β) [128,129,130]. SCFAs produced by gut microbiota have been shown to influence the migration and homing of Tregs to sites of inflammation and tissue damage by regulating the expression of chemokine receptors on Tregs, particularly the key receptor CCR4 [131] thereby guiding their migration to specific tissues or lymphoid organs [132]. The dysregulation of chemokine receptors has been implicated in various autoimmune diseases, including multiple sclerosis, rheumatoid arthritis, type 1 diabetes, autoimmune thyroiditis, Graves’ disease and Addison’s disease [133,134,135,136]. These receptors and their ligands play a critical role in the recruitment and trafficking of immune cells to affected organs, contributing to the pathogenesis of these diseases. Studies have demonstrated the importance of Tregs in the maintenance of tolerance to MPO in the experimental model of MPO-AAV [137]. However, the role of chemokine receptors on Tregs in kidney disease has not been investigated. It is possible that the dysregulation of chemokine receptors via SCFAs interactions in AAV may impact Treg function, contributing to the pathogenesis of the disease.
9. Use of Animal Models in Microbiome Studies - Considerations for Translation into Human Studies
Animal models play a crucial role in microbiome studies, providing controlled systems to investigate the complex interactions between microbial changes and disease. The microbiome composition and function can vary significantly between animal models and humans. Thus, selecting appropriate animal models and experimental designs is imperative to closely reflect human pathophysiology. Animal models are often used to test the efficacy of potential microbiome-targeted therapies, such as dietary intervention [7,10], antibiotic treatment [138], FMT [139] and administration of microbial metabolites such as SCFAs [127]. These interventions can be investigated for their ability to modulate microbiome and ameliorate disease severity. Animal models allow for the induction of specific diseases or conditions and study of how alterations in the microbiome may contribute to disease pathogenesis.
9.1. Differences between Human and Mouse Gut Anatomy as Gut Microbiota Diversity
Although the gastrointestinal tracts in both species are similar it is important to understand the critical differences when studying the gut microbiome in mice digestive tracts. In terms of intestinal surface area versus body surface area, both species are similar. However, where the difference occurs is over the anatomical structure of the gut. For example, the length ratio of the small intestine to colon is 7 in humans compared to 2.5 in mice with the mouse cecum being larger than the human cecum which allows them to process extra nutrients from indigestible fibre with greater fermentation capacity [140]. The location of fermentation of fibre by gut bacteria in the mouse system is compartmentalized in the cecum in comparison to the fermentation of gut bacteria in humans which occurs in the colon [141]. The physiological consequence of this is that mice have an expanded cecum capacity allowing them to digest significantly more undigestible fibres in their diet relative to humans. The differences do not end there, in terms of the histological structure of the intestinal wall, mice have much more elongated villi than that of humans hence increasing surface area, allowing for enhanced absorption of nutrients [141].
Given the anatomical differences between mice and humans it is not surprising that this would provide different ecological niches for gut bacteria favouring the growth of some families over others. Despite this the two species share 90% similarities in phyla and 89% in genera [142]. However, it is important to be aware that a large number of bacterial families found in mice are not present in humans. Mice microbiota composition favours a greater Bacteroidetes:Firmicutes ratio and humans have a greater Firmicutes:Bacteroidetes ratio [143].
9.2. Use of Germ Free/Gnotobiotic Animal Models
Germ-free or gnotobiotic animal models, which are animals that are completely lacking gut microbiota or with defined microbial communities allows for the study of the impact of microbiota on various diseases. Changes in the microbiome composition can be assessed by administrating prebiotics, probiotics, antibiotics or FMT to tease out the effect certain families of bacteria have on disease outcomes. FMT can be used to colonize germ-free or antibiotic-treated animals to explore the ability of gut microbiota in modulating disease severity as demonstrated by FMT from patients with hypertension [144] and end stage renal disease [145] and mice with colitis [146] to germ-free mice.
Germ-free or gnotobiotic animal models are a critical resource for the study of the intestinal microbiota. Germ-free mice as the name suggests have had all their gut microbiota removed. This in itself is technically challenging requiring a dedicated facility where mice pups are removed under sterile conditions from their mothers to prevent contamination with the mother’s bacteria flora [147]. In contrast gnotobiotic animals allow for mice to be colonized with specific microbiota allowing researchers to dissect out the specific characteristics and importance of specific strains of bacteria. The most common use of germ-free mice is to use them as a recipient for FMT studies. However, these models are not without their limitations. Mice raised in a sterile microisolator can lead to abnormalities in their development as well as have an abnormal immune system [148]. Work carried out in these germ-free facilities comes at a great cost as they require dedicated staff, and space for the microisolators. Given the cost, an important alternative is to use antibiotics to deplete the gut bacteria. There are different combinations used by different laboratories but the most common cocktails use varying concentrations of ampicillin, vancomycin and gentomycin [149,150]. Some of the issues with this approach is side effects from the antibiotics which can affect metabolic processes and disturbances in immune cell functions [151].
9.3. Housing Consideration When Using Mouse Models for Microbiota Studies
Experimentation in animal models requires controlled experimental conditions, allowing for standardized protocols and reproducibility of results. However, factors such as housing, age, sample collection site, littermate effect, bedding, supplier effect may influence consistency and reproducibility of experiments [152].
Each individual cage mice are housed in, creates a unique environment. This is due to close contact with other mice and coprophagy (consumption of other mouse faeces). To reduce agistment costs in animal houses it is common practice to put 4-5 mice in the same cage. Studies have shown that reducing housing density to 2 per cage has an effect on the gut microbiota providing more effective antibiotic treatment than 4-5 mice housed together [153]. Furthermore, fluctuations in temperature not only effect gut microbiome composition but reduce reproducibility for microbiome experiments. Examples of this is mice housed at 22 degrees Celsius have an enrichment of Lachnospiraceae compared to mice housed at 30 degrees Celsius which are enriched for Prevotellaceae [154].
9.4. The Requirement for Guidelines for Studying the Microbiome
The role of the gut microbiome in varying diseases has become more apparent in recent years gaining traction as an appealing strategy for treatment of varying diseases. With this comes an increasing need to ensure there are guidelines governing this that allows reproducibility. A critical factor for consideration in this area of research is determining causation versus association. In terms of dissecting out the role of the microbiome in renal diseases there is an additional confounding issue in that the relationship between kidney function and the gut microbiome is reciprocal. Urea that accumulates systemically as renal disease progresses is able to translocate across the gut epithelia where it has a direct effect on the gut microbiota. Conversely, ammonia which is released as a biproduct of urea hydrolysis released by bacterial urease is utilized as a source of nitrogen for bacteria to synthesis critical amino acids, puramines and sugars [155]. Therefore, when conducting microbiome studies to determine causation versus association reverse experiments should be conducted where gut bacteria from the diseases groups of interest is transplanted via FMT into germ free mice [145].
FMT itself needs strict guidelines. Guidelines for reporting on animal faecal transplantation (GRAFT) have been published outlining the importance of having standard protocols for the collection, downstream processing and minus 80 degrees Celsius storage of the samples before transplant. One of the critical factors in preparation of the samples for transplant is the vehicle in which it is prepared paired with the conditions in which it is prepared. For example, if the faecal samples are not prepared under anaerobic conditions many of the anaerobic bacterial families will have decreased viability [156].
This gives impetus to ensuring findings in mouse studies are replicated in well-designed human clinical trials. Special considerations should be given to the microbiome targeted interventions such as high fibre diets that have been utilised in mouse studies, to ensure effective translation into human studies.
10. Modulation of Gut Microbiota as Potential Therapy
Dietary fibre is critical for a healthy gut. Non-digestible fibres are fermented by gut bacteria which produce SCFAs which in turn provide an energy source for colonocytes to enhance gut barrier integrity. The epithelium of the colon receives the majority of its energy from SCFAS (over 70%), of which butyrate is the most critical. A complication with the treatment of patients with CKD is they are often prescribed low-fibre diets to limit potassium intake due to the risk of hyperkalaemia [157]. Therefore, the gut microbial composition becomes altered due to the lack of fibre.
10.1. Dietary Intervention
A western diet is characterized by a high intake of processed food containing fat, sucrose, low intake of dietary fibers, minerals and vitamins, linked with several metabolic disorders and inflammatory disease [158]. Dietary intervention has an obvious impact on health but the composition can also modulate the microbiota profile, thereby dysregulating the host immunity. Several studies have shown diet, microbiota and autoimmune diseases are highly interconnected [159]. As a result, prebiotics such as dietary fiber are metabolized into SCFAs and other products which have both a direct and indirect effect to our health [160] (Figure 2).
One of the dietary recommendations is fiber, which is a non-digestible carbohydrate that bypass’s degradation by the digestive enzyme and reaches the large intestine intact [161]. Resistant starch in particular is fermented slowly over time therefore, compared to other fermentable oligosaccharides, disaccharides, monosaccharides and polyols (FODMAPS) which contribute to unwanted side effects such as bloating and gas, RS is much more palatable as a choice. Evidence from clinical trials using high amylose resistant starch that has been enriched with both acetate and butyrate (HAMSAB) have shown that the immune profiles can be altered in Type 1 Diabetes with as little as 6 weeks supplementation. Those subjects which had the highest concentrations of SCFAs in the blood had better glycemic control [162]. Short term studies with HAMSAB for the control of hypertension have also showed positive results with a reduction in 24-hour systolic blood pressure. Importantly, HAMSAB supplementations were associated with a change in the gut microbial diversity by creating an ecological niche that favored the growth of SCFA-producing bacteria[163].
Dietary fiber intervention has a positive effect by stimulating the production of SCFA metabolites and promoting beneficial gut microbiota that leads to a decreased production of uremic toxin, creatinine, and inflammatory response in people with CKD [164]. A prospective observational study involving 61 Belgian children (age <18 years) with all stages of CKD (stage 1-5) including recipients of transplant, and different etiologies (congenital anomalies of the kidneys and urinary tracts, polycystic disease and other non-glomerular diseases) demonstrated an opposite relationship between the rise in fiber intake (g/day) and serum concentrations of different gut-derived protein-bound waste products, namely indole acetic acid (free and total), indoxyl sulfate, free p-cresyl sulfate, as well as p-cresyl glucuronide (free and total) [165].
Analysis of a nationwide database reported that among 14,543 participants, 25.7% CKD subpopulation had elevated serum c-reactive protein (CRP) (>3 mg/l) compared with the non-CKD subpopulation. Additionally, DF intake (10 g/day) (total, soluble, and insoluble DF) was significantly associated with lower serum CRP in patients in CKD patients [166]. Another cohort of a 1110 elderly Swedish men (age 70–71 years) with CKD were given a 7-day dietary fiber intake of 10 g/day higher than their usual intake. This resulted in an increase of eGFR with an average of <60 ml/min per 1.73m and a reduced mortality. Notably, participants in higher dietary fiber quartiles had lower levels of CRP (<3 mg/l) and IL-6 level [167].
