
Submitted:
25 March 2024
Posted:
27 March 2024
You are already at the latest version
Abstract
Rotavirus (RV) replicates within viroplasms, membrane-less electron-dense globular cytosolic inclusions with liquid-liquid phase properties. In these structures occur the virus transcription, replication, and packaging of the virus genome in newly assembled double-layered particles. The viroplasms are composed of virus proteins (NSP2, NSP5, NSP4, VP1, VP2, VP3, and VP6), single and double-stranded virus RNAs, and host components such as microtubules, perilipin-1 and chaperonins. The formation, coalescence, maintenance, and perinuclear localization of viroplasms rely on their association with the cytoskeleton. A stabilized microtubule network involving microtubules and kinesin Eg5 and dynein molecular motors is associated with NSP5, NSP2, and VP2, facilitating dynamic processes such as viroplasm coalescence and perinuclear localization. Key post-translation modifications, particularly phosphorylation events of RV proteins NSP5 and NSP2, play pivotal roles in orchestrating these interactions. Actin filaments also contribute, triggering the formation of the viroplasms through the association of soluble cytosolic VP4 with actin and the molecular motor myosin. This review explores the evolving understanding of RV replication, emphasizing the host requirements essential for viroplasm formation and highlighting their dynamic interplay within the host cell.
Keywords:
rotavirus
; viroplasm
; cytoskeleton
; microtubule
; actin
; molecular motors
; lipid droplets
; NSP5
; NSP2
; VP2
; VP4
# These authors contributed equally to this work
1. Rotavirus
Rotavirus (RV) was initially observed in 1963 from electron microscopy of feces samples of young monkeys and mice presenting diarrhea [1]. In humans, the virus was first described in 1973 in the duodenal mucosa of infants with acute non-bacterial gastroenteritis [2]. Fifty years later, RV infections are the leading cause of severe gastroenteritis and dehydration in infants and young animals [3]. In 2008, before worldwide RV vaccine programs, RV-gastroenteritis led to 435'000 deaths worldwide, mainly in developing countries, and high-cost hospitalization in developed countries [4]. The introduction of vaccine programs reduced the disease burden by 85% in developed countries [5]; however, developing countries show a much more modest reduction in disease burden [6].
RV is a non-enveloped virus belonging to the order of Reovirales within the family Sedoreoviridae, where it belongs to the genus Rotavirus [7,8]. The genus Rotavirus currently entails nine different RV species, A-D and F-J, distinguished by serological criteria, host range, and sequence analysis [9]. The strains of RV species E were lost and removed from the species list by the ICTV in 2019 [10]. Moreover, recent reports indicate the identification and sequencing of RV species K and L [11,12]. However, the ICTV has not yet approved them.
The mature RV virion is a non-enveloped, icosahedral (T=13), triple-layered particle (TLP) [13] of about 100 nm in diameter [14]. The virion encapsidates one copy of each of the eleven double-stranded RNA (dsRNA) genome segments (gs). Each gs encodes for one protein, six structural proteins (VP1, VP2, VP3, VP4, VP6, and VP7), which are incorporated into the mature virion, and five (or six) non-structural proteins (NSP1, NSP2, NSP3, NSP4, NSP5, and in certain strains also NSP6 [15].
VP8* initiates RV cell entry by attaching to various cellular glycans [16], among them terminal sialic acids [17,18] and histo-blood group antigens (HBGAs) [19]. After cell binding, RV favors entry to the cell via clathrin-mediated endocytosis, although some strains also use clathrin-independent pathways [20,21].
A common step in RV entry is the localization of the virus particles into early endosomes, where they are exposed to environmental changes, such as an acidic pH, low calcium concentrations, or other lysosomal components. Those factors seem to be involved in the entrance of the virus into the cytosol [22], but they appear to be strain-dependent, as some strains seem to profit from maturing endosomes, while others use late endosomes [23]. It is thought that VP5* is involved in forming pore-like structures in the endosomes, eventually allowing endosomal escape [24].
In this process, the outer layer of the virion is detached, and the DLPs are released in the cytosol. These DLPs become transcriptionally active [25] entailing into the cytosol capped, non-polyadenylated (+)ssRNAs [26] for direct translation of the virus proteins required to i) block the innate immune response of the host (NSP1, VP3), ii) shut-off host translation (NSP3), and iii) build viroplasms (NSP5, NSP2 and VP2) [27,28,29,30].
Specific RV proteins accumulate within specialized cytosolic inclusions called viroplasms, where the RV genome and DLPs come together. Subsequently, DLPs exit the viroplasms via a not fully understood pathway. The current mechanism involves the association of VP6 with NSP4 embedded in the membrane of the endoplasmic reticulum (ER) [31,32]. Simultaneously, the spike protein VP4 localized between the viroplasm and the ER, associating with NSP4 [33]. These associations with NSP4 bring VP6 and VP4 in proximity, leading to the formation of a transiently enveloped DLP (eDLP) in the lumen of the ER [34]. In fact, eDLP reconstructions appear as DLPs with 60 trimeric VP4 spikes, which connect the particles to the transient envelope where VP7 and NSP4 are not discernible in these images [34].
2. Viroplasms
2.1. Spatial and Temporal Organization
Viroplasms are membrane-less globular electron-dense cytosolic inclusions (Figure 1a). These structures are responsible for virus genome replication and the generation of new rotavirus virions. So far, only RVA has been shown experimentally to induce viroplasm formation; their formation in other species remains to be elucidated. The viroplasms comprise NSP5, NSP4, NSP2, VP6, VP1, VP2, VP3, virus single and double-stranded RNA, and host components such as tubulin, perilipin, the host proteasome, and cellular chaperonins [25,39,40,41,42,43,44]. Finally, viroplasms are surrounded by the ER, enriched in VP4 and VP7 [39,45].
Viroplasms are detected as early as 2 hours post-infection (hpi), and their size steadily increases [46,47]. Interestingly, the number of viroplasms per cell decreases at 6 hpi, indicating coalescence between the structures and their dynamic nature [46,48]. Indeed, it has recently been shown that viroplasms are liquid-like inclusions [49]. Furthermore, during the infection, viroplasms move toward the perinuclear region of the cell in a process that is dependent on the microtubule (MT) cytoskeleton [48].
Remarkably, the expression of NSP5 in the presence of NSP2 or VP2 induces the formation of viroplasm-like structures (VLS) [50]. VLSs closely resemble the morphology of RV viroplasms (Figure 1b-d). Due to their simplicity, VLSs are a valuable model for studying viroplasms within a host using an in-vivo approach, as they share characteristics such as coalescence and perinuclear condensation [46,50].
2.2. Replication Steps within Viroplasms
The exact mechanism of genome packaging and assembly of virions within the viroplasms is unclear. The current model assumes that pre-genomic (+)ssRNAs are organized sequence-specifically through the assistance of NSP2 [51] in the viroplasms and packaged in the assembling core while simultaneously being replicated into dsRNA [52]. The filled cores move towards the periphery of the viroplasms, which are rich in VP6 for converting the cores into DLPs. These DLPs would produce i) more (+)ssRNA [53] or ii) migrate to the ER to become mature TLPs [54].
2.2. NSP5
Inhibition of NSP5 expression by RNA interference completely abolishes viroplasm formation and synthesis of genomic dsRNA as well as progeny virus, revealing the essential role of NSP5 in viroplasm formation [55]. While the complete structure of NSP5 remains unknown, it has been shown to form dimers and oligomers [56,57]. NSP5 is a hyperphosphorylated protein in infected cells, achieved by phosphorylation by cellular kinases, such as CK1α, and regulated by autoregulation and interaction with NSP2 and VP2 [58,59,60,61,62,63]. The phosphorylation cascade is critically dependent on the presence of a serine at position 67 [58]. A recombinant RV (rRV) harboring a point mutation in NSP5 in serine 67 to alanine (S67A) shows impaired globular viroplasms. This observation suggests that hyperphosphorylation of NSP5 is crucial for viroplasm morphology [64]. Additionally, the study highlights the significance of the NSP5 tail region in the phosphorylation cascade and viroplasm formation [59,64]. Despite the initial notion that autokinase activity could be described, no kinase activity could be attributed to NSP5, which, in addition to being the primary driver of viroplasm formation, displays ATPase activity [57,59,65].
