
Submitted:
24 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
CD44 is a type I transmembrane glycoprotein, which is associated with poor prognosis in various solid tumors. Since CD44 plays critical roles in tumor development by regulating cell adhesion, survival, proliferation, and stemness, it has been considered a target for tumor therapy. Anti-CD44 monoclonal antibodies (mAbs) have been developed and applied to antibody-drug conjugates and chimeric antigen receptor-T cell therapy. We previously developed anti‐pan-CD44 mAbs, C44Mab-5 and C44Mab-46, which can recognize both CD44 standard (CD44s) and variant isoforms. In the present study, a defucosylated mouse IgG2a version of the anti-pan-CD44 mAbs (5-mG2a-f and C44Mab-46-mG2a-f) was generated to evaluate the antitumor activities against CD44-positive cells. Both 5-mG2a-f and C44Mab-46-mG2a-f recognized CD44s-overexpressed CHO-K1 (CHO/CD44s) cells and esophageal tumor cell line (KYSE770) in flow cytometry. Furthermore, both 5-mG2a-f and C44Mab-46-mG2a-f could activate effector cells in the presence of CHO/CD44s cells and exhibited the complement‐dependent cytotoxicity against both CHO/CD44s and KYSE770 cells. Furthermore, the administration of 5-mG2a-f and C44Mab-46-mG2a-f significantly suppressed CHO/CD44s and KYSE770 xenograft tumor development compared with the control defucosylated mouse IgG2a. These results indicate that 5-mG2a-f and C44Mab-46-mG2a-f could exert antitumor activities against CD44-positive cancers and could be a promising therapeutic regimen for tumors.
Keywords:
CD44
; monoclonal antibody
; ADCC
; CDC
; esophageal tumor
; antitumor activity
1. Introduction
Overexpression of CD44 is observed in various solid tumors, which are involved in the tumor malignant progression through the promotion of cellular proliferation, invasiveness, and stemness via specific signaling pathways [1]. The diversity of CD44 molecular function is mediated by the alternative splicing [2]. CD44 is encoded in 20 exons. The first five (1–5) and the last five (16–20) are constant exons that generate the shortest CD44 standard (CD44s) isoform. The exons 6–15 are alternatively spliced and inserted into the CD44s as variant exons [3]. The CD44 splice variants with variant exons are designated as CD44 variant (CD44v) isoforms. The inclusion of CD44v exons in various combinations is regulated by receptor tyrosine kinase signaling and splicing activators of pre-mRNA [1].
CD44 possesses an extracellular domain (ectodomain), a transmembrane domain, and an intracellular cytoplasmic domain [4]. The ectodomain contains a hyaluronic acid (HA)-binding domain (HABD) that mediate cellular homing, adhesion, migration, and proliferation [4]. Both CD44s and CD44v isoforms have the HABD, and the HA binding causes conformational changes of the CD44, which results in the promotion of intracellular signaling pathways to regulate cell migration and proliferation [5]. In CD44v isoforms, the variant exons-encoding sequences form stem region which provides a co-receptor for various growth factors and cytokines. These functions activate specific signaling pathways to promote invasion and stemness [6]. Therefore, different isoforms of CD44v exhibit different functions according to the inserted variant exons in the CD44 ectodomain.
The relationship between CD44 expression and prognosis in cancer patients has been evaluated and showed both poor and favorable outcomes [7]. Currently, increased evidence suggests that overexpression of CD44 and its isoforms is an unfavorable indicator in cancer patients [7]. Conclusive evidence was obtained by meta-analyses which provided a predictive value of CD44 expression in cancer patients. In an analysis of nine studies including 583 cases of pancreatic cancer, CD44 overexpression was predictive of poor five-year overall survival, a more advanced stage, and more lymph node invasion but was not associated with differentiation, tumor size, and distance metastasis [8]. Furthermore, another meta-analysis investigated the prognostic significance of cancer stem cell markers including CD44 from 52 studies of ovarian cancer. The study concluded that CD44 overexpression was predictive of worse disease-free survival and resistance to chemotherapy [9].
Several anti-CD44 monoclonal antibodies (mAbs) have been developed for preclinical and clinical research for tumor therapy. An anti-pan-CD44 mAb (H4C4) reduced tumor growth, metastasis, and post−radiation recurrence in a human pancreatic tumor xenograft model [10]. A humanized anti-pan-CD44 mAb, RG7356, showed the cytotoxicity for B cell leukemia, but no cytotoxicity on normal B cells. Administration of RG7356 to immunodeficient mice engrafted with chronic lymphocytic leukemia cells resulted in complete clearance of engrafted leukemia cells [11]. Phase I clinical trials with RG7356 were conducted in patients with acute myeloid leukemia [12] and advanced CD44-positive solid tumor patients [13]. RG7356 showed an acceptable safety profile. However, the studies were not continued due to the lack of a clinical and/or pharmacodynamic dose-response relationship with RG7356 [13].
Therapies using anti-CD44v6 mAbs were considered since CD44v6 plays critical roles in the malignant progression of tumors [14]. Humanized anti-CD44v6 mAbs including BIWA-4 and BIWA-8 were developed. These mAbs labeled with 186Re were evaluated in head and neck squamous cell carcinoma xenograft-bearing mice and exhibited therapeutic efficacy [15]. Furthermore, the BIWA-4 was developed into a humanized version-drug conjugate, bivatuzumab-mertansine (anti-tubulin agent) which was evaluated in clinical trials [16]. However, the clinical trials were terminated because of the severe skin toxicity such as lethal epidermal necrolysis [17]. Since CD44v6 is expressed in normal skin epithelium, the toxicity of mertansine in the skin was most likely responsible for the high toxicity [17,18]. Recently, a mutated version of BIWA-4, called BIWA-8, was developed for increasing binding affinity by two amino acid substitutions of the light chain [15]. The BIWA-8 was further developed to chimeric antigen receptors (CARs). The BIWA-8 CAR-T showed antitumor activities against multiple myeloma or acute myeloid leukemia engrafted with immunodeficient mice [19]. Furthermore, the BIWA-8 CAR-T exhibited efficacy in lung and ovarian carcinomas xenograft models [20], which is expected for an application toward solid tumors.
Antibody-dependent cellular cytotoxicity (ADCC) is elicited by NK cells or macrophages upon the binding of Fcγ receptors (FcγRs) to the Fc region of mAbs [21]. However, the N-linked glycosylation in the Fc region impairs the ADCC [22]. In particular, a lack of core fucose on the Fc N-glycan enhances the Fc binding to the FcγRs on effector cells [23]. Fucosyltransferase 8 (FUT8) is the only α1,6-fucosyltransferase that transfers fucose from GDP-fucose to N-acetylglucosamine via an α1,6 linkage [24]. Therefore, FUT8 knockout-Chinese hamster ovary (CHO) is an ideal host cell for the production of completely defucosylated mAbs for therapeutic use [25]. Moreover, the complement family is also activated by the Fc domain, which exerts complement-dependent cytotoxicity (CDC) [26,27].
