
Submitted:
22 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Understanding the genotype x environment (GE) interaction for traget trait is valuable for growers, agronomists, breeders and the seed industry. The objective of this study was to see and explore the genotype x environment interaction for grain yield of wheat to better guide genotype selection and breeding. The genotype main effect and GE interaction effect (GGE) biplots resolved the following three genotypes: 85 (ETW17-377), 82 (ETW17-374) and 13 (ETW17-281) for being the best yielding and stable. The Kulumsa site planted on 4th of July 2017 (environment 7=E7) and the DebreZeit site on 20th of July 2017 (E3) were the most effective environments in distinguishing the test genotypes and are thus suitable environments for future evaluation and selection with specific adaptation. Melkasa site planted on 4th July (E9) was a representative environment where genotypes can be evaluated for wide adpaptation. The genetic covariate biplot indicated that thousand kernel weight (TKW) and plant height (PH) had significant positive associations with grain yield implying the possible concurrent indirect selection of both traits for exploiting GE interaction for grain yield of wheat.
Keywords:
biplot method
; covariate traits
; genotype x environment interaction
; grain yield
; Triticum aestivum
; indirect selection
1. Introduction
Wheat (Triticum aestivum L., AABBDD, 2n = 6x = 42) is among the most widely cultivated cereal crop in the world including maize (Zea mays L.) and rice (Oryza sativa L.) [1]. Globally, China (with >115 million tons per annum) is the largest producer of wheat, followed by India (>82 million tons), the Russian Federation (>53 million tons), the USA (>50 million tons), France (>36 million tons), Canada (>26 million tons), Germany (>22 million tons), Pakistan (>22 million tons), Australia (>21 million tons) and Ukraine (>19 million tons) [2]. Wheat has C3 photosynthesis but adapts to growing under diverse agroecological conditions [3]. Ethiopia is the largest wheat producer in sub-Saharan Africa [4], with rising production areas of 1.7 million hectares in 2018 to 2.3 million in 2022. Wheat production has increased from 483 million tons in 2018 to 7 million tons in 2022 [2]. The national government has been spearheading local production and productivity of wheat through a dedicated extension system, expansion of irrigated wheat production to lowlands and clustered farming. Currently, wheat yields has increased on average from 1.8 tons ha-1 to 3 tons ha-1.
Global productivity of wheat has been threatened by several stresses such as biotic (e.g., rust diseases and insect pests), abiotic (e.g., recurrent drought, heat, cold and aluminium toxicity) and socio-economic constraints (e.g., limited access and affordability to production inputs) [5]. In sub-Saharan Africa, wheat is mainly cultivated under rainfed and drought-prone conditions. In these environments, the crop is affected by random and recurrent drought and heat stresses associated with climate change [5,6,7]. In Eastern Africa, 50-70% of crop yield is expected to decline due to climate change [8]. Climate-smart and stable-yielding improved cultivars and modern production technologies can bolster global wheat production and productivity under the changing climate conditions [5].
Well-characterized germplasm breeding populations are valuable genetic resources as a source of genetic variation for new and pipeline breeding programs [9,10]. Diverse wheat germplasm has previously been evaluated under varied environmental conditions by manipulating the sowing date [11]. However, genotype x environment interaction for target trait such as grain yield has not been studied. In this regard, multi-environment trials (MET) such as this are commonly studied to select and recommend genotypes suitable for target growing conditions with specific or broad adaptation. Understanding the GE interaction for grain yield is valuable for growers, agronomists, breeders and the seed industry. Several statistical methods are used to assess GE effects in crop improvement programs [12,13,14,15,16]. The most commonly applied methods are the Additive Main Effects and Multiplicative Interaction (AMMI) and the genotype main effect and GE interaction effect (GGE) models [17]. However, integrating biplot analysis system helps to realize and explore GE interaction for target trait such as grain yield [18]. The objective of this study was therefore to assess and explore the genotype x environment interaction for grain yield of wheat to better guide genotype selection and breeding.
2. Materials and methods
The present study used the dataset of one hundred and twenty bread wheat genotypes that were genotypically evaluated in 2017 at five locations, namely, AlemTena, DebreZeit, Dera, Kulumsa and Melkasa, using two planting dates following the onset of the main rainy season at each location, resulting in a total of 10 test environments (5 locations x 2 planting dates). The two planting dates represented two different moisture levels; early/optimum planting time corresponds to non-stressed, while late planting was drought-stressed. The non-stressed trial was planted on 14th (E1), 20th (E3), 5th (E5),4th (E7) and 16th (E9) of July 2017 at AlemTena, DebreZeit, Dera, Kulumsa and Melkasa, respectively, whereas the drought-stressed trial was planted on 13th (E2), 20th (E4), 3rd (E6),2nd (E8), and 15th (E10) of August 2017 at each location, in that order. Data on grain yield and yield related traits were collected at various stages of the crop cycle. Detailed of the experiment can be found in Semahegn et al. [11].
Data Analysis
A pooled analysis of variance was conducted using the GLM procedure in SAS version 9.3 (SAS Institute, Cary, NC, USA). The biplots were generated using the model for a GGE biplot based on singular value decomposition (SVD) of two-way data containing G and GE. The genotype x trait biplot was also produced using the same model, except that environments are replaced with traits, and it is based on the SVD of the trait-standardized dataset. The model is:
where Yij is the mean of the ith genotype in the jth environment; μ is the overall mean; ẟ is the effect of jth environment; t= the rank of the biplot (number of PC required, with t= min (g, e-1) for the full GGE model; λk ‘s (λk ≥ λk+1) are singular values that are partitioned into the singular vectors for genotypes (αik) and for environments (γjk) to approximate the dataset for biplot construction. ԑij is the residual error assumed NID (0, σ2/r), where r is the number of replications with in an environment.
A covariate–effect biplot was produced using a dataset of a two-way table of 120 rows for the genotypes and 7 columns for the explanatory traits plus 10 columns for the yield data in each environment. A 7 by 10 trait x environment two-way genetic covariable effects table was created based on the correlation coefficients between yield and the genetic values of each trait (data not shown). The two-way data was then decomposed into principal components through SVD, and the principal components were used to construct a biplot to visualize the covariate effect [18]. The model used was:
where rij is the correlation coefficient between yield and trait i in environment j, the other terms are the same as the above, except the genotype replaced with the trait. Detailed of the above models can be found in [17,18,19]. All the biplots were constructed using GEA-R software.