2.2. NSP2
Another critical protein in viroplasm formation is NSP2. Similarly to NSP5, inhibition of NSP2 expression also leads to impairment of viroplasm formation [66]. NSP2 self-assembles into donut-shaped octamers, as denoted by crystallographic analysis for species A, B, and C [67,68,69,70]. These multimers can interact with the RNA-dependent RNA polymerase VP1 and viral RNA [71,72]. Furthermore, it has been demonstrated that NSP2 is an RNA chaperone, capable of binding to RNA transcripts and consequently controlling their interaction and unfolding [73]. NSP2 has been linked to several enzymatic activities, among them a nucleoside diphosphate (NDP) kinase-like activity [74], RNA-helix-destabilizing activities [74], and nucleoside triphosphatase (NTPase) activity [67]. NSP2 plays a direct role in viroplasm coalescence events [46]. Notably, the phosphorylation of NSP2 has also been implicated in viroplasm formation, as evidenced by the delayed formation of viroplasms observed in a rRV harboring an NSP2 S313D phosphomimetic mutant [75]. Additionally, studies using a mutant NSP2 harboring a lysine-to-glutamic acid change in the C-terminal region revealed the importance of a flexible tail in viroplasm biogenesis and coalescence properties [76].
2.5. VP2
An often-overlooked protein in the context of viroplasms is VP2. Silencing of VP2 expression in infected cells reduces the number of viroplasms per cell [77]. VP2, primarily studied as the main structural core protein, is also an inducer of VLS formation when co-expressed with NSP5 [40]. It has been shown that VLS formation is critically dependent on the presence of the three amino acids, L124, V865, and I878, residues highly conserved in VP2 of RV species A-H[78]. Previous studies have demonstrated that NSP2 [60] and VP2 [40,78] trigger the hyperphosphorylation cascade of NSP5. Additionally, VP2 has been implicated in modifying viroplasm perinuclear localization [48].
3. Host Cell Cytoskeleton
3.1. Microtubules
MTs are a significant component of the cytoskeletal network in eukaryotic cells, forming a dynamic network of polymeric filaments distributed throughout the cytoplasm. MTs play pivotal roles in numerous cellular processes, such as cell division, intracellular transport, motility, and organelle positioning. MTs are hollowed-out tubes formed from α-tubulin and β-tubulin (αβ-tubulin) heterodimers that are polarized and typically oriented toward the cell periphery [79]. The polarity, a crucial requirement for MT function, results from the head-to-tail polymerization of tubulin dimers with α-tubulin at the minus end and β-tubulin at the plus end [80]. Notably, individual filaments can reach up to 5000 µm persistence length in vitro, much longer than actin filaments, which can only reach persistence lengths of 15 – 20 µm [81].
An exciting feature of tubulins is their ability to undergo various reversible post-translational modifications (PTMs), such as acetylation, phosphorylation, poly-glycylation, poly-glutamylation, (de)tyrosination, and palmitoylation [82,83]. Most PTMs occur in the carboxy-terminal tails of tubulin, with the notable exception of acetylation [83]. Acetylation mainly occurs after the assembly of MTs and is associated with stabilizing the MT structure [83]. In addition, acetylation can improve the binding and transport of molecular motors, such as kinesin-1 or dynein [84,85]. Another way to regulate MT functions is by non-motor MT-associated proteins (MAPs), classified as MT-stabilizers, destabilizers, or plus-end tracking proteins [86,87]. MAPs also play a major role in MT bundling, a process that further regulates the stability of MT filaments [88,89].
3.1. MT-Dependent Molecular Motors
Two main classes of molecular motors specialize in transport along the MT network, corresponding to kinesin and dynein motors. Kinesin motors move towards the MT plus-end in what is known as anterograde transport. The diverse cargoes can either associate directly with the heavy chain or bind to specific regions in the C-terminus of the light chain [95]. In contrast, the molecular motor dynein moves towards the MT minus end, performing retrograde transport [96]. The cargo can bind to dynein in numerous ways, allowing for a wide range of client proteins [97,98]. Viruses, as cargoes, exploit cytoplasmic dynein to facilitate their motion within the cell [95].
3.1. Actin
Actin is the most abundant protein in many eukaryotic cells. Accordingly, several viruses subvert the actin cytoskeleton for long-range dissemination and transport [99].
Actin is expressed as a globular monomer known as G-actin [100]. When it polymerizes, it forms F-actin, filamentous structures that can form spontaneously in physiological conditions. Actin fibers play a fundamental role in many cellular processes, including motility, morphogenesis, cytokinesis, or endocytosis [101]. Actin- bundling-proteins can crosslink actin filaments into actin bundles, which are the main components of the actin network [102]. When smaller filaments are organized into microvilli in the plasma membrane protrusions and tightly packed into arrays, the filaments are referred to as brush borders [103]. Within the cells, the force of actin is produced by myosin molecular motors that move along the long actin domains, referred to as stress fibers [101,104]. These stress fibers are often anchored to focal adhesions corresponding to complex structures responsible for crucial scaffolding interactions with actin [105].
3.4. Actin-Dependent Molecular Motors
Over forty classes of myosins are expressed in eukaryotes, divided into muscle and non-muscle myosins [106]. Known as "conventional myosin," non-muscle myosin-2 (NM2) is present in almost every cell type, existing in three variants [107]. Therefore, it is not surprising that NM2 has been shown to play a role in the life cycle of numerous viruses [106].
3.5. Intermediate Filaments
The intermediate filaments (IFs) are the third component of the eukaryotic cytoskeletal network and are less studied than MTs and actin [108]. One reason is that the IFs are polymers of two, three, or more different proteins. These proteins include, among others, keratins, vimentin, lamins, and nestin, which form six subtypes of filaments [108]. Vimentin and nestin play a role in cell migration, but other proteins have diverse functions depending on the cell context [109]. Interestingly, IFs are formed in the cytoplasm and the nucleus [110]. So far, no motor proteins have been identified moving along IFs.
3.6. Viroplasm Interaction with Host Components
Aside from the viral components, viroplasms interact with many cellular components, including lipid droplets, proteins, and host nucleic acids. In this context, viroplasms are found to recruit components of lipid droplets (LDs) during the replication cycle [111]. LDs are spherical organelles that play a significant role in lipid homeostasis and contain mostly perilipins [112]. Associations with LDs appear required to form viroplasms and infectious virus progeny by serving as a scaffold for viroplasm assembly and allowing the association between viroplasms and ER membranes [41,111,113].
However, the viroplasms are also found to interact with many elements of the host cell cytoskeleton. All three primary cytoskeletal components (actin, MTs, and IFs) are restructured during RV infection. The formation of viroplasms relies on several of these reorganizations [114,115,116,117,118,119].
The reorganization of the MT-cytoskeleton has been shown to directly influence the coalescence and localization of viroplasms [48], which seems a trait common among many viruses inducing the formation of membrane-less replication compartments, such as birnaviruses, reoviruses, or African swine fever viruses [92,120,121,122,123]. In this sense, MT depolymerization drugs harm both perinuclear condensation and coalescence of the viroplasms. On the other hand, MT stabilizing drugs, such as taxol, showed no effect. In fact (Fig 2 a and b), RV infection increases stabilized MTs, as denoted by the rise of acetylated tubulin in viroplasms [48]. Collectively, RV can subvert the cytoskeleton to assemble and maintain viroplasms.
Indeed, RV NSP2 and NSP5 have been implicated in directly interacting with tubulin in co-immunoprecipitation assays followed by western blot or mass spectrometry. However, while the interaction of NSP2 with tubulin appears very stable, the interaction with NSP5 is not consistent [48,124,125,126]. NSP5 has been pulled down with tubulin as a contaminant in RV-infected cells due to its ability to bind to NSP2 [46,56]. It seems that NSP5 and tubulin compete for binding to the same positively charged grooves on the NSP2 octamer [125]. Interestingly, despite significant MT reorganization induced by NSP2 transfection, this study does not observe considerable colocalization of NSP2 and tubulin in NSP2-transfected cells. [125]. Furthermore, NSP2 exhibits a robust binding to non-acetylated tubulin compared to acetylated tubulin. [124]. Still, acetylated tubulin seems to accumulate in mature viroplasms [48].