In a recent randomized trial, a total of 162 participants with end-stage renal disease were instructed to consume 10g of DF (potato starch) orally each day for 8 weeks. Lactobacillus, Bifidobacterium adolescentis, and lactobacillaceae were significantly increased in the DF group. Interestingly, SCFAs namely butyric acid, isovaleric acid, isobutyric acid, hexanoic acid, and valeric acid were also significantly increased whilst there was no significant change for either propionic acid and acetic acid levels [168].
An observational study in Germany comprising 3193 moderately severe CKD participants utilized a food frequency questionnaire to score adherence to dietary recommendations. The study revealed that increased consumption of fiber and potassium was linked to higher eGFR and lower CRP. Conversly, higher intake of sodium, cholesterol, total protein, and sugar, had the opposite effect [169]. In a follow up study on potassium intake in a South Korean cohort, studied the effect of dietary potassium. Among 5064 participants with stage 2 CKD (aged ≥40 years), the study found that those with high potassium intake (Q3 1765.603–2364.251 and Q4>2364.251 mg/day) had a lower risk of CKD development and decrease in eGFR (<60 mL/min/1.73 m2) [170].
RS has been proposed as a treatment for CKD patients to modulate the microbiota in the gut by promoting the proliferation of favourable gut bacteria and increase the production of metabolites including SCFAs [5]. It can be degraded by a diverse group of microbes such as primary degraders (Ruminococcus bromii and Bifidobacterium adolescentis) and secondary degraders (Eubacterium rectale, Roseburia and Butyrivibrio) as well as Bacteroides thetaiotaomicron which can generate metabolites like butyrate, lactate and acetate [171].
Studies in experimental models of disease and humans with kidney diseases have shown that RS intervention increases the concentrations of SCFAs and diversity of gut microbes. For instance, rats fed a diet containing high amylose maize starch (5% RS type 2) compared to a caloried control group, showed an increase in fecal SCFA after a 2-week intervention. Additionally, lower levels of pH and blood urea were observed [172]. Similar interventional studies of RS [high amylose maize RS (59%)] for a duration of 28 days in a mouse model of CKD (via nephrectomy) demonstrated a slow progression of CKD. The gut bacterial diversity was also higher in RS groups. However, BUN/creatinine levels were not significantly altered in CKD mice supplemented with RS compared to control mice fed a calorie-matched diet [173].
A randomized trial in 75 patients (18-80 years) with early diabetic nephropathy were treated with 50 g of high RS for 12 weeks. This resulted in a reduction in glucose levels and an increase in serum uric acid and superoxide dismutase levels. However, the diet was unable to reduce inflammation markers IL-6 and TNF-α [174].
Likewise, in a randomized control trial conducted in Iran, 20 patients undergoing haemodialysis or end stage kidney disease were given biscuits comprising 20 and 25 g/d of high-amylose maize RS type 2, during the first and second month respectively. Interestingly, the 16s DNA sequencing from fecal samples revealed a significant increase of Faecalibacterium bacteria but not with the genera Parabacteroides, Bifidobacteria, Ruminococcus and Prevotella [175]. As seen in the previous studies, neither TNFα nor IL-6 were not significantly altered with this diet.
10.2. SCFAs as Potential Therapeutic Agents
SCFAs namely acetate, propionate, and butyrate represent the main metabolites generated by the gut microbiota, all of which have immunomodualtory functions [176]. As mentioned earlier, the anaerobic fermentation of DF and RS by gut microbiota produce SCFA. As a result, dietary fiber is strongly associated with increased SCFA production [177]. Thus, dietary intervention, SCFAs and gut microbiota profile, function and metabolism are highly interconnected with each other. SCFAs warrant investigation as potential therapeutics to either restore gut homeostasis via protective effects of the gut barrier, or to modulate inflammatory cells (Figure 2).
Gut bacteria produced SCFAs provide protection to the gut epithelium. SCFAs are recongised by specific GPCRs in the colon. Via these receptors butyrate for example is able to aid in enhancing the mucus layer by increasing production of Mucin 2. Acetate produced by Bifidobacterium a SCFA prouducing bacteria provides energy for both propionate and butyrate producing bacteria, an example of a postive feed back loop where by one bacteria releases SCFA to enhance the growth of another [178]. A healthy adult blood is estimated to have a concentration of 100-150 µmoL/L for acetate, 4- 5 µmoL/L for propionate, and 1-3 µmoL/L for butyrate [179] with a molar ratio of 60 % acetate, 20% butyrate and 20% propionate in the large intestines [180]. Acetate and propionate are mainly induced by Bacteroidetes (gram-negative), while butyrate is predominantly triggered by Firmicutes (gram-positive) gut microbial phyla [181].
In vitro butyrate and propionate are able to promote the production of Tregs from naïve T cells in co-culture with DCs. However Th1, Th2, and Th17 cytokines (IL-4, IL-13, IL-17, IFN-γ) remained unchanged upon butyrate treatment [35]. Butyrate has potent anti-inflammatory effects that are involved in intestinal homeostasis and energy metabolism [182,183]. SCFAs also regulate mTOR–S6K pathway and Tregs by promoting IL-1, IFN, and/or IL-10 [128]. Interestingly, a reduction in anti-CD3-induced inflammation occurred in the acetate-treated group, in an IL-10-dependent manner [128].
Li et al., demonstrated that butyrate significantly inhibits the formation of NETs and neutrophil-associated inflammatory mediators including chemokines (CXCL1, CCL3, CCL4, and IL-8), CCL20, CXCL9, and MPO which were significantly decreased in patients with IBD [117]. Nephrectomised rats receiving sodium butyrate (400mg/kg/day) in drinking water had markedly improved insulin resistance and significantly reduced circulating LPS levels. This was accompanied with restoration of decreased IL-10 levels compared to the control groups. Markers of kidney injury were also significantly decreased with lower levels of serum urea and proteinuria in the treatment groups, however there was no effect on creatinine levels [184]. This suggests that treatment with SCFAs, in particular butyrate may be of potential therapeutic benefit in kidney diseases.
An in human study has shown that SCFAs given as apple cider vinegar or through RS inulin (to induce bacteria to release SCFAs) can modulate immune cells in healthy individuals. After 3 weeks of the intervetion B cells were lower on the High SCFA diet compared to those on a low SCFA diet. Likewise Th1 cells were lower in the High SCFA diet compared to the low SCFA diet [185]. This provides proof of concept that SCFAs can influence different populations of immune cells, and may be of potential theruapeutic benefit in the treatment of inflammatory kidney diseases. Although a high fibre diet contain RS can produce large amounts of anti inflammatory SCFAs, it is difficult for patients to maintain these diets. This is evidenced by the increaing level of obesity in the developed world. Although its is known changing your diet will reduce weight and encourage a healthy gut many patients are unable or unwilling to make these changes. Therefore administration of SCFAs themselves rather than via diet may be an attractive alternative for many non-compliant patients who do not follow prescribed diets.
10.3. Fecal Microbiota Transplants as Therapy
FMT has become an emerging attractive therapeutic agent with the successes seen in treatment for IBD. There is now growing interest in extending these findings to target autoimmune inflammatory diseases and cancer. FMT has been demonstrated to restore altered microbiota and reconstruct the gut micro ecosystem and modulate both the innate and adaptive arms of the immune systed, leading to therapeutic outcomes for different diseases [186]. Among these, it has been very successful and extensively used for treating Clostridioides difficile infection implicated in IBD [187].
The principle behind FMT as a form of therapy is to restore altered gut microbial ecology by replacing it with fecal microbiota from a healthy individual. The donor is heaily screened and can be chosen by the patient (e.g., a family member) or from a biobank. Screening includes esuring none of the donors have infections (particulary Hepatititis A, B and C, HIV or syphillis, helminth or other parasie infections), or have a gastrointestinal disorder. Importantly donors are screened to make sure they have not had any antibiotic treatment which can deplete healthy gut bacteria.
There have been multiple animal studies of kidney disease that demonstrate that FMT can alleviated renal disease endpoints (kidney histology and functional injury evidenced by lower serum creatine or albuminuria/proteinuria).The gut microbiota composition affects the intestinal barrier integrity and accumulation of harmful metabolites such as urea, indoxyl sulfate (IS), and p-cresyl sulfate (PCS), leads to accumulation of immune complex’s that cause damage to the renal parenchyma in renal diseases such as lupus nephritis [188]. FMT, once transferred has been shown to rapidly normalize the microbial community structure within 24 hours after administration to the recipient [189]. A preclinical trial on humanized mice demonstrated that FMT modulates IgA nephropathy phenotype and inflammation by increasing serum B cell activation factor (BAFF), decreasing CD89 cell surface expression, both of which are associated with IgA1 mesangial deposits [190].
A recent study aimed to improve CKD patients renal dysfunction by administering a washed microbiota transplanst WMT for 3 consecutive days. Unlike conventional FMT this study used a wash protocol in the preparation of the WMT. This consisited of repeated saline washes, microbiota purification and centrifuging, prior to transfer via either transendoscopic or nasojeunal tube. This method resulted in significant differences in microbial diversity and an improvement in renal function evidenced by an improvement in blood urea nitrogen and serum creatine. Interestingly, some gut genera, including Eubacterium coprostanoligenes, Anaerostipes, Monoglobus, and Dorea, demonstrated a significant increase while Hungatella genera was decreased after FMT [191].
Despite tremendous achievements have been made so far, optimization of protocols and the potential metabolic and immunological consequences during host-microbiota interaction is not clearly established [189]. Additionally, a disease-specific approach may be needed for donor selection and FMT in CKD patients such as characteristics of ideal donors, dose, frequency, route of administration, etiology and stage of kidney disease and the need of the aforementioned recommended diets [192]. This highlights the importance of conducting standardized research as outlined in the GRAFT guidelines.
10.4. Transfer of Immunomodulatory Microbial Isolates as Therapy
FMT has an obvious beneficial effect for several inflammatory diseases, based on the level of success treating patients with IBD. However, the administration of whole fecal product may have a risk of introducing unexpected pathogenic bacteria in the gut. To reduce such risk, researchers have been investigating an alternative approach by administering specific bacterial strains extracted from healthy stool [193]. As discussed earlier [191] it can also be hypothesized that certain gut microbiota may not provide benefits, as indicated by a reduction in specific genera following FMT rather than an increase in protective immunomodulatory familes of bacteria.
Although there are limited studies for specific microbial isolates in patients, there has been a randomized clinical trial aimed to see the modulation of the intestinal microbiota in healthy adults. Consumption of the probiotic strain Bifidobacterium bifidum for 4 weeks isolated from fecal samples obtained from a healthy woman, resulted in a significant increase in the Ruminococcaceae and Rikenellaceae bacterial families, but a decreased Prevotellaceae family following administration [194]. In contrast, mono-colonization with segmented filamentous bacteria in a mouse model of MS exacerbated autoimmune demyelination through the triggering of myelin-specific CD4+ T cells and autoantibody-producing B cells [195].