3. Result
3.1. Analysis of Variance
Pooled analysis of variance showed that environment, genotype and environment X genotype effects were significant (p < 0.0001) for grain yield of the tested genotypes (Table 1). Environment explained 85.2% of the total variation in the data while genotype and GE attributed to 1.5 and 6.1% of the variation, respectively. The tested genotypes had an overall mean of 3.8 tons ha-1.
3.2. Test Environment
The GGE biplot based on the 120-genotype x 10-environment yield dataset is depicted in Figure 1. Environment-focused singular value partitioning allows visualization of the relationships among environments. In this figure, the environment-vector view of the GGE biplot explained about 67% of the total variation of the environment–centred dataset. Environment vectors were drawn from the biplot origin (0, 0), indicating the test environments. The correlation between the environments were estimated using the cosine of the angle between their vectors. The Kulumsa site planted on 4th July (E7) and the same site planted on 2nd August (E8) had an acute angle and were positively correlated. E7 and the Dera site planted on 5th July (E5) had an obtuse angle and were negatively correlated, while E7 and the DebreZeit site planted on 20th July (E3) had minimal relation (with a right angle). Among the ten test environments, E7 was the most discriminating, followed by E3, E8 and E5. The AlemTena site planted on 13th August (E2) and the Dera site planted on 3rd August (E6) were the least discriminating in selecting the best genotype. Therefore, only E3, E5, E7, E8 and the Melkasa site planted on 16th July (E9) were the best environments for further genotype evaluation and analysis.
The ideal test environment should be the most discriminating/informative and representative of the target environment. The average-environment axis is a single-arrowed line that passes through the biplot origin (0,0) and the average environment (Figure 2). This biplot explained 74.4% of the total variation. E9 was close to the average environment and can be considered as most representative. The ideal test environment is the centre of the concentric circles in this figure. It is a point on the AEA (“most representative”) with a distance to the biplot origin equal to the longest vector of all environments (“most informative”). E7 and E8 were found to be relatively close to the ideal environment.
3.3. Genotype Stability
An ideal genotype should have high mean performance and stability across environments. Figure 3 depicts an “ideal” genotype (the centre of the concentric circles) to be a point on the average environment axis (AEA) (“absolutely stable”) and has a vector length equal to the longest vectors of the genotypes on the side of the AEA (“highest mean performance”). Therefore, genotypes closer to the ‘ideal genotype’ are more desirable than others. Based on this biplot view, genotypes designated as 85 (named as ETW17-377), 82 (ETW17-374) and 13 (ETW17-281) were the highest-yielding and stable, while 60 (ETW17-350), 77 (ETW17-368) and 119 (Dereselegn) were the lowest yielding and unstable. Genotype 90 (ETW17-383) was one of the most unstable, though it is high-yielding (4.83 tons ha-1).
The which-won-where GGE biplot in Figure 4 shows the winning genotypes on the vertices of the polygon and genotypes insensitive to changes in the environment in the polygon close to the origin of the biplot. The five environments fall into three sectors where genotypes 2 (ETW17-269) and 36 (ETW17-306) were the winner at Kulumsa site planted on 4th July (E7) and at the same site planted on 2nd August (E8) while genotypes 13 (ETW17-281) and 90 (ETW17-383) were the winner at DebreZeit site planted on 20th July (E3) and 118 (Ogolcho) was the winner at Dera site planted on 4th July (E5).
3.4. Genotype by Trait Analysis
Figure 5 represents the genotype x trait biplot that explained 64.4% of the total variation of the trait-centred genotype x trait two-way table. This biplot approximately displays the genetic correlations among the traits. The seven traits fell into three groups. TKW and PH constitute one group with positive associations among them. DH, MD, SS and KS constitute another group. These two groups were found to be negatively correlated; this relationship suggests that it is possible to identify and develop early maturing and high-yielding varieties by considering TKW and PH simultaneously in a breeding program. SL is intermediate between the two groups of traits.
3.5. Genetic Covariate by Environment
The covariate x environment biplot is shown in Figure 6. This biplot explained 48.8% of the total variation of the covariate-centred dataset. All traits except KS had stronger associations with grain yield (longer vectors). TKW and PH had consistently positive associations with yield, as evidenced by the acute angles between the vectors of these traits and the vectors of all environments except E5. Conversly, DH and MD showed consistently negative associations with yield, supported by the obtuse angles between their vectors and the vectors of all environments except E5.
4. Discussion
4.1. Analysis of Variance
The selection of superior genotypes for a target population of environments can be enanced through the proper utilization of GE. The proportion of GE was higher than G which may imply for possible existence of different mega-environments in testing environments [20,21]. Environment component normally accounts for 80% of the total variation and G and GE each accounts for about 10% of the total variation [22]. In this dataset, the analysis of variance revealed 85.2% of contribution to the total variation. The G and GE together accounted for 7.6% of the total variability.
4.2. Best Environments for Genotype Evaluation
The relationship among the test environments is inferred through the environment-vector view of the biplot with the cosine of the angles between two vectors estimating their correlation [18]. This analysis showed that the test environments are closely related except E7 and E8. Among the test environments, the Kulumsa site planted on 4th July (E7) was the most discriminating, followed by the DebreZeit site planted on 20th July (E3), the Kulumsa site planted on 2nd August (E8) and the Dera site planted on 5th July (E5). No environment was found perfectly aligned to the ideal environment; however, E7 and E8 were relatively close to the ideal environment. These environments are valuable to distinguish the test genotypes and for future evaluation and selection with specific adaptation [18]. The Melkasa site planted on 4th July (E9) was a representative environment where genotypes can be evaluated for wide environments. E7 and E3/E5 are vital to select wheat genotypes adapted to specific environments, given that the target environments can be divided into mega-environments. Several authors reported alike in different crops [23,24,25,26]. This will minimize breeding costs and maximise gain through better seed production. However, additional years are required to provide plausible conclusions. In sub-Saharan Africa, including Ethiopia, rainfall and temperature vary considerably across years. This study allowed genotype selection to be necessary in focused environments and thus reduced breeding cost.
