
Submitted:
25 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Background: Histone deacetylases (HDACs) are implicated in carcinogenesis, and HDAC inhibitors (HDACis) are explored as a therapeutic tool in several tumors. The aim of this study was to evaluate the clinical significance of HDAC -2, -4, and -5 expression in epithelial ovarian carcinoma (EOC). Methods: HDAC-2, -4, and -5 immunohistochemical expression was examined in 92 EOC tissue specimens and was correlated with clinicopathological characteristics. Results: HDAC-2 was the most frequently (94.4%) expressed isoform, being marginally higher in serous tumors compared with other types (p=0,08). HDAC-5 was the less frequently expressed (28.1%), being positively associated with HDAC-4. HDAC-4 positivity was associated with lower FIGO-stage (p=0,045) and T-category (p=0.,043) and the absence of lymph node (p=0,05) or distant metastasis (p=0,09) in serous carcinomas. HDAC-2 positivity was correlated with absence of lymph node metastasis in serous tumors (p=0,045). On the contrary, HDAC-5 nuclear positivity was correlated with lymph node metastasis in the entire cohort (p=0,048). HDAC-4 positivity was marginally associated with favorable prognosis in serous carcinomas in univariate survival analysis (p=0,086), a correlation which was not significant in multivariate analysis. Conclusion: These findings suggest a differential expression of HDAC-2, -4 and -5, providing evidence for a potential role in the pathobiology of EOC.
Keywords:
HDAC
; ovarial adenocarcinoma
; immunohistochemistry
; prognosis
1. Introduction
Epigenetic alterations are essential regulators of gene transcription and have been reportedly correlated with carcinogenesis and tumor progression. These post-translational modifications include acetylation, methylation, phosphorylation, ubiquitination, sumoylation and ADP-ribosylation of the long N-terminal extensions of the DNA-bound core histones [1]. Acetylation is regulated by the balance of two opposite classes of enzymes: histone acetyl transferases (HATs), which transfer the acetyl moiety from acetyl coenzyme A to specific lysine residues of histones and histone deacetylaces (HDACs), which remove acetyl groups from histone specific lysine residues [2,3]. The latter establishes a positive charge on the N-termini of the histones therefore strengthening histone tail-DNA interactions, and blocking access of the transcriptional machinery to the DNA template, thus suppressing gene transcription [2,3].
The members of the HDAC protein family are divided into four classes according to the corresponding homology to the yeast S. Cerevisiae. Among them class I HDACs encompasses HDAC-1, -2, -3 and -8, with homology with the yeast proteins Rpd3, Hos1 and Hos2. HDAC-1 and -2 are the most closely related members of the class I family and seem to play a role in the regulation of cell cycle genes. Class I and II members share a common enzymatic mechanism, the Zn-catalyzed hydrolysis of the acetyl-lysine amide bond. Class II HDACs is divided in two different subclasses (IIa and IIb) and comprise of HDAC-4, -5, -6, 7, -9 and 10, which share homology with the yeast proteins HDA1 and Hos3. The class IIa HDACs, (HDAC- 4, -5, -7, -9), are associated with transcription factors of the MEF and Runx families. Class IIb HDACs (HDAC-6 and -10) have a duplication of their catalytic domains. HDAC-6 is the only deacetylase known to act on tubulin, which is required for disposal of misfolded proteins in aggresomes. Class III HDACs (sirtuins) are structurally distinct from class I and II HDACs and homologues of the yeast silent information regulator 2 (Sir2). Sirtuins catalyze the transfer of the acetyl group onto the sugar moiety of Nicotinamide adenine dinucleotide, namely NAD. Class IV consists of an additional Zn-dependent HDAC (HDAC-11) that is phylogenetically different from class I and II enzymes [1,4,5,6].
HDAC proteins have been recently shown to be overexpressed in a wide range of human malignancies, being associated with tumor initiation and disease progression as well as patients’ prognosis [6]. In various tumor types a higher level of HDACs is associated with advanced disease and poor outcome, presumably due to the loss of tumor suppressor functions. There are, however, multiple reports of suggesting an important role in the prevention of metastasis being associated with better clinical outcomes [7,8]. The mechanisms by which individual members of the HDAC protein family can regulate tumor development are quite diverse and several reports suggest a distinct type-specific role of HDACs overexpression [9]. HDACs induce a range of cellular and molecular effects through hyperacetylation of histone and nonhistone substrates and therefore could either repress tumor suppressor gene expression or regulate the oncogenic cell-signaling pathway via modification of key molecules [10].
There is to the best of our knowledge limited information available to date regarding the clinical significance of different classes of HDACs expression in ovarian adenocarcinomas and in particular Class II members, even though the role of HDAC inhibitors in the treatment of ovarian adenocarcinoma is already being thoroughly discussed [11]. In view of the above considerations, the present study aims to assess retrospectively the immunohistochemical expression of several members of HDAC protein family classes I and IIa in specimens of ovarian adenocarcinomas, in association with clinicopathological parameters as well as patients’ overall survival.
2. Materials and Methods
2.1. Patients Collective
This is a study of archival histopathological material from 92 patients with ovarian carcinoma diagnosed during 2016–2018 at the Department of Pathology, in Klinikum Nuremberg, Germany, for whom medical records were available. All patients underwent a surgical resection and none of them had received any kind of neoadjuvant radio- or chemotherapy prior to surgery. Patients were assigned a clinical stage according to the International Federation of Obstetrics and Gynecology (FIGO) standards. Surgical and pathological findings and postoperative abdominopelvic computerized tomography scans were used to determine the FIGO stage for the ovarian adenocarcinomas and the residual disease after initial surgery. The study was conducted in accordance with the Declaration of Helsinki and was approved by the internal institutional Bioethics Committee of the Paracelsus University, Nuremberg, Germany. Due to the retrospective nature of the studies, as well as the lack of impact on the treatment of patients, it was not necessary to obtain informed consent.
2.2. Immunohistochemical Analysis
Immunohistochemistry (IHC) for HDAC-2, -4, and -5 was performed on formalin-fixed, paraffin-embedded tissue sections using the following antibodies: rabbit polyclonal anti-HDAC-2 (Y461, ab32117, Abcam, Cambridge, United Kingdom), mouse monoclonal anti-HDAC-4 (ab16339, Abcam, Cambridge, United Kingdom), and mouse monoclonal anti-HDAC-5 (ab55403, Abcam, Cambridge, United Kingdom). Immunostaining was performed using Autostainer 48 (DAKO, Glostrup, Denmark). Liquid Permanent Red (DAKO) was used as a detection system. Red chromogen visualization kit enabled visualization in tissues containing a large amount of melanin. Suitable negative and positive controls were used as appropriate [12].
