
Submitted:
26 March 2024
Posted:
26 March 2024
You are already at the latest version
Abstract
Rice blast and brown planthopper (BPH) are two of the most destructive diseases and pests in rice (Oryza sativa L.) production in China. The development of multi-resistant varieties is widely recognized as the most efficient and environmentally friendly approach to controlling crop diseases and pests. Functional molecular markers (FMMs) have been developed from functional variants in the genic region associated with trait variation, greatly enhancing the efficiency of identifying and pyramiding valuable genes in crop breeding. In this study, two FMMs and a multiplex PCR amplification system were developed for two major broad-spectrum BPH resistance genes, Bph6 and Bph9. By assistance of FMMs in multi-resistant rice restorer line development pipeline, two lines (Huahui7713 and Huahui3006) with blast and BPH resistance were developed by pyramiding three resistance genes Pigm, Bph6 and Bph9. Huahui7713 and Huahui3006 as male lines, two new hybrid rice varieties (WeiLiangYou7713 and Xuanliangyou3006) were certificated and commercial released in China. WeiLiangYou7713 and Xuanliangyou3006 showed improved blast and BPH resistance, and no penalty on yield and grain-quality. Annual promotion area of WeiLiangYou7713 has exceeded 53.3 thousand hectares, indicating its widespread adoption and success in the market. Huahui7713 and Huahui3006 could have great potential for utilization in blast and BPH resistance rice breeding.
Keywords:
blast
; brown planthopper
; resistance gene
; marker-assisted selection
; multiplex PCR
; hybrid rice
1. Introduction
Rice is a vital food crop globally and a staple in China. However, its yield and quality are frequently and severely threatened by various diseases and pests, such as rice blast and brown planthopper (BPH). Rice blast, caused by the filamentous ascomycete fungus Magnaporthe oryzae B.C. Couch (anamorph: Pyricularia oryzae Cavara) is the most devastating rice disease, leading to reduced yields and lower grain quality by damaging leaves, stalks, and spikes. Each year, rice blast causes estimated global yield of 10-30% of the total harvest which is enough to feed 60 million people annually [1]. BPH is a notorious rice pest that feeds on the phloem sap of rice leaf sheaths using needle-like mouthparts, causing wilting, yellowing, and plant death. Moreover, it transmits viral diseases such as rice grassy stunt virus (RGSV), rice ragged stunt virus (RRSV), and rice wilted stunt virus (RWSV), leading to significant yield loss and even complete crop failure [2]. Therefore, developing rice varieties resistant to both pests and diseases is crucial for ensuring food security and enhancing agricultural productivity.
The utilization of resistant germplasm and resistance genes is widely accepted as one of the most efficient approaches to combat crop pests and diseases in crops. To date, more than 40 BPH resistance genes have been identified from cultivated or wild rice germplasm, with 17 of them isolated through map-based cloning [3,4,5]. These genes serve as invaluable resources for developing BPH-resistant rice varieties. As populations of BPH continue to evolve and new BPH biotypes emerge, rice varieties that possess a single BPH resistance gene tend to lose their resistance over time. For example, the BPH-resistant variety IR26, developed in the 1970s by the International Rice Research Institute, experienced a collapse in its resistance just 3 years after widespread cultivation [6]. This loss of resistance was mainly attributed to the emergence of BPH biotype 2. Similarly, other varieties with single BPH resistance genes, such as Bph2 or Bph3, have also experienced a decline in their resistance effectiveness over time. This decline can be attributed to the adaptation of BPH or the emergence of new biotypes [7]. To address this rapid failure in resistance, it is recommended to pyramid BPH resistance genes or QTLs when developing rice varieties. By incorporating multiple resistance genes, these varieties are better equipped to withstand the evolving BPH populations and the emergence of new biotypes, ensuring long-term effectiveness against BPH infestations [8]. A study conducted by Myint et al. [9] found that the pyramided line carrying both Bph25 and Bph26 exhibited significantly higher levels of resistance compared to lines with single resistance genes. Furthermore, the introduction of three BPH resistance genes (Bph14, Bph15, and Bph18) into the elite indica rice variety 93-11 resulted in superior resistance compared to the lines carrying two or single resistance genes [10]. In the case of rice blast disease, more than 100 major resistance genes have been identified and documented on the 12 rice chromosomes, excluding chromosome 3 [11]. Among these genes, at least 39 have been cloned [12,13].
Molecular marker-assisted selection (MAS) is a technique used in rice breeding to help select genotypes based on their genetic composition at the genome level. It offers a fast, efficient, and precise way to combine multiple genes of interest [14]. Effective implementation of MAS relies on the availability of appropriate molecular markers specific to target genes. The decreasing costs of next-generation sequencing technologies have significantly facilitated the acquisition of genome sequence variations. Numerous rice genome sequencing projects have revealed a wide range of single nucleotide polymorphisms (SNPs) distributed throughout the entire genome [15]. SNP marker systems are particularly attractive and effective when SNPs are present in the target gene region between donor and recipient cultivars. Among the available SNP genotyping methods, the amplification refractory mutation system (ARMS) PCR method stands out due to its ability to detect SNPs with minimal laboratory equipment requirements [16]. It allows the identification of different genotypes through a single PCR amplification, making it a simple, reliable and affordable option for rice molecular breeding [17,18,19].
Bph9, located on chromosome 12, is a dominant BPH resistance gene that confers resistance to BPH biotypes 1, 2, and 3. It encodes a rare type of nucleotide-binding and leucine-rich repeat (NLR) gene. Eight genes (Bph1, bph2, bph7, Bph9, Bph10, Bph18, Bph21, and Bph26), previously identified and mapped on the long arm of chromosome 12, are in fact multiple alleles of the same locus, which is designated as BPH1/9. These alleles can be classified into four allelic types based on their sequences: BPH1/9-1 (Bph1/Bph10/Bph18/Bph21), BPH1/9-2 (Bph2/Bph26), BPH1/9-7 (Bph7) and BPH1/9-9 (Bph9) [20]. Bph6, which was mapped to the long arm of rice chromosome 4, was later identified as a gene encoding an exocyst-localized protein. It provides resistance to BPH biotype 4 and biotype 2, which are prevalent in Bangladesh and China, respectively [21]. Wang et al. developed a functional dominant marker for Bph9, and successfully pyramided Bph6 and Bph9 into a 93-11 genetic background using MAS [22]. The resulting improved line, Luoyang69, exhibited enhanced antixenotic and antibiosis effects on BPH compared to the single gene introgression lines. Luoyang69 is a valuable resource for developing rice varieties with durable broad-spectrum high resistance to BPH. However, the availability of specific, precise, and efficient molecular markers for utilizing the Bph6 and Bph9 genes in BPH-resistant rice breeding is still limited. Pigm, a prominent rice blast resistance gene, is located on rice chromosome 6 and consists of multiple NBS-LRR-like disease resistance genes. This gene cluster has been extensively utilized in Chinese rice production [23]. In our previous research, we developed functional markers for the Pigm gene and selected a series of rice blast resistant varieties [24]. The objective of this study is to develop Bph6 and Bph9 functional molecular markers (FMMs) and an efficient detection system for breeding practice. Additionally, we aim to select restorer lines with resistance to both rice blast and BPH, and their derived hybrid combinations, by pyramiding Bph6, Bph9, and Pigm through MAS combined with phenotypic selection (PS).