A newly identified variant of NSP2 displays varying interactions with NSP5 and acetylated tubulin, depending on the phosphorylation status of NSP2 [124]. These two NSP2 conformations have been distinguished using two different monoclonal antibodies targeting different regions of NSP2. One conformation corresponds to viroplasmic NSP2 (vNSP2), which localizes in viroplasms. The second conformation is a cytosolic dispersed pool of NSP2 (dNSP2), which is phosphorylated at its C-terminus. Additionally, dNSP2 is weakly colocalizing with NSP5 and vNSP2 in viroplasms. Interestingly, dNSP2 resulted in a precursor of vNSP2 and can bind when phosphorylated to acetylated tubulin and hypophosphorylated NSP5. On the other hand, vNSP2 interacts with phosphorylated NSP5 and only weakly with tubulin [124]. This outcome suggests a mechanism of viroplasm formation and assembly coordinated by phosphorylation and tubulin acetylation. In this model, dNSP2 could bind to hypophosphorylated NSP5, triggering NSP5 phosphorylation at Ser67, leading to a nucleation event and viroplasm formation. These events concomitantly initiate the reorganization of the MT network to induce favorable conditions. Following this model, the destabilization of MTs during the early stages of infection hinders the coalescence of viroplasms [48].
Interestingly, the interaction with the MT network is not only based on NSP2-tubulin associations. In experiments using VLSs induced by co-expression of NSP5 with either NSP2 or VP2 and treated with an MT-destabilizing drug, it was shown that NSP2 confers the coalescence properties while VP2 mediates the perinuclear condensation properties. Additional research evidence that transfected NSP4 also binds and reorganizes the MT network [127,128,129]. Overall, the interaction of NSP5 and NSP2 with tubulin and their phosphorylation-dependent effects on viroplasm formation remain to be fully discovered.
Some studies point to the involvement of dynein-mediated transport in the coalescence of viroplasms [130]. NSP2 can interact with the dynein intermediate chain (DIC), mediating the ability of the viroplasm to coalesce. These findings resemble measles virus replication compartments, whose liquid-liquid phase separated replication organelles depend on dynein-mediated transport to form large inclusion and viral replication [131]--suggesting a conserved reliance on dynein-mediated transport among diverse viruses to organize replication structures. In addition, it has been shown that viroplasms can no longer coalesce or move to the perinuclear region when the molecular motor Eg5 of the kinesin-5 family is inhibited [48]. So far, however, no direct interaction partner has been identified, as VLS properties seem to be independent of the Eg5 function, regardless of VLS induction by NSP2 or VP2 [48]. Moreover, RV infection halts the host cell cycle in the S/G2 phase [132], a stage that correlates with a stabilized MT network [133]. The RV-induced cell cycle arrest relies on the kinesin motor Eg5 and the actin and MT networks. This connection underscores the significance of a stabilized MT network for viroplasm formation, linking it with the cell cycle arrest and, consequently, RV replication [132].
The actin cytoskeleton plays an additional important role in viroplasm dynamic and formation. In this context, actin has mainly been found to interact with VP4, but NSP4 has likewise been shown to induce actin remodeling [134,135,136,137]. VP4 is predominantly known as a structural spike protein but is also expressed as a soluble protein in the cytosol [134]. The interaction of VP4 and actin is well known [38,134,136]. It has been found that VP4 can induce actin-remodeling when expressed in the absence of other virus proteins [136]. Thus, VP4 has an actin-binding domain (ABD, amino acid region 713 to 773) at its C-terminus and a coiled-coil domain, allowing association to actin filaments. The VP4 ABD is buried in the assembled particle, pointing to the importance of soluble VP4 in the cytoplasm [134]. The use of a recombinant RV harboring a BAP tag in the VP8 region of VP4 (rRV/VP4-BAP) (Figure 2 c and d) demonstrated that cytosolic VP4 plays a critical role, either directly or indirectly, in interacting with actin filaments to facilitate viroplasm formation [137]. Similarly, as observed for Negri bodies in rabies virus (RABV)-infected cells [138], the treatment with cytochalasin D leads to a reduced number of viroplasms in RV-infected cells.
Additional studies proved that VP4 associates with MTs, potentially in an early step of virus release [139]. Studies have also shown that VP4 colocalizes with β-tubulin in both RV-infected and VP4-transfected cells, an interaction susceptible to disruption through MT depolymerization [139]. Moreover, it has been hypothesized that VP4 is transported to the plasma membrane by MT molecular motors [139]. It is plausible that the VP4 intracellular transport is differentially regulated, depending on the specific component of the cytoskeleton. It is well known that various viruses, such as flaviviruses [140] or influenza viruses [141] shift from actin-mediated transport to MT-associated transport at different steps of their life cycle. Transportation along the actin cytoskeleton may direct VP4 towards viroplasms to facilitate viroplasm formation. This process might involve the regulation of actin filaments and stress fiber formation by VP4, which are necessary for initiating viroplasm assembly. In contrast, the MT network may transport VP4 away from viroplasms for incorporation in the plasma membrane in an alternative TLP assembly pathway [139]. The potential of VP4 to undergo differential transport opens new questions regarding the regulation of host cell factors.
Only sparse research is available on the interplay of RV infection and the intermediate filaments. Infection with RV induces substantial restructuring of vimentin in adherent kidney cells, whereas such reorganization is not observed in differentiated human intestinal epithelial cells. Conversely, differentiated human intestinal epithelial cells display rearrangement of other cytoskeletal elements, a phenomenon not observed in undifferentiated human intestinal epithelial cells. [116,142]. Further research on the role of intermediate filaments is needed, as this is a relatively unexplored area.
Despite significant progress in the research on the assembly and maintenance of viroplasms, there are still gaps in our understanding of the precise molecular mechanisms involved in their formation and organization, as well as the interplay between different cytoskeletal components and their regulatory mechanisms.
4. Viral Factories Interaction with Host Components of Other dsRNA Viruses
Studies on other dsRNA viruses apart from RV, such as reoviruses or bluetongue virus, have revealed similar interactions between their viral factories and the host cell's cytoskeleton. Notably, research on mammalian reovirus (MRV) viral factories indicates their reliance on the MT network for their perinuclear condensation, movement, and structural assembly [92], properties observable in RV viroplasm formation as well [48]. It was found that both filamentous and globular MRV viral factories need an intact MT network for proper function with dynein localizing in both viral factories [92]. Moreover, the MT network is essential for forming large globular perinuclear inclusions by MRV non-structural protein μNS as nocodazole treatment, a tubulin depolymerizing agent, was shown to disperse the filamentous viral factories into smaller inclusions [143]. These results appear consistent with studies on RV showing inhibition of perinuclear condensation and coalescence upon treatment with nocodazole, suggesting similarities in the interaction of viroplasms with the host cytoskeleton in other dsRNA viruses [48].
Further, the interaction between reovirus core protein μ2 and MTs stabilized by bundling and hyperacetylation of α-tubulin determined the filamentous shape of reovirus inclusion bodies, highlighting the dependency of a stabilized MT network for the distribution of MRV viral factories in cells [121,144]. The direct association between MRV and spindle tubules observed in L2 cells could explain the aggregates of the virus in extensive perinuclear inclusions, although this association is not necessary for viral replication [145]. Furthermore, studies on both RV and MRV have shown that their infection disrupts and reorganizes vimentin filaments without affecting MTs or microfilament bundles [116,146] – Likewise, bluetongue virus associates with the cytoskeleton. Linear arrays of virus particles around viral inclusion bodies were found to be formed upon treatment with a chemical compound (colchicine), leading to aggregation of the vimentin filament network in the perinuclear region, suggesting an association of the viral inclusion bodies with the intermediate filaments [147].
5. Concluding Remarks
This review delineates the crucial interactions between RV proteins and the cellular cytoskeleton. It has become clear that phosphorylation, particularly the sequence of phosphorylation events, and other PTMs play a critical role in regulating the interaction between RV proteins and the cytoskeleton, particularly between NSP5, NSP2, and tubulin. Additionally, the new role of VP4 in regulating viroplasm formation, through its interaction with actin filaments as previously described, underscores the multifunctionality of RV proteins. This highlights the significance of host cell factors on the dynamics of viroplasms and virus replication.
In conclusion, this review underlines promising research areas and aims to enrich the ongoing discussion surrounding viroplasm assembly and maintenance. Addressing these unresolved questions and conducting further studies in these areas will deepen our comprehension of the complex interplay between RV and the host cell cytoskeleton, potentially leading to the development of novel therapeutic strategies for combating RV infections.
