
Submitted:
24 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
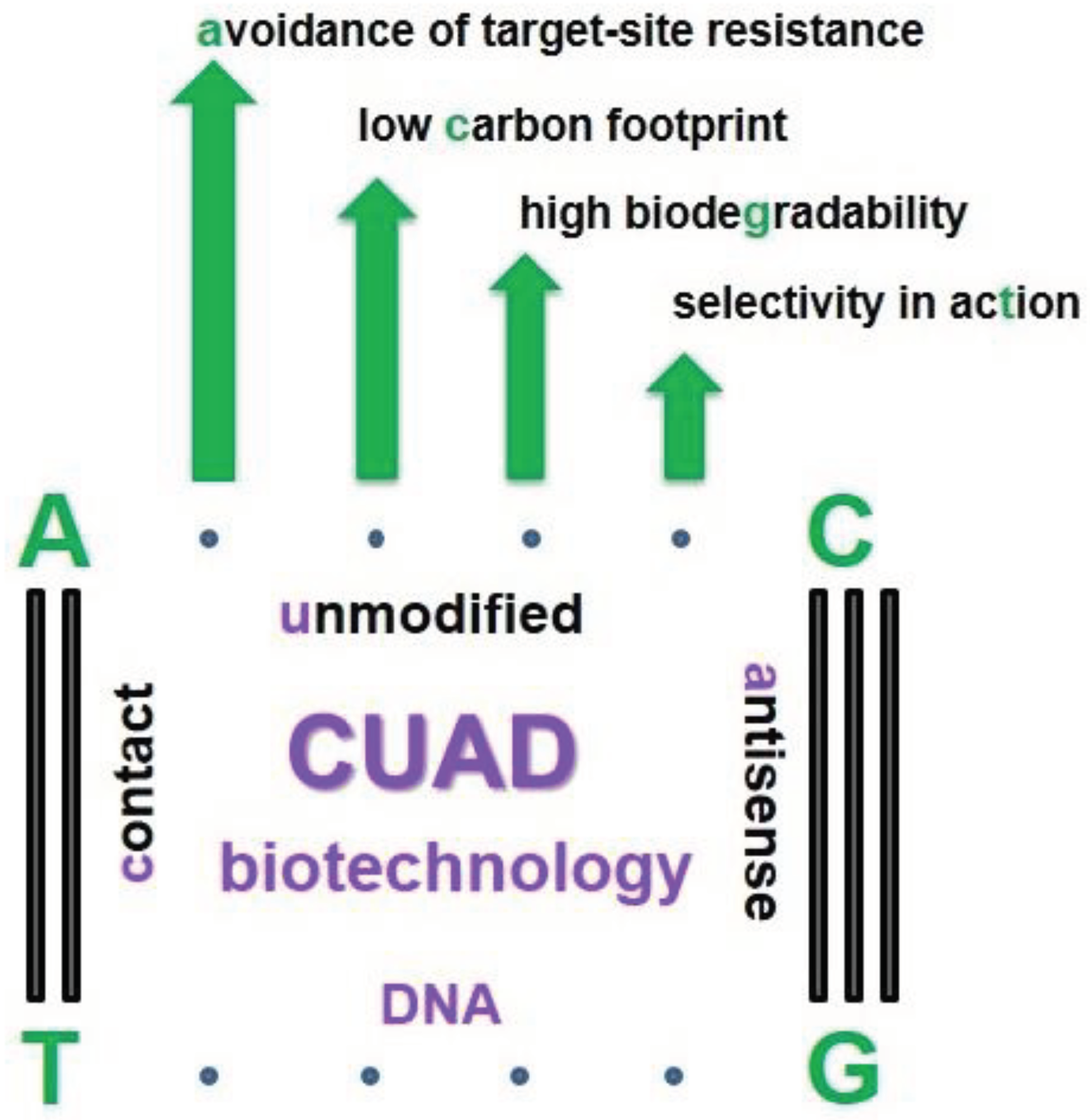
The idea of CUAD (contact unmodified antisense DNA) biotechnology and the use of oligonucleotide insecticides (briefly, olinscides, or DNA insecticides) was proposed on lepidopterans and gradually disseminated on hemipteran pests. As the next-generation platform for creation of insecticides, it has a number of unique characteristics (short unmodified antisense DNA as contact insecticide, insect rRNAs as target, DNA containment as mechanism of action) that distinguish it from existing classes of chemical insecticides. Oligonucleotide insecticides possess low carbon footprint, high safety for non-target organisms, rapid biodegradability, avoidance of target-site resistance, ensuring prosperity for agricultural countries that will implement this innovation. As the next-generation class of insecticides olinscides create opportunities that have never existed before in crop protection. Using flexible CUAD platform, any farmer, botanical garden or pest control company is capable of creating its own unique arsenal of insecticides, well-tailored for a particular population of insect pest. Most technological innovations start out very expensive and don't work well, but eventually start working well and become very cheap. CUAD biotechnology has come a long way for 16 years and is very close to the point to be implemented on a large scale against certain groups of pests. In this article we provide basic knowledge of use of oligonucleotide pesticides and list pests (mainly from suborder Sternorrhyncha, Hemiptera) which were successfully controlled by us recently.
Keywords:
CUAD (contact unmodified antisense DNA) biotechnology
; oligonucleotide insecticides
; ribosomal RNA
; DNA containment mechanism
; plant protection
Introduction
