
Submitted:
26 March 2024
Posted:
27 March 2024
You are already at the latest version
Abstract
The gastrointestinal tract is colonized by trillions of different microorganisms, named the gut microbiota, which is key to degrade undigested food such as dietary fibers. The fermentation of these food components leads to the production of short-chain fatty acids (SCFA) acetate, propionate, and butyrate, which exploit several beneficial roles for the host’s health. Their production and absorption happen in different ways in the human intestine and depend on the type of dietary fiber reaching the gut and the microorganisms involved in the fermentation. The supplementation of SCFAs, mostly butyrate, in treating gastrointestinal, metabolic, cardiovascular, and gut-brain-related diseases has been reported in the medical literature. This review aims to give an overview of the production and absorption dynamics of acetate, propionate, and butyrate in the human gut, with a final focus on the role played by these SCFAs on gastrointestinal and metabolic health and the present therapeutic implications.
Keywords:
intestinal microbiota
; short-chain fatty acids (SCFAs)
; dietary fiber fermentation
; gastrointestinal and metabolic health
; therapeutic implication
1. Introduction
The human gastrointestinal tract is heavily colonized by trillions of microbes that include hundreds of species endowed with a wide range of hydrolases involved in the fermentation of indigestible carbohydrates[1]. Microbial fermentation of polysaccharides is highest in the colon, reaching a daily production rate of 300 mmol/day, of which only 10 mmol/day are excreted [2]. The main volatile short-chain fatty acids (SCFA) produced are acetate, propionate, and butyrate in a ratio of 60:25:15 [3]. Butyrate acts as a key SCFA in the intestine. It serves as the primary energy for the metabolism of colonocytes, improving the integrity of the epithelial tissue, alleviating mucosal inflammation, and stimulating the absorption of electrolytes [4]. Furthermore, dissociated butyric acid can freely penetrate the cytoplasm, inhibiting DNA replication and dissociating the nutrient transport system from bacteria, leading to a broad-spectrum antibacterial effect [5]. Propionate is thought to benefit the gut environment, such as lowering lipogenesis, cholesterol levels and carcinogenesis [6]. Finally, studies demonstrate that acetate can beneficially affect host energy and substrate metabolism in the gut by stimulating the secretion of gut hormones such as the glucagon-like peptide-1 and peptide YY [7]. Thanks to their beneficial properties, SCFA is often used as a supplement to treat different diseases. However, the pharmaceutical formulation of short-chain fatty acid supplements fundamentally impacts their delivery and absorption. After looking at the production of SCFA acetate, propionate, and butyrate in the intestinal environment in the first part of this review, the second part will focus on the absorption of these SCFA. It will give an overview of SCFA supplements used in clinical trials, with particular attention to their formulations. Moreover, the importance of SCFAs in gastrointestinal and metabolic health will be discussed, concluding with the existing therapeutic implications.
2. Production of SCFA in the Gastrointestinal Tract
2.1. Cross-Feeding and Production of SCFA in the Human Intestine
Microbial communities are shaped by positive and negative interactions ranging from competition to mutualism. Millions of microbial inhabitants are present in the mammalian gut, and the interactions between these microbes produce synergistic responses [8]. Many ecological dynamics are mediated by diffusible metabolites, which can act as nutrient sources, inhibitory compounds, or signaling molecules [8]. Cross-feeding is the exchange of metabolites as energy and nutrients among different specimens or strains of microorganisms [9]. Other types of cross-feeding within the gut microbiome exist parasitism, commensalism, and mutualism. Parasitism occurs when one microbe benefits from a substrate produced by a partner organism while changing the environment to harm this producer. On the other hand, mutualism cross-feeding results when one consumer feeds on metabolites produced by another microbe with no impact on the latter. Finally, commensalism cross-feeding occurs when two species feed on a metabolite produced by the other or when one microbe feeds on a metabolite from another and modifies the environment to benefit the producer. It is essential to mention that for many species, cross-feeding fermentative intermediates is an integral part of their lifestyle in the gut. Important fermentative intermediates are SCFAs and carboxylic acids with a short aliphatic tail of 6 carbons, especially acetate (C2), propionate (C3), and butyrate (C4). Some species of bacteria produce These metabolites under anaerobic conditions upon fermentation of dietary fibers, mainly oligofructose, arabinoxylan, inulin, and pectin [10]. However, other factors, such as the low pH (5.5), are likely to beneficially affect the colon's community structure and microbial activities [11,12]. This consideration may be necessary, for example, to allow butyrate-producing bacteria to compete against carbohydrate-utilizing bacteria, such as Bacteroides spp., which, on the other hand, prefer a pH of 6.5 [13].
2.2. Production of Acetate by the Intestinal Microbiota
Acetate is a net fermentation product for most gut anaerobes and almost invariably achieves the highest concentration among SCFAs in the gut lumen [14]. Microbial-derived acetate production is yielded by the fermentation of indigestible foods, especially foods of acetogenic fibers such as galacto-oligosaccharides (GOS) and inulin [15]. Microbial fermentation of acetogenic fibers generates acetate production via two metabolic pathways: acetogenesis and carbon fixation. The former consists in the production of acetate, mediated by homoacetogenic bacteria or acetogens, which are able to produce acetate from H2 and CO2; while the carbon fixation pathway produces acetate from CO2 as a precursor, and it is also known as the WOOd-Ljungdahl pathway [16]. In particular, this trend is accompanied by an increase of the Firmicutes/Bacteroidetes ratio and cross-feeding mechanisms, as reported by the upregulation of pyruvate fermentation pathways to acetate and lactate by Lactobacillus reuteri and other unclassified bacteria [17]. Other studies support these findings, describing increased abundance of important acetate producers, such as Akkermansia muciniphila, during human fasting and caloric restriction interventions [18,19]. This intermediate is of particular interest because it can be further metabolized by acetate-consumers, such as Faecalibacterium prausnitzii and Roseburia intestinalis/Eubacterium rectale to produce butyrate [20]. More specifically, it has been reported that acetate is a growth requirement for these bacteria [20,21]. Thus, it is an essential intermediate in the intestine.
2.3. Production of Propionate by the Intestinal Microbiota
Propionate is a SCFA that can derive mainly from two essential pathways through the fermentation of different carbohydrates by gut bacteria: i) the succinate pathway consists of the fermentation of hexose and pentose sugars with the production of propionate, while ii) the propanediol pathway produces propionate through fermentation of fructose and rhamnose. The former is found mainly in Bacetroidetes and in the Negativicutes class of Firmicutes [21], and it is the primary route for propionate formation from dietary carbohydrates fermentation as the abundant Bacteroidetes drive it. In particular, succinate is a precursor of propionate, and its conversion to propionate requires vitamin B12 [22]. Propionate formation from rhamnose and fructose has been reported in gut bacteria belonging to the Lachnospiraceae family, including Roseburia inulinivorans and Blautia species [21]. Besides carbohydrates, peptides and amino acids can also be precursors for propionate formation. However, amino acid-fermenting bacteria have been estimated to constitute less than 1% of the large intestinal microbiota. In particular, Bacteroidetes are responsible for propionate formation by proteolysis from peptides and amino acids [23]. More specifically, in vitro, incubations of fecal slurries with individual amino acids reported that propionate derived mainly from aspartate, alanine, threonine, and methionine [24]. Finally, cross-feeding between different commensal gut bacteria is also essential in propionate production. Indeed, bacteria belonging to Bacteroides species, Escherichia coli, and Anaerostipes rhamnosivorans can degrade deoxy sugars by producing the pathway intermediate 1,2 propanediol as the final product. E. halii and Lactobacillus reuteri can further degrade this intermediate with propionate formation [25]. Although propionate is less frequently studied compared to other microbial metabolites, such as butyrate, it has been reported that it also has some distinct health-promoting properties, including cholesterol-lowering [26] and antilipogenic [27] effects, stimulation of satiety [28] and protection against colorectal cancer in particular [29]. Its beneficial effect in the contest of gastrointestinal diseases, particularly inflammatory bowel diseases (IBD) and inflammatory bowel syndrome (IBS), will be discussed in more detail in the third part of this review.
2.4. Production of Butyrate by the Intestinal Microbiota
Production of butyrate can derive from different pathways: butyrate formation by synthesis from acetoacetyl-CoA, which is formed by the reaction of two molecules acetyl-CoA: Butyryl-CoA, acetate CoA-transferase converts butyryl-CoA to generate butyrate. This has been observed in Eubacterium, Roseburia, Anaerostipes and Faecalibacterium prausnitzii [3]. Another route of butyrate formation is through the phosphotransbutylase and butyrate kinase. For example, specific Coprococcus species and numerous Clostridium species in the Firmicutes family have butyrate kinase to generate butyrate [30]. Among Firmicutes, the two most abundant gut bacteria families of butyrate producers are Ruminococcaceae and Lachnospiraceae. Faecalibacterium prausnitzii belongs to the family of Ruminococcaceae and is one of the most abundant species in the healthy human microbiota [23]. As previously mentioned, this bacteria produces butyrate via butyryl-CoA: acetate CoA transferase with net consumption of acetate, which stimulates its growth on carbohydrate energy sources [20]. In particular, this bacterium has gained increasing interest in recent years thanks to evidence reporting its anti-inflammatory properties in the intestine. Due to its beneficial properties, it has attracted interest as a potential therapeutic for patients suffering from IBD, whose microbiota usually are deprived of this bacteria [31].
Butyrate-producing Lachnospiraceae show considerable divergence in their phylogeny, gene organization, and physiology [32]. Eubacterium rectale and Roseburia species are closely related and constitute a significant group of butyrate-producing Firmicutes through the butyryl-CoA: acetate CoA transferase route. It is of interest to note that in some Roseburia strains, at mildly acidic pH, butyrate is almost the sole fermentation acid produced, with net consumption of acetate accompanying the formation of butyrate. On the other hand, other strains also produce formate and lactate in addition to butyrate [32]. Moreover, certain Lachnospiraceae, including A. hardus and E. halii, have the ability to grow in the presence of lactate and produce butyrate [33]
Also, butyrate can be formed through the fermentation of peptides and amino acids. An example is Intestinimonas AF211, which ferments lysine to butyrate [34]. Moreover, several distinct pathways exist for glutamate degradation to butyrate in butyrate-producing bacteria. These intermediates enter the main butyrate pathway either via pyruvate (Fusobacterium spp, Clostridium limosum) or crotonyl-CoA (found in different Firmicutes, including Acidamincoccus symbiosum, Clostridium sporosphaeroides, Clostridium symbiosum, etc). As it concerns the fermentation routes of other amino acids, these are less well characterized [35]. However, there is evidence that histidine is converted to glutamate, which is further fermented to butyrate by the intestinal microbiota [36,37]. An overview of the production of the three different SCFA and the metabolic pathways and bacteria involved is represented in Table 1.
2.5. Cross-Feeding Lays the Basis of Butyrate Production by Intestinal Microbiota
As previously mentioned, the production of SCFAs and other intermediates depends on the dietary fibers and, in lower amounts, on peptides and amino acids metabolized by the intestinal bacteria. These food components belong to the prebiotics category: non-digestible food ingredients that beneficially affect the consumer by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon [38]. Examples of industrial prebiotics are, for example, inulin-type fructans, GOS, and fructooligosaccharides (FOS) [39]. Inulin-type fructans are naturally present in different vegetables, such as onions, garlic, leek, bananas, and chicory roots. These compounds encompass short and long polymers of fructose with varying degrees of polymerization. Because of their β-linkages, inulin-type fructans are not digested nor absorbed in the human gastrointestinal tract [40]. Once these fibers reach the colon, they are selectively fermented by the microbiota, mainly by the bifidobacterial communities. During the complex colon fermentation process, they are primarily converted to SCFAs and other organic acids (e.g., lactate and succinate) and gases (hydrogen gas and carbon dioxide) [41]. In particular, the presence of unfermented inulin-type fructans in the intestine elicits the growth of intestinal Bifidobacteria, the so-called bifidogenic effect [42], as well as an enhancement of colonic butyrate production, the so-called butyrogenic effect [42], promoted by Bifidobacteria, and thanks to the cross-feeding phenomenon [11]. In the case of inulin-type fructans, two main types of cross-feeding have been reported: one involving short oligosaccharides and monosaccharides released by Bifidobacterium from the prebiotic substrate; the other one using consumption of end-products of bifidobacterial fructans fermentation, including acetate and lactate [43]. For example, E. halii DSM 17630 has been shown to efficiently convert lactate and acetate produced by B.adolescentis DSM 20083 into butyrate, when growing in co-culture with oligofructose [43]; moreover A. caccae DSM 14662 in co-culture with B.longum BB536 convert acetate and fructose produced by the latter strain during substrate breakdown [44]. Although clostridiales species form a minor fraction of the human colon microbiota (5-10%), butyrate formation by strict anaerobic bacteria, including Clostridium genus has been known for a while. In particular, more than 90% of the colonic butyrate-producing bacteria are represented by Faecalibacterium prausnitizii (Clostridium leptum cluster) and Eubacterium/Roseburia spp (Clostriudium coccoides cluster). The rate of butyrate formation by acetate-consumers (e.g., F. prausnitzii and Roseburia intestinalis) can vary depending on the species of butyrate-producing bacterium and the type of fermentable carbohydrate [8]. Indeed, besides inulin-type fructans, the butyrogenic effect is already known in the case of resistant starch fermentation [45], and in vitro studies have reported an efficient butyrate production in a co-culture of B.longum JCM 1217 and Eubacterium limosum JCM 6421 on germinated barley. In this case, the former strain produces lactate upon starch degradation, while the latter uses lactate for butyrate production [46]. Another study recently reported an efficient cross-feeding between R. intestinalis (a butyrate producer) and Ruminococcus hydrogenotrophicus (an acetate producer) when growing on xylan. In this context, xylan degradation first occurs thanks to R. intestinalis, which produces carbon dioxide and hydrogen gas, which serve as substrates for R. hydrogenotrophicus to grow, along with the production of acetate. This SCFA is then an indispensable co-substrate for butyrate production [47]. An overview of the production of SCFAs in the gut by the microbiota is represented in Figure 1.
As previously mentioned, researchers have lately been drawn to the study of butyrate because of its beneficial properties in the intestinal environment. Indeed, butyrate has been reported to have essential effects on intestinal cell development and gene expression [4,48] and is generally thought to play a protective role against colorectal cancer and colitis. Its beneficial roles in the context of gastrointestinal and metabolic diseases will be discussed in more detail in the third part of this review.
3. Absorption of SCFAs in the Intestine and SCFA Supplements
3.1. Absorption of Butyrate
For many years, it was believed that the primary mechanism of butyrate absorption was passive diffusion in its liposoluble form [48]. Today, considerable evidence suggests that SCFAs, including butyrate, are predominantly absorbed through a facilitated process involving a series of transport proteins. The characterization of several transmembrane proteins has led to the identification of two well-defined absorption pathways, both involving monocarboxylate transporters: MCT1 and MCT4 [49,50], two hydrogen-coupled transporters, and SMCT1, a sodium-coupled transporter [51].
From the early studies conducted by Thibault et al., assessing butyrate absorption in diseased colon tissue from Inflammatory Bowel disease (IBD), Familial Adenomatous Polyposis (FAP), and colorectal cancer (CRC), it was highlighted that MCT1 mRNA was drastically reduced in diseased tissues and correlated with the degree of inflammation. Functionally, this was demonstrated by a reduction in butyrate absorption and metabolism [52]. In cancerous tissue (CRC), however, there are peculiarities: while MCT1 expression decreases during the transition from normal to malignancy, being downregulated in the early stages of carcinogenesis [53], a subsequent upregulation of MCT1 has been described in advanced metastatic CRC tumors. In these tumors, MCT1 and MCT4 transporters play a crucial role in lactate transport and, consequently, intracellular pH regulation. Inhibiting MCT1 reduces intracellular pH, leading to tumor cell death. In this context, MCT1 and MCT4 can be potential therapeutic targets in cancer treatment [54,55,56]. Butyrate has been previously approved for clinical use in CRC treatment [57], as it is a substrate for MCT1 and MCT4, is well metabolized, and no side effects have been reported until now [58]. In contrast to the MCT1 receptor, knowledge regarding the regulation of SMCT1 at the intestinal level is still limited. SMCT1 is downregulated during intestinal inflammation, and its expression is often silenced in aberrant crypt foci, colon adenomas, colon tumors, and colon cancer cell lines, suggesting that STMC1 silencing is an early event in colon tumorigenesis [59,60,61,62]. It has been proposed that SMCT1 functions as a tumor suppressor, and its ability to mediate butyrate entry into colonocytes underlies its potential tumor-suppressive function [63].
Also noteworthy among the control and interaction systems with butyrate are the so-called efflux transporters, capable of removing butyrate from cells. Among these, Breast Cancer Resistance Protein (BCRP) is believed to limit drug absorption, bioavailability, and toxicity. Butyrate is a substrate for BCRP [64], and the inhibition of BCRP has significantly potentiated the inhibitory effect of butyrate on cell proliferation [65]. After absorption, butyrate signals through three membrane G-protein-coupled receptors (GPCRs), GPR41, GPR-43, and GPR 109A, present on the surface of colon cells, adipocytes, and immune cells. These receptors modulate cytokine levels and various signaling pathways when activated, promoting the anti-inflammatory response [66].
3.1.1. Butyrate Supplements
The literature generally presents studies conducted using two different formulations of butyrate: calcium butyrate (CaBu) and sodium butyrate (NaBu). Sodium butyrate and calcium butyrate are salts of butyric acid but differ in the metal to which they are bound. Calcium and sodium are the primary cations in the extracellular space, with calcium exhibiting reduced water solubility compared to sodium [67]. The choice of butyrate formulation with associated metal could be significant in treating patients with specific conditions or deficiencies [68]. The formulation of CaBu associated with vitamin D may be particularly interesting, especially for cancer prevention [69]. NaBu formulations, depending on the inflammatory context, may contribute to protective immunity relative to the associated ion [68]. It has been demonstrated that sodium storage in tissues improves defense against invasive pathogens [70]. However, immune activation induced by sodium salt may also negatively influence wound healing [71]. It should be noted that the concentration of salts coupled with butyrate is generally in the order of a few milligrams, depending on the formulation examined.
In addition to the formulation, the type of pharmaceutical form used for product delivery should also be carefully evaluated based on the site of action and the desired effect. Although several studies have reported beneficial effects of butyrate in the colon [100], some in vitro studies [101] and others conducted on mice have shown that butyrate enemas administered for three consecutive days induced concentration-dependent colon hypersensitivity (from 3-8 up to 1000 mmol/L) and mechanical hyperalgesia but no macroscopic and histological modification of the colon mucosa. This condition mimics the clinical condition observed in patients with IBS and is used as a model of chronic non-inflammatory colon hypersensitivity [102]. However, in human subjects, a comparable administration of butyrate in the distal colon decreases pain and discomfort, sharply contrasting with studies in rats. Some scholars attribute this difference to the different modulation of butyrate-coupled receptors in rats and humans and the concentration of butyrate present in the colon after exogenous administration compared to that naturally present given endogenous butyrate production [103]. It is reasonable to think that the pharmaceutical formulation may be functional in the butyrate concentration in the colon and have a pharmacological effect on the examined pathology [94]. Oral formulations with gastro-resistant capsules, microencapsulation, or enemas (see Table 2) may have different effects in different parts of the body, as the concentration of butyrate reaching the tissues may vary depending on the pharmaceutical form's ability to release the product [104]. Lipid microencapsulation rather than protection in gastro-resistant capsules allows n-butyric acid compounds to first not diffuse the unpleasant odor of rancid butter that characterizes the product and secondly not be readily hydrolyzed by gastric acids and thus reach the small intestine and colon where they can exert their function. New cream formulations based solely on butyrate [105] and free of corticosteroids are currently on the market and could prove helpful in controlling local inflammations, preventing the side effects associated with corticosteroids [106]; however, to date, there are no clinical studies evaluating their effectiveness.
3.2. Absorption of Propionate
Propionate has been associated with reducing lipogenesis and serum cholesterol levels[107]. It elicits strong effects on weight control and eating behavior [108]. It has also been demonstrated that, like butyrate, propionate exerts an antiproliferative effect on colon tumor cells [109,110]. The production of propionate by intestinal bacteria involves the transformation of prebiotic compounds such as L-rhamnose, D-tagatose, inulin, resistant starch, polydextrose, and arabinoxylans [6]. However, it isn't easy to perform a comparative assessment of the modulatory effects of propionate of such compounds due to the heterogeneity of the experimental setup of the studies. It should be emphasized that a direct connection between the production of SCFAs and their concentration in the intestinal lumen can only be established in an in vitro context without intestinal absorption.
The mechanisms of propionate production involve specific fermenting bacteria that use specific metabolic strategies, as previously mentioned. The propionate thus produced is easily transported systemically [111,112], passing through the liver. In general, propionate and acetate can activate GPR41 and GPR43 cell surface receptors but can also be easily absorbed at the level of any cell, bypassing SCFA receptors on the cellular surface. It has been shown that propionate also enhances the differentiation of T cells into effector cells such as Th1 and Th17 in favor of regulatory T cells producing anti-inflammatory IL-10 [113]. This regulation is essential for maintaining intestinal homeostasis and preventing chronic IBD.
3.2.1. Propionate Supplements
Unlike butyrate, formulations of propionate used for supplementation have been rarely studied in clinical trials (especially in obesity, diabetes, and cardiovascular disease, see Table 3). Recently, supplementation of propionic acid administered twice daily with 500 mg capsules for a 14-day treatment period in patients with multiple sclerosis (MS) has shown a significant increase (30%) compared to baseline in Treg cells and a reduction in Th17 cells [114], correlated with the attenuation of clinical symptoms (reduction in relapse and stabilization of disability). In a crossover RCT, administering an inulin-propionate ester formulation for 24 weeks to overweight adult subjects confirmed that increased propionate in the colon prevents weight gain in enrolled subjects [115]. Currently, two clinical trials are evaluating the effect of sodium propionate in subjects with various types of pathologies, but none of them in the gastroenterological field (Table 3a). Given the growing clinical evidence on the immunomodulatory effects of propionate, an increase in well-structured clinical studies is hoped, particularly in the context of chronic intestinal inflammations.