4.3. Yield Stability
Changes in the environment may cause differential genotype response, which inturn result in inconsistent performance i.e., genotype x environment interaction [27]. The effect of GE can thus confound the selection of superior genotypes for a target population and environment. Conversely, this scenario helps identify broadly adapted and locally adapted genotypes [28]. High-yielding and stable performance of genotypes are desirable goals of plant breeding to attain sustainability in the food system [29]. GGE biplot is a vital tool to analyse MET data and detect the GE pattern [18,30]. Genotypes 85 (ETW17-377), 82 (ETW17-374) and 13 (ETW17-281) were found to be high-yielding and stable, while 90 (ETW17-383) was the most high-yielding but unstable genotype. Genotypes with both high performance and stability across test environments contribute to the success of breeding in reducing the impact of GE, thereby developing cultivars that perform better in drought stress and non-stress conditions [29]. Similary genotypes were differenciated based on their yield performance and stability in different crops, including bread wheat [24], durum wheat [26], sorghum [25] and oat [31]. The which-won-where view of the GGE biplot showed which genotypes performed best in which environments. According to this biplot, most genotypes placed close to the center, indicating stable performance of the genotypes across the selected test environments [18]. The selected five environments fall into three sectors where genotype 2 (ETW17-269) and 36 (ETW17-306) were the winner genotypes at Kulumsa site planted on 4th July (E7) and at the same site planted on 2nd August (E8) while 13 (ETW17-281) and 90 (ETW17-383) were the winner at the DebreZeit site planted on 20th July (E3) and 118 (Check released cultivar “Ogolcho”) was the winner genotype at the Dera site planted on 5th July (E5). This pattern suggests that the target environment may consist of three mega-environments and that different cultivars should be selected and deployed for each. However, a representative set of environments across years is required to see reliable classification of mega-environments [32].
4.4. Traits Relationships
Yield component traits are governed by different genes, resulting in variation of genotype stability [21]. The genotype x trait biplot estimates the genetic correlations among the traits [17]. In this analysis, trait relationships showed that TKW and PH as important indirect selection criteria for identifying and developing early maturing and high-yielding varieties (Figure 5). This result corroborated the correlation and path analyses made on the same genotypes that revealed TKW and PH as the most yield contributing traits under drought and non-stress environmental regimes [33]. The effect of TKW on grain yield performance of the genotypes conforms with the result reported by Annicchiarico and Pecetti [34]. Plant height is a farmer-prefered trait along with the grain yield in sorghum [25]. Similarly, plant height was reported as an important trait contributing to yield in maize under drought stress [35]. Another interesting feature of a graphical view of the biplot, a genetic covariate by environment biplot enables interpretation of G and GE of the target trait, such as grain yield in terms of covariate by environment interactions, where other traits are considered as explanatory traits or covariates [32]. Based on this analysis, selection for larger TKW and earliness may lead to increased yield in all environments except in the Dera site planted on 5th July (E5). The outcomes of the biplots are in conformity with Badu-Apraku et al. [35], who reported ear aspect, plant aspect and Striga damage as the most reliable traits for indirect selection for improved grain yield under Striga infestation in maize. TKW and PH constitute one group of traits (had acute angle) with similar effects on yield. DH, MD, KS and SS constitute another group. The effects of the two groups were opposite, as indicated by the obtuse angles between them. This trait association could result from the genetic correlation among traits for which indirect selection can be devised to explore the GE interaction for grain yield [18].
5. Conclusions
The study assessed the how to utilize GE interaction of target trait. Based on the GGE biplot analysis, genotypes 85 (ETW17-377), 82 (ETW17-374) and 13 (ETW17-281) are the best yielding and stable. The Kulumsa site planted on 4th of July 2017 (environment 7=E7) and the DebreZeit site on 20th of July 2017 (E3) corresponding to early or optimum plantings, were the most effective environments in distinguishing genotypes and can thus be suitable environments for future evaluation and selection with specific adaptation. The Melkasa site planted on 4th July (E9) was a representative environment where genotypes can be evaluated for wide adaptation. TKW and PH are essential traits for indirect selection to exploit the GE interaction for grain yield of bread wheat. However, there is a need to include data sets across years and further statistical tests to validate the current results and make reliable inferences.
Authors contributions: YS analyzed the data and drafted the manuscript. HS reviwed and edited the manuscript.
Funding
The study was funded by the Alliance for Green Revolution in Africa (AGRA) through the African Centre for Crop Improvement of University of KwaZulu-Natal.
Acknowledgments
We thank the Kulumsa Agricultural Research Centre (KARC), Melekassa Agricultural Research Centre (MARC) and DebreZeit Agricultural Research Centre (DZARC) for providing technical support.
Conflicts of Interest
No conflict of interest is declared.
References
- Erenstein, O.; Jaleta, M.; Sonder Kai. ; Mottaleb K.; Prasanna, B.M. Global maize production, consumption and trade: trends and R&D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar]
- FAO. Food and Agriculture of the United Nations. Rome. Italy. 2024.
- Gupta, V.; Kumar, M.; Singh, V.; Chaudhary, L.; Yashveer, S.; Sheoran, R.; Dalal, M.S.; Nain, A.; Lamba, K.; Gangadharaiah, N.; Sharma, R.; Nagpal, S. Genotype by environment interaction analysis for grain yield of wheat (Triticum aestivum (L.) em.Thell) genotypes. Agric. 2022, 12, 1002. [Google Scholar] [CrossRef]
- USDA. World Agricultural Production. 2023.
- Tadesse, W.; Solh, M.; Braun, H.J.; Oweis, T.; Baum, M. Approaches and strategies for sustainable wheat production. International Centre for Agricultural Research in the Dry Areas (ICARDA), Beirut, Lebanon. 2016.
- Farshadfar, E.; Mohammadi, R.; Aghaee, M.; Vaisi, Z. GGE biplot analysis of genotype × environment interaction in wheat-barley disomic addition lines. Aust J Crop Sci. 2012, 6, 1074–1079. [Google Scholar]
- Adhikari, U.; Nejadhashemi, A.P.; Woznicki, S.A. Climate change and eastern Africa: a review of impact on major crops. Food Energy Secur. 2015, 4, 110–132. [Google Scholar] [CrossRef]
- FAO. The Eastern Horn of Africa faces an exceptional prolonged and persistent agro-pastoral drought sequence. Reports contributed by Global Information and Early Warning System on Food and Agriculture (GIEWS), Rome, Italy. 2021.