10.5. Bacterial Exosomes
Extracellular vesicles (EVs) in health and disease have been an area of increasing interest. EVs are a heterogeneous group of lipid-membrane-bound vesicles (30- to 5,000-nm) secreted by almost all cell types [196]. EVs are generally categorized into three subtypes including microvesicles, exosomes, and apoptotic bodies. This classification is based on their their size, biogenesis, release pathways, conten, and function [197]. The cargo of EVs comprise functional proteins, nucleic acids, lipids, and other bioactive molecules, thereby modulating the behaviour of recipient cells and playing a role in different physiological and pathological processes [198]. Due to their role in immune system modulation, the applications of EVs as therapeutic targets, novel drug delivery agents and stand alone therapeutics are being actively explored.
Exosomes have been widely proposed as natural nanocarriers for delivering functional RNAs, proteins, and synthetic drugs which can be used therapeutically. This is attributed to their compatibility, reduced toxicity, biocompatible, minimally immunogenic, and retaining targeting capacity properties [199]. However, studies are very limited for their ability to regulate the gut microbiome as a therapy in autoimmune and inflammatory kidney disease. This limited understanding could be due to the highly dynamic structure and complexity of EVs as well as the secretions by different cells and tissues in both physiological and pathological conditions.
While certain studies have shown a promising therapeutic potential of renal-derived exosomes, the critical role of gut bacterial-derived exosomes and the associated molecular mechanisms have received little attention. However, it is clear that bacteria-host interactions mediated by exosomes could affect the expression level of either inflammatory or anti-inflammatory factors depending on the environment [200].
Proteomics studies have discovered that glomerular podocytes and tubular cells are the principal cells secreting exososomes in the nephron [201]. Studies have indicated that hypoxia, along with hypoxia-inducible factor-1 (HIF-1) promote exosome production in tubule epithelial cells, thereby modulating the fate of adjacent cells and consequently the extent of kidney damage [202]. Thus, it is reasonable to speculate that tubular exosomes could communicate with gut microbiota and gut bacterial exosome in turn have a direct therapeutic effect during pathological conditions of chronic kidney inflammation.
The gut bacterium Akkermansia muciniphila-derived EVs were shown to regulate mucosal inflammation by inhibiting the production of IL-6 from colon epithelial cells in IBD mice model [203]. Lactobacillus EVs are also reported as a promising therapy for IBD and autoimmune diseases [204].
In summary, most of the specific gut microbiota EV cargo components responsible for the therapeutic approach in autoimmune and inflammatory kidney disease have not been fully explored. Therefore, deeper understanding to determine the molecular mechanisms of complex cargoes carried by several bacterial exosomes along with different types of kidney diseases and exploring how exosomes interact with receptor cells using multi-omics approach would be of great interest.
11. Conclusion
The role of gut microbiome in autoimmune kidney disease is becoming an area of growing research. Given the lack of current treatments that are both effective and without toxic side effects, exploring mechanisms in which we can control inflammation and mitigate disease progression through the manipulation of gut microbiome presents a potential for novel therapeutic approach. Nevertheless, numerous questions persist regarding the utilization of gut microbiome manipulation as therapy.
References
- Marino, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat Immunol 2017, 18, 552–562. [Google Scholar] [CrossRef]
- Valiente, G.R.; Munir, A.; Hart, M.L.; Blough, P.; Wada, T.T.; Dalan, E.E.; Willis, W.L.; Wu, L.C.; Freud, A.G.; Jarjour, W.N. Gut dysbiosis is associated with acceleration of lupus nephritis. Sci Rep 2022, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.D.; Fernandes, G.R.; Calderaro, D.C.; Mendonca, S.M.S.; Silva, J.M.; Albiero, M.L.; Cunha, F.Q.; Xiao, E.; Ferreira, G.A.; Teixeira, A.L.; et al. Oral microbial dysbiosis linked to worsened periodontal condition in rheumatoid arthritis patients. Sci Rep 2019, 9, 8379. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Mackay, C.R.; Kaye, D.M. Beyond gut feelings: how the gut microbiota regulates blood pressure. Nat Rev Cardiol 2018, 15, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Kellow, N.J.; Coughlan, M.T. Modulation of the Gut Microbiota by Resistant Starch as a Treatment of Chronic Kidney Diseases: Evidence of Efficacy and Mechanistic Insights. Adv Nutr 2019, 10, 303–320. [Google Scholar] [CrossRef]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review article: short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment Pharmacol Ther 2018, 48, 15–34. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of Prebiotic Fiber and Insufficient Signaling Through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Tanti, J.F.; Ceppo, F.; Jager, J.; Berthou, F. Implication of inflammatory signaling pathways in obesity-induced insulin resistance. Front Endocrinol (Lausanne) 2012, 3, 181. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr Rev 2009, 67, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Chen, X.; Kwan, T.K.; Loh, Y.W.; Singer, J.; Liu, Y.; Ma, J.; Tan, J.; Macia, L.; Mackay, C.R.; et al. Dietary Fiber Protects against Diabetic Nephropathy through Short-Chain Fatty Acid-Mediated Activation of G Protein-Coupled Receptors GPR43 and GPR109A. J Am Soc Nephrol 2020, 31, 1267–1281. [Google Scholar] [CrossRef]
- Chen, J.; Yue, Y.; Wang, L.; Deng, Z.; Yuan, Y.; Zhao, M.; Yuan, Z.; Tan, C.; Cao, Y. Altered gut microbiota correlated with systemic inflammation in children with Kawasaki disease. Sci Rep 2020, 10, 14525. [Google Scholar] [CrossRef] [PubMed]
- R, R.M.; Marques, F.Z. Diet-related gut microbial metabolites and sensing in hypertension. J Hum Hypertens 2021, 35, 162–169. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Im, S.H. Of men in mice: the development and application of a humanized gnotobiotic mouse model for microbiome therapeutics. Exp Mol Med 2020, 52, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Tan, S.; Clarke, R.; de Pasquale, C.; Thallas-Bonke, V.; Nguyen, T.; Penfold, S.; Harcourt, B.; Sourris, K.; Lindblom, R.; et al. Processed Foods drive Intestinal Barrier permeability and Microvascular Diseases. Science Advances 2021, 7. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Kusnadi, Y.; Saleh, M.I.; Ali, Z.; Hermansyah, H.; Murti, K.; Hafy, Z.; Yuristo, N.S.E. Firmicutes/Bacteroidetes Ratio of Gut Microbiota and Its Relationships with Clinical Parameters of Type 2 Diabetes Mellitus: A Systematic Review. Open Access Macedonian Journal of Medical Sciences 2023, 11, 67–72. [Google Scholar] [CrossRef]
- Polidori, I.; Marullo, L.; Ialongo, C.; Tomassetti, F.; Colombo, R.; di Gaudio, F.; Calugi, G.; Marrone, G.; Noce, A.; Bernardini, S.; et al. Characterization of Gut Microbiota Composition in Type 2 Diabetes Patients: A Population-Based Study. International Journal of Environmental Research and Public Health 2022, 19, 15913. [Google Scholar] [CrossRef]
- Chow, J.; Tang, H.; Mazmanian, S.K. Pathobionts of the gastrointestinal microbiota and inflammatory disease. Curr Opin Immunol 2011, 23, 473–480. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, L.; Ren, Q.; Feng, H.; Tao, S.; Cheng, L.; Ma, L.; Gou, S.J.; Fu, P. Understanding the Gut-Kidney Axis in Antineutrophil Cytoplasmic Antibody-Associated Vasculitis: An Analysis of Gut Microbiota Composition. Front Pharmacol 2022, 13, 783679. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Kusuda, T.; Nakashima, Y.; Murata, K.; Kanno, S.; Nishio, H.; Saito, M.; Tanaka, T.; Yamamura, K.; Sakai, Y.; Takada, H.; et al. Kawasaki disease-specific molecules in the sera are linked to microbe-associated molecular patterns in the biofilms. PLoS One 2014, 9, e113054. [Google Scholar] [CrossRef]
- Park, J.; Goergen, C.J.; HogenEsch, H.; Kim, C.H. Chronically Elevated Levels of Short-Chain Fatty Acids Induce T Cell-Mediated Ureteritis and Hydronephrosis. J Immunol 2016, 196, 2388–2400. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur J Pharmacol 2018, 831, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Coady, M.J.; Chang, M.H.; Charron, F.M.; Plata, C.; Wallendorff, B.; Sah, J.F.; Markowitz, S.D.; Romero, M.F.; Lapointe, J.Y. The human tumour suppressor gene SLC5A8 expresses a Na+-monocarboxylate cotransporter. J Physiol 2004, 557, 719–731. [Google Scholar] [CrossRef]
- Luu, M.; Pautz, S.; Kohl, V.; Singh, R.; Romero, R.; Lucas, S.; Hofmann, J.; Raifer, H.; Vachharajani, N.; Carrascosa, L.C.; et al. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat Commun 2019, 10, 760. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.A.; Andrusaite, A.; Andersen, P.; Lawson, M.; Alcon-Giner, C.; Leclaire, C.; Caim, S.; Le Gall, G.; Shaw, T.; Connolly, J.P.R.; et al. Antibiotics induce sustained dysregulation of intestinal T cell immunity by perturbing macrophage homeostasis. Sci Transl Med 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, E.; Kantartzi, K.; Tsigalou, C.; Konstantinidis, T.; Romanidou, G.; Voidarou, C.; Bezirtzoglou, E. Focus on the Gut-Kidney Axis in Health and Disease. Front Med (Lausanne) 2020, 7, 620102. [Google Scholar] [CrossRef]
- Cummings, J.H.; Hill, M.J.; Bone, E.S.; Branch, W.J.; Jenkins, D.J. The effect of meat protein and dietary fiber on colonic function and metabolism. II. Bacterial metabolites in feces and urine. Am J Clin Nutr 1979, 32, 2094–2101. [Google Scholar] [CrossRef]