IHC evaluation was performed by counting at least 1000 tumor cells in each case independently by two experienced pathologists (S.T., G.L.), blinded to clinical information, with complete interobserver compliance. Nuclear and cytoplasmic immunoreactivity was evaluated separately. The extent of various HDACs expression was calculated by the percentage of positive tumor cells to the total number of tumor cells. The staining intensity was estimated in four categories: 0 (no reaction), 1 (mild reaction), 2 (moderate reaction), and 3 (intense reaction).
2.3. Statistical Analysis
Statistical analysis was performed by a MSc biostatistician (GL). The association between the IHC expression of HDAC-2, -4 and -5 and clinicopathological characteristics was examined using non-parametric tests with correction for multiple comparisons, as appropriate. Survival analysis was performed using Kaplan–Meier survival curves and the differences between the curves were compared with log-rank test. Cox regression analysis was used to evaluate to potential prognostic value of each parameter independent of the remaining available parameters. A p-value of <0.05 was considered statistically significant. A p-value of > 0.05 but lower of < 0.10 was considered of marginal significance. The analysis was performed with the statistical package STATA 11.0/SE for Windows.Interventionary studies involving animals or humans, and other studies that require ethical approval, must list the authority that provided approval and the corresponding ethical approval code.
3. Results
3.1. Patients’ Characteristics
The patients’ characteristics are shown in Table 1. The median age was 62 years (range 31-92 years). There were 74 serous carcinomas, 3 mucinous carcinomas, 5 endometrioid carcinomas, 2 clear cell carcinomas, 1 mixed mucinous-endometrioid carcinoma, 2 carcinosarcomas, and 6 serous borderline tumors. Tumor stage according to FIGO was as follows: 30% has a FIGO-stage I, 11.1% a FIGO-stage II, 40% a FIGO-stage III and 18.9% a FIGO-stage IV. For two cases there was not any information regarding tumor stage. Among carcinomas eight patients had a low grade (LG) and 68 had a high grade (HG) carcinoma. Follow-up information was available in 80 patients, 27 of whom died of their disease within a period of 0.1-77.9 months. Twenty-five patients had a lymph node metastasis and 18 a metastatic disease at diagnosis.
3.2. HDAC-2 Expression and Associations with Clinicopathological Characteristics
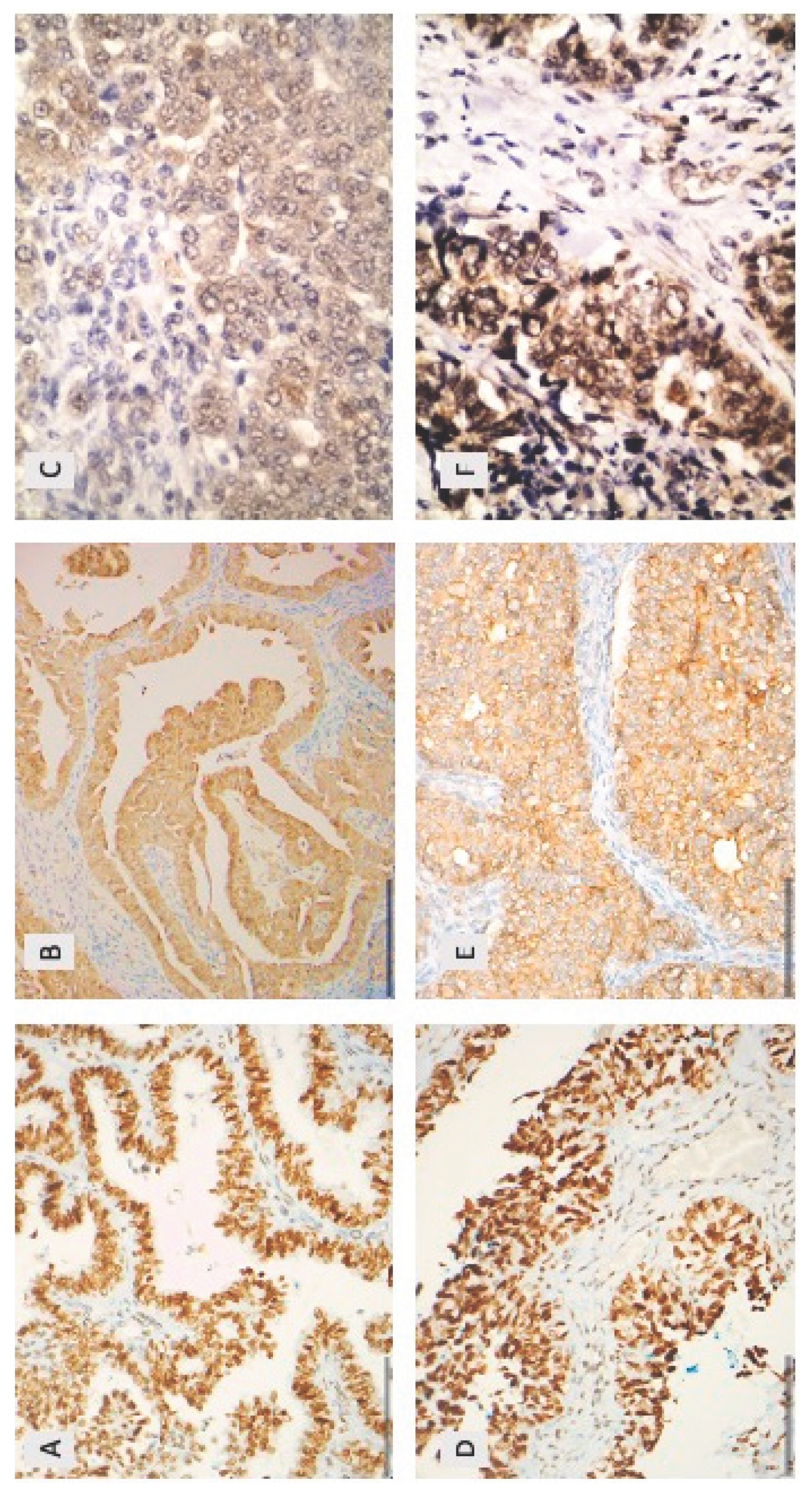
HDAC-2 immunopositivity was observed in 94.4% of the examined cases, all cases showing a nuclear staining (Figure 1A,D, Table 2). The majority of the cases displayed an increased immunoexpression (67.8% of the cases with a positive expression in >80% of the tumor population) with a strong staining intensity (61% of the cases).