Author Contributions
Conceptualization, JV, ML, and CE; methodology, JV and ML; software, JV, ML, and CE; resources, JV, ML, and CE; data curation, JV, ML, and CE; writing—original draft preparation, JV, ML, and CE.; writing—review and editing, JV, ML, and CE; visualization, JV, ML, and CE; supervision, CE.; project administration, CE.; funding acquisition: CE. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Zurich
Institutional Review Board Statement
Not applicable
Acknowledgments
To permit open access, the authors have applied for a CC BY public copyright license for any version of this manuscript when accepted for publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Adams WR, Kraft LM: Epizootic Diarrhea of Infant Mice: Identification of the Etiologic Agent. Science 1963, 141:359-360. [CrossRef]
- Bishop RF, Davidson GP, Holmes IH, Ruck BJ: Virus particles in epithelial cells of duodenal mucosa from children with acute non-bacterial gastroenteritis. The Lancet 1973, 302:1281-1283. [CrossRef]
- Crawford SE, Ramani S, Tate JE, Parashar UD, Svensson L, Hagbom M, Franco MA, Greenberg HB, O'Ryan M, Kang G, et al: Rotavirus infection. Nature Reviews Disease Primers 2017, 3:1-16.
- Tate JE, Burton AH, Boschi-Pinto C, Steele AD, Duque J, Parashar UD: 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: a systematic review and meta-analysis. The Lancet Infectious Diseases 2012, 12:136-141. [CrossRef]
- Vesikari T, Karvonen A, Prymula R, Schuster V, Tejedor JC, Cohen R, Meurice F, Han HH, Damaso S, Bouckenooghe A: Efficacy of human rotavirus vaccine against rotavirus gastroenteritis during the first 2 years of life in European infants: randomised, double-blind controlled study. The Lancet 2007, 370:1757-1763.
- Hallowell BD, Tate J, Parashar U: An overview of rotavirus vaccination programs in developing countries. Expert Review of Vaccines 2020, 19:529-537. [CrossRef]
- Fauquet CM: Taxonomy, Classification and Nomenclature of Viruses. Encyclopedia of Virology 2008.
- Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA: Virus taxonomy: VIIIth report of the International Committee on Taxonomy of Viruses. Academic Press; 2005.
- Lefkowitz EJ, Dempsey DM, Hendrickson RC, Orton RJ, Siddell SG, Smith DB: Virus taxonomy: the database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Research 2018, 46:D708-D717. [CrossRef]
- Matthijnssens J, Theuns S: Minutes of the 7th Rotavirus Classification Working Group (RCWG) Meeting. In 12th International Double Stranded RNA Virus Symposium; Goa Marriott Beach Resort & Spa, Goa, India. 2015.
- Johne R, Tausch SH, Ulrich RG, Schilling-Loeffler K: Genome analysis of the novel putative rotavirus species K. Virus Research 2023, 334:199171. [CrossRef]
- Johne R, Schilling-Loeffler K, Ulrich RG, Tausch SH: Whole Genome Sequence Analysis of a Prototype Strain of the Novel Putative Rotavirus Species L. In Viruses, vol. 142022. [CrossRef]
- Asensio-Cob D, Rodríguez JM, Luque D: Rotavirus Particle Disassembly and Assembly In Vivo and In Vitro. Viruses 2023, 15. [CrossRef]
- Prasad BVV, Wang GJ, Clerx JPM, Chiu W: Three-dimensional structure of rotavirus. Journal of Molecular Biology 1988, 199:269-275. [CrossRef]
- Pesavento JB, Crawford SE, Estes MK, Venkataram Prasad BV: Rotavirus Proteins: Structure and Assembly. In Reoviruses: Entry, Assembly and Morphogenesis. Edited by Roy P. Berlin, Heidelberg: Springer Berlin Heidelberg; 2006: 189-219.
- Dormitzer PR, Sun ZY, Wagner G, Harrison SC: The rhesus rotavirus VP4 sialic acid binding domain has a galectin fold with a novel carbohydrate binding site. EMBO J 2002, 21:885-897. [CrossRef]
- Isa P, López S, Segovia L, Arias CF: Functional and structural analysis of the sialic acid-binding domain of rotaviruses. Journal of Virology 1997, 71:6749-6756. [CrossRef]
- Isa P, Arias CF, López S: Role of sialic acids in rotavirus infection. Glycoconjugate Journal 2006, 23:27-37. [CrossRef]
- Huang P, Xia M, Tan M, Zhong W, Wei C, Wang L, Morrow A, Jiang X: Spike Protein VP8* of Human Rotavirus Recognizes Histo-Blood Group Antigens in a Type-Specific Manner. Journal of Virology 2012, 86:4833-4843. [CrossRef]
- Gutiérrez M, Isa P, Sánchez-San Martin C, Pérez-Vargas J, Espinosa R, Arias CF, López S: Different Rotavirus Strains Enter MA104 Cells through Different Endocytic Pathways: the Role of Clathrin-Mediated Endocytosis. Journal of Virology 2010, 84:9161-9169. [CrossRef]
- Li B, Ding S, Feng N, Mooney N, Ooi YS, Ren L, Diep J, Kelly MR, Yasukawa LL, Patton JT, et al: Drebrin restricts rotavirus entry by inhibiting dynamin-mediated endocytosis. Proc Natl Acad Sci U S A 2017, 114:E3642-E3651.
- Arias CF, Silva-Ayala D, López S: Rotavirus Entry: a Deep Journey into the Cell with Several Exits. Journal of Virology 2015, 89:890-893. [CrossRef]
- Díaz-Salinas Marco A, Silva-Ayala D, López S, Arias Carlos F: Rotaviruses Reach Late Endosomes and Require the Cation-Dependent Mannose-6-Phosphate Receptor and the Activity of Cathepsin Proteases To Enter the Cell. Journal of Virology 2014, 88:4389-4402.
- Golantsova Nina E, Gorbunova Elena E, Mackow Erich R: Discrete Domains within the Rotavirus VP5* Direct Peripheral Membrane Association and Membrane Permeability. Journal of Virology 2004, 78:2037-2044.
- Patton JT, Silvestri LS, Tortorici MA, Carpio V-D, Taraporewala ZF: Rotavirus genome replication and morphogenesis: role of the viroplasm. Reoviruses: entry, assembly and morphogenesis 2006:169-187.
- Lawton JA, Estes MK, Prasad BVV: Three-dimensional visualization of mRNA release from actively transcribing rotavirus particles. Nature Structural Biology 1997, 4:118-121. [CrossRef]
- Petrie BL, Graham DY, Hanssen H, Estes MK: Localization of rotavirus antigens in infected cells by ultrastructural immunocytochemistry. Journal of General Virology 1982, 63:457-467. [CrossRef]
- Arnold Michelle M: The Rotavirus Interferon Antagonist NSP1: Many Targets, Many Questions. Journal of Virology 2016, 90:5212-5215.
- Padilla-Noriega L, Paniagua O, Guzmán-León S: Rotavirus protein NSP3 shuts off host cell protein synthesis. Virology 2002, 298:1-7. [CrossRef]
- Morelli M, Ogden KM, Patton JT: Silencing the alarms: Innate immune antagonism by rotavirus NSP1 and VP3. Virology 2015, 479-480:75-84. [CrossRef]
- Taylor JA, O'Brien JA, Lord VJ, Meyer JC, Bellamy AR: The RER-Localized Rotavirus Intracellular Receptor: A Truncated Purified Soluble Form Is Multivalent and Binds Virus Particles. Virology 1993, 194:807-814. [CrossRef]
- López T, Camacho M, Zayas M, Nájera R, Sánchez R, Arias Carlos F, López S: Silencing the Morphogenesis of Rotavirus. Journal of Virology 2005, 79:184-192. [CrossRef]
- Trask SD, McDonald SM, Patton JT: Structural insights into the coupling of virion assembly and rotavirus replication. Nat Rev Microbiol 2012, 10:165-177. [CrossRef]
- Shah PNM, Gilchrist JB, Forsberg BO, Burt A, Howe A, Mosalaganti S, Wan W, Radecke J, Chaban Y, Sutton G, et al: Characterization of the rotavirus assembly pathway in situ using cryoelectron tomography. Cell Host Microbe 2023, 31:604-615.e604.
