
Submitted:
25 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Oncolytic virotherapy is a promising immunotherapy approach for cancer treatment, utilizing viruses to preferentially infect and eliminate cancer cells while stimulating immune responses. In this review we synthesize current literature on the molecular circuits of immune sensing and response to oncolytic virotherapy; focusing on viral DNA or RNA sensing by infected cells, cytokine and danger-associated signal sensing by neighboring cells, and subsequent downstream activation of immune pathways. The sequential sense-and-response mechanisms involve the triggering of molecular sensors by viruses or infected cells to activate transcription factors and related genes for a breadth of immune responses. We describe how molecular signals induced in the tumor upon virotherapy can trigger diverse immune signaling pathways, activating both antigen-presenting cell-based innate and T cell-based adaptive immune responses. Insights into these complex mechanisms provide valuable knowledge for enhancing oncolytic virotherapy strategies.
Keywords:
molecular pathways
; oncolytic virus
; pattern recognition receptors
; transcription factors
1. Introduction
Oncolytic virotherapy is a novel class of immunotherapy for cancer treatment. It employs the use of natively occurring or genetically modified viruses, with the ability to preferentially infect and eliminate cancer cells [1]. Virus-induced infection and cancer cell death further initiate a cascade of events that not only reduce the tumor burden but also stimulate potent immune responses. This dual mode of action is regulated through the release of infection-related danger signals, activating diverse innate and adaptive immune pathways [2]. The intricate network of molecular signals and pathways, comparable to a ‘molecular circuit’, coordinates the activation of the immune response against cancer cells.
At a molecular level, infection by oncolytic viruses triggers signaling pathways of the immune response through the release of diverse immunogenic signals. For instance, viruses can trigger target cells directly through pathogen-associated molecular patterns (PAMP) or indirectly through infection-induced cell death, releasing cytokines or danger-associated molecular patterns (DAMP) [3,4]. These signals are sensed at a molecular level by the infected cell or neighboring immune cells. Upon detecting the presence of PAMPs or DAMPs, these responder cells release supplementary cytokines and chemokines, fostering a microenvironment conducive to robust antitumor immune responses [5]. As oncolytic virotherapies differ in genetic design and type, specific virus-cell interactions in terms of molecular sensing and response may shape immune-associated signaling in the tumor. Therefore a better understanding of specific molecular triggers induced by specific viruses can potentially help in optimizing their efficacy. As an illustration, talimogene laherparepvec (T-VEC) is an FDA and EMA-approved oncolytic virotherapy for advanced melanoma [6,7]. T-VEC employs a genetically modified herpes simplex virus encoding granulocyte-macrophage colony-stimulating factor (GM-CSF). GM-CSF, a key cytokine, plays a crucial role in dendritic cell activation and antigen presentation for effective anticancer T cell responses. Thus, understanding the molecular pathway of GM-CSF-induced immune responses has been crucial for optimizing T-VEC's design and efficacy. Following this rationale, various virotherapy candidates are currently being evaluated in preclinical and clinical stages for their ability to effectively induce durable antitumor immune responses [8,9].
In the present review, we aim to provide a comprehensive synthesis of the current literature surrounding the molecular circuits of immune sensing and response to oncolytic virotherapy. To focus on molecular signaling, our review will not assess additional factors that may influence the efficacy of virotherapy such as virus tropism, dosage/administration methods, or patient-specific factors like genetic variations. Moreover, we will not focus on virus-induced signaling that leads to therapeutic resistance as this has been reviewed earlier [10,11]. By exploring the diverse pathways triggered by oncolytic viruses and understanding the end response factors, our review aims to summarize key mechanisms associated with the induction of immune responses. To achieve this, we will review literature providing evidence on how oncolytic viruses and related signals are sensed by (i) infected cells, (ii) neighboring immune cells, and how this subsequently leads to (iii) activation of immune response in the tumor. Additionally, we review how various oncolytic viruses, distinguished by their genetic composition uniquely activate the immune system.
2. Molecular Sensing and Response to Oncolytic Viruses by Infected Cells
Cells, cancerous or healthy, utilize intricate molecular mechanisms to detect and respond to infection with oncolytic viruses. Upon infection, oncolytic viruses release their genetic material, either RNA or DNA depending on the virus type. RNA viruses like reovirus or picornavirus generate double-stranded RNA (dsRNA) during replication, while viruses such as measles or influenza produce single-stranded RNA (ssRNA). Conversely, DNA viruses like adenovirus or herpes simplex virus release their genetic material during replication. The entry of viral DNA into the cytoplasm during infection can occur through various mechanisms. Viruses with DNA genomes may exploit viral proteins or hijack host cellular machinery to transport DNA from the nucleus to the cytoplasm, facilitating viral replication and detection by cytoplasmic DNA sensors. Additionally, certain viruses may induce host cell damage or modify cellular membrane permeability, leading to the release of viral DNA into the cytoplasm. Amidst these complex interactions, pattern recognition receptors within infected cells, including toll-like receptors (TLRs), retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs), and cyclic GMP-AMP synthase (cGAS), play pivotal roles in sensing viral nucleic acids. These receptors serve as key sensors of the host immune system, initiating a wide range of immune responses.
2.1. Sensing Viral RNA
Sensing and responding to RNA-based oncolytic viruses is a multifaceted process crucial for the initiation of immune responses within host cells [3]. The cytoplasmic sensors RIG-I and MDA5 play pivotal roles in detecting both single-stranded and double-stranded RNA within infected cells (Figure 1A). Activation of these sensors elicits a cascade of events leading to the activation of transcription factors like nuclear factor kappa-light-chain-enhancer of activated B cells (NFΚB1) and interferon regulatory factors 3 and 7 (IRF3 and IRF7) [12,13]. Consequently, the expression of type-1 interferons-α and β (IFNα, IFNβ), pro-inflammatory cytokines like interleukin-18 and -12 (IL-18 and IL-12), tumor necrosis factor (TNF), and chemokines like C-X-C motif chemokine ligand 10 (CXCL10) is induced, contributing to the establishment of an antiviral state within the host cell. For instance, upon infection by oncolytic alphaviruses, proteins like RIG-I and TNF receptor-associated factor-6 (TRAF-6) are engaged, leading to the amplification of antiviral responses. Consequently, pro-inflammatory cytokines including IL-1beta, TNF-alpha, IL-6, and CXCL9 are expressed, enhancing the antitumor milieu and suppressing interferon-stimulated genes (ISGs) partly through zinc-finger antiviral protein (ZAP) expression [14,15,16,17,18,19,20,21]. Similarly, infection by coxsackievirus activates the RIG-I, triggering both innate and adaptive immune responses against tumors. This activation leads to the upregulation of IFN-inducible genes and Th1-associated chemokines, facilitating effector T-cell recruitment to the tumor microenvironment [22]. In hormone-refractory prostate cancer, Sendai virus also triggers the activation of RIG-I, leading to upregulation of IFN-related genes and subsequent activation of the Janus kinase and signal transducer and activator of transcription proteins (JAK/STAT) pathway, ultimately inducing apoptosis in cancer cells [23]. Virus-induced oncolysis mediated by the RIG-I signaling pathway, can also occur by upregulation of TNF-related apoptosis-inducing ligand, making it a promising target for cancer therapy [24]. This indicates the importance of viral RNA sensing pathways as a key mechanism to induce immune responses in the tumor.
2.2. Sensing Viral DNA
Various cellular sensors are also involved in sensing the DNA of oncolytic viruses. Cytoplasmic DNA sensing is facilitated by cGAS and Z-DNA binding protein 1 (ZBP1). Upon detection of cytoplasmic DNA, cGAS catalyzes the production of cyclic GMP-AMP (cGAMP), initiating downstream signaling events that culminate in the activation of transcription factors NFΚB1, IRF3, and IRF7 (Figure 1B) [25,26]. This results in the expression of type-1 IFNs (IFNα, IFNβ), pro-inflammatory cytokines (IL6), and chemokine motif ligands 5 and 4L1 (CCL5, CCL4L1), bolstering the cellular defense against viral invasion. Similarly, ZBP1 activation leads to the induction of IFNs (IFNα, IFNβ), contributing to the antiviral response elicited upon cytoplasmic DNA detection (Figure 1C). The cytoplasmic sensing of viral signals has been described to potentially induce innate immune responses. For example, cGAS-mediated sensing and activation of immune responses are crucial for the therapeutic efficacy of oncolytic herpes virus [27]. Additionally, it has been shown that epigenetic downregulation of cGAS-mediated signaling in ovarian cancers also leads to a downregulation of cytokine expression and related immune responses [28]. In this case, compensatory signaling mechanisms such as those activated by dsRNA through RIG-I/MDA5, remain largely unaffected and generate an immune response [28].
The recognition of methylated (CpG) DNA by endosomal toll-like receptor 9 (TLR9) activates the transcription factor NFκB1 (Figure 1D), triggering the expression of pro-inflammatory cytokines (TNF, IL1B, IL6, IL12A) and chemokines (CXCL8, CCL5, CCL3L3, CCL4L1) [12,29]. TLR-9-mediated recognition of viral signals can potentially induce immune responses in the tumor. This has inspired the development of genetically engineered oncolytic parvovirus that induces TLR-9 activation [30]. Incorporation of immunostimulatory CpG motifs into parvoviruses variants like JabCG1 and JabCG2, has been found to boost their adjuvant capacity. These variants trigger TLR-9-mediated signaling, leading to enhanced immunogenicity in animal models of cancer. Notably, JabCG2 demonstrated superior antitumor activity, inducing markers of cellular immunity and dendritic cell activation, thus reducing metastatic rates compared to other treatments. Alternatively, the loss of TLR9-mediated virus recognition has been shown to cause dysfunctional innate immune responses against oncolytic adenoviruses [28].
3. Molecular Sensing and Response to Infected Cells by neighboring Cells
Oncolytic virus-infected cells become active contributors to the intricate molecular signaling within the tumor microenvironment. A myriad of immunostimulatory molecules are released by virus-infected or dying tumor cells due to the diverse signaling pathways triggered by oncolytic viruses [31,32,33,34,35,36,37,38].