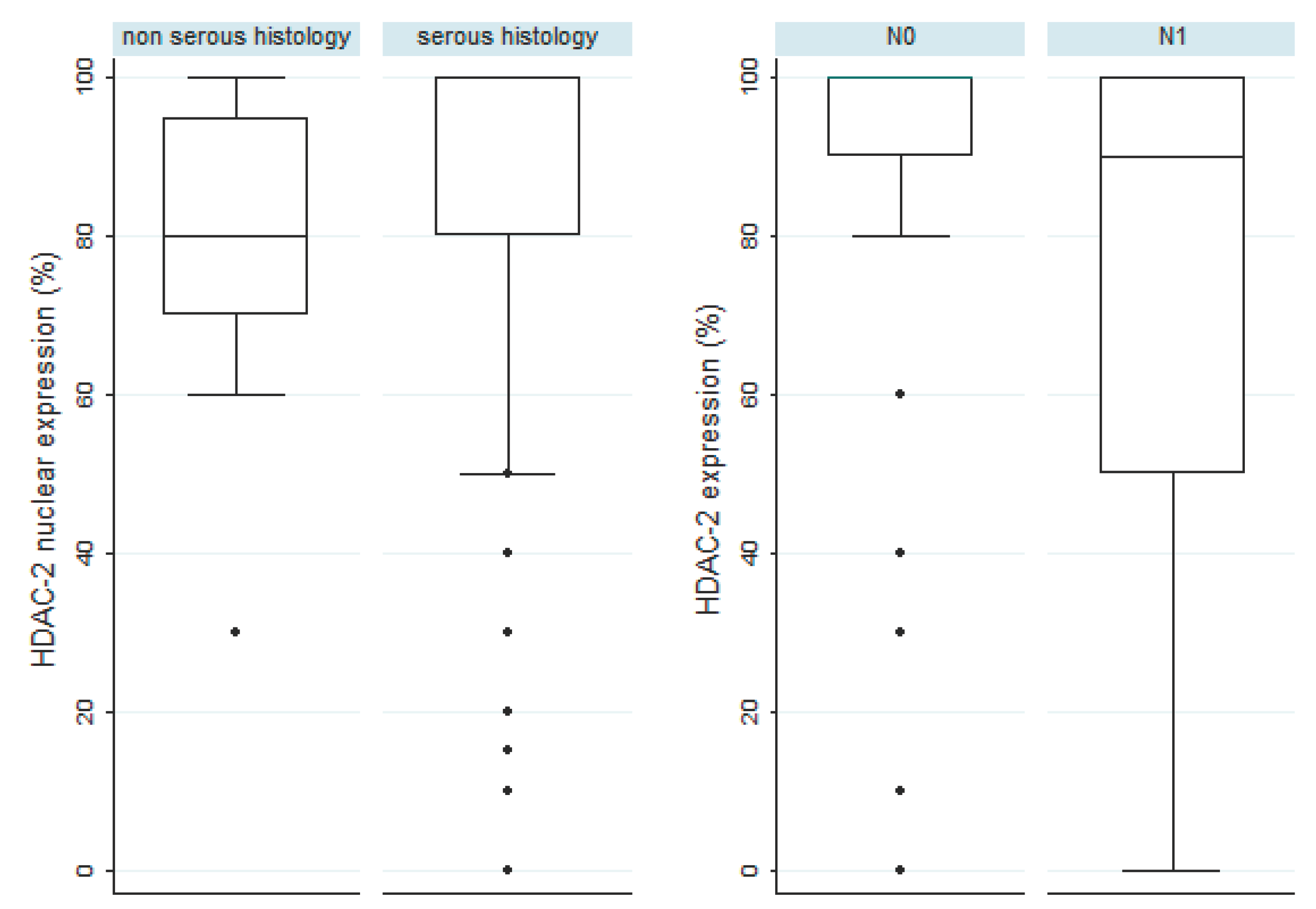
Patients with increased HDAC-2 nuclear immunoexpression were slightly older than those with lower HDAC-2 expression (64 versus 58 years old, Mann Whitney U test, p=0.045). Serous histology was mostly associated with higher levels of HDAC-2 expression, a correlation which was of marginal significance (median HDAC-2 expression 80% in non-serous tumors versus 100% in serous tumors, Mann-Whitney U test, p=0.08, Figure 2). There was however not any significant difference in HDAC-2 expression between patients with serous borderline tumors and serous carcinomas (p>0,10). Moreover, there was not any statistically significant association between HDAC-2 expression and FIGO stage, T-status, histological grade, the presence of metastasis or relapse, not only in the whole cohort but also in an analysis restricted to serous tumors. Interestingly, the presence of lymph node metastasis was associated with lower levels of HDAC-2 expression, an association which was stronger in tumors with serous histology (Mann Whitney U test, p=0.045, Figure 2) and was of borderline significance in the whole cohort (p=0.081).
3.3. HDAC-4 Expression and Associations with Clinicopathological Characteristics
HDAC-4 immunopositivity was cytoplasmic and was observed in 80.4% of the investigated cases (median value of immunoexpression 30%) (Figure 1B,E, Table 2). The vast majority of the positive cases (77,2%) had a relatively weak staining intensity, whereas only 22.8% displayed a moderate and none of the cases a strong staining intensity.
Figure 1.
Immunohistochemical expression of HDAC-2 (A, D), HDAC-4 (B, E) and HDAC-5 (C, F) in epithelial ovarian carcinomas. (A) A low grade serous ovarian carcinoma with nuclear HDAC-2 expression (x200) (B) a low grade serous ovarian carcinoma with cytoplasmic HDAC-4 expression (x200) (C) high grade serous ovarian carcinoma with cytoplasmic HDAC-5 expression (x200) (D) high grade serous ovarian carcinoma with nuclear HDAC-2 expression (x200) (E) high grade serous ovarian carcinoma with cytoplasmic HDAC-4 expression (x400) (F) a high grade serous ovarian carcinoma with nuclear and cytoplasmic HDAC-4 expression (x400).
Figure 1.
Immunohistochemical expression of HDAC-2 (A, D), HDAC-4 (B, E) and HDAC-5 (C, F) in epithelial ovarian carcinomas. (A) A low grade serous ovarian carcinoma with nuclear HDAC-2 expression (x200) (B) a low grade serous ovarian carcinoma with cytoplasmic HDAC-4 expression (x200) (C) high grade serous ovarian carcinoma with cytoplasmic HDAC-5 expression (x200) (D) high grade serous ovarian carcinoma with nuclear HDAC-2 expression (x200) (E) high grade serous ovarian carcinoma with cytoplasmic HDAC-4 expression (x400) (F) a high grade serous ovarian carcinoma with nuclear and cytoplasmic HDAC-4 expression (x400).