3.3. Absorption of Acetate
Although less studied than butyrate, acetate could also be of interest as it is less toxic to epithelial cells, stimulates bacteria that produce butyrate through cross-feeding, and has anti-inflammatory and protective properties [119]. Receptors like GPR43, which play an essential role in calcium homeostasis, are susceptible to acetate and propionate [120]. The probiotic activity of Saccharomyces cerevisiae var. boulardii is believed to be closely associated with its unusually high production of acetate [121]. The mechanism of action of acetate on intestinal cells is less known, but an interesting aspect is the positive effect that acetate has shown on body weight control. In mice, acetate administration can impact body weight control through effects on energy intake and expenditure [122]. In humans, studies on long-term oral acetate supplementation or endovenous/gastric infusion in the colon with weight loss and energy expenditure as primary outcomes are limited [7], and cross-sectional/cohort analyses have shown inconsistent results with obesity and adiposity [80]. The primary acetate source remains dietary integration through dairy products, pasta, bread, eggs, smoked fish, and coffee [123]. Other significant sources include ethanol, vinegar, and microbial production obtained from fermentation of indigestible carbohydrates (particularly acetogenic fibers like inulin and galactooligosaccharides [15].
3.3.1. Acetate Supplements
The most commonly used formulations in clinical studies are inulin acetate ester and sodium acetate, administered in the proximal colon by enema. As already reported for propionate, clinical studies regarding the use of acetate do not directly involve gastrointestinal disorders but rather the effect of oral supplementation of fermented foods for weight control [122]. Table 4 reports the impact of acetate interventions on hyperinsulinemic females.
4. Implications of SCFAs in Human Gastrointestinal and Metabolic Health
Several studies have indicated the involvement of SCFA in human GI and metabolic health. SCFAs are thought to have pleiotropic effects on gastrointestinal and metabolic health. The identified signaling mechanisms of SCFAs may function through two main mechanisms. The first is via interactions with GPCRs, as previously described, expressed in various organs, including the intestine, kidney, and heart [128,129,130]. These receptors are expressed in various cell types within the gastrointestinal tract, including enterocytes, enteroendocrine cells, immune cells, and neuronal cells, mediating a range of physiological responses [130]. The second acts as a histone deacetylase (HDAC) inhibitor [131,132], promoting gene expression and regulating cell metabolism, differentiation, and proliferation by inhibiting specific gene transcription [133,134,135].
4.1. Gastrointestinal Diseases
SCFAs play a critical role in maintaining gut health and have been implicated in various gastrointestinal diseases, including IBD, CRC, and disorders of the gut-brain axis. The supposed mechanisms of SCFA are summarized in Table 5.
4.1.1. Inflammatory Bowel Disease
The interaction between SCFAs and IBD is multifaceted, involving the interplay among gut microbiota, immune responses, and the integrity of the gut epithelial barrier [136,137]. Butyrate, a primary energy source for colonocytes, exerts anti-inflammatory effects by inhibiting the activation of the nuclear factor kappa B and reducing proinflammatory gene expression [138]. A decline in SCFA-producing bacteria characterizes IBD patients, notably butyrate producers like Faecalibacterium prausnitizii and Roseburia hominis [139,140,141]. This results in reduced colonic SCFA levels linked to compromised gut barrier function in IBD [142,143].
SCFAs protect against IBD-associated intestinal inflammation through various mechanisms [144]. They enhance the intestinal epithelial barrier by promoting mucus production and tightening tight junctions between epithelial cells [144]. Additionally, SCFAs modulate immune responses by influencing the differentiation and function of Tregs, suppressing excessive immune reactions [145]. Several pathways are involved in SCFA-mediated immune regulation, including GPCRs, HDACs, and the regulation of innate immune sensors like Toll-like receptors (TLRs) and Nod-like receptor family pyrin domain containing 3 (NLRP3) inflammasome. SCFAs inhibit the progression of IBD by regulating innate immune sensors, TLRs, and NLRP3 inflammasomes. SCFAs protect the intestinal barrier; acetate, propionate, and butyrate stimulate the intestinal NLRP3 inflammasome, increasing IL-18 secretion and enhancing intestinal barrier integrity [146]. Moreover, SCFAs engage with GPR43 and GPR109A receptors essential for regulating intestinal immunity, stimulating the production of Treg. This has been demonstrated in preclinical studies, where controlling colonic Treg levels and function in a GPR43-dependent manner has been shown to mitigate inflammation, as seen in SCFA-mediated protection against colitis in GPR43-deficient (Gpr43(−/−)) mice [147,148]. Furthermore, SCFAs promote the differentiation of Tregs by inhibiting HDAC activity, and Tregs secrete protective cytokines, such as IL10, to suppress inflammation [149]. SCFAs not only inhibit TLR signaling, but butyrate acts as an HDAC inhibitor to suppress TLR4 expression and the TLR2-mediated release of inflammatory factors [150,151,152]. Finally, SCFAs participate in tissue repair processes within the gut, promoting the proliferation and differentiation of epithelial cells, thus facilitating the healing of damaged tissues caused by inflammation in IBD [153].
A recent study investigated the utility of fecal SCFA concentrations as surrogate markers for gut microbiota diversity in patients with IBD and primary sclerosing cholangitis (PSC) [154]. Results decreased fecal isobutyrate levels compared to healthy controls. Fecal acetate and butyrate positively correlated with fecal calprotectin and serum C-reactive protein in ulcerative colitis (UC) patients. Furthermore, UC patients with higher fecal calprotectin levels exhibited elevated fecal acetate, butyrate, and propionate levels. These findings suggest potential associations between SCFA levels and disease activity in UC patients.
Although SCFA concentrations are decreased in IBD patients, SCFA supplementation through diet or probiotics shows promise as an adjunct therapy, with minimal adverse effects reported [139,153,155,156,157,158]. However, the exact mechanisms underlying the therapeutic effects of SCFAs in IBD require further elucidation, highlighting the complexity of their relationship with the disease. Figure 2 illustrates the mechanism of action of SCFAs.
4.1.2. Colorectal Cancer
Colorectal cancer (CRC) ranks among the top three causes of cancer-related mortality worldwide, with increasing recognition of the microbiota's contribution to its pathogenesis [159]. Various factors contribute to CRC, including a high-fat diet, stress, antibiotics, synthetic food additives, a sedentary lifestyle, and environmental factors [160]. High-fat diet, especially prevalent in Western diets featuring high red and processed meat consumption, high fructose corn syrup, and unhealthy cooking methods, significantly contributes to CRC [161]. Current research has explored the protective role of dietary fibers in reducing the risk of CRC [162,163].
A systematic review and meta-analysis by Alvandi et al. explored the role of fecal SCFAs in CRC incidence and risk stratification [164]. The study, encompassing seventeen case-control and six cross-sectional studies, revealed that individuals with lower concentrations of acetic, propionic, and butyric acid are at a higher risk of CRC. Although these findings suggest a potential association between decreased fecal SCFA concentrations and CRC susceptibility, emphasizing the importance of gut microbiota and bacterial metabolites in CRC prevention, their exact role in CRC prevention remains poorly understood. SCFAs, notably butyrate and propionate, are thought to influence CRC by regulating gene expression, expressing immunomodulatory effects, promoting immune cell differentiation, and mitigating inflammation. Moreover, compelling evidence underscores the role of SCFAs, including butyrate and propionate, in directly influencing intestinal epithelial cell transformation and inhibiting CRC by regulating tumor suppressor gene expression, promoting apoptosis, and modulating CRC cell proliferation and metabolism [165,166,167]. Butyrate is an energy metabolite and supports normal colon cell proliferation. In CRC cells, butyrate reprograms cell metabolism by promoting pyruvate kinase isozyme 2 (PKM2) activity, inhibiting the Warburg effect and enhancing energy metabolism, therefore inhibiting cancerous colonocytes, which rely on glucose due to the Warburg effect [133,168]. SCFAs act as an HDAC inhibitor, fostering apoptosis in cancer cells [166,169,170,171,172,173]. Additionally, SCFAs play a pivotal anti-inflammatory role in regulating local and systemic immune cells, contributing to their antitumor efficacy [174]. SCFAs mitigate inflammation by inhibiting NF-κB activation, decreasing pro-inflammatory cytokine expression like tumor necrosis factor-alpha (TNF-α), promoting anti-inflammatory cytokines such as IL-10, and transforming growth factor-beta, and facilitating the differentiation of naïve T cells into Tregs, thereby dampening immune responses [175]. They promote antimicrobial compound production, neutrophil and macrophage inhibition, Treg activation, and dendritic cell induction of tolerogenic properties [174]. In a recent in vitro experiment by Mowat et al. CRC cells treated with SCFAs induced much greater activation of CD8+ T cells than untreated CRC cells [175]. Surprisingly, the butyrate-producing bacterium Fusobacterium nucleatum does not consistently inhibit colon cancer; instead, it may promote cancer progression via mechanisms such as TLR4/myeloid differentiation primary response 88 (MYD88)/NF-κB signaling [176]. Furthermore, despite the anticipated decrease in DNA damage within cancer cells, numerous reports suggest that SCFAs might exacerbate DNA damage accumulation in CRC cells by disrupting DNA repair mechanisms [173,177,178,179,180]. Hence, the antitumorigenic effects of SCFAs likely involve intricate mechanisms extending beyond the tumor cells themselves, particularly significant in CRC cells with underlying DNA repair defects, such as microsatellite instability-high (MSI) CRC subset known for its heightened immunogenicity. Given inflammation's potent role in tumor progression, these effects likely contribute to SCFAs' antitumor efficacy. However, as tumor-targeted T-cell responses are crucial for antitumor immunity and treatment efficacy, SCFAs like butyrate may suppress such responses, potentially fueling tumor progression and compromising treatment outcomes [181,182,183,184].
Tian et al. investigated the potential protective role of SCFAs in the development of colitis-associated CRC using a mouse model induced by azoxymethane (AOM) and dextran sodium sulfate (DSS) [185]. The researchers administered a mix of SCFAs in the drinking water throughout the study. They found that the SCFA mix significantly reduced tumor incidence and size in the mice with colitis-associated colorectal cancer. Additionally, the SCFA mix improved colon inflammation and disease activity index score and suppressed the expression of proinflammatory cytokines such as IL-6, TNF-α, and IL-17. These findings suggest that SCFA mix administration could prevent tumor development and attenuate colonic inflammation, indicating its potential as an agent for the prevention and treatment of colitis-associated colorectal cancer. Further investigation is warranted to determine if supplementing with dietary butyrate or consuming foods rich in butyrate-producing bacteria, such as omega-3 polyunsaturated fatty acids, can effectively hinder colorectal cancer and lower its occurrence.
4.1.3. Disorders of the Gut-Brain Axis
The gut-brain axis facilitates bidirectional communication between the gastrointestinal and nervous systems through a complex signaling pathway network [186,187,188]. This intricate system encompasses connections such as the enteric nervous system, vagus nerve, immune system, endocrine signals, microbiota, and metabolites. Disruption of communication along the gut-brain axis is increasingly recognized as a significant contributor to neuroinflammation, which is considered a common feature of several neurodegenerative diseases, including Alzheimer's and Parkinson's diseases, characterized by chronic and debilitating conditions marked by the progressive degeneration of neurons [189,190,191,192,193,194,195]. Recent research suggests that neurodegenerative diseases may originate in the intestinal epithelium before affecting the brain via the gut-brain axis [196,197,198,199,200,201]. Various studies have documented the accumulation of protein aggregates, characteristic pathologies of neurodegenerative diseases such as Alzheimer's and Parkinson's, in enteric neurons or the gastrointestinal epithelium years before their detection in the central nervous system [194,202,203,204,205]. Functional studies illuminate major microbiota components' roles in the gut-brain axis [206,207,208,209]. An important aspect is the observed close correlation between alteration in the microbiota, mucosal immunity, and intestinal vascular impairment, potentially leading to the gradual release of systemic inflammatory mediators and bacterial components such as LPS, thereby initiating or exacerbating the development of neurological disorders [210,211,212]. Evidence suggests that microbial and systemic inflammatory molecules could contribute to cerebral vascular impairment, microglial activation, neuronal dysfunction, and pre- and post-synaptic activity imbalances. The microbiome of patients with Parkinson’s and Alzheimer's disease exhibits a reduction in SCFA-producing bacteria [210,213]. Recent research has highlighted their importance for learning and memory, with cuts in SCFAs associated with inflammation in Multiple Sclerosis patients and compromised neuronal function in various neurodegenerative diseases [214,215]. Furthermore, SCFAs appear to have neuroprotective roles, affecting the brain indirectly or directly by acting as ligands for GPCRs or as epigenetic modulators of HDAC to control transcriptional changes that affect neuronal functions [216,217,218,219,220]. The diminished concentration of SCFAs is suggested to be a critical factor in disrupting gut-brain balance, but the role of SCFA in this context is under active investigation. These SCFAs can cross the blood-brain barrier, likely through the monocarboxylate transport system, influence brain function, and regulate blood flow, with dietary butyrate demonstrating an anti-inflammatory effect in the brain by influencing blood–brain barrier permeability [221,222]. SCFAs have also been implicated in maintaining gut and immune homeostasis in mammalian systems, highlighting their neuro-immunoendocrine regulatory role in the brain [221,223]. In Parkinson's disease, the decline in butyrate levels is thought to lead to intestinal barrier integrity impairment, release of LPS and other pro-inflammatory molecules into the bloodstream, and trigger microglial activation [135,224]. Furthermore, reduced SCFAs and microbiota alterations result in decreased circulating GLP-1 levels. The lowered SCFA-mediated secretion of GLP-1 may activate pro-inflammatory pathways and depressive symptoms in PD patients [225,226]. Additionally, butyrate can induce epigenetic modifications in the genome of neurodegenerative disorder patients. Methylation analysis on blood samples from Parkinson’s disease patients and controls revealed a correlation between alterations in butyrate-producing bacterial taxa and epigenetic changes in genes containing butyrate-associated methylation sites. Notably, these modified sites coincide with genes implicated in psychiatric and gastrointestinal disorders [227].
In a study by Kong et al., 16S ribosomal RNA gene sequencing and gas chromatography-mass spectrometry analyses in a Drosophila model of Alzheimer's disease revealed a decrease in Lactobacillus and Acetobacter species correlating with a dramatic reduction in acetate [228]. Similarly, in Drosophila models of Parkinson's disease, administration of sodium-butyrate reduced degeneration of dopaminergic neurons and improved locomotor defects in a pan-neuronal transgenic fly model expressing mutant-human-α-Synuclein [222]. The SCFA composition derived from microbes also clinically correlates with neural activity and brain structure, as evidenced by functional and structural magnetic resonance imaging [229]. Recently, Muller et al. examined the fecal SCFA profile of patients with a major depressive disorder/generalized anxiety disorder, comparing it with nuclear magnetic resonance spectroscopy and self-reported depressive and gut symptoms. The severity of depressive symptoms positively correlated with acetate levels and negatively correlated with butyrate levels [230]. In preclinical studies focusing on Alzheimer’s disease, prebiotic and probiotic supplementation appear advantageous, although limited data is available specifically on SCFA. Bonfili et al. demonstrated the positive impacts of SLAB51 treatment on 8-week-old transgenic Alzheimer’s disease model mice over four months [231,232,233]. SLAB51 administration enhanced performance in the novel object recognition test, reduced brain damage, decreased Aβ plaques, elevated SCFAs, and lowered plasma cytokine levels [233]. Additionally, prebiotics have shown efficacy in Alzheimer’s disease amyloid models. Liu et al. treated 5XFAD transgenic Alzheimer’s disease model mice with prebiotic mannan oligosaccharide for eight weeks starting from birth. They observed improvements in cognitive deficits, reduction in Aβ plaques, decreased oxidative stress, diminished microglial activation, and alterations in the gut microbiome. Interestingly, gut microbiome-induced changes in the brain appeared to be mediated by SCFAs, as supplementation with SCFAs produced similar effects [234]. Finally, a case report demonstrated that FMT improved cognitive function, microbiota diversity, and SCFA production in an Alzheimer's patient [235].
Several studies have investigated the administration of probiotics in both murine models and human subjects with Parkinson’s disease, exploring their impact on gastrointestinal and neurological symptoms [236,237,238,239,240,241,242]. A pilot study regarding FMT use in Parkinson’s patients has recently been published, with promising data [243]. However, only a few studies have evaluated SCFA's role. Specifically, Bifidobacterium has been demonstrated to be effective in modulating the host microbiota in a murine model induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [244]. In mice overexpressing α-synuclein, a prebiotic diet altered the activation of microglia and motor deficits by changing the composition of the gut microbiome and levels of SCFAs [245]. Combining polymannuronic acid with Lacticaseibacillus rhamnosus GG demonstrated more potent neuroprotective effects against Parkinson’s disease than either treatment alone, suggesting the therapeutic promise of synbiotics in Parkinson’s disease [246]. Oral administration of B. breve CCFM1067 to MPTP-induced Parkinson’s disease mice led to a reduction in intestinal microbial alterations, marked by a decline in pathogenic bacteria (Escherichia-Shigella) and an increase in Bifidobacterium and Akkermansia. This intervention also restored SCFA production (butyrate and acetate), which may account for the observed local and cerebral anti-inflammatory effects. Recently, Bifidobacterium animalis subsp. Lactic Probio-M8 (Probio-M8) was examined to evaluate its additional beneficial effects and mechanisms when used as an adjunct treatment alongside conventional therapy (Benserazide and dopamine agonists) in patients with Parkinson’s. This investigation was conducted over three months in a randomized, double-blind, placebo-controlled trial [247]. Clinical outcomes were assessed by analyzing changes in various clinical indices, gut microbiome composition, and serum metabolome profiles before, during, and after the intervention. The findings revealed that co-administration of Probio-M8 resulted in additional benefits, including improved sleep quality, reduced anxiety, and alleviated gastrointestinal symptoms. Metagenomic analysis demonstrated significant modifications in the participants' gut microbiome and serum metabolites following the intervention. The serum concentration of acetic acid was notably higher in the probiotic group.
IBS is a Disorder of Gut-Brain Interaction (DGBI) characterized by abdominal pain and changes in stool consistency or frequency. According to the Rome IV criteria, IBS can be divided into four subtypes based on the primary clinical features: IBS with diarrhea (IBS-D), IBS with constipation (IBS-C), IBS with mixed stool patterns (IBS-M), and unclassified IBS [248,249,250]. SCFAs play a pivotal role in IBS, with reported findings indicating that patients with IBS exhibit significantly elevated levels of acetate, propionate, and total SCFAs in fecal samples, with the severity of symptoms correlating positively [251]. Alterations in SCFA levels are subtype-specific, with reduced levels in IBS-C and increased levels in IBS-D compared to controls [252,253]. Treem et al. sought to investigate whether patients with IBS-D exhibit a distinct pattern and pace of carbohydrate and fiber fermentation in SCFA in vitro studies of fecal homogenates compared to controls. The fecal SCFA profile of IBS-D patients revealed diminished concentrations of total SCFA, acetate, and propionate alongside elevated levels and proportion of n-butyrate [254]. Fredericks et al., in 2021, examined gut microbiota, concentrations of SCFA, and mRNA expression of monocarboxylate transporters in individuals with IBS-C, IBS-D, and healthy controls. They observed changes in fecal SCFA ratios in both IBS groups, with a decrease in all three measured SCFAs in IBS-C and a reduction specifically in acetic acid in IBS-D [255]. Similarly, Undseth et al. aimed to compare colonic fermentation between individuals with IBS and healthy counterparts by examining serum SCFA concentrations before and 90 minutes after ingesting lactulose, an unabsorbable yet fermentable carbohydrate. They found that reduced serum SCFA levels post-lactulose ingestion may indicate compromised colonic fermentation in IBS patients [256]. The dysregulated SCFA levels in feces are linked to shifts in intestinal bacterial composition in IBS patients, characterized by higher counts of acetate and propionate-producing bacteria like Veillonella and Lactobacillus and lower counts of butyrate-producing bacteria like Roseburia-Eubacterium rectale group [251,257,258]. Zhou et al. recently set out to investigate how linaclotide affects the gut microbiota and pinpoint essential bacterial genera that could influence linaclotide's effectiveness. Interestingly, they discovered a direct link between higher levels of Blautia and SCFA concentrations and the amelioration of clinical symptoms in patients with IBS-C [259].
SCFAs, particularly propionate and butyrate, show promise as non-invasive biomarkers for diagnosing IBS, with diagnostic properties consistent across all IBS subgroups. Farup et al. 2016 examined fecal SCFA as a potential diagnostic indicator for IBS in a study involving 25 IBS subjects and 25 controls. They assessed total SCFA levels and individual SCFA amounts to identify the most effective diagnostic approach. Their findings revealed that the discrepancy between propionic and butyric acid levels demonstrated superior diagnostic performance using a threshold of 0.015 mmol/l to indicate IBS, independent of the IBS subgroup [260].
Several potential mechanisms exist through which SCFAs could influence the pathophysiology of IBS, many of which have been previously examined in the IBD section of this review. As already described, SCFAs interact with specific receptors, such as GPR41, GPR43, and GPR109A, expressed in various gastrointestinal cell types, modulating physiological responses. They play a multifaceted role in immunity and inflammation, influencing inflammatory mediator production, immune cell differentiation, and intestinal barrier integrity [261,262,263,264,265]. Additionally, SCFAs influence the differentiation of immune cells, including T cells and Tregs, and suppress intestinal inflammation [145,266,267]. They also contribute to the integrity of the intestinal barrier by promoting mucin secretion and enhancing tight junction assembly [268,269,270,271,272,273].
Furthermore, SCFAs impact gut motility through various mechanisms, including modulation of neural activity, neurotransmitter release, and regulation of calcium signaling and smooth muscle contractility [274,275,276,277,278,279,280,281,282,283,284,285]. The effects of SCFAs on colonic motility are nuanced and context-dependent, varying based on SCFA concentration and colonic segment [274,275,276,277,278,279,280,281,282,283,284,285]. Waseem et al., in their recent prospective observational study, investigated the associations between fecal SCFAs, colonic transit time, fecal bile acids, and dietary intake in individuals with IBS and healthy controls [286]. They found that fecal SCFAs were inversely correlated with overall and segmental colonic transit time, with similar patterns observed in both IBS and healthy control groups. Additionally, the acetate-to-butyrate ratio was associated with slower transit times. Logistic regression analyses demonstrated that acetate could accurately predict delayed colonic transit time and BAD. These findings suggest that fecal SCFAs and dietary factors may play a role in the IBS pathophysiology and serve as diagnostic markers for bowel transit disorders [286].