Partial possibilities of use of modified antisense oligonucleotides was first found by Paul Zamechnik and Mary Stephenson in 1978 on Rous sarcoma virus (Zamecnik and Stephenson 1978). One of eventual mechanisms of action of antisense oligonucleotides was detected a year later when Donis-Keller presented results showing that RNase H cleaves the RNA strand in RNA-DNA heteroduplexes in a site-specific manner (Donis-Keller 1979). It took 3 decades for unmodified antisense oligonucleotides to be conceptually applied in the form of CUAD (contact unmodified antisense DNA) biotechnology (Oberemok et al. 2008) and use of oligonucleotide insecticides (briefly, olinscides, or DNA insecticides) in plant protection (Manju et al. 2022, TriLink BioTechnologies 2024; Gal’chinsky et al., 2024) (Figure 1). In 2008 an equal sign was put between unmodified antisense DNA oligonucleotides and contact insecticides (Oberemok 2008). By that time, the development of phosphoramidite DNA synthesis (Hoose et al. 2023) made it possible to synthesize and test antisense DNA fragments on a large number of pests at an affordable price. Oligonucleotide insecticides were tested for the first time on spongy moth Lymantria dispar. Contact application of antisense DNA oligonucleotides targeting IAP genes has shown its effectiveness on both baculovirus-free and LdMNPV-infected gypsy moth caterpillars (Oberemok et al. 2016, Oberemok et al. 2017, Kumar et al. 2022).
In 2019, three important changes occurred that significantly advanced the development of CUAD biotechnology. First, rRNAs of insect pests began to be used as targets for oligonucleotide insecticides (this led to increase in the efficiency of oligonucleotide insecticides, since rRNA makes up 80% of all RNA in the cell) (Oberemok et al. 2019). Second, the length of the oligonucleotide insecticides was successfully reduced to 11 nucleotides providing sufficient selectivity in action (this helped to decrease the cost of oligonucleotide insecticides, since the yield of DNA synthesis for short DNA sequences is higher). Third, representatives of the suborder Sternorrhyncha, serious pests of agriculture and forestry all over the world, were found to be highly sensitive to oligonucleotide insecticides (Gal'chinsky et al. 2020, Useinov et al. 2020, Oberemok et al. 2022, Oberemok et al. 2023, Gal'chinsky et al. 2023, Puzanova et al. 2023).