In our previous studies, an anti-pan-CD44 mAb, C44Mab-5 (IgG1, kappa) was developed using the Cell-Based Immunization and Screening (CBIS) method [28]. Another mAb (C44Mab-46) [29] was developed by immunization of the CD44v3–10 ectodomain. Both C44Mab-5 and C44Mab-46 have the epitopes within the constant exon 2- and 5-encoded sequences [30,31,32] and could be applied to immunohistochemistry in oral squamous cell carcinoma (OSCC) [28] and esophageal squamous cell carcinoma (ESCC) [29], respectively. Furthermore, we have developed various anti-CD44v mAbs, including C44Mab-6 (an anti-CD44v3 mAb), C44Mab-108 (an anti-CD44v4 mAb), C44Mab-3 (an anti-CD44v5 mAb), C44Mab-9 (an anti-CD44v6 mAb), C44Mab-34 (an anti-CD44v7/8 mAb), C44Mab-1 (an anti-CD44v9 mAb), and C44Mab-18 (an anti-CD44v10 mAb) (http://www.med-tohoku-antibody.com/topics/001_paper_antibody_PDIS.htm#CD44v3). The generation of anti-CD44 mAbs, which recognize all variant exons, is important for the comprehensive analysis of human tumors.
In this study, we produced a class-switched and defucosylated type of recombinant C44Mab-5 (5-mG2a-f) and C44Mab-46 (C44Mab-46-mG2a-f) using FUT8-deficient ExpiCHO-S cells and investigated the antitumor activity in xenograft-transplanted mice.
2. Materials and Methods
2.1. Cell Lines and Cell Culture
ESCC cell line, KYSE770 was obtained from the Japanese Collection of Research Bioresources (Osaka, Japan). CHO-K1 was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). CHO/CD44s was previously established by transfecting pCAG-Ble/PA16-CD44s into CHO-K1 cells using a Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA, USA) [29].
2.2. Antibodies
Anti-pan-CD44 mAbs, C44Mab-5 and C44Mab-46 were previously established [28,29]. To generate recombinant mAbs, VH cDNAs of C44Mab-5 or C44Mab-46 and CH of mouse IgG2a were cloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). VL cDNA of C44Mab-5 or C44Mab-46 and CL cDNA of mouse kappa light chain was also subcloned into the pCAG-Neo vector (FUJIFILM Wako Pure Chemical Corporation). The vectors were transduced into BINDS-09 (FUT8-knockout ExpiCHO-S) cells [33] using the ExpiCHO Expression System (Thermo Fisher Scientific, Inc). 281-mG2a-f (defucosylated anti-hamster podoplanin mAb, control mouse IgG2a) was previously described [34].
2.3. Flow Cytometry
Cells were isolated using 0.25% trypsin and 1 mM ethylenediamine tetraacetic acid (EDTA; Nacalai Tesque, Inc.) treatment. The cells were treated with 5-mG2a-f and C44Mab-46-mG2a-f, or blocking buffer [0.1% bovine serum albumin in phosphate-buffered saline (PBS)] (control) for 30 min at 4˚C. Subsequently, the cells were incubated in Alexa Fluor 488-conjugated anti-mouse IgG (1:2,000; Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 min at 4˚C. Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan) and analyzed using SA3800 software ver. 2.05 (Sony Corp.).
2.4. ADCC Reporter Bioassay
The ADCC reporter bioassay was performed using an ADCC Reporter Bioassay kit (Promega Corporation, Madison, WI, USA), according to the manufacturer’s instructions.
2.5. CDC
We labeled target cells (CHO/CD44s and KYSE770) using 10 µg/mL Calcein AM (Thermo Fisher Scientific, Inc.). The target cells were plated in 96-well plates (1 × 104 cells/well) and mixed with rabbit complement (final dilution 1:10, Low-Tox-M Rabbit Complement; Cedarlane Laboratories, Hornby, ON, Canada) and 100 μg/ml of control 281-mG2a-f, C44Mab-5, or C44Mab-46. The Calcein release to the supernatant from each well was measured after a 4-hour incubation. Fluorescence intensity was determined using a microplate reader (Power Scan HT; BioTek Instruments, Winooski, VT, USA).
2.6. Antitumor Activity of 5-mG2a-f and C44Mab-46-mG2a-f in Xenografts of CHO/CD44s and KYSE770
CHO/CD44s and KYSE770 cells (5 × 106 cells) suspended with BD Matrigel Matrix Growth Factor Reduced (BD Biosciences, Franklin Lakes, NJ, USA) were inoculated into the left flank of female BALB/c nude mice subcutaneously. On day 7 after the inoculation, 100 μg of 5-mG2a-f (n=8), C44Mab-46-mG2a-f (n=8), or control 281-mG2a-f (n=8) in 100 μl PBS were injected intraperitoneally. Additional antibody injections were performed on days 14 and 21. The tumor volume was measured on days 7, 9, 14, 17, 21, and 23.
In the KYSE770 xenograft experiment (500 μg dosage), 500 μg of 5-mG2a-f (n=8), C44Mab-46-mG2a-f (n=8), or control 281-mG2a-f (n=8) in 100 μl PBS were injected intraperitoneally on days 8 and 13 after the inoculation. The tumor volume was measured on days 8, 12, and 19. The xenograft tumors were carefully removed from the sacrificed mice and then weighed immediately.
3. Results
3.1. Flow Cytometric Analysis against CHO/CD44s and KYSE770 Cells Using 5-mG2a-f and C44Mab-46-mG2a-f
In our previous study, anti-pan-CD44 mAbs (C44Mab-5 and C44Mab-46) were established and were shown to be available for flow cytometry and immunohistochemistry [28,29]. In the present study, a defucosylated form of C44Mab-5 and C44Mab-46 (5-mG2a-f and C44Mab-46-mG2a-f) were produced by combining VH and VL of both mAbs with CH and CL of mouse IgG2a, respectively. We first confirm the reactivity of CD44s-overexpressed CHO-K1 cells (CHO/CD44s) and an endogenous CD44-expressing esophageal tumor cell line (KYSE770). 5-mG2a-f and C44Mab-46-mG2a-f detected CHO/CD44s and not parental CHO-K1 cells in a concentration-dependent manner (Figure 1A,B). 5-mG2a-f and C44Mab-46-mG2a-f also detected KYSE770 cells in a concentration-dependent manner (Figure 1C). These results indicated that 5-mG2a-f and C44Mab-46-mG2a-f recognize exogenous and endogenous CD44.
3.2. 5-mG2a-f and C44Mab-46-mG2a-f-Mediated ADCC Pathway Activation in the Presence of CHO/CD44s and KYSE770 Cells
The ADCC reporter bioassay is a bioluminescent reporter gene assay for the quantification of the biological activity of the antibody via FcγRIIIa-mediated pathway activation [35]. To compare the ADCC pathway activation by 5-mG2a-f and C44Mab-46-mG2a-f, the CHO/CD44s cells were treated with serially diluted mAbs and then incubated with effector Jurkat cells, which express the human FcγRIIIa receptor and an NFAT response element driving Firefly luciferase. As demonstrated in Figure 2A, 5-mG2a-f and C44Mab-46-mG2a-f activated the effector in a concentration-dependent manner (EC50; 6.4×10−8 g/mL and 5.4×10−8 g/mL, respectively). In the presence of KYSE770 cells, 5-mG2a-f activated the effector (EC50; 2.5×10−7 g/mL, Figure 2B). However, C44Mab-46-mG2a-f did not (Figure 2B).
3.3. The CDC by 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s and KYSE770 Cells
We next examined the CDC mediated by 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s and KYSE770 cells. As shown in Figure 3, both 5-mG2a-f and C44Mab-46-mG2a-f significantly exerted CDC against CHO/CD44s cells (67% and 88% cytotoxicity, respectively) and KYSE770 (27% and 31% cytotoxicity, respectively) compared with those induced by control defucosylated mouse IgG2a (281-mG2a-f). These results demonstrated that 5-mG2a-f and C44Mab-46-mG2a-f exhibited potent CDC activities against CHO/CD44s and KYSE770 cells.