2. Materials and Methods
2.1. Plant Materials
A total of 478 rice varieties were utilized for the development and validation of FFMs. These varieties included BPH resistance germplasm Mudgo (Bph18/Bph1/Bph10/Bph21), T12 (Bph7), ADR52 (Bph26/Bph2) and Luoyang69 (Bph6 and Bph9), as well as BPH susceptible varieties TN1, Nipponbare and 02428. R6888, a donor for the rice blast resistance gene Pigm, was also used. Two thermo-sensitive genic male sterile (TGMS) lines, Huawei338S and Huaxuan302S, were employed for hybrid rice development. The materials were arranged in a randomized group design and grown under natural conditions at the Guanshan experimental field of Longping Hi-Tech Seed Scientific Research Institute. The experiment consisted of three replications, with each plot containing four rows of eight plants. The spaceing between plants within and between the rows was 16.5 cm. For newly developed restorer lines, main agronomic traits were measured, including grain yield (GY), days to heading (DTH), plant height (PH), number of grains per panicle (NGP), panicle length (PL), spikelet fertility (SF), thousand-grain weight (TGW), and rice quality traits such as apparent amylose content (AAC), gel consistency (GC), gelatinization temperature (GT), chalkiness degree (CD), head rice rate (HRR), and length-width ratio (LWR). These measurements were taken for the five plants in the middle of the central row of each plot.
2.2. Primer Design of the ARMS Markers of Bph6 and Bph9
The genome sequences of Bph6, Bph9, Bph7, Bph18, and Bph26 were obtained from the NCBI (www.ncbi.nlm.nih.gov/) database (accession numbers KX818198, KU216221, KU221258, KJ850252, and AB910360, respectively). Additionally, the corresponding genomic sequences of Bph6 and Bph9 in the varieties Nipponbare and 93-11 were also obtained. Sequences alignment and analysis of polymorphic sites within genic regions were performed using DNAman software. Based on ARMS-PCR, two strategies were employed for designing the markers. Strategy A involved using two allelic sequences of a polymorphic site as the 3' ends of both the forward and reverse inner primers. Two outer primers were designed to pair with the inner primers, forming a four-primer marker system. The genotyping of samples was done by detecting the fragment size of the amplification products of the marker system. Strategy B focused on designing two pairs of dominant markers to specifically amplify the two alleles, targeting two co-separated polymorphic sites. These two pairs of markers were then used in combination to identify different genotypes in the same amplification system (Figure 1).
2.3. Evaluation of BPH and Blast Resistance
BPH resistance was assessed using the modified bulked seedling test [22], with Luoyang69 and TN1 serving as the resistant and susceptible controls, respectively. The resistance level of a seedling was determined by the resistance score (RS = 0, 1, 3, 5, 7, or 9), and the resistance level of a line was determined by the average RS of 16 seedlings. Rice blast resistance was evaluated following the method described by Qin et al. [24]. For leaf blast resistance, a scoring system ranging from 0 to 9 was utilized. Scores of 0 to 3 indicated a resistant reaction, 4 to 5 were considered moderately resistant, 6 indicated moderate susceptibility, and scores of 7 to 9 represented susceptibility.
2.4. MAS of New Hybrid Rice Restorer Lines with High-Level BPH and Blast Resistance
In order to pyramid rice blast and BPH resistance genes into the same rice variety, R6888 (Pigm) and Luoyang69 (Bph6 and Bph9) were crossed. The resulting F1 plants were self-crossed to obtain the F2 segregating population. We genotyped the F2 population for Pigm, Bph6 and Bph9 using the functional marker GM [24] and the newly developed markers in this study. Additionally, we conducted phenotypic evaluations of agronomic traits to further screen the population. When the population reached the advanced generation F6, lines were evaluated for BPH and blast resistance, as well as for main agronomic and grain quality traits. The genetically stable and best performing lines were then crossed with TGMS lines to produce hybrids.
2.5. Evaluation of Main Agronomic and Grain Quality Traits
To evaluate the F1 hybrid combinations, newly selected rice restorer lines were crossed with commonly used sterile lines Huawei338S and Huaxuan302S. The multi-location evaluation was conducted by the Research and Development Center of Yuan Longping High-Tech Agriculture Company, following the guidelines provided by the National Committee of Crop Variety Certification (http://www.china riceinfo.com/).
3. Results
3.1. Development and Validation of Molecular Markers for Bph6 and Bph9
Sequence alignment revealed the presence of two polymorphic regions within Bph6 (Bph6-SNP and Bph6-MNP) and three within Bph9 (Bph9-MNP1, Bph9-MNP2 and Bph9-MNP3) (Figure 1a). Two strategies were employed to develop PCR primers targeting these polymorphic regions through ARMS technology (Figure 1b). In strategy A, a set of tetra-primers (B9) was designed to target Bph9-MNP1. Amplification of the Bph9 allele resulted in a 230-bp band, while the bph9 allele produced a 316-bp band (Table1, Figure 2b,e). In strategy B, two polymorphic sites, Bph6-SNP and Bph6-MNP, were used as targets for the developing of a set of tetra-primers (B6). B6 enabled the amplification of the Bph6 allele, generating a 283-bp band, while the bph6 allele produced a 361-bp band (Figure 2a,d). Additionally, Strategy B was also used to develop tetra-primers (B1/9) for BPH1/9, targeting the polymorphic Bph9-MNP2 and Bph9-MNP3. Primers B1/9-RR and B1/9-RF were capable of specifically amplifying BPH1/9 resistant alleles (Bph7, Bph9, Bph18 and Bph26), generating a 545-bp band (Figure 2c,f). The tetra-primers B1/9 and B6 can be used for multiplex PCR to simultaneously detect Bph6 and Bph9 genotypes (Figure 2g). These tetra-primers are applicable for Bph6 and Bph9 genotyping in breeding programs.

Table 1.
Primers for Bph6, Bph9 and Pigm.
| Target gene | Marker name | Primer name | Sequence (5'-3') | Product size (bp) |
|---|---|---|---|---|
| Bph6 | B6 | SF | AGGGCCTCTGGCGCTCTAC | 361(-) / 283(+) |
| SR | AATGTGAAAGTGCAATTAGAAGGT | |||
| RF | ATAGTGAAGTTGAATCCGAAGG | |||
| RR | AGTGACTCAGCCTTGTGTTTCG | |||
| Bph9 | B9 | IF | ACCATTGTTAGGCAGTTGTTCA | 316(-) / 230(+) |
| OR | ATTCGACTCCCTTTCTTGTTATCT | |||
| OF | AATGTCGCACCCAGCAGC | |||
| IR | CAGCCTCCTGAAGAGATCTTTCA | |||
| BPH1/9 | B1/9 | SF | GGCGAGTTGATGAAATAAAGG | 473(-) / 545(+) |
| SR | GTTCTTTGCATCGCTGTCTC | |||
| RR | TTGCTCCCACCGAAGTCA | |||
| RF | AGGGAATGGTGTAGTAGCA | |||
| Pigm | GM | OF | TGCTGAACAAGGTGTAGAGGTA | 448(-) / 621(+) |
| OR | GCCAGGTCCTACTTCCACAAAA | |||
| IF | GTGACATCCAGTCCTACACAATCTAA | |||
| IR | CACGGAACTCGTTTTCGTC |
“+” indicates the size of the PCR product of the resistant allele; “-” indicates the size of the PCR product of the susceptible allele; The underlined primers B1/9-RR and B1/9-RF can be used as universal dominant markers for Bph7, Bph9, Bph18 and Bph26.