In the course of research, we discovered that oligonucleotide insecticides act through DNA containment (DNAc) mechanism, which consists of two steps. At first step of DNAc, antisense DNA oligonucleotide (oligonucleotide insecticide) complementarily interacts with target rRNA (in other words, ‘arrests’ target rRNA) and interferes with normal functioning of ribosomes (‘arrested’ ribosomes); this process is accompanied with substantial insect pest mortality. After that we see target rRNA hypercompensation by the DNA-dependent RNA polymerase as the only way for the insect cell to fight for life in such a situation when target rRNAs and/or polycistronic rRNA transcripts are ‘arrested’ by antisense DNA oligonucleotides. At second step, RNase H cleaves target rRNA and substantial decrease in its concentration occurs; this step is also accompanied with substantial insect pest mortality (Gal'chinsky et al. 2024, Oberemok and Gal'chinsky 2024).
Oligonucleotide insecticides can be designed using DNAInsector program (dnainsector.com) or manually using sequences of pest rRNAs found in GenBank database. Phosphoramidite solid-phase and liquid-phase methods of oligonucleotide synthesis are used to obtain olinscides. Oligonucleotide insecticides are generally dissolved in nuclease-free water and usual concentration is 1 mg of olinscides per m2 of plant leaves containing insect pests. We believe that contact delivery of unmodified antisense DNA (CUADs) is much more efficient than oral delivery of unmodified antisense DNA (ODUADs) because of active DNases present in digestive tract of insects. Oligonucleotide insecticides are applied using hand sprayers or cold fog generators. Olinscides possess high selectivity in action, safety for non-target organisms, low carbon footprint, rapid biodegradability, and create opportunity for creation of insecticides with a long operational life based on conservative sequences of pest ribosomal RNA genes (Oberemok et al. 2019, Oberemok et al. 2022, Gal'chinsky et al. 2023, Puzanova et al. 2023, Gal'chinsky et al. 2024). The purpose of this article is a brief overview of experiments using oligonucleotide insecticides, which will help to understand the simplicity and effectiveness of CUAD biotechnology, as well as evaluate its high potential for green agriculture.
Lymantria dispar (Linnaeus, 1758)
The spongy moth (formerly known as gypsy moth) Lymantria dispar L. (Lepidoptera: Erebidae) is the notorious invasive polyphagous pest of the Holarctic region, infesting over 500 plant species and causing widespread loss of leaves in forests in Europe, Asia, North America, and parts of Africa (Martemyanov et al. 2019, Boukouvala et al. 2022). In Europe and North America for L. dispar the preferred hosts vary by region, but include species of Quercus, Salix, Populus, and Betula (Boukouvala et al. 2022).
Oligonucleotide insecticide oligoRIBO-11 (5′-TGC-GTT-CGA-AA-3′) targeting 5.8S rRNA in concentration 72 ng/µL leads to 46.9 ± 9.3% mortality rate of spongy moth larvae on the 6th day (Oberemok et al. 2019a).
Unaspis euonymi(Comstock, 1881)
The euonymous scale Unaspis euonymi Comstock (Hemiptera: Diaspididae) is the most common and most serious pest found on Euonymus plants (Salisbury et al. 2013). It is frequently encountered in dendrological nurseries, parks, and ornamental gardens.
Oligonucleotide insecticide oligoUE-11 (5′-ATA-CCG-ACG-AT-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 99.24 ± 1.32 % mortality rate of euonymous scale larvae on the 10th day (Oberemok et al. 2019b, Gal'chinsky et al. 2020).
Dynaspidiotus britannicus(Newstead, 1898)
The holly scale Dynaspidiotus britannicus Newstead (Hemiptera: Diaspididae) is often found on species of Buxus, Hedera, Ilex and Laurus (Nakahara 1982), on conifers (Ülgentürk et al. 2012, Kaydan et al. 2014), and also as a minor pest of olive trees, palms and ornamentals.
Oligonucleotide insecticide oligoDB-11 (5′-ATA-CCG-ACG-AT-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 82.44 ± 15.62 % mortality rate of holly scale larvae on the 10th day (Gal'chinsky et al. 2020).