- Musalem C, Espejo RT: Release of Progeny Virus from Cells Infected with Simian Rotavirus SA11. Journal of General Virology 1985, 66:2715-2724. [CrossRef]
- Cevallos Porta D, López S, Arias CF, Isa P: Polarized rotavirus entry and release from differentiated small intestinal cells. Virology 2016, 499:65-71. [CrossRef]
- Gardet A, Breton M, Trugnan G, Chwetzoff S: Role for actin in the polarized release of rotavirus. J Virol 2007, 81:4892-4894. [CrossRef]
- Trejo-Cerro Ó, Eichwald C, Schraner EM, Silva-Ayala D, López S, Arias CF: Actin-Dependent Nonlytic Rotavirus Exit and Infectious Virus Morphogenetic Pathway in Nonpolarized Cells. Journal of Virology 2018, 92. [CrossRef]
- Altenburg BC, Graham DY, Kolb Estes M: Ultrastructural study of rotavirus replication in cultured cells. Journal of General Virology 1980, 46:75-85. [CrossRef]
- Contin R, Arnoldi F, Campagna M, Burrone OR: Rotavirus NSP5 orchestrates recruitment of viroplasmic proteins. J Gen Virol 2010, 91:1782-1793. [CrossRef]
- Cheung W, Gill M, Esposito A, Kaminski Clemens F, Courousse N, Chwetzoff S, Trugnan G, Keshavan N, Lever A, Desselberger U: Rotaviruses Associate with Cellular Lipid Droplet Components To Replicate in Viroplasms, and Compounds Disrupting or Blocking Lipid Droplets Inhibit Viroplasm Formation and Viral Replication. Journal of Virology 2010, 84:6782-6798. [CrossRef]
- Contin R, Arnoldi F, Mano M, Burrone OR: Rotavirus replication requires a functional proteasome for effective assembly of viroplasms. J Virol 2011, 85:2781-2792. [CrossRef]
- Lopez T, Silva-Ayala D, Lopez S, Arias CF: Replication of the Rotavirus Genome Requires an Active Ubiquitin-Proteasome System. Journal of Virology 2011, 85:11964-11971. [CrossRef]
- Vetter J, Papa G, Tobler K, Rodriguez Javier M, Kley M, Myers M, Wiesendanger M, Schraner Elisabeth M, Luque D, Burrone Oscar R, et al: The recruitment of TRiC chaperonin in rotavirus viroplasms correlates with virus replication. mBio 2024, 0:e00499-00424.
- Gonzalez RA, Espinosa R, Romero P, Lopez S, Arias CF: Relative localization of viroplasmic and endoplasmic reticulum-resident rotavirus proteins in infected cells. Archives of virology 2000, 145:1963-1973. [CrossRef]
- Eichwald C, Rodriguez JF, Burrone OR: Characterization of rotavirus NSP2/NSP5 interactions and the dynamics of viroplasm formation. J Gen Virol 2004, 85:625-634.
- Carreño-Torres JJ, Gutiérrez M, Arias CF, López S, Isa P: Characterization of viroplasm formation during the early stages of rotavirus infection. Virol J 2010, 7:350. [CrossRef]
- Eichwald C, Arnoldi F, Laimbacher AS, Schraner EM, Fraefel C, Wild P, Burrone OR, Ackermann M: Rotavirus viroplasm fusion and perinuclear localization are dynamic processes requiring stabilized microtubules. PLoS One 2012, 7:e47947. [CrossRef]
- Geiger F, Acker J, Papa G, Wang X, Arter WE, Saar KL, Erkamp NA, Qi R, Bravo JPK, Strauss S, et al: Liquid–liquid phase separation underpins the formation of replication factories in rotaviruses. The EMBO Journal 2021, 40:e107711.
- Fabbretti E, Afrikanova I, Vascotto F, Burrone OR: Two non-structural rotavirus proteins, NSP2 and NSP5, form viroplasm-like structures in vivo. J Gen Virol 1999, 80 ( Pt 2):333-339.
- Borodavka A, Dykeman EC, Schrimpf W, Lamb DC: Protein-mediated RNA folding governs sequence-specific interactions between rotavirus genome segments. eLife 2017, 6:e27453.
- Patton JT, Jones MT, Kalbach AN, He YW, Xiaobo J: Rotavirus RNA polymerase requires the core shell protein to synthesize the double-stranded RNA genome. J Virol 1997, 71:9618-9626. [CrossRef]
- Periz J, Celma C, Jing B, Pinkney JN, Roy P, Kapanidis AN: Rotavirus mRNAS are released by transcript-specific channels in the double-layered viral capsid. Proc Natl Acad Sci U S A 2013, 110:12042-12047.
- Ruiz MC, Leon T, Diaz Y, Michelangeli F: Molecular Biology of Rotavirus Entry and Replication. TheScientificWorldJOURNAL 2009, 9:879851. [CrossRef]
- Campagna M, Eichwald C, Vascotto F, Burrone OR: RNA interference of rotavirus segment 11 mRNA reveals the essential role of NSP5 in the virus replicative cycle. J Gen Virol 2005, 86:1481-1487. [CrossRef]
- Martin D, Ouldali M, Ménétrey J, Poncet D: Structural organisation of the rotavirus nonstructural protein NSP5. J Mol Biol 2011, 413:209-221. [CrossRef]
- Martin D, Charpilienne A, Parent A, Boussac A, D'Autreaux B, Poupon J, Poncet D: The rotavirus nonstructural protein NSP5 coordinates a [2Fe-2S] iron-sulfur cluster that modulates interaction to RNA. FASEB J 2013, 27:1074-1083. [CrossRef]
- Eichwald C, Jacob G, Muszynski B, Allende JE, Burrone OR: Uncoupling substrate and activation functions of rotavirus NSP5: phosphorylation of Ser-67 by casein kinase 1 is essential for hyperphosphorylation. Proc Natl Acad Sci U S A 2004, 101:16304-16309. [CrossRef]
- Eichwald C, Vascotto F, Fabbretti E, Burrone OR: Rotavirus NSP5: mapping phosphorylation sites and kinase activation and viroplasm localization domains. J Virol 2002, 76:3461-3470. [CrossRef]
- Afrikanova I, Fabbretti E, Miozzo MC, Burrone OR: Rotavirus NSP5 phosphorylation is up-regulated by interaction with NSP2. J Gen Virol 1998, 79 ( Pt 11):2679-2686. [CrossRef]
- Campagna M, Budini M, Arnoldi F, Desselberger U, Allende JE, Burrone OR: Impaired hyperphosphorylation of rotavirus NSP5 in cells depleted of casein kinase 1alpha is associated with the formation of viroplasms with altered morphology and a moderate decrease in virus replication. J Gen Virol 2007, 88:2800-2810. [CrossRef]
- Torres-Vega MA, González RA, Duarte M, Poncet D, López S, Arias CF: The C-terminal domain of rotavirus NSP5 is essential for its multimerization, hyperphosphorylation and interaction with NSP6. J Gen Virol 2000, 81:821-830. [CrossRef]
- Arnoldi F, Campagna M, Eichwald C, Desselberger U, Burrone OR: Interaction of rotavirus polymerase VP1 with nonstructural protein NSP5 is stronger than that with NSP2. J Virol 2007, 81:2128-2137. [CrossRef]
- Papa G, Venditti L, Arnoldi F, Schraner EM, Potgieter C, Borodavka A, Eichwald C, Burrone OR: Recombinant rotaviruses rescued by reverse genetics reveal the role of NSP5 hyperphosphorylation in the assembly of viral factories. Journal of Virology 2020, 94:1-23. [CrossRef]
- Bar-Magen T, Spencer E, Patton JT: An ATPase activity associated with the rotavirus phosphoprotein NSP5. Virology 2007, 369:389-399. [CrossRef]
- Silvestri LS, Taraporewala ZF, Patton JT: Rotavirus replication: plus-sense templates for double-stranded RNA synthesis are made in viroplasms. J Virol 2004, 78:7763-7774. [CrossRef]
- Taraporewala Z, Chen D, Patton JT: Multimers formed by the rotavirus nonstructural protein NSP2 bind to RNA and have nucleoside triphosphatase activity. J Virol 1999, 73:9934-9943. [CrossRef]
- Jayaram H, Taraporewala Z, Patton JT, Prasad BVV: Rotavirus protein involved in genome replication and packaging exhibits a HIT-like fold. Nature 2002, 417:311-315. [CrossRef]
- Taraporewala Zenobia F, Jiang X, Vasquez-Del Carpio R, Jayaram H, Prasad BVV, Patton John T: Structure-Function Analysis of Rotavirus NSP2 Octamer by Using a Novel Complementation System. Journal of Virology 2006, 80:7984-7994. [CrossRef]
- Chamera S, Wycisk K, Czarnocki-Cieciura M, Nowotny M: Cryo-EM structure of rotavirus B NSP2 reveals its unique tertiary architecture. Journal of Virology 2024, 0:e01660-01623. [CrossRef]
- Kattoura MD, Clapp LL, Patton JT: The rotavirus nonstructural protein, NS35, possesses RNA-binding activity in vitro and in vivo. Virology 1992, 191:698-708. [CrossRef]
- Kattoura MD, Chen X, Patton JT: The Rotavirus RNA-Binding Protein NS35 (NSP2) Forms 10S Multimers and Interacts with the Viral RNA Polymerase. Virology 1994, 202:803-813. [CrossRef]
- Bravo JPK, Bartnik K, Venditti L, Acker J, Gail EH, Colyer A, Davidovich C, Lamb DC, Tuma R, Calabrese AN, Borodavka A: Structural basis of rotavirus RNA chaperone displacement and RNA annealing. Proc Natl Acad Sci U S A 2021, 118. [CrossRef]
- Kumar M, Jayaram H, Vasquez-Del Carpio R, Jiang X, Taraporewala Zenobia F, Jacobson Raymond H, Patton John T, Prasad BVV: Crystallographic and Biochemical Analysis of Rotavirus NSP2 with Nucleotides Reveals a Nucleoside Diphosphate Kinase-Like Activity. Journal of Virology 2007, 81:12272-12284.