We conducted a comprehensive analysis of 478 core rice varieties using newly developed markers. Surprisingly, we found that only Luoyang69 possessed the Bph6 and Bph9 genotypes, while the other varieties did not have these genotypes. Furthermore, by utilizing the B1/9 dominant marker B1/9-RF/B1/9-RR, we identified 115 out of the 478 rice varieties that carried BPH1/9 resistant alleles (Bph7, Bph9, Bph18, and Bph26). Through further re-sequencing analysis, we determined that all 115 rice cultivars carried one of the four resistance genes, with Bph18 being the most prevalent (71), followed by Bph26 (42), and only one instance each of Bph7 and Bph9 (Supplementary Table S1). These findings align with the widespread use of Bph1 and Bph2 in rice breeding programs.
3.2. Development of New Restorer Lines with BPH and Blast Resistance by MAS
Luoyang69, a BPH resistance improved line, incorporates the Bph6 and Bph9 gene into the background of 93-11, an elite indica restorer line of two-line hybrid rice in China. R6888, harboring Pigm gene, is an elite indica restorer line of two-line hybrid rice developed by Yuan Longping High-Tech Agriculture Co., Ltd.. A cross (Luoyang69/R6888) was generated to produce an F2 population. Out of approximately 2400 F2 plants, a subset of 560 plants exhibiting homozygous resistance alleles for Pigm, Bph6 and Bph9 by MAS were specifically selected for transplanting. The plants with superior performance in terms of yield and ideotype were harvested for underwent subsequent selfing, enabling further detailed observation of agronomic characteristics, yield potential, resistance and grain quality. By the F6 generation, 24 outstanding new lines exhibiting enhanced resistance against blast and BPH, along with superior agronomic traits, were selected for test-crossing trials. These lines were categorized into three groups: Gm69, Gm6 and Gm9. Gm69 group consisted of 6 lines that were pyramided with Bph6, Bph9, and Pigm. Gm6 group consisted of 8 lines pyramided with Bph6 and Pigm, while Gm9 group consisted of 10 lines pyramided with Bph9 and Pigm (Table 2). The average seedling blast resistance score for Gm69, Gm6 and Gm9 group lines was 3.76, 3.62 and 3.43, respectively, which was significantly higher than that of the parental Luoyang69 with a score of 8.23. Similarly, the average seedling BPH resistance score for Gm69, Gm6 and Gm9 group lines was 2.75, 3.56 and 3.26, respectively, showing significant enhancement compared to the parental R6888 with a score of 8.20. Furthermore, the BPH resistance of Gm69 group lines were significantly higher than that of Gm6 and Gm9 group lines, indicating that pyramiding Bph6 and Bph9 can effectively enhances BPH resistance than pyramiding single Bph6 or Bph9 gene. After test-crossing trials on these lines, two elite Gm69 lines, namely Huahui3006 and Huahui7713, demonstrated the highest combining ability and heterosis, displaying outstanding agronomic performance in hybrids (Table 3, Figure 3). Both Huahui3006 and Huahui7713 carry Pigm, Bph6 and Bph9, and exhibit high-level resistance to both blast disease and BPH. In contrast, the parental lines, Luoyang69 and R6888, were highly susceptible to blast disease and BPH, respectively (Table 4, Figure 4).
3.3. New Hybrid Rice Varieties Development with Blast and BPH Resistance
Huahui7713 and Huahui3006 were crossed with the elite sterile lines Huawei338S and Huaxuan302S, respectively, which were newly developed by Yuan Longping High-Tech Agriculture Co., Ltd.. The derived hybrid rice varieties, named as Weiliangyou7713 and Xuanliangyou3006, were evaluated in the state’s regional trials for new variety of middle-season indica in middle and lower reaches of Yangtze River in 2019 and 2020, respectively. In the state’s regional trials, both Weiliangyou7713 and Xuanliangyou3006 showed significantly improved resistance to rice blast and BPH. Additionally, both hybrid rice varieties displayed superior yield performance compared to the control variety. Notably, Weiliangyou7713 showed a remarkable yield increase of 7.52% (Table 5). Furthermore, both varieties showed good rice quality with second-class grain quality according to the standard of superior quality rice industry in China (NY/T593-2013). Due to the outstanding performance, Weiliangyou7713 and Xuanliangyou3006 got new variety certifications in 2021 and 2022, respectively. Additionally, Weiliangyou7713 was recognized as super-rice by Ministry of Agriculture and Rural Affairs of China (MARAC) in 2023, is also the first super-rice conferring both blast and BPH resistance. Weiliangyou7713 achieved an average yield of 16.26 tons/ha and 15.55 tons/ha in demonstration plot trials over 66.67-hectare in Hunan province in 2021 and 2022, respectively. By 2023, the planting area of Weiliangyou7713 had surpassed 53.3 thousand hectares.