Ceroplastes japonicus(Green, 1921)
The Japanese wax scale Ceroplastes japonicus Green (Hemiptera: Sternorrhyncha: Coccidae) is polyphagous pest of soft and hardwood trees, fruit trees, citrus trees, and ornamentals (Morales et al. 2016). The host plants most commonly infested by these insects are Citrus, Diospyros, Ilex, and Hedera (Pellizzari and Germain 2010).
Oligonucleotide insecticide oligoCJ-11 (5′-CGA-CCG-ACG-AA-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 78.82 ± 18.60 % mortality rate of Japanese wax scale larvae on the 10th day (Useinov et al. 2020).
Diaspis echinocacti(Bouche, 1833)
The invasive scale insect Diaspis echinocacti Bouché (Hemiptera: Diaspididae) is a major and specific pest of cactus pear species worldwide, recorded from 74 countries (Asia, Europe, North America and Africa) (García Morales et al. 2016). This pest was recorded on 58 plant species to the family Cactaceae: most commonly on Opuntia dillenii and O. ficus-indica (Mrabti et al. 2022, Aalaoui and Sbaghi 2023).
Oligonucleotide insecticide Cactus-NBG (5′-ATC-GCT-GCG-GA-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 84.2 ± 2.2 % mortality rate of D. echinocacti larvae on the 14th day (Plugatar et al. 2021).
Coccus hesperidum(Linnaeus, 1758)
The soft scale insect Coccus hesperidum L. (Hemiptera: Coccoidae) is cosmopolitan and polyphagous pest species (Villanueva et al. 2021, Kapranas et al. 2007) causing significant damage to citrus crops, mango, guava, and lychee (Kapranas et al. 2007). C. hesperidum may have the capacity to affect approximately 125 plant families (García Morales 2016).
Oligonucleotide insecticide Coccus-11 (5′-CGA-CCG-ACG-AA-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 95.59 ± 1.63 % mortality rate of C. hesperidum larvae on the 12th day (Oberemok et al. 2022).
Trioza alacris(Flor, 1861)
The bay sucker Trioza alacris Flor (Triozidae: Hemiptera) is oligophagous pest commonly feeding on economically valuable plants from the family Lauraceae: Laurus nobilis, Laurus azorica, Laurus novocanariensis, Cinnamomum camphora, and Laurus indicia, particularly in Mediterranean areas but also occurs in North and South America (Zeity 2018).
Oligonucleotide insecticide Alacris-11 (5′-CCA-CCG-GGT-AG-3′) targeting ITS2 of polycistronic rRNA transcript in concentration 100 ng/µL leads to 71.02 ± 5.21 % mortality rate of the bay sucker larvae on the 9th day.
Oligonucleotide insecticide Laura-11 (5′-GAC-ACG-CGC-GC-3′) targeting ITS2 of polycistronic rRNA transcript in concentration 100 ng/µL leads to 72.39 ± 6.48 % mortality rate of the bay sucker larvae on the 9th day (Novikov et al. 2022).
Icerya purchasi(Maskell, 1878)
The cottony cushion scale Icerya purchasi Maskell (Hemiptera: Monophlebidae) is a cosmopolitan pest native to Australia and New Zealand and is known to have affected over 200 different plant species (Kollar, et al. 2016). It is pest in several ornamentals and crops, such as Citrus reticulata, Artocarpus heterophyllus, Magnolia denudata, and Ficus altissima (Liu and Shi 2020).
Oligonucleotide insecticide oligoICER-11 (5′-ACA-CCG-ACG-AC-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 70.55 ± 0.77 % mortality rate of the cottony cushion scale larvae on the 10th day (Gal'chinsky et al. 2023).
Macrosiphoniella sanborni(Gillette, 1908)
The chrysanthemum aphid Macrosiphoniella sanborni Gillette (Hemiptera: Aphididae) is a major destructive oligophagous pest for chrysanthemums (Zhong et al. 2022).