There was not any significant difference in the levels of HDAC-4 expression between tumors with or without serous histology (p<0.10). Similarly, the expression in borderline tumors seemed to be comparable to that observed in carcinomas (p<0.10), and there was not any difference among low- or high-grade carcinomas (p>0.10). Advanced FIGO stage tumors (IV) were more frequently negative for HDAC-4 (35,3%) compared to the tumors with lower FIGO stage (I-III, 16,2%), a relationship which was of marginal significance in the entire cohort (Chi square test, p=0.07), but was proven to be significant in tumors with serous histology (borderline and carcinomas, p=0.05) and especially in serous carcinomas (p=0.045). A similar association was observed regarding tumor T-status especially in the subset of serous carcinomas, where all T1-T3 cases were positive for HDAC-4 compared to 23.6% of T4 cases (Chi-square test, p=0.043), whereas the respective relationship was of marginal significance in tumors with serous histology (borderline and carcinomas, p=0.078) and failed to attain statistical significance when the analysis was performed in the entire cohort (p>0,10). Likewise, serous carcinomas presenting without a metastatic disease at diagnosis displayed more frequently a positive immunoreactivity for HDAC-4 (86.8% versus 66.7% for cases with metastatic disease, Chi-square test, p=0.05), an association which was of marginal significance when the analysis was performed in the entire cohort of carcinomas (p=0.09). Similarly, tumors without lymph node metastasis were more frequently positive for HDAC-4 compared to tumors with lymph node metastasis, an association which was however of borderline significance in serous carcinomas as well as in the entire cohort, Chi-square test p=0.09.
Figure 2.
Associations of HDAC-2 nuclear expression with serous histology (first panel) and with the presence of lymph node metastasis at the time of diagnosis in serous carcinomas (second panel).
Figure 2.
Associations of HDAC-2 nuclear expression with serous histology (first panel) and with the presence of lymph node metastasis at the time of diagnosis in serous carcinomas (second panel).
3.4. HDAC-5 Expression and Associations with Clinicopathological Characteristics
HDAC-5 immunoreactivity in epithelial ovarian carcinomas was mainly cytoplasmic with some cases displaying an additional nuclear immunostaining. Cytoplasmic HDAC-5 expression was observed in 28.1% of the examined cases (25/89) and nuclear in only 10% of the cases (9/89) (Figure 1C,F, Table 2). The vast majority of the investigated cases did not show any positive expression of HDAC-5 (78.9%). The staining intensity was weak in most of the positive cases (76% of the positive cases), the remaining cases showing a moderate staining intensity. There was a positive correlation between nuclear and cytoplasmic HDAC-5 expression (Spearman’s correlation coefficient, rho=0,53, p<0.001, Table 3).
There was no significant difference in the levels of HDAC-5 expression (cytoplasmic or nuclear) between tumors with or without serous histology (p<0.10). HDAC-5 expression did not differ between borderline tumors and carcinomas (p<0.10. Moreover, there was not any statistically significant association between HDAC-5 expression and FIGO stage, T-status, histological grade, the presence of metastasis or relapse not only in the whole cohort but also when the analysis was restricted to serous tumors. Interestingly, the presence of lymph node metastasis was associated with nuclear HDAC-5 positivity in the entire cohort (Fisher’s exact test, p=0.048).
3.5. Associations between HDAC-2, HDAC-4 and HDAC-5
There was a positive correlation between HDAC-4 and HDAC-5 expression (Spearman correlation coefficient, rho=0.27, p=0.012 for nuclear HDAC-5 and rho=0.24 p=0.029 for cytoplasmic HDAC-5). These correlations remained significant when the analysis was restricted to serous tumors (data not shown). The associations among HDAC-2 and HDAC-4 or HDAC-5 were not significant. The associations between the investigated HDACs are presented in Table 3.

Table 3.
Associations among the investigated HDACs in epithelial ovarian carcinomas. Star denotes a statistically significant result.
Table 3.
Associations among the investigated HDACs in epithelial ovarian carcinomas. Star denotes a statistically significant result.
| Nuclear HDAC-2 | Cytoplasmic HDAC-4 | Cytoplasmic HDAC-5 | |
|---|---|---|---|
| Cytoplasmic HDAC-4 | rho=-0.09 p=0.390 | ||
| Cytoplasmic HDAC-5 | rho=-0.03 p=0.781 | rho=0.24 p=0.029* | |
| Nuclear HDAC-5 | rho=-0.03 p=0.792 | rho=0.27 p=0.012* | rho=0.5274 p<0.001* |
3.6. Survival Analysis
In univariate survival analysis for overall survival only HDAC-4 immunopositivity in serous carcinomas seemed to be associated with better OS, a correlation which was only of a marginal significance (log-rank test, p=0.086, Figure 3). The respective associations of HDAC-2 and HDAC-5 (nuclear or cytoplasmic) with OS in the entire cohort as well as in serous tumors or serous carcinomas was not significant (log rank test, p>0.10). In our cohort tumor T-category (p=0.004), FIGO stage (p=0.004), the presence of relapse (p=0.007) and the presence of residual disease (p=0.035) were associated with an adverse patient prognosis. Table 4 illustrates the results of univariate survival analysis in our cohort.
However, in multivariate survival analysis none of the investigated HDACs remained significant, after adjustment to the prognostic parameters, which were proven to be significant in univariate survival analysis.
4. Discussion
Epithelial ovarian cancer (EOC) is one of the leading causes of cancer mortality in women, being characterized by late-stage at presentation and poor patient prognosis [11]. Despite the significant progress in the therapeutic approaches for cancer over the past 20 years, which have undoubtedly revolutionized anti-ovarian-cancer therapy, 5-year relative survival rates amount less than 40% [13]. The first line treatment encompasses surgery aiming not only at the reduction of tumor volume but also at staging the disease. Cytotoxic chemotherapy with a platinum agent and a taxane, which can be given before or after surgery is a mandatory element of treatment [14]. Some cases recur within 6 months after completion of initial chemotherapy and are considered “platinum-resistant”. Chemoresistance is the main challenge for ovarian carcinoma, being responsible for treatment failure and unfavorable clinical outcomes. The dismal prognosis of EOC along with the limitations of the available therapeutic modalities have set the stage for the investigation of the potential use of novel epigenetic therapies, including HDAC-inhibitors, either alone or in combination with other therapies.
The word “epigenetics” has been derived from the Greek prefix epi (“in annexation, on the top of, all over’’), which refers to features that are either “on the extrinsic surface” or “in annexation” to the genetic basis of inheritance. Several epigenetic alterations have been observed in EOC, mainly altered DNA methylation, in terms of pan-hypomethylation of heterochromatin, and regional CpG island methylation [15], with some reports also focusing on the role of Histone acetylation [16,17]. In this context, Caslini et al. studied how histone modifications affected expression of GATA transcription factors on five ovarian cancer cell lines and more specifically GATA4 and GATA6 gene silencing was found to correlate with hypoacetylation of histones H3 and H4 [16]. Genome-wide studies have also revealed that genetic alterations affecting expression of histone-modifying genes are present in ovarian carcinoma [18].