4. Discussion
Developing rice varieties with broad-spectrum or persistent resistance has been a significant goal for breeders. However, it has been revealed that BPH-resistance genes, like disease resistance genes, encode immune receptors to perceive the insect effector [25]. It is commonly observed that one resistance gene provides resistance to a specific pathogenic microbe or insect biotype [26]. As pathogenic microbes mutate or insect biotypes evolve, the resistance of rice varieties carrying a single resistance gene may diminish over time. Therefore, breeders are actively seeking ways to reduce selection pressure on specific pathogens or insects. One approach is to utilize broad-spectrum resistance or pyramiding multiple resistance genes, which can reduce the frequency of pathogen variation or insect evolution. The ultimate aim is to develop rice varieties with durable and stable resistance. In recent years, most studies on improving rice resistance through MAS have focused on either diseases or insects, with fewer studies targeting both disease and insect resistance [27,28,29,30]. There are also limited reports on the commercial-scale promotion of varieties resistant to both rice blast and BPH. In this study, we successfully pyramided the BPH resistance genes Bph6, Bph9, and the broad-spectrum rice blast resistance gene Pigm, resulting in the development of two new restorer lines with high-level resistance to both blast disease and BPH. Their derived hybrid combinations also exhibit resistance to rice blast and BPH, as well as excellent performance in terms of yield and rice quality. Up to now, two derived new hybrid rice varieties (Weiliangyou7713 and Xuanliangyou3006) have been certificated and commercial released in China. Notably, both hybrid rice varieties demonstrated enhanced resistance against rice blast and BPH, without compromising yield and grain quality. By 2023, the planting area of super-rice Weiliangyou7713 had surpassed 53.3 thousand hectares, which will significantly reduce the utilization of fungicides and pesticides, and play a critical role in preventing and controlling the outbreak of rice blast and BPH. This provides strong evidence that the utilization of Huahui7713 and Huahui3006 is highly effective in rice breeding programs aimed at improving resistance against blast and BPH. The Bph6 and Bph9 genes, found in tropical rice cultivars, have less undesirable linkage drag and are more likely to be effectively used in rice breeding. These genes are dominant, which means they provide a higher level of resistance in heterozygous genotypes, making them valuable for hybrid rice breeding. However, our analysis of 478 rice varieties suggested that Bph6 and Bph9 have not been utilized in breeding. To control BPH damage, it is important to accelerate the application of Bph6 and Bph9 in breeding and pyramid them to develop BPH-resistant rice varieties. Routine breeding method for pyramiding multiple genes based on phenotype evaluation of insect resistance is impracticable due to a huge amount of work and difficulties in distinguishing gene effects. In our study, the combination of MAS with conventional breeding methods has successfully pyramided two BPH-resistance genes along with a blast-resistance gene into the high yield and superior-quality background. Although molecular marker technology has been extensively used in rice breeding practice, breeders are still primarily concerned with improving the accuracy and efficiency of molecular markers and reducing the cost of detection. One approach to achieve this is through the development of functional markers or gene-specific molecular markers, which are based on gene-specific sites. Here we developed functional molecular markers for Bph6 and Bph9. These markers are capable to distinguish the allele variation of the selected genes, and consequently enhance the accuracy and efficiency of MAS [31,32,33]. Another strategy to improve the efficiency of molecular marker detection is the utilization of multiplex PCR technology. This technology allows for the simultaneous identification of multiple markers in a single PCR amplification system, and is therefore an important avenue to enhance the efficiency of molecular marker detection [34,35,36]. In this experiment, ARMS-PCR was applied for detecting SNPs that is based on allele-specific amplification PCR. The conventional strategy, known as strategy A, involves designing four primers with a single site as the target. However, this approach has limitations as both the upstream and downstream sequences of the target site need to satisfy the primer design, resulting in a limited success rate. In contrast, strategy B offers better flexibility and a higher success rate by using dual targets to design allele-specific amplification primers separately. Additionally, strategy B allows for the design of amplification products of different sizes for multiple sites of different genes, making it relatively easy to achieve multiplex PCR amplification and improve detection efficiency. In this study, we obtained several representative variations of Bph6 and Bph9, but most of them did not meet the requirements for primer design. Among the selected representative variations, we designed gene-specific markers for Bph9 and Bph6 based on strategy A and strategy B, respectively. These markers can accurately identify rice varieties carrying Bph6 or Bph9, making them valuable for screening BPH-resistant germplasm resources. Additionally, we developed a multiplex PCR amplification genotyping system for Bph6 and Bph9, which allows for simultaneous detection of their genotypes through 1.5-3% agarose gel electrophoresis. This system is fast, simple, and cost-effective, and will contributes to pyramiding multiple genes and breeding varieties of high yield, superior quality and durable resistance.
5. Conclusions
The development of multi-resistant varieties is widely recognized as an efficient and environmentally friendly approach to controlling crop diseases and pests. In this study, we reported the development of two functional molecular markers for two major BPH resistance genes (Bph6 and Bph9), which can be used in molecular design breeding. Two new elite male lines (Huahui7713 and Huahui3006) for two-line hybrid rice were also developed by pyramiding Bph6, Bph9 and major blast resistance gene Pigm. Huahui7713 and Huahui3006 exhibit enhanced resistance to both BPH and blast, along with high-yielding and superior quality performance. Furthermore, we have successfully developed two Huahui7713 and Huahui3006 derived two-line hybrid rice varieties (Weiliangyou7713 and Xuanliangyou3006) with BPH and blast resistance. These hybrids performed excellently in state-wide trials in China, have been approved for commercial release and a gained acceptance in the market with an annual promotion area of 53.3 thousand hectares in the three years since their releasing. Notably, these varieties provide the dual benefits of high yield and improved resistance to both BPH and blast, thereby potentially minimizing the use of pesticides and fungicides in rice production. This study exemplifies the practical application of molecular design in rice breeding programs, particularly for enhancing disease and pest resistance. The developed markers and breeding strategies provide valuable tools for future BPH and blast resistance breeding in rice, and the commercial release of resistance varieties can contribute significantly to ensuring food security.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Y.Y. and K.W. conceived and designed experiments. Z.D. and P.Q. contributed to data analysis and the first draft of the manuscript. K.L. and N.J. and contributed to data collection. T.Y. and X.Z. were in charge of filed management. K.W., Y.Y., C.F. and G.H. supervised and complemented the manuscript writing. All authors read and approved the final manuscript.
Funding
This research was funded by the Science and Technology Innovation Program of Hunan (2023NK1010, 2022NK1010), Shenzhen Science and Technology Project (202320D314).
Data Availability Statement
The data that support this study are available upon reasonable request from the corresponding author.