3.4. Antitumor Effects of 5-mG2a-f and C44Mab-46-mG2a-f in the Mouse Xenografts of CHO/CD44s
In the CHO/CD44s xenograft tumors, 100 µg of 5-mG2a-f, C44Mab-46-mG2a-f, or control mouse IgG2a were injected into mice intraperitoneally on days 7, 14, and 21, following CHO/CD44s inoculation. On days 7, 9, 14, 17, 21, and 23, the tumor volume was measured. The 5-mG2a-f and C44Mab-46-mG2a-f administration resulted in a significant reduction in tumor volume on days 17 (p < 0.05), 21 (p < 0.01), and 23 (p < 0.01) compared with that of the control mouse IgG2a (Figure 4A). The 5-mG2a-f and C44Mab-46-mG2a-f administration resulted in 46% and 48% reductions in tumor volume compared with that of the control mouse IgG2a on day 23, respectively.
The weight of CHO/CD44s tumors treated with 5-mG2a-f and C44Mab-46-mG2a-f was significantly lower than that of tumors treated with the control mouse IgG2a (53% and 55% reduction, respectively; p < 0.05; Figure 4B). The loss of body weight was not observed in the CHO/CD44s tumor-implanted mice during the treatments (Figure 4C).
3.5. Antitumor Effects of 5-mG2a-f and C44Mab-46-mG2a-f on KYSE770 Xenografts
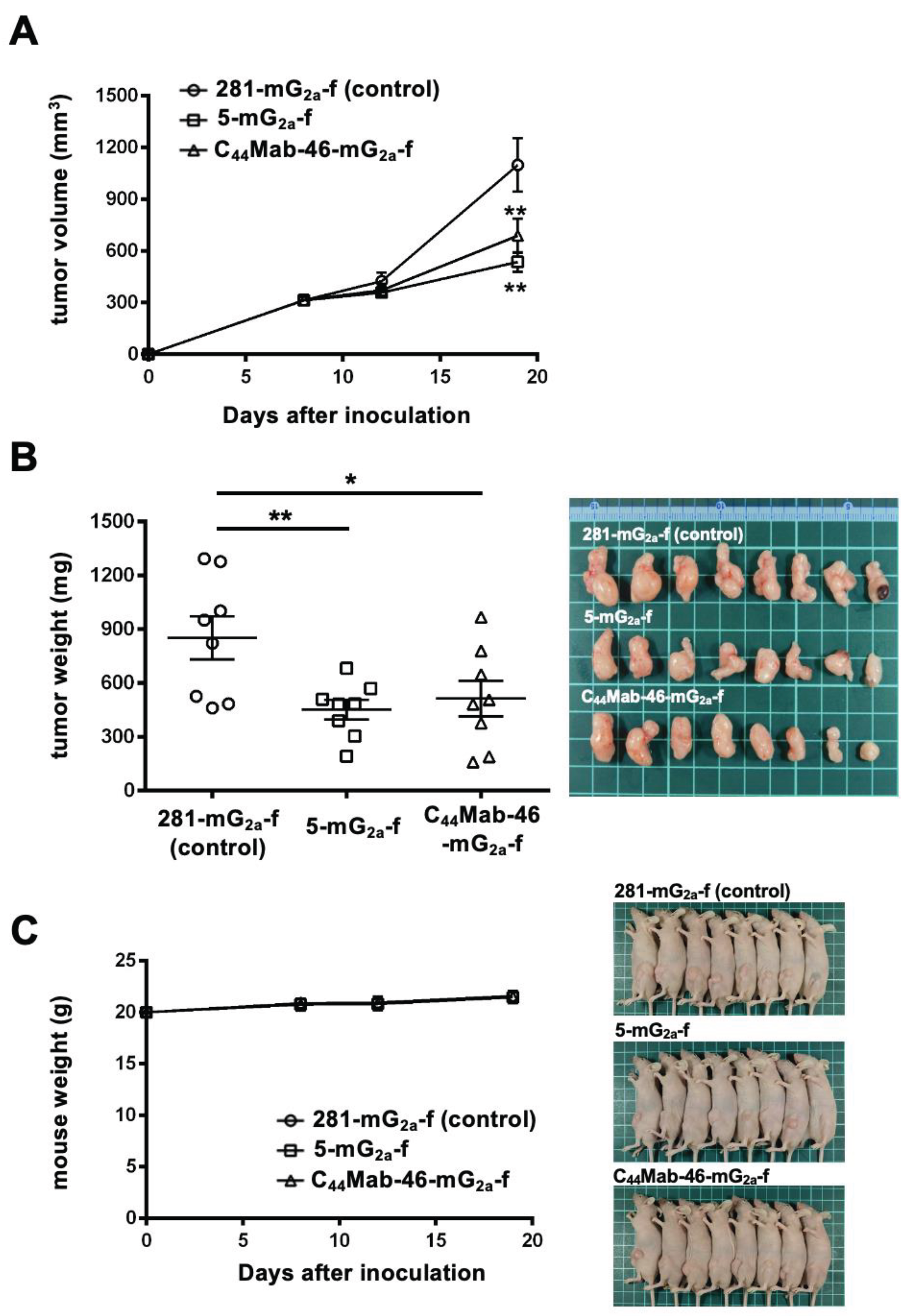
In the KYSE770 xenograft models, 100 µg of 5-mG2a-f and C44Mab-46-mG2a-f and control mouse IgG2a were injected into mice on days 7, 14, and 21, following KYSE770 inoculation. Although we observed the tendency of the reduction of tumor volume by the treatment with 5-mG2a-f and C44Mab-46-mG2a-f, the significant differences were not obtained (Supplementary Figure 1). We next increased the dosage (500 µg) and investigated the antitumor effects. As shown in Figure 5, 500 µg of 5-mG2a-f, C44Mab-46-mG2a-f, and control mouse IgG2a were injected into mice on days 8 and 13, following KYSE770 inoculation. On days 8, 12, and 19, the tumor volume was measured. The 5-mG2a-f and C44Mab-46-mG2a-f administration resulted in a significant reduction in tumor volume on day 19 (p < 0.01) compared with that of the control mouse IgG2a (Figure 5A). The 5-mG2a-f and C44Mab-46-mG2a-f administration resulted in 51% and 37% reduction of tumor volume compared with that of the control mouse IgG2a on day 19, respectively.
The weight of CHO/CD44s tumors treated with 5-mG2a-f and C44Mab-46-mG2a-f was significantly lower than that of tumors treated with the control mouse IgG2a [47% (p < 0.01) and 40% (p < 0.05) reduction, respectively; Figure 5B]. The loss of body weight was not observed in the CHO/CD44s tumor-implanted mice during the treatments (Figure 5C).
4. Discussion
In the present study, we produced defucosylated mouse IgG2a type anti-pan-CD44 mAbs (5-mG2a-f and C44Mab-46-mG2a-f) and evaluated antitumor activity against CHO/CD44s and KYSE770 xenograft tumors. Both 5-mG2a-f and C44Mab-46-mG2a-f showed comparable reactivity in flow cytometry (Figure 1), effector activation (Figure 2), and antitumor activity (Figure 4) against CHO/CD44s. Against KYSE770 cells, 5-mG2a-f exhibited superior reactivity in flow cytometry (Figure 1) and effector activation (Figure 2) compared to C44Mab-46-mG2a-f. However, both mAbs showed similar antitumor activity to KYSE770 xenograft in high dosage (Figure 5).