3.1. Sensing Cytokines from Infected Cells
Production of cytokines, particularly interferons, by infected cells serves as a critical component of the antiviral defense mechanism. One of the pivotal mechanisms involves the recognition of extracellular interferon by interferon-alpha receptors (IFNAR) on adjacent cells (Figure 2A) [39]. This interaction initiates a cascade of events that includes the activation of transcription factors STAT1, STAT2, or IRF9, ultimately leading to the induction of various genes such as CXCL9, CXCL10, CXCL11, major histocompatibility complex-I and II (MHC-I, and MHC-II) [40]. Therefore, interferons not only exert direct antiviral effects but also facilitate the activation of neighboring immune and other stromal cells. For instance, oncolytic adenoviruses and alphaviruses, induce interferon responses through various mechanisms. Adenoviruses trigger interferon signaling by activating the STING pathway and promoting the release of type-1 IFNs (IFNα, IFNβ) in infected tumor cells, which subsequently stimulate an antitumor immune response [41]. Similarly, alphaviruses induce interferon responses by upregulating autophagy and endoplasmic reticulum stress, leading to enhanced therapeutic efficacy through increased apoptosis and reduced tumor proliferation [42,43].
3.2. Sensing Pathogen-Associated Signals from Infected Cells
Molecules such as viral PAMPs (e.g. nucleic acids) serve as potent danger signals, triggering the activation of innate immune receptors in neighboring cells, thereby amplifying the antiviral response. For example, endocytosed or phagocytosed debris containing viral RNA is detected by Toll-like receptors-3, 7, or 8 (TLR-3, TLR7, or TLR-8) within antigen-presenting cells like plasmacytoid dendritic cells (pDCs) and macrophages [29]. The recognition of double-stranded viral RNA in the endosomes is mediated through TLR3 (Figure 2B). Upon encountering dsRNA, TLR3 activates a signaling cascade leading to the activation of transcription factors IRF3 and IRF7 [44]. Consequently, the expression of pivotal immune mediators such as interferons (IFNα and IFNβ), as well as various chemokines (CXCL9, CXCL10, CXCL11), and co-stimulatory molecules like clusters of differentiation 40, 80 or 86 (CD40, CD80, CD86) is induced, orchestrating a robust antiviral response. Single-stranded RNA derived from oncolytic viruses is sensed by endosomal TLR7 and/or 8 (Figure 2B-C) [12]. Activation of these receptors triggers downstream signaling pathways, culminating in the activation of the transcription factor NFΚB1. This activation prompts the expression of pro-inflammatory cytokines (TNF, IL1B, IL6, IL12A) and chemokines (CXCL8, CCL5, CCL3L3, CCL4L1), fostering an inflammatory milieu conducive to innate immune responses. This orchestrated response enhances the immune surveillance against viral infections and contributes to the shaping of an immune response in the tumor milieu. For example, Newcastle disease virus infection activates various signaling mechanisms, including TLR-signaling in tumors to regulate immune response and tumor-susceptibility markers [45]. Oncolytic parvovirus exploits TLR-signaling to induce human immune responses, enhancing dendritic cell maturation and stimulating NFκB-dependent activation of the adaptive immune system, thereby priming immune responses against tumors [46]. It has also been shown that immune responses induced by viral infection also synergize to enhance oncolysis. For example, IL-24 enhances apoptosis induced by influenza A virus via TLR3 and caspase-8 pathways, sensitizing cancer cells to TLR-mediated apoptosis [47]. Similarly, measles virus-based therapies induce oncolysis by activating plasmacytoid dendritic cells, resulting in the production of interferon-alpha and cross-presentation of tumor antigens, thereby facilitating antigen-specific immune responses against tumors [48]. Alternatively, some variants of parvoviruses escape TLR/RIG-I mediated immune sensing by tumor cells, resulting in lower IFN production and subsequent immune responses [49].
3.2. Sensing Danger-Associated Signals from Infected Cells
The process of viral replication and cell lysis leads to the release of cellular DAMPs from dying cancer cells, acting as an endogenous danger signal. For example, extracellular high-mobility group box 1 (HMGB1), is a DAMP sensed by neighboring antigen-presenting cells (Figure 2D) [50]. Interaction with Toll-like receptor 4 (TLR4) sets off a signaling cascade that involves the activation of transcription factors NFΚB1 or IRF5 [51]. This activation, in turn, leads to the expression of a plethora of pro-inflammatory and immunomodulatory molecules including IFNα, IFNβ, TNF, IL1B, IL 6, IL 12A, CXCL8, CCL5, CCL3L3, and CCL4L1. Thus, the recognition and response to DAMPs further amplify the immune response against viral infections and contribute to the overall activation of immune responses in the tumor microenvironment. For instance, upon infection with oncolytic adenoviruses, tumor cells release DAMPs such as HMGB1 and ATP, which activate dendritic cells and promote tumor-specific T cell responses, contributing to the antitumor immune response [41]. Similarly, oncolytic parvovirus induces cell death in glioma cells, activating dendritic cells and microglia, thereby breaking tumor tolerance and inducing long-term memory responses against tumors [27].
4. Molecular Signaling Response for Immune Activation
In response to signals emanating from viruses or infected cells, various components of the immune system, including macrophages, natural killer (NK) cells, T cells, dendritic cells, and B cells, engage in intricate molecular signaling pathways [5], culminating in the activation of both innate and adaptive immune responses [52].
4.1. Innate Immune Signaling
Activation of innate immune responses by oncolytic viruses plays a pivotal role in eliciting antitumor effects [4]. Innate immune signaling is regulated through diverse chemokines recognized by their respective chemokine receptors, triggering the activation of transcription factors such as Forkhead box O (FOXO), NFΚB1, and STAT, among others (Figure 3A). This activation results in the expression of a wide array of genes promoting immune cell proliferation, differentiation, and migration to sites of action [4,5]. For example, IL-2 or IL-4 sensed by their receptors, IL-2 receptor and IL-4 receptor respectively, activate the transcription factor STAT5, leading to the autocrine expression of IL-2, IL-4, IL-2 receptor, IL-4 receptor, and other cytokines (Figure 3B) [53,54]. Similarly, interferon-gamma (IFNγ) sensed by the interferon-gamma receptor (IFNGR) receptor triggers the activation of transcription factors STAT1 and STAT4, inducing the expression of IFNγ, T-box transcription factor (T-bet), and interleukin-12 receptor (IL12RB1) (Figure 3C) [55]. Activation of innate immune responses by oncolytic viruses plays a pivotal role in eliciting antitumor effects. Replication-competent Sendai viruses (rSeV) have been observed to activate dendritic cells through the RIG-I pathway, inducing the production of type I IFNs which contribute to their antimetastatic effects [56]. Similarly, the measles virus exploits its receptors CD150 and CD46 on tumor cells to trigger immune responses, leading to increased IFNγ levels and a favorable immune milieu for tumor clearance [57]. Vesicular stomatitis virus induces type I IFN responses, enhances dendritic cell maturation, and promotes antigen presentation, thus facilitating the initiation of adaptive immune responses against tumors [58,59,60].
4.2. Adaptive Immune Signaling
In addition to innate immune activation, oncolytic viruses also stimulate adaptive immune responses crucial for sustained antitumor effects. In the context of antigen-specific or target-specific immune activation, viral or tumor antigens presented by MHC-I and II molecules and sensed by T-cell receptors initiate signaling cascades [61,62]. This activation leads to the engagement of transcription factors like extracellular signal-regulated kinases 1 and 2 (ERK1, ERK2), and nuclear factor of activated T cells (NFAT), prompting the expression of perforin, granzyme, and FAS-ligand (FAS-L), ultimately resulting in the killing of target cells (Figure 4A). Similarly, antigens presented by MHC molecules and sensed by T-cell receptors activate transcription factors NFΚB1, NFATC1, protein c-Fos (FOS), and protein c-Jun (JUN), resulting in the expression of IFNγ, T-bet, TNF, GM-CSF, IL-2, IL-4, IL-5, IL-10, and IL-13 (Figure 4B) [61,62]. Moreover, the loss or downregulation of MHC molecules or the presence of unconventional MHC molecules, recognized by NK cell receptors, activate transcription factors ERK1, ERK2, and NFAT, leading to the expression of perforin, granzyme, and FAS-ligand, thereby facilitating target cell killing (Figure 4C) [63,64,65]. Overall it has been observed that various oncolytic virotherapy facilitates the activation of antigen-specific immune responses. For instance, Newcastle disease virus (NDV) oncolysis induces immunogenic cell death, leading to tumor infiltration by effector T lymphocytes and long-term tumor-specific immunological memory responses [66,67]. H-1 parvovirus selectively activates helper and not regulatory CD4+ T cells, thus demonstrating its potential as an anticancer treatment without exhibiting immunosuppressive effects [68]. The activation of helper CD4+ T cell responses by H-1 parvovirus further underscores its immunotherapeutic potential. These findings highlight the dual role of oncolytic viruses in triggering both innate and adaptive immune responses, thereby enhancing their efficacy as cancer therapeutics.
5. Strategies to Exploit Immune Signaling In Favor of Oncolytic Virotherapy
Various avenues have been explored to improve the safety and efficacy of oncolytic virotherapy. Inspired by the molecular mechanisms employed by virotherapy to trigger immune response pathways, efforts have been made to boost their immunogenic potential. These strategies include engineering viruses to enhance immune signaling, targeting tumors specifically, and employing combinatorial approaches with immunomodulatory agents or other viral therapies to amplify antitumor effects.