- Jennette, J.C.; Falk, R.J. Pathogenesis of antineutrophil cytoplasmic autoantibody-mediated disease. Nat Rev Rheumatol 2014, 10, 463–473. [Google Scholar] [CrossRef]
- Jennette, J.C.; Falk, R.J. Small-vessel vasculitis. N Engl J Med 1997, 337, 1512–1523. [Google Scholar] [CrossRef]
- Jennette, J.C.; Hoidal, J.R.; Falk, R.J. Specificity of anti-neutrophil cytoplasmic autoantibodies for proteinase 3. Blood 1990, 75, 2263–2264. [Google Scholar] [CrossRef] [PubMed]
- Couser, W.G. Glomerulonephritis. Lancet 1999, 353, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Pearce, F.A.; Craven, A.; Merkel, P.A.; Luqmani, R.A.; Watts, R.A. Global ethnic and geographic differences in the clinical presentations of anti-neutrophil cytoplasm antibody-associated vasculitis. Rheumatology (Oxford) 2017, 56, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- Kitching, A.R.; Anders, H.J.; Basu, N.; Brouwer, E.; Gordon, J.; Jayne, D.R.; Kullman, J.; Lyons, P.A.; Merkel, P.A.; Savage, C.O.S.; et al. ANCA-associated vasculitis. Nat Rev Dis Primers 2020, 6, 71. [Google Scholar] [CrossRef] [PubMed]
- Ball, G.V. The history of ANCA-associated vasculitis. Rheum Dis Clin North Am 2010, 36, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Jayne, D. Review article: Progress of treatment in ANCA-associated vasculitis. Nephrology (Carlton) 2009, 14, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Hamour, S.; Salama, A.D.; Pusey, C.D. Management of ANCA-associated vasculitis: Current trends and future prospects. Ther Clin Risk Manag 2010, 6, 253–264. [Google Scholar] [CrossRef]
- Salama, A.D. Pathogenesis and treatment of ANCA-associated systemic vasculitis. J R Soc Med 1999, 92, 456–461. [Google Scholar] [CrossRef]
- Little, M.A.; Smyth, C.L.; Yadav, R.; Ambrose, L.; Cook, H.T.; Nourshargh, S.; Pusey, C.D. Antineutrophil cytoplasm antibodies directed against myeloperoxidase augment leukocyte-microvascular interactions in vivo. Blood 2005, 106, 2050–2058. [Google Scholar] [CrossRef]
- Nachman, P.H.; Hogan, S.L.; Jennette, J.C.; Falk, R.J. Treatment response and relapse in antineutrophil cytoplasmic autoantibody-associated microscopic polyangiitis and glomerulonephritis. J Am Soc Nephrol 1996, 7, 33–39. [Google Scholar] [CrossRef]
- Menahem, S.; Hiremagalur, B.; Mudge, D.; Toussaint, N.; Walters, G.; Caring for Australians with Renal, I. The CARI guidelines. Induction and maintenance therapy in ANCA-associated systemic vasculitis. Nephrology (Carlton) 2008, 13 Suppl 2, S24–S36. [Google Scholar] [CrossRef]
- Keogh, K.A.; Wylam, M.E.; Stone, J.H.; Specks, U. Induction of remission by B lymphocyte depletion in eleven patients with refractory antineutrophil cytoplasmic antibody-associated vasculitis. Arthritis Rheum 2005, 52, 262–268. [Google Scholar] [CrossRef]
- de Lind van Wijngaarden, R.A.; Hauer, H.A.; Wolterbeek, R.; Jayne, D.R.; Gaskin, G.; Rasmussen, N.; Noel, L.H.; Ferrario, F.; Waldherr, R.; Hagen, E.C.; et al. Clinical and histologic determinants of renal outcome in ANCA-associated vasculitis: A prospective analysis of 100 patients with severe renal involvement. J Am Soc Nephrol 2006, 17, 2264–2274. [Google Scholar] [CrossRef]
- Jayne, D.R.; Gaskin, G.; Rasmussen, N.; Abramowicz, D.; Ferrario, F.; Guillevin, L.; Mirapeix, E.; Savage, C.O.; Sinico, R.A.; Stegeman, C.A.; et al. Randomized trial of plasma exchange or high-dosage methylprednisolone as adjunctive therapy for severe renal vasculitis. J Am Soc Nephrol 2007, 18, 2180–2188. [Google Scholar] [CrossRef]
- O’Sullivan, K.M.; Snelson, M.; Nguyen, J.; Le, A.C.; Coughlan, M. Resistant starch supplementation alters the gut microbial consortium and attenuates kidney inflammation in an experimental model of autoimmune vasculitis. The Journal of Immunology 2023, 210, 165.08. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci Transl Med 2009, 1, 6ra14. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis Model Mech 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Linnenbrink, M.; Wang, J.; Hardouin, E.A.; Kunzel, S.; Metzler, D.; Baines, J.F. The role of biogeography in shaping diversity of the intestinal microbiota in house mice. Mol Ecol 2013, 22, 1904–1916. [Google Scholar] [CrossRef]
- Moroni, G.; Ponticelli, C. Rapidly progressive crescentic glomerulonephritis: Early treatment is a must. Autoimmun Rev 2014, 13, 723–729. [Google Scholar] [CrossRef]
- Reynolds, J.; Preston, G.A.; Pressler, B.M.; Hewins, P.; Brown, M.; Roth, A.; Alderman, E.; Bunch, D.; Jennette, J.C.; Cook, H.T.; et al. Autoimmunity to the alpha 3 chain of type IV collagen in glomerulonephritis is triggered by ‘autoantigen complementarity’. J Autoimmun 2015, 59, 8–18. [Google Scholar] [CrossRef]
- Marques, C.; Plaisier, E.; Cacoub, P.; Cadranel, J.; Saadoun, D. [Review on anti-glomerular basement membrane disease or Goodpasture’s syndrome]. Rev Med Interne 2020, 41, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gu, Q.-h.; Cui, Z.; Zhao, M.-h.; Jia, X.-y. Short-chain fatty acids ameliorate experimental anti-glomerular basement membrane disease. Clinical Immunology 2024, 259, 109903. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Huang, X.; Yu, G.; Qiao, J.; Cheng, J.; Wu, J.; Chen, J. Pathogenesis of IgA Vasculitis: An Up-To-Date Review. Front Immunol 2021, 12, 771619. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Tu, L.; Zou, J.S.; Zhu, S.Q.; Zhao, Y.J.; Qin, Y.H. The Involvement of Neutrophil Extracellular Traps in Disease Activity Associated With IgA Vasculitis. Front Immunol 2021, 12, 668974. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.S.; Zandbergen, M.; Wolterbeek, R.; Baelde, H.J.; van Es, L.A.; de Fijter, J.W.; Bruijn, J.A.; Bajema, I.M. Complement-mediated microangiopathy in IgA nephropathy and IgA vasculitis with nephritis. Mod Pathol 2019, 32, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Tsai, I.J.; Chang, C.J.; Chuang, Y.H.; Hsu, H.Y.; Chiang, B.L. The interaction between circulating complement proteins and cutaneous microvascular endothelial cells in the development of childhood Henoch-Schönlein Purpura. PLoS One 2015, 10, e0120411. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, R.; Khan, M.A.; Echaide, M.; Pérez-Gil, J.; Palaniyar, N. SP-D attenuates LPS-induced formation of human neutrophil extracellular traps (NETs), protecting pulmonary surfactant inactivation by NETs. Communications Biology 2019, 2, 470. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Su, X.; Pan, P.; Zhang, L.; Hu, Y.; Tan, H.; Wu, D.; Liu, B.; Li, H.; Li, H.; et al. Neutrophil extracellular traps are indirectly triggered by lipopolysaccharide and contribute to acute lung injury. Scientific Reports 2016, 6, 37252. [Google Scholar] [CrossRef] [PubMed]
- Aubé, F.A.; Bidias, A.; Pépin, G. Who and how, DNA sensors in NETs-driven inflammation. Front Immunol 2023, 14, 1190177. [Google Scholar] [CrossRef]
- Mallavia, B.; Liu, F.; Lefrancais, E.; Cleary, S.J.; Kwaan, N.; Tian, J.J.; Magnen, M.; Sayah, D.M.; Soong, A.; Chen, J.; et al. Mitochondrial DNA Stimulates TLR9-Dependent Neutrophil Extracellular Trap Formation in Primary Graft Dysfunction. Am J Respir Cell Mol Biol 2020, 62, 364–372. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Lin, X.; Bian, X.; Jing, R.; Frelinger, A.; Zhang, A. Comparison and Analysis of Gut Microbiota in Children With IgA Vasculitis With Different Clinical Symptoms. Front Pediatr 2021, 9, 800677. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhao, C.; Zhao, L.; Sheng, D.; Chen, B.; Zhao, G.; Wang, Q.; Zhang, L. Taxonomic and functional shifts of gut microbiome in immunoglobulin A vasculitis children and their mothers. Front Pediatr 2024, 12, 1356529. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jia, X.; Lai, P.; Wang, K.; Bao, Y.; Li, X. Relevance of Intestinal Microbiota in Immunoglobulin A Vasculitis With Abdominal Involvement. Front Pediatr 2022, 10, 943267. [Google Scholar] [CrossRef] [PubMed]
- Rizk, D.V.; Saha, M.K.; Hall, S.; Novak, L.; Brown, R.; Huang, Z.Q.; Fatima, H.; Julian, B.A.; Novak, J. Glomerular Immunodeposits of Patients with IgA Nephropathy Are Enriched for IgG Autoantibodies Specific for Galactose-Deficient IgA1. J Am Soc Nephrol 2019, 30, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Moldoveanu, Z.; Suzuki, H.; Reily, C.; Satake, K.; Novak, L.; Xu, N.; Huang, Z.Q.; Knoppova, B.; Khan, A.; Hall, S.; et al. Experimental evidence of pathogenic role of IgG autoantibodies in IgA nephropathy. J Autoimmun 2021, 118, 102593. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Montemurno, E.; Piccolo, M.; Vannini, L.; Lauriero, G.; Maranzano, V.; Gozzi, G.; Serrazanetti, D.; Dalfino, G.; Gobbetti, M.; et al. Microbiota and metabolome associated with immunoglobulin A nephropathy (IgAN). PLoS One 2014, 9, e99006. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Tan, J.; Tan, L.; Tang, Y.; Qiu, Z.; Pei, G.; Qin, W. Modifications of gut microbiota are associated with the severity of IgA nephropathy in the Chinese population. Int Immunopharmacol 2020, 89, 107085. [Google Scholar] [CrossRef]
- Shah, N.B.; Nigwekar, S.U.; Kalim, S.; Lelouvier, B.; Servant, F.; Dalal, M.; Krinsky, S.; Fasano, A.; Tolkoff-Rubin, N.; Allegretti, A.S. The Gut and Blood Microbiome in IgA Nephropathy and Healthy Controls. Kidney360 2021, 2, 1261–1274. [Google Scholar] [CrossRef]
- Anders, H.J.; Saxena, R.; Zhao, M.H.; Parodis, I.; Salmon, J.E.; Mohan, C. Lupus nephritis. Nat Rev Dis Primers 2020, 6, 7. [Google Scholar] [CrossRef]
- Zhang, L.; Qing, P.; Yang, H.; Wu, Y.; Liu, Y.; Luo, Y. Gut Microbiome and Metabolites in Systemic Lupus Erythematosus: Link, Mechanisms and Intervention. Front Immunol 2021, 12, 686501. [Google Scholar] [CrossRef]
- Zhang, S.X.; Wang, J.; Chen, J.W.; Zhang, M.X.; Zhang, Y.F.; Hu, F.Y.; Lv, Z.Q.; Gao, C.; Li, Y.F.; Li, X.F. The level of peripheral regulatory T cells is linked to changes in gut commensal microflora in patients with systemic lupus erythematosus. Ann Rheum Dis 2021, 80, e177. [Google Scholar] [CrossRef]