4.2. Metabolic Diseases
Metabolic diseases, spanning conditions like obesity, type 2 diabetes (T2D), and metabolic dysfunction-associated steatotic liver disease (MASLD), present significant health challenges globally [287,288,289]. Central to the pathophysiology of these conditions is the intricate interplay between the gut microbiota and SCFAs, which profoundly influence host metabolism. An imbalance in gut microbial communities is a critical contributor to the development of common metabolic disorders in humans [290]. Nevertheless, the emerging evidence underscores the promising therapeutic potential of targeting the gut microbiota and its metabolites for managing various metabolic conditions, extending beyond the well-established associations with obesity, T2D, and MASLD. Mechanisms of SCFA in metabolic disorders are summarized in Table 6.
4.2.1. Obesity
Obesity poses a significant risk for various chronic conditions, including T2D, insulin resistance, MASLD, and cardiovascular disease, among others [291]. Interestingly, obese individuals have been associated with altered fecal SCFA concentrations, particularly propionate. A study involving Mexican children revealed that those with excess weight and obesity exhibited lower concentrations of fecal propionate and butyrate compared to their normal-weight counterparts [292]. A recently published study examined African-origin groups from different regions and discovered variations in gut microbiota composition and predicted functions linked to population obesity and geography [293]. The study found that fecal SCFA concentrations are inversely correlated with microbial diversity and obesity. However, the prediction of obesity from microbiota varied by country: Prevotella-rich microbiota dominates in traditionally non-western groups, while Bacteroides-rich microbiota is found in high-income countries. Conversely, other studies have associated obese individuals with higher fecal SCFA concentrations than lean individuals [294,295]. A study in the Netherlands found that overweight and obese individuals had elevated fecal SCFA concentrations compared to lean counterparts, suggesting enhanced microbial energy extraction [294]. Indeed, a previous survey of 441 adults published by Cuesta-Zuluaga et al. in 2018 revealed a correlation between higher fecal SCFA levels and obesity [296]. The excessive production of SCFA may contribute to weight gain due to increased energy storage despite its typically beneficial effects on well-being [294,297,298,299,300].
However, these findings are debatable due to possible fluctuations in SCFA concentrations and broader microbiota alterations within the intestinal microbial community [301]. Numerous studies have investigated the role of SCFAs in adiposity, examining human subjects and conducting in vitro and in vivo animal studies. In vitro studies have demonstrated that acetate and propionate treatment can induce expressions of vital metabolic regulators, promoting lipolysis metabolism [302,303].
Animal studies have shown that SCFA supplementation can counteract weight and adiposity gain, with treatments like sodium butyrate inducing weight loss by enhancing energy expenditure and fat oxidation [304,305]. In mice on a high-fat diet, butyrate supplementation increases the expression of peroxisome proliferator-activated receptor-γ (PPARγ) coactivator-1 alpha (PGC-1α), activates AMP Kinase (AMPK) and p38, and improves insulin sensitivity inducing weight loss by enhancing energy expenditure and fat oxidation [306]. This finding was observed when the functioning of adipose and hepatic PPARγ pathways were intact. Dietary supplementation with SCFAs has been found to upregulate GPR43 and GPR41 expressions in adipose tissue, enhance triglyceride hydrolysis, promote free fatty acid oxidation in adipose tissue, leading to brown fat production, and reduce body weight in HFD-fed mouse models [307]. Ganoderma lucidum, a medicinal mushroom with a long history of use in Asian countries, has been shown to increase SCFA production and GPR43 expression in C57BL/6 J mice, enhance ileal tight junction proteins and antibacterial peptides expression, mitigate endotoxemia, and attenuate HFD-induced upregulation of TLR4/Myd88/NF-κB signaling in adipose tissue [308,309].
Overall, while growing evidence supports the role of SCFAs in obesity treatment, comprehensive mechanistic studies are needed to elucidate their precise mechanisms of action and optimize their therapeutic potential.
4.2.2. Type 2 Diabetes
Research involving individuals from various ethnic backgrounds has revealed that those with T2D exhibit diminished levels of SCFA-producing bacteria. This is implicated in insulin resistance and the progression of T2D and can contribute to gut inflammation [310].
Regarding microbial metabolites, SCFAs exhibit diverse effects across various sites regulating glucose metabolism. In vitro and in vivo studies have shown that SCFAs act as potent secretagogues for glucagon-like peptide 1 (GLP-1) and peptide YY (PYY), thereby enhancing feelings of satiety via the gut-brain axis. Consequently, they may indirectly decrease appetite and subsequent food intake, thus mitigating the risk of weight gain, a known predisposing factor for T2D [218]. Research has revealed that acetate can reduce hormone-sensitive lipase phosphorylation in human multipotent adipose tissue-derived stem adipocytes in a Gi-coupled manner [311]. Acetate and butyrate activate GPR43 and GPR41 on rat intestinal cells, stimulating insulin, GLP-1, and peptide YY secretion, modulating blood lipid metabolism and lowering peripheral blood glucose levels, slowing intestinal transit, decreasing gastric emptying, food intake, and intestinal motility [312]. Acetate and butyrate activate GPR43 and GPR41 receptors on intestinal cells, promoting the secretion of insulin, GLP-1, and peptide YY, which helps modulate blood lipid metabolism and lower peripheral blood glucose levels [313,314]. In the liver, SCFAs have been observed to inhibit glycolysis and gluconeogenesis while enhancing glycogen synthesis and fatty acid oxidation [218,315,316,317]. Additionally, SCFAs have been shown to improve glucose uptake in skeletal muscle and adipose tissue by upregulating the expression of GLUT4 through AMPK activation. Furthermore, in skeletal muscle, SCFAs reduce glycolysis, leading to the accumulation of glucose-6-phosphate and increased glycogen synthesis [306,315,316,317,318,319,320]. In preclinical models, ingesting soluble dietary fibers prompts the production of SCFAs, particularly propionate, and butyrate, which activate intestinal gluconeogenesis (IGN), a process crucial for glucose and energy homeostasis [321]. SCFAs play a role in promoting IGN production to mitigate metabolic diseases in mice [322]. Butyrate triggers IGN gene expression via a cAMP-dependent mechanism. At the same time, propionate, as an IGN substrate, enhances gene expression through activation of the gut-brain neural circuit [321], thereby exerting beneficial effects on glucose regulation, energy balance, and body weight control. In rabbits, acetate could curb lipid accumulation, promoting lipolysis and fatty acid oxidation and inhibiting synthesis [323].
Regarding the microbiota populations, T2D patients exhibit a higher abundance of Proteobacteria and a skewed Firmicutes/Bacteroidetes ratio compared to healthy individuals, alongside reduced SCFA-producing Bacteroides [324,325,326]. Acetate and butyrate improved intestinal barrier function and increased the number of Bacteroides species in NOD model mice, which helped to inhibit T1D [327].
As a result of the role of SCFAs in human glucose metabolism, intervention studies involving the supplementation of propionate and butyrate have been conducted. A recent meta-analysis has shown that probiotic intervention can significantly improve the homeostatic model assessment of insulin resistance (HOMA-IR) and considerably decrease glycated hemoglobin HbA1c levels and fasting blood glucose levels in T2DM patients compared to placebo [328,329]. However, the evidence remains inconclusive due to the limited number of studies conducted in small cohorts. Nevertheless, these studies suggest that inulin-propionate supplementation (10 g/day) increases GLP-1 and PYY levels while reducing food intake, contributing to body weight regulation [330,331]. Additionally, sodium butyrate supplementation (4 g/day) enhances insulin sensitivity solely in lean individuals and not those with metabolic syndrome [332]. Despite these promising findings, the optimal doses and exposure durations for SCFA treatment in T2D remain undefined, and further research is needed to elucidate their time- and dose-dependent effects. Additionally, studies have focused on translating fecal microbiota from lean donors to recipients with metabolic syndrome to enhance insulin sensitivity [333,334].
Moreover, adopting a low-calorie, low-protein, low-carbohydrate HFD as a fast-mimicking diet has shown promise in promoting cell regeneration, reducing protein kinase A and mTOR activity, inducing the expression of Sox2 and Ngn3, and restoring insulin production, secretion, and glucose homeostasis in both T2D mouse models and type 1 diabetes patients [335].
4.2.3. Metabolic Dysfunction–Associated Steatotic Liver Disease
The transition from NAFLD to MAFLD and MASLD marks a significant shift in the understanding and classification of metabolic liver diseases, aiming to reflect their pathophysiology better and reduce social stigma [289]. This evolution in terminology and diagnostic criteria, supported by international experts and widely accepted in clinical practice guidelines, emphasizes the link between metabolic dysfunction and liver health, paving the way for improved disease identification and management strategies. The connection between MASLD and its advancement to steatohepatitis and cirrhosis has previously been associated with the gut microbiome via multiple pathways. This correlation could stem from gut microbiota alterations and the systemic impact of metabolites derived from it, such as SCFA [336].
Notably, the gut microbiota of patients with the formerly known non-alcoholic fatty liver disease (NAFLD) exhibits a significantly reduced abundance of SCFA-producing bacteria such as Bacteroides, Lactobacillus curvatus, and L. plantarum [337,338,339,340]. As described in this review, previous studies have suggested that individuals with obesity and MASLD tend to have higher levels of fecal SCFAs [294,296,341]. However, it is unclear whether there is a relationship between circulating SCFA levels and MASLD and other metabolic disorders [298,342,343,344]. While some studies have found no significant differences between control groups and MASLD patients, others have reported lower SCFA levels in MASLD cirrhosis or higher levels in patients with hepatocellular carcinoma and cirrhosis linked to MASLD [342,344,345,346]. These conflicting conclusions may result from differences in study design, such as variations in the selection criteria for control and MASLD patients or discrepancies in the severity of underlying MASLD conditions.
The mechanisms linking SCFAs and MASLD may involve alterations in glucose homeostasis, lipid metabolism, and inflammatory and immune responses [341,347]. The gut-liver axis plays a crucial role in this process, as evidenced by the reciprocal relationship between gut microbiota, gut-derived metabolites, and liver function [348].
Although the precise role of these SCFAs in MASLD remains unclear, insights may be gleaned from research on other metabolic disorders as previously described in this review. Previous studies have associated acetate with greater gut microbiota diversity, reduced visceral fat, and less severe MASLD cases [349,350]. Consistent with these findings, our study observed lower acetate levels in MASLD patients than healthy controls. Propionate, when present in adequate concentrations, is also linked to positive health outcomes and the regulation of gut hormones influencing appetite and fullness [350]. However, conflicting evidence exists, as evidenced by a study on early MASLD patients where higher levels of SCFA-producing bacteria and fecal acetate and propionate were associated with an elevated TH17/Treg ratio, suggesting a potential contribution to low-grade inflammation [341].
In a recent study, Thing et al. investigated the association between plasma SCFAs and MASLD. The results showed higher plasma concentrations of propionate, formate, valerate, and α-methylbutyrate but lower plasma acetate concentrations in MASLD patients compared to healthy controls. Moreover, among MASLD patients, significant fibrosis was positively associated with several SCFAs [351].
Animal studies have shown that supplementation with SCFAs such as sodium acetate and sodium butyrate can protect against hepatic steatosis induced by nicotine and metabolic factors [352,353]. In MASLD patients, downregulation of the GLP-1 receptor in the liver is observed, with butyrate supplementation in MASLD mice enhancing GLP-1 receptor expression by inhibiting HDAC-2, consequently promoting energy metabolism and inhibiting lipid accumulation [354]. Butyrate also improves insulin sensitivity, activates AMPK to induce the expression of fatty acid oxidation genes in hepatocytes, and reduces fat deposition in MASLD mice [355]. The MASLD mouse model increases the abundance of beneficial bacteria in the intestine, such as Christensenellaceae, Blautia, and Lactobacillus, establishing a positive feedback loop by augmenting butyric acid production [356,357]. Additionally, butyrate attenuates MASLD-induced intestinal mucosal injury by upregulating ZO-1 expression in the intestinal tract of mice, thereby preventing enterotoxin migration to the liver and suppressing liver inflammation [358].
Overall, these findings underscore the therapeutic potential of SCFAs in preventing and managing MASLD by targeting multiple pathways involved in its pathogenesis. Emerging evidence underscores the pathogenic role of microbe-derived metabolites, including trimethylamine, secondary bile acids, SCFAs, and ethanol, in MASLD pathogenesis [348].
4.3. Therapeutic Implications
4.3.1. Fecal Microbiota Transplantation
FMT is a therapeutic approach involving the transfer of a fecal suspension from a healthy donor to the patient's gastrointestinal tract to restore average microbial composition and function [359,360]. It is recommended by guidelines and consensus from international societies for the treatment of recurrent Clostridioides difficile infection (rCDI) [361,362,363,364,365]. Encouraging results indicate that FMT might also potentially treat additional conditions linked to disruptions in gut microbiota composition, including IBD and disturbances of the gut-brain axis, like anorexia [359,361,366,367,368,369,370,371,372]. The efficacy of FMT largely depends on the donor's microbiota, with "super donors" possessing favorable bacterial characteristics crucial for successful outcomes [373]. Advancements in frozen stool processing have facilitated the establishment of FMT libraries for clinical applications [369,374]. However, the specific bacterial composition of FMTs and the underlying treatment mechanisms remain unclear, necessitating further research to better understand this promising therapeutic approach [375].
Metabolite levels linked to gut microbiota, including SCFAs and bile acids, show improvement following FMT. Paramsothy et al. found that patients with UC achieving remission after FMT exhibited enrichment of Eubacterium hallii and Roseburia inulinivorans, along with elevated levels of SCFA biosynthesis and secondary bile acids, compared to non-responders [376]. FMT administration is thought to elevate SCFA levels in the colon and regulate the NF-κB pathway to reduce inflammation [377,378]. In a study conducted by Osaki et al. in 2021, the effectiveness of FMT was evaluated along with its impact on fecal microbiota and SCFA levels in patients with IBD and rCDI. The analysis of fecal microbiota showed changes in bacterial composition after FMT, with modifications in specific bacterial taxa associated with clinical response. In UC patients, fecal SCFA levels remained unchanged post-FMT, regardless of treatment response. However, responders showed a significant increase in fecal butyric acid levels in CD patients at eight weeks post-FMT compared to donors, while rCDI patients had lower pre-FMT butyric acid levels than donors. Furthermore, fecal propionic acid levels significantly increased at eight weeks post-FMT in rCDI patients, while acetic acid and butyric acid levels showed a non-significant increase [379]. Conversely, Seekatz et al. observed increased butyrate, acetate, and propionate levels and recovery of secondary bile acids like deoxycholate and lithocholic in rCDI patients post-FMT [380].
A 2021 RCT conducted by El-Salhy and colleagues investigated the impact of FMT on fecal SCFA levels in patients with IBS. The study included 142 participants from a previous study. The results showed that individuals who received FMT had increased levels of butyric acid, especially in the 30-g and 60-g FMT groups. In addition, the 60-g FMT group had higher levels of total SCFAs and several other SCFA types. Significantly, higher butyric acid levels were associated with symptom improvement in FMT responders [381].
4.3.2. Dietary Intervention
Dietary composition exerts a significant influence on gut microbes [382,383]. Various diets can alter microbial composition, increase the ratio of harmful bacteria to beneficial metabolites, and contribute to the development of chronic metabolic diseases such as obesity and T2D [384,385]. The potential role of dietary interventions in diseases from cognitive impairment to IBD has brought new studies on the connection between diet and microbiota [386,387,388]. Adopting healthy eating habits with a diet rich in fresh fruits, vegetables, and whole grains can reduce the risk of cardiovascular and metabolic diseases and cancer. On the other hand, consuming refined and processed foods such as sugary treats, fried foods, processed meats, and refined grains may increase their likelihood [385,389].
Dietary fiber is an essential component of food, and soluble fiber is resistant to gastrointestinal digestive enzymes and is utilized by the anaerobic intestinal microbiota to produce SCFAs [390]. In a recent systematic review examining the impact of dietary fibers on SCFA production and gut microbiota composition in healthy adults, a total of forty-four human intervention studies on confirmed and candidate prebiotics were included. Among them, inulin was the most extensively studied dietary fiber. While specific studies indicated notable rises in total SCFAs after dietary fiber intervention, others observed no significant alterations, indicating that the influence of nutritional fibers on SCFA levels may be influenced by variables such as dosage, fiber type, and baseline gut microbiota composition [391,392]. To analyze the potential mechanisms of the role of the ketogenic diet in epilepsy, a recent study by Gudan et al. examined the impact of this on the synthesis of intestinal SCFAs in healthy adults [393]. The analysis highlighted that cruciferous and leaf vegetables, berries, and nuts consumption on a ketogenic diet have been linked to a positive impact on the profile of SCFAs. The LIBRE trial investigated the effect of the Mediterranean diet in 260 women and found that adherence to the Mediterranean diet led to increased fecal SCFA levels, particularly propionate and butyrate [394].
Dietary fibers play a crucial role in modulating intestinal SCFA levels, preserving mucosal homeostasis, enhancing intestinal epithelial integrity, fostering the growth of Tregs, and suppressing the expression of inflammatory cytokines to prevent or alleviate disease [395]. Supplementation with wheat bran, rich in arabinoxylan oligosaccharides, elevated butyrate, acetate, and propionate levels, along with total SCFA concentrations in a human trial [396]. However, the increased fecal bulking and reduced transit time associated with higher dietary fiber intake could decrease colonic SCFA absorption, potentially explaining the observed rise in fecal SCFA concentrations in studies with increased fiber content.
According to two studies, consumption of barley-kernel-based bread rich in β-glucan fibers for three days can result in an increase in the levels of Prevotella and a decrease in the levels of Bacteroides and intensified fermentation activity, SCFA serum levels, and gut hormone secretion (GLP-1, PYY, and GLP-2) in healthy adults, enhancing insulin sensitivity [397,398]. These results were observed among healthy participants, and they suggest that certain foods can have a significant impact on the gut microbiome. This shift was linked to a decrease in postprandial glucose response, corresponding to an increase in total serum SCFA concentration. Another study indicated that a supplement containing three grams per day of high molecular weight β-glucan altered the gut microbiota composition, increasing Bacteroides and decreasing Firmicutes, with correlations observed between changes in these bacteria and cardiovascular disease risk factors [399]. These findings suggest that high molecular weight β-glucan fibers can induce microbiota shifts, potentially explaining their metabolic benefits.
In 2020, Farup and Valeur conducted a study to investigate the impact of weight-loss interventions on fecal SCFA levels in people with obesity. They studied ninety subjects with morbid obesity and measured their fecal SCFA levels before and after a six-month conservative weight-loss intervention followed by bariatric surgery. The study found a reduction in total fecal SCFA levels post-surgery, accompanied by a decrease in the main straight-chain SCFAs such as acetic-, propionic-, and butyric-acids, and an increase in branched-chain SCFAs like isobutyric-, isovaleric-, and isocaproic-acids. This indicated a shift towards a proteolytic fermentation pattern. Interestingly, SCFA levels were associated with diet but not metabolic markers or fecal microbiota composition. This suggests that dietary interventions can potentially mitigate these effects [400].
4.3.3. Prebiotic and Probiotic Applications
In recent years, there has been a surge of interest in prebiotics and probiotics [401], with their mechanisms of action being intricate and diverse, often specific to particular strains and compounds [402]. Probiotics can alter the gastrointestinal microenvironment, outcompete pathogenic bacteria for nutrients, and hinder pathogenic growth by producing antimicrobial compounds unique to each strain [403,404]. Probiotics' safety and potential roles in diseases where gut microbiota is considered part of the pathophysiology have fueled research in this area [405,406]. SCFAs have the potential to regulate cognitive abilities and influence mental function via the gut-brain axis [407].
In 2015, Sawin et al. investigated the prebiotic properties of glycomacropeptide (GMP), a glycophosphopeptide. Using mouse models, researchers found that GMP reduced the abundance of Desulfovibrio bacteria, increased levels of cecal SCFA, and exhibited anti-inflammatory effects compared to casein and amino acid diets [408].
Holmes et al. conducted a six-week, three-period prebiotic intervention study on forty-one healthy adults to analyze personalized responses to different prebiotics, inulin, GOS, and dextrin. They found that the proportional increase in butyrogenic response to prebiotics was inversely correlated with regular dietary fiber intake [409]. The study suggested that individuals' gut microbiota may have a limited capacity to produce SCFAs from fiber, and their responsiveness to prebiotic treatment could be predicted based on diet and baseline SCFA levels in the stool.
A systematic review and meta-analysis focusing on dietary fiber interventions in individuals with type 2 diabetes revealed improvements in the relative abundance of Bifidobacterium and total SCFA. They enhanced glycated hemoglobin levels [410]. This review included an intervention study involving 16 g per day of inulin-type fructans for six weeks, notably increasing bifidobacteria concentrations [411]. Although the prebiotic treatment boosted fecal SCFA concentrations, including total SCFA, acetate, and propionate, it had no discernible impact on butyrate or overall bacterial diversity. Moreover, it did not positively influence glucose levels, insulin, gut hormones, appetite, or energy intake [412,413].
Inulin-type fructans possess a prebiotic effect, elevating Bifidobacterium, Lactobacillus, and Faecalibacterium prausnitzii abundances. The benefits reported include improved intestinal barrier function, insulin sensitivity, lipid profile, mineral absorption, and satiety [413]. However, the effects on blood glucose, cholesterol, and triglyceride concentrations appear favorable primarily in individuals with prediabetes and diabetes [414].
In a randomized, double-blind, placebo-controlled study assessing the impact of the probiotic intervention on fecal SCFAs, a multi-strain probiotic formula was administered to 56 postmenopausal obese women [415]. The study revealed a positive effect on their cardiometabolic health, with the higher probiotic dose group showing elevated levels of fecal SCFAs [415]. Another recent study investigated the impact of a low-carbohydrate diet compared to a habitual diet on fecal SCFA levels and serum inflammatory markers in obese women undergoing an energy-restricted diet [416]. After adjusting for baseline parameters, the two diet groups observed significant differences in fecal levels of butyric, propionic, and acetic acid.