5.1. Engineering Viruses to Trigger Immune Signaling
Various oncolytic viruses have been engineered to bolster the immune response against cancer [69,70]. One approach involves encoding cytokines and chemokines within these viruses to enhance immune stimulation. For instance, adenoviruses can be armed with molecules such as GM-CSF and B7-1 to activate dendritic cells and facilitate T cell infiltration into tumors, thereby priming tumor-specific cytotoxic T lymphocytes [71]. Additionally, interleukin-12 expressed by adenoviruses promotes Th1-type immune responses and enhances NK cell and cytotoxic T cell activity [72]. Furthermore, molecules like CCL5 and beta-defensin-2 recruit and activate immune cells within the tumor microenvironment [73,74]. Incorporating CD40 ligand (CD40L) and IL-24 further amplifies immune-mediated tumor cell killing [75,76]. Adenoviruses encoding combinations of cytokines such as IL-12 and IL-18 could synergistically enhance antitumor immunity [77]. In HSV vectors, encoding immunomodulatory cytokines like IL-2, IL-12, and GM-CSF enhances tumor regression mediated by CD4+ and CD8+ lymphocytes [78,79,80]. Methods such as the HSVQuik system expedite the generation of oncolytic HSV vectors expressing immunomodulators for cancer gene therapy [81]. Similarly, oncolytic influenza A viruses armed with immune-stimulating molecules like OX40L exhibit potent oncolytic effects, selectively destroying tumor tissues and enhancing Th1-dominant immune responses [82]. Measles virus strains have demonstrated oncolytic capabilities in hepatocellular carcinoma and glioblastoma cells, inhibiting tumor growth and improving survival rates [83,84,85]. Recombinant Newcastle disease virus strains expressing cytokines such as IL-2, IL-15, or TRAIL stimulate tumor-specific CTL responses, inducing CD4+ and CD8+ T cell proliferation, leading to tumor regression [86,87]. Genetically engineered Sendai Virus carrying the IL-2 gene stimulates antitumor effects by modulating immune cell populations [88]. Oncolytic vaccinia viruses, armed with immunomodulatory molecules play a crucial role in activating molecular immune responses against cancer [89,90,91,92,93,94,95,96,97,98]. These viruses enhance antitumor efficacy by stimulating the activation of CD4+ and CD8+ T cells, promoting immune cell infiltration, and augmenting immune-based antitumoral activity through the delivery of chemokines and immunomodulatory antibodies. VSV engineered to encode therapeutic genes like IFN-beta enhances antitumor effects [99], although certain molecules like CD40L may not significantly improve efficacy [100].
5.2. Engineering Viruses for Tumor Targeting
Tumor targeting strategies aim to enhance the immunotherapeutic potential of oncolytic viruses by selectively targeting components of the tumor microenvironment. For instance, targeted depletion of tumor-associated macrophages by oncolytic adenovirus leads to microenvironmental repolarization towards a pro-inflammatory state, facilitating antitumor immune responses [101]. Utilizing tumor-specific promoters such as CXCR4-promoter allows for tumor-specific transgene expression, driving high expression of therapeutic genes like GM-CSF and B7-1, thereby enhancing immune cell activation and infiltration into tumors [71,102]. Various delivery strategies, including gelatin gel-mediated co-delivery and mesenchymal stem cell-mediated delivery, improve the sustained release and tumor targeting of oncolytic adenoviruses, thereby enhancing their therapeutic efficacy [103,104].
5.3. Combinatorial Approach to Boost Viral Immunogenicity
Combination strategies play a crucial role in enhancing the efficacy of oncolytic viruses in cancer therapy. One approach involves leveraging immune signaling pathways, exemplified by the combination of oncolytic adenoviral therapy with immune checkpoint inhibitors. Adenoviruses engineered to express TNF-alpha and IL-2, when coupled with anti-PD-1 therapy, exhibit augmented tumor control through increased CD8+ T cell infiltration and reduced immunosuppressive cell populations [72,105]. Additionally, combining oncolytic virotherapy with conventional treatments such as gemcitabine shows promise in enhancing oncolytic activity and eliciting systemic antitumor immunity by reducing myeloid-derived suppressor cells and promoting tumor regression [106]. Furthermore, combining oncolytic vaccinia viruses with immune checkpoint blockade amplifies antitumor immunity by increasing effector T cell infiltration, inducing PD-L1 expression, and reducing exhaustion markers [107,108].
Immunomodulatory agents further enhance the efficacy of oncolytic viruses. For instance, the combination of oncolytic virotherapy with the DTA-1 monoclonal antibody enhances tumor growth inhibition by fostering CD8+ T cell accumulation and diminishing regulatory T cells [109]. Similarly, blocking TNFα, which can impede oncolytic herpes virus replication by macrophages and microglia, augments virus replication and improves survival rates in glioblastoma models [110]. Augmenting dendritic cell populations through FMS-like tyrosine kinase ligand (Flt3L) administration improves tumor antigen cross-presentation and CD8+T cell responses, thereby enhancing the efficacy of oncolytic Newcastle disease virus [111]. Finally, synergistic antitumor effects are observed when combining oncolytic virotherapy with other viral therapies. Combining Newcastle disease virotherapy with influenza HA2 gene therapy or viral sensitizer-mediated therapy enhances immune responses and yields heightened antitumor effects [112,113].
6. Conclusion
In summary, our review delves into the intricate interplay between virus mechanisms of molecular sensing and the immune response across diverse oncolytic viruses. We systematically reviewed the literature concerning various molecular sensors, transcription factors, and immune signals associated with different oncolytic virotherapies (Figure 5). We summarize a sequential sense-and-response mechanism wherein diverse molecular sensors such as TLRs, RIG-I, and others detect viral genetic material or signals emanating from infected cells, ultimately triggering a cascade of innate and antigen-specific immune responses. This intricate process is finely regulated by the activation of various transcription factors, which in turn induce the expression of immune response genes. Notably, our analysis provides comprehensive insights into these molecular interactions, shedding light on the complex mechanisms underlying the efficacy of oncolytic virotherapies.
Author Contributions
Conceptualization, D.K.B and T.D.; data curation, D.K.B.; writing—original draft preparation, D.K.B; writing—review and editing, D.K.B and T.D; visualization, D.K.B; supervision, funding acquisition, T.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic Viruses: A New Class of Immunotherapy Drugs. Nature Reviews Drug Discovery 2015, 14, 642–662. [Google Scholar] [CrossRef] [PubMed]
- Lemos De Matos, A.; Franco, L.S.; McFadden, G. Oncolytic Viruses and the Immune System: The Dynamic Duo. Molecular Therapy - Methods & Clinical Development 2020, 17, 349–358. [Google Scholar] [CrossRef]
- Luan, X.; Wang, L.; Song, G.; Zhou, W. Innate Immune Responses to RNA: Sensing and Signaling. Front Immunol 2024, 15, 1287940. [Google Scholar] [CrossRef] [PubMed]