After the phase I trial of RG7356 in solid tumors, the developer Roche group reported that the CD44s expression is associated with HA production and predicts response to treatment with RG7356 in both xenograft tumor models and clinical response [36]. The group also showed that overexpression of CD44s stimulates the HA production [36]. These results suggest the close interplay between CD44s and HA and a potential biomarker to enrich patient responses to anti-CD44 mAb therapy in the clinic. In our study, we used CD44s-overexpressed CHO-K1 cells and KYSE770 cells which mainly express CD44v [29]. Due to the expression levels of total CD44 (Figure 1) and the abovementioned reason, a high dosage (500 µg) of mAbs was needed to suppress the KYSE770 xenograft.
As shown in Figure 2B, C44Mab-46-mG2a-f did not activate the effector cells in the presence of KYSE770 but exerted the antitumor effect in vivo (Figure 5). The CDC is an important antitumor mechanism by mAbs [26,27]. Some tendency was observed that C44Mab-46-mG2a-f showed a larger CDC effect compared to 5-mG2a-f (Figure 3). The difference could be sufficient to exert similar antitumor effects in vivo by 5-mG2a-f and C44Mab-46-mG2a-f. Since the epitopes of 5-mG2a-f and C44Mab-46-mG2a-f are different [30,31,32], the contribution of ADCC and CDC effects on antitumor activity could be different. We should clarify the reason why 5-mG2a-f and C44Mab-46-mG2a-f possess different ADCC and CDC activities in vitro.
Complement has been considered as an adjunctive component that potentiates the antibody-mediated cytolytic effects. However, complement is currently considered an important effector of tumor cytotoxic responses of antibody-based immunotherapy, which is guiding new therapeutic options [27]. Through the attempts to show the involvement with the cytolytic capacity of complement in mAb-based cancer immunotherapy, rituximab, a chimeric anti-CD20 mAb developed for the treatment of B cell lymphomas, achieved the full realization of the antitumor effects of complement [37,38]. The tumor destructive role of complement has been shown in not only anti-CD20 but also anti-CD38 and CD52 immunotherapies that are highly expressed on B cell or T cell-derived tumors [38,39,40]. A growing body of evidence suggests that complement plays critical roles in not only mAb-mediated tumor cytolysis but also several immunomodulatory functions in tumor immunosurveillance and anti-tumor immunity [41,42]. By the complicated crosstalk of complement effectors with cellular pathways that drive B cell and T cell responses, T helper/effector T cell survival, differentiation, and B cell activation are influenced. Therefore, the involvement of complement in our experimental system and/or immunocompetent mouse models should be investigated in future studies.
Most therapeutic mAbs exhibit adverse effects by the recognition of antigens in normal cells [43]. Clinical trials of an anti-CD44v6 mAb (BIWA-4) bivatuzumab-mertansine drug conjugate to solid tumors failed because of the skin toxicities [17,18]. Therefore, tumor-selective or specific mAbs would reduce the adverse effects. We have developed cancer-specific mAbs (CasMabs) against HER2 (H2Mab-214 [44] and H2Mab-250 [45]), podocalyxin (PcMab-6 [46] and PcMab-60 [47]), and podoplanin (LpMab-2 [48] and LpMab-23 [49]) and evaluated the reactivity to cancer and normal cells in flow cytometry. We also reported the antitumor effect in mouse xenograft models using the defucosylated mouse IgG2a or human IgG1 types recombinant mAbs [44,46,48,49,50,51,52]. Some anti-CD44 mAbs which exhibit cancer specificity have been reported [53]. Among them, the 4C8 mAb recognizes aberrantly O-glycosylated CD44v6 with Tn (GalNAcα1-O-Ser/Thr) antigen. The 4C8 chimeric antigen receptor (CAR)-T cells demonstrated target-specific cytotoxicity in vitro, significant tumor regression, and prolonged survival in vivo [54]. In our CasMab development against CD44, we screened our established anti-CD44 mAbs by comparing the reactivity against cancer and normal cells. Anti-CD44 CasMabs could be applicable for designing the modalities including antibody-drug conjugates and CAR-T cells.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Antitumor activity of 5-mG2a-f and C44Mab-46-mG2a-f against KYSE770 xenograft (low dosage).
Author Contributions
K.I., H.S., T.O., T.N., M.Y., G.L., and T.T. performed the experiments. A.O. and Y.Katori supervised the first author. M.K., M.K.K., and Y.Kato designed the experiments. H.S. and Y.Kato. analyzed the data. H.S. and Y.Kato wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP23ama121008 (to Y.Kato), JP23am0401013 (to Y.Kato), JP23bm1123027 (to Y.Kato), and JP23ck0106730 (to Y.Kato), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.S.), 21K20789 (to T.T.), 21K07168 (to M.K.K.), and 22K07224 (to Y.Kato).
Institutional Review Board Statement
Animal experiments were approved by the Institutional Committee for Experiments of the Institute of Microbial Chemistry (approval nos. 2023-037 and 2023-054).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article and supplementary material.
Acknowledgments
The authors would like to thank Shun-ichi Ohba and Akiko Harakawa (Institute of Microbial Chemistry [BIKAKEN], Numazu, the Microbial Chemistry Research Foundation) for technical assistance with animal experiments.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: from adhesion molecules to signalling regulators. Nat Rev Mol Cell Biol 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Zöller, M. CD44: can a cancer-initiating cell profit from an abundantly expressed molecule? Nat Rev Cancer 2011, 11, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, L.; Tesarik, R.; Turanek, J. Regulation of alternative splicing of CD44 in cancer. Cell Signal 2014, 26, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yang, C.; Gao, F. The state of CD44 activation in cancer progression and therapeutic targeting. Febs j 2021. [Google Scholar] [CrossRef] [PubMed]