Acknowledgments
Thank you to all those who helped with this study and to the research projects that sponsored it.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Skamnioti, P.; Gurr, S.J. Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27(3), 141–150. [Google Scholar] [CrossRef] [PubMed]
- Dyck, V.A.; Thomas, B. The brown planthopper problem, inBrown Planthopper: Threat to Rice Production in Asia. International Rice Research Institute 1979, Laguna, Philippines, 1. [Google Scholar]
- Cheng, X.; Zhu, L.; He, G. Towards understanding of molecular interactions between rice and the brown planthopper. Mol. Plant 2013, 6(3), 621–634. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Zhao, Y.; Du, B.; Chen, R.; Zhu, L.; He, G. Genomics of interaction between the brown planthopper and rice. Curr Opin Insect Sci. 2017, 19, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, H.; Nie, L.; Tan, D.; Zhou, C.; Zhang, Q.; Li, Y.; Du, B.; Guo, J.; Huang, J.; Wu, D.; Zheng, X.; Guan, W.; Shan, J.; Zhu, L.; Chen, R.; Xue, L.; Walling, L.; He, G. Bph30 confers resistance to brown planthopper by fortifying sclerenchyma in rice leaf sheaths. Mol. Plant 2021, 14(10), 1714–1732. [Google Scholar] [CrossRef]
- Khush, G.S. Breeding for resistance in rice. Ann Ny Acad Sci. 1977, 296, 308. [Google Scholar] [CrossRef]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breeding 2007, 19, 35–44. [Google Scholar] [CrossRef]
- Cohen, M.B; Alam, S.N.; Medina, E.B.; Bernal, C.C. Brown planthopper, Nilaparvata lugens, resistance in rice cultivar IR64: mechanism and role in successful N. lugens management in Central Luzon. Philippines. lugens management in Central Luzon. Philippines. Entomol. Exp. Appl. 1997, 85(3), 221–9. [Google Scholar] [CrossRef]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 2012, 124(3), 495–504. [Google Scholar] [CrossRef]
- Hu, J.; Cheng, M.; Gao, G.; Zhang, Q.; Xiao, J.; He, Y. Pyramiding and evaluation of three dominant brown planthopper resistance genes in the elite indica rice 9311 and its hybrids. Pest Manag. Sci. 2013, 69(7), 802–808. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, P.K.; Arya, M.; et al. Molecular screening of blast resistance genes in rice using SSR markers. The Plant Pathology Journal 2015, 31(1), 12. [Google Scholar] [CrossRef]
- Devanna, B.N.; Jain, P.; Solanke, A.U.; et al. Understanding the dynamics of blast resistance in rice-Magnaporthe oryzae interactions. J. Fungi 2022, 8(6), 584. [Google Scholar] [CrossRef]
- Sugihara, Y.; Abe, Y.; Takagi, H.; et al. Disentangling the complex gene interaction networks between rice and the blast fungus identifies a new pathogen effector. Plos Biol. 2023, 21(1), e 3001945. [CrossRef]
- Nogoy, F.M.; Song, J.Y.; Ouk, S.; Rahimi, S.; Kwon, S.W.; Kang, K.K.; Cho, Y.G. Current applicable DNA markers for marker assisted breeding in abiotic and biotic stress tolerance in rice (Oryza sativa L.). Plant Breeding and Biotechnology, 2016, 4(3), 271-284. [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; Xing, Y.; Chen, L.; Xie, W. RiceVarMap: a comprehensive database of rice genomic variations. Nucleic Acids Res. 2015, 43(D1), D1018–D1022. [Google Scholar] [CrossRef]
- Ye, S.; Dhillon, S.; Ke, X.; Collins, A.R.; Day, I.N.M. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Res. 2001, 29(17), e88–e88. [Google Scholar] [CrossRef]
- Ramkumar, G.; Sivaranjani, A.K.P.; Pandey, M.K.; Sakthivel, K.; Rani, N.S.; Sudarshan, I.; Prasad, G.S.V.; Neeraja, C.N.; Sundaram, R.M.; Viraktamath, B.C.; Madhav, M.S. Development of a PCR-based SNP marker system for effective selection of kernel length and kernel elongation in rice. Mol. Breeding 2010, 26(4), 735–740. [Google Scholar] [CrossRef]
- Ramkumar, G.; Prahalada, G.D.; Hechanova, S.L.; Vinarao, R.; Jena, K.K. Development and validation of SNP-based functional co-dominant markers for two major disease resistance genes in rice (O. sativa L.). Mol. Breeding. [CrossRef]
- Liu, Y.; Zhang, A.; Wang, F.; Wang, J.; Bi, J.; Kong, D.; Zhang, F.; Luo, L.; Liu, G.; Yu, X. Development and validation of a PCR-based functional marker system for identifying the low amylose content-associated gene Wxhp in rice. Breeding Sci. 2019, 69(4), 702–706. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Wang, Z.; Jing, S.; Wang, Y.; Ouyang, Y.; Cai, B.; Xin, X.; Liu, X.; Zhang, C.; Pan, Y.; Ma, Y.; Li, Q.; Jiang, W.; Zeng, Y.; Shangguan, X.; Wang, H.; Du, B.; Zhu, L.; Xu, X.; Feng, Y.; He, S.; Chen, R.; Zhang, Q.; He, G. Allelic diversity in an NLR gene BPH9 enables rice to combat planthopper variation. PNAS 2016, 113(45), 12850–12855. [Google Scholar] [CrossRef]
- Guo, J.; Xu, C.; Wu, D.; et al. Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice. Net. Genet. 2018, 50(2), 297–306. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Liu, H.; Zeng, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Marker assisted pyramiding of Bph6 and Bph9 into elite restorer line 93-11 and development of functional marker for Bph9. Rice 2017, 10(1), 1–13. [Google Scholar] [CrossRef]
- Xing, J.; Deng, H.; Yuan, L. The Utilization of Rice Blast Resistance Genes in Hybrid Rice Breeding in China. Protecting Rice Grains in the Post-Genomic Era. ntechOpen 2019. [CrossRef]
- Qin, P.; Jiang, N.; Hu, X.; Zhu, Y.; Deng, Z.; Bai, Z.; Fu, J.; Zhang, X.; Wang, K.; Chen, S.; Yang, Y. Breeding a series of new rice restorer lines with blast resistance and low-to-intermediate apparent amylose contents by integrating marker-assisted and phenotypic selection. J. Phytopathol. 2021, 169(11-12), 667-677. [CrossRef]
- Guo, J.; Wang, H.; Guan, W.; Guo, Q.; Wang, J.; Yang, J.; Peng, Y.; Shan, J.; Gao, M.; Shi, S.; Shangguan, X.; Liu, B.; Jing, S.; Zhang, J.; Xu, C.; Huang, J.; Rao, W.; Zheng, X.; Wu, D.; Zhou, C.; Du, B.; Chen, R.; Zhu, L.; et al. A tripartite rheostat controls self-regulated host plant resistance to insects. Nature 2023, 618, 799–807. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M. Current status of brown planthopper (BPH) resistance and genetics. Rice 2010, 3(2), 161–171. [Google Scholar] [CrossRef]
- Hu. J.; Li, X.; Wu, C.; Yang, C.; Hua, H.; Gao, G.; Xiao, J.; He, Y. Pyramiding and evaluation of the brown planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Breeding 2012, 29(1), 61-69. [CrossRef]
- Jiang, H.; Feng, Y.; Bao, L.; Li, X.; Gao, G.; Zhang, Q.; He, Y. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol. Breeding 2012, 30(4), 1679–1688. [Google Scholar] [CrossRef]
- Wang, H.; Ye, S.; Mou, T. Molecular breeding of rice restorer lines and hybrids for brown planthopper (BPH) resistance using the Bph14 and Bph15 genes. Rice 2016, 9(1), 53. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, T.; Zhang, A.; Ong, K.; Luo, Z.; Li, Z.; Yang, J.; Yin, Z. Marker-assisted breeding of Chinese elite rice cultivar 9311 for disease resistance to rice blast and bacterial blight and tolerance to submergence. Mol. Breeding 2017, 37(8), 106. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, Z.; Lang, X.; Du, B.; Liu, K.; Yang, G.; Hu, G.; Li, S.; He, G.; You, A. Development and validation of a PCR-based functional marker system for the brown planthopper resistance gene Bph14 in rice. Breeding Sci. 2013, 63(3), 347–352. [Google Scholar] [CrossRef]
- Tian, D.; Chen, Z.; Chen, Z.; Zhou, Y.; Wang, Z.; Wang, F.; Chen, S. Allele-specific marker-based assessment revealed that the rice blast resistance genes Pi2 and Pi9 have not been widely deployed in Chinese indica rice cultivars. Rice 2016, 9(1), 1–11. [Google Scholar] [CrossRef]
- Pranathi, K.; Viraktamath, B.C.; Neeraja, C.N.; et al. Development and validation of candidate gene-specific markers for the major fertility restorer genes, Rf4 and Rf3 in rice. Mol. Breeding 2016, 36(10), 1–14. [Google Scholar] [CrossRef]
- Yap, R.; Hsu, Y.; Wu, Y.; Lin, Y.; Kuo, C. Multiplex PCR genotyping for five bacterial blight resistance genes applied to marker-assisted selection in rice (Oryza sativa). Plant Breeding 2016, 135(3), 309–317. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Li, M.; Zhao, W.; Sun, H.; Zhao, Q. A multiplex PCR system for detection of wide compatibility allele S5-n and erect panicle allele dep1 in rice. Crop Breed. Appl. Biot. 2017, 17, 250–258. [Google Scholar] [CrossRef]
- Hayashi, K.; Yoshida, H.; Ashikawa, I. Development of PCR-based allele-specific and InDel marker sets for nine rice blast resistance genes. Theor. Appl. Genet. 2006, 113(2), 251–260. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Development of molecular markers targeting Bph6 and Bph9 genes. Identification of nucleotide polymorphisms in Bph6 and Bph9 (a). Schematic illustration of two strategies for ARMS primers design for Bph6 and Bph9 (b).