- Mealiea, D.; McCart, J.A. Cutting Both Ways: The Innate Immune Response to Oncolytic Virotherapy. Cancer Gene Ther 2022, 29, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Pol, J.G.; Workenhe, S.T.; Konda, P.; Gujar, S.; Kroemer, G. Cytokines in Oncolytic Virotherapy. Cytokine Growth Factor Rev 2020, 56, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Bommareddy, P.K.; Patel, A.; Hossain, S.; Kaufman, H.L. Talimogene Laherparepvec (T-VEC) and Other Oncolytic Viruses for the Treatment of Melanoma. Am J Clin Dermatol 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.; Collichio, F.A.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.; Spitler, L.; Puzanov, I.; Agarwala, S.; Milhem, M. Final Planned Overall Survival (OS) from OPTiM, a Randomized Phase III Trial of Talimogene Laherparepvec (T-VEC) versus GM-CSF for the Treatment of Unresected Stage IIIB/C/IV Melanoma (NCT00769704). J. Immunotherapy Cancer 2014, 2, P263. [Google Scholar] [CrossRef]
- Bhatt, D.K.; Wekema, L.; Carvalho Barros, L.R.; Chammas, R.; Daemen, T. A Systematic Analysis on the Clinical Safety and Efficacy of Onco-Virotherapy. Molecular Therapy - Oncolytics, 2021; S2372770521001364. [Google Scholar] [CrossRef]
- Macedo, N.; Miller, D.M.; Haq, R.; Kaufman, H.L. Clinical Landscape of Oncolytic Virus Research in 2020. J Immunother Cancer 2020, 8, e001486. [Google Scholar] [CrossRef]
- Bhatt, D.K.; Chammas, R.; Daemen, T. Resistance Mechanisms Influencing Oncolytic Virotherapy, a Systematic Analysis. Vaccines 2021, 9, 1166. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Ghorbani, E.; Khazaei, M.; Avan, A.; Ryzhikov, M.; Azadmanesh, K.; Hassanian, S.M. Interferon-Mediated Tumor Resistance to Oncolytic Virotherapy. J Cell Biochem 2017, 118, 1994–1999. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Toll-like Receptor and RIG-I-like Receptor Signaling. Ann N Y Acad Sci 2008, 1143, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Thoresen, D.; Wang, W.; Galls, D.; Guo, R.; Xu, L.; Pyle, A.M. The Molecular Mechanism of RIG-I Activation and Signaling. Immunol Rev 2021, 304, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhu, W.; Lin, Y.; Zhang, S.; Chen, X.; Gong, S.; He, S.; Hu, J.; Yan, G.; Liang, J. Systematic Characterization of the Biodistribution of the Oncolytic Virus M1. Hum Gene Ther 2020, 31, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, Y.; Lin, Y.; Liang, J.-K.; Zhong, W.-W.; Li, K.; Huang, W.-T.; Wang, D.-J.; Yan, G.-M.; Zhu, W.-B.; et al. Intravenous Injections of the Oncolytic Virus M1 as a Novel Therapy for Muscle-Invasive Bladder Cancer. Cell Death Dis 2018, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Hu, C.; Xing, F.; Gao, M.; Liang, J.; Xiao, X.; Cai, J.; Tan, Y.; Hu, J.; Zhu, W. Deficiency of the IRE1α-Autophagy Axis Enhances the Antitumor Effects of the Oncolytic Virus M1. J Virol 2018, 92, e01331-17. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, H.; Liang, J.; Li, K.; Zhu, W.; Fu, L.; Wang, F.; Zheng, X.; Shi, H.; Wu, S.; et al. Identification and Characterization of Alphavirus M1 as a Selective Oncolytic Virus Targeting ZAP-Defective Human Cancers. Proc Natl Acad Sci U S A 2014, 111, E4504–4512. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, K.; Zhu, W.-B.; Zhang, H.; Huang, W.-T.; Liu, X.-C.; Lin, Y.; Cai, J.; Yan, G.-M.; Qiu, J.-G.; et al. Suppression of CCDC6 Sensitizes Tumor to Oncolytic Virus M1. Neoplasia 2021, 23, 158–168. [Google Scholar] [CrossRef]
- Sun, S.; Liu, Y.; He, C.; Hu, W.; Liu, W.; Huang, X.; Wu, J.; Xie, F.; Chen, C.; Wang, J.; et al. Combining NanoKnife with M1 Oncolytic Virus Enhances Anticancer Activity in Pancreatic Cancer. Cancer Lett 2021, 502, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Toribio, R.; Díaz-López, I.; Berlanga, J.J.; Molina-Jiménez, F.; Majano, P.; Ventoso, I. Naturally Occurring and Engineered Alphaviruses Sensitive to Double-Stranded-RNA-Activated Protein Kinase Show Restricted Translation in Mammalian Cells, Increased Sensitivity to Interferon, and Marked Oncotropism. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, H.; Zou, H.; Tian, X.; Hu, J.; Qiu, P.; Hu, H.; Yan, G. Liposome Encapsulation of Oncolytic Virus M1 To Reduce Immunogenicity and Immune Clearance in Vivo. Mol Pharm 2019, 16, 779–785. [Google Scholar] [CrossRef]
- Annels, N.E.; Mansfield, D.; Arif, M.; Ballesteros-Merino, C.; Simpson, G.R.; Denyer, M.; Sandhu, S.S.; Melcher, A.A.; Harrington, K.J.; Davies, B.; et al. Phase I Trial of an ICAM-1-Targeted Immunotherapeutic-Coxsackievirus A21 (CVA21) as an Oncolytic Agent Against Non Muscle-Invasive Bladder Cancer. Clin Cancer Res 2019, 25, 5818–5831. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Miyamoto, Y.; Inoue, T.; Kaneda, Y. Efficient Eradication of Hormone-Resistant Human Prostate Cancers by Inactivated Sendai Virus Particle. Int J Cancer 2009, 124, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Matsushima-Miyagi, T.; Hatano, K.; Nomura, M.; Li-Wen, L.; Nishikawa, T.; Saga, K.; Shimbo, T.; Kaneda, Y. TRAIL and Noxa Are Selectively Upregulated in Prostate Cancer Cells Downstream of the RIG-I/MAVS Signaling Pathway by Nonreplicating Sendai Virus Particles. Clin Cancer Res 2012, 18, 6271–6283. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xu, P. Cellular Functions of cGAS-STING Signaling. Trends Cell Biol 2023, 33, 630–648. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, D.; Zhang, J.; Xiang, P.; Zeng, Z.; Xiong, W.; Shi, L. cGAS-STING Signaling in the Tumor Microenvironment. Cancer Lett 2023, 577, 216409. [Google Scholar] [CrossRef] [PubMed]
- Froechlich, G.; Caiazza, C.; Gentile, C.; D’Alise, A.M.; De Lucia, M.; Langone, F.; Leoni, G.; Cotugno, G.; Scisciola, V.; Nicosia, A.; et al. Integrity of the Antiviral STING-Mediated DNA Sensing in Tumor Cells Is Required to Sustain the Immunotherapeutic Efficacy of Herpes Simplex Oncolytic Virus. Cancers (Basel) 2020, 12, 3407. [Google Scholar] [CrossRef] [PubMed]
- Ahtiainen, L.; Mirantes, C.; Jahkola, T.; Escutenaire, S.; Diaconu, I.; Osterlund, P.; Kanerva, A.; Cerullo, V.; Hemminki, A. Defects in Innate Immunity Render Breast Cancer Initiating Cells Permissive to Oncolytic Adenovirus. PLoS One 2010, 5, e13859. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Liu, Y.-J. Regulation of TLR7/9 Signaling in Plasmacytoid Dendritic Cells. Protein Cell 2013, 4, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Raykov, Z.; Grekova, S.; Leuchs, B.; Aprahamian, M.; Rommelaere, J. Arming Parvoviruses with CpG Motifs to Improve Their Oncosuppressive Capacity. Int J Cancer 2008, 122, 2880–2884. [Google Scholar] [CrossRef]
- Efferson, C.L.; Tsuda, N.; Kawano, K.; Nistal-Villán, E.; Sellappan, S.; Yu, D.; Murray, J.L.; García-Sastre, A.; Ioannides, C.G. Prostate Tumor Cells Infected with a Recombinant Influenza Virus Expressing a Truncated NS1 Protein Activate Cytolytic CD8+ Cells to Recognize Noninfected Tumor Cells. J Virol 2006, 80, 383–394. [Google Scholar] [CrossRef]
- Elankumaran, S.; Rockemann, D.; Samal, S.K. Newcastle Disease Virus Exerts Oncolysis by Both Intrinsic and Extrinsic Caspase-Dependent Pathways of Cell Death. J Virol 2006, 80, 7522–7534. [Google Scholar] [CrossRef] [PubMed]
- Puhlmann, J.; Puehler, F.; Mumberg, D.; Boukamp, P.; Beier, R. Rac1 Is Required for Oncolytic NDV Replication in Human Cancer Cells and Establishes a Link between Tumorigenesis and Sensitivity to Oncolytic Virus. Oncogene 2010, 29, 2205–2216. [Google Scholar] [CrossRef] [PubMed]
- Okemoto, K.; Wagner, B.; Meisen, H.; Haseley, A.; Kaur, B.; Chiocca, E.A. STAT3 Activation Promotes Oncolytic HSV1 Replication in Glioma Cells. PLoS One 2013, 8, e71932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, X.; Yuan, Y.; Gong, X.; Chen, Z.; Xu, X. IPS-1 Plays a Dual Function to Directly Induce Apoptosis in Murine Melanoma Cells by Inactivated Sendai Virus. Int J Cancer 2014, 134, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-W.; Nishikawa, T.; Kaneda, Y. An RNA Molecule Derived From Sendai Virus DI Particles Induces Antitumor Immunity and Cancer Cell-Selective Apoptosis. Mol Ther 2016, 24, 135–145. [Google Scholar] [CrossRef]