To date the available information regarding the role of HDACs in EOC is limited to four articles, three of which investigated only members of class I, namely HDACs-1, -2 and -3 [9,19,20]. All three studies report an increased expression of all three class I HDACs (mRNA or immunohistochemical) in ovarian carcinomas, being higher than that observed in normal tissue [9,19,20]. Interestingly, Weichert et al. suggest a lower expression of these class I HDACs in endometrioid ovarian carcinomas compared to serous carcinomas [20]. The third study focuses on a member of class IIa, namely HDAC-4 and presents overexpression of this molecule in EOC, suggesting also a role in repression of p21 [21]. However, to the best of our knowledge there is not any investigation regarding the expression of HDAC-5 in EOC. Similarly, there is not any published information regarding the potential correlation or interplay of members of different classes HDACs.
In accordance with the previous studies investigating HDAC-2 in EOC [9,19,20], we also observed in our study HDAC-2 immunopositivity in the vast majority of the examined cases (94.4%). A similar rate of HDAC-2 positivity is also reported by Weichert et al. [20]. Moreover HDAC-2 was the most extensively expressed HDAC among the examined HDACs in our cohort, with 67.8% of the cases having a positive expression in >80% of the neoplastic cells, exhibiting a strong staining intensity in 61% of the positive cases. These results are in alignment with previous investigations in uveal melanoma [8,22]. HDAC-4 was also expressed in 80% of the cases, having however a relatively weak staining intensity in most of the cases, none of them displayed a strong staining intensity. On the contrary, HDAC-5 was the least expressed HDAC in our cohort, being positive in 28.1% of the examined cases.
In this study, we observed a nuclear HDAC-2 staining, in consistency with previous reports in other malignancies as well as in studies on ovarian carcinoma and in keeping with the fact that class I HDACs are reported to be ubiquitously located in cell nucleus, due to a lack of a nuclear export signal [8,20,23]. Most of the published investigations in several human malignancies focus on the role of class I HDACs in the nucleus. Some studies also report a cytoplasmic immunoreactivity, the function of which remains unclear [12,24]. In this context, Hayashi et al. observed a positive HDAC-2 in both the cytoplasm and the nucleus of the cell [9], an observation which however was not repeated in our cohort. On the other hand, HDAC-4 exhibited only cytoplasmic and HDAC-5 mainly cytoplasmic immunoreactivity, in keeping with the reported staining patterns of these proteins [8]. This concomitant nuclear and cytoplasmic immunolocalization of HDAC-5 according to the literature can be attributed to the, capability of class II HDACs of nucleocytoplasmic shuttling in response to certain cellular signals [25].
Importantly, we found a different expression of HDAC-2 among different histological types. In this context, serous tumors (borderline and carcinomas) displayed an increased HDAC-2 immunoreactivity, a correlation which was however of marginal significance, probably due to the low number of non-serous tumors included in our investigation. This result is in alignment with the observations reported by Weichert et al. [20], who present an increased expression of HDAC-2 in serous carcinomas compared to endometrioid carcinomas, the latter showing the lowest expression levels among all histological types. On the contrary, we did not find any significant difference in the expression of HDAC-4 and -5 between serous histology and non-serous histology.
Interestingly we observed an inverse association of HDAC-4 expression with FIGO stage and tumor T-status. In particular, the presence of HDAC-4 immunopositivity was associated with lower FIGO stage (I-III) and T-status (T1-T3), both associations being stronger in the subgroup of serous carcinomas. Similarly, HDAC-4 immunopositivity was correlated with the absence of metastatic disease at diagnosis in serous carcinomas. A similar result, although of marginal significance was observed regarding lymph node metastasis, the HDAC-4 negative cases presenting more frequently with lymph node metastasis at diagnosis. This observation is in contrast with the results of Shen et al., who report increased HDAC-4 expression in ovarian cancer specimens with advanced stage disease, without however specifying the histological subtype of the investigated cases in their cohort [21]. This inverse association of HDAC-4 observed in our cohort could possibly be an event presenting especially in the subset of serous tumors. According to the literature, HDACs can induce a broad range of cellular and molecular effects through hyperacetylation of histone and nonhistone substrates and in this context to regulate and modify a wide range of signaling pathways [10], being therefore able to induce different events in different types of tumors. A similar negative association of HDAC-4 expression with the presence of distant metastasis has also been reported in human pancreatic adenocarcinomas [26].
In our study not only lower HDAC-4 but also lower HDAC-2 expression was associated with the presence of lymph node metastasis, a correlation that was stronger in serous tumors and of marginal significance in the whole cohort, as illustrated in Figure 2. The presence of a similar association of HDAC-2 and -4 with lymph node metastasis is interesting, especially since there was not any significant association between HDAC-2 and HDAC-4. However, the respective associations of HDAC-2 with FIGO stage, tumor T-category and presence of distant metastasis at diagnosis failed to attain statistical significance. HDAC-2 is the most investigated member of the HDAC family in the literature, showing very frequently associations with parameters correlated with patient prognosis. There are only isolated examples in which HDAC-2 expression levels were not correlated with clinicopathological parameters, as for example in the investigation by Giaginis et al. in pancreatic adenocarcinomas [26]. Moreover, Hayashi et al. report an increased expression of HDAC-2 in carcinomas as compared to borderline tumors in a cohort comprising of both serous and mucinous tumors [9], a result which however was not repeated in our investigation.
Regarding the associations of HDAC-5 expression with clinicopathological parameters, most of the associations were not proven to be statistically significant, which is probably in accordance with the limited number of positive cases observed in our investigation. Interestingly, the presence of nuclear HDAC-5 immunohistochemical positivity was associated with the presence of lymph nodes metastasis. Similarly, Peng et al. observed that the suppression of miR-671-5p or promotion of HDAC5 expression encouraged ovarian carcinoma tumor growth in animal models [27].
Moreover, we found significant positive associations between the immunohistochemical expression of HDAC-4 and HDAC-5 (nuclear or cytoplasmic), both being class IIa HDACs. This is the first investigation in the literature in which more than one member of class II family has been investigated. Our result implicates a possible interplay between the two molecules. Further investigations are however needed in order to confirm and explain this observation.