Figure 1.
Development of molecular markers targeting Bph6 and Bph9 genes. Identification of nucleotide polymorphisms in Bph6 and Bph9 (a). Schematic illustration of two strategies for ARMS primers design for Bph6 and Bph9 (b).
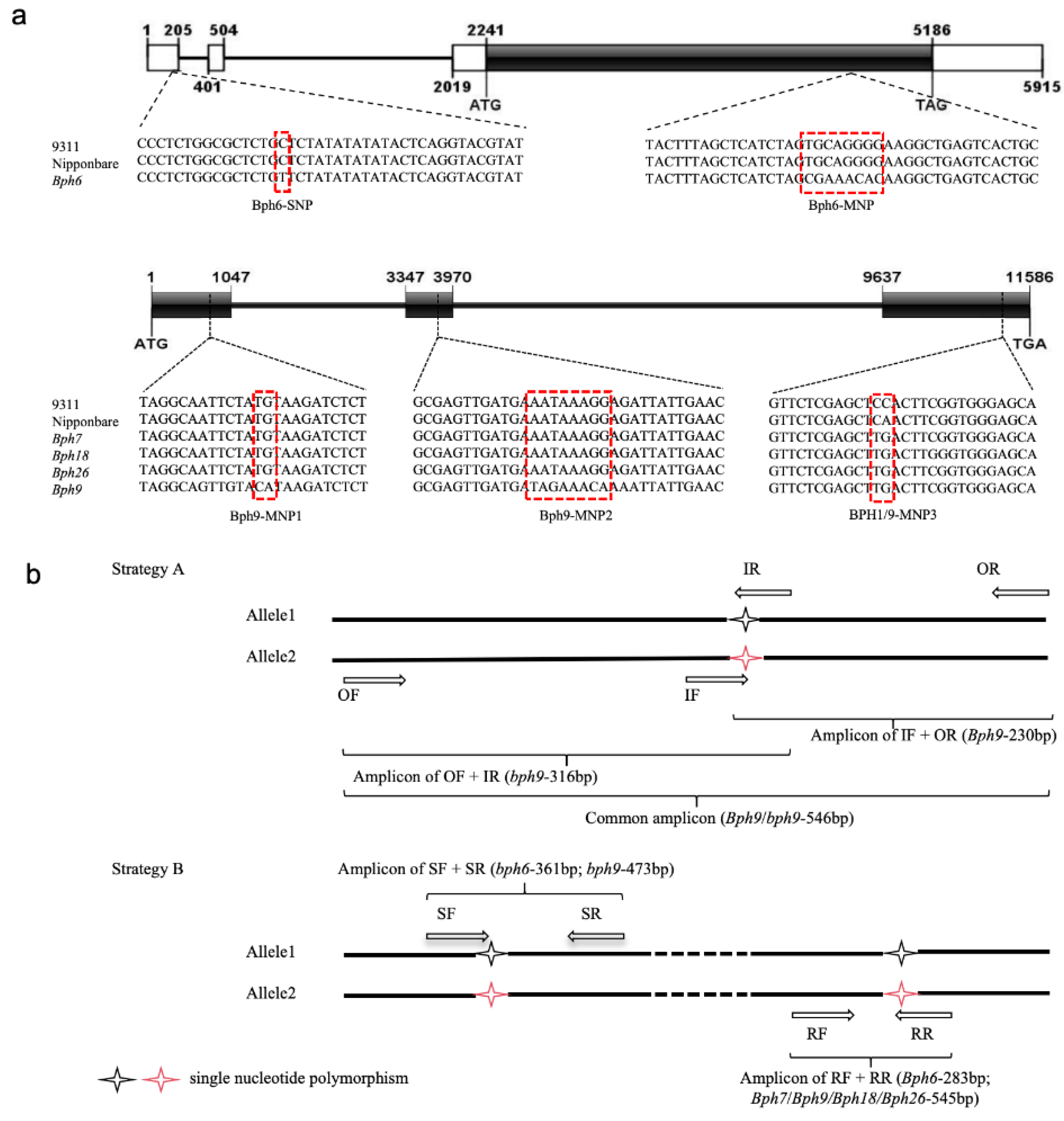
Figure 2.
Validation of the developed markers. Validation of markers B6 (a), B9 (b) and B1/9 (c) with rice varieties. M: DL1000 DNA marker, lane1-16: Luoyang69, Mudgo, T12, ADR52, 9311, Nipponbare, Huazhan, 02428, IRTA129, Huabao, Qiuguang, Chujing28, Chenghui19, Mianhui725, Nongxiang39, Fudao88. Validation of markers B6 (d), B9 (e) and B1/9 (f) using segregating population. lane1-16: individual plants from the F2 of R6888/Luoyang69. g, amplification pattern of Bph6 and Bph9 multiplex PCR amplification system in F2 population, “+” indicates carrying Bph6 or Bph9, “-” indicates without Bph6 or Bph9, H indicates heterozygous genotype.
Figure 2.
Validation of the developed markers. Validation of markers B6 (a), B9 (b) and B1/9 (c) with rice varieties. M: DL1000 DNA marker, lane1-16: Luoyang69, Mudgo, T12, ADR52, 9311, Nipponbare, Huazhan, 02428, IRTA129, Huabao, Qiuguang, Chujing28, Chenghui19, Mianhui725, Nongxiang39, Fudao88. Validation of markers B6 (d), B9 (e) and B1/9 (f) using segregating population. lane1-16: individual plants from the F2 of R6888/Luoyang69. g, amplification pattern of Bph6 and Bph9 multiplex PCR amplification system in F2 population, “+” indicates carrying Bph6 or Bph9, “-” indicates without Bph6 or Bph9, H indicates heterozygous genotype.