- Azzouz, D.; Omarbekova, A.; Heguy, A.; Schwudke, D.; Gisch, N.; Rovin, B.H.; Caricchio, R.; Buyon, J.P.; Alekseyenko, A.V.; Silverman, G.J. Lupus nephritis is linked to disease-activity associated expansions and immunity to a gut commensal. Ann Rheum Dis 2019, 78, 947–956. [Google Scholar] [CrossRef]
- Vahidi, Z.; Samadi, M.; Mahmoudi, M.; RezaieYazdi, Z.; Sahebari, M.; Tabasi, N.; Esmaeili, S.-A.; Sahebkar, A.; Rastin, M. Lactobacillus rhamnosus and Lactobacillus delbrueckii ameliorate the expression of miR-155 and miR-181a in SLE patients. Journal of Functional Foods 2018, 48, 228–233. [Google Scholar] [CrossRef]
- Jang, H.R.; Gandolfo, M.T.; Ko, G.J.; Satpute, S.; Racusen, L.; Rabb, H. Early exposure to germs modifies kidney damage and inflammation after experimental ischemia-reperfusion injury. Am J Physiol Renal Physiol 2009, 297, F1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Emal, D.; Rampanelli, E.; Stroo, I.; Butter, L.M.; Teske, G.J.; Claessen, N.; Stokman, G.; Florquin, S.; Leemans, J.C.; Dessing, M.C. Depletion of Gut Microbiota Protects against Renal Ischemia-Reperfusion Injury. J Am Soc Nephrol 2017, 28, 1450–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.G.; Cho, W.Y.; Chung, S.M.; Choi, Y.E.; Fang, Y.; Park, M.S.; Park, S.J.; Ko, Y.S.; Lee, H.Y.; Yang, J.; et al. Altered gut microbiome plays an important role in AKI to CKD transition in aged mice. Front Med (Lausanne) 2023, 10, 1238960. [Google Scholar] [CrossRef] [PubMed]
- Gharaie, S.; Lee, K.; Newman-Rivera, A.M.; Xu, J.; Patel, S.K.; Gooya, M.; Arend, L.J.; Raj, D.S.; Pluznick, J.; Parikh, C.; et al. Microbiome modulation after severe acute kidney injury accelerates functional recovery and decreases kidney fibrosis. Kidney International 2023, 104, 470–491. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; de Pasquale, C.; Ekinci, E.I.; Coughlan, M.T. Gut microbiome, prebiotics, intestinal permeability and diabetes complications. Best Practice & Research Clinical Endocrinology & Metabolism 2021, 35, 101507. [Google Scholar] [CrossRef]
- Tao, S.; Li, L.; Li, L.; Liu, Y.; Ren, Q.; Shi, M.; Liu, J.; Jiang, J.; Ma, H.; Huang, Z.; et al. Understanding the gut-kidney axis among biopsy-proven diabetic nephropathy, type 2 diabetes mellitus and healthy controls: an analysis of the gut microbiota composition. Acta Diabetol 2019, 56, 581–592. [Google Scholar] [CrossRef]
- He, X.; Sun, J.; Liu, C.; Yu, X.; Li, H.; Zhang, W.; Li, Y.; Geng, Y.; Wang, Z. Compositional Alterations of Gut Microbiota in Patients with Diabetic Kidney Disease and Type 2 Diabetes Mellitus. Diabetes Metab Syndr Obes 2022, 15, 755–765. [Google Scholar] [CrossRef]
- Du, X.; Liu, J.; Xue, Y.; Kong, X.; Lv, C.; Li, Z.; Huang, Y.; Wang, B. Alteration of gut microbial profile in patients with diabetic nephropathy. Endocrine 2021, 73, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Salguero, M.V.; Al-Obaide, M.A.I.; Singh, R.; Siepmann, T.; Vasylyeva, T.L. Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp Ther Med 2019, 18, 3461–3469. [Google Scholar] [CrossRef]
- Xu, K.-Y.; Xia, G.-H.; Lu, J.-Q.; Chen, M.-X.; Zhen, X.; Wang, S.; You, C.; Nie, J.; Zhou, H.-W.; Yin, J. Impaired renal function and dysbiosis of gut microbiota contribute to increased trimethylamine-N-oxide in chronic kidney disease patients. Scientific Reports 2017, 7, 1445. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; DeSantis, T.Z.; Ni, Z.; Nguyen, T.-H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney International 2013, 83, 308–315. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of Urease- and Uricase-Containing, Indole- and p-Cresol-Forming and Contraction of Short-Chain Fatty Acid-Producing Intestinal Microbiota in ESRD. American Journal of Nephrology 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.B.; Allegretti, A.S.; Nigwekar, S.U.; Kalim, S.; Zhao, S.; Lelouvier, B.; Servant, F.; Serena, G.; Thadhani, R.I.; Raj, D.S.; et al. Blood Microbiome Profile in CKD : A Pilot Study. Clin J Am Soc Nephrol 2019, 14, 692–701. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunology 2017, 10, 18–26. [Google Scholar] [CrossRef]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The Role of Enterobacteriaceae in Gut Microbiota Dysbiosis in Inflammatory Bowel Diseases. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Stanford, J.; Charlton, K.; Stefoska-Needham, A.; Ibrahim, R.; Lambert, K. The gut microbiota profile of adults with kidney disease and kidney stones: a systematic review of the literature. BMC Nephrology 2020, 21, 215. [Google Scholar] [CrossRef]
- Hoyles, L.; Fernandez-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat Med 2018, 24, 1070–1080. [Google Scholar] [CrossRef]
- Salosensaari, A.; Laitinen, V.; Havulinna, A.S.; Meric, G.; Cheng, S.; Perola, M.; Valsta, L.; Alfthan, G.; Inouye, M.; Watrous, J.D.; et al. Taxonomic signatures of cause-specific mortality risk in human gut microbiome. Nature Communications 2021, 12, 2671. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, B.R.; Rocchetti, M.T.; De Angelis, M.; Cosola, C.; Marzocco, S.; Di Micco, L.; di Bari, I.; Accetturo, M.; Vacca, M.; Gobbetti, M.; et al. Nutritional Therapy Modulates Intestinal Microbiota and Reduces Serum Levels of Total and Free Indoxyl Sulfate and P-Cresyl Sulfate in Chronic Kidney Disease (Medika Study). J Clin Med 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, Y.; Tamura, H.; Furuie, K.; Kuraoka, S.; Nagata, H.; Nakazato, H. Cyclosporine therapy could be considered for membranoproliferative glomerulonephritis with immunoglobulin A deposits: a case report. BMC Nephrology 2022, 23, 358. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, D.; Law, H.K.-W.; Wu, Y.; Zhu, G.-h.; Huang, W.-y.; Kang, Y. Integrative Analysis of Gut Microbiota and Fecal Metabolites in Rats after Prednisone Treatment. Microbiology Spectrum 2021, 9, e00650–00621. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wang, L.; Bhat, O.M.; Lohner, H.; Li, P.L. Differential effects of short chain fatty acids on endothelial Nlrp3 inflammasome activation and neointima formation: Antioxidant action of butyrate. Redox Biol 2018, 16, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-Chain Fatty Acids Manifest Stimulative and Protective Effects on Intestinal Barrier Function Through the Inhibition of NLRP3 Inflammasome and Autophagy. Cell Physiol Biochem 2018, 49, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Wu, S.; Yan, C.; Zhao, C.; Jin, H.; Yan, N.; Xu, J.; Wu, Y.; Li, C.; Shao, Q.; et al. Butyrate upregulates the TLR4 expression and the phosphorylation of MAPKs and NK-kappaB in colon cancer cell in vitro. Oncol Lett 2018, 16, 4439–4447. [Google Scholar] [CrossRef]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr Res 2008, 28, 321–328. [Google Scholar] [CrossRef]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-chain fatty acids suppress lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines through inhibition of NF-kappaB pathway in RAW264.7 cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef]
- Lepenies, J.; Eardley, K.S.; Kienitz, T.; Hewison, M.; Ihl, T.; Stewart, P.M.; Cockwell, P.; Quinkler, M. Renal TLR4 mRNA expression correlates with inflammatory marker MCP-1 and profibrotic molecule TGF-beta(1) in patients with chronic kidney disease. Nephron Clin Pract 2011, 119, c97–c104. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, T.; Lawson, B.R. Toll-like receptors and kidney diseases. Inflamm Allergy Drug Targets 2009, 8, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.A.; Steinmetz, O.M.; Ooi, J.D.; Gan, P.Y.; O’Sullivan, K.M.; Visvanathan, K.; Akira, S.; Kitching, A.R.; Holdsworth, S.R. Toll-like receptor 9 enhances nephritogenic immunity and glomerular leukocyte recruitment, exacerbating experimental crescentic glomerulonephritis. Am J Pathol 2010, 177, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.A.; van der Veen, B.S.; O’Sullivan, K.M.; Gan, P.Y.; Ooi, J.D.; Heeringa, P.; Satchell, S.C.; Mathieson, P.W.; Saleem, M.A.; Visvanathan, K.; et al. Intrinsic renal cell and leukocyte-derived TLR4 aggravate experimental anti-MPO glomerulonephritis. Kidney international 2010, 78, 1263–1274. [Google Scholar] [CrossRef]
- O’Sullivan, K.M.; Ford, S.L.; Longano, A.; Kitching, A.R.; Holdsworth, S.R. Intrarenal Toll-like receptor 4 and Toll-like receptor 2 expression correlates with injury in antineutrophil cytoplasmic antibody-associated vasculitis. Am J Physiol Renal Physiol 2018, 315, F1283–F1294. [Google Scholar] [CrossRef]
- Rutting, S.; Xenaki, D.; Malouf, M.; Horvat, J.C.; Wood, L.G.; Hansbro, P.M.; Oliver, B.G. Short-chain fatty acids increase TNFalpha-induced inflammation in primary human lung mesenchymal cells through the activation of p38 MAPK. Am J Physiol Lung Cell Mol Physiol 2019, 316, L157–L174. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota metabolite butyrate constrains neutrophil functions and ameliorates mucosal inflammation in inflammatory bowel disease. Gut microbes 2021, 13, 1968257. [Google Scholar] [CrossRef]
- Zhang, D.; Frenette, P.S. Cross talk between neutrophils and the microbiota. Blood 2019, 133, 2168–2177. [Google Scholar] [CrossRef]
- Ruth, A.J.; Kitching, A.R.; Kwan, R.Y.; Odobasic, D.; Ooi, J.D.; Timoshanko, J.R.; Hickey, M.J.; Holdsworth, S.R. Anti-neutrophil cytoplasmic antibodies and effector CD4+ cells play nonredundant roles in anti-myeloperoxidase crescentic glomerulonephritis. J Am Soc Nephrol 2006, 17, 1940–1949. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J Nutr Biochem 2011, 22, 849–855. [Google Scholar] [CrossRef]
- Hamam, H.J.; Khan, M.A.; Palaniyar, N. Histone Acetylation Promotes Neutrophil Extracellular Trap Formation. Biomolecules 2019, 9. [Google Scholar] [CrossRef]
- O’Sullivan, K.M.; Holdsworth, S.R. Neutrophil Extracellular Traps: A Potential Therapeutic Target in MPO-ANCA Associated Vasculitis? Front Immunol 2021, 12, 635188. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly, Y.M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PLoS One 2011, 6, e21205. [Google Scholar] [CrossRef] [PubMed]