5. Conclusion
A significant disparity has been highlighted in the clinical studies conducted with butyrate compared to those with propionate and acetate. Indeed, propionate and acetate play a secondary role compared to butyrate, although the evidence suggests that acetate and propionate are equally applicable fatty acids as butyrate and that gut bacteria sometimes utilize them to produce butyrate [8,299,302,304,305]. Similarly, it has been noted that the availability of propionate/acetate-based supplements is deficient compared to butyrate-based supplements. An additional emerging idea is that any therapeutic, pharmacological, integrative, or nutritional intervention must consider the role played by the intestinal microbiota. Human health is correlated with the ecology of microbial communities living inside and outside our bodies [417], and health itself evolves within an "ecological" perspective that involves the interaction between living beings and the environment. This important concept has been highlighted by different clinical[418,419] and pre-clinical [392] studies where individual microbiota composition and functions have been reported to influence dietary fiber supplementation, with varying consequences on intestinal health. For this reason, recognizing the importance of focusing on the microbiome leads to a "before" and "after" in health research and innovation, with the perspective nowadays to develop personalized medicine for patients, taking into account individual microbiota features when indicating the most appropriate treatment [361,420]. The relationship between nutrition, production/utilization of SCFA, and variation of the microbiota is just a small example to explain this ecological perspective that is gradually evolving towards the more philosophical concept of One Health, and perhaps in the not too distant future towards the more holistic concept of "Microbiome One Health"[421].
Author Contributions
Conceptualization, EB, SF, LB, and EVS.; methodology, EB, SF, LB, and EVS.; writing—original Draft Preparation, EB, SF, and LB; writing—review & Editing, EB, SF, LB, GL, CDB, BB, FZ, DM, MS, CR, IA, and EVS; supervision, EVS.
Funding
This research received no external funding
Conflicts of Interest
Edoardo Vincenzo Savarino has served as speaker for Abbvie, Agave, AGPharma, Alfasigma, Aurora Pharma, CaDiGroup, Celltrion, Dr Falk, EG Stada Group, Fenix Pharma, Fresenius Kabi, Galapagos, Janssen, JB Pharmaceuticals, Innovamedica/Adacyte, Malesci, MayolyBiohealth, Omega Pharma, Pfizer, Reckitt Benckiser, Sandoz, SILA, Sofar, Takeda, Tillots, Unifarco; has served as a consultant for Abbvie, Agave, Alfasigma, Biogen, Bristol-Myers Squibb, Celltrion, DiademaFarmaceutici, Dr Falk, Fenix Pharma, Fresenius Kabi, Janssen, JB Pharmaceuticals, Merck & Co, Nestlè, Reckitt Benckiser, Regeneron, Sanofi, SILA, Sofar, Synformulas GmbH, Takeda, Unifarco; he received research support from Pfizer, Reckitt Benckiser, SILA, Sofar, Unifarco, Zeta Farmaceutici. Sonia Facchin has served as a consultant for SILA, Unifarco, and Zeta Farmaceutici and has served as a speaker for Unifarco and SILA. Fabiana Zingone has served as a speaker for EG Stada Group, Fresenius Kabi, Janssen, Pfizer, Takeda, Unifarco, Malesci, and Kedrion and has served as a consultant for Galapagos. Brigida Barberio has served as a speaker for Abbvie, Agave, Alfasigma, AGpharma, Janssen, MSD, Procise, Sofar, Takeda, Unifarco. The other authors declare no conflict of interest.
References
- Musso, G.; Gambino, R.; Cassader, M. Gut Microbiota as a Regulator of Energy Homeostasis and Ectopic Fat Deposition: Mechanisms and Implications for Metabolic Disorders. Curr. Opin. Lipidol. 2010, 21, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Høverstad, T. Studies of Short-Chain Fatty Acid Absorption in Man. Scand. J. Gastroenterol. 1986, 21, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of Short-Chain Fatty Acid Production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.T.; Cresci, G.A.M. The Immunomodulatory Functions of Butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the Gut to the Peripheral Tissues: The Multiple Effects of Butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van De Wiele, T. Propionate as a Health-Promoting Microbial Metabolite in the Human Gut: Nutrition Reviews® Vol. 69(5):245-258. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Hernández; Canfora; Jocken; Blaak The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11, 1943. [CrossRef] [PubMed]
- Culp, E.J.; Goodman, A.L. Cross-Feeding in the Gut Microbiome: Ecology and Mechanisms. Cell Host Microbe 2023, 31, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Germerodt, S.; Bohl, K.; Lück, A.; Pande, S.; Schröter, A.; Kaleta, C.; Schuster, S.; Kost, C. Pervasive Selection for Cooperative Cross-Feeding in Bacterial Communities. PLOS Comput. Biol. 2016, 12, e1004986. [Google Scholar] [CrossRef]
- Rasouli-Saravani, A.; Jahankhani, K.; Moradi, S.; Gorgani, M.; Shafaghat, Z.; Mirsanei, Z.; Mehmandar, A.; Mirzaei, R. Role of Microbiota Short-Chain Fatty Acids in the Pathogenesis of Autoimmune Diseases. Biomed. Pharmacother. 2023, 162, 114620. [Google Scholar] [CrossRef]
- Belenguer, A.; Duncan, S.H.; Holtrop, G.; Anderson, S.E.; Lobley, G.E.; Flint, H.J. Impact of pH on Lactate Formation and Utilization by Human Fecal Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 6526–6533. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Duncan, S.H.; McWilliam Leitch, E.C.; Child, M.W.; Flint, H.J. pH and Peptide Supply Can Radically Alter Bacterial Populations and Short-Chain Fatty Acid Ratios within Microbial Communities from the Human Colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Interactions and Competition within the Microbial Community of the Human Colon: Links between Diet and Health. Environ. Microbiol. 2007, 9, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Hosmer, J.; McEwan, A.G.; Kappler, U. Bacterial Acetate Metabolism and Its Influence on Human Epithelia. Emerg. Top. Life Sci. 2023. [Google Scholar]
- Wong, J.M.W.; De Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic Health: Fermentation and Short Chain Fatty Acids: J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Schug, Z.T.; Vande Voorde, J.; Gottlieb, E. The Metabolic Fate of Acetate in Cancer. Nat. Rev. Cancer 2016, 16, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T.; et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metab. 2017, 26, 672–685. [Google Scholar] [CrossRef]
- Remely, M.; Hippe, B.; Geretschlaeger, I.; Stegmayer, S.; Hoefinger, I.; Haslberger, A. Increased Gut Microbiota Diversity and Abundance of Faecalibacterium Prausnitzii and Akkermansia after Fasting: A Pilot Study. Wien. Klin. Wochenschr. 2015, 127, 394–398. [Google Scholar] [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia Muciniphila and Improved Metabolic Health during a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef]
- Duncan, S.H. Growth Requirements and Fermentation Products of Fusobacterium Prausnitzii, and a Proposal to Reclassify It as Faecalibacterium Prausnitzii Gen. Nov., Comb. Nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic Distribution of Three Pathways for Propionate Production within the Human Gut Microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Strobel, H.J. Vitamin B12-Dependent Propionate Production by the Ruminal Bacterium Prevotella Ruminicola 23. Appl. Environ. Microbiol. 1992, 58, 2331–2333. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Macfarlane, G.T. Enumeration of Amino Acid Fermenting Bacteria in the Human Large Intestine: Effects of pH and Starch on Peptide Metabolism and Dissimilation of Amino Acids. FEMS Microbiol. Ecol. 1998, 25, 355–368. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic Metabolism Revisited: Metabolism of Lactic Acid Bacteria in Food Fermentations and Food Spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Beaulieu, K.E.; McBurney, M.I. Changes in Pig Serum Lipids, Nutrient Digestibility and Sterol Excretion during Cecal Infusion of Propionate, J. Nutr. 1992, 122, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Todesco, T.; Rao, A.; Bosello, O.; Jenkins, D. Propionate Lowers Blood Glucose and Alters Lipid Metabolism in Healthy Subjects. Am. J. Clin. Nutr. 1991, 54, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Martin, R.J.; Tulley, R.T.; Raggio, A.M.; McCutcheon, K.L.; Shen, L.; Danna, S.C.; Tripathy, S.; Hegsted, M.; Keenan, M.J. Dietary Resistant Starch Upregulates Total GLP-1 and PYY in a Sustained Day-Long Manner through Fermentation in Rodents. Am. J. Physiol. - Endocrinol. Metab. 2008, 295, E1160–E1166. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Belzacq, A.-S.; Haouzi, D.; Rouault, A.; Kroemer, G.; Brenner, C. Propionibacteria Induce Apoptosis of Colorectal Carcinoma Cells via Short-Chain Fatty Acids Acting on Mitochondria. Cell Death Differ.
- Louis, P.; Duncan, S.H.; McCrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted Distribution of the Butyrate Kinase Pathway among Butyrate-Producing Bacteria from the Human Colon. J. Bacteriol. 2004, 186, 2099–2106. [Google Scholar] [CrossRef]
- Quévrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an Anti-Inflammatory Protein from Faecalibacterium Prausnitzii, a Commensal Bacterium Deficient in Crohn’s Disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, Metabolism and Microbial Ecology of Butyrate-Producing Bacteria from the Human Large Intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oliphant, K.; Parreira, V.R.; Cochrane, K.; Allen-Vercoe, E. Drivers of Human Gut Microbial Community Assembly: Coadaptation, Determinism and Stochasticity. ISME J. 2019, 13, 3080–3092. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Ritari, J.; Boeren, S.; De Waard, P.; Plugge, C.M.; De Vos, W.M. Production of Butyrate from Lysine and the Amadori Product Fructoselysine by a Human Gut Commensal. Nat. Commun. 2015, 6, 10062. [Google Scholar] [CrossRef] [PubMed]
- Buckel, W. Unusual Enzymes Involved in Five Pathways of Glutamate Fermentation. Appl. Microbiol. Biotechnol. 2001, 57, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, J.; White, R.L.; Bearne, S.L. Proteomic Investigation of Amino Acid Catabolism in the Indigenous Gut Anaerobe Fusobacterium Varium. PROTEOMICS 2008, 8, 2691–2703. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a Reference Resource for Gene and Protein Annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary Fiber and Prebiotics and the Gastrointestinal Microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Updating the Concept of Prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-Feeding between Bifidobacteria and Butyrate-Producing Colon Bacteria Explains Bifdobacterial Competitiveness, Butyrate Production, and Gas Production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef]
- Molis, C.; Flourié, B.; Ouarne, F.; Gailing, M.; Lartigue, S.; Guibert, A.; Bornet, F.; Galmiche, J. Digestion, Excretion, and Energy Value of Fructooligosaccharides in Healthy Humans. Am. J. Clin. Nutr. 1996, 64, 324–328. [Google Scholar] [CrossRef]
- Roberfroid, M.B.; Van Loo, J.A.E.; Gibson, G.R. The Bifidogenic Nature of Chicory Inulin and Its Hydrolysis Products. J. Nutr. 1998, 128, 11–19. [Google Scholar] [CrossRef]
- Belenguer, A.; Duncan, S.H.; Calder, A.G.; Holtrop, G.; Louis, P.; Lobley, G.E.; Flint, H.J. Two Routes of Metabolic Cross-Feeding between Bifidobacterium Adolescentis and Butyrate-Producing Anaerobes from the Human Gut. Appl. Environ. Microbiol. 2006, 72, 3593–3599. [Google Scholar] [CrossRef]
- Falony, G.; Vlachou, A.; Verbrugghe, K.; Vuyst, L.D. Cross-Feeding between Bifidobacterium Longum BB536 and Acetate-Converting, Butyrate-Producing Colon Bacteria during Growth on Oligofructose. Appl. Environ. Microbiol. 2006, 72, 7835–7841. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the Effects of Diet on Bacterial Metabolism in the Large Intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef]
- Kanauchi, O.; Fujiyama, Y.; Mitsuyama, K.; Araki, Y.; Ishii, T.; Nakamura, T.; Hitomi, Y.; Agata, K.; Saiki, T.; Andoh, A.; et al. Increased Growth of Bifidobacterium and Eubacterium by Germinated Barley Foodstuff, Accompanied by Enhanced Butyrate Production in Healthy Volunteers. Int. J. Mol. Med. 1999. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C.; Bernalier-Donadille, A. H 2 and Acetate Transfers during Xylan Fermentation between a Butyrate-Producing Xylanolytic Species and Hydrogenotrophic Microorganisms from the Human Gut. FEMS Microbiol. Lett. 2006, 254, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, O.C.; Lederer, H.M.; Rombeau, J.L. Butyrate and the Colonocyte. Production, Absorption, Metabolism, and Therapeutic Implications. Adv. Exp. Med. Biol. 1997, 427, 123–134. [Google Scholar]
- Garcia, C.K.; Goldstein, J.L.; Pathak, R.K.; Anderson, R.G.; Brown, M.S. Molecular Characterization of a Membrane Transporter for Lactate, Pyruvate, and Other Monocarboxylates: Implications for the Cori Cycle. Cell 1994, 76, 865–873. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2018, 8, 299–314. [Google Scholar] [CrossRef]
- Coady, M.J.; Chang, M.-H.; Charron, F.M.; Plata, C.; Wallendorff, B.; Sah, J.F.; Markowitz, S.D.; Romero, M.F.; Lapointe, J.-Y. The Human Tumour Suppressor Gene SLC5A8 Expresses a Na+-Monocarboxylate Cotransporter. J. Physiol. 2004, 557, 719–731. [Google Scholar] [CrossRef]
- Thibault, R.; Blachier, F.; Darcy-Vrillon, B.; de Coppet, P.; Bourreille, A.; Segain, J.-P. Butyrate Utilization by the Colonic Mucosa in Inflammatory Bowel Diseases: A Transport Deficiency. Inflamm. Bowel Dis. 2010, 16, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Lambert, D.W.; Wood, I.S.; Ellis, A.; Shirazi-Beechey, S.P. Molecular Changes in the Expression of Human Colonic Nutrient Transporters during the Transition from Normality to Malignancy. Br. J. Cancer 2002, 86, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Chiche, J.; Ricci, J.-E.; Pouysségur, J. Tumor Hypoxia and Metabolism -- towards Novel Anticancer Approaches. Ann. Endocrinol. 2013, 74, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.P.; Schulze, A. Targeting Cancer Metabolism--Aiming at a Tumour’s Sweet-Spot. Drug Discov. Today 2012, 17, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Marchiq, I.; Pouysségur, J. Hypoxia, Cancer Metabolism and the Therapeutic Benefit of Targeting Lactate/H(+) Symporters. J. Mol. Med. Berl. Ger. 2016, 94, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Bolden, J.E.; Peart, M.J.; Johnstone, R.W. Anticancer Activities of Histone Deacetylase Inhibitors. Nat. Rev. Drug Discov. 2006, 5, 769–784. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, W.N. Lactate Transporter Defect: A New Disease of Muscle. Science 1986, 234, 1254–1256. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Myeroff, L.; Smiraglia, D.; Romero, M.F.; Pretlow, T.P.; Kasturi, L.; Lutterbaugh, J.; Rerko, R.M.; Casey, G.; Issa, J.-P.; et al. SLC5A8, a Sodium Transporter, Is a Tumor Suppressor Gene Silenced by Methylation in Human Colon Aberrant Crypt Foci and Cancers. Proc. Natl. Acad. Sci. 2003, 100, 8412–8417. [Google Scholar] [CrossRef]
- Zhang, Y.; Bao, Y.L.; Wu, Y.; Yu, C.L.; Sun, Y.; Li, Y.X. Identification and Characterization of the Human SLC5A8 Gene Promoter. Cancer Genet. Cytogenet. 2010, 196, 124–132. [Google Scholar] [CrossRef]
- Rodriguez, A.-M.; Perron, B.; Lacroix, L.; Caillou, B.; Leblanc, G.; Schlumberger, M.; Bidart, J.-M.; Pourcher, T. Identification and Characterization of a Putative Human Iodide Transporter Located at the Apical Membrane of Thyrocytes. J. Clin. Endocrinol. Metab. 2002, 87, 3500–3503. [Google Scholar] [CrossRef]
- Ganapathy, V.; Thangaraju, M.; Gopal, E.; Martin, P.M.; Itagaki, S.; Miyauchi, S.; Prasad, P.D. Sodium-Coupled Monocarboxylate Transporters in Normal Tissues and in Cancer. AAPS J. 2008, 10, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Martin, P.M.; Prasad, P.D.; Ganapathy, V. SLC5A8 (SMCT1)-Mediated Transport of Butyrate Forms the Basis for the Tumor Suppressive Function of the Transporter. Life Sci. 2006, 78, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C.G.; Vehr, A.-K.; Martin, I.V.; Gassler, N.; Rath, T.; Roeb, E.; Schmitt, J.; Trautwein, C.; Geier, A. Downregulation of Breast Cancer Resistance Protein in Colon Adenomas Reduces Cellular Xenobiotic Resistance and Leads to Accumulation of a Food-Derived Carcinogen. Int. J. Cancer 2011, 129, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.; Gregório, I.; Martel, F. The Short-Chain Fatty Acid Butyrate Is a Substrate of Breast Cancer Resistance Protein. Am. J. Physiol. Cell Physiol. 2011, 301, C984–994. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Chen, S.; Deng, B.; Tan, C.; Deng, J.; Zhu, G.; Yin, Y.; Ren, W. Implication of G Protein-Coupled Receptor 43 in Intestinal Inflammation: A Mini-Review. Front. Immunol. 2018, 9, 1434. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Makova, M.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Rhodes, C.J.; Valko, M. Essential Metals in Health and Disease. Chem. Biol. Interact. 2022, 367, 110173. [Google Scholar] [CrossRef]
- Wilck, N.; Balogh, A.; Markó, L.; Bartolomaeus, H.; Müller, D.N. The Role of Sodium in Modulating Immune Cell Function. Nat. Rev. Nephrol. 2019, 15, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Gaschott, T.; Stein, J. Short-Chain Fatty Acids and Colon Cancer Cells: The Vitamin D Receptor—Butyrate Connection. In Vitamin D Analogs in Cancer Prevention and Therapy; Reichrath, J., Tilgen, W., Friedrich, M., Eds.; Springer: Berlin, Heidelberg, 2003; ISBN 978-3-642-62435-3. [Google Scholar]
- Jantsch, J.; Schatz, V.; Friedrich, D.; Schröder, A.; Kopp, C.; Siegert, I.; Maronna, A.; Wendelborn, D.; Linz, P.; Binger, K.J.; et al. Cutaneous Na+ Storage Strengthens the Antimicrobial Barrier Function of the Skin and Boosts Macrophage-Driven Host Defense. Cell Metab. 2015, 21, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Byles, V.; Covarrubias, A.J.; Ben-Sahra, I.; Lamming, D.W.; Sabatini, D.M.; Manning, B.D.; Horng, T. The TSC-mTOR Pathway Regulates Macrophage Polarization. Nat. Commun. 2013, 4, 2834. [Google Scholar] [CrossRef]
- Guillemot, F.; Colombel, J.F.; Neut, C.; Verplanck, N.; Lecomte, M.; Romond, C.; Paris, J.C.; Cortot, A. Treatment of Diversion Colitis by Short-Chain Fatty Acids: Prospective and Double-Blind Study. Dis. Colon Rectum 1991, 34, 861–864. [Google Scholar] [CrossRef]
- Scheppach, W.; Sommer, H.; Kirchner, T.; Paganelli, G.M.; Bartram, P.; Christl, S.; Richter, F.; Dusel, G.; Kasper, H. Effect of Butyrate Enemas on the Colonic Mucosa in Distal Ulcerative Colitis. Gastroenterology 1992, 103, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Steinhart, A.H.; Brzezinski, A.; Baker, J.P. Treatment of Refractory Ulcerative Proctosigmoiditis with Butyrate Enemas. Am. J. Gastroenterol. 1994, 89, 179–183. [Google Scholar] [CrossRef] [PubMed]
- VERNIA, P.; MARCHEGGIANO, A.; CAPRILLI, R.; FRIERI, G.; CORRAO, G.; VALPIANI, D.; PAOLO, M.C.D.; PAOLUZI, P.; TORSOLI, A. Short-chain Fatty Acid Topical Treatment in Distal Ulcerative Colitis. Aliment. Pharmacol. Ther. 1995, 9, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Steinhart, A.H.; Hiruki, T.; Brzezinski, A.; Baker, J.P. Treatment of Left-Sided Ulcerative Colitis with Butyrate Enemas: A Controlled Trial. Aliment. Pharmacol. Ther. 1996, 10, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W.; Christl, S.U.; Bartram, H.P.; Richter, F.; Kasper, H. Effects of Short-Chain Fatty Acids on the Inflamed Colonic Mucosa. Scand. J. Gastroenterol. Suppl. 1997, 32. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.; Fidalgo, P.; Cravo, M.; Midões, J.; Chaves, P.; Rosa, J.; Birto, M.D.A.; Leitão, C.N. Short Chain Fatty Acids Are Effective in Short-Term Treatment of Chronic Radiation Proctitis: Randomized, Double-Blind, Controlled Trial. Dis. Colon Rectum 1999, 42, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Monteleone, G.; Grandinetti, G.; Villotti, G.; Di Giulio, E.; Frieri, G.; Marcheggiano, A.; Pallone, F.; Caprilli, R.; Torsoli, A. Combined Oral Sodium Butyrate and Mesalazine Treatment Compared to Oral Mesalazine Alone in Ulcerative Colitis: Randomized, Double-Blind, Placebo- Controlled Pilot Study. Dig. Dis. Sci. 2000, 45, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Fracasso, P.; Casale, V.; Villotti, G.; Marcheggiano, A.; Stigliano, V.; Pinnaro, P.; Bagnardi, V.; Caprilli, R. Topical Butyrate for Acute Radiation Proctitis: Randomised, Crossover Trial. The Lancet 2000, 356, 1232–1235. [Google Scholar] [CrossRef]