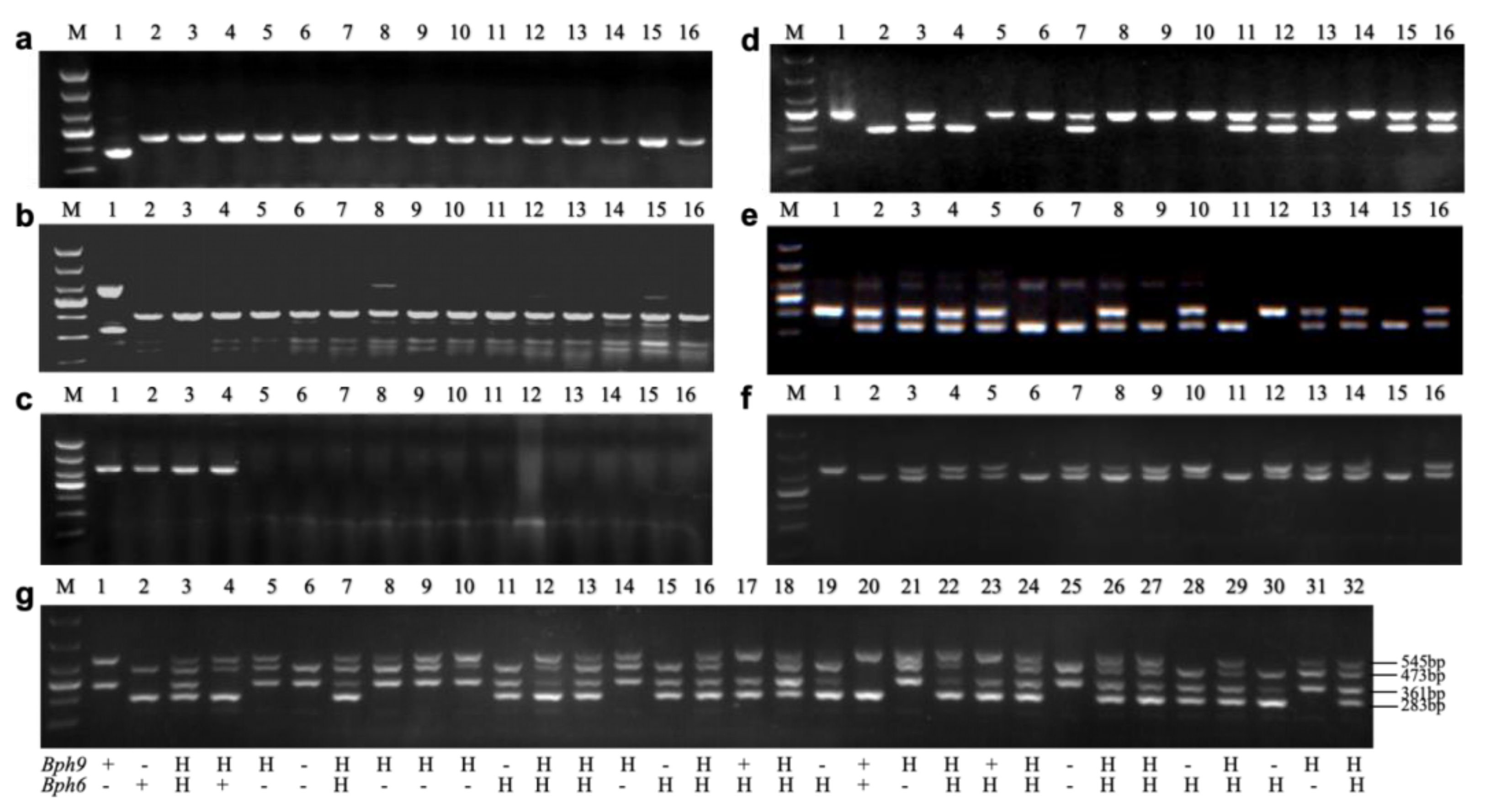
Figure 3.
The breeding process of new restorer lines and hybrids with improved blast and BPH resistance.
Figure 3.
The breeding process of new restorer lines and hybrids with improved blast and BPH resistance.
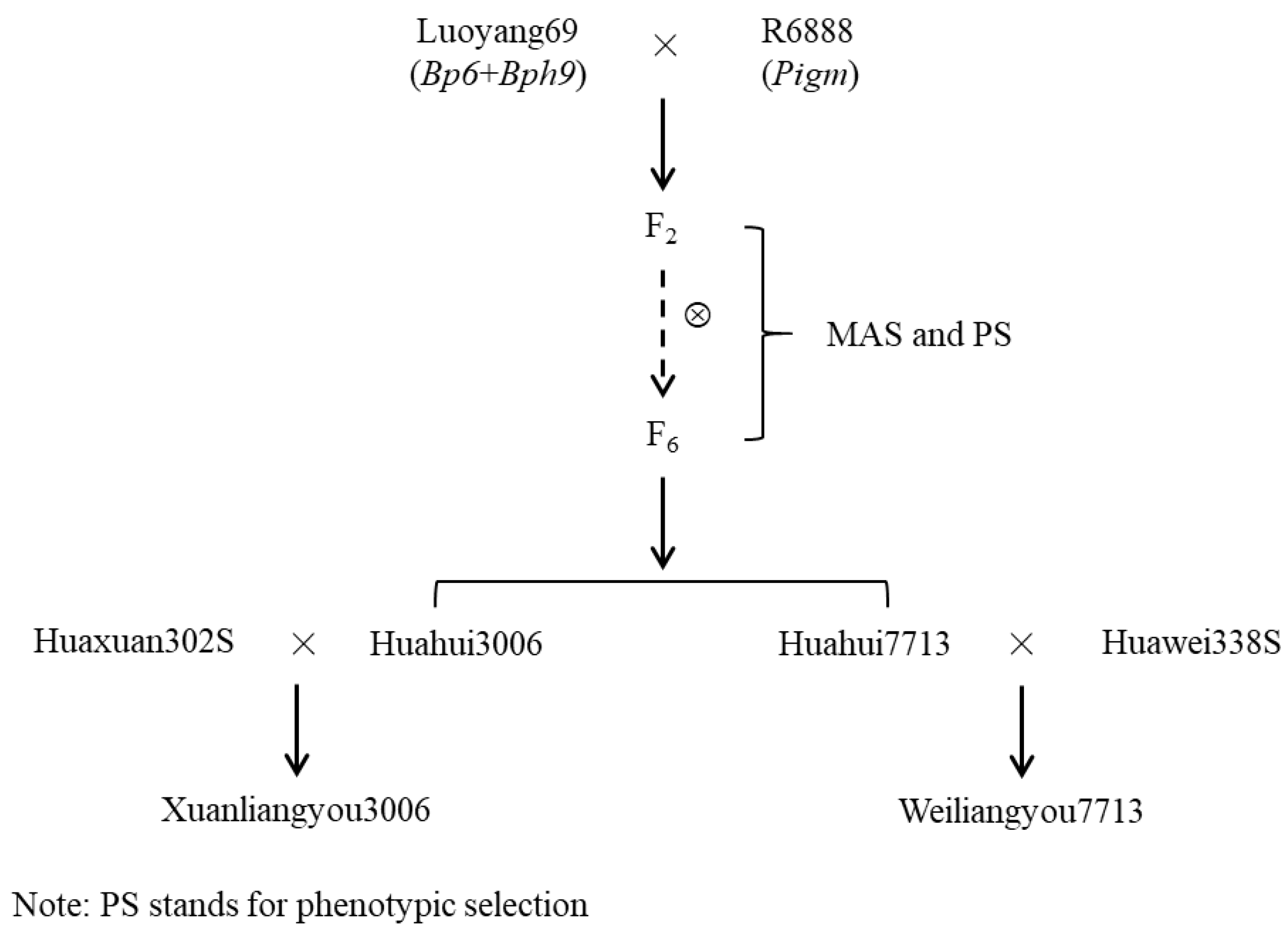
Figure 4.