- Kamp, M.E.; Shim, R.; Nicholls, A.J.; Oliveira, A.C.; Mason, L.J.; Binge, L.; Mackay, C.R.; Wong, C.H. G Protein-Coupled Receptor 43 Modulates Neutrophil Recruitment during Acute Inflammation. PLoS One 2016, 11, e0163750. [Google Scholar] [CrossRef]
- Kobayashi, M.; Mikami, D.; Kimura, H.; Kamiyama, K.; Morikawa, Y.; Yokoi, S.; Kasuno, K.; Takahashi, N.; Taniguchi, T.; Iwano, M. Short-chain fatty acids, GPR41 and GPR43 ligands, inhibit TNF-alpha-induced MCP-1 expression by modulating p38 and JNK signaling pathways in human renal cortical epithelial cells. Biochem Biophys Res Commun 2017, 486, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem 2003, 278, 25481–25489. [Google Scholar] [CrossRef]
- Liu, J.; Gu, Q.H.; Cui, Z.; Zhao, M.H.; Jia, X.Y. Short-chain fatty acids ameliorate experimental anti-glomerular basement membrane disease. Clin Immunol 2024, 259, 109903. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal immunology 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Asarat, M.; Apostolopoulos, V.; Vasiljevic, T.; Donkor, O. Short-Chain Fatty Acids Regulate Cytokines and Th17/Treg Cells in Human Peripheral Blood Mononuclear Cells in vitro. Immunol Invest 2016, 45, 205–222. [Google Scholar] [CrossRef]
- Schiweck, C.; Edwin Thanarajah, S.; Aichholzer, M.; Matura, S.; Reif, A.; Vrieze, E.; Weigert, A.; Visekruna, A. Regulation of CD4(+) and CD8(+) T Cell Biology by Short-Chain Fatty Acids and Its Relevance for Autoimmune Pathology. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Yoshie, O.; Matsushima, K. CCR4 and its ligands: from bench to bedside. Int Immunol 2015, 27, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Veldkamp, C.T.; Seibert, C.; Peterson, F.C.; Sakmar, T.P.; Volkman, B.F. Recognition of a CXCR4 sulfotyrosine by the chemokine stromal cell-derived factor-1alpha (SDF-1alpha/CXCL12). J Mol Biol 2006, 359, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Strazza, M.; Azoulay-Alfaguter, I.; Silverman, G.J.; Mor, A. T cell chemokine receptor patterns as pathogenic signatures in autoimmunity. Discovery medicine 2015, 19 103, 117–125. [Google Scholar]
- García-Cuesta, E.M.; Santiago, C.A.; Vallejo-Díaz, J.; Juarranz, Y.; Rodríguez-Frade, J.M.; Mellado, M. The Role of the CXCL12/CXCR4/ACKR3 Axis in Autoimmune Diseases. Frontiers in Endocrinology 2019, 10. [Google Scholar] [CrossRef]
- Fallahi, P.; Ferrari, S.M.; Ragusa, F.; Ruffilli, I.; Elia, G.; Paparo, S.R.; Antonelli, A. Th1 chemokines in autoimmune endocrine disorders. The Journal of clinical endocrinology and metabolism 2019. [Google Scholar] [CrossRef] [PubMed]
- Sato, W. Chemokine receptors on T cells in multiple sclerosis. Clinical and Experimental Neuroimmunology 2014, 5. [Google Scholar] [CrossRef]
- Tan, D.S.; Gan, P.Y.; O’Sullivan, K.M.; Hammett, M.V.; Summers, S.A.; Ooi, J.D.; Lundgren, B.A.; Boyd, R.L.; Scott, H.S.; Kitching, A.R.; et al. Thymic deletion and regulatory T cells prevent antimyeloperoxidase GN. J Am Soc Nephrol 2013, 24, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Mendez-Garcia, C.; Rojo, D.; Barbas, C.; Moya, A. Antibiotic use and microbiome function. Biochem Pharmacol 2017, 134, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.F.; Aden, L.A.; Barbaro, N.R.; Van Beusecum, J.P.; Xiao, L.; Simmons, A.J.; Warden, C.; Pasic, L.; Himmel, L.E.; Washington, M.K.; et al. High dietary salt-induced dendritic cell activation underlies microbial dysbiosis-associated hypertension. JCI Insight 2019, 5. [Google Scholar] [CrossRef]
- Treuting, P.M.; Dintzis, S.; Montine, K.S. Comparative anatomy and histology: a mouse, rat, and human atlas; Academic Press, 2017. [Google Scholar]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Krych, L.; Hansen, C.H.; Hansen, A.K.; van den Berg, F.W.; Nielsen, D.S. Quantitatively different, yet qualitatively alike: a meta-analysis of the mouse core gut microbiome with a view towards the human gut microbiome. PLoS One 2013, 8, e62578. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Wang, S.; Solberg Woods, L.C.; Seshie, O.; Chung, S.T.; Shively, C.A.; Register, T.C.; Craft, S.; McClain, D.A.; Yadav, H. Comparative Microbiome Signatures and Short-Chain Fatty Acids in Mouse, Rat, Non-human Primate, and Human Feces. Front Microbiol 2018, 9, 2897. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, S.; Li, S.; Zhao, L.; Hao, Y.; Qin, J.; Zhang, L.; Zhang, C.; Bian, W.; Zuo, L.; et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut 2020, 69, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, S.; Kim, B.; Holzapfel, W.H.; Hyun, C.K. Disturbance of lipid metabolism in germ-free mice transplanted with gut microbiota of DSS-induced colitis mice. PLoS One 2023, 18, e0280850. [Google Scholar] [CrossRef] [PubMed]
- Luczynski, P.; McVey Neufeld, K.A.; Oriach, C.S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Growing up in a Bubble: Using Germ-Free Animals to Assess the Influence of the Gut Microbiota on Brain and Behavior. Int J Neuropsychopharmacol 2016, 19. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, C.E.; Ritz, N.L.; Martin, J.A.; Wardill, H.R.; Cryan, J.F.; Clarke, G. Investigating causality with fecal microbiota transplantation in rodents: applications, recommendations and pitfalls. Gut Microbes 2021, 13, 1941711. [Google Scholar] [CrossRef] [PubMed]
- Staley, C.; Kaiser, T.; Beura, L.K.; Hamilton, M.J.; Weingarden, A.R.; Bobr, A.; Kang, J.; Masopust, D.; Sadowsky, M.J.; Khoruts, A. Stable engraftment of human microbiota into mice with a single oral gavage following antibiotic conditioning. Microbiome 2017, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Nicolson, S.W.; Jackson, S. The effect of different oral antibiotics on the gastrointestinal microflora of a wild rodent (Aethomys namaquensis). Comp Biochem Physiol A Mol Integr Physiol 2004, 138, 475–483. [Google Scholar] [CrossRef]
- Shekhar, S.; Petersen, F.C. The Dark Side of Antibiotics: Adverse Effects on the Infant Immune Defense Against Infection. Front Pediatr 2020, 8, 544460. [Google Scholar] [CrossRef]
- Muralitharan, R.R.; Snelson, M.; Meric, G.; Coughlan, M.T.; Marques, F.Z. Guidelines for microbiome studies in renal physiology. Am J Physiol Renal Physiol 2023, 325, F345–F362. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.; Copio, J.N.; Shi, Y.; Kang, S.; Franklin, C.L.; Ericsson, A.C. Reduced housing density improves statistical power of murine gut microbiota studies. Cell Rep 2022, 39, 110783. [Google Scholar] [CrossRef] [PubMed]
- Hylander, B.L.; Qiao, G.; Cortes Gomez, E.; Singh, P.; Repasky, E.A. Housing temperature plays a critical role in determining gut microbiome composition in research mice: Implications for experimental reproducibility. Biochimie 2023, 210, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Shen, T.D.; Chen, E.Z.; Bittinger, K.; Bailey, A.; Roggiani, M.; Sirota-Madi, A.; Friedman, E.S.; Chau, L.; Lin, A.; et al. A role for bacterial urease in gut dysbiosis and Crohn’s disease. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Secombe, K.R.; Al-Qadami, G.H.; Subramaniam, C.B.; Bowen, J.M.; Scott, J.; Van Sebille, Y.Z.A.; Snelson, M.; Cowan, C.; Clarke, G.; Gheorghe, C.E.; et al. Guidelines for reporting on animal fecal transplantation (GRAFT) studies: recommendations from a systematic review of murine transplantation protocols. Gut Microbes 2021, 13, 1979878. [Google Scholar] [CrossRef] [PubMed]
- Picard, K.; Barreto Silva, M.I.; Mager, D.; Richard, C. Dietary Potassium Intake and Risk of Chronic Kidney Disease Progression in Predialysis Patients with Chronic Kidney Disease: A Systematic Review. Adv Nutr 2020, 11, 1002–1015. [Google Scholar] [CrossRef]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Vieira, S.M.; Pagovich, O.E.; Kriegel, M.A. Diet, microbiota and autoimmune diseases. Lupus 2014, 23, 518–526. [Google Scholar] [CrossRef]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacological research 2013, 69, 52–60. [Google Scholar] [CrossRef]
- Cigarrán Guldris, S.; Latorre Catalá, J.A.; Sanjurjo Amado, A.; Menéndez Granados, N.; Piñeiro Varela, E. Fibre Intake in Chronic Kidney Disease: What Fibre Should We Recommend? Nutrients 2022, 14, 4419. [Google Scholar] [CrossRef]
- Bell, K.J.; Saad, S.; Tillett, B.J.; McGuire, H.M.; Bordbar, S.; Yap, Y.A.; Nguyen, L.T.; Wilkins, M.R.; Corley, S.; Brodie, S.; et al. Metabolite-based dietary supplementation in human type 1 diabetes is associated with microbiota and immune modulation. Microbiome 2022, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Jama, H.A.; Rhys-Jones, D.; Nakai, M.; Yao, C.K.; Climie, R.E.; Sata, Y.; Anderson, D.; Creek, D.J.; Head, G.A.; Kaye, D.M.; et al. Prebiotic intervention with HAMSAB in untreated essential hypertensive patients assessed in a phase II randomized trial. Nature Cardiovascular Research 2023, 2, 35–43. [Google Scholar] [CrossRef]
- Su, G.; Qin, X.; Yang, C.; Sabatino, A.; Kelly, J.T.; Avesani, C.M.; Carrero, J.J. Fiber intake and health in people with chronic kidney disease. Clin Kidney J 2022, 15, 213–225. [Google Scholar] [CrossRef] [PubMed]
- El Amouri, A.; Snauwaert, E.; Foulon, A.; Vande Moortel, C.; Van Dyck, M.; Van Hoeck, K.; Godefroid, N.; Glorieux, G.; Van Biesen, W.; Vande Walle, J.; et al. Dietary Fibre Intake Is Associated with Serum Levels of Uraemic Toxins in Children with Chronic Kidney Disease. Toxins (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Krishnamurthy, V.M.; Wei, G.; Baird, B.C.; Murtaugh, M.; Chonchol, M.B.; Raphael, K.L.; Greene, T.; Beddhu, S. High dietary fiber intake is associated with decreased inflammation and all-cause mortality in patients with chronic kidney disease. Kidney Int 2012, 81, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Huang, X.; Riserus, U.; Krishnamurthy, V.M.; Cederholm, T.; Ärnlöv, J.; Lindholm, B.; Sjögren, P.; Carrero, J.J. Dietary fiber, kidney function, inflammation, and mortality risk. Clinical journal of the American Society of Nephrology: CJASN 2014, 9, 2104. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, M.; Song, J.; Liu, S.; Wang, Y.; Su, X.; Wei, K.; Xu, Z.; Li, H.; Wang, Z. The prebiotic effects of soluble dietary fiber mixture on renal anemia and the gut microbiota in end-stage renal disease patients on maintenance hemodialysis: a prospective, randomized, placebo-controlled study. J Transl Med 2022, 20, 599. [Google Scholar] [CrossRef] [PubMed]