Another interesting finding of the present investigation is that the presence of HDAC-4 expression in serous carcinomas connotes a better survival probability. To the best of our knowledge, this appears to be the first report investigating the potential prognostic role of class II HDACs in epithelial ovarian carcinomas. HDAC-2 and -5 expression does not seem to be informative in this regard. According to a comprehensive review of Weichert, class I HDAC isoforms are mostly expected to be associated with poor patient survival, whereas high expression of class II isoforms seems to predict a better patient outcome [28]. However, the correlation of HDAC-4 with favorable prognosis failed to remain significant in our cohort in multivariate survival analysis, probably indicating that this association observed in univariate analysis reflects the respective association of HDAC-4 expression with FIGO stage and tumor T-status.
The present investigation is the first study analyzing concomitantly the expression of members of different classes of the HDAC protein family. It has however, some limitations that need to be taken into consideration. For example, our cohort encompasses mainly tumors with serous histology, comprising 86.9% of the investigated cases. However, it should be mentioned that the results of survival analysis in our investigation recapitulate many of the traditional parameters that have been proposed as important determinants of clinical outcome in epithelial ovarian carcinomas, namely FIGO stage and tumor T-category as well as the presence of relapse or residual disease after surgery, supporting the validity of statistical analysis and denoting that our cohort is representative.
In the last decade, a variety of compounds that can block the deacetylase activity of HDACs have been recognized, and synthetic or natural molecules targeting class I, II, and IV HDACs have been developed [29]. Moreover, preclinical studies have found that HDAC inhibitors (HDACis) are able to inhibit ovarian cancer cell growth in vitro and in vivo by inhibiting the cell cycle and inducing mitotic defects through histone-mediated and histone-independent interactions [30]. Recently, numerous clinical trials investigate the role of HDACis alone or in combination with other drugs in patients with epithelial ovarian cancer, showing encouraging anti-tumor effects [29], especially when combined with other chemotherapeutics. This combination seems to show chemosensitizing or synergistic antitumor efficacy, which may be due to their ability to overcome particular mechanisms associated with drug resistance [31]. However, additional studies are needed to determine the efficacy of these therapies.
5. Conclusions
The present investigation is the first study which simultaneously investigates the immunohistochemical expression of HDAC members of different classes and shows clearly that different members of the HDAC family are differentially expressed in EOC, HDAC-2 being the most frequently and HDAC-5 the least frequently expressed isoform among the investigated HDACs. Moreover, HDAC molecules seem to have a different role in serous tumors compared with other histological types. Additionally, significant positive associations between HDAC-4 and HDAC-5 expression levels were observed, implicating a potential interplay between members of class II. HDAC-4 expression was mostly correlated with clinicopathological parameters, being associated with lower FIGO stage and tumor T-category as well as the absence of lymph node metastasis or distant metastasis. These findings provide evidence for a potential role of HDACs in the biological mechanisms governing epithelial ovarian cancer evolution and progression. Further studies offering a deeper understanding on the possible role of HDACs and consequently of the mechanisms involved in the antitumor activity of HDACis are warranted.
Author Contributions
G.L. Conceptualization, Writing—Original Draft Preparation, Formal analysis, Supervision; D.A. Investigation (Data collection), Resources; D. I. B. Investigation (Data collection), Resources; R.M. Data collection; C.V. M. B. Investigation, Methodology; T. P. Investigation (Data collection); S. T. Conceptualization, Writing—Review and Editing. Each author has approved the submitted version and agrees to be personally accountable for the author’s own contributions and for ensuring that questions related to the accuracy or integrity of any part of the work, even ones in which the author was not personally involved, are appropriately investigated, resolved, and documented in the literature. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and it was approved by the Bioethics Committee of the Paracelsus University of Nuremberg, Germany (IRB-2023-16, 12th October 2023).
Informed Consent Statement
Not applicable. Due to the retrospective nature of the studies and the lack of impact on the treatment of patients, it was not necessary to obtain informed consent.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gallinari, P.; Marco, S.D.; Jones, P.; Pallaoro, M.; Steinkühler, C. HDACs, Histone Deacetylation and Gene Transcription: From Molecular Biology to Cancer Therapeutics. Cell Res 2007, 17, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.Y.; Denu, J.M.; Allis, C.D. Histone Acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef] [PubMed]
- Marks, P. Histone Deacetylases. Current Opinion in Pharmacology 2003, 3, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Blander, G.; Guarente, L. The Sir2 Family of Protein Deacetylases. Annu. Rev. Biochem. 2004, 73, 417–435. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, A.J.M.; Gennip, A.H. van; Caron, H.N.; Kemp, S.; Kuilenburg, A.B.P. van Histone Deacetylases (HDACs): Characterization of the Classical HDAC Family. Biochemical Journal 2003, 370, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Goutas, D.; Theocharis, S.; Tsourouflis, G. Unraveling the Epigenetic Role and Clinical Impact of Histone Deacetylases in Neoplasia. Diagnostics 2021, 11, 1346. [Google Scholar] [CrossRef]
- Hu, X.; Xing, W.; Zhao, R.; Tan, Y.; Wu, X.; Ao, L.; Li, Z.; Yao, M.; Yuan, M.; Guo, W.; et al. HDAC2 Inhibits EMT-Mediated Cancer Metastasis by Downregulating the Long Noncoding RNA H19 in Colorectal Cancer. J Exp Clin Cancer Res 2020, 39, 270. [Google Scholar] [CrossRef] [PubMed]