- Hummel, J.L.; Safroneeva, E.; Mossman, K.L. The Role of ICP0-Null HSV-1 and Interferon Signaling Defects in the Effective Treatment of Breast Adenocarcinoma. Mol Ther 2005, 12, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Nanni, P.; Gatta, V.; Menotti, L.; De Giovanni, C.; Ianzano, M.; Palladini, A.; Grosso, V.; Dall’ora, M.; Croci, S.; Nicoletti, G.; et al. Preclinical Therapy of Disseminated HER-2+ Ovarian and Breast Carcinomas with a HER-2-Retargeted Oncolytic Herpesvirus. PLoS Pathog 2013, 9, e1003155. [Google Scholar] [CrossRef]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of Type I Interferon Responses. Nat Rev Immunol 2014, 14, 36–49. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu Rev Immunol 2014, 32, 513–545. [Google Scholar] [CrossRef]
- Di Somma, S.; Iannuzzi, C.A.; Passaro, C.; Forte, I.M.; Iannone, R.; Gigantino, V.; Indovina, P.; Botti, G.; Giordano, A.; Formisano, P.; et al. The Oncolytic Virus Dl922-947 Triggers Immunogenic Cell Death in Mesothelioma and Reduces Xenograft Growth. Front Oncol 2019, 9, 564. [Google Scholar] [CrossRef]
- Abraham, R.; Mudaliar, P.; Padmanabhan, A.; Sreekumar, E. Induction of Cytopathogenicity in Human Glioblastoma Cells by Chikungunya Virus. PLoS One 2013, 8, e75854. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Tan, J.; Zhang, Y.; Wong, C.-W.; Lin, Z.; Liu, X.; Sander, M.; Yang, X.; Liang, L.; et al. Necroptotic Virotherapy of Oncolytic Alphavirus M1 Cooperated with Doxorubicin Displays Promising Therapeutic Efficacy in TNBC. Oncogene 2021, 40, 4783–4795. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like Receptor 3 (TLR3) Regulation Mechanisms and Roles in Antiviral Innate Immune Responses. J Zhejiang Univ Sci B 2021, 22, 609–632. [Google Scholar] [CrossRef] [PubMed]
- Balogh, A.; Bátor, J.; Markó, L.; Németh, M.; Pap, M.; Sétáló, G.; Müller, D.N.; Csatary, L.K.; Szeberényi, J. Gene Expression Profiling in PC12 Cells Infected with an Oncolytic Newcastle Disease Virus Strain. Virus Res 2014, 185, 10–22. [Google Scholar] [CrossRef]
- Sieben, M.; Schäfer, P.; Dinsart, C.; Galle, P.R.; Moehler, M. Activation of the Human Immune System via Toll-like Receptors by the Oncolytic Parvovirus H-1. Int J Cancer 2013, 132, 2548–2556. [Google Scholar] [CrossRef]
- Weiss, R.; Sachet, M.; Zinngrebe, J.; Aschacher, T.; Krainer, M.; Hegedus, B.; Walczak, H.; Bergmann, M. IL-24 Sensitizes Tumor Cells to TLR3-Mediated Apoptosis. Cell Death Differ 2013, 20, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Guillerme, J.-B.; Boisgerault, N.; Roulois, D.; Ménager, J.; Combredet, C.; Tangy, F.; Fonteneau, J.-F.; Gregoire, M. Measles Virus Vaccine–Infected Tumor Cells Induce Tumor Antigen Cross-Presentation by Human Plasmacytoid Dendritic Cells. Clin Cancer Res 2013, 19, 1147–1158. [Google Scholar] [CrossRef]
- Angelova, A.; Pierrard, K.; Detje, C.N.; Santiago, E.; Grewenig, A.; Nüesch, J.P.F.; Kalinke, U.; Ungerechts, G.; Rommelaere, J.; Daeffler, L. Oncolytic Rodent Protoparvoviruses Evade a TLR- and RLR-Independent Antiviral Response in Transformed Cells. Pathogens 2023, 12, 607. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front Immunol 2020, 11, 484. [Google Scholar] [CrossRef]
- Sims, G.P.; Rowe, D.C.; Rietdijk, S.T.; Herbst, R.; Coyle, A.J. HMGB1 and RAGE in Inflammation and Cancer. Annu Rev Immunol 2010, 28, 367–388. [Google Scholar] [CrossRef]
- Engeland, C.E.; Bell, J.C. Introduction to Oncolytic Virotherapy. Methods Mol Biol 2020, 2058, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu Rev Immunol 2018, 36, 411–433. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. T Helper 2 (Th2) Cell Differentiation, Type 2 Innate Lymphoid Cell (ILC2) Development and Regulation of Interleukin-4 (IL-4) and IL-13 Production. Cytokine 2015, 75, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Schoenborn, J.R.; Wilson, C.B. Regulation of Interferon-Gamma during Innate and Adaptive Immune Responses. Adv Immunol 2007, 96, 41–101. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Ueda, Y.; Kinoh, H.; Yoneyama, Y.; Matsunaga, A.; Komaru, A.; Harada, Y.; Suzuki, H.; Komiya, A.; Shibata, S.; et al. RIG-I Helicase-Independent Pathway in Sendai Virus-Activated Dendritic Cells Is Critical for Preventing Lung Metastasis of AT6.3 Prostate Cancer. Neoplasia 2010, 12, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Heinzerling, L.; Künzi, V.; Oberholzer, P.A.; Kündig, T.; Naim, H.; Dummer, R. Oncolytic Measles Virus in Cutaneous T-Cell Lymphomas Mounts Antitumor Immune Responses in Vivo and Targets Interferon-Resistant Tumor Cells. Blood 2005, 106, 2287–2294. [Google Scholar] [CrossRef]
- Ahmed, M.; Brzoza, K.L.; Hiltbold, E.M. Matrix Protein Mutant of Vesicular Stomatitis Virus Stimulates Maturation of Myeloid Dendritic Cells. J Virol 2006, 80, 2194–2205. [Google Scholar] [CrossRef] [PubMed]
- Willmon, C.L.; Saloura, V.; Fridlender, Z.G.; Wongthida, P.; Diaz, R.M.; Thompson, J.; Kottke, T.; Federspiel, M.; Barber, G.; Albelda, S.M.; et al. Expression of IFN-Beta Enhances Both Efficacy and Safety of Oncolytic Vesicular Stomatitis Virus for Therapy of Mesothelioma. Cancer Res 2009, 69, 7713–7720. [Google Scholar] [CrossRef] [PubMed]
- Wongthida, P.; Diaz, R.M.; Galivo, F.; Kottke, T.; Thompson, J.; Melcher, A.; Vile, R. VSV Oncolytic Virotherapy in the B16 Model Depends upon Intact MyD88 Signaling. Mol Ther 2011, 19, 150–158. [Google Scholar] [CrossRef]
- Gaud, G.; Lesourne, R.; Love, P.E. Regulatory Mechanisms in T Cell Receptor Signalling. Nat Rev Immunol 2018, 18, 485–497. [Google Scholar] [CrossRef]
- Brownlie, R.J.; Zamoyska, R. T Cell Receptor Signalling Networks: Branched, Diversified and Bounded. Nat Rev Immunol 2013, 13, 257–269. [Google Scholar] [CrossRef]
- Prager, I.; Watzl, C. Mechanisms of Natural Killer Cell-Mediated Cellular Cytotoxicity. J Leukoc Biol 2019, 105, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.A.; Yagita, H.; Takeda, K.; van Dommelen, S.L.H.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK Cell Cytotoxicity. Mol Immunol 2005, 42, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Nunès, J.A.; Vély, F. Natural Killer Cell Signaling Pathways. Science 2004, 306, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Long, Y.; Liu, B.; Yang, D.; Li, C.; Chen, T.; Wang, X.; Liu, C.; Zhu, H. ISG12a Mediates Cell Response to Newcastle Disease Viral Infection. Virology 2014, 462–463, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Koks, C.A.; Garg, A.D.; Ehrhardt, M.; Riva, M.; Vandenberk, L.; Boon, L.; De Vleeschouwer, S.; Agostinis, P.; Graf, N.; Van Gool, S.W. Newcastle Disease Virotherapy Induces Long-Term Survival and Tumor-Specific Immune Memory in Orthotopic Glioma through the Induction of Immunogenic Cell Death. Int J Cancer 2015, 136, E313–325. [Google Scholar] [CrossRef] [PubMed]
- Moralès, O.; Richard, A.; Martin, N.; Mrizak, D.; Sénéchal, M.; Miroux, C.; Pancré, V.; Rommelaere, J.; Caillet-Fauquet, P.; de Launoit, Y. Activation of a Helper and Not Regulatory Human CD4+ T Cell Response by Oncolytic H-1 Parvovirus. PLoS One 2012, 7, e32197. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.T.; Bell, J.C. Oncolytic Virus Combination Therapy: Killing One Bird with Two Stones. Molecular Therapy 2018, 26, 1414–1422. [Google Scholar] [CrossRef] [PubMed]
- Twumasi-Boateng, K.; Pettigrew, J.L.; Kwok, Y.Y.E.; Bell, J.C.; Nelson, B.H. Oncolytic Viruses as Engineering Platforms for Combination Immunotherapy. Nature Reviews Cancer 2018. [Google Scholar] [CrossRef]