- Lührs, H.; Gerke, T.; Müller, J.G.; Melcher, R.; Schauber, J.; Boxberge, F.; Scheppach, W.; Menzel, T. Butyrate Inhibits NF-kappaB Activation in Lamina Propria Macrophages of Patients with Ulcerative Colitis. Scand. J. Gastroenterol. 2002, 37, 458–466. [Google Scholar] [CrossRef]
- Vernia, P.; Annese, V.; Bresci, G.; D’Albasio, G.; D’Inc??, R.; Giaccari, S.; Ingrosso, M.; Mansi, C.; Riegler, G.; Valpiani, D.; et al. Topical Butyrate Improves Efficacy of 5-ASA in Refractory Distal Ulcerative Colitis: Results of a Multicentre Trial. Eur. J. Clin. Invest. 2003, 33, 244–248. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Morera, R.; Ciccocioppo, R.; Cazzola, P.; Gotti, S.; Tinozzi, F.P.; Tinozzi, S.; Corazza, G.R. Oral Butyrate for Mildly to Moderately Active Crohn’s Disease. Aliment. Pharmacol. Ther. 2005, 22, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Assisi, R.F.; GISDI Study Group Combined Butyric Acid/Mesalazine Treatment in Ulcerative Colitis with Mild-Moderate Activity. Results of a Multicentre Pilot Study. Minerva Gastroenterol. Dietol. 2008, 54, 231–238. [Google Scholar] [PubMed]
- Vanhoutvin, S.A.L.W.; Troost, F.J.; Kilkens, T.O.C.; Lindsey, P.J.; Hamer, H.M.; Jonkers, D.M.A.E.; Venema, K.; Brummer, R.J.M. The Effects of Butyrate Enemas on Visceral Perception in Healthy Volunteers. Neurogastroenterol. Motil. 2009, 21. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.M.A.E.; Vanhoutvin, S.A.L.W.; Troost, F.J.; Rijkers, G.; de Bruïne, A.; Bast, A.; Venema, K.; Brummer, R.J.M. Effect of Butyrate Enemas on Inflammation and Antioxidant Status in the Colonic Mucosa of Patients with Ulcerative Colitis in Remission. Clin. Nutr. 2010, 29, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Banasiewicz, T.; Krokowicz, L.; Stojcev, Z.; Kaczmarek, B.F.; Kaczmarek, E.; Maik, J.; Marciniak, R.; Krokowicz, P.; Walkowiak, J.; Drews, M. Microencapsulated Sodium Butyrate Reduces the Frequency of Abdominal Pain in Patients with Irritable Bowel Syndrome. Colorectal Dis. Off. J. Assoc. Coloproctology G. B. Irel. 2013, 15, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Krokowicz, L.; Stojcev, Z.; Kaczmarek, B.F.; Kociemba, W.; Kaczmarek, E.; Walkowiak, J.; Krokowicz, P.; Drews, M.; Banasiewicz, T. Microencapsulated Sodium Butyrate Administered to Patients with Diverticulosis Decreases Incidence of Diverticulitis - A Prospective Randomized Study. Int. J. Colorectal Dis. 2014, 29, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Krokowicz, L.; Kaczmarek, B.F.; Krokowicz, P.; Stojcev, Z.; Mackiewicz, J.; Walkowiak, J.; Drews, M.; Banasiewicz, T. Sodium Butyrate and Short Chain Fatty Acids in Prevention of Travellers’ Diarrhoea: A Randomized Prospective Study. Travel Med. Infect. Dis. 2014, 12, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Maggio, A.; Magli, A.; Rancati, T.; Fiorino, C.; Valvo, F.; Fellin, G.; Ricardi, U.; Munoz, F.; Cosentino, D.; Cazzaniga, L.F.; et al. Daily Sodium Butyrate Enema for the Prevention of Radiation Proctitis in Prostate Cancer Patients Undergoing Radical Radiation Therapy: Results of a Multicenter Randomized Placebo-Controlled Dose-Finding Phase 2 Study. Int. J. Radiat. Oncol. 2014, 89, 518–524. [Google Scholar] [CrossRef]
- Luceri, C.; Femia, A.P.; Fazi, M.; Di Martino, C.; Zolfanelli, F.; Dolara, P.; Tonelli, F. Effect of Butyrate Enemas on Gene Expression Profiles and Endoscopic/Histopathological Scores of Diverted Colorectal Mucosa: A Randomized Trial. Dig. Liver Dis. 2016, 48, 27–33. [Google Scholar] [CrossRef]
- Roshanravan, N.; Mahdavi, R.; Alizadeh, E.; Jafarabadi, M.; Hedayati, M.; Ghavami, A.; Alipour, S.; Alamdari, N.; Barati, M.; Ostadrahimi, A. Effect of Butyrate and Inulin Supplementation on Glycemic Status, Lipid Profile and Glucagon-Like Peptide 1 Level in Patients with Type 2 Diabetes: A Randomized Double-Blind, Placebo-Controlled Trial. Horm. Metab. Res. 2017, 49, 886–891. [Google Scholar] [CrossRef]
- Vernero, M.; De Blasio, F.; Ribaldone, D.G.; Bugianesi, E.; Pellicano, R.; Saracco, G.M.; Astegiano, M.; Caviglia, G.P. The Usefulness of Microencapsulated Sodium Butyrate Add-On Therapy in Maintaining Remission in Patients with Ulcerative Colitis: A Prospective Observational Study. J. Clin. Med. 2020, 9, 3941. [Google Scholar] [CrossRef] [PubMed]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Buda, A.; Romualdi, C.; Pohl, D.; Perini, B.; Lorenzon, G.; Marinelli, C.; D’Incà, R.; et al. Microbiota Changes Induced by Microencapsulated Sodium Butyrate in Patients with Inflammatory Bowel Disease. Neurogastroenterol. Motil. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.F.; Nikolic, T.; Imangaliyev, S.; Bekkering, S.; Duinkerken, G.; Keij, F.M.; Herrema, H.; Winkelmeijer, M.; Kroon, J.; Levin, E.; et al. Oral Butyrate Does Not Affect Innate Immunity and Islet Autoimmunity in Individuals with Longstanding Type 1 Diabetes: A Randomised Controlled Trial. Diabetologia 2020, 63, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Coppola, S.; Nocerino, R.; Paparo, L.; Bedogni, G.; Calignano, A.; Di Scala, C.; De Giovanni Di Santa Severina, A.F.; De Filippis, F.; Ercolini, D.; Berni Canani, R. Therapeutic Effects of Butyrate on Pediatric Obesity: A Randomized Clinical Trial. JAMA Netw. Open 2022, 5, e2244912. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, A.; Banasiuk, M.; Szczepanik, M.; Borys-Iwanicka, A.; Pytrus, T.; Walkowiak, J.; Banaszkiewicz, A. Sodium Butyrate Effectiveness in Children and Adolescents with Newly Diagnosed Inflammatory Bowel Diseases—Randomized Placebo-Controlled Multicenter Trial. Nutrients 2022, 14, 3283. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, Z.; Hadi, A.; Tutunchi, H.; Asghari-Jafarabadi, M.; Naeinie, F.; Roshanravan, N.; Ostadrahimi, A.; Fadel, A. The Effects of Butyrate Supplementation on Glycemic Control, Lipid Profile, Blood Pressure, Nitric Oxide Level and Glutathione Peroxidase Activity in Type 2 Diabetic Patients: A Randomized Triple -Blind, Placebo-Controlled Trial. Clin. Nutr. ESPEN 2022, 49, 79–85. [Google Scholar] [CrossRef]
- Qaisar, R.; Karim, A.; Muhammad, T.; Ahmad, F. Butyrate Supplementation Reduces Sarcopenia by Repairing Neuromuscular Junction in Patients with Chronic Obstructive Pulmonary Disease. Respir. Med. 2024, 222, 107510. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, K.; El Abbar, F.; Dobranowski, P.; Manoogian, J.; Butcher, J.; Figeys, D.; Mack, D.; Stintzi, A. Butyrate’s Role in Human Health and the Current Progress towards Its Clinical Application to Treat Gastrointestinal Disease. Clin. Nutr. 2023, 42, 61–75. [Google Scholar] [CrossRef]
- Rhodes, J.M. Nutrition and Gut Health: The Impact of Specific Dietary Components – It’s Not Just Five-a-Day. Proc. Nutr. Soc. 2021, 80, 9–18. [Google Scholar] [CrossRef]
- Bourdu, S.; Dapoigny, M.; Chapuy, E.; Artigue, F.; Vasson, M.-P.; Dechelotte, P.; Bommelaer, G.; Eschalier, A.; Ardid, D. Rectal Instillation of Butyrate Provides a Novel Clinically Relevant Model of Noninflammatory Colonic Hypersensitivity in Rats. Gastroenterology 2005, 128, 1996–2008. [Google Scholar] [CrossRef]
- Kannampalli, P.; Shaker, R.; Sengupta, J.N. Colonic Butyrate- Algesic or Analgesic?: Colonic Butyrate Function. Neurogastroenterol. Motil. 2011, 23, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Anshory, M.; Effendi, R.M.R.A.; Kalim, H.; Dwiyana, R.F.; Suwarsa, O.; Nijsten, T.E.C.; Nouwen, J.L.; Thio, H.B. Butyrate Properties in Immune-Related Diseases: Friend or Foe? Fermentation 2023, 9, 205. [Google Scholar] [CrossRef]
- Schwarz, A.; Bruhs, A.; Schwarz, T. The Short-Chain Fatty Acid Sodium Butyrate Functions as a Regulator of the Skin Immune System. J. Invest. Dermatol. 2017, 137. [Google Scholar] [CrossRef] [PubMed]
- Stacey, S.K.; McEleney, M. Topical Corticosteroids: Choice and Application. Am. Fam. Physician 2021, 103, 337–343. [Google Scholar] [PubMed]
- Delzenne, N.M.; Williams, C.M. Prebiotics and Lipid Metabolism: Curr. Opin. Lipidol. 2002, 13, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Oba, M.; Allen, M.S. Intraruminal Infusion of Propionate Alters Feeding Behavior and Decreases Energy Intake of Lactating Dairy Cows. J. Nutr. 2003, 133, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Belzacq, A.-S.; Haouzi, D.; Rouault, A.; Kroemer, G.; Brenner, C. Propionibacteria Induce Apoptosis of Colorectal Carcinoma Cells via Short-Chain Fatty Acids Acting on Mitochondria. Cell Death Differ.
- Li, C.J.; Elsasser, T.H. Butyrate-Induced Apoptosis and Cell Cycle Arrest in Bovine Kidney Epithelial Cells: Involvement of Caspase and Proteasome Pathways1. J. Anim. Sci. 2005, 83, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kamp, F.; Hamilton, J.A. How Fatty Acids of Different Chain Length Enter and Leave Cells by Free Diffusion. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, H.W.J.; Naylor, C.P.E.; MacFarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-Chain Fatty Acids Induce Both Effector and Regulatory T Cells by Suppression of Histone Deacetylases and Regulation of the mTOR–S6K Pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Duscha, A.; Gisevius, B.; Hirschberg, S.; Yissachar, N.; Stangl, G.I.; Eilers, E.; Bader, V.; Haase, S.; Kaisler, J.; David, C.; et al. Propionic Acid Shapes the Multiple Sclerosis Disease Course by an Immunomodulatory Mechanism. Cell 2020, 180. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of Targeted Delivery of Propionate to the Human Colon on Appetite Regulation, Body Weight Maintenance and Adiposity in Overweight Adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef]
- Chambers, E.S.; Byrne, C.S.; Morrison, D.J.; Murphy, K.G.; Preston, T.; Tedford, C.; Garcia-Perez, I.; Fountana, S.; Serrano-Contreras, J.I.; Holmes, E.; et al. Dietary Supplementation with Inulin-Propionate Ester or Inulin Improves Insulin Sensitivity in Adults with Overweight and Obesity with Distinct Effects on the Gut Microbiota, Plasma Metabolome and Systemic Inflammatory Responses: A Randomised Cross-over Trial. Gut 2019, 68, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Malkova, D.; Polyviou, T.; Rizou, E.; Gerasimidis, K.; Chambers, E.S.; Preston, T.; Tedford, M.C.; Frost, G.; Morrison, D.J. Moderate Intensity Exercise Training Combined with Inulin-Propionate Ester Supplementation Increases Whole Body Resting Fat Oxidation in Overweight Women. Metabolism 2020, 104, 154043. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; Zimmermann, F.; Schumann, P.; Jasina, A.; Roessler, J.; Schmidt, D.; Heinze, P.; Kaisler, J.; Nageswaran, V.; Aigner, A.; et al. Propionate Attenuates Atherosclerosis by Immune-Dependent Regulation of Intestinal Cholesterol Metabolism. Eur. Heart J. 2021, 43, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short Chain Fatty Acids and Its Producing Organisms: An Overlooked Therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review Article: Short Chain Fatty Acids as Potential Therapeutic Agents in Human Gastrointestinal and Inflammatory Disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Offei, B.; Vandecruys, P.; De Graeve, S.; Foulquié-Moreno, M.R.; Thevelein, J.M. Unique Genetic Basis of the Distinct Antibiotic Potency of High Acetic Acid Production in the Probiotic Yeast Saccharomyces Cerevisiae Var. Boulardii. Genome Res. 2019, 29, 1478–1494. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kishi, M.; Fushimi, T.; Kaga, T. Acetic Acid Upregulates the Expression of Genes for Fatty Acid Oxidation Enzymes in Liver To Suppress Body Fat Accumulation. J. Agric. Food Chem. 2009, 57, 5982–5986. [Google Scholar] [CrossRef]
- Evaluation of Certain Food Additives: Fifty-Third Report of the Joint FAO/WHO Expert Committee on Food Additives; Joint FAO/WHO Expert Committee on Food Additives, World Health Organization, Eds.; WHO technical report series; World Health Organization: Geneva, 2000; ISBN 978-92-4-120896-3.
- Freeland, K.R.; Wolever, T.M.S. Acute Effects of Intravenous and Rectal Acetate on Glucagon-like Peptide-1, Peptide YY, Ghrelin, Adiponectin and Tumour Necrosis Factor-α. Br. J. Nutr. 2010, 103, 460–466. [Google Scholar] [CrossRef]
- Fernandes, J.; Vogt, J.; Wolever, T.M. Intravenous Acetate Elicits a Greater Free Fatty Acid Rebound in Normal than Hyperinsulinaemic Humans. Eur. J. Clin. Nutr. 2012, 66, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Van Der Beek, C.M.; Canfora, E.E.; Lenaerts, K.; Troost, F.J.; Olde Damink, S.W.M.; Holst, J.J.; Masclee, A.A.M.; Dejong, C.H.C.; Blaak, E.E. Distal, Not Proximal, Colonic Acetate Infusions Promote Fat Oxidation and Improve Metabolic Markers in Overweight/Obese Men. Clin. Sci. 2016, 130, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.M.; Lenaerts, K.; Dejong, C.H.C.; Blaak, E.E. Colonic Infusions of Short-Chain Fatty Acid Mixtures Promote Energy Metabolism in Overweight/Obese Men: A Randomized Crossover Trial. Sci. Rep. 2017, 7, 2360. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota Metabolite Short Chain Fatty Acids, GPCR, and Inflammatory Bowel Diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Blad, C.C.; Tang, C.; Offermanns, S. G Protein-Coupled Receptors for Energy Metabolites as New Therapeutic Targets. Nat. Rev. Drug Discov. 2012, 11, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Candido, E.P.; Reeves, R.; Davie, J.R. Sodium Butyrate Inhibits Histone Deacetylation in Cultured Cells. Cell 1978, 14, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Sealy, L.; Chalkley, R. The Effect of Sodium Butyrate on Histone Modification. Cell 1978, 14, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Visekruna, A.; Luu, M. The Role of Short-Chain Fatty Acids and Bile Acids in Intestinal and Liver Function, Inflammation, and Carcinogenesis. Front. Cell Dev. Biol. 2021, 9, 703218. [Google Scholar] [CrossRef]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, Neuroepigenetics and the Gut Microbiome: Can a High Fiber Diet Improve Brain Health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Chen, Y.; Ma, Z.; Zhang, X.; Shi, D.; Khan, J.A.; Liu, H. Gut Microbiota-Derived Short Chain Fatty Acids Are Potential Mediators in Gut Inflammation. Anim. Nutr. Zhongguo Xu Mu Shou Yi Xue Hui 2022, 8, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Barberio, B.; Facchin, S.; Patuzzi, I.; Ford, A.C.; Massimi, D.; Valle, G.; Sattin, E.; Simionati, B.; Bertazzo, E.; Zingone, F.; et al. A Specific Microbiota Signature Is Associated to Various Degrees of Ulcerative Colitis as Assessed by a Machine Learning Approach. Gut Microbes 14. [CrossRef]
- Segain, J.; de la Bletiere, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottiere, H.; Galmiche, J. Butyrate Inhibits Inflammatory Responses through NFκB Inhibition: Implications for Crohn’s Disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A Decrease of the Butyrate-Producing Species Roseburia Hominis and Faecalibacterium Prausnitzii Defines Dysbiosis in Patients with Ulcerative Colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; Di Stasi, L.C.; Sassaki, L.Y. Gut Microbiota, Inflammatory Bowel Disease and Colorectal Cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572.e5. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; Raingeard de la Blétière, D.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M.; Galmiche, J.P. Butyrate Inhibits Inflammatory Responses through NFkappaB Inhibition: Implications for Crohn’s Disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef]
- Zhang, Y.; Si, X.; Yang, L.; Wang, H.; Sun, Y.; Liu, N. Association between Intestinal Microbiota and Inflammatory Bowel Disease. Anim. Models Exp. Med. 2022, 5, 311–322. [Google Scholar] [CrossRef]
- Ma, J.; Piao, X.; Mahfuz, S.; Long, S.; Wang, J. The Interaction among Gut Microbes, the Intestinal Barrier and Short Chain Fatty Acids. Anim. Nutr. Zhongguo Xu Mu Shou Yi Xue Hui 2022, 9, 159–174. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, Receptor for Niacin and the Commensal Metabolite Butyrate, Suppresses Colonic Inflammation and Carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the Gut Microbiota on Host Adiposity Are Modulated by the Short-Chain Fatty-Acid Binding G Protein-Coupled Receptor, Gpr41. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of Inflammatory Responses by Gut Microbiota and Chemoattractant Receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, J.J.; Tillinger, W.; Hofer, J.; Hölzl, M.A.; Heinzl, H.; Saemann, M.D.; Zlabinger, G.J. Impaired Anti-Inflammatory Efficacy of n-Butyrate in Patients with IBD. Eur. J. Clin. Invest. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Li, A.-L.; Ni, W.-W.; Zhang, Q.-M.; Li, Y.; Zhang, X.; Wu, H.-Y.; Du, P.; Hou, J.-C.; Zhang, Y. Effect of Cinnamon Essential Oil on Gut Microbiota in the Mouse Model of Dextran Sodium Sulfate-Induced Colitis. Microbiol. Immunol. 2020, 64, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Kazemi Sefat, N.A.; Mohammadi, M.M.; Hadjati, J.; Talebi, S.; Ajami, M.; Daneshvar, H. Sodium Butyrate as a Histone Deacetylase Inhibitor Affects Toll-Like Receptor 4 Expression in Colorectal Cancer Cell Lines. Immunol. Invest. 2019, 48, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Tews, H.C.; Elger, T.; Gunawan, S.; Fererberger, T.; Sommersberger, S.; Loibl, J.; Huss, M.; Liebisch, G.; Müller, M.; Kandulski, A.; et al. Fecal Short Chain Fatty Acids and Urinary 3-Indoxyl Sulfate Do Not Discriminate between Patients with Crohn´s Disease and Ulcerative Colitis and Are Not of Diagnostic Utility for Predicting Disease Severity. Lipids Health Dis. 2023, 22, 164. [Google Scholar] [CrossRef]
- Chen, G.; Ran, X.; Li, B.; Li, Y.; He, D.; Huang, B.; Fu, S.; Liu, J.; Wang, W. Sodium Butyrate Inhibits Inflammation and Maintains Epithelium Barrier Integrity in a TNBS-Induced Inflammatory Bowel Disease Mice Model. EBioMedicine 2018, 30, 317–325. [Google Scholar] [CrossRef]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota Metabolite Butyrate Constrains Neutrophil Functions and Ameliorates Mucosal Inflammation in Inflammatory Bowel Disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium Prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [PubMed]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short Chain Fatty Acids and Its Producing Organisms: An Overlooked Therapy for IBD? eBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal Cancer. Lancet Lond. Engl. 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Hofseth, L.J.; Hebert, J.R.; Chanda, A.; Chen, H.; Love, B.L.; Pena, M.M.; Murphy, E.A.; Sajish, M.; Sheth, A.; Buckhaults, P.J.; et al. Early-Onset Colorectal Cancer: Initial Clues and Current Views. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wei, H.; Zhou, Y.; Szeto, C.-H.; Li, C.; Lin, Y.; Coker, O.O.; Lau, H.C.H.; Chan, A.W.H.; Sung, J.J.Y.; et al. High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites. Gastroenterology 2022, 162, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Kim, H.; Lee, D.H.; Lee, A.; Giovannucci, E.L.; Kang, S.-S.; Keum, N. Different Dietary Fibre Sources and Risks of Colorectal Cancer and Adenoma: A Dose-Response Meta-Analysis of Prospective Studies. Br. J. Nutr. 2019, 122, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Gianfredi, V.; Salvatori, T.; Villarini, M.; Moretti, M.; Nucci, D.; Realdon, S. Is Dietary Fibre Truly Protective against Colon Cancer? A Systematic Review and Meta-Analysis. Int. J. Food Sci. Nutr. 2018, 69, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Alvandi, E.; Wong, W.K.M.; Joglekar, M.V.; Spring, K.J.; Hardikar, A.A. Short-Chain Fatty Acid Concentrations in the Incidence and Risk-Stratification of Colorectal Cancer: A Systematic Review and Meta-Analysis. BMC Med. 2022, 20, 323. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The Microbiome and Human Cancer. Science 2021, 371, eabc4552. [Google Scholar] [CrossRef]