- Criglar Jeanette M, Crawford Sue E, Zhao B, Smith Hunter G, Stossi F, Estes Mary K: A Genetically Engineered Rotavirus NSP2 Phosphorylation Mutant Impaired in Viroplasm Formation and Replication Shows an Early Interaction between vNSP2 and Cellular Lipid Droplets. Journal of Virology 2020, 94:10.1128/jvi.00972-00920.
- Nichols SL, Nilsson EM, Brown-Harding H, LaConte LEW, Acker J, Borodavka A, McDonald Esstman S: Flexibility of the Rotavirus NSP2 C-Terminal Region Supports Factory Formation via Liquid-Liquid Phase Separation. J Virol 2023, 97:e0003923. [CrossRef]
- Montero H, Rojas M, Arias Carlos F, López S: Rotavirus Infection Induces the Phosphorylation of eIF2α but Prevents the Formation of Stress Granules. Journal of Virology 2008, 82:1496-1504. [CrossRef]
- Buttafuoco A, Michaelsen K, Tobler K, Ackermann M, Fraefel C, Eichwald C: Conserved Rotavirus NSP5 and VP2 Domains Interact and Affect Viroplasm. J Virol 2020, 94. [CrossRef]
- Akhmanova A, Steinmetz MO: Microtubule minus-end regulation at a glance. Journal of Cell Science 2019, 132:jcs227850. [CrossRef]
- Mitchison TJ: Localization of an Exchangeable GTP Binding Site at the Plus End of Microtubules. Science 1993, 261:1044-1047.
- Gittes F, Mickey B, Nettleton J, Howard J: Flexural rigidity of microtubules and actin filaments measured from thermal fluctuations in shape. Journal of Cell Biology 1993, 120:923-934. [CrossRef]
- Janke C, Bulinski JC: Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat Rev Mol Cell Biol 2011, 12:773-786. [CrossRef]
- Westermann S, Weber K: Post-translational modifications regulate microtubule function. Nature Reviews Molecular Cell Biology 2003, 4:938-948. [CrossRef]
- Reed NA, Cai D, Blasius TL, Jih GT, Meyhofer E, Gaertig J, Verhey KJ: Microtubule Acetylation Promotes Kinesin-1 Binding and Transport. Current Biology 2006, 16:2166-2172. [CrossRef]
- Alper Joshua D, Decker F, Agana B, Howard J: The Motility of Axonemal Dynein Is Regulated by the Tubulin Code. Biophysical Journal 2014, 107:2872-2880. [CrossRef]
- Goodson HV, Jonasson EM: Microtubules and Microtubule-Associated Proteins. Cold Spring Harb Perspect Biol 2018, 10.
- Bodakuntla S, Jijumon AS, Villablanca C, Gonzalez-Billault C, Janke C: Microtubule-Associated Proteins: Structuring the Cytoskeleton. Trends in Cell Biology 2019, 29:804-819.
- Brandt R, Lee G: Orientation, assembly, and stability of microtubule bundles induced by a fragment of tau protein. Cell Motility 1994, 28:143-154. [CrossRef]
- Walczak CE, Shaw SL: A MAP for Bundling Microtubules. Cell 2010, 142:364-367. [CrossRef]
- Greber UF, Way M: A Superhighway to Virus Infection. Cell 2006, 124:741-754. [CrossRef]
- Walsh D, Naghavi MH: Exploitation of Cytoskeletal Networks during Early Viral Infection. Trends in Microbiology 2019, 27:39-50. [CrossRef]
- Eichwald C, Ackermann M, Nibert ML: The dynamics of both filamentous and globular mammalian reovirus viral factories rely on the microtubule network. Virology 2018, 518:77-86. [CrossRef]
- Iwamoto M, Cai D, Sugiyama M, Suzuki R, Aizaki H, Ryo A, Ohtani N, Tanaka Y, Mizokami M, Wakita T, et al: Functional association of cellular microtubules with viral capsid assembly supports efficient hepatitis B virus replication. Scientific Reports 2017, 7:10620.
- Jouvenet N, Monaghan P, Way M, Wileman T: Transport of African Swine Fever Virus from Assembly Sites to the Plasma Membrane Is Dependent on Microtubules and Conventional Kinesin. Journal of Virology 2004, 78:7990-8001. [CrossRef]
- Dodding MP, Way M: Coupling viruses to dynein and kinesin-1. The EMBO Journal 2011, 30:3527-3539. [CrossRef]
- Roberts AJ: Emerging mechanisms of dynein transport in the cytoplasm versus the cilium. Biochemical Society Transactions 2018, 46:967-982. [CrossRef]
- Roberts AJ, Kon T, Knight PJ, Sutoh K, Burgess SA: Functions and mechanics of dynein motor proteins. Nature reviews Molecular cell biology 2013, 14:713-713. [CrossRef]
- King SM: The dynein microtubule motor. In Biochimica et Biophysica Acta - Molecular Cell Research, vol. 1496. pp. 60-75: Elsevier; 2000: 60-75.
- Taylor MP, Koyuncu OO, Enquist LW: Subversion of the actin cytoskeleton during viral infection. Nature Reviews Microbiology 2011 9:6 2011, 9:427-439. [CrossRef]
- Dominguez R, Holmes KC: Actin Structure and Function. Annual Review of Biophysics 2011, 40:169-186.
- Tojkander S, Gateva G, Lappalainen P: Actin stress fibers – assembly, dynamics and biological roles. Journal of Cell Science 2012, 125:1855-1864.
- Rajan S, Kudryashov DS, Reisler E: Actin Bundles Dynamics and Architecture. In Biomolecules, vol. 132023. [CrossRef]
- Meenderink LM, Gaeta IM, Postema MM, Cencer CS, Chinowsky CR, Krystofiak ES, Millis BA, Tyska MJ: Actin dynamics drive microvillar motility and clustering during brush border assembly. Developmental cell 2019, 50:545-556. e544.
- Lehtimäki JI, Rajakylä EK, Tojkander S, Lappalainen P: Generation of stress fibers through myosin-driven reorganization of the actin cortex. eLife 2021, 10:e60710.
- Geiger B, Spatz JP, Bershadsky AD: Environmental sensing through focal adhesions. Nature Reviews Molecular Cell Biology 2009, 10:21-33.
- Matozo T, Kogachi L, de Alencar BC: Myosin motors on the pathway of viral infections. Cytoskeleton 2022, 79:41-63.