- Choi, K.-J.; Kim, J.-H.; Lee, Y.-S.; Kim, J.; Suh, B.-S.; Kim, H.; Cho, S.; Sohn, J.-H.; Kim, G.E.; Yun, C.-O. Concurrent Delivery of GM-CSF and B7-1 Using an Oncolytic Adenovirus Elicits Potent Antitumor Effect. Gene Ther 2006, 13, 1010–1020. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Kim, J.-H.; Choi, K.-J.; Choi, I.-K.; Kim, H.; Cho, S.; Cho, B.C.; Yun, C.-O. Enhanced Antitumor Effect of Oncolytic Adenovirus Expressing Interleukin-12 and B7-1 in an Immunocompetent Murine Model. Clin Cancer Res 2006, 12, 5859–5868. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, N.; Aldrich, M.; Rollins, L.; Ren, W.; Goltsova, T.; Chen, S.-Y.; Huang, X.F. Attraction and Activation of Dendritic Cells at the Site of Tumor Elicits Potent Antitumor Immunity. Mol Ther 2009, 17, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, N.; Aldrich, M.; Weksberg, D.; Rollins, L.; Goltsova, T.; Chen, S.-Y.; Huang, X.F. Targeting the Intratumoral Dendritic Cells by the Oncolytic Adenoviral Vaccine Expressing RANTES Elicits Potent Antitumor Immunity. J Immunother 2009, 32, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.S.; Gomes, E.M.; Butcher, L.D.; Hernandez-Alcoceba, R.; Chang, D.; Kansopon, J.; Newman, J.; Stone, M.J.; Tong, A.W. Growth Inhibition of Human Multiple Myeloma Cells by an Oncolytic Adenovirus Carrying the CD40 Ligand Transgene. Clin Cancer Res 2009, 15, 4847–4856. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wei, R.; Liu, X.; Zeng, Y.; Huang, H.; Ding, M.; Zhang, K.; Liu, X.-Y. Cancer Targeting Gene-Viro-Therapy Specific for Liver Cancer by α-Fetoprotein-Controlled Oncolytic Adenovirus Expression of SOCS3 and IL-24. Acta Biochim Biophys Sin (Shanghai) 2011, 43, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-K.; Lee, J.-S.; Zhang, S.-N.; Park, J.; Sonn, C.H.; Lee, K.-M.; Yun, C.-O. Oncolytic Adenovirus Co-Expressing IL-12 and IL-18 Improves Tumor-Specific Immunity via Differentiation of T Cells Expressing IL-12Rβ2 or IL-18Rα. Gene Ther 2011, 18, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Zager, J.S.; Delman, K.A.; Malhotra, S.; Ebright, M.I.; Bennett, J.J.; Kates, T.; Halterman, M.; Federoff, H.; Fong, Y. Combination Vascular Delivery of Herpes Simplex Oncolytic Viruses and Amplicon Mediated Cytokine Gene Transfer Is Effective Therapy for Experimental Liver Cancer. Mol Med 2001, 7, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.J.; Chan, M.-K.; Yu, Z.; Kim, T.H.; Bhargava, A.; Stiles, B.M.; Horsburgh, B.C.; Shah, J.P.; Ghossein, R.A.; Singh, B.; et al. Effective Intravenous Therapy of Murine Pulmonary Metastases with an Oncolytic Herpes Virus Expressing Interleukin 12. Clin Cancer Res 2004, 10, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Kim, T.; Zager, J.; Bennett, J.; Ebright, M.; D’Angelica, M.; Fong, Y. Use of an Oncolytic Virus Secreting GM-CSF as Combined Oncolytic and Immunotherapy for Treatment of Colorectal and Hepatic Adenocarcinomas. Surgery 2007, 141, 520–529. [Google Scholar] [CrossRef]
- Terada, K.; Wakimoto, H.; Tyminski, E.; Chiocca, E.A.; Saeki, Y. Development of a Rapid Method to Generate Multiple Oncolytic HSV Vectors and Their in Vivo Evaluation Using Syngeneic Mouse Tumor Models. Gene Ther 2006, 13, 705–714. [Google Scholar] [CrossRef]
- Yang, H.; Lei, G.; Deng, Z.; Sun, F.; Tian, Y.; Cheng, J.; Yu, H.; Li, C.; Bai, C.; Zhang, S.; et al. An Engineered Influenza a Virus Expressing the Co-Stimulator OX40L as an Oncolytic Agent Against Hepatocellular Carcinoma. J Hepatocell Carcinoma 2024, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Blechacz, B.; Splinter, P.L.; Greiner, S.; Myers, R.; Peng, K.-W.; Federspiel, M.J.; Russell, S.J.; LaRusso, N.F. Engineered Measles Virus as a Novel Oncolytic Viral Therapy System for Hepatocellular Carcinoma. Hepatology 2006, 44, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Paraskevakou, G.; Allen, C.; Nakamura, T.; Zollman, P.; James, C.D.; Peng, K.W.; Schroeder, M.; Russell, S.J.; Galanis, E. Epidermal Growth Factor Receptor (EGFR)-Retargeted Measles Virus Strains Effectively Target EGFR- or EGFRvIII Expressing Gliomas. Mol Ther 2007, 15, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.; Paraskevakou, G.; Iankov, I.; Giannini, C.; Schroeder, M.; Sarkaria, J.; Schroeder, M.; Puri, R.K.; Russell, S.J.; Galanis, E. Interleukin-13 Displaying Retargeted Oncolytic Measles Virus Strains Have Significant Activity against Gliomas with Improved Specificity. Mol Ther 2008, 16, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Niu, Z.; Tian, H.; Li, S.; Lv, Z.; Zhang, T.; Ren, G.; Li, D. Genetically Engineered Newcastle Disease Virus Expressing Interleukin 2 Is a Potential Drug Candidate for Cancer Immunotherapy. Immunol Lett 2014, 159, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Bai, F.; Sun, T.; Tian, H.; Yu, D.; Yin, J.; Li, S.; Li, T.; Cao, H.; Yu, Q.; et al. Recombinant Newcastle Disease Virus Expressing IL15 Demonstrates Promising Antitumor Efficiency in Melanoma Model. Technol Cancer Res Treat 2015, 14, 607–615. [Google Scholar] [CrossRef]
- Takehara, Y.; Satoh, T.; Nishizawa, A.; Saeki, K.; Nakamura, M.; Masuzawa, M.; Kaneda, Y.; Katayama, I.; Yokozeki, H. Anti-Tumor Effects of Inactivated Sendai Virus Particles with an IL-2 Gene on Angiosarcoma. Clin Immunol 2013, 149, 1–10. [Google Scholar] [CrossRef]
- Wang, L.-C.S.; Lynn, R.C.; Cheng, G.; Alexander, E.; Kapoor, V.; Moon, E.K.; Sun, J.; Fridlender, Z.G.; Isaacs, S.N.; Thorne, S.H.; et al. Treating Tumors with a Vaccinia Virus Expressing IFNβ Illustrates the Complex Relationships between Oncolytic Ability and Immunogenicity. Mol Ther 2012, 20, 736–748. [Google Scholar] [CrossRef]
- Fend, L.; Yamazaki, T.; Remy, C.; Fahrner, C.; Gantzer, M.; Nourtier, V.; Préville, X.; Quéméneur, E.; Kepp, O.; Adam, J.; et al. Immune Checkpoint Blockade, Immunogenic Chemotherapy or IFN-α Blockade Boost the Local and Abscopal Effects of Oncolytic Virotherapy. Cancer Res 2017, 77, 4146–4157. [Google Scholar] [CrossRef]
- John, L.B.; Howland, L.J.; Flynn, J.K.; West, A.C.; Devaud, C.; Duong, C.P.; Stewart, T.J.; Westwood, J.A.; Guo, Z.S.; Bartlett, D.L.; et al. Oncolytic Virus and Anti-4-1BB Combination Therapy Elicits Strong Antitumor Immunity against Established Cancer. Cancer Res 2012, 72, 1651–1660. [Google Scholar] [CrossRef]
- Li, J.; O’Malley, M.; Sampath, P.; Kalinski, P.; Bartlett, D.L.; Thorne, S.H. Expression of CCL19 from Oncolytic Vaccinia Enhances Immunotherapeutic Potential While Maintaining Oncolytic Activity. Neoplasia 2012, 14, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Chard, L.S.; Maniati, E.; Wang, P.; Zhang, Z.; Gao, D.; Wang, J.; Cao, F.; Ahmed, J.; El Khouri, M.; Hughes, J.; et al. A Vaccinia Virus Armed with Interleukin-10 Is a Promising Therapeutic Agent for Treatment of Murine Pancreatic Cancer. Clin Cancer Res 2015, 21, 405–416. [Google Scholar] [CrossRef]
- Rojas, J.J.; Sampath, P.; Bonilla, B.; Ashley, A.; Hou, W.; Byrd, D.; Thorne, S.H. Manipulating TLR Signaling Increases the Anti-Tumor T Cell Response Induced by Viral Cancer Therapies. Cell Rep 2016, 15, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Kowalsky, S.J.; Liu, Z.; Feist, M.; Berkey, S.E.; Ma, C.; Ravindranathan, R.; Dai, E.; Roy, E.J.; Guo, Z.S.; Bartlett, D.L. Superagonist IL-15-Armed Oncolytic Virus Elicits Potent Antitumor Immunity and Therapy That Are Enhanced with PD-1 Blockade. Mol Ther 2018, 26, 2476–2486. [Google Scholar] [CrossRef] [PubMed]
- Riederer, S.; Fux, R.; Lehmann, M.H.; Volz, A.; Sutter, G.; Rojas, J.J. Activation of Interferon Regulatory Factor 3 by Replication-Competent Vaccinia Viruses Improves Antitumor Efficacy Mediated by T Cell Responses. Mol Ther Oncolytics 2021, 22, 399–409. [Google Scholar] [CrossRef] [PubMed]
- DePeaux, K.; Rivadeneira, D.B.; Lontos, K.; Dean, V.G.; Gunn, W.G.; Watson, M.J.; Yao, T.; Wilfahrt, D.; Hinck, C.; Wieteska, L.; et al. An Oncolytic Virus-Delivered TGFβ Inhibitor Overcomes the Immunosuppressive Tumor Microenvironment. J Exp Med 2023, 220, e20230053. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sheng, Y.; Hou, W.; Sampath, P.; Byrd, D.; Thorne, S.; Zhang, Y. CCL5-Armed Oncolytic Virus Augments CCR5-Engineered NK Cell Infiltration and Antitumor Efficiency. J Immunother Cancer 2020, 8, e000131. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.R.; Jacobson, B.A.; Ji, Y.; Drees, J.; Tang, S.; Xiong, K.; Wang, H.; Prigge, J.E.; Dash, A.S.; Kratzke, A.K.; et al. Vesicular Stomatitis Virus Expressing Interferon-β Is Oncolytic and Promotes Antitumor Immune Responses in a Syngeneic Murine Model of Non-Small Cell Lung Cancer. Oncotarget 2015, 6, 33165–33177. [Google Scholar] [CrossRef] [PubMed]