- van der Beek, C.M.; Dejong, C.H.C.; Troost, F.J.; Masclee, A.A.M.; Lenaerts, K. Role of Short-Chain Fatty Acids in Colonic Inflammation, Carcinogenesis, and Mucosal Protection and Healing. Nutr. Rev. 2017, 75, 286–305. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.; Greathouse, K.L. Targeting Dietary and Microbial Tryptophan-Indole Metabolism as Therapeutic Approaches to Colon Cancer. Nutrients 2021, 13, 1189. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cao, L.; Tian, Y.; Zhang, P.; Ding, C.; Lu, W.; Jia, C.; Shao, C.; Liu, W.; Wang, D.; et al. Butyrate Suppresses the Proliferation of Colorectal Cancer Cells via Targeting Pyruvate Kinase M2 and Metabolic Reprogramming. Mol. Cell. Proteomics MCP 2018, 17, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D. Diet, Microorganisms and Their Metabolites, and Colon Cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Bachem, A.; Makhlouf, C.; Binger, K.J.; de Souza, D.P.; Tull, D.; Hochheiser, K.; Whitney, P.G.; Fernandez-Ruiz, D.; Dähling, S.; Kastenmüller, W.; et al. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8+ T Cells. Immunity 2019, 51, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, B.F.; Meng, S.; Wu, J.T.; Archer, S.Y.; Hodin, R.A. The Effects of Short-Chain Fatty Acids on Human Colon Cancer Cell Phenotype Are Associated with Histone Hyperacetylation. J. Nutr. 2002, 132, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.J. Metabolism: Warburg behind the Butyrate Paradox? Nat. Rev. Cancer 2012, 12, 798. [Google Scholar] [CrossRef]
- Li, L.; Sun, Y.; Liu, J.; Wu, X.; Chen, L.; Ma, L.; Wu, P. Histone Deacetylase Inhibitor Sodium Butyrate Suppresses DNA Double Strand Break Repair Induced by Etoposide More Effectively in MCF-7 Cells than in HEK293 Cells. BMC Biochem. 2015, 16, 2. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, X.; Wang, J. Small Molecules in the Big Picture of Gut Microbiome-Host Cross-Talk. EBioMedicine 2022, 81, 104085. [Google Scholar] [CrossRef]
- Mowat, C.; Dhatt, J.; Bhatti, I.; Hamie, A.; Baker, K. Short Chain Fatty Acids Prime Colorectal Cancer Cells to Activate Antitumor Immunity. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Yang, Y.; Weng, W.; Peng, J.; Hong, L.; Yang, L.; Toiyama, Y.; Gao, R.; Liu, M.; Yin, M.; Pan, C.; et al. Fusobacterium Nucleatum Increases Proliferation of Colorectal Cancer Cells and Tumor Development in Mice by Activating Toll-Like Receptor 4 Signaling to Nuclear Factor-κB, and Up-Regulating Expression of MicroRNA-21. Gastroenterology 2017, 152, 851–866. [Google Scholar] [CrossRef] [PubMed]
- Stoilov, L.; Darroudi, F.; Meschini, R.; van der Schans, G.; Mullenders, L.H.; Natarajan, A.T. Inhibition of Repair of X-Ray-Induced DNA Double-Strand Breaks in Human Lymphocytes Exposed to Sodium Butyrate. Int. J. Radiat. Biol. 2000, 76, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, T.; Ibuki, Y. Histone Deacetylase Inhibitor Sodium Butyrate Enhances the Cell Killing Effect of Psoralen plus UVA by Attenuating Nucleotide Excision Repair. Cancer Res. 2009, 69, 3492–3500. [Google Scholar] [CrossRef] [PubMed]
- Koprinarova, M.; Botev, P.; Russev, G. Histone Deacetylase Inhibitor Sodium Butyrate Enhances Cellular Radiosensitivity by Inhibiting Both DNA Nonhomologous End Joining and Homologous Recombination. DNA Repair 2011, 10, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Robert, T.; Vanoli, F.; Chiolo, I.; Shubassi, G.; Bernstein, K.A.; Rothstein, R.; Botrugno, O.A.; Parazzoli, D.; Oldani, A.; Minucci, S.; et al. HDACs Link the DNA Damage Response, Processing of Double-Strand Breaks and Autophagy. Nature 2011, 471, 74–79. [Google Scholar] [CrossRef]
- Luu, M.; Riester, Z.; Baldrich, A.; Reichardt, N.; Yuille, S.; Busetti, A.; Klein, M.; Wempe, A.; Leister, H.; Raifer, H.; et al. Microbial Short-Chain Fatty Acids Modulate CD8+ T Cell Responses and Improve Adoptive Immunotherapy for Cancer. Nat. Commun. 2021, 12, 4077. [Google Scholar] [CrossRef]
- Mowat, C.; Mosley, S.R.; Namdar, A.; Schiller, D.; Baker, K. Anti-Tumor Immunity in Mismatch Repair-Deficient Colorectal Cancers Requires Type I IFN-Driven CCL5 and CXCL10. J. Exp. Med. 2021, 218, e20210108. [Google Scholar] [CrossRef]
- Pedrosa, L.; Esposito, F.; Thomson, T.M.; Maurel, J. The Tumor Microenvironment in Colorectal Cancer Therapy. Cancers 2019, 11, 1172. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Gebhardt, T.; Mackay, L.K. Tissue-Resident Memory T Cells in Cancer Immunosurveillance. Trends Immunol. 2019, 40, 735–747. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, Q.; Sun, L.; Ye, Y.; Ji, G. Short-Chain Fatty Acids Administration Is Protective in Colitis-Associated Colorectal Cancer Development. J. Nutr. Biochem. 2018, 57, 103–109. [Google Scholar] [CrossRef]
- Gracie, D.J.; Hamlin, P.J.; Ford, A.C. The Influence of the Brain-Gut Axis in Inflammatory Bowel Disease and Possible Implications for Treatment. Lancet Gastroenterol. Hepatol. 2019, 4, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Agirman, G.; Yu, K.B.; Hsiao, E.Y. Signaling Inflammation across the Gut-Brain Axis. Science 2021, 374, 1087–1092. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Emmi, A.; Sandre, M.; Russo, F.P.; Tombesi, G.; Garrì, F.; Campagnolo, M.; Carecchio, M.; Biundo, R.; Spolverato, G.; Macchi, V.; et al. Duodenal Alpha-Synuclein Pathology and Enteric Gliosis in Advanced Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2023, 38, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Onyango, I.G.; Jauregui, G.V.; Čarná, M.; Bennett, J.P.; Stokin, G.B. Neuroinflammation in Alzheimer’s Disease. Biomedicines 2021, 9, 524. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Liu, X. The Microbiota-Gut-Brain Axis and Neurodevelopmental Disorders. Protein Cell 2023, 14, 762–775. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Lu, T.; Chen, W.; Yan, W.; Yuan, K.; Shi, L.; Liu, X.; Zhou, X.; Shi, J.; et al. The Microbiota-Gut-Brain Axis in Sleep Disorders. Sleep Med. Rev. 2022, 65, 101691. [Google Scholar] [CrossRef] [PubMed]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef]
- Romano, S.; Savva, G.M.; Bedarf, J.R.; Charles, I.G.; Hildebrand, F.; Narbad, A. Meta-Analysis of the Parkinson’s Disease Gut Microbiome Suggests Alterations Linked to Intestinal Inflammation. NPJ Park. Dis. 2021, 7, 27. [Google Scholar] [CrossRef]
- Castelli, V.; d’Angelo, M.; Quintiliani, M.; Benedetti, E.; Cifone, M.G.; Cimini, A. The Emerging Role of Probiotics in Neurodegenerative Diseases: New Hope for Parkinson’s Disease? Neural Regen. Res. 2021, 16, 628–634. [Google Scholar] [CrossRef]
- Shannon, K.M.; Keshavarzian, A.; Dodiya, H.B.; Jakate, S.; Kordower, J.H. Is Alpha-Synuclein in the Colon a Biomarker for Premotor Parkinson’s Disease? Evidence from 3 Cases. Mov. Disord. Off. J. Mov. Disord. Soc. 2012, 27, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Brudek, T. Inflammatory Bowel Diseases and Parkinson’s Disease. J. Park. Dis. 2019, 9, S331–S344. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-Q.; Haikal, C.; Li, W.; Li, J.-Y. Gut Inflammation in Association With Pathogenesis of Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 218. [Google Scholar] [CrossRef]
- Mertsalmi, T.H.; Pekkonen, E.; Scheperjans, F. Antibiotic Exposure and Risk of Parkinson’s Disease in Finland: A Nationwide Case-Control Study. Mov. Disord. Off. J. Mov. Disord. Soc. 2020, 35, 431–442. [Google Scholar] [CrossRef]
- Challis, C.; Hori, A.; Sampson, T.R.; Yoo, B.B.; Challis, R.C.; Hamilton, A.M.; Mazmanian, S.K.; Volpicelli-Daley, L.A.; Gradinaru, V. Gut-Seeded α-Synuclein Fibrils Promote Gut Dysfunction and Brain Pathology Specifically in Aged Mice. Nat. Neurosci. 2020, 23, 327–336. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. A Timeline for Parkinson’s Disease. Parkinsonism Relat. Disord. 2010, 16, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; Bruley des Varannes, S.; Naveilhan, P.; Nguyen, J.-M.; et al. Colonic Inflammation in Parkinson’s Disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef]
- Chalazonitis, A.; Rao, M. Enteric Nervous System Manifestations of Neurodegenerative Disease. Brain Res. 2018, 1693, 207–213. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Knox, E.G.; Lynch, C.M.K.; Lee, Y.S.; O’Driscoll, C.M.; Clarke, G.; Cryan, J.F.; Aburto, M.R. The Gut Microbiota Is Important for the Maintenance of Blood–Cerebrospinal Fluid Barrier Integrity. Eur. J. Neurosci. 2023, 57, 233–241. [Google Scholar] [CrossRef]
- Sochocka, M.; Donskow-Łysoniewska, K.; Diniz, B.S.; Kurpas, D.; Brzozowska, E.; Leszek, J. The Gut Microbiome Alterations and Inflammation-Driven Pathogenesis of Alzheimer’s Disease-a Critical Review. Mol. Neurobiol. 2019, 56, 1841–1851. [Google Scholar] [CrossRef]
- Lin, C.-H.; Chen, C.-C.; Chiang, H.-L.; Liou, J.-M.; Chang, C.-M.; Lu, T.-P.; Chuang, E.Y.; Tai, Y.-C.; Cheng, C.; Lin, H.-Y.; et al. Altered Gut Microbiota and Inflammatory Cytokine Responses in Patients with Parkinson’s Disease. J. Neuroinflammation 2019, 16, 129. [Google Scholar] [CrossRef]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Glass, C.K.; Mazmanian, S.K. Microbiome-Microglia Connections via the Gut-Brain Axis. J. Exp. Med. 2019, 216, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liang, H.; Hu, Y.; Lu, L.; Zheng, C.; Fan, Y.; Wu, B.; Zou, T.; Luo, X.; Zhang, X.; et al. Gut Bacterial Profiles in Parkinson’s Disease: A Systematic Review. CNS Neurosci. Ther. 2022, 29, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Trinder, M.; Daisley, B.A.; Dube, J.S.; Reid, G. Drosophila Melanogaster as a High-Throughput Model for Host-Microbiota Interactions. Front. Microbiol. 2017, 8, 751. [Google Scholar] [CrossRef]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.-F. Impact of Microbiota on Central Nervous System and Neurological Diseases: The Gut-Brain Axis. J. Neuroinflammation 2019, 16, 53. [Google Scholar] [CrossRef]
- Stilling, R.M.; Dinan, T.G.; Cryan, J.F. Microbial Genes, Brain & Behaviour - Epigenetic Regulation of the Gut-Brain Axis. Genes Brain Behav. 2014, 13, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Galland, L. The Gut Microbiome and the Brain. J. Med. Food 2014, 17, 1261–1272. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Barnes, S.; Demark-Wahnefried, W.; Morrow, C.; Salvador, C.; Skibola, C.; Tollefsbol, T.O. Influences of Diet and the Gut Microbiome on Epigenetic Modulation in Cancer and Other Diseases. Clin. Epigenetics 2015, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The Neuropharmacology of Butyrate: The Bread and Butter of the Microbiota-Gut-Brain Axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Meng, L.; Shen, L. Multiple Roles of Short-Chain Fatty Acids in Alzheimer Disease. Nutr. Burbank Los Angel. Cty. Calif 2022, 93, 111499. [Google Scholar] [CrossRef] [PubMed]
- St Laurent, R.; O’Brien, L.M.; Ahmad, S.T. Sodium Butyrate Improves Locomotor Impairment and Early Mortality in a Rotenone-Induced Drosophila Model of Parkinson’s Disease. Neuroscience 2013, 246, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Unger, M.M.; Spiegel, J.; Dillmann, K.-U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.-H. Short Chain Fatty Acids and Gut Microbiota Differ between Patients with Parkinson’s Disease and Age-Matched Controls. Parkinsonism Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Manfready, R.A.; Forsyth, C.B.; Voigt, R.M.; Hall, D.A.; Goetz, C.G.; Keshavarzian, A. Gut-Brain Communication in Parkinson’s Disease: Enteroendocrine Regulation by GLP-1. Curr. Neurol. Neurosci. Rep. 2022, 22, 335–342. [Google Scholar] [CrossRef]
- Gerhardt, S.; Mohajeri, M.H. Changes of Colonic Bacterial Composition in Parkinson’s Disease and Other Neurodegenerative Diseases. Nutrients 2018, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Xie, A.; Ensink, E.; Li, P.; Gordevičius, J.; Marshall, L.L.; George, S.; Pospisilik, J.A.; Aho, V.T.E.; Houser, M.C.; Pereira, P.A.B.; et al. Bacterial Butyrate in Parkinson’s Disease Is Linked to Epigenetic Changes and Depressive Symptoms. Mov. Disord. Off. J. Mov. Disord. Soc. 2022, 37, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Jiang, B.; Luo, X. Gut Microbiota Influences Alzheimer’s Disease Pathogenesis by Regulating Acetate in Drosophila Model. Future Microbiol. 2018, 13, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Real, J.-M.; Serino, M.; Blasco, G.; Puig, J.; Daunis-i-Estadella, J.; Ricart, W.; Burcelin, R.; Fernández-Aranda, F.; Portero-Otin, M. Gut Microbiota Interacts With Brain Microstructure and Function. J. Clin. Endocrinol. Metab. 2015, 100, 4505–4513. [Google Scholar] [CrossRef] [PubMed]
- Müller, B.; Rasmusson, A.J.; Just, D.; Jayarathna, S.; Moazzami, A.; Novicic, Z.K.; Cunningham, J.L. Fecal Short-Chain Fatty Acid Ratios as Related to Gastrointestinal and Depressive Symptoms in Young Adults. Psychosom. Med. 2021, 83, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-Transgenic Model of Alzheimer’s Disease with Plaques and Tangles: Intracellular Abeta and Synaptic Dysfunction. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.; Feher, J.; Scuderi, G.L.; Szabo, D.; Dobolyi, A.; Cservenak, M.; Juhasz, J.; Ligeti, B.; Pongor, S.; Gomez-Cabrera, M.C.; et al. Exercise and Probiotics Attenuate the Development of Alzheimer’s Disease in Transgenic Mice: Role of Microbiome. Exp. Gerontol. 2019, 115, 122–131. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A.M. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Xi, Y.; Wang, Q.; Liu, J.; Li, P.; Meng, X.; Liu, K.; Chen, W.; Liu, X.; Liu, Z. Mannan Oligosaccharide Attenuates Cognitive and Behavioral Disorders in the 5xFAD Alzheimer’s Disease Mouse Model via Regulating the Gut Microbiota-Brain Axis. Brain. Behav. Immun. 2021, 95, 330–343. [Google Scholar] [CrossRef]
- Park, S.-H.; Lee, J.H.; Shin, J.; Kim, J.-S.; Cha, B.; Lee, S.; Kwon, K.S.; Shin, Y.W.; Choi, S.H. Cognitive Function Improvement after Fecal Microbiota Transplantation in Alzheimer’s Dementia Patient: A Case Report. Curr. Med. Res. Opin. 2021, 37, 1739–1744. [Google Scholar] [CrossRef]
- Ibrahim, A.; Ali, R.A.R.; Manaf, M.R.A.; Ahmad, N.; Tajurruddin, F.W.; Qin, W.Z.; Desa, S.H.M.; Ibrahim, N.M. Multi-Strain Probiotics (Hexbio) Containing MCP BCMC Strains Improved Constipation and Gut Motility in Parkinson’s Disease: A Randomised Controlled Trial. PloS One 2020, 15, e0244680. [Google Scholar] [CrossRef]
- Georgescu, D.; Ancusa, O.E.; Georgescu, L.A.; Ionita, I.; Reisz, D. Nonmotor Gastrointestinal Disorders in Older Patients with Parkinson’s Disease: Is There Hope? Clin. Interv. Aging 2016, 11, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhou, X.; Miao, Y.; Han, Y.; Wei, J.; Chen, T. Therapeutic Effect of GLP-1 Engineered Strain on Mice Model of Alzheimer’s Disease and Parkinson’s Disease. AMB Express 2020, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Cassani, E.; Privitera, G.; Pezzoli, G.; Pusani, C.; Madio, C.; Iorio, L.; Barichella, M. Use of Probiotics for the Treatment of Constipation in Parkinson’s Disease Patients. Minerva Gastroenterol. Dietol. 2011, 57, 117–121. [Google Scholar]
- Barichella, M.; Pacchetti, C.; Bolliri, C.; Cassani, E.; Iorio, L.; Pusani, C.; Pinelli, G.; Privitera, G.; Cesari, I.; Faierman, S.A.; et al. Probiotics and Prebiotic Fiber for Constipation Associated with Parkinson Disease: An RCT. Neurology 2016, 87, 1274–1280. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Taghizadeh, M.; Daneshvar Kakhaki, R.; Kouchaki, E.; Bahmani, F.; Borzabadi, S.; Oryan, S.; Mafi, A.; Asemi, Z. Clinical and Metabolic Response to Probiotic Administration in People with Parkinson’s Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Nutr. Edinb. Scotl. 2019, 38, 1031–1035. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.-Y.; Chong, K.K.; A Manap, M.A.A.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson Disease: A Randomized Placebo-Controlled Study. Neurology 2021, 96, e772–e782. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L.; Suescun, J.; Jiang, Z.-D.; Brown, E.L.; Essigmann, H.T.; Alexander, A.S.; DuPont, A.W.; Iqbal, T.; Utay, N.S.; Newmark, M.; et al. Fecal Microbiota Transplantation in Parkinson’s Disease—A Randomized Repeat-Dose, Placebo-Controlled Clinical Pilot Study. Front. Neurol. 2023, 14, 1104759. [Google Scholar] [CrossRef]
- Li, T.; Chu, C.; Yu, L.; Zhai, Q.; Wang, S.; Zhao, J.; Zhang, H.; Chen, W.; Tian, F. Neuroprotective Effects of Bifidobacterium Breve CCFM1067 in MPTP-Induced Mouse Models of Parkinson’s Disease. Nutrients 2022, 14, 4678. [Google Scholar] [CrossRef]
- Abdel-Haq, R.; Schlachetzki, J.C.M.; Boktor, J.C.; Cantu-Jungles, T.M.; Thron, T.; Zhang, M.; Bostick, J.W.; Khazaei, T.; Chilakala, S.; Morais, L.H.; et al. A Prebiotic Diet Modulates Microglial States and Motor Deficits in α-Synuclein Overexpressing Mice. eLife 2022, 11, e81453. [Google Scholar] [CrossRef]
- Liu, X.; Du, Z.R.; Wang, X.; Sun, X.R.; Zhao, Q.; Zhao, F.; Wong, W.T.; Wong, K.H.; Dong, X.-L. Polymannuronic Acid Prebiotic plus Lacticaseibacillus Rhamnosus GG Probiotic as a Novel Synbiotic Promoted Their Separate Neuroprotection against Parkinson’s Disease. Food Res. Int. Ott. Ont 2022, 155, 111067. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, F.; Liu, Y.; Ma, T.; Jin, H.; Quan, K.; Leng, B.; Zhao, J.; Yuan, X.; Li, Z.; et al. Probiotics Synergized with Conventional Regimen in Managing Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Lacy, B.E.; Patel, N.K. Rome Criteria and a Diagnostic Approach to Irritable Bowel Syndrome. J. Clin. Med. 2017, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Cremon, C.; Bellini, M.; Corsetti, M.; Di Nardo, G.; Falangone, F.; Fuccio, L.; Galeazzi, F.; Iovino, P.; Sarnelli, G.; et al. Italian Guidelines for the Management of Irritable Bowel Syndrome: Joint Consensus from the Italian Societies of: Gastroenterology and Endoscopy (SIGE), Neurogastroenterology and Motility (SINGEM), Hospital Gastroenterologists and Endoscopists (AIGO), Digestive Endoscopy (SIED), General Medicine (SIMG), Gastroenterology, Hepatology and Pediatric Nutrition (SIGENP) and Pediatrics (SIP). Dig. Liver Dis. 2023, 55, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Savarino, E.; Zingone, F.; Barberio, B.; Marasco, G.; Akyuz, F.; Akpinar, H.; Barboi, O.; Bodini, G.; Bor, S.; Chiarioni, G.; et al. Functional Bowel Disorders with Diarrhoea: Clinical Guidelines of the United European Gastroenterology and European Society for Neurogastroenterology and Motility. United Eur. Gastroenterol. J. 2022, 10, 556–584. [Google Scholar] [CrossRef] [PubMed]
- Tana, C.; Umesaki, Y.; Imaoka, A.; Handa, T.; Kanazawa, M.; Fukudo, S. Altered Profiles of Intestinal Microbiota and Organic Acids May Be the Origin of Symptoms in Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2010, 22, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Jia, Q.; Song, L.; Duan, L. Alterations in Fecal Short-Chain Fatty Acids in Patients with Irritable Bowel Syndrome. Medicine (Baltimore) 2019, 98, e14513. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Taverniti, V.; Gardana, C.; Cremon, C.; Canducci, F.; Pagano, I.; Barbaro, M.R.; Bellacosa, L.; Castellazzi, A.M.; Valsecchi, C.; et al. Fecal Clostridiales Distribution and Short-Chain Fatty Acids Reflect Bowel Habits in Irritable Bowel Syndrome. Environ. Microbiol. 2018, 20, 3201–3213. [Google Scholar] [CrossRef] [PubMed]