- Kaesler, N.; Baid-Agrawal, S.; Grams, S.; Nadal, J.; Schmid, M.; Schneider, M.P.; Eckardt, K.U.; Floege, J.; Bergmann, M.M.; Schlieper, G.; et al. Low adherence to CKD-specific dietary recommendations associates with impaired kidney function, dyslipidemia, and inflammation. European journal of clinical nutrition 2021, 75, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Mun, K.H.; Yu, G.I.; Choi, B.Y.; Kim, M.K.; Shin, M.H.; Shin, D.H. Association of Dietary Potassium Intake with the Development of Chronic Kidney Disease and Renal Function in Patients with Mildly Decreased Kidney Function: The Korean Multi-Rural Communities Cohort Study. Med Sci Monit 2019, 25, 1061–1070. [Google Scholar] [CrossRef]
- Dobranowski, P.A.; Stintzi, A. Resistant starch, microbiome, and precision modulation. Gut microbes 2021, 13, 1926842. [Google Scholar] [CrossRef]
- Kalmokoff, M.; Zwicker, B.; O’Hara, M.; Matias, F.; Green, J.; Shastri, P.; Green-Johnson, J.; Brooks, S.P. Temporal change in the gut community of rats fed high amylose cornstarch is driven by endogenous urea rather than strictly on carbohydrate availability. J Appl Microbiol 2013, 114, 1516–1528. [Google Scholar] [CrossRef]
- Karaduta, O.; Glazko, G.; Dvanajscak, Z.; Arthur, J.; Mackintosh, S.; Orr, L.; Rahmatallah, Y.; Yeruva, L.; Tackett, A.; Zybailov, B. Resistant starch slows the progression of CKD in the 5/6 nephrectomy mouse model. Physiol Rep 2020, 8, e14610. [Google Scholar] [CrossRef]
- Meng, Y.; Bai, H.; Yu, Q.; Yan, J.; Zhao, L.; Wang, S.; Li, Z.; Wang, Q.; Chen, L. High-Resistant Starch, Low-Protein Flour Intervention on Patients With Early Type 2 Diabetic Nephropathy: A Randomized Trial. J Ren Nutr 2019, 29, 386–393. [Google Scholar] [CrossRef]
- Laffin, M.R.; Tayebi Khosroshahi, H.; Park, H.; Laffin, L.J.; Madsen, K.; Kafil, H.S.; Abedi, B.; Shiralizadeh, S.; Vaziri, N.D. Amylose resistant starch (HAM-RS2) supplementation increases the proportion of Faecalibacterium bacteria in end-stage renal disease patients: Microbial analysis from a randomized placebo-controlled trial. Hemodialysis International 2019, 23, 343–347. [Google Scholar] [CrossRef]
- Machado, M.G.; Sencio, V.; Trottein, F. Short-Chain Fatty Acids as a Potential Treatment for Infections: a Closer Look at the Lungs. Infection and immunity 2021, 89, e0018821. [Google Scholar] [CrossRef]
- Lin, X.B.; Farhangfar, A.; Valcheva, R.; Sawyer, M.B.; Dieleman, L.; Schieber, A.; Ganzle, M.G.; Baracos, V. The role of intestinal microbiota in development of irinotecan toxicity and in toxicity reduction through dietary fibres in rats. PLoS One 2014, 9, e83644. [Google Scholar] [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are Short Chain Fatty Acids in Gut Microbiota Defensive Players for Inflammation and Atherosclerosis? Journal of Atherosclerosis and Thrombosis 2017, 24, 660–672. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol 2019, 10, 277. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv Nutr 2018, 9, 21–29. [Google Scholar] [CrossRef]
- Gonzalez, A.; Krieg, R.; Massey, H.D.; Carl, D.; Ghosh, S.; Gehr, T.W.B.; Ghosh, S.S. Sodium butyrate ameliorates insulin resistance and renal failure in CKD rats by modulating intestinal permeability and mucin expression. Nephrol Dial Transplant 2019, 34, 783–794. [Google Scholar] [CrossRef]
- Gill, P.A.; Muir, J.G.; Gibson, P.R.; van Zelm, M.C. A randomized dietary intervention to increase colonic and peripheral blood SCFAs modulates the blood B- and T-cell compartments in healthy humans. The American Journal of Clinical Nutrition 2022, 116, 1354–1367. [Google Scholar] [CrossRef]
- Yang, R.; Chen, Z.; Cai, J. Fecal microbiota transplantation: Emerging applications in autoimmune diseases. J Autoimmun 2023, 141, 103038. [Google Scholar] [CrossRef]
- Zhang, X.; Ishikawa, D.; Ohkusa, T.; Fukuda, S.; Nagahara, A. Hot topics on fecal microbiota transplantation for the treatment of inflammatory bowel disease. Frontiers in medicine 2022, 9, 1068567. [Google Scholar] [CrossRef]
- Chi, M.; Ma, K.; Wang, J.; Ding, Z.; Li, Y.; Zhu, S.; Liang, X.; Zhang, Q.; Song, L.; Liu, C. The Immunomodulatory Effect of the Gut Microbiota in Kidney Disease. Journal of immunology research 2021, 2021, 5516035. [Google Scholar] [CrossRef]
- Khoruts, A.; Sadowsky, M.J. Understanding the mechanisms of faecal microbiota transplantation. Nature reviews. Gastroenterology & hepatology 2016, 13, 508–516. [Google Scholar] [CrossRef]
- Lauriero, G.; Abbad, L.; Vacca, M.; Celano, G.; Chemouny, J.M.; Calasso, M.; Berthelot, L.; Gesualdo, L.; De Angelis, M.; Monteiro, R.C. Fecal Microbiota Transplantation Modulates Renal Phenotype in the Humanized Mouse Model of IgA Nephropathy. Front Immunol 2021, 12, 694787. [Google Scholar] [CrossRef]
- Zhong, H.J.; Xie, X.; Chen, W.J.; Zhuang, Y.P.; Hu, X.; Cai, Y.L.; Zeng, H.L.; Xiao, C.; Li, Y.; Ding, Y.; et al. Washed microbiota transplantation improves renal function in patients with renal dysfunction: a retrospective cohort study. J Transl Med 2023, 21, 740. [Google Scholar] [CrossRef]
- Benech, N.; Koppe, L. Is there a place for faecal microbiota transplantation in chronic kidney disease? Nephrol Dial Transplant 2022, 37, 2303–2306. [Google Scholar] [CrossRef]
- Bian, J.; Liebert, A.; Bicknell, B.; Chen, X.-M.; Huang, C.; Pollock, C.A. Faecal Microbiota Transplantation and Chronic Kidney Disease. Nutrients 2022, 14, 2528. [Google Scholar] [CrossRef]
- Gargari, G.; Taverniti, V.; Balzaretti, S.; Ferrario, C.; Gardana, C.; Simonetti, P.; Guglielmetti, S. Consumption of a Bifidobacterium bifidum Strain for 4 Weeks Modulates Dominant Intestinal Bacterial Taxa and Fecal Butyrate in Healthy Adults. Appl Environ Microbiol 2016, 82, 5850–5859. [Google Scholar] [CrossRef]
- Berer, K.; Mues, M.; Koutrolos, M.; Rasbi, Z.A.; Boziki, M.; Johner, C.; Wekerle, H.; Krishnamoorthy, G. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature 2011, 479, 538–541. [Google Scholar] [CrossRef]
- Marar, C.; Starich, B.; Wirtz, D. Extracellular vesicles in immunomodulation and tumor progression. Nat Immunol 2021, 22, 560–570. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8. [Google Scholar] [CrossRef]
- Zara, M.; Guidetti, G.F.; Camera, M.; Canobbio, I.; Amadio, P.; Torti, M.; Tremoli, E.; Barbieri, S.S. Biology and Role of Extracellular Vesicles (EVs) in the Pathogenesis of Thrombosis. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Ailuno, G.; Baldassari, S.; Lai, F.; Florio, T.; Caviglioli, G. Exosomes and Extracellular Vesicles as Emerging Theranostic Platforms in Cancer Research. Cells 2020, 9. [Google Scholar] [CrossRef]
- Peng, Y.; Yin, S.; Wang, M. Extracellular vesicles of bacteria as potential targets for immune interventions. Hum Vaccin Immunother 2021, 17, 897–903. [Google Scholar] [CrossRef]
- Thongboonkerd, V. Roles for Exosome in Various Kidney Diseases and Disorders. Frontiers in pharmacology 2019, 10, 1655. [Google Scholar] [CrossRef]
- Lv, L.L.; Feng, Y.; Tang, T.T.; Liu, B.C. New insight into the role of extracellular vesicles in kidney disease. J Cell Mol Med 2019, 23, 731–739. [Google Scholar] [CrossRef]
- Kang, C.S.; Ban, M.; Choi, E.J.; Moon, H.G.; Jeon, J.S.; Kim, D.K.; Park, S.K.; Jeon, S.G.; Roh, T.Y.; Myung, S.J.; et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One 2013, 8, e76520. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; et al. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Exp Mol Med 2020, 52, 423–437. [Google Scholar] [CrossRef]
Figure 1.
The mechanism of the effects of SCFAs in the gut-kidney axis and immune cells. Dietary fibre is made up of insoluble and soluble components. The uptake of dietary fibre by intestinal microbiota increases the concentration of SCFAs in the intestine and encourages the growth of SCFA-producing bacteria. Acetate, butyrate and propionate are the most abundant SCFAs produced by gut bacteria in the large intestines, with a molar ratio of 60:20:20 respectively [38]. Renal cells, immune cells, and intestinal epithelial cells have SCFA-sensing GPCR, and upon binding can alter metabolic pathways (tricarboxylic acid cycle, β-oxidation, oxidative phosphorylation), epigenetic changes through histone deacetylase (HDAC) inhibition, phenotypes (M1 to M2 macrophages), and expression of inflammatory cytokines. SCFAs may indirectly induce renoprotection through their immunomodulatory properties such as reducing leukocyte recruitment, inflammation, and kidney injury.
Figure 1.
The mechanism of the effects of SCFAs in the gut-kidney axis and immune cells. Dietary fibre is made up of insoluble and soluble components. The uptake of dietary fibre by intestinal microbiota increases the concentration of SCFAs in the intestine and encourages the growth of SCFA-producing bacteria. Acetate, butyrate and propionate are the most abundant SCFAs produced by gut bacteria in the large intestines, with a molar ratio of 60:20:20 respectively [38]. Renal cells, immune cells, and intestinal epithelial cells have SCFA-sensing GPCR, and upon binding can alter metabolic pathways (tricarboxylic acid cycle, β-oxidation, oxidative phosphorylation), epigenetic changes through histone deacetylase (HDAC) inhibition, phenotypes (M1 to M2 macrophages), and expression of inflammatory cytokines. SCFAs may indirectly induce renoprotection through their immunomodulatory properties such as reducing leukocyte recruitment, inflammation, and kidney injury.