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS One 2013, 8, e67280. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Dube, E.; Laing, M.D.; Tsilo, T.J. Breeding wheat for drought tolerance: Progress and technologies. J Integr Agric. 2016, 15, 935–943. [Google Scholar] [CrossRef]
- Semahegn, Y.; H. Shimelis; M. Laing; Mathew I. Evaluation of bread wheat (Triticum aestivum L.) genotypes for yield and related traits under drought-stressed conditions. Acta Agr Scand B-S-P. 2020, 70, 474–484. [Google Scholar]
- Lin, C.S.; Binns, M.R.; Lefkovitch, L.P. Stability analysis: where do we stand? Crop Sci. 1986, 26, 894–900. [Google Scholar] [CrossRef]
- Finlay, K.W.; Wilkinson, G.N. The analysis of adaptation in a plant breeding programme. Aust J Agric Res. 1963, 14, 742–752. [Google Scholar] [CrossRef]
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Gauch, H.G. Model selection and validation for yield trials with interaction. Biom. 1988, 44, 705–715. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar evaluation and mega environment investigation based on the GGE biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Yang, R.C.; Crossa, J.; Cornelius, P.L.; Burgueno, J. Biplot analysis of genotype x environment interaction: Proceed with caution. Crop Sci. 2009, 49, 1564–1576. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. An integrated biplot analysis system for displaying, interpreting, and exploring genotype x environment interaction. Crop Sci. 2005, 45, 1004–1016. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A. Biplot analysis of multi-Environment trial data In. Quantitative Genetics, Genomics and Plant Breeding. Kang, M. S., (Ed.), CABI Publishing, New York. 2002, 289-303.
- Mohammadi, R.; Amri, A. Analysis of genotype x environment interaction in rain-fed durum wheat of Iran using GGE-biplot an non-parametric methods. Can J Plant Sci. 2012, 92, 757–770. [Google Scholar] [CrossRef]
- Rakshit, S.; Ganapathy, K.N.; Gomashe, S.S.; Rathore, A.; Ghorade, R.B. , Nagesh Kumar, M.V.; Ganesmurthy, K.; Jain, S.K.; Kamtar, M.Y.; Sachan, J.S.; Ambekar, S.S.; Ranwa, B.R.; Kanawade, D.G.; Balusamy, M.; Kadam, D.; Sarkar, A.; Tonapi, V.A.; Patil, J.V. GGE biplot analysis to evaluate genotype, environment and their interactions in sorghum multi-location data. Euphytica 2012, 185, 465–479. [Google Scholar]
- Yan, W.; Hunt, L.A. Biplot analysis of multi-environment trial data In. Quantitative Genetics, Genomics and Plant Breeding. Kang, M. S., (Ed.), CABI Publishing, New York. 2002, 289-303.
- Abakemal, D.; Shimelis, H.; Derera, J. Analyses of combining ability and genotype-by-environment interaction of quality protein maize inbred lines adapted to tropical-highlands. Crop Sci. 2016, 56, 1–12. [Google Scholar] [CrossRef]
- Alemu, G.; Dabi, A.; Geleta, N.; Solomon, T.; Delessa, A.; Duga, R.; Zegeye, H.; Asnake, D.; Getamesay, A.; Asefa, B.; Zewdu, D.; Abeyo, B.G.; Badebo, A.; Girma, B. Development of bread wheat (Triticum aestivum L.) varieties for high moisture areas of Ethiopia: A G x E interaction and stability analysis for grain yield. Ethiop. J Crop Sci. 2020, 8, 87–104. [Google Scholar]
- Enyew, M.; Feyissa, T.; Geleta, M.; Tesfaye, K.; Hammenhag, C.; Carlsson, A.S. Genotype by environment interaction, correlation, AMMI, GGE biplot and cluster analysis for grain yield and other agronomic traits in sorghum (Sorghum bicolor L. Moench). PLoS ONE 2021, 16, e0258211. [Google Scholar] [CrossRef] [PubMed]
- Mulugeta, B.; Tesfaye, K.; Geleta, M.; Johansson, E.; Hailesilassie, T.; Hammenhag, C.; Hailu, F.; Ortiz, R. Multivariate analyses of Ethiopian durum wheat revealed stable and high yielding genotypes. PLoS ONE 2022, 17, e0273008. [Google Scholar] [CrossRef] [PubMed]
- Bocianowski, J.; Warzecha, T.; Nowosad, K.; Bathelt, R. Genotype by environment interaction using AMMI model and estimation of additive and epistasis gene effects for 1000-kernel weight in spring barley (Hordeum vulgare L.). J Appl Genet. 2019, 60, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Antelo, J.L.; Benitez-Riquelme, I.; Vargas-Hernandez, M.; Huerta-Espino, J.; Bentley, A. R.; Villasenor-Mir, H. E.; Pinera-Chavez, F. J. Multi-location trials identify stable high-yielding spring bread and durum wheat cultivars in Mexico. Crop Sci. 2023, 63, 2103–2114. [Google Scholar] [CrossRef]
- Stella, T.; Webber, H.; Rezaei, E.E.; Asseng, S.; Martre, P.; Dueri, S.; Guarin, J.R.; Pequeno, D.N.L.; Calderini, D.F.; Reynolds, M.; Molero, G.; Miralles, D.; Garcia, G.; Slafer, G.; Giunta, F.; Kim, Y.; Wang, C.; Ruane, A.C.; Ewert, F. Wheat crop traits conferring high yield potential may also improve yield stability under climate change. in silico Plants 2023, 5, 1–16. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–655. [Google Scholar] [CrossRef]
- Kebede, G.; Worku, W.; Jifar, H.; Feyissa, F. GGE biplot analysis of genotype by environment interaction and grain yield stability of oat (Avena sativa L.) in Ethiopia. Agrosyst Geosci Environ. 2023, 6, e20410. [Google Scholar] [CrossRef]
- Yan, W.K.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can J Plant Sci. 2005, 86, 623–645. [Google Scholar] [CrossRef]
- Semahegn, Y.; Shimelis, H.; Laing, M.; Mathew, I. Genetic variability and association of yield and yield components among bread wheat genotypes under drought-stressed conditions. Aust J Crop Sci. 2021, 15, 863–870. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Pecetti, L. Contribution of some agronomic traits to durum wheat performance in a dry Mediterranean region of Northern Syria. Agronomie 1993, 13, 25–34. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, B.; Akinwale, R.; Annor, B.; Adewale, S.; Toyinbo, J.; Akintibu, S. 2020. Application of the GGE Biplot as a statistical tool in the breeding and testing of early and extra-early maturing maize in Sub-Saharan Africa. Crop Breed Genet Genom. 2020, 2, e200012. [Google Scholar] [CrossRef]
Figure 1.
Relationship among test environments in discriminating the genotypes using the environment-vector view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
Figure 1.