Oligonucleotide insecticide Macsan-11 (5′-TGT-GTT-CGT-TA-3′) targeting ITS2 of polycistronic rRNA transcript gene in concentration 100 ng/µL leads to 97.38 ± 2.49 % mortality rate of the chrysanthemum aphid larvae on the 7th day (Puzanova et al. 2023).
Pseudococcus viburni(Signoret, 1875)
The mealybug Pseudococcus viburni Signoret (formerly known as Pseudococcus affinis) (Hemiptera: Pseudococcidae) is a polyphagous insect recorded from 236 host-plant genera in 89 families (García Morales et al. 2017). It is a pest of several temperate fruits, including grapes (Correa et al. 2012, Dapoto et al. 2011), apples (Ciampolini et al. 2011), and pears (Dapoto et al. 2011).
Oligonucleotide insecticide Alpha-11 (5′-GGT-CGC-GAC-GT-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 63.42 ± 3.1 % mortality rate of the mealybug larvae on the 14th day.
Oligonucleotide insecticide Beta-11 (5′-GGA-ATC-GAA-CC-3′) targeting 18S rRNA in concentration 100 ng/µL leads to 78.31 ± 4.5 % mortality rate of the mealybug larvae on the 3rd day.
Oligonucleotide insecticide Gamma-11 (5′-CCT-CAG-ACA-GG-3′) targeting 5.8S rRNA in concentration 100 ng/µL leads to 66.96 ± 2.9 % mortality rate of the mealybug larvae on the 14th day (Novikov et al. 2023).
Aonidia lauri(Bouche, 1833)
The laurel scale Aonidia lauri Bouché (Hemiptera: Diaspididae) occurs in almost all regions of the world, except Australia and the Pacific Islands (Danzig and Pellizzari 1998). It is a monophagous species that infests Laurus sp. and causes serious damage in all laurel-growing areas (Miller and Davidson 1990).
Oligonucleotide insecticide oligoAL-11 (5′-ATG-CCA-ACG-AT-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 98.19 ± 3.12 % mortality rate of laurel scale larvae on the 14th day (Gal'chinsky et al. 2024).
Tetranychus urticae (Koch, 1836), a case for oligonucleotide acaricides
The two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) is a polyphagous pest recorded from 1275 host plants from 70 genera representing several dozen botanical families (Migeon and Dorkeld 2021), either wild or cultivated, including vegetables, ornamental plants, crops, fruit trees and shrubs (El-Sayed et al. 2022).
Oligonucleotide acaricide Tur-3 (5′-AAA-ACA-TCA-AG-3′) targeting ITS2 of polycistronic rRNA transcript in concentration 100 ng/µL leads to 72.85 ± 4.55 % mortality rate of the mite on the 3rd day.
Oligonucleotide acaricide Turka (5′-AGC-GAC-GTC-GC-3′) targeting 28S rRNA in concentration 100 ng/µL leads to 77 ± 0.4 % mortality rate of the mite on the 3rd day (Novikov et al. 2023).
Conclusion
For the first time in the 155-year history of deoxyribonucleic acid, we can confidently say that DNA is also a contact insecticide with unique and advanced characteristics for agriculture and forestry. The simplicity, flexibility, and effectiveness of CUAD platform for sap-feeding pests (aphids, psyllids, soft scales, armored scales, mealybugs, whiteflies, etc.) is amazing. The use of unique conservative sequences of target rRNAs of insect pests will minimize the occurrence of target-site resistance in insect pests. Certain combinations of nitrogenous bases in an oligonucleotide insecticide will make it well-tailored to a single pest. The use of unmodified DNA as a natural polymer will minimize the toxicity load on ecosystems. Liquid-phase synthesis of DNA makes CUAD biotechnology very cheap already today. Obviously, more complex formulations of oligonucleotide insecticides with auxiliary substances will help enhance the effect of oligonucleotide insecticides on representatives from other orders of insects. Oligonucleotide insecticides are solving many problems of the 21st century in agriculture and countries that implement this innovation will prosper.