- Treem, W.R.; Ahsan, N.; Kastoff, G.; Hyams, J.S. Fecal Short-Chain Fatty Acids in Patients with Diarrhea-Predominant Irritable Bowel Syndrome: In Vitro Studies of Carbohydrate Fermentation. J. Pediatr. Gastroenterol. Nutr. 1996, 23, 280–286. [Google Scholar] [CrossRef]
- Fredericks, E.; Theunissen, R.; Roux, S. Short Chain Fatty Acids and Monocarboxylate Transporters in Irritable Bowel Syndrome. Turk. J. Gastroenterol. 2020, 31, 840–847. [Google Scholar] [CrossRef]
- Undseth, R.; Jakobsdottir, G.; Nyman, M.; Berstad, A.; Valeur, J. Low Serum Levels of Short-Chain Fatty Acids after Lactulose Ingestion May Indicate Impaired Colonic Fermentation in Patients with Irritable Bowel Syndrome. Clin. Exp. Gastroenterol. 2015, 8, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C.; Dapoigny, M.; Scott, K.P.; Crouzet, L.; Del’homme, C.; Marquet, P.; Martin, J.C.; Pickering, G.; Ardid, D.; Eschalier, A.; et al. Functional Dysbiosis within the Gut Microbiota of Patients with Constipated-Irritable Bowel Syndrome. Aliment. Pharmacol. Ther. 2012, 35, 828–838. [Google Scholar] [CrossRef]
- Pozuelo, M.; Panda, S.; Santiago, A.; Mendez, S.; Accarino, A.; Santos, J.; Guarner, F.; Azpiroz, F.; Manichanh, C. Reduction of Butyrate- and Methane-Producing Microorganisms in Patients with Irritable Bowel Syndrome. Sci. Rep. 2015, 5, 12693. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wei, H.; Zhou, A.; Xiao, X.; Xie, X.; Tang, B.; Lin, H.; Tang, L.; Meng, R.; Yuan, X.; et al. The Gut Microbiota Participates in the Effect of Linaclotide in Patients with Irritable Bowel Syndrome with Constipation (IBS-C): A Multicenter, Prospective, Pre-Post Study. J. Transl. Med. 2024, 22, 98. [Google Scholar] [CrossRef] [PubMed]
- Farup, P.G.; Rudi, K.; Hestad, K. Faecal Short-Chain Fatty Acids - a Diagnostic Biomarker for Irritable Bowel Syndrome? BMC Gastroenterol. 2016, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-Sensing Receptors GPR43 and GPR109A Facilitate Dietary Fibre-Induced Gut Homeostasis through Regulation of the Inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-Chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.H.; Boyd, K.L.; Vogel, P.; Kastan, M.B.; Lamkanfi, M.; Kanneganti, T.-D. The NLRP3 Inflammasome Protects against Loss of Epithelial Integrity and Mortality during Experimental Colitis. Immunity 2010, 32, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Hirota, S.A.; Ng, J.; Lueng, A.; Khajah, M.; Parhar, K.; Li, Y.; Lam, V.; Potentier, M.S.; Ng, K.; Bawa, M.; et al. NLRP3 Inflammasome Plays a Key Role in the Regulation of Intestinal Homeostasis. Inflamm. Bowel Dis. 2011, 17, 1359–1372. [Google Scholar] [CrossRef]
- Dunsmore, G.; Koleva, P.; Ghobakhloo, N.; Sutton, R.; Ambrosio, L.; Meng, X.; Hotte, N.; Nguyen, V.; Madsen, K.L.; Dieleman, L.A.; et al. Lower Abundance and Impaired Function of CD71+ Erythroid Cells in Inflammatory Bowel Disease Patients During Pregnancy. J. Crohns Colitis 2019, 13, 230–244. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-Chain Fatty Acids Induce Both Effector and Regulatory T Cells by Suppression of Histone Deacetylases and Regulation of the mTOR-S6K Pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C.; Chen, F.; Xiao, Y.; Zhao, Y.; Ma, C.; et al. Microbiota-Derived Short-Chain Fatty Acids Promote Th1 Cell IL-10 Production to Maintain Intestinal Homeostasis. Nat. Commun. 2018, 9, 3555. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The Regulation of Intestinal Mucin MUC2 Expression by Short-Chain Fatty Acids: Implications for Epithelial Protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef]
- Hatayama, H.; Iwashita, J.; Kuwajima, A.; Abe, T. The Short Chain Fatty Acid, Butyrate, Stimulates MUC2 Mucin Production in the Human Colon Cancer Cell Line, LS174T. Biochem. Biophys. Res. Commun. 2007, 356, 599–603. [Google Scholar] [CrossRef]
- Augenlicht, L.; Shi, L.; Mariadason, J.; Laboisse, C.; Velcich, A. Repression of MUC2 Gene Expression by Butyrate, a Physiological Regulator of Intestinal Cell Maturation. Oncogene 2003, 22, 4983–4992. [Google Scholar] [CrossRef]
- Willemsen, L.E.M.; Koetsier, M.A.; van Deventer, S.J.H.; van Tol, E. a. F. Short Chain Fatty Acids Stimulate Epithelial Mucin 2 Expression through Differential Effects on Prostaglandin E(1) and E(2) Production by Intestinal Myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef]
- Finnie, I.A.; Dwarakanath, A.D.; Taylor, B.A.; Rhodes, J.M. Colonic Mucin Synthesis Is Increased by Sodium Butyrate. Gut 1995, 36, 93–99. [Google Scholar] [CrossRef]
- Miao, W.; Wu, X.; Wang, K.; Wang, W.; Wang, Y.; Li, Z.; Liu, J.; Li, L.; Peng, L. Sodium Butyrate Promotes Reassembly of Tight Junctions in Caco-2 Monolayers Involving Inhibition of MLCK/MLC2 Pathway and Phosphorylation of PKCβ2. Int. J. Mol. Sci. 2016, 17, 1696. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Ha, S.E.; Wei, L.; Singh, R.; Zogg, H.; Clemmensen, B.; Heredia, D.J.; Gould, T.W.; Sanders, K.M.; Ro, S. Colonic Motility Is Improved by the Activation of 5-HT2B Receptors on Interstitial Cells of Cajal in Diabetic Mice. Gastroenterology 2021, 161, 608–622.e7. [Google Scholar] [CrossRef] [PubMed]
- Tharayil, V.S.; Wouters, M.M.; Stanich, J.E.; Roeder, J.L.; Lei, S.; Beyder, A.; Gomez-Pinilla, P.J.; Gershon, M.D.; Maroteaux, L.; Gibbons, S.J.; et al. Lack of Serotonin 5-HT2B Receptor Alters Proliferation and Network Volume of Interstitial Cells of Cajal in Vivo. Neurogastroenterol. Motil. 2010, 22, 462–469. [Google Scholar] [CrossRef]
- Wouters, M.M.; Gibbons, S.J.; Roeder, J.L.; Distad, M.; Ou, Y.; Strege, P.R.; Szurszewski, J.H.; Farrugia, G. Exogenous Serotonin Regulates Proliferation of Interstitial Cells of Cajal in Mouse Jejunum through 5-HT2B Receptors. Gastroenterology 2007, 133, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Karaki, S.; Kuwahara, A. Short-Chain Fatty Acids Decrease the Frequency of Spontaneous Contractions of Longitudinal Muscle via Enteric Nerves in Rat Distal Colon. Jpn. J. Physiol. 2004, 54, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Grider, J.R.; Kuemmerle, J.F.; Jin, J.G. 5-HT Released by Mucosal Stimuli Initiates Peristalsis by Activating 5-HT4/5-HT1p Receptors on Sensory CGRP Neurons. Am. J. Physiol. 1996, 270, G778–782. [Google Scholar] [CrossRef] [PubMed]
- Foxx-Orenstein, A.E.; Kuemmerle, J.F.; Grider, J.R. Distinct 5-HT Receptors Mediate the Peristaltic Reflex Induced by Mucosal Stimuli in Human and Guinea Pig Intestine. Gastroenterology 1996, 111, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, F.A.; Keenan, C.M.; Wallace, L.E.; Woods, C.; Cavin, J.-B.; Flockton, A.R.; Macklin, W.B.; Belkind-Gerson, J.; Hirota, S.A.; Sharkey, K.A. Intestinal Microbiota Shapes Gut Physiology and Regulates Enteric Neurons and Glia. Microbiome 2021, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Soret, R.; Chevalier, J.; De Coppet, P.; Poupeau, G.; Derkinderen, P.; Segain, J.P.; Neunlist, M. Short-Chain Fatty Acids Regulate the Enteric Neurons and Control Gastrointestinal Motility in Rats. Gastroenterology 2010, 138, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Suply, E.; de Vries, P.; Soret, R.; Cossais, F.; Neunlist, M. Butyrate Enemas Enhance Both Cholinergic and Nitrergic Phenotype of Myenteric Neurons and Neuromuscular Transmission in Newborn Rat Colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Shaidullov, I.F.; Sorokina, D.M.; Sitdikov, F.G.; Hermann, A.; Abdulkhakov, S.R.; Sitdikova, G.F. Short Chain Fatty Acids and Colon Motility in a Mouse Model of Irritable Bowel Syndrome. BMC Gastroenterol. 2021, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Tan, W.; Ren, H.; Yan, L.; Wang, Y.; Luo, H. The Effects of Short-Chain Fatty Acids on Rat Colonic Hypermotility Induced by Water Avoidance Stress. Drug Des. Devel. Ther. 2020, 14, 4671–4684. [Google Scholar] [CrossRef]
- Hurst, N.R.; Kendig, D.M.; Murthy, K.S.; Grider, J.R. The Short Chain Fatty Acids, Butyrate and Propionate, Have Differential Effects on the Motility of the Guinea Pig Colon. Neurogastroenterol. Motil. 2014, 26, 1586–1596. [Google Scholar] [CrossRef]
- Waseem, M.R.; Shin, A.; Siwiec, R.; James-Stevenson, T.; Bohm, M.; Rogers, N.; Wo, J.; Waseem, L.; Gupta, A.; Jarrett, M.; et al. Associations of Fecal Short Chain Fatty Acids With Colonic Transit, Fecal Bile Acid, and Food Intake in Irritable Bowel Syndrome. Clin. Transl. Gastroenterol. 2023, 14, e00541. [Google Scholar] [CrossRef] [PubMed]
- Day, E.A.; Ford, R.J.; Steinberg, G.R. AMPK as a Therapeutic Target for Treating Metabolic Diseases. Trends Endocrinol. Metab. TEM 2017, 28, 545–560. [Google Scholar] [CrossRef]
- Dabke, K.; Hendrick, G.; Devkota, S. The Gut Microbiome and Metabolic Syndrome. J. Clin. Invest. 2019, 129, 4050–4057. [Google Scholar] [CrossRef] [PubMed]
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. A Multisociety Delphi Consensus Statement on New Fatty Liver Disease Nomenclature. J. Hepatol. 2023, 79, 1542–1556. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Waalen, J. The Genetics of Human Obesity. Transl. Res. J. Lab. Clin. Med. 2014, 164, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, S.; Ulloa-Martínez, M.; Martínez-Rojano, H.; Galván-Rodríguez, F.M.; Miranda-Brito, C.; Romano, M.C.; Piña-Escobedo, A.; Pizano-Zárate, M.L.; Hoyo-Vadillo, C.; García-Mena, J. Study of the Diversity and Short-Chain Fatty Acids Production by the Bacterial Community in Overweight and Obese Mexican Children. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2015, 34, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Ecklu-Mensah, G.; Choo-Kang, C.; Maseng, M.G.; Donato, S.; Bovet, P.; Viswanathan, B.; Bedu-Addo, K.; Plange-Rhule, J.; Oti Boateng, P.; Forrester, T.E.; et al. Gut Microbiota and Fecal Short Chain Fatty Acids Differ with Adiposity and Country of Origin: The METS-Microbiome Study. Nat. Commun. 2023, 14, 5160. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in Lean and Overweight Healthy Subjects. Obes. Silver Spring Md 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.S.; Comelli, E.M. Adiposity, Gut Microbiota and Faecal Short Chain Fatty Acids Are Linked in Adult Humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed]
- Tirosh, A.; Calay, E.S.; Tuncman, G.; Claiborn, K.C.; Inouye, K.E.; Eguchi, K.; Alcala, M.; Rathaus, M.; Hollander, K.S.; Ron, I.; et al. The Short-Chain Fatty Acid Propionate Increases Glucagon and FABP4 Production, Impairing Insulin Action in Mice and Humans. Sci. Transl. Med. 2019, 11, eaav0120. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate Mediates a Microbiome-Brain-β Cell Axis Promoting Metabolic Syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, T.F.S.; Grześkowiak, Ł.; Franceschini, S.C.C.; Bressan, J.; Ferreira, C.L.L.F.; Peluzio, M.C.G. Higher Level of Faecal SCFA in Women Correlates with Metabolic Syndrome Risk Factors. Br. J. Nutr. 2013, 109, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Royall, D.; Wolever, T.M.; Jeejeebhoy, K.N. Clinical Significance of Colonic Fermentation. Am. J. Gastroenterol. 1990, 85, 1307–1312. [Google Scholar] [PubMed]
- Reshef, L.; Niv, J.; Shapiro, B. Effect of Propionate on Lipogenesis in Adipose Tissue. J. Lipid Res. 1967, 8, 682–687. [Google Scholar] [CrossRef]
- Wolever, T.M.; Brighenti, F.; Royall, D.; Jenkins, A.L.; Jenkins, D.J. Effect of Rectal Infusion of Short Chain Fatty Acids in Human Subjects. Am. J. Gastroenterol. 1989, 84, 1027–1033. [Google Scholar]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PloS One 2012, 7, e35240. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The Short-Chain Fatty Acid Acetate Reduces Appetite via a Central Homeostatic Mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short Chain Fatty Acids Prevent High-Fat-Diet-Induced Obesity in Mice by Regulating G Protein-Coupled Receptors and Gut Microbiota. Sci. Rep. 2016, 6, 37589. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.S.; Kao, C.H.J.; Xu, Y.; Glucina, M.P.; Paterson, R.R.M.; Ferguson, L.R. From 2000years of Ganoderma Lucidum to Recent Developments in Nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.; Guo, C.; Guo, D.; Wu, J.; Wang, Y.; Wang, Y.; Chen, J.; Chen, C.; Wu, K.; Na, K.; et al. Suppression of Obesity and Inflammation by Polysaccharide from Sporoderm-Broken Spore of Ganoderma Lucidum via Gut Microbiota Regulation. Carbohydr. Polym. 2021, 256, 117594. [Google Scholar] [CrossRef] [PubMed]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Pedersen, H.K.; et al. Disentangling Type 2 Diabetes and Metformin Treatment Signatures in the Human Gut Microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Jocken, J.W.E.; González Hernández, M.A.; Hoebers, N.T.H.; van der Beek, C.M.; Essers, Y.P.G.; Blaak, E.E.; Canfora, E.E. Short-Chain Fatty Acids Differentially Affect Intracellular Lipolysis in a Human White Adipocyte Model. Front. Endocrinol. 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The Impact of Short-Chain Fatty Acids on GLP-1 and PYY Secretion from the Isolated Perfused Rat Colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef] [PubMed]
- Veprik, A.; Laufer, D.; Weiss, S.; Rubins, N.; Walker, M.D. GPR41 Modulates Insulin Secretion and Gene Expression in Pancreatic β-Cells and Modifies Metabolic Homeostasis in Fed and Fasting States. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2016, 30, 3860–3869. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Villa, S.R.; Fuller, M.; Wicksteed, B.; Mackay, C.R.; Alquier, T.; Poitout, V.; Mancebo, H.; Mirmira, R.G.; Gilchrist, A.; et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. Baltim. Md 2015, 29, 1055–1066. [Google Scholar] [CrossRef]
- Fushimi, T.; Tayama, K.; Fukaya, M.; Kitakoshi, K.; Nakai, N.; Tsukamoto, Y.; Sato, Y. Acetic Acid Feeding Enhances Glycogen Repletion in Liver and Skeletal Muscle of Rats. J. Nutr. 2001, 131, 1973–1977. [Google Scholar] [CrossRef]
- Li, H.; Gao, Z.; Zhang, J.; Ye, X.; Xu, A.; Ye, J.; Jia, W. Sodium Butyrate Stimulates Expression of Fibroblast Growth Factor 21 in Liver by Inhibition of Histone Deacetylase 3. Diabetes 2012, 61, 797–806. [Google Scholar] [CrossRef]
- Li, X.; Chen, H.; Guan, Y.; Li, X.; Lei, L.; Liu, J.; Yin, L.; Liu, G.; Wang, Z. Acetic Acid Activates the AMP-Activated Protein Kinase Signaling Pathway to Regulate Lipid Metabolism in Bovine Hepatocytes. PloS One 2013, 8, e67880. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Maruta, H.; Jozuka, M.; Kimura, R.; Iwabuchi, H.; Yamato, M.; Saito, T.; Fujisawa, K.; Takahashi, Y.; Kimoto, M.; et al. Effects of Acetate on Lipid Metabolism in Muscles and Adipose Tissues of Type 2 Diabetic Otsuka Long-Evans Tokushima Fatty (OLETF) Rats. Biosci. Biotechnol. Biochem. 2009, 73, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H. Biological Function of Acetic Acid-Improvement in Obesity and Glucose Tolerance by Acetic Acid in Type 2 Diabetic Rats. Crit. Rev. Food Sci. Nutr. 2016, 56 Suppl 1, S171–175. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Generated Metabolites Promote Metabolic Benefits via Gut-Brain Neural Circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Vily-Petit, J.; Soty, M.; Silva, M.; Micoud, M.; Bron, C.; Guérin-Deremaux, L.; Mithieux, G. Improvement of Energy Metabolism Associated with NUTRIOSE® Soluble Fiber, a Dietary Ingredient Exhibiting Prebiotic Properties, Requires Intestinal Gluconeogenesis. Food Res. Int. Ott. Ont 2023, 167, 112723. [Google Scholar] [CrossRef]
- Liu, L.; Fu, C.; Li, F. Acetate Affects the Process of Lipid Metabolism in Rabbit Liver, Skeletal Muscle and Adipose Tissue. Anim. Open Access J. MDPI 2019, 9, 799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lou, H.; Peng, Y.; Chen, S.; Zhang, Y.; Li, X. Comprehensive Relationships between Gut Microbiome and Faecal Metabolome in Individuals with Type 2 Diabetes and Its Complications. Endocrine 2019, 66, 526–537. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human Gut Microbes Impact Host Serum Metabolome and Insulin Sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Perera, D.; Kleinstein, S.E.; Hanson, B.; Hasturk, H.; Eveloff, R.; Freire, M.; Ramsey, M. Impaired Host Response and the Presence of Acinetobacter Baumannii in the Serum Microbiome of Type-II Diabetic Patients. iScience 2021, 24, 101941. [Google Scholar] [CrossRef] [PubMed]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut Microbial Metabolites Limit the Frequency of Autoimmune T Cells and Protect against Type 1 Diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef]
- Sanna, S.; van Zuydam, N.R.; Mahajan, A.; Kurilshikov, A.; Vich Vila, A.; Võsa, U.; Mujagic, Z.; Masclee, A.A.M.; Jonkers, D.M.A.E.; Oosting, M.; et al. Causal Relationships among the Gut Microbiome, Short-Chain Fatty Acids and Metabolic Diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Ayesha, I.E.; Monson, N.R.; Klair, N.; Patel, U.; Saxena, A.; Patel, D.; Venugopal, S. Probiotics and Their Role in the Management of Type 2 Diabetes Mellitus (Short-Term Versus Long-Term Effect): A Systematic Review and Meta-Analysis. Cureus 15, e46741. [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of Targeted Delivery of Propionate to the Human Colon on Appetite Regulation, Body Weight Maintenance and Adiposity in Overweight Adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Preston, T.; Tedford, C.; Brignardello, J.; Garcia-Perez, I.; Holmes, E.; Wallis, G.A.; Morrison, D.J.; Frost, G.S. Effects of Inulin Propionate Ester Incorporated into Palatable Food Products on Appetite and Resting Energy Expenditure: A Randomised Crossover Study. Nutrients 2019, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- Bouter, K.; Bakker, G.J.; Levin, E.; Hartstra, A.V.; Kootte, R.S.; Udayappan, S.D.; Katiraei, S.; Bahler, L.; Gilijamse, P.W.; Tremaroli, V.; et al. Differential Metabolic Effects of Oral Butyrate Treatment in Lean versus Metabolic Syndrome Subjects. Clin. Transl. Gastroenterol. 2018, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of Intestinal Microbiota from Lean Donors Increases Insulin Sensitivity in Individuals with Metabolic Syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L. The Microbiome and Risk for Obesity and Diabetes. JAMA 2017, 317, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, X.; Adams, H.; Kubena, K.; Guo, S. Etiology of Metabolic Syndrome and Dietary Intervention. Int. J. Mol. Sci. 2018, 20, 128. [Google Scholar] [CrossRef]
- Mouzaki, M.; Comelli, E.M.; Arendt, B.M.; Bonengel, J.; Fung, S.K.; Fischer, S.E.; McGilvray, I.D.; Allard, J.P. Intestinal Microbiota in Patients with Nonalcoholic Fatty Liver Disease. Hepatol. Baltim. Md 2013, 58, 120–127. [Google Scholar] [CrossRef]
- Ding, Y.; Yanagi, K.; Cheng, C.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Interactions between Gut Microbiota and Non-Alcoholic Liver Disease: The Role of Microbiota-Derived Metabolites. Pharmacol. Res. 2019, 141, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Schoeler, M.; Caesar, R. Dietary Lipids, Gut Microbiota and Lipid Metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef]
- Xie, C.; Halegoua-DeMarzio, D. Role of Probiotics in Non-Alcoholic Fatty Liver Disease: Does Gut Microbiota Matter? Nutrients 2019, 11, 2837. [Google Scholar] [CrossRef]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary Cholesterol Drives Fatty Liver-Associated Liver Cancer by Modulating Gut Microbiota and Metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Rau, M.; Rehman, A.; Dittrich, M.; Groen, A.K.; Hermanns, H.M.; Seyfried, F.; Beyersdorf, N.; Dandekar, T.; Rosenstiel, P.; Geier, A. Fecal SCFAs and SCFA-Producing Bacteria in Gut Microbiome of Human NAFLD as a Putative Link to Systemic T-Cell Activation and Advanced Disease. United Eur. Gastroenterol. J. 2018, 6, 1496–1507. [Google Scholar] [CrossRef] [PubMed]