Relationship among test environments in discriminating the genotypes using the environment-vector view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
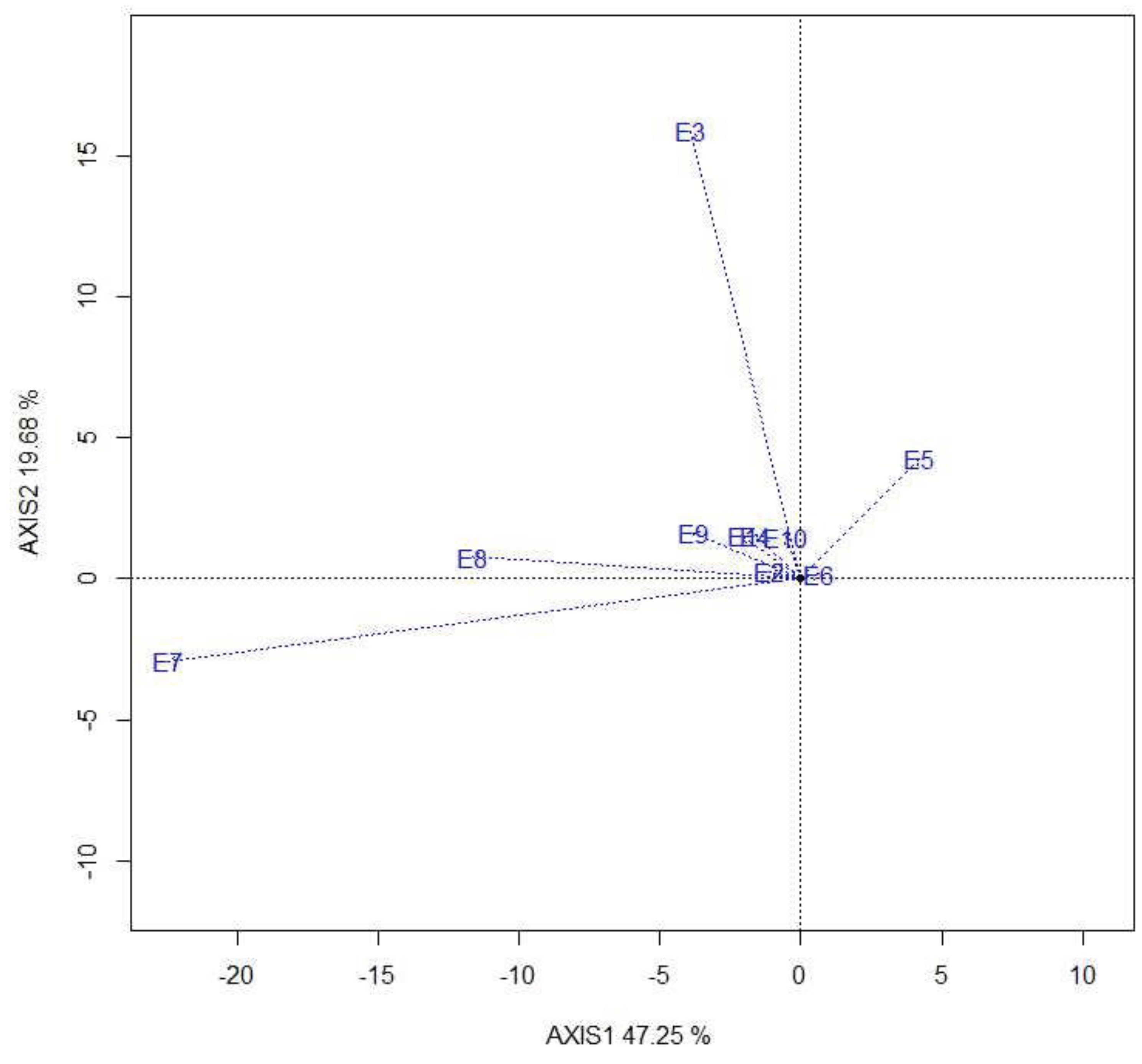
Figure 2.
Ranking environments based on both discriminating ability and representativeness using the environment-vector view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
Figure 2.
Ranking environments based on both discriminating ability and representativeness using the environment-vector view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
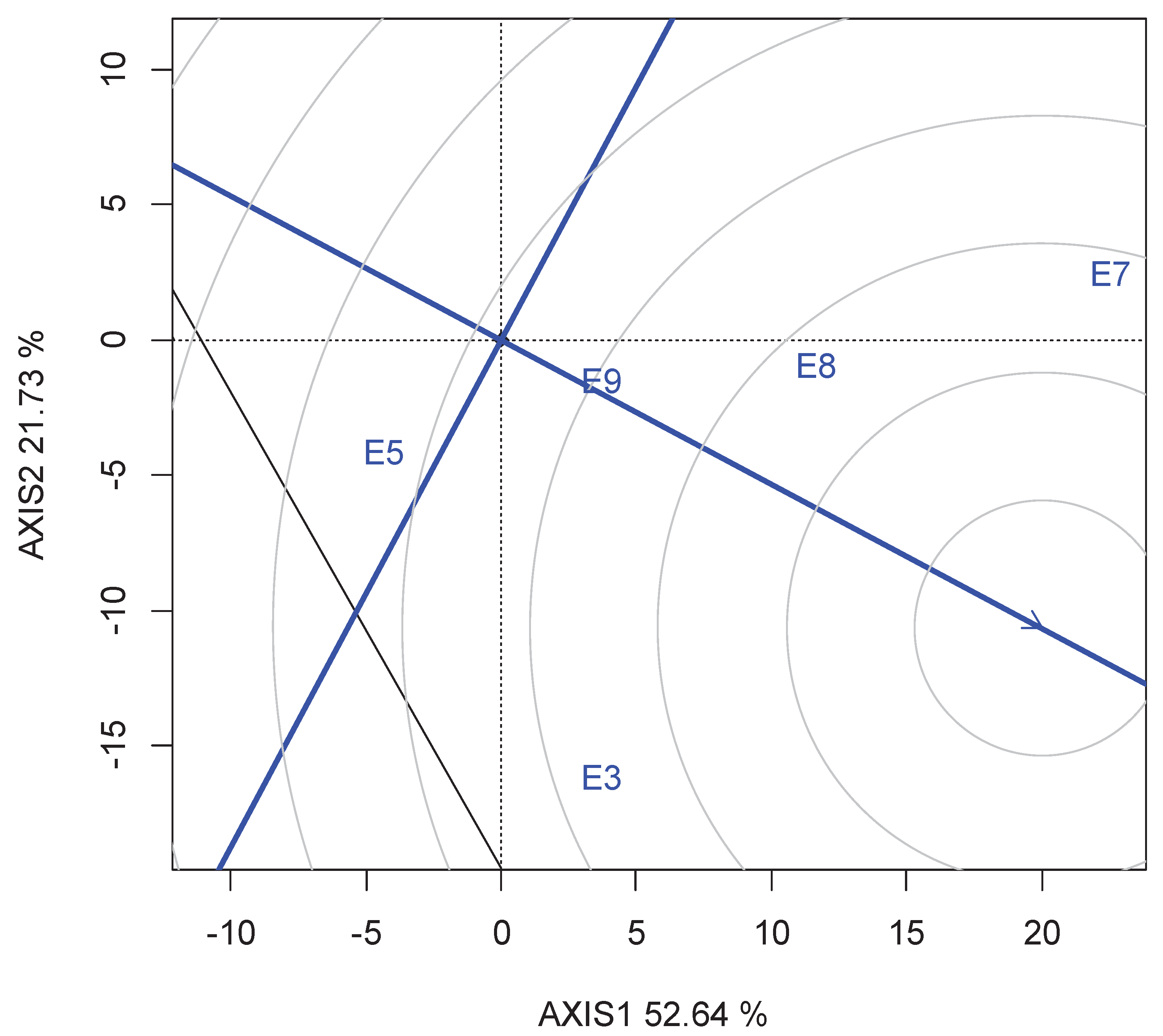
Figure 3.