Funding
Research results obtained within the framework of a state assignment V.I. Vernadsky Crimean Federal University for 2024 and the planning period of 2024–2026 No. FZEG-2024–0001.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aalaoui EI, Sbaghi M. 2023. Potential of parasitoids to control Diaspis echinocacti (Bouché) (Hemiptera: Diaspididae) on Opuntia spp. cactus pear. Egypt J. Biol. Pest. Control. 33:57. [CrossRef]
- Boukouvala MC, Kavallieratos NG, Skourti A et al. 2022. Lymantria dispar (L.) (Lepidoptera: Erebidae): Current Status of Biology, Ecology, and Management in Europe with Notes from North America. Insects. 13:854. [CrossRef]
- Ciampolini M, Lupi D, Süss L. 2002. Pseudococcus viburni (Signoret) (Hemiptera: Coccoidea) nocivo in frutticoltura nell’Italia centrale Boll. Zool. Agrar. Bachic. 34:97-108.
- Correa MCG, Germain J-F, Malausa T et al. 2012. Molecular and morphological characterization of mealybugs (Hemiptera: Pseudococcidae) from Chilean vineyards Bull. Entomol. Res. 102:524-530.
- Danzig EM, Pellizzari G. 1998. Diaspididae. In: Kozár, F. (Eds.), Catalogue of Palaearctic Coccoidea. Budapest, Hungary: Hungarian Academy of Sciences, 172-370 p.
- Dapoto GL, Olave A, Bondoni M et al. 2011. Obscure mealybug (Pseudococcus viburni) in pear trees in the Alto Valle of Rio Negro and Neuquen, Argentina Acta Hortic. 909:497-504.
- Donis-Keller, H. 1979. Site specific enzymatic cleavage of RNA. Nucleic Acids Res. 7(1):179-192. 10.1093/nar/7.1.179.
- El-Sayed SM, Ahmed N, Selim S et al. 2022. Acaricidal and antioxidant activities of anise oil (Pimpinella anisum) and the oil’s effect on protease and acetylcholinesterase in the two-spotted spider mite (Tetranychus urticae Koch). Agriculture. 12:224.
- Gal'chinsky N, Useinov R, Yatskova E et al. 2020. A breakthrough in the efficiency of contact DNA insecticides: rapid high mortality rates in the sap-sucking insects Dynaspidiotus britannicus Comstock and Unaspis euonymi Newstead. Journal of Plant Protection Research. 60(2):220-223. [CrossRef]
- Gal'chinsky NV, Yatskova EV, Novikov IA et al. 2023. Icerya purchasi Maskell (Hemiptera: Monophlebidae) Control Using Low Carbon Footprint Oligonucleotide Insecticides. Int J Mol Sci. 24(14):11650. [CrossRef]
- Gal'chinsky NV, Yatskova EV, Novikov IA et al. 2024. Mixed insect pest populations of Diaspididae species under control of oligonucleotide insecticides: 3′-end nucleotide matters. Pesticide Biochemistry and Physiology. 105838. [CrossRef]
- García Morales M, Denno B, Miller DR et al. 2017. ScaleNet: a literature-based model of scale insect biology and systematics. www.scalenet.info (accessed 19 Jul 2017).
- García Morales M, Denno BD, Miller DR et al. 2016. ScaleNet: A literature-based model of scale insect biology and systematics. Database J. Biol. Database Curation. bav118. [CrossRef]
- Hoose A, Vellacott R, Storch M et al. 2023. DNA synthesis technologies to close the gene writing gap. Nat Rev Chem. 7:144–161. [CrossRef]
- Kapranas A, Morse JG, Pacheco P et al. 2007. Survey of brown soft scale Coccus hesperidum L. parasitoids in southern California citrus. Biol. Control. 42:288–299.
- Kaydan M, Ülgentürk S, Özdemir I et al. 2014. Coccoidea (Hemiptera) species in Bartın and Kastamonu Provinces. Bulletin of Plant Protection. 54(1):11–44.