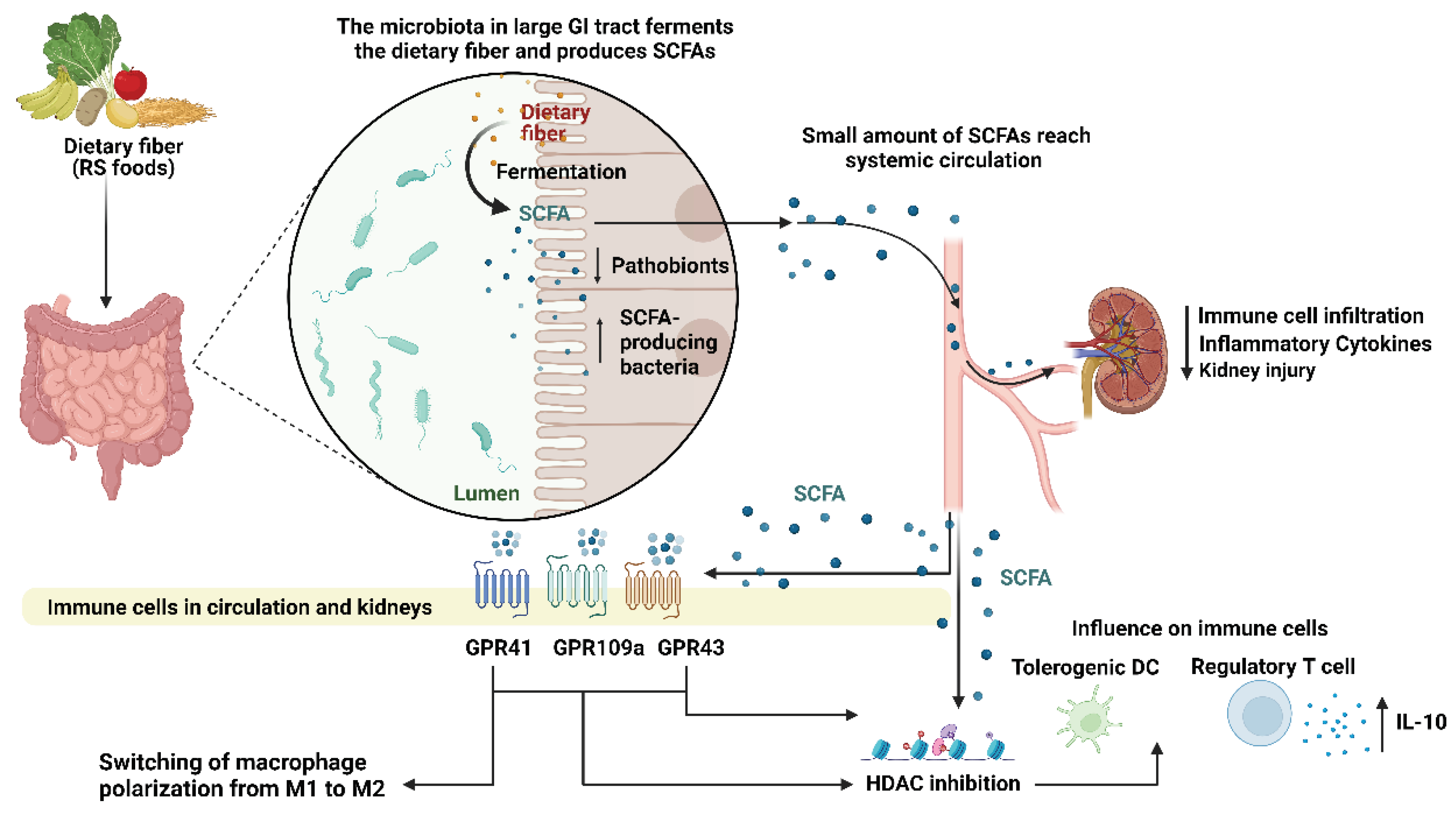
Figure 2.
Strategies for modulating the inflammatory response during autoimmune kidney disease. This includes dietary fiber intake, SCFAs (namely acetate, propionate, and butyrate), the transfer of gut microbial isolate, faecal microbial transplantation, and gut bacterial exosome therapy. (1) Dietary fibers and starches that are resistant to digestion in the small intestine promote gut microbiota, and the metabolized product (SCFAs) is in turn associated with gut microbiota homeostasis. (2) The transfer of immunomodulatory microbial isolate(s) can modulate inflammation and reduce the risk of introducing pathogenic bacteria. (3) Faecal microbiota transplantation from a healthy donor also introduces gut microbiota into the patient, thereby modulating inflammation. (4) Isolated and characterized exosomes from gut bacteria with known immunomodulatory properties can also serve as therapeutic targets. (5) As a result, all these potential targeted therapies could modulate CD4+ T cell differentiation, induce the activity of Treg responses, regulate B cell activity, and regulate neutrophils and other immune cells, all dampening the host immune responses. Type 2 responses suppress the Th1 responses against the disease and skew cytokine profiles, resulting in a dominant IL-4, IL-5, IL-9, and IL-13 response produced by expanded populations of alternatively activated macrophages. Abbreviations: FMT: Faecal Microbiota Transplantation, IFN-γ: Interferon-gamma, IL: Interleukin, M2: Alternative type Macrophage, NETosis: Formation of Neutrophil extracellular traps, SCFA: Short Chain Fatty Acids, Th: T helper cell, TNF-α: Tumour Necrosis Factor alpha. The figure was created using https://app.biorender.com.
Figure 2.
Strategies for modulating the inflammatory response during autoimmune kidney disease. This includes dietary fiber intake, SCFAs (namely acetate, propionate, and butyrate), the transfer of gut microbial isolate, faecal microbial transplantation, and gut bacterial exosome therapy. (1) Dietary fibers and starches that are resistant to digestion in the small intestine promote gut microbiota, and the metabolized product (SCFAs) is in turn associated with gut microbiota homeostasis. (2) The transfer of immunomodulatory microbial isolate(s) can modulate inflammation and reduce the risk of introducing pathogenic bacteria. (3) Faecal microbiota transplantation from a healthy donor also introduces gut microbiota into the patient, thereby modulating inflammation. (4) Isolated and characterized exosomes from gut bacteria with known immunomodulatory properties can also serve as therapeutic targets. (5) As a result, all these potential targeted therapies could modulate CD4+ T cell differentiation, induce the activity of Treg responses, regulate B cell activity, and regulate neutrophils and other immune cells, all dampening the host immune responses. Type 2 responses suppress the Th1 responses against the disease and skew cytokine profiles, resulting in a dominant IL-4, IL-5, IL-9, and IL-13 response produced by expanded populations of alternatively activated macrophages. Abbreviations: FMT: Faecal Microbiota Transplantation, IFN-γ: Interferon-gamma, IL: Interleukin, M2: Alternative type Macrophage, NETosis: Formation of Neutrophil extracellular traps, SCFA: Short Chain Fatty Acids, Th: T helper cell, TNF-α: Tumour Necrosis Factor alpha. The figure was created using https://app.biorender.com.
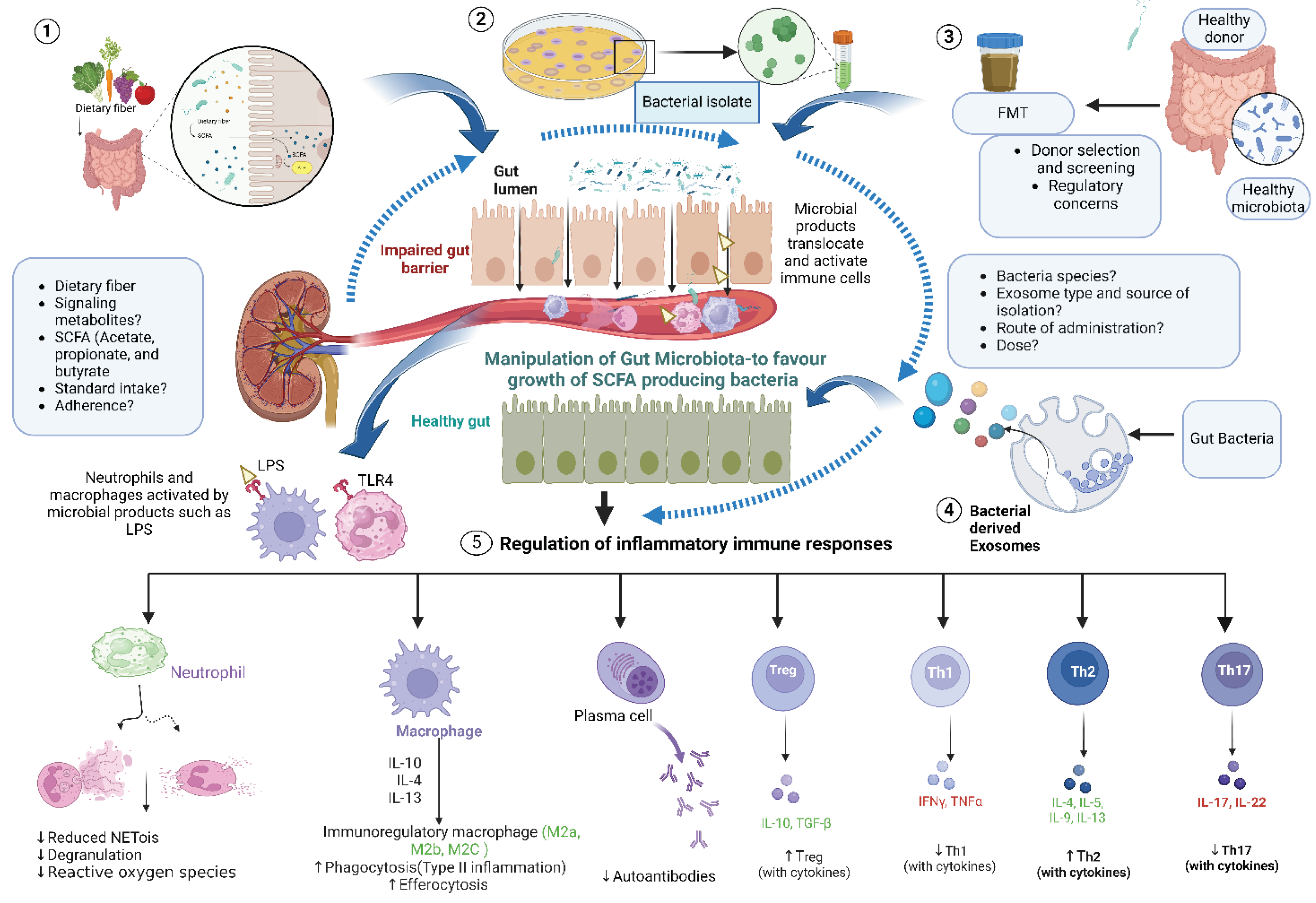
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