Ranking genotypes relative to an ideal genotype (the center of the concentric circles) using the average-environment coordination view (AEC) of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is genotype-metric preserving (SVP = 1). Genotypes coded as YS-1, YS-2… in the prevous study [11] designated here as 1,2…,respectively.
Figure 3.
Ranking genotypes relative to an ideal genotype (the center of the concentric circles) using the average-environment coordination view (AEC) of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is genotype-metric preserving (SVP = 1). Genotypes coded as YS-1, YS-2… in the prevous study [11] designated here as 1,2…,respectively.
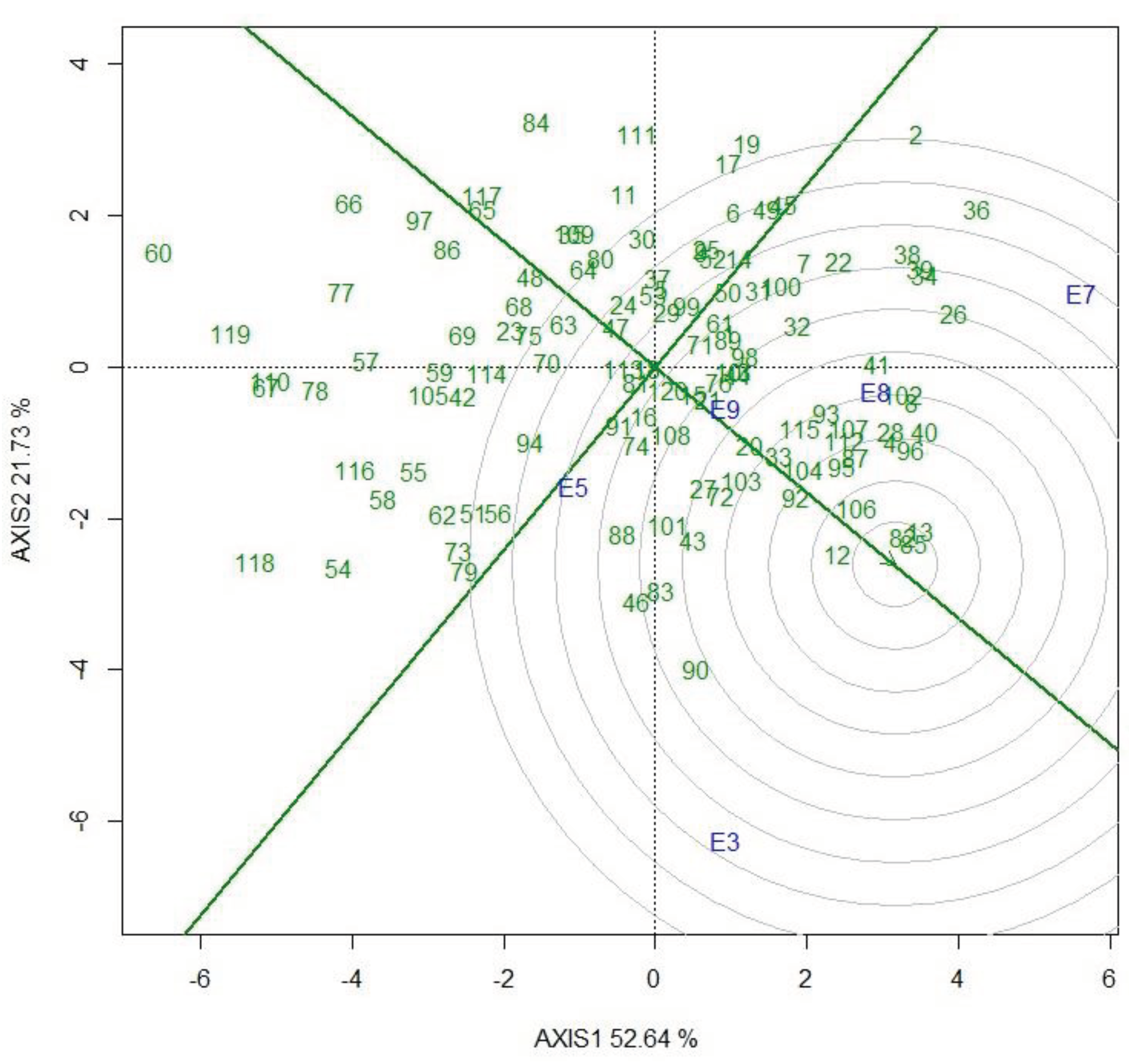
Figure 4.
The “which-won-where” view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
Figure 4.
The “which-won-where” view of the GGE biplot based on an environment-centered (Centering = 2) genotype x environment dataset without any scaling (scaling = 0), and it is environment-metric preserving (SVP = 2).