- Kidd NAC. 1988. The Large Pine Aphid on Scots Pine in Britain. In: Berryman, A.A. (Eds), Dynamics of Forest Insect Populations. Population Ecology. Springer, Boston, MA. [CrossRef]
- Kollar J, Bakay L, Pastor M. 2016. First record of cottony cushion scale Icerya purchasi (Hemiptera, Monophlebidae) in Slovakia. Plant. Prot. Sci. 52:217–219.
- Kumar H, Sharma M, Chandel A. 2022. DNA Insecticides: Future of Crop Protection. ICAR- Indian Agricultural Research Institute, New Delhi-110012. Article ID: 37444. www.agrifoodmagazine.co.in (accessed 28 Feb 2024).
- Kuzio J, Pearson MN, Harwoo SH et al. 1999. Sequence and analysis of the genome of a baculovirus pathogenic for Lymantria Dispar. Virology. 253:17–34.
- Liu Y, Shi J. 2020. Predicting the Potential Global Geographical Distribution of Two Icerya Species under Climate Change. Forests. 11:684.
- Manju M, Nirosha V, Tullika T et al. 2022. DNA insecticides: an emerging tool in pest management. 4(9). https://agriallis.com/issue/volume-4-issue-9-september-2022/ (accessed 28 Feb 2024).
- Martemyanov V, Bykov R, Demenkova M et al. 2019. Genetic evidence of broad spreading of Lymantria dispar in the West Siberian Plain. PLOS ONE. 14(8):e0220954. [CrossRef]
- Migeon A, Dorkeld F. 2022. Spider Mites Web: a comprehensive database for the Tetranychidae. Available from https://www1.montpellier.inrae.fr/CBGP/spmweb/index.php (accessed 5 Dec 2022).
- Miller D, Davidson J. 1990. Armored scale insects as pests. In: Rosen, D. (Eds.), Armored scale insects, their biology, natural enemies and control. In World Crop Pests, 299-306 p.
- Morales MG, Denno BD, Miller DR et al. 2016. ScaleNet: A literature-based model of scale insect biology and systematics. Database, 2016, article ID bav118. [CrossRef]
- Mrabti I, Haddad N, Moghaddam M et al. 2022. Current status of the cactus scale Diaspis echinocacti Bouché (Hemiptera: Diaspididae) on Opuntia ficus-indica and its first morphological and microscopic description in Morocco. EPPO Bulletin. 52:718-724. 10.1111/epp.12902.
- Novikov A, Yatskova E, Bilyk A et al. 2023. Efficient Control of the Obscure Mealybug Pseudococcus viburni with DNA Insecticides. In Vitro Cellular & Developmental Biology-Animal; Springer: New York, NY, USA. 59:92–108. [CrossRef]
- Novikov IA, Yatskova EV, Useinov RZ et al. 2022. Efficient Bay Sucker (Trioza Alacris) Control with DNA Insecticides. In Vitro Cellular & Developmental Biology-Animal; Springer: New York, NY, USA. 58:43. [CrossRef]
- Oberemok VV, Gal’chinsky NV, Useinov RZ et al. 2023. Four Most Pathogenic Superfamilies of Insect Pests of Suborder Sternorrhyncha: Invisible Superplunderers of Plant Vitality. Insects. 14:462. [CrossRef]
- Oberemok VV, Gal’chinsky NV. 2024. Oligonucleotide insecticides (contact unmodified antisense DNA biotechnology) and RNA biocontrols (double-stranded RNA technology): newly born fraternal twins in plant protection. bioRxiv 584797. [CrossRef]
- Oberemok VV, Laikova KV, Gal'chinsky NV et al. 2019a. DNA insecticide developed from the Lymantria dispar 5.8S ribosomal RNA gene provides a novel biotechnology for plant protection. Sci Rep. 9(1):6197. [CrossRef]
- Oberemok VV, Laikova KV, Useinov RZ et al. 2019b. Insecticidal activity of three 10–12 nucleotides long antisense sequences from 5.8S ribosomal RNA gene of gypsy moth Lymantria dispar L. against its larvae. Journal of Plant Protection Research. 59(4):561-564. [CrossRef]
- Oberemok VV, Laikova KV, Zaitsev A.S et al. 2017. Molecular Alliance of Lymantria dispar Multiple Nucleopolyhedrovirus and a Short Unmodified Antisense Oligonucleotide of Its Anti-Apoptotic IAP-3 Gene: A Novel Approach for Gypsy Moth Control. Int. J. Mol. Sci. 18:2446. [CrossRef]
- Oberemok VV, Laikova KV, Zaitsev AS et al. 2016. The RING for gypsy moth control: Topical application of fragment of its nuclear polyhedrosis virus anti-apoptosis gene as insecticide. Pest. Biochem. Physiol. 131:32–39.