- Galivo, F.; Diaz, R.M.; Thanarajasingam, U.; Jevremovic, D.; Wongthida, P.; Thompson, J.; Kottke, T.; Barber, G.N.; Melcher, A.; Vile, R.G. Interference of CD40L-Mediated Tumor Immunotherapy by Oncolytic Vesicular Stomatitis Virus. Hum Gene Ther 2010, 21, 439–450. [Google Scholar] [CrossRef]
- Scott, E.M.; Jacobus, E.J.; Lyons, B.; Frost, S.; Freedman, J.D.; Dyer, A.; Khalique, H.; Taverner, W.K.; Carr, A.; Champion, B.R.; et al. Bi- and Tri-Valent T Cell Engagers Deplete Tumour-Associated Macrophages in Cancer Patient Samples. J Immunother Cancer 2019, 7, 320. [Google Scholar] [CrossRef]
- Stoff-Khalili, M.A.; Stoff, A.; Rivera, A.A.; Banerjee, N.S.; Everts, M.; Young, S.; Siegal, G.P.; Richter, D.F.; Wang, M.; Dall, P.; et al. Preclinical Evaluation of Transcriptional Targeting Strategies for Carcinoma of the Breast in a Tissue Slice Model System. Breast Cancer Res 2005, 7, R1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Oh, J.-E.; Hong, J.; Chung, Y.; Lee, Y.; Park, K.D.; Kim, S.; Yun, C.-O. Optimized Biodegradable Polymeric Reservoir-Mediated Local and Sustained Co-Delivery of Dendritic Cells and Oncolytic Adenovirus Co-Expressing IL-12 and GM-CSF for Cancer Immunotherapy. J Control Release 2017, 259, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.-T.; Wu, M.-H.; Chen, M.-J.; Lin, S.-P.; Yen, Y.-T.; Hung, S.-C. Combination of Mesenchymal Stem Cell-Delivered Oncolytic Virus with Prodrug Activation Increases Efficacy and Safety of Colorectal Cancer Therapy. Biomedicines 2021, 9, 548. [Google Scholar] [CrossRef]
- Cervera-Carrascon, V.; Quixabeira, D.C.A.; Santos, J.M.; Havunen, R.; Milenova, I.; Verhoeff, J.; Heiniö, C.; Zafar, S.; Garcia-Vallejo, J.J.; van Beusechem, V.W.; et al. Adenovirus Armed With TNFa and IL2 Added to aPD-1 Regimen Mediates Antitumor Efficacy in Tumors Refractory to aPD-1. Front Immunol 2021, 12, 706517. [Google Scholar] [CrossRef] [PubMed]
- Esaki, S.; Goshima, F.; Kimura, H.; Murakami, S.; Nishiyama, Y. Enhanced Antitumoral Activity of Oncolytic Herpes Simplex Virus with Gemcitabine Using Colorectal Tumor Models. Int J Cancer 2013, 132, 1592–1601. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ravindranathan, R.; Kalinski, P.; Guo, Z.S.; Bartlett, D.L. Rational Combination of Oncolytic Vaccinia Virus and PD-L1 Blockade Works Synergistically to Enhance Therapeutic Efficacy. Nat Commun 2017, 8, 14754. [Google Scholar] [CrossRef] [PubMed]
- Mistarz, A.; Komorowski, M.P.; Graczyk, M.A.; Gil, M.; Jiang, A.; Opyrchal, M.; Rokita, H.; Odunsi, K.O.; Kozbor, D. Recruitment of Intratumoral CD103+ Dendritic Cells by a CXCR4 Antagonist-Armed Virotherapy Enhances Antitumor Immunity. Mol Ther Oncolytics 2019, 14, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, M.; Seo, N.; Mitsui, J.; Muraoka, D.; Tanaka, M.; Mineno, J.; Ikeda, H.; Shiku, H. Systemic CD8+ T Cell-Mediated Tumoricidal Effects by Intratumoral Treatment of Oncolytic Herpes Simplex Virus with the Agonistic Monoclonal Antibody for Murine Glucocorticoid-Induced Tumor Necrosis Factor Receptor. PLoS One 2014, 9, e104669. [Google Scholar] [CrossRef] [PubMed]
- Meisen, W.H.; Wohleb, E.S.; Jaime-Ramirez, A.C.; Bolyard, C.; Yoo, J.Y.; Russell, L.; Hardcastle, J.; Dubin, S.; Muili, K.; Yu, J.; et al. The Impact of Macrophage- and Microglia-Secreted TNFα on Oncolytic HSV-1 Therapy in the Glioblastoma Tumor Microenvironment. Clin Cancer Res 2015, 21, 3274–3285. [Google Scholar] [CrossRef]
- Svensson-Arvelund, J.; Cuadrado-Castano, S.; Pantsulaia, G.; Kim, K.; Aleynick, M.; Hammerich, L.; Upadhyay, R.; Yellin, M.; Marsh, H.; Oreper, D.; et al. Expanding Cross-Presenting Dendritic Cells Enhances Oncolytic Virotherapy and Is Critical for Long-Term Anti-Tumor Immunity. Nat Commun 2022, 13, 7149. [Google Scholar] [CrossRef]
- Miri, S.M.; Ebrahimzadeh, M.S.; Abdolalipour, E.; Yazdi, M.; Hosseini Ravandi, H.; Ghaemi, A. Synergy between Hemagglutinin 2 (HA2) Subunit of Influenza Fusogenic Membrane Glycoprotein and Oncolytic Newcastle Disease Virus Suppressed Tumor Growth and Further Enhanced by Immune Checkpoint PD-1 Blockade. Cancer Cell Int 2020, 20, 380. [Google Scholar] [CrossRef] [PubMed]
- McAusland, T.M.; van Vloten, J.P.; Santry, L.A.; Guilleman, M.M.; Rghei, A.D.; Ferreira, E.M.; Ingrao, J.C.; Arulanandam, R.; Major, P.P.; Susta, L.; et al. Combining Vanadyl Sulfate with Newcastle Disease Virus Potentiates Rapid Innate Immune-Mediated Regression with Curative Potential in Murine Cancer Models. Mol Ther Oncolytics 2021, 20, 306–324. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Direct sensing of oncolytic viruses and related immune responses. The figure illustrates essential pathways involved in molecular sensing and response during viral infection. (A) Cytoplasmic receptors RIG-I and MDA5 detect single or double-stranded RNA, activating transcription factors NFΚB1, IRF3, and IRF7, leading to the production of interferons and pro-inflammatory cytokines. (B) Cytoplasmic DNA sensed by cGAS triggers the activation of transcription factors NFΚB1, IRF3, and IRF7, leading to the expression of key cytokines and chemokines. (C) Cytoplasmic DNA detection by ZBP1 results in the activation of transcription factors IRF3 and IRF7, inducing the expression of interferons. (D) Methylated CpG DNA recognition by endosomal TLR9 activates NFΚB1, leading to the expression of pro-inflammatory mediators.
Figure 1.
Direct sensing of oncolytic viruses and related immune responses. The figure illustrates essential pathways involved in molecular sensing and response during viral infection. (A) Cytoplasmic receptors RIG-I and MDA5 detect single or double-stranded RNA, activating transcription factors NFΚB1, IRF3, and IRF7, leading to the production of interferons and pro-inflammatory cytokines. (B) Cytoplasmic DNA sensed by cGAS triggers the activation of transcription factors NFΚB1, IRF3, and IRF7, leading to the expression of key cytokines and chemokines. (C) Cytoplasmic DNA detection by ZBP1 results in the activation of transcription factors IRF3 and IRF7, inducing the expression of interferons. (D) Methylated CpG DNA recognition by endosomal TLR9 activates NFΚB1, leading to the expression of pro-inflammatory mediators.
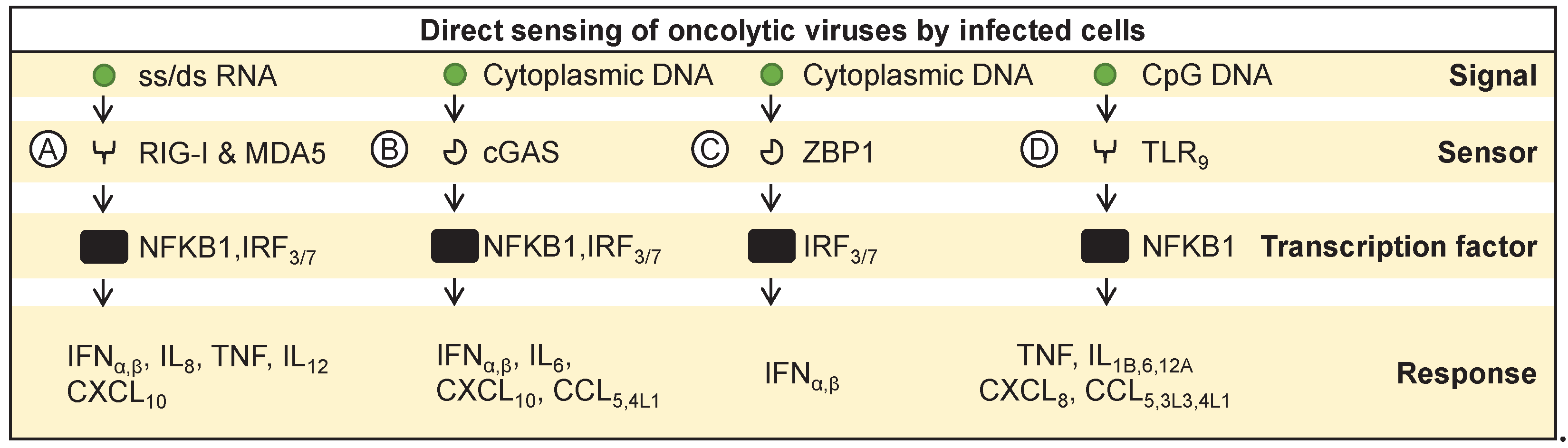
Figure 2.
Sensing of immunostimulatory signals released by infected cells. (A) Interferon-alpha and -beta are recognized by IFNAR receptors, inducing expression of various antiviral genes and antigen presentation. Endocytosed or phagocytosed debris containing viral PAMPs (e.g. double or single-stranded-RNA) activates transcription factors like (B) IRF3 and IRF7 or (C) NFΚB1 in antigen-presenting cells, leading to proinflammatory cytokine production and upregulate antigen presentation. (D) HMGB1, a DAMP, is sensed by TLR4, triggering immune responses.
Figure 2.
Sensing of immunostimulatory signals released by infected cells. (A) Interferon-alpha and -beta are recognized by IFNAR receptors, inducing expression of various antiviral genes and antigen presentation. Endocytosed or phagocytosed debris containing viral PAMPs (e.g. double or single-stranded-RNA) activates transcription factors like (B) IRF3 and IRF7 or (C) NFΚB1 in antigen-presenting cells, leading to proinflammatory cytokine production and upregulate antigen presentation. (D) HMGB1, a DAMP, is sensed by TLR4, triggering immune responses.
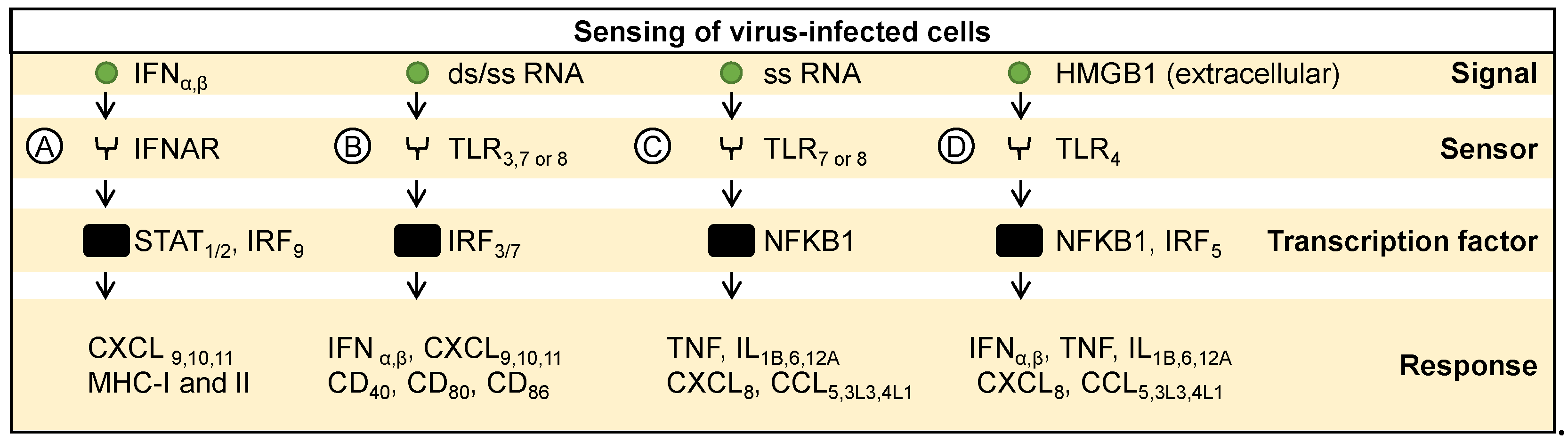
Figure 3.