- Levidou, G.; Gajdzis, P.; Cassoux, N.; Donizy, P.; Masaoutis, C.; Gajdzis, M.; Gardrat, S.; Pergaris, A.; Danas, E.; Klijanienko, J.; et al. Histone Deacetylase (HDAC)-1, -2, -4, and -6 in Uveal Melanomas: Associations with Clinicopathological Parameters and Patients’ Survival. Cancers 2021, 13, 4763. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Horiuchi, A.; Kikuchi, N.; Hayashi, T.; Fuseya, C.; Suzuki, A.; Konishi, I.; Shiozawa, T. Type-specific Roles of Histone Deacetylase (HDAC) Overexpression in Ovarian Carcinoma: HDAC1 Enhances Cell Proliferation and HDAC3 Stimulates Cell Migration with Downregulation of E-cadherin. Intl Journal of Cancer 2010, 127, 1332–1346. [Google Scholar] [CrossRef]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb Perspect Med 2016, 6, a026831. [Google Scholar] [CrossRef]
- Moufarrij, S.; Dandapani, M.; Arthofer, E.; Gomez, S.; Srivastava, A.; Lopez-Acevedo, M.; Villagra, A.; Chiappinelli, K.B. Epigenetic Therapy for Ovarian Cancer: Promise and Progress. Clin Epigenet 2019, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Pouloudi, D.; Manou, M.; Sarantis, P.; Tsoukalas, N.; Tsourouflis, G.; Dana, E.; Karamouzis, M.V.; Klijanienko, J.; Theocharis, S. Clinical Significance of Histone Deacetylase (HDAC)-1, -2, -4 and -6 Expression in Salivary Gland Tumors. Diagnostics 2021, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Psilopatis, I.; Schaefer, J.I.; Arsenakis, D.; Bolovis, D.; Levidou, G. SOX11 and Epithelial-Mesenchymal Transition in Metastatic Serous Ovarian Cancer. Biomedicines 2023, 11, 2540. [Google Scholar] [CrossRef]
- Falzone, L.; Scandurra, G.; Lombardo, V.; Gattuso, G.; Lavoro, A.; Distefano, A.; Scibilia, G.; Scollo, P. A Multidisciplinary Approach Remains the Best Strategy to Improve and Strengthen the Management of Ovarian Cancer (Review). Int J Oncol 2021, 59, 53. [Google Scholar] [CrossRef] [PubMed]
- Qazi, S.; Sharma, A.; Raza, K. The Role of Epigenetic Changes in Ovarian Cancer: A Review. Indian J Gynecol Oncolog 2021, 19, 27. [Google Scholar] [CrossRef]
- Caslini, C.; Capo-chichi, C.D.; Roland, I.H.; Nicolas, E.; Yeung, A.T.; Xu, X.-X. Histone Modifications Silence the GATA Transcription Factor Genes in Ovarian Cancer. Oncogene 2006, 25, 5446–5461. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.W.Y.; Huang, Y.-W.; Hartman-Frey, C.; Kuo, C.-T.; Deatherage, D.; Qin, H.; Cheng, A.S.L.; Yan, P.S.; Davuluri, R.V.; Huang, T.H.-M.; et al. Aberrant Transforming Growth Factor Β1 Signaling and SMAD4 Nuclear Translocation Confer Epigenetic Repression of ADAM19 in Ovarian Cancer. Neoplasia 2008, 10, 908–IN2. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.H.M.; Afroze, S.; Osuji, G.; Kayani, S.; Colon, N.; Pantho, A.; Kuehl, T.; Pilkinton, K.; Uddin, M. Epigenetic Modifications in Ovarian Cancer: A Review. J Cancer Treat & Diagnosis 2020, 4, 17–35. [Google Scholar] [CrossRef]
- Jin, K.L.; Pak, J.H.; Park, J.-Y.; Choi, W.H.; Lee, J.-Y.; Kim, J.-H.; Nam, J.-H. Expression Profile of Histone Deacetylases 1, 2 and 3 in Ovarian Cancer Tissues. J Gynecol Oncol 2008, 19, 185. [Google Scholar] [CrossRef]
- Weichert, W.; Denkert, C.; Noske, A.; Darb-Esfahani, S.; Dietel, M.; Kalloger, S.E.; Huntsman, D.G.; Köbel, M. Expression of Class I Histone Deacetylases Indicates Poor Prognosis in Endometrioid Subtypes of Ovarian and Endometrial Carcinomas. Neoplasia 2008, 10, 1021–1027. [Google Scholar] [CrossRef]
- Shen, Y.-F.; Wei, A.-M.; Kou, Q.; Zhu, Q.-Y.; Zhang, L. Histone Deacetylase 4 Increases Progressive Epithelial Ovarian Cancer Cells via Repression of P21 on Fibrillar Collagen Matrices. Oncology Reports 2016, 35, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Levinzon, L.; Madigan, M.; Nguyen, V.; Hasic, E.; Conway, M.; Cherepanoff, S. Tumour Expression of Histone Deacetylases in Uveal Melanoma. Ocul Oncol Pathol 2019, 5, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Mottet, D.; Castronovo, V. Histone Deacetylases: Target Enzymes for Cancer Therapy. Clin Exp Metastasis 2008, 25, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.; Nielsen, T.O. Histone Deacetylase 1 and 2 in Mesenchymal Tumors. Modern Pathology 2012, 25, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Mutze, K.; Langer, R.; Becker, K.; Ott, K.; Novotny, A.; Luber, B.; Hapfelmeier, A.; Göttlicher, M.; Höfler, H.; Keller, G. Histone Deacetylase (HDAC) 1 and 2 Expression and Chemotherapy in Gastric Cancer. Ann Surg Oncol 2010, 17, 3336–3343. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Damaskos, C.; Koutsounas, I.; Zizi-Serbetzoglou, A.; Tsoukalas, N.; Patsouris, E.; Kouraklis, G.; Theocharis, S. Histone Deacetylase (HDAC)-1, −2, −4 and −6 Expression in Human Pancreatic Adenocarcinoma: Associations with Clinicopathological Parameters, Tumor Proliferative Capacity and Patients’ Survival. BMC Gastroenterol 2015, 15, 148. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Wu, T.; Wang, J.; Huang, J.; Zheng, L.; Wang, P.; Li, J.; Wu, L.; Luo, M. microRNA-671-5p Reduces Tumorigenicity of Ovarian Cancer via Suppressing HDAC5 and HIF-1α Expression. Chemico-Biological Interactions 2022, 355, 109780. [Google Scholar] [CrossRef] [PubMed]
- Weichert, W. HDAC Expression and Clinical Prognosis in Human Malignancies. Cancer Letters 2009, 280, 168–176. [Google Scholar] [CrossRef] [PubMed]
- 2Xu, L.; Yan, X.; Wang, J.; Zhao, Y.; Liu, Q.; Fu, J.; Shi, X.; Su, J. The Roles of Histone Deacetylases in the Regulation of Ovarian Cancer Metastasis. IJMS 2023, 24, 15066. [Google Scholar] [CrossRef]
- Garrett, L.A.; Growdon, W.B.; Rueda, B.R.; Foster, R. Influence of a Novel Histone Deacetylase Inhibitor Panobinostat (LBH589) on the Growth of Ovarian Cancer. J Ovarian Res 2016, 9, 58. [Google Scholar] [CrossRef]
- Zhou, M.; Yuan, M.; Zhang, M.; Lei, C.; Aras, O.; Zhang, X.; An, F. Combining Histone Deacetylase Inhibitors (HDACis) with Other Therapies for Cancer Therapy. European Journal of Medicinal Chemistry 2021, 226, 113825. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
Kaplan-Meier survival curves according to cytoplasmic HDAC-4 positivity in serous carcinomas.
Figure 3.
Kaplan-Meier survival curves according to cytoplasmic HDAC-4 positivity in serous carcinomas.