- Oberemok VV, Useinov RZ, Skorokhod OA 2022. Oligonucleotide Insecticides for Green Agriculture: Regulatory Role of Contact DNA in Plant–Insect Interactions. Int. J. Mol. Sci. 23:15681. [CrossRef]
- Oberemok, VV. 2008. Method of Elimination of Phyllophagous Insects from Order Lepidoptera. Ukraine Patent UA # 36, 445.
- Pellizzari G, Germain J. 2010. Scales (Hemiptera, Superfamily Coccoidea). Chapter 9.3. BioRisk 4:475-510. [CrossRef]
- Plugatar YuV, Chichkanova ES, Yatskova EV et al. 2021. An innovative method of Diaspis echinocacti Bouche control using DNA insecticide on Opuntia ficus-indica (L.) Mill. in the Nikitsky Botanical Garden, Crimea. South of Russia: ecology, development. 16(2):119–128. [CrossRef]
- Puzanova YV, Novikov IA, Bilyk AI et al. 2023. Perfect Complementarity Mechanism for Aphid Control: Oligonucleotide Insecticide Macsan-11 Selectively Causes High Mortality Rate for Macrosiphoniella sanborni Gillette. Int. J. Mol. Sci. 24:11690. [CrossRef]
- Salisbury A, Malumphy C, Halstead AJ. 2013. Euonymus scale Unaspis euonymi (Hemiptera:Diaspididae); an introduced pest of spindle (Euonymus)in Britain. Brit. J. Entomol. Nat. Hist. 26:211‒218.
- TriLink BioTechnologies, 2023. Feasibility of Antisense Oligonucleotides as DNAInsecticides. https://www.trilinkbiotech.com/blog/feasibility-of-antisense-oligonucleotides-as-dna-insecticides/ (accessed 28 Feb 2024).
- Ülgentürk S, Şahin Ö, Ayhan B et al. 2012. Scale insects species of Taurus cedar in Turkey. Turkish Journal of Entomology. 36:113–121.
- Useinov RZ, Gal'chinsky N, Yatskova E et al. 2020. To bee or not to bee: creating DNA insecticides to replace non-selective organophosphate insecticides for use against the soft scale insect Ceroplastes japonicus Green. Journal of Plant Protection Research. 60(4):406-409. [CrossRef]
- Villanueva RT, Gauthier N, Ahmed ZM. 2021. First Record of Coccus hesperidum L. (Hemiptera: Coccidae) in Industrial Hemp in Kentucky. Fla. Entomol. 103:514–515.
- Zamecnik PC, Stephenson ML. 1978. Inhibition of Rous sarcoma virus replication and cell transformation by a specific oligodeoxynucleotide. Proc. Natl Acad. Sci. 75:280–284.
- Zeity, M. 2018. First record of the bay sucker Trioza alacris Flor (Triozidae: Hemiptera) in Syria. EPPO Bulletin. 48(3):586–588. [CrossRef]
- Zhong J, Wang Y, Lu Y et al. 2022. Identification and Expression Analysis of Chemosensory Genes in the Antennal Transcriptome of Chrysanthemum Aphid Macrosiphoniella sanborni. Insects. 13:597. [CrossRef]
Figure 1.
Advanced features of CUAD biotechnology.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