The BPH and blast resistance performance of improved restorer lines and their hybrid combinations. Performance of new restorer lines for resistance to BPH (a) and rice blast (b) at seedling stage. BPH resistance performance of hybrid combinations derived from Xuanliangyou3006 (c) and Weiliangyou7713 (d).
Figure 4.
The BPH and blast resistance performance of improved restorer lines and their hybrid combinations. Performance of new restorer lines for resistance to BPH (a) and rice blast (b) at seedling stage. BPH resistance performance of hybrid combinations derived from Xuanliangyou3006 (c) and Weiliangyou7713 (d).
Table 2.
Resistance analysis of pyramiding lines and their parents to rice blast and BPH.
| Lines | Genes of pyramiding | Number of Lines | Seedling BPH resistance score | Seedling blast resistance score |
|---|---|---|---|---|
| Gm69 lines | Bph6+Bph9+Pigm | 6 | 2.75 ± 0.37c | 3.76 ± 0.41a |
| Gm6 lines | Bph6+Pigm | 8 | 3.56 ± 0.54b | 3.62 ± 0.42a |
| Gm9 lines | Bph9+Pigm | 10 | 3.26 ± 0.19b | 3.43 ± 0.52a |
| R6888 | Pigm | 8.20 ± 0.26a | 3.63 ± 0.41a | |
| Luoyang69 | Bph6+Bph9 | 2.73 ± 0.15c | 8.23 ± 0.15b | |
| 9311 | 7.90 ± 0.16a | 8.33 ± 0.33b |
Different characters of superscripts (a, b and c) indicate significant differences by one-way ANOVA analysis.
Table 3.
The main agronomic traits and rice quality of parents and improved new lines.
| Line | GY (t/ha) | DTH (d) | PH (cm) | PL (cm) | NGP | SF (%) | TGW (g) | AAC (%) | GC (mm) | GT | HRR (%) | LWR | CD (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Huahui3006 | 8.43 | 125 | 114.7 | 23.8 | 220.6 | 88.90 | 27.5 | 16.10 | 62 | 6.0 | 64.20 | 3.4 | 3.60 |
| Huahui7713 | 8.67 | 126 | 114.3 | 23.2 | 230.6 | 88.20 | 27.8 | 16.40 | 63 | 6.0 | 63.10 | 3.3 | 4.60 |
| R6888 | 8.31 | 129 | 117.6 | 22.3 | 258.2 | 85.80 | 27.3 | 15.50 | 65 | 3.0 | 64.10 | 3.3 | 4.41 |
| Luoyang69 | 8.52 | 120 | 112.3 | 23.8 | 171.4 | 88.01 | 29.4 | 17.30 | 65 | 6.2 | 69.50 | 2.9 | 7.05 |
Abbreviations: GY, grain yield; AAC, apparent amylose content; CD, chalkiness degree; DTH, days to heading; GC, gel consistency; GT, gelatinization temperature; GY, grain yield; HRR, head rice rate; LWR, length-width ratio; NGP, number of grains per panicle; PH, plant height; PL, panicle length; SF, spikelet fertility; TGW, thousand-grain weight.
Table 4.
The genotypic and resistance performances of parents, improved lines and derived hybrid combinations.
Table 4.
The genotypic and resistance performances of parents, improved lines and derived hybrid combinations.
| Line | Genotype | Blast resistance | BPH resistance | |||||||
| Bph6 | Bph9 | Pigm | Seedling | Panicle blast severity index | Incidence of infected panicles | Integrated resistance index | ||||
| R6888 | - | - | + | 3 | 3 | 3 | 3 | HS | ||
| Luoyang69 | + | + | - | 9 | 9 | 9 | 9 | R | ||
| Huahui3006 | + | + | + | 3 | 3 | 3 | 3 | R | ||
| Huahui7713 | + | + | + | 3 | 3 | 3 | 3 | R | ||
| XuanLiangYou3006 | +/- | +/- | +/- | 3 | 3 | 3 | 3 | R | ||
| WeiLiangYou7713 | +/- | +/- | +/- | 3 | 3 | 3 | 3 | R | ||
Abbreviations: HS, high susceptible; R, resistance.
Table 5.
The agronomic trait performance, BPH and blast resistance of new hybrid rice varieties in state new variety trails.
Table 5.
The agronomic trait performance, BPH and blast resistance of new hybrid rice varieties in state new variety trails.
| Variety | Year | Agronomic traits | Grain quality traits | Blast resistance | BPH resistance | |||||||||||
| GY | DTM | AAC (%) | GC (mm) | CD (%) | HRR (%) | IDI | HSPBS | |||||||||
| GY (t/ha) | ±CK (%) | DTM (d) | ±CK (d) | |||||||||||||
| Weiliangyou7713 | 2019 | 10.22 | 6.27 | 138.0 | 4.6 | 15 | 82 | 0.8 | 68.9 | 3.5 | 5 | R | ||||
| 2020 | 9.61 | 8.76 | 135.9 | 2.8 | 14.3 | 82 | 0.9 | 65.9 | 3 | 5 | R | |||||
| Average | 9.91 | 7.52 | 136.9 | 3.7 | 14.65 | 82 | 0.85 | 67.4 | 3.25 | 5 | R | |||||
| Xuanliangyou3006 | 2020 | 9.13 | 3.24 | 137.5 | 5.6 | 16.8 | 60 | 1.4 | 66.3 | 3.4 | 3 | MR | ||||
| 2021 | 9.39 | 1.89 | 139.0 | 8.1 | 15.4 | 61 | 7 | 64.9 | 2.6 | 3 | MR | |||||
| Average | 9.26 | 2.56 | 138.3 | 6.9 | 16.1 | 60.5 | 4.2 | 65.6 | 3 | 3 | MR | |||||
Abbreviations: GY, grain yield; DTM, days to mature; CK, control check variety in state new variety trail; AAC, apparent amylose content; GC, gel consistency; CD, chalkiness degree; HRR, head rice rate; IDI, integrated disease index; HSPBS, highest scale of panicle blast severity; R, resistance; MR, medium resistance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