- Sellers JR, Heissler SM: Nonmuscle myosin-2 isoforms. Current Biology 2019, 29:R275-R278. [CrossRef]
- Leduc C, Etienne-Manneville S: Intermediate filaments in cell migration and invasion: the unusual suspects. Current Opinion in Cell Biology 2015, 32:102-112. [CrossRef]
- Eriksson JE, Dechat T, Grin B, Helfand B, Mendez M, Pallari H-M, Goldman RD: Introducing intermediate filaments: from discovery to disease. The Journal of Clinical Investigation 2009, 119:1763-1771.
- Herrmann H, Bär H, Kreplak L, Strelkov SV, Aebi U: Intermediate filaments: from cell architecture to nanomechanics. Nature Reviews Molecular Cell Biology 2007, 8:562-573.
- Desselberger U: The significance of lipid droplets for the replication of rotaviruses and other RNA viruses. J Biol Todays World 2020, 9:001-003.
- Murphy S, Martin S, Parton RG: Lipid droplet-organelle interactions; sharing the fats. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2009, 1791:441-447.
- Martínez JL, Eichwald C, Schraner EM, López S, Arias CF: Lipid metabolism is involved in the association of rotavirus viroplasms with endoplasmic reticulum membranes. Virology 2022, 569:29-36. [CrossRef]
- Mattion NM, Cohen J, Aponte C, Estes MK: Characterization of an oligomerization domain and RNA-binding properties on rotavirus nonstructural protein NS34. Virology 1992, 190:68-83. [CrossRef]
- Hua J, Patton JT: The Carboxyl-Half of the Rotavirus Nonstructural Protein NS53 (NSP1) Is Not Required for Virus Replication. Virology 1994, 198:567-576. [CrossRef]
- Weclewicz K, Kristensson K, Svensson L: Rotavirus causes selective vimentin reorganization in monkey kidney CV-1 cells. Journal of General Virology 1994, 75:3267-3271. [CrossRef]
- Brunet J-P, Cotte-Laffitte J, Linxe C, Quero A-M, Géniteau-Legendre M, Servin A: Rotavirus infection induces an increase in intracellular calcium concentration in human intestinal epithelial cells: role in microvillar actin alteration. Journal of virology 2000, 74:2323-2332. [CrossRef]
- Jourdan N, Brunet Jean P, Sapin C, Blais A, Cotte-Laffitte J, Forestier F, Quero A-M, Trugnan G, Servin Alain L: Rotavirus Infection Reduces Sucrase-Isomaltase Expression in Human Intestinal Epithelial Cells by Perturbing Protein Targeting and Organization of Microvillar Cytoskeleton. Journal of Virology 1998, 72:7228-7236. [CrossRef]
- Weclewicz K, Svensson L, Billger M, Holmberg K, Wallin M, Kristensson K: Microtubule-associated protein 2 appears in axons of cultured dorsal root ganglia and spinal cord neurons after rotavirus infection. Journal of Neuroscience Research 1993, 36:173-182. [CrossRef]
- Campbell Elle A, Reddy Vishwanatha RAP, Gray Alice G, Wells J, Simpson J, Skinner Michael A, Hawes Philippa C, Broadbent Andrew J: Discrete Virus Factories Form in the Cytoplasm of Cells Coinfected with Two Replication-Competent Tagged Reporter Birnaviruses That Subsequently Coalesce over Time. Journal of Virology 2020, 94:10.1128/jvi.02107-02119.
- Parker John SL, Broering Teresa J, Kim J, Higgins Darren E, Nibert Max L: Reovirus Core Protein μ2 Determines the Filamentous Morphology of Viral Inclusion Bodies by Interacting with and Stabilizing Microtubules. Journal of Virology 2002, 76:4483-4496.
- Heath CM, Windsor M, Wileman T: Aggresomes Resemble Sites Specialized for Virus Assembly. Journal of Cell Biology 2001, 153:449-456. [CrossRef]
- Lahaye X, Vidy A, Pomier C, Obiang L, Harper F, Gaudin Y, Blondel D: Functional Characterization of Negri Bodies (NBs) in Rabies Virus-Infected Cells: Evidence that NBs Are Sites of Viral Transcription and Replication. Journal of Virology 2009, 83:7948-7958. [CrossRef]
- Criglar Jeanette M, Hu L, Crawford Sue E, Hyser Joseph M, Broughman James R, Prasad BVV, Estes Mary K: A Novel Form of Rotavirus NSP2 and Phosphorylation-Dependent NSP2-NSP5 Interactions Are Associated with Viroplasm Assembly. Journal of Virology 2014, 88:786-798.
- Martin D, Duarte M, Lepault J, Poncet D: Sequestration of free tubulin molecules by the viral protein NSP2 induces microtubule depolymerization during rotavirus infection. J Virol 2010, 84:2522-2532. [CrossRef]
- Dhillon P, Tandra Varsha N, Chorghade Sandip G, Namsa Nima D, Sahoo L, Rao CD: Cytoplasmic Relocalization and Colocalization with Viroplasms of Host Cell Proteins, and Their Role in Rotavirus Infection. Journal of Virology 2018, 92:10.1128/jvi.00612-00618.
- Zambrano JL, Sorondo O, Alcala A, Vizzi E, Diaz Y, Ruiz MC, Michelangeli F, Liprandi F, Ludert JE: Rotavirus infection of cells in culture induces activation of RhoA and changes in the actin and tubulin cytoskeleton. PLoS One 2012, 7:e47612. [CrossRef]
- Zhang M, Zeng Carl QY, Morris Andrew P, Estes Mary K: A Functional NSP4 Enterotoxin Peptide Secreted from Rotavirus-Infected Cells. Journal of Virology 2000, 74:11663-11670.
- Xu A, Bellamy AR, Taylor JA: Immobilization of the early secretory pathway by a virus glycoprotein that binds to microtubules. The EMBO Journal 2000, 19:6465-6474. [CrossRef]
- Jing Z, Shi H, Chen J, Shi D, Liu J, Guo L, Tian J, Wu Y, Dong H, Ji Z, et al: Rotavirus Viroplasm Biogenesis Involves Microtubule-Based Dynein Transport Mediated by an Interaction between NSP2 and Dynein Intermediate Chain. J Virol 2021, 95:e0124621. [CrossRef]
- Zhou Y, Su Justin M, Samuel Charles E, Ma D: Measles Virus Forms Inclusion Bodies with Properties of Liquid Organelles. Journal of Virology 2019, 93:10.1128/jvi.00948-00919.
- Glück S, Buttafuoco A, Meier AF, Arnoldi F, Vogt B, Schraner EM, Ackermann M, Eichwald C: Rotavirus replication is correlated with S/G2 interphase arrest of the host cell cycle. PLoS One 2017, 12:e0179607. [CrossRef]
- Akhmanova A, Steinmetz MO: Control of microtubule organization and dynamics: two ends in the limelight. Nature Reviews Molecular Cell Biology 2015, 16:711-726.