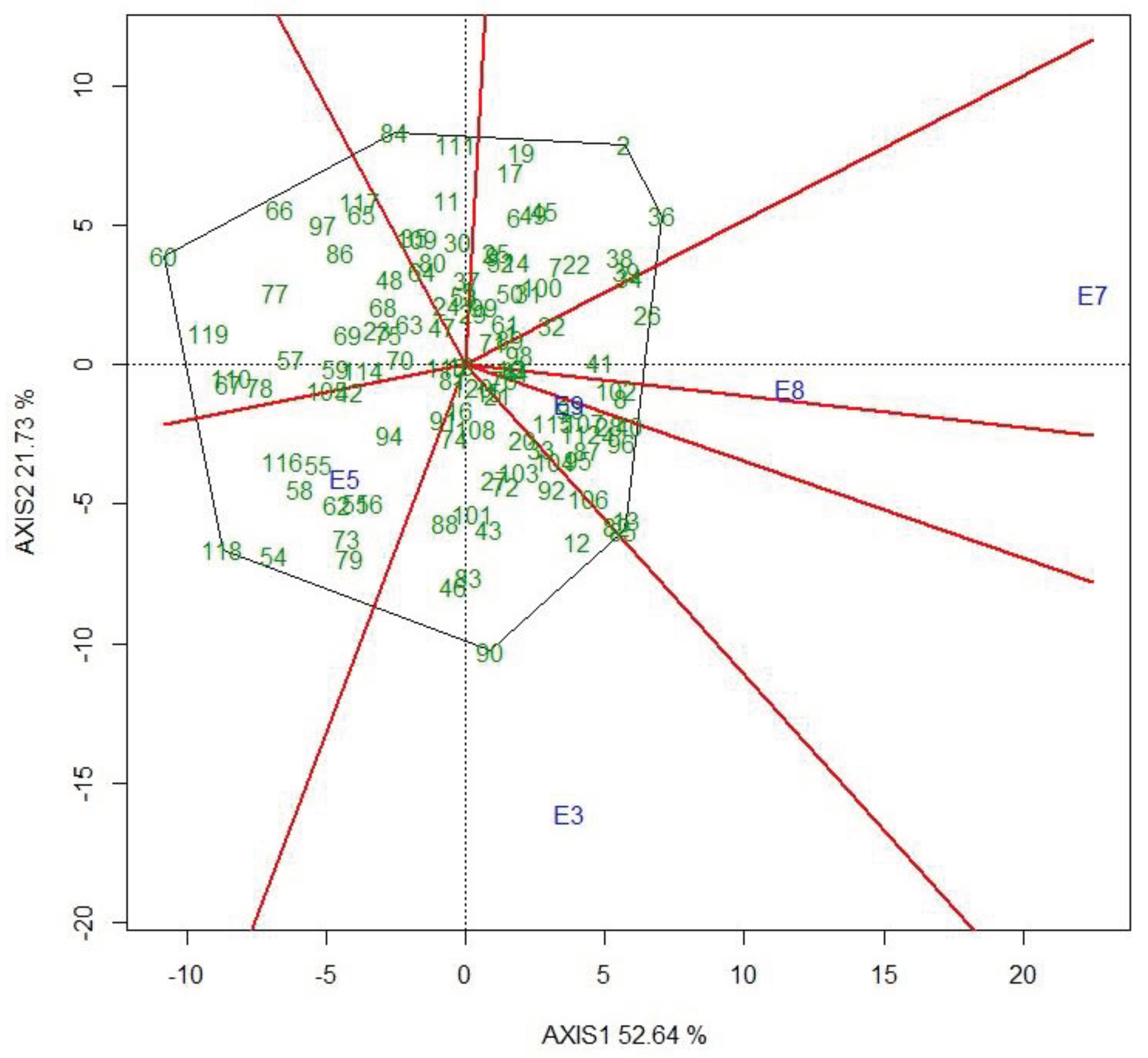
Figure 5.
Genotype by trait biplot showing the genetic correlations among seven traits across genotypes using the trait-vector view of the GGE biplot based on a trait-centered (Centering = 2) genotype x trait dataset with scaling (scaling = 1), and it is trait-metric preserving (SVP = 2). DM-days to maturity; PH-plant height (cm); SL-spike length (cm); SS-number of spikelet per spike; KS-number of kernel per spike; TKW-thousand kernel weight.
Figure 5.
Genotype by trait biplot showing the genetic correlations among seven traits across genotypes using the trait-vector view of the GGE biplot based on a trait-centered (Centering = 2) genotype x trait dataset with scaling (scaling = 1), and it is trait-metric preserving (SVP = 2). DM-days to maturity; PH-plant height (cm); SL-spike length (cm); SS-number of spikelet per spike; KS-number of kernel per spike; TKW-thousand kernel weight.
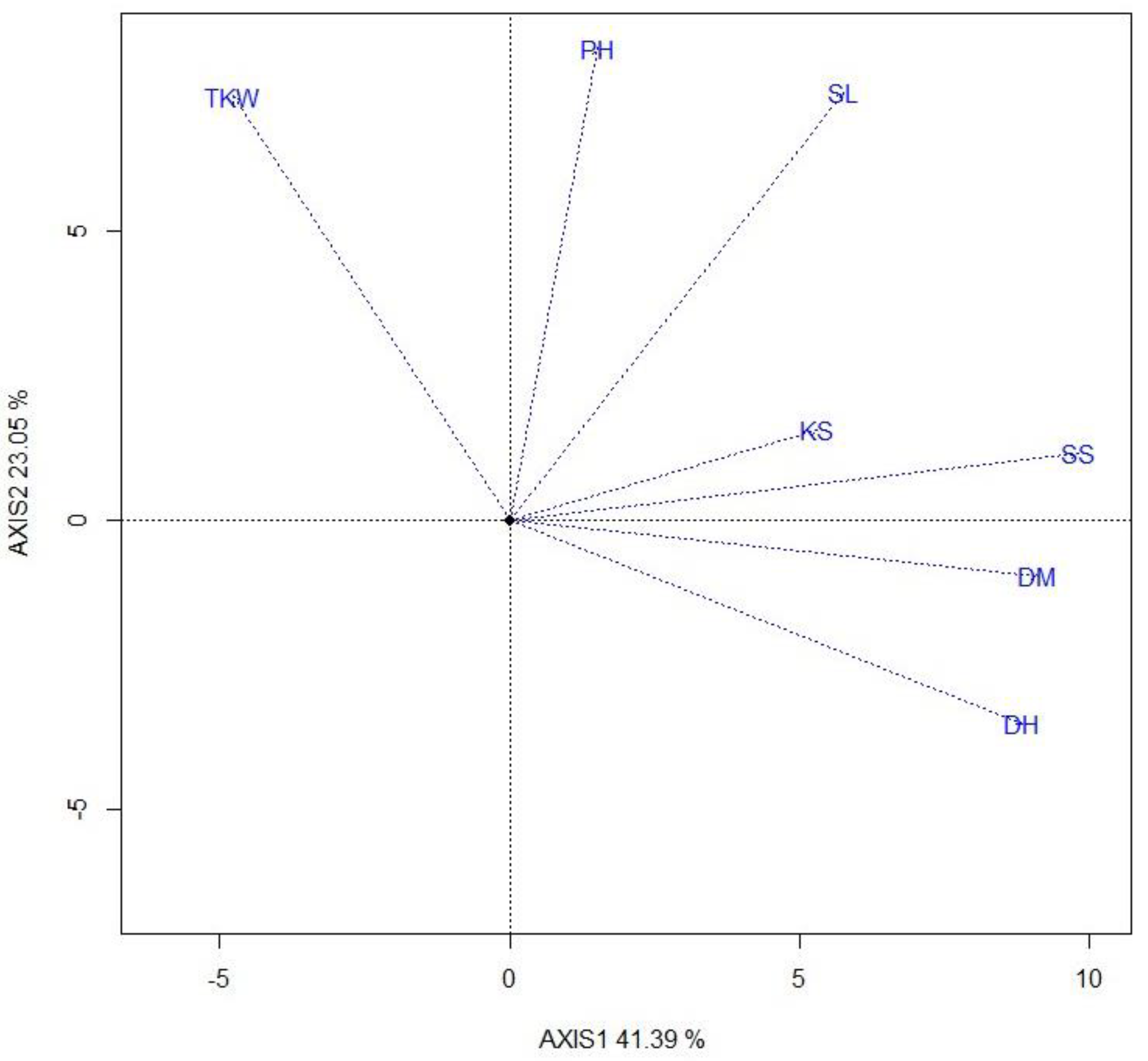
Figure 6.