- Zöller, M. CD44, Hyaluronan, the Hematopoietic Stem Cell, and Leukemia-Initiating Cells. Front Immunol 2015, 6, 235. [Google Scholar] [CrossRef] [PubMed]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, N. The Hyaluronan/CD44 Axis: A Double-Edged Sword in Cancer. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, T.; Lu, D.; Zhen, J.; Zhang, L. CD44 overexpression related to lymph node metastasis and poor prognosis of pancreatic cancer. Int J Biol Markers 2018, 33, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Li, H.; Huang, R.; Mo, D.; Zeng, T.; Fang, M.; Li, M. Clinicopathological and Prognostic Significance of Cancer Stem Cell Markers in Ovarian Cancer Patients: Evidence from 52 Studies. Cell Physiol Biochem 2018, 46, 1716–1726. [Google Scholar] [CrossRef]
- Li, L.; Hao, X.; Qin, J.; Tang, W.; He, F.; Smith, A.; Zhang, M.; Simeone, D.M.; Qiao, X.T.; Chen, Z.N.; et al. Antibody against CD44s inhibits pancreatic tumor initiation and postradiation recurrence in mice. Gastroenterology 2014, 146, 1108–1118. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, C.C.; Fecteau, J.F.; Cui, B.; Chen, L.; Zhang, L.; Wu, R.; Rassenti, L.; Lao, F.; Weigand, S.; et al. Targeting chronic lymphocytic leukemia cells with a humanized monoclonal antibody specific for CD44. Proc Natl Acad Sci U S A 2013, 110, 6127–6132. [Google Scholar] [CrossRef]
- Vey, N.; Delaunay, J.; Martinelli, G.; Fiedler, W.; Raffoux, E.; Prebet, T.; Gomez-Roca, C.; Papayannidis, C.; Kebenko, M.; Paschka, P.; et al. Phase I clinical study of RG7356, an anti-CD44 humanized antibody, in patients with acute myeloid leukemia. Oncotarget 2016, 7, 32532–32542. [Google Scholar] [CrossRef]
- Menke-van der Houven van Oordt, C.W.; Gomez-Roca, C.; van Herpen, C.; Coveler, A.L.; Mahalingam, D.; Verheul, H.M.; van der Graaf, W.T.; Christen, R.; Rüttinger, D.; Weigand, S.; et al. First-in-human phase I clinical trial of RG7356, an anti-CD44 humanized antibody, in patients with advanced, CD44-expressing solid tumors. Oncotarget 2016, 7, 80046–80058. [Google Scholar] [CrossRef]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell 2014, 14, 342–356. [Google Scholar] [CrossRef]
- Verel, I.; Heider, K.H.; Siegmund, M.; Ostermann, E.; Patzelt, E.; Sproll, M.; Snow, G.B.; Adolf, G.R.; van Dongen, G.A. Tumor targeting properties of monoclonal antibodies with different affinity for target antigen CD44V6 in nude mice bearing head-and-neck cancer xenografts. Int J Cancer 2002, 99, 396–402. [Google Scholar] [CrossRef]
- Orian-Rousseau, V.; Ponta, H. Perspectives of CD44 targeting therapies. Arch Toxicol 2015, 89, 3–14. [Google Scholar] [CrossRef]
- Tijink, B.M.; Buter, J.; de Bree, R.; Giaccone, G.; Lang, M.S.; Staab, A.; Leemans, C.R.; van Dongen, G.A. A phase I dose escalation study with anti-CD44v6 bivatuzumab mertansine in patients with incurable squamous cell carcinoma of the head and neck or esophagus. Clin Cancer Res 2006, 12, 6064–6072. [Google Scholar] [CrossRef]
- Riechelmann, H.; Sauter, A.; Golze, W.; Hanft, G.; Schroen, C.; Hoermann, K.; Erhardt, T.; Gronau, S. Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral Oncol 2008, 44, 823–829. [Google Scholar] [CrossRef]
- Casucci, M.; Nicolis di Robilant, B.; Falcone, L.; Camisa, B.; Norelli, M.; Genovese, P.; Gentner, B.; Gullotta, F.; Ponzoni, M.; Bernardi, M.; et al. CD44v6-targeted T cells mediate potent antitumor effects against acute myeloid leukemia and multiple myeloma. Blood 2013, 122, 3461–3472. [Google Scholar] [CrossRef]
- Porcellini, S.; Asperti, C.; Corna, S.; Cicoria, E.; Valtolina, V.; Stornaiuolo, A.; Valentinis, B.; Bordignon, C.; Traversari, C. CAR T Cells Redirected to CD44v6 Control Tumor Growth in Lung and Ovary Adenocarcinoma Bearing Mice. Front Immunol 2020, 11, 99. [Google Scholar] [CrossRef]
- Tsao, L.C.; Force, J.; Hartman, Z.C. Mechanisms of Therapeutic Antitumor Monoclonal Antibodies. Cancer Res 2021, 81, 4641–4651. [Google Scholar] [CrossRef]
- Pereira, N.A.; Chan, K.F.; Lin, P.C.; Song, Z. The "less-is-more" in therapeutic antibodies: Afucosylated anti-cancer antibodies with enhanced antibody-dependent cellular cytotoxicity. MAbs 2018, 10, 693–711. [Google Scholar] [CrossRef]
- Shinkawa, T.; Nakamura, K.; Yamane, N.; Shoji-Hosaka, E.; Kanda, Y.; Sakurada, M.; Uchida, K.; Anazawa, H.; Satoh, M.; Yamasaki, M.; et al. The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem 2003, 278, 3466–3473. [Google Scholar] [CrossRef]
- Miyoshi, E.; Noda, K.; Yamaguchi, Y.; Inoue, S.; Ikeda, Y.; Wang, W.; Ko, J.H.; Uozumi, N.; Li, W.; Taniguchi, N. The alpha1-6-fucosyltransferase gene and its biological significance. Biochim Biophys Acta 1999, 1473, 9–20. [Google Scholar] [CrossRef]
- Yamane-Ohnuki, N.; Kinoshita, S.; Inoue-Urakubo, M.; Kusunoki, M.; Iida, S.; Nakano, R.; Wakitani, M.; Niwa, R.; Sakurada, M.; Uchida, K.; et al. Establishment of FUT8 knockout Chinese hamster ovary cells: an ideal host cell line for producing completely defucosylated antibodies with enhanced antibody-dependent cellular cytotoxicity. Biotechnol Bioeng 2004, 87, 614–622. [Google Scholar] [CrossRef]
- Golay, J.; Taylor, R.P. The Role of Complement in the Mechanism of Action of Therapeutic Anti-Cancer mAbs. Antibodies (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Reis, E.S.; Mastellos, D.C.; Ricklin, D.; Mantovani, A.; Lambris, J.D. Complement in cancer: untangling an intricate relationship. Nat Rev Immunol 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C(44)Mab-5. Biochem Biophys Rep 2018, 14, 64–68. [Google Scholar] [CrossRef]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using Alanine-Scanning Mutagenesis and Surface Plasmon Resonance. Monoclon Antib Immunodiagn Immunother 2021, 40, 219–226. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Takei, J.; Tateyama, N.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 156–161. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 162–167. [Google Scholar] [CrossRef]