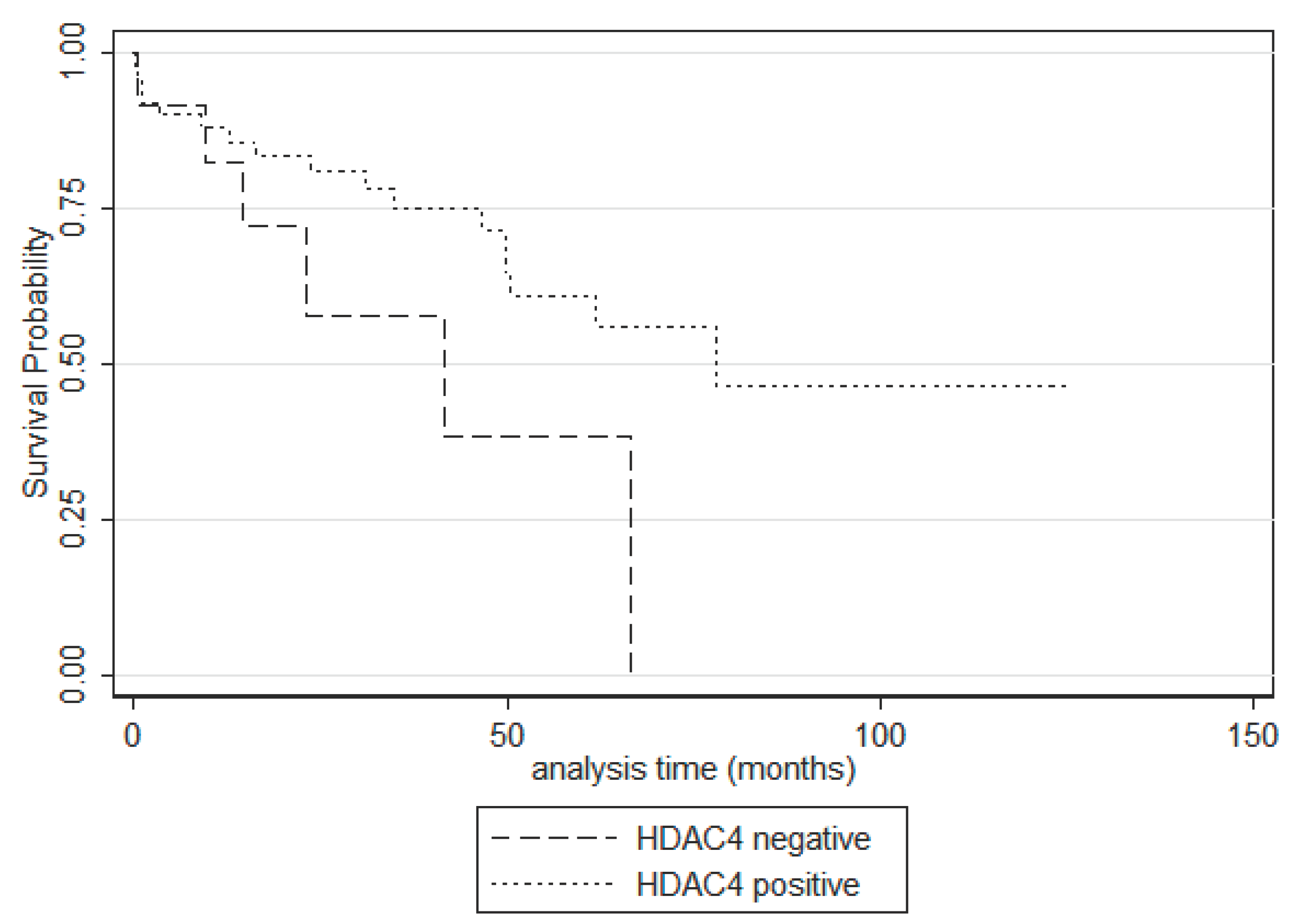
Table 1.
Patients’ characteristics.
| Patients’ characteristics | Median value | Range |
|---|---|---|
| Age (in years) | 62 | 31-92 |
| Number of patients | Percentage | |
| Histological subtype | ||
| Serous carcinoma | 74 | 80.4% |
| Mucinous carcinoma | 3 | 3.3% |
| Endometrioid carcinoma | 5 | 5.4% |
| Clear cell carcinoma | 1 | 1.1% |
| Mixed mucinous-endometrioid carcinoma | 1 | 1.1% |
| Carcinosarcoma | 2 | 2.2% |
| Serous borderline tumor | 6 | 6.5% |
| FIGO stage | ||
| I | 27 | 30% |
| II | 10 | 11.1% |
| III | 36 | 10% |
| IV | 17 | 18.9% |
| Tumor grade | ||
| Low grade | 18 | 21%% |
| High grade | 68 | 79% |
| T-status | ||
| T1 | 29 | 32% |
| T2 | 11 | 12% |
| T3 | 50 | 56% |
| N-status | ||
| N0 | 39 | 60.9% |
| N1 | 25 | 39.1% |
| Metastasis | ||
| Metastatic cancer | 18 | 19.6% |
| Non-metastatic cancer | 74 | 80.4%% |
| Residual disease | ||
| None | 25 | 29.1%% |
| Present | 10 | 11.6% |
| Unknown | 51 | 59.3% |
| Event | ||
| Death of disease | 27/80 (follow-up: 0.1-77.9 months) | 28% |
| Censored | 53/80 (follow-up: 0.3-125.2 months) | 71% |
| Relapse | ||
| Present | 36 | 58.1% |
Table 2.
Expression of the investigated HDACs in epithelial ovarian carcinomas.
| Positivity rate | median | Range | |
|---|---|---|---|
| Nuclear HDAC2 | 94.4% | 100 | 0-100 |
| Cytoplasmic HDAC4 | 80,4% | 30 | 0-95 |
| Cytoplasmic HDAC5 | 28,1% | 0 | 0-55 |
| Nuclear HDAC5 | 10% | 0 | 0-45 |
Table 4.
Univariate survival analysis in our cohort (log-rank test, p-value).
| Parameter | Entire cohort | Serous carcinomas |
|---|---|---|
| Age | ||
| FIGO stage | 0.005 | 0.015 |
| T-status | 0.005 | 0.045 |
| Histological Grade | 0.368 | 0.043 |
| Serous histology | 0.060 | - |
| Presence of metastasis | 0.116 | 0.097 |
| Presence of lymph node metastasis | 0.104 | 0.175 |
| Presence of residual disease | 0.035 | 0.027 |
| HDAC-2 positivity | 0.788 | 0.802 |
| HDAC-4 positivity | 0.105 | 0.086 |
| HDAC-5 cytoplasmic positivity | 0.967 | 0.940 |
| HDAC-5 nuclear positivity | 0.324 | 0.573 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