A genetic covariate by environment view using the GGE biplot based on a covariate-centered (Centering = 2) covariate x environment dataset with scaling (scaling = 1), and it is covariate-metric preserving (SVP = 1).
Figure 6.
A genetic covariate by environment view using the GGE biplot based on a covariate-centered (Centering = 2) covariate x environment dataset with scaling (scaling = 1), and it is covariate-metric preserving (SVP = 1).
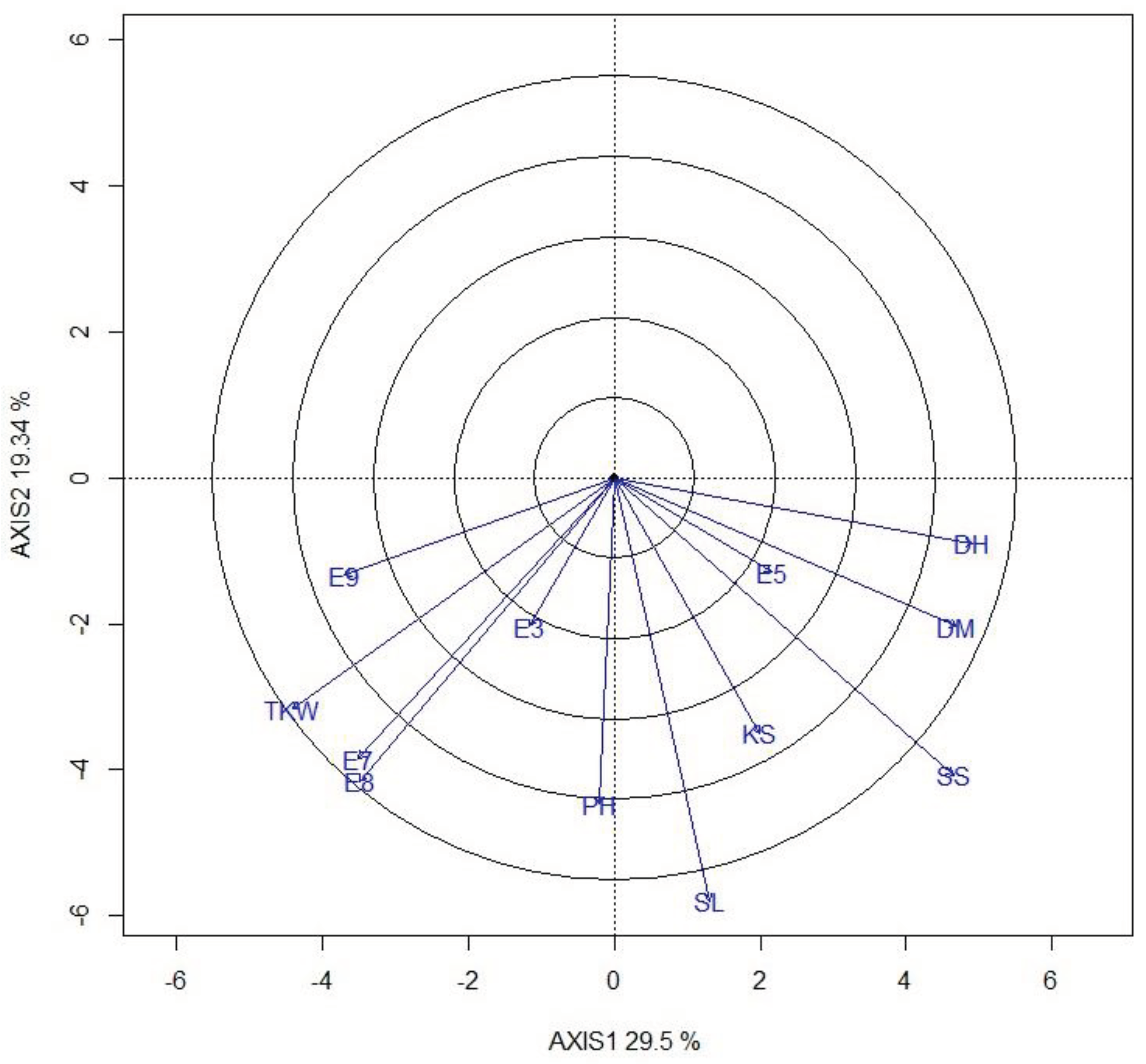

Table 1.
Pooled analysis of variance for grain yield of 120 genotypes tested across 10 test environments in 2017.
Table 1.
Pooled analysis of variance for grain yield of 120 genotypes tested across 10 test environments in 2017.
| Source | DF | Mean Square | % SST |
|---|---|---|---|
| Environment (E) | 9 | 3094.09*** | 85.2 |
| Replication(E) | 10 | 12.31*** | 0.38 |
| Block(E*Replication) | 180 | 2.50*** | 1.38 |
| Genotype (G) | 119 | 4.17*** | 1.52 |
| GE interaction | 1071 | 1.87*** | 6.14 |
| Error | 1008 | 1.31 | 4.04 |
| Trial statistics | |||
| Mean | 3.79 | ||
| LSD | 0.71 | ||
| CV (%) | 30.2 | ||
| R2 | 0.96 |
DF-degree of freedom; %SST-percentage relative to the sum of squares total; LSD-least significant difference; CV-coefficient of variation; R2-coefficient of determination.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