- Li, G.; Suzuki, H.; Ohishi, T.; Asano, T.; Tanaka, T.; Yanaka, M.; Nakamura, T.; Yoshikawa, T.; Kawada, M.; Kaneko, M.K.; et al. Antitumor activities of a defucosylated anti-EpCAM monoclonal antibody in colorectal carcinoma xenograft models. Int J Mol Med 2023, 51. [Google Scholar] [CrossRef]
- Nanamiya, R.; Suzuki, H.; Takei, J.; Li, G.; Goto, N.; Harada, H.; Saito, M.; Tanaka, T.; Asano, T.; Kaneko, M.K.; et al. Development of Monoclonal Antibody 281-mG(2a)-f Against Golden Hamster Podoplanin. Monoclon Antib Immunodiagn Immunother 2022, 41, 311–319. [Google Scholar] [CrossRef]
- Garvin, D.; Stecha, P.; Gilden, J.; Wang, J.; Grailer, J.; Hartnett, J.; Fan, F.; Cong, M.; Cheng, Z.J. Determining ADCC Activity of Antibody-Based Therapeutic Molecules using Two Bioluminescent Reporter-Based Bioassays. Curr Protoc 2021, 1, e296. [Google Scholar] [CrossRef]
- Birzele, F.; Voss, E.; Nopora, A.; Honold, K.; Heil, F.; Lohmann, S.; Verheul, H.; Le Tourneau, C.; Delord, J.P.; van Herpen, C.; et al. CD44 Isoform Status Predicts Response to Treatment with Anti-CD44 Antibody in Cancer Patients. Clin Cancer Res 2015, 21, 2753–2762. [Google Scholar] [CrossRef]
- Taylor, R.P.; Lindorfer, M.A. Cytotoxic mechanisms of immunotherapy: Harnessing complement in the action of anti-tumor monoclonal antibodies. Semin Immunol 2016, 28, 309–316. [Google Scholar] [CrossRef]
- Reff, M.E.; Carner, K.; Chambers, K.S.; Chinn, P.C.; Leonard, J.E.; Raab, R.; Newman, R.A.; Hanna, N.; Anderson, D.R. Depletion of B cells in vivo by a chimeric mouse human monoclonal antibody to CD20. Blood 1994, 83, 435–445. [Google Scholar] [CrossRef]
- de Weers, M.; Tai, Y.T.; van der Veer, M.S.; Bakker, J.M.; Vink, T.; Jacobs, D.C.; Oomen, L.A.; Peipp, M.; Valerius, T.; Slootstra, J.W.; et al. Daratumumab, a novel therapeutic human CD38 monoclonal antibody, induces killing of multiple myeloma and other hematological tumors. J Immunol 2011, 186, 1840–1848. [Google Scholar] [CrossRef]
- Zent, C.S.; Secreto, C.R.; LaPlant, B.R.; Bone, N.D.; Call, T.G.; Shanafelt, T.D.; Jelinek, D.F.; Tschumper, R.C.; Kay, N.E. Direct and complement dependent cytotoxicity in CLL cells from patients with high-risk early-intermediate stage chronic lymphocytic leukemia (CLL) treated with alemtuzumab and rituximab. Leuk Res 2008, 32, 1849–1856. [Google Scholar] [CrossRef]
- Schmudde, I.; Laumonnier, Y.; Köhl, J. Anaphylatoxins coordinate innate and adaptive immune responses in allergic asthma. Semin Immunol 2013, 25, 2–11. [Google Scholar] [CrossRef]
- Carroll, M.C.; Isenman, D.E. Regulation of humoral immunity by complement. Immunity 2012, 37, 199–207. [Google Scholar] [CrossRef]
- Gogia, P.; Ashraf, H.; Bhasin, S.; Xu, Y. Antibody-Drug Conjugates: A Review of Approved Drugs and Their Clinical Level of Evidence. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Arimori, T.; Mihara, E.; Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Takagi, J.; Kato, Y. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Kato, Y. Establishment of a Novel Cancer-specific Anti-HER2 Monoclonal Antibody H2Mab-250/H2CasMab-2 for breast cancers. Monoclon Antib Immunodiagn Immunother 2024, in press. [Google Scholar]
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Monoclonal Antibody against Podocalyxin Exerted Antitumor Activities in Pancreatic Cancer Xenografts. Int J Mol Sci 2023, 25. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Ohishi, T.; Kawada, M.; Kato, Y. A cancer-specific anti-podocalyxin monoclonal antibody (60-mG(2a)-f) exerts antitumor effects in mouse xenograft models of pancreatic carcinoma. Biochem Biophys Rep 2020, 24, 100826. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci Rep 2014, 4, 5924. [Google Scholar] [CrossRef]
- Yamada, S.; Ogasawara, S.; Kaneko, M.K.; Kato, Y. LpMab-23: A Cancer-Specific Monoclonal Antibody Against Human Podoplanin. Monoclon Antib Immunodiagn Immunother 2017, 36, 72–76. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Suzuki, H.; Ohishi, T.; Nakamura, T.; Tanaka, T.; Kato, Y. A Cancer-Specific Monoclonal Antibody against HER2 Exerts Antitumor Activities in Human Breast Cancer Xenograft Models. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. A Humanized and Defucosylated Antibody against Podoplanin (humLpMab-23-f) Exerts Antitumor Activities in Human Lung Cancer and Glioblastoma Xenograft Models. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Yamada, S.; Nakamura, T.; Abe, S.; Nishioka, Y.; Kunita, A.; Fukayama, M.; Fujii, Y.; Ogasawara, S.; Kato, Y. Antitumor activity of chLpMab-2, a human-mouse chimeric cancer-specific antihuman podoplanin antibody, via antibody-dependent cellular cytotoxicity. Cancer Med 2017, 6, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Lodewijk, I.; Dueñas, M.; Paramio, J.M.; Rubio, C. CD44v6, STn & O-GD2: promising tumor associated antigens paving the way for new targeted cancer therapies. Front Immunol 2023, 14, 1272681. [Google Scholar] [CrossRef] [PubMed]
- Aasted, M.K.M.; Groen, A.C.; Keane, J.T.; Dabelsteen, S.; Tan, E.; Schnabel, J.; Liu, F.; Lewis, H.S.; Theodoropulos, C.; Posey, A.D.; et al. Targeting Solid Cancers with a Cancer-Specific Monoclonal Antibody to Surface Expressed Aberrantly O-glycosylated Proteins. Mol Cancer Ther 2023, 22, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow cytometry using 5-mG2a-f and C44Mab-46-mG2a-f. CHO/CD44s (A) CHO-K1 (B), and KYSE770 (C) cells were treated with 10-0.01 µg/mL of 5-mG2a-f and C44Mab-46-mG2a-f (red) or buffer control (black), followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were analyzed using the SA3800 Cell Analyzer.
Figure 1.
Flow cytometry using 5-mG2a-f and C44Mab-46-mG2a-f. CHO/CD44s (A) CHO-K1 (B), and KYSE770 (C) cells were treated with 10-0.01 µg/mL of 5-mG2a-f and C44Mab-46-mG2a-f (red) or buffer control (black), followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were analyzed using the SA3800 Cell Analyzer.
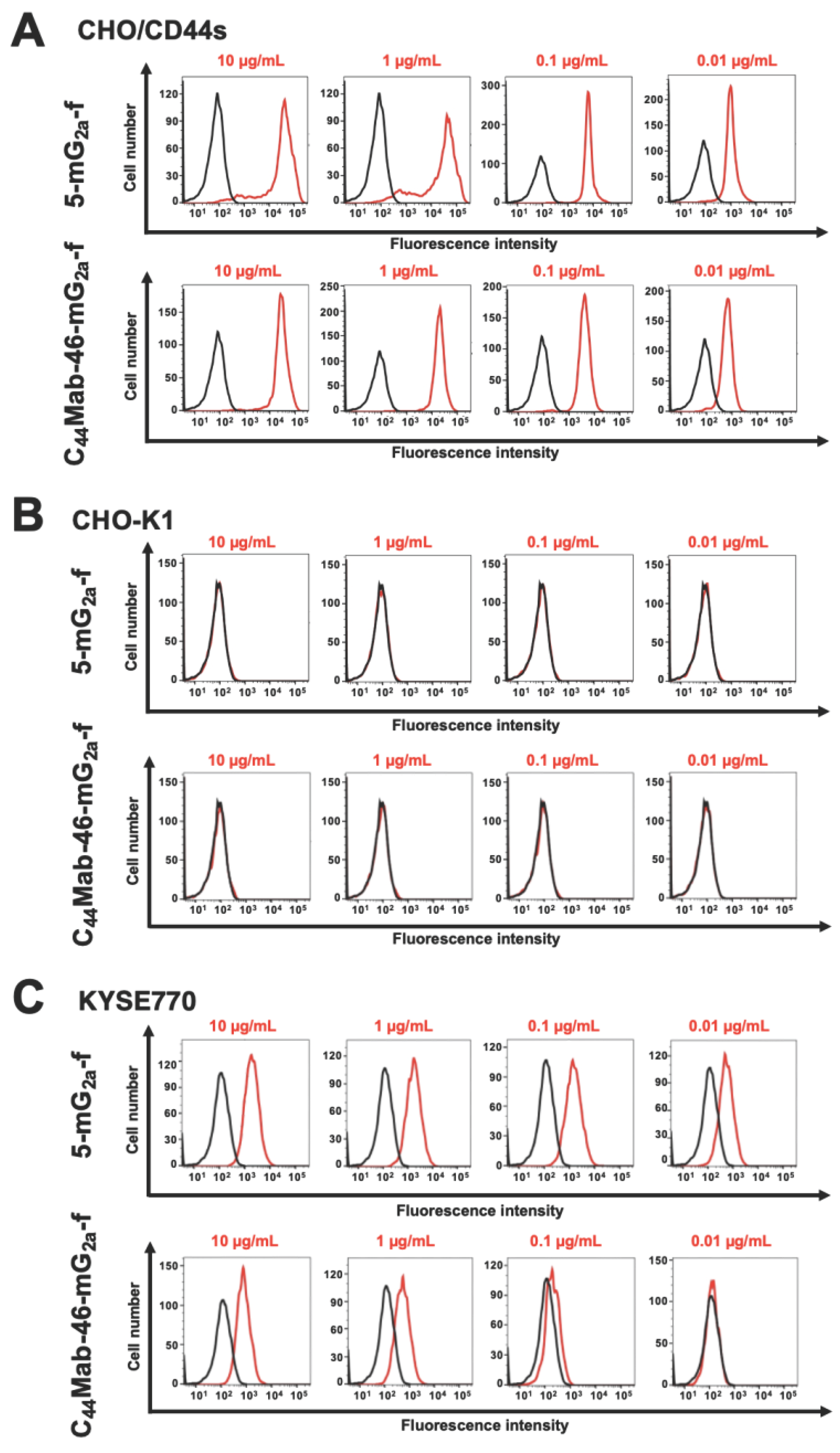
Figure 2.