Non-specific immune activation. (A) Chemokines, recognized by their respective receptors, activate various transcription factors promoting immune cell proliferation, differentiation, and migration. For example, (B) IL2 or IL4 activate STAT5, inducing autocrine and paracrine expression of cytokines and receptors. (C) Similarly, IFN gamma activates STAT1 and STAT4, inducing expression of immune-related genes. Overall, these pathways regulate immune responses against infections and cancer.
Figure 3.
Non-specific immune activation. (A) Chemokines, recognized by their respective receptors, activate various transcription factors promoting immune cell proliferation, differentiation, and migration. For example, (B) IL2 or IL4 activate STAT5, inducing autocrine and paracrine expression of cytokines and receptors. (C) Similarly, IFN gamma activates STAT1 and STAT4, inducing expression of immune-related genes. Overall, these pathways regulate immune responses against infections and cancer.
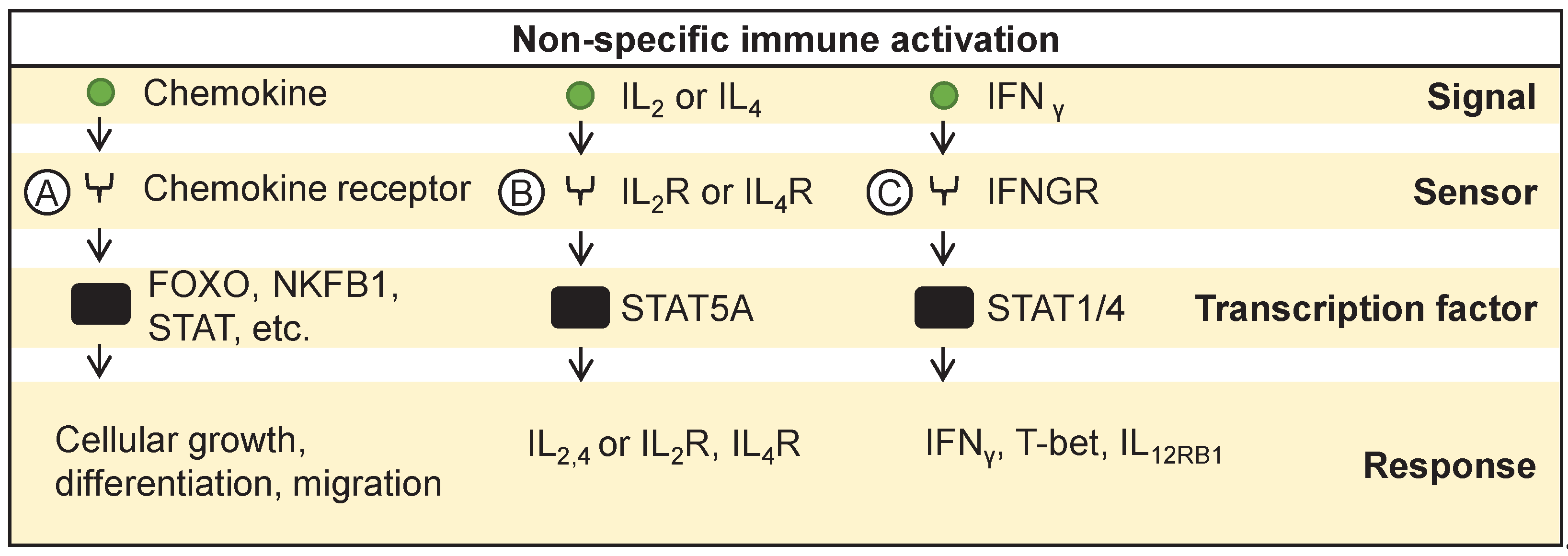
Figure 4.
Target specific immune activation. Antigenic exposure to T cells activates various transcription factors promoting (A) cytotoxic activity towards target cells and (B) release of various proinflammatory molecules promoting antigen presentation and cancer killing. (C) Similarly, NK cells recognize loss or downregulation of MHC molecules on target cells and activate cytotoxic activity towards target cells.
Figure 4.
Target specific immune activation. Antigenic exposure to T cells activates various transcription factors promoting (A) cytotoxic activity towards target cells and (B) release of various proinflammatory molecules promoting antigen presentation and cancer killing. (C) Similarly, NK cells recognize loss or downregulation of MHC molecules on target cells and activate cytotoxic activity towards target cells.
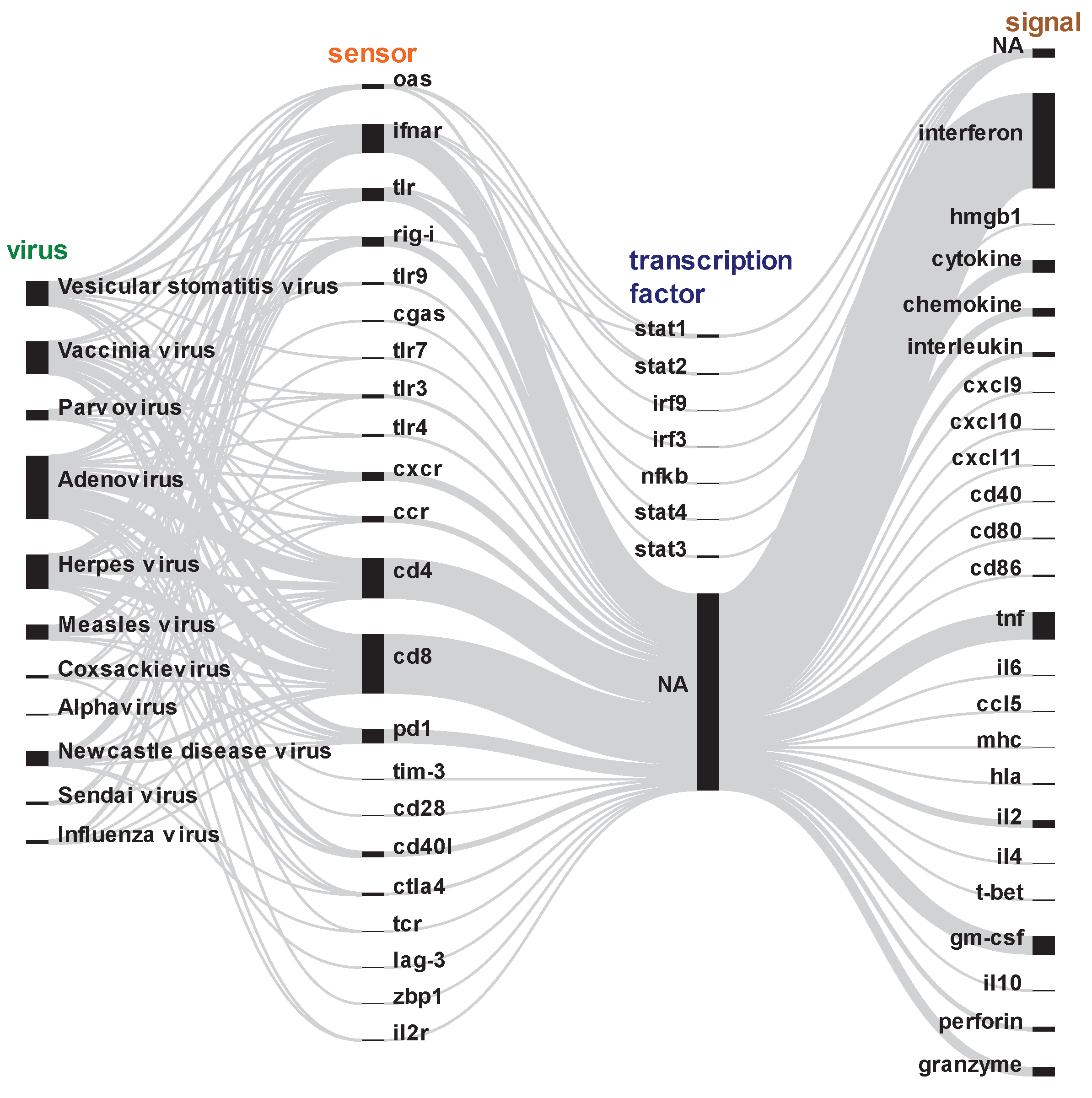
Figure 5.
Literature-based association of virus mechanisms of molecular sensing and immune response to diverse oncolytic viruses. We collected abstracts of scientific articles from PubMed and looked for the presence of molecular evidence on various sensors, transcription factors and immune signals that were associated with different oncolytic virotherapies. In a sequential sense-and-response manner, diverse molecular sensors like TLRs, RIG-I, etc. detect virus genetic material or signals from infected cells to ultimately induce various innate and antigen-specific immune responses. This is regulated by the activation of various transcription factors that induce the expression of immune response genes. NA=not available. Each line in the figure represents connections between different components involved in the molecular sensing and immune response to oncolytic viruses. These connections visually illustrate how various sensors, transcription factors, and immune signals influence each other in initiating and regulating the immune response. For instance, lines can associate the activation of transcription factors by sensors, or the regulation of immune response genes by transcription factors.
Figure 5.
Literature-based association of virus mechanisms of molecular sensing and immune response to diverse oncolytic viruses. We collected abstracts of scientific articles from PubMed and looked for the presence of molecular evidence on various sensors, transcription factors and immune signals that were associated with different oncolytic virotherapies. In a sequential sense-and-response manner, diverse molecular sensors like TLRs, RIG-I, etc. detect virus genetic material or signals from infected cells to ultimately induce various innate and antigen-specific immune responses. This is regulated by the activation of various transcription factors that induce the expression of immune response genes. NA=not available. Each line in the figure represents connections between different components involved in the molecular sensing and immune response to oncolytic viruses. These connections visually illustrate how various sensors, transcription factors, and immune signals influence each other in initiating and regulating the immune response. For instance, lines can associate the activation of transcription factors by sensors, or the regulation of immune response genes by transcription factors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