- Condemine W, Eguether T, Couroussé N, Etchebest C, Gardet A, Trugnan G, Chwetzoff S: The C Terminus of Rotavirus VP4 Protein Contains an Actin Binding Domain Which Requires Cooperation with the Coiled-Coil Domain for Actin Remodeling. Journal of Virology 2018, 93. [CrossRef]
- Berkova Z, Crawford SE, Blutt SE, Morris AP, Estes MK: Expression of rotavirus NSP4 alters the actin network organization through the actin remodeling protein cofilin. Journal of virology 2007, 81:3545-3553. [CrossRef]
- Gardet A, Breton M, Fontanges P, Trugnan G, Chwetzoff S: Rotavirus spike protein VP4 binds to and remodels actin bundles of the epithelial brush border into actin bodies. J Virol 2006, 80:3947-3956. [CrossRef]
- Vetter J, Papa G, Seyffert M, Gunasekera K, De Lorenzo G, Wiesendanger M, Reymond JL, Fraefel C, Burrone OR, Eichwald C: Rotavirus Spike Protein VP4 Mediates Viroplasm Assembly by Association to Actin Filaments. J Virol 2022:e0107422. [CrossRef]
- Nikolic J, Le Bars R, Lama Z, Scrima N, Lagaudrière-Gesbert C, Gaudin Y, Blondel D: Negri bodies are viral factories with properties of liquid organelles. Nature Communications 2017, 8:1-12. [CrossRef]
- Nejmeddine M, Trugnan G, Sapin C, Kohli E, Svensson L, Lopez S, Cohen J: Rotavirus spike protein VP4 is present at the plasma membrane and is associated with microtubules in infected cells. J Virol 2000, 74:3313-3320. [CrossRef]
- Zhang Y, Gao W, Li J, Wu W, Jiu Y: The Role of Host Cytoskeleton in Flavivirus Infection. Virologica Sinica 2019, 34:30-41. [CrossRef]
- Zhang L-J, Xia L, Liu S-L, Sun E-Z, Wu Q-M, Wen L, Zhang Z-L, Pang D-W: A “Driver Switchover” Mechanism of Influenza Virus Transport from Microfilaments to Microtubules. ACS Nano 2018, 12:474-484. [CrossRef]
- Brunet J-P, Jourdan N, Cotte-Laffitte J, Linxe C, Géniteau-Legendre M, Servin A, Quéro A-M: Rotavirus Infection Induces Cytoskeleton Disorganization in Human Intestinal Epithelial Cells: Implication of an Increase in Intracellular Calcium Concentration. Journal of Virology 2000, 74:10801-10806. [CrossRef]
- Broering TJ, Parker JSL, Joyce PL, Kim J, Nibert ML: Mammalian Reovirus Nonstructural Protein μNS Forms Large Inclusions and Colocalizes with Reovirus Microtubule-Associated Protein μ2 in Transfected Cells. Journal of Virology 2002, 76:8285-8297. [CrossRef]
- Bussiere LD, Choudhury P, Bellaire B, Miller CL: Characterization of a Replicating Mammalian Orthoreovirus with Tetracysteine-Tagged μNS for Live-Cell Visualization of Viral Factories. J Virol 2017, 91. [CrossRef]
- Dales S: Association Between The Spindle Apparatus and Reovirus. Proceedings of the National Academy of Sciences of the United States of America 1963, 50:268-275.
- Sharpe AH, Chen LB, Fields BN: The interaction of mammalian reoviruses with the cytoskeleton of monkey kidney CV-1 cells. Virology 1982, 120:399-411. [CrossRef]
- Eaton BT, Hyatt AD: Association of Bluetongue Virus with the Cytoskeleton. Volume 151989: 233-273.
Figure 1.
comparison of viroplasms and VLSs. (a) Electron micrograph of viroplasm (V) at 6 hpi. The electron dense viroplasm structure is surrounded of the reticulum endoplasmic membrane filled with TLPs at diverse stage of maturation. Immunofluorescence of images of viroplasm (b), VLS(NSP2)i (c) and VLS (VP2)i (d) immunostained with anti-NSP5 (green). Nuclei are stained with DAPI (blue). Scale bar is 10 µm.
Figure 1.
comparison of viroplasms and VLSs. (a) Electron micrograph of viroplasm (V) at 6 hpi. The electron dense viroplasm structure is surrounded of the reticulum endoplasmic membrane filled with TLPs at diverse stage of maturation. Immunofluorescence of images of viroplasm (b), VLS(NSP2)i (c) and VLS (VP2)i (d) immunostained with anti-NSP5 (green). Nuclei are stained with DAPI (blue). Scale bar is 10 µm.
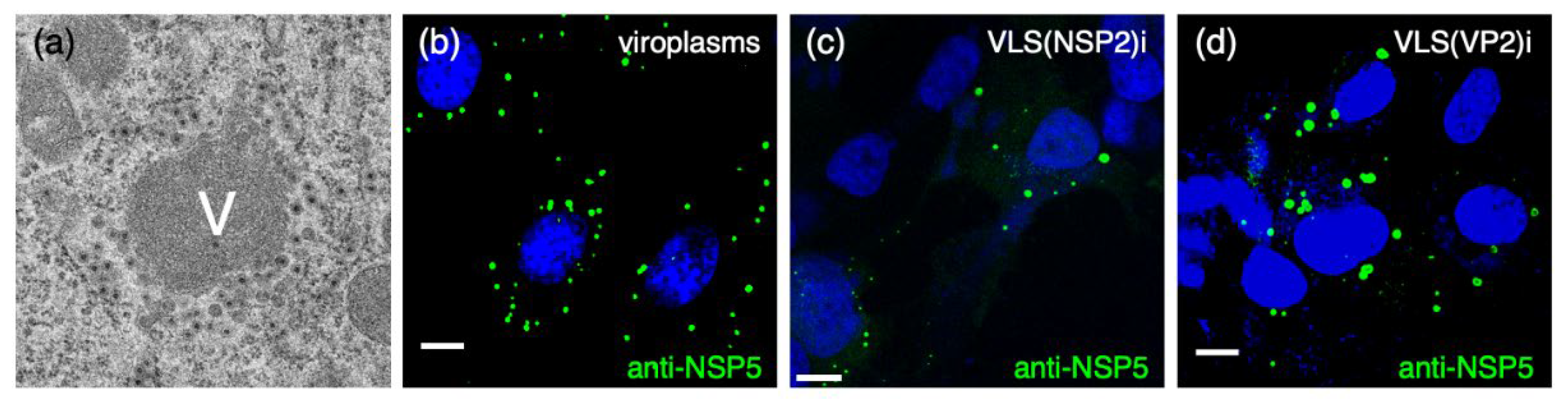
Figure 2.
Association of viroplasms with microtubules and actin. (a) Electron microscopy of SA11-infected MA104 cells at 8 hpi, showing viroplasm. Black arrowheads indicate the MT-bundles; viroplasms (V). Scale bar is 0.5 µm. (b) Immunofluorescence of SA11-infected MA104 cells at 6 hpi showing viroplasms (anti-NSP5, green), acetylated tubulin (mAb anti-acetylated tubulin, red) and nucleus (DAPI, blue), upper left image. The white-boxed area shows an enlarged photomicrograph indicating the localization of the hyper-acetylated MTs (white arrowheads) in the viroplasm region. Scale bar is 15 µm. From Eichwald et al., 2012. (c) Immunostaining of noninfected and SA11-infected MA104 cells. At 6 hpi, cells were fixed with methanol and immunostained to detect viroplasms (anti-NSP5, green) and actin cytoskeleton (anti-actin, cyan). Nuclei were stained with DAPI (blue). The scale bar is 20 μm. Immunostaining of noninfected and rRV/wt- or rRV/VP4-BAP-infected MA104 cells. At 6 hpi, cells were fixed with methanol and immunostained for detection of (d) VP4 (anti-VP4, green) and actin cytoskeleton (anti-actin, cyan). Nuclei were stained with DAPI (blue). The scale bar is 20 μm. Open yellow and red arrowheads point to stress fibers in the actin cytoskeleton and VP4 fibers, respectively. From Vetter et al., 2022.
Figure 2.
Association of viroplasms with microtubules and actin. (a) Electron microscopy of SA11-infected MA104 cells at 8 hpi, showing viroplasm. Black arrowheads indicate the MT-bundles; viroplasms (V). Scale bar is 0.5 µm. (b) Immunofluorescence of SA11-infected MA104 cells at 6 hpi showing viroplasms (anti-NSP5, green), acetylated tubulin (mAb anti-acetylated tubulin, red) and nucleus (DAPI, blue), upper left image. The white-boxed area shows an enlarged photomicrograph indicating the localization of the hyper-acetylated MTs (white arrowheads) in the viroplasm region. Scale bar is 15 µm. From Eichwald et al., 2012. (c) Immunostaining of noninfected and SA11-infected MA104 cells. At 6 hpi, cells were fixed with methanol and immunostained to detect viroplasms (anti-NSP5, green) and actin cytoskeleton (anti-actin, cyan). Nuclei were stained with DAPI (blue). The scale bar is 20 μm. Immunostaining of noninfected and rRV/wt- or rRV/VP4-BAP-infected MA104 cells. At 6 hpi, cells were fixed with methanol and immunostained for detection of (d) VP4 (anti-VP4, green) and actin cytoskeleton (anti-actin, cyan). Nuclei were stained with DAPI (blue). The scale bar is 20 μm. Open yellow and red arrowheads point to stress fibers in the actin cytoskeleton and VP4 fibers, respectively. From Vetter et al., 2022.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