Evaluation of ADCC activity. The ADCC reporter assay by 5-mG2a-f and C44Mab-46-mG2a-f in the presence of CHO/CD44s (A) and KYSE770 (B) cells. Values are presented as the mean ± SD. N.D., not determined. ADCC, Antibody-dependent cellular cytotoxicity.
Figure 2.
Evaluation of ADCC activity. The ADCC reporter assay by 5-mG2a-f and C44Mab-46-mG2a-f in the presence of CHO/CD44s (A) and KYSE770 (B) cells. Values are presented as the mean ± SD. N.D., not determined. ADCC, Antibody-dependent cellular cytotoxicity.
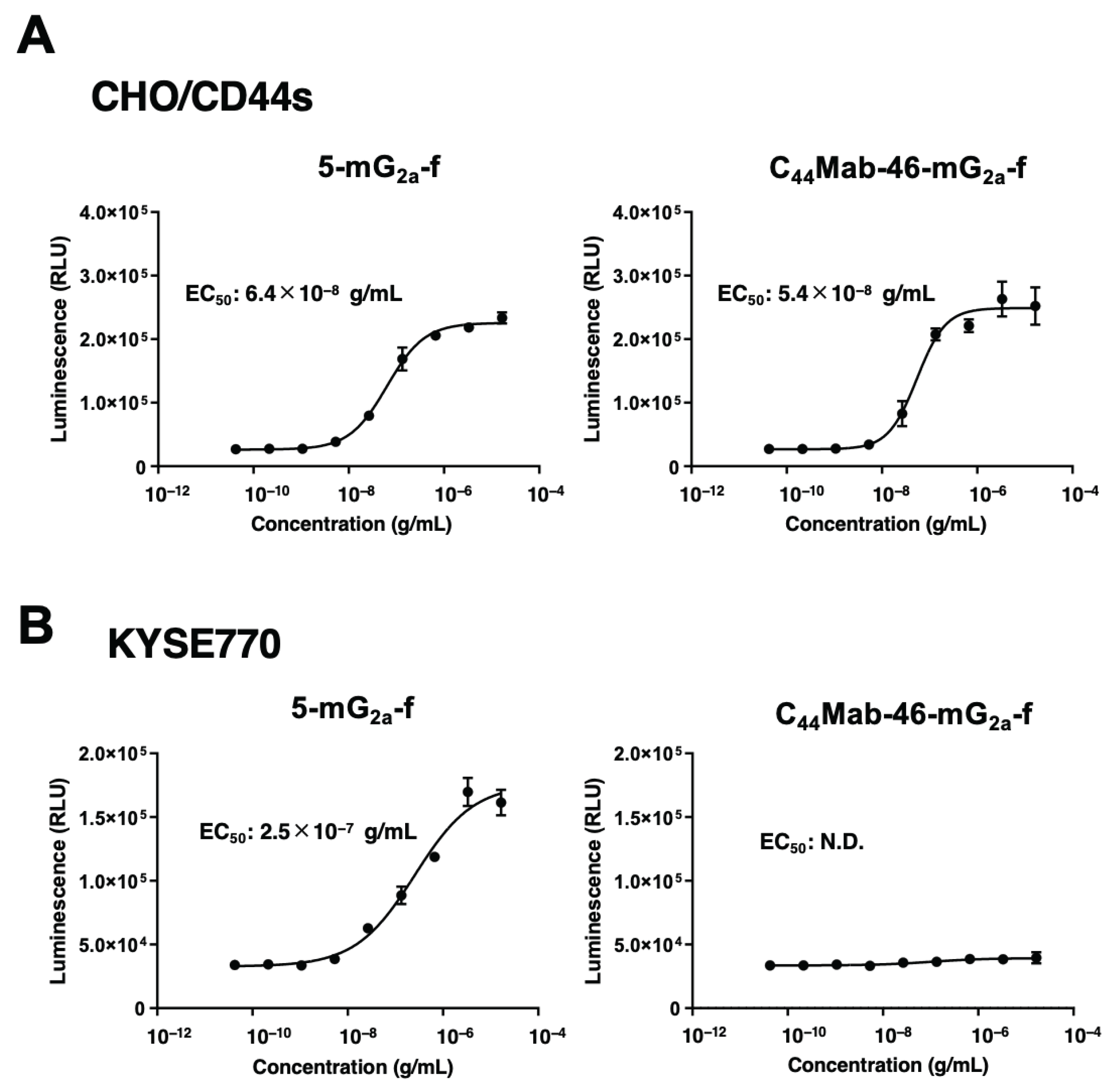
Figure 3.
Evaluation of CDC activity by 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s and KYSE770 cells. The CDC induced by 5-mG2a-f, C44Mab-46-mG2a-f, or control defucosylated mouse IgG2a (281-mG2a-f) against CHO/CD44s (A) and KYSE770 (B) cells. Values are shown as mean ± SEM. Asterisks indicate statistical significance (**P < 0.01 and *P < 0.05; one-way ANOVA with Tukey's multiple comparisons test). CDC, complement-dependent cytotoxicity. SEM, standard error of the mean.
Figure 3.
Evaluation of CDC activity by 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s and KYSE770 cells. The CDC induced by 5-mG2a-f, C44Mab-46-mG2a-f, or control defucosylated mouse IgG2a (281-mG2a-f) against CHO/CD44s (A) and KYSE770 (B) cells. Values are shown as mean ± SEM. Asterisks indicate statistical significance (**P < 0.01 and *P < 0.05; one-way ANOVA with Tukey's multiple comparisons test). CDC, complement-dependent cytotoxicity. SEM, standard error of the mean.

Figure 4.
Antitumor activity of 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s xenograft tumor. (A) Measurement of tumor volume in CHO/CD44s xenograft. CHO/CD44s cells (5 × 106 cells) were injected into mice subcutaneously. On day 7, 100 μg of 5-mG2a-f and C44Mab-46-mG2a-f or control mouse IgG2a (281-mG2a-f) were injected into mice intraperitoneally. On days 14 and 21, additional antibodies were injected. On days 7, 9, 14, 17, 21, and 23 following the inoculation, the tumor volume was measured. Values are presented as the mean ± SEM. *P < 0.05 and **P < 0.01 (ANOVA with Sidak’s multiple comparisons test). (B) The weight (left) and appearance (right) of excised CHO/CD44s xenografts on day 23. Values are presented as the mean ± SEM. *P<0.05 (Two-tailed unpaired t-test). (C) The body weight (left) and appearance (right) of control mouse IgG2a, 5-mG2a-f, and C44Mab-46-mG2a-f-treated mice. SEM, standard error of the mean.
Figure 4.
Antitumor activity of 5-mG2a-f and C44Mab-46-mG2a-f against CHO/CD44s xenograft tumor. (A) Measurement of tumor volume in CHO/CD44s xenograft. CHO/CD44s cells (5 × 106 cells) were injected into mice subcutaneously. On day 7, 100 μg of 5-mG2a-f and C44Mab-46-mG2a-f or control mouse IgG2a (281-mG2a-f) were injected into mice intraperitoneally. On days 14 and 21, additional antibodies were injected. On days 7, 9, 14, 17, 21, and 23 following the inoculation, the tumor volume was measured. Values are presented as the mean ± SEM. *P < 0.05 and **P < 0.01 (ANOVA with Sidak’s multiple comparisons test). (B) The weight (left) and appearance (right) of excised CHO/CD44s xenografts on day 23. Values are presented as the mean ± SEM. *P<0.05 (Two-tailed unpaired t-test). (C) The body weight (left) and appearance (right) of control mouse IgG2a, 5-mG2a-f, and C44Mab-46-mG2a-f-treated mice. SEM, standard error of the mean.
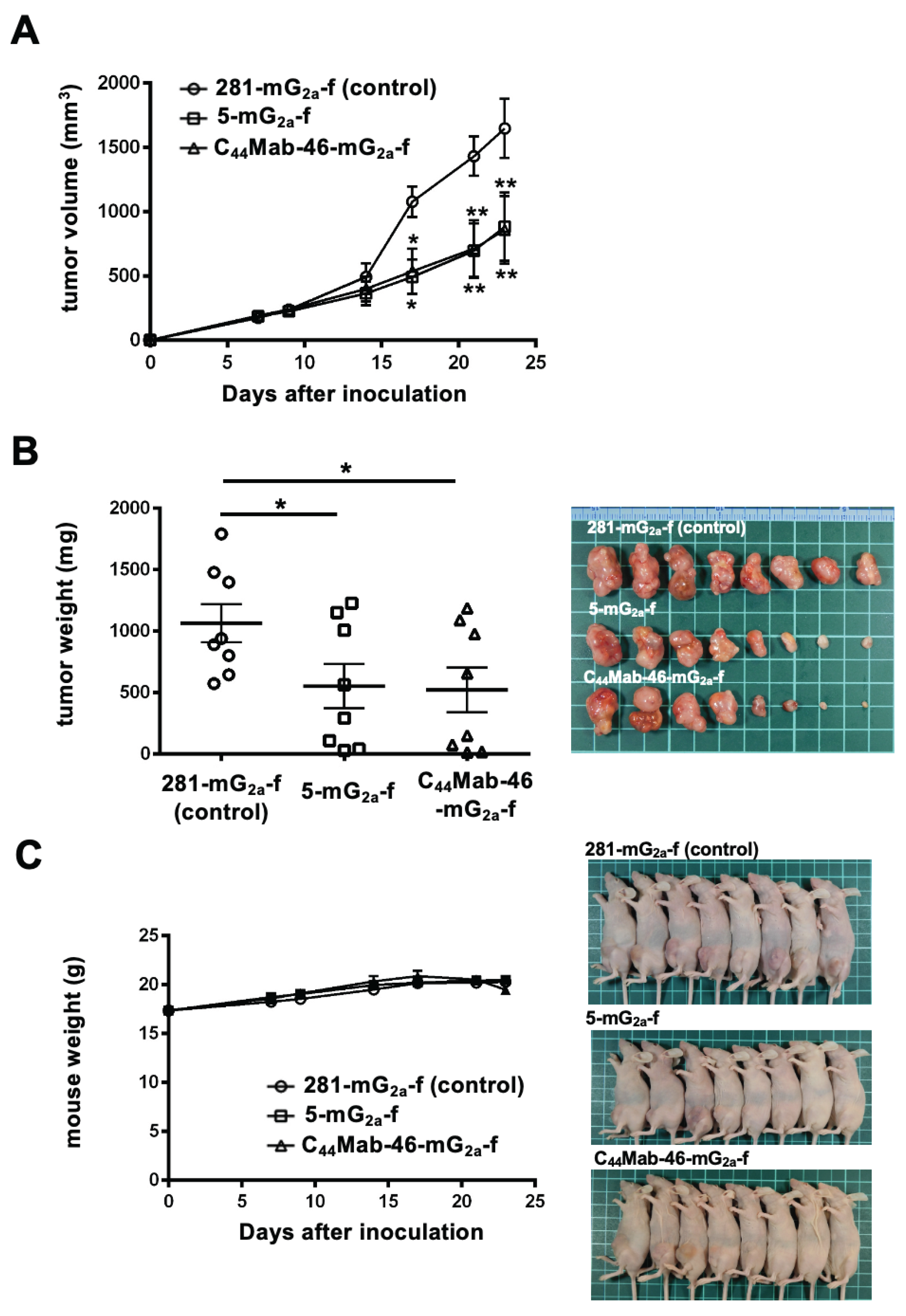
Figure 5.
Antitumor activity of 5-mG2a-f and C44Mab-46-mG2a-f against KYSE770 xenograft tumor. (A) Measurement of tumor volume in KYSE770 xenograft. KYSE770 cells (5 × 106 cells) were injected into mice subcutaneously. On day 8, 500 μg of 5-mG2a-f and C44Mab-46-mG2a-f or control mouse IgG2a (281-mG2a-f) were injected into mice intraperitoneally. On day 13, additional antibodies were injected. On days 8, 12, and 19 following the inoculation, the tumor volume was measured. Values are presented as the mean ± SEM. **P < 0.01 (ANOVA with Sidak’s multiple comparisons test). (B) The weight (left) and appearance (right) of the excised KYSE770 xenografts on day 19. Values are presented as the mean ± SEM. *P < 0.05 and **P < 0.01 (Two-tailed unpaired t-test). (C) The body weight (left) and appearance (right) of control mouse IgG2a, 5-mG2a-f, and C44Mab-46-mG2a-f-treated mice. SEM, standard error of the mean.
Figure 5.
Antitumor activity of 5-mG2a-f and C44Mab-46-mG2a-f against KYSE770 xenograft tumor. (A) Measurement of tumor volume in KYSE770 xenograft. KYSE770 cells (5 × 106 cells) were injected into mice subcutaneously. On day 8, 500 μg of 5-mG2a-f and C44Mab-46-mG2a-f or control mouse IgG2a (281-mG2a-f) were injected into mice intraperitoneally. On day 13, additional antibodies were injected. On days 8, 12, and 19 following the inoculation, the tumor volume was measured. Values are presented as the mean ± SEM. **P < 0.01 (ANOVA with Sidak’s multiple comparisons test). (B) The weight (left) and appearance (right) of the excised KYSE770 xenografts on day 19. Values are presented as the mean ± SEM. *P < 0.05 and **P < 0.01 (Two-tailed unpaired t-test). (C) The body weight (left) and appearance (right) of control mouse IgG2a, 5-mG2a-f, and C44Mab-46-mG2a-f-treated mice. SEM, standard error of the mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