- Aragonès, G.; Colom-Pellicer, M.; Aguilar, C.; Guiu-Jurado, E.; Martínez, S.; Sabench, F.; Antonio Porras, J.; Riesco, D.; Del Castillo, D.; Richart, C.; et al. Circulating Microbiota-Derived Metabolites: A "liquid Biopsy? Int. J. Obes. 2005 2020, 44, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Adler, G.K.; Hornik, E.S.; Murray, G.; Bhandari, S.; Yadav, Y.; Heydarpour, M.; Basu, R.; Garg, R.; Tirosh, A. Acute Effects of the Food Preservative Propionic Acid on Glucose Metabolism in Humans. BMJ Open Diabetes Res. Care 2021, 9, e002336. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Chen, X.; Zhao, Z.; Liao, Y.; Zhou, T.; Xiang, Q. A Potential Link between Plasma Short-Chain Fatty Acids, TNF-α Level and Disease Progression in Non-Alcoholic Fatty Liver Disease: A Retrospective Study. Exp. Ther. Med. 2022, 24, 598. [Google Scholar] [CrossRef]
- Behary, J.; Amorim, N.; Jiang, X.-T.; Raposo, A.; Gong, L.; McGovern, E.; Ibrahim, R.; Chu, F.; Stephens, C.; Jebeili, H.; et al. Gut Microbiota Impact on the Peripheral Immune Response in Non-Alcoholic Fatty Liver Disease Related Hepatocellular Carcinoma. Nat. Commun. 2021, 12, 187. [Google Scholar] [CrossRef]
- Tsai, H.-J.; Hung, W.-C.; Hung, W.-W.; Lee, Y.-J.; Chen, Y.-C.; Lee, C.-Y.; Tsai, Y.-C.; Dai, C.-Y. Circulating Short-Chain Fatty Acids and Non-Alcoholic Fatty Liver Disease Severity in Patients with Type 2 Diabetes Mellitus. Nutrients 2023, 15, 1712. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Nogal, A.; Louca, P.; Zhang, X.; Wells, P.M.; Steves, C.J.; Spector, T.D.; Falchi, M.; Valdes, A.M.; Menni, C. Circulating Levels of the Short-Chain Fatty Acid Acetate Mediate the Effect of the Gut Microbiome on Visceral Fat. Front. Microbiol. 2021, 12, 711359. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-Invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Thing, M.; Werge, M.P.; Kimer, N.; Hetland, L.E.; Rashu, E.B.; Nabilou, P.; Junker, A.E.; Galsgaard, E.D.; Bendtsen, F.; Laupsa-Borge, J.; et al. Targeted Metabolomics Reveals Plasma Short-Chain Fatty Acids Are Associated with Metabolic Dysfunction-Associated Steatotic Liver Disease. BMC Gastroenterol. 2024, 24, 43. [Google Scholar] [CrossRef] [PubMed]
- Dangana, E.O.; Omolekulo, T.E.; Areola, E.D.; Olaniyi, K.S.; Soladoye, A.O.; Olatunji, L.A. Sodium Acetate Protects against Nicotine-Induced Excess Hepatic Lipid in Male Rats by Suppressing Xanthine Oxidase Activity. Chem. Biol. Interact. 2020, 316, 108929. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.J.; Sellmann, C.; Engstler, A.J.; Ziegenhardt, D.; Bergheim, I. Supplementation of Sodium Butyrate Protects Mice from the Development of Non-Alcoholic Steatohepatitis (NASH). Br. J. Nutr. 2015, 114, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Fan, J.-G. Microbial Metabolites in Non-Alcoholic Fatty Liver Disease. World J. Gastroenterol. 2019, 25, 2019–2028. [Google Scholar] [CrossRef]
- Deng, M.; Qu, F.; Chen, L.; Liu, C.; Zhang, M.; Ren, F.; Guo, H.; Zhang, H.; Ge, S.; Wu, C.; et al. SCFAs Alleviated Steatosis and Inflammation in Mice with NASH Induced by MCD. J. Endocrinol. 2020, 245, 425–437. [Google Scholar] [CrossRef]
- Zhao, Z.-H.; Wang, Z.-X.; Zhou, D.; Han, Y.; Ma, F.; Hu, Z.; Xin, F.-Z.; Liu, X.-L.; Ren, T.-Y.; Zhang, F.; et al. Sodium Butyrate Supplementation Inhibits Hepatic Steatosis by Stimulating Liver Kinase B1 and Insulin-Induced Gene. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 857–871. [Google Scholar] [CrossRef]
- Zhou, D.; Pan, Q.; Xin, F.-Z.; Zhang, R.-N.; He, C.-X.; Chen, G.-Y.; Liu, C.; Chen, Y.-W.; Fan, J.-G. Sodium Butyrate Attenuates High-Fat Diet-Induced Steatohepatitis in Mice by Improving Gut Microbiota and Gastrointestinal Barrier. World J. Gastroenterol. 2017, 23, 60–75. [Google Scholar] [CrossRef]
- Liu, W.; Luo, X.; Tang, J.; Mo, Q.; Zhong, H.; Zhang, H.; Feng, F. A Bridge for Short-Chain Fatty Acids to Affect Inflammatory Bowel Disease, Type 1 Diabetes, and Non-Alcoholic Fatty Liver Disease Positively: By Changing Gut Barrier. Eur. J. Nutr. 2021, 60, 2317–2330. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, I.; Paramsothy, S.; Doron, I.; Semon, A.; Kaakoush, N.O.; Clemente, J.C.; Faith, J.J.; Borody, T.J.; Mitchell, H.M.; Colombel, J.-F.; et al. Fungal Trans-Kingdom Dynamics Linked to Responsiveness to Fecal Microbiota Transplantation (FMT) Therapy in Ulcerative Colitis. Cell Host Microbe 2020, 27, 823–829.e3. [Google Scholar] [CrossRef] [PubMed]
- de Groot, P.; Scheithauer, T.; Bakker, G.J.; Prodan, A.; Levin, E.; Khan, M.T.; Herrema, H.; Ackermans, M.; Serlie, M.J.M.; de Brauw, M.; et al. Donor Metabolic Characteristics Drive Effects of Faecal Microbiota Transplantation on Recipient Insulin Sensitivity, Energy Expenditure and Intestinal Transit Time. Gut 2020, 69, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.F.; Kelly, C.R.; Kao, D.; Vaughn, B.P.; Lebwohl, B.; Singh, S.; Imdad, A.; Altayar, O. AGA Clinical Practice Guideline on Fecal Microbiota–Based Therapies for Select Gastrointestinal Diseases. Gastroenterology 2024, 166, 409–434. [Google Scholar] [CrossRef]
- Mullish, B.H.; Quraishi, M.N.; Segal, J.P.; McCune, V.L.; Baxter, M.; Marsden, G.L.; Moore, D.J.; Colville, A.; Bhala, N.; Iqbal, T.H.; et al. The Use of Faecal Microbiota Transplant as Treatment for Recurrent or Refractory Clostridium Difficile Infection and Other Potential Indications: Joint British Society of Gastroenterology (BSG) and Healthcare Infection Society (HIS) Guidelines. Gut 2018, 67, 1920–1941. [Google Scholar] [CrossRef] [PubMed]
- Gweon, T.-G.; Lee, Y.J.; Kim, K.O.; Yim, S.K.; Soh, J.S.; Kim, S.Y.; Park, J.J.; Shin, S.Y.; Lee, T.H.; Choi, C.H.; et al. Clinical Practice Guidelines for Fecal Microbiota Transplantation in Korea. J. Neurogastroenterol. Motil. 2022, 28, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Ianiro, G.; Tilg, H.; Rajilić-Stojanović, M.; Kump, P.; Satokari, R.; Sokol, H.; Arkkila, P.; Pintus, C.; Hart, A.; et al. European Consensus Conference on Faecal Microbiota Transplantation in Clinical Practice. Gut 2017, 66, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Barberio, B.; Facchin, S.; Mele, E.; D’Incà, R.; Sturniolo, G.C.; Farinati, F.; Zingone, F.; Quagliariello, A.; Ghisa, M.; Massimi, D.; et al. Faecal Microbiota Transplantation in Clostridioides Difficile Infection: Real-Life Experience from an Academic Italian Hospital. Ther. Adv. Gastroenterol. 2020, 13, 1756284820934315. [Google Scholar] [CrossRef]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective Effects of Fecal Microbiota Transplantation on MPTP-Induced Parkinson’s Disease Mice: Gut Microbiota, Glial Reaction and TLR4/TNF-α Signaling Pathway. Brain. Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Smits, L.P.; Bouter, K.E.C.; de Vos, W.M.; Borody, T.J.; Nieuwdorp, M. Therapeutic Potential of Fecal Microbiota Transplantation. Gastroenterology 2013, 145, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, J.-Y.; Li, Q.-H.; Su, H.; Sun, X. Lactobacillus Rhamnosus GG Induced Protective Effect on Allergic Airway Inflammation Is Associated with Gut Microbiota. Cell. Immunol. 2018, 332, 77–84. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Hausken, T.; Hatlebakk, J.G. Increasing the Dose and/or Repeating Faecal Microbiota Transplantation (FMT) Increases the Response in Patients with Irritable Bowel Syndrome (IBS). Nutrients 2019, 11, 1415. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.C.; Derraik, J.G.B.; Albert, B.B.; Leong, K.S.W.; Tweedie-Cullen, R.Y.; Creagh, C.; Depczynski, M.; Edwards, T.; Vatanen, T.; Thabrew, H.; et al. An Open-Label Pilot Trial of Faecal Microbiome Transfer to Restore the Gut Microbiome in Anorexia Nervosa: Protocol. BMJ Open 2023, 13, e070616. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Støving, R.K.; Berreira Ibraim, S.; Hyötyläinen, T.; Thirion, F.; Arora, T.; Lyu, L.; Stankevic, E.; Hansen, T.H.; Déchelotte, P.; et al. The Gut Microbiota Contributes to the Pathogenesis of Anorexia Nervosa in Humans and Mice. Nat. Microbiol. 2023, 8, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lv, L.; Wang, C. Efficacy of Fecal Microbiota Transplantation in Irritable Bowel Syndrome: A Meta-Analysis of Randomized Controlled Trials. Front. Cell. Infect. Microbiol. 2022, 12, 827395. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.C.; Vatanen, T.; Cutfield, W.S.; O’Sullivan, J.M. The Super-Donor Phenomenon in Fecal Microbiota Transplantation. Front. Cell. Infect. Microbiol. 2019, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Dorsaz, S.; Charretier, Y.; Girard, M.; Gaïa, N.; Leo, S.; Schrenzel, J.; Harbarth, S.; Huttner, B.; Lazarevic, V. Changes in Microbiota Profiles After Prolonged Frozen Storage of Stool Suspensions. Front. Cell. Infect. Microbiol. 2020, 10, 77. [Google Scholar] [CrossRef]
- Varga, A.; Kocsis, B.; Sipos, D.; Kása, P.; Vigvári, S.; Pál, S.; Dembrovszky, F.; Farkas, K.; Péterfi, Z. How to Apply FMT More Effectively, Conveniently and Flexible – A Comparison of FMT Methods. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated With Response to Fecal Microbiota Transplantation in Patients With Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454. [Google Scholar] [CrossRef]
- Joseph, J.; Depp, C.; Shih, P.-A.B.; Cadenhead, K.S.; Schmid-Schönbein, G. Modified Mediterranean Diet for Enrichment of Short Chain Fatty Acids: Potential Adjunctive Therapeutic to Target Immune and Metabolic Dysfunction in Schizophrenia? Front. Neurosci. 2017, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Guo, R.; Wang, W.; Ju, Y.; Wang, Q.; Ma, Q.; Sun, Q.; Fan, Y.; Xie, Y.; Yang, Z.; et al. Transplantation of Microbiota from Drug-Free Patients with Schizophrenia Causes Schizophrenia-like Abnormal Behaviors and Dysregulated Kynurenine Metabolism in Mice. Mol. Psychiatry 2020, 25, 2905–2918. [Google Scholar] [CrossRef]
- Osaki, H.; Jodai, Y.; Koyama, K.; Omori, T.; Horiguchi, N.; Kamano, T.; Funasaka, K.; Nagasaka, M.; Nakagawa, Y.; Shibata, T.; et al. Clinical Response and Changes in the Fecal Microbiota and Metabolite Levels after Fecal Microbiota Transplantation in Patients with Inflammatory Bowel Disease and Recurrent Clostridioides Difficile Infection. Fujita Med. J. 2021, 7, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Seekatz, A.M.; Theriot, C.M.; Rao, K.; Chang, Y.-M.; Freeman, A.E.; Kao, J.Y.; Young, V.B. Restoration of Short Chain Fatty Acid and Bile Acid Metabolism Following Fecal Microbiota Transplantation in Patients with Recurrent Clostridium Difficile Infection. Anaerobe 2018, 53, 64–73. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Valeur, J.; Hausken, T.; Gunnar Hatlebakk, J. Changes in Fecal Short-Chain Fatty Acids Following Fecal Microbiota Transplantation in Patients with Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2021, 33, e13983. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Shock, T.; Badang, L.; Ferguson, B.; Martinez-Guryn, K. The Interplay between Diet, Gut Microbes, and Host Epigenetics in Health and Disease. J. Nutr. Biochem. 2021, 95, 108631. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.L.; Weir, T.L. The Gut Microbiota at the Intersection of Diet and Human Health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Castro-Barquero, S.; Ruiz-León, A.M.; Sierra-Pérez, M.; Estruch, R.; Casas, R. Dietary Strategies for Metabolic Syndrome: A Comprehensive Review. Nutrients 2020, 12, 2983. [Google Scholar] [CrossRef]
- Shi, H.; Ge, X.; Ma, X.; Zheng, M.; Cui, X.; Pan, W.; Zheng, P.; Yang, X.; Zhang, P.; Hu, M.; et al. A Fiber-Deprived Diet Causes Cognitive Impairment and Hippocampal Microglia-Mediated Synaptic Loss through the Gut Microbiota and Metabolites. Microbiome 2021, 9, 223. [Google Scholar] [CrossRef]
- Ristori, M.V.; Quagliariello, A.; Reddel, S.; Ianiro, G.; Vicari, S.; Gasbarrini, A.; Putignani, L. Autism, Gastrointestinal Symptoms and Modulation of Gut Microbiota by Nutritional Interventions. Nutrients 2019, 11, 2812. [Google Scholar] [CrossRef] [PubMed]
- Adolph, T.E.; Zhang, J. Diet Fuelling Inflammatory Bowel Diseases: Preclinical and Clinical Concepts. Gut 2022, 71, 2574–2586. [Google Scholar] [CrossRef] [PubMed]
- Opie, R.S.; O’Neil, A.; Jacka, F.N.; Pizzinga, J.; Itsiopoulos, C. A Modified Mediterranean Dietary Intervention for Adults with Major Depression: Dietary Protocol and Feasibility Data from the SMILES Trial. Nutr. Neurosci. 2018, 21, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Pylkas, A.M.; Juneja, L.R.; Slavin, J.L. Comparison of Different Fibers for in Vitro Production of Short Chain Fatty Acids by Intestinal Microflora. J. Med. Food 2005, 8, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Vinelli, V.; Biscotti, P.; Martini, D.; Del Bo’, C.; Marino, M.; Meroño, T.; Nikoloudaki, O.; Calabrese, F.M.; Turroni, S.; Taverniti, V.; et al. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients 2022, 14, 2559. [Google Scholar] [CrossRef] [PubMed]
- Bonazzi, E.; Bretin, A.; Vigué, L.; Hao, F.; Patterson, A.D.; Gewirtz, A.T.; Chassaing, B. Individualized Microbiotas Dictate the Impact of Dietary Fiber on Colitis Sensitivity. Microbiome 2024, 12, 5. [Google Scholar] [CrossRef]
- Gudan, A.; Skonieczna-Żydecka, K.; Palma, J.; Drozd, A.; Stachowska, E. Effects of Dietary Components on Intestinal Short-Chain Fatty Acids (SCFAs) Synthesis in Healthy Adult Persons Following a Ketogenic Diet. Rocz. Panstw. Zakl. Hig. 2022, 73, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Seethaler, B.; Nguyen, N.K.; Basrai, M.; Kiechle, M.; Walter, J.; Delzenne, N.M.; Bischoff, S.C. Short-Chain Fatty Acids Are Key Mediators of the Favorable Effects of the Mediterranean Diet on Intestinal Barrier Integrity: Data from the Randomized Controlled LIBRE Trial. Am. J. Clin. Nutr. 2022, 116, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; Macia, L.; Mackay, C.R. Dietary Fiber and SCFAs in the Regulation of Mucosal Immunity. J. Allergy Clin. Immunol. 2023, 151, 361–370. [Google Scholar] [CrossRef]
- François, I.E.J.A.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a Wheat Bran Extract Containing Arabinoxylan Oligosaccharides on Gastrointestinal Health Parameters in Healthy Adult Human Volunteers: A Double-Blind, Randomised, Placebo-Controlled, Cross-over Trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.C.; Johansson-Boll, E.V.; Björck, I.M.E. Increased Gut Hormones and Insulin Sensitivity Index Following a 3-d Intervention with a Barley Kernel-Based Product: A Randomised Cross-over Study in Healthy Middle-Aged Subjects. Br. J. Nutr. 2015, 114, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ames, N.P.; Tun, H.M.; Tosh, S.M.; Jones, P.J.; Khafipour, E. High Molecular Weight Barley β-Glucan Alters Gut Microbiota Toward Reduced Cardiovascular Disease Risk. Front. Microbiol. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Farup, P.G.; Valeur, J. Changes in Faecal Short-Chain Fatty Acids after Weight-Loss Interventions in Subjects with Morbid Obesity. Nutrients 2020, 12, 802. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion Mechanisms Mediated by Probiotics and Prebiotics and Their Potential Impact on Human Health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.-M.; Dequenne, I.; de Timary, P.; Cani, P.D. How Probiotics Affect the Microbiota. Front. Cell. Infect. Microbiol. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.-W.; Li, R.-Q.; An, J.-X.; Xie, T.-Q.; Han, Z.-Y.; Xu, R.; Fang, Y.; Zhang, X.-Z. Prebiotics-Encapsulated Probiotic Spores Regulate Gut Microbiota and Suppress Colon Cancer. Adv. Mater. Deerfield Beach Fla 2020, 32, e2004529. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota-Gut-Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide Is a Prebiotic That Reduces Desulfovibrio Bacteria, Increases Cecal Short-Chain Fatty Acids, and Is Anti-Inflammatory in Mice. Am. J. Physiol. - Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef] [PubMed]
- Holmes, Z.C.; Villa, M.M.; Durand, H.K.; Jiang, S.; Dallow, E.P.; Petrone, B.L.; Silverman, J.D.; Lin, P.-H.; David, L.A. Microbiota Responses to Different Prebiotics Are Conserved within Individuals and Associated with Habitual Fiber Intake. Microbiome 2022, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Feng, Q.-Q.; Ojo, O.O.; Wang, X.-H. The Role of Dietary Fibre in Modulating Gut Microbiota Dysbiosis in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2020, 12, 3239. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, E.; Gharagozlian, S.; Birkeland, K.I.; Valeur, J.; Måge, I.; Rud, I.; Aas, A.-M. Prebiotic Effect of Inulin-Type Fructans on Faecal Microbiota and Short-Chain Fatty Acids in Type 2 Diabetes: A Randomised Controlled Trial. Eur. J. Nutr. 2020, 59, 3325–3338. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, E.; Gharagozlian, S.; Gulseth, H.L.; Birkeland, K.I.; Hartmann, B.; Holst, J.J.; Holst, R.; Aas, A.-M. Effects of Prebiotics on Postprandial GLP-1, GLP-2 and Glucose Regulation in Patients with Type 2 Diabetes: A Randomised, Double-Blind, Placebo-Controlled Crossover Trial. Diabet. Med. J. Br. Diabet. Assoc. 2021, 38, e14657. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.L.; Alvarado, D.A.; Swanson, K.S.; Holscher, H.D. The Prebiotic Potential of Inulin-Type Fructans: A Systematic Review. Adv. Nutr. Bethesda Md 2022, 13, 492–529. [Google Scholar] [CrossRef]
- Li, L.; Li, P.; Xu, L. Assessing the Effects of Inulin-Type Fructan Intake on Body Weight, Blood Glucose, and Lipid Profile: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Food Sci. Nutr. 2021, 9, 4598–4616. [Google Scholar] [CrossRef] [PubMed]
- Łoniewski, I.; Szulińska, M.; Kaczmarczyk, M.; Podsiadło, K.; Styburski, D.; Skonieczna-Żydecka, K.; Bogdański, P. Multispecies Probiotic Affects Fecal Short-Chain Fatty Acids in Postmenopausal Women with Obesity: A Post Hoc Analysis of a Randomized, Double-Blind, Placebo-Controlled Study. Nutrition 2023, 114, 112109. [Google Scholar] [CrossRef]
- Rad, Z.A.; Mousavi, S.N.; Chiti, H. A Low-Carb Diet Increases Fecal Short-Chain Fatty Acids in Feces of Obese Women Following a Weight-Loss Program: Randomized Feeding Trial. Sci. Rep. 2023, 13, 18146. [Google Scholar] [CrossRef]
- Cong, J.; Zhou, P.; Zhang, R. Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease. Nutrients 2022, 14, 1977. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, H.K.; Bording-Jorgensen, M.; Santer, D.M.; Zhang, Z.; Valcheva, R.; Rieger, A.M.; Sung-Ho Kim, J.; Dijk, S.I.; Mahmood, R.; Ogungbola, O.; et al. Unfermented β-Fructan Fibers Fuel Inflammation in Select Inflammatory Bowel Disease Patients. Gastroenterology 2023, 164, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Valcheva, R.; Armstrong, H.; Kovic, O.; Bording-Jorgensen, M.; Veniamin, S.; Pérez-Muñoz, M.E.; Haskey, N.; Silva, M.; Peerani, F.; Wong, K.; et al. Double Blind Placebo-Controlled Trial for the Prevention of Ulcerative Colitis Relapses by β-Fructan Prebiotics: Efficacy and Metabolomic Analysis; Gastroenterology, 2022.
- Gulliver, E.L.; Young, R.B.; Chonwerawong, M.; D’Adamo, G.L.; Thomason, T.; Widdop, J.T.; Rutten, E.L.; Rossetto Marcelino, V.; Bryant, R.V.; Costello, S.P.; et al. Review Article: The Future of Microbiome-based Therapeutics. Aliment. Pharmacol. Ther. 2022, 56, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Khan, N.A. Microbiome and One Health: Potential of Novel Metabolites from the Gut Microbiome of Unique Species for Human Health. Microorganisms 2023, 11, 481. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Production and absorption of SCFAs acetate, propionate, and butyrate in the human intestine. Effect on gastrointestinal and metabolic health.
Figure 1.
Production and absorption of SCFAs acetate, propionate, and butyrate in the human intestine. Effect on gastrointestinal and metabolic health.
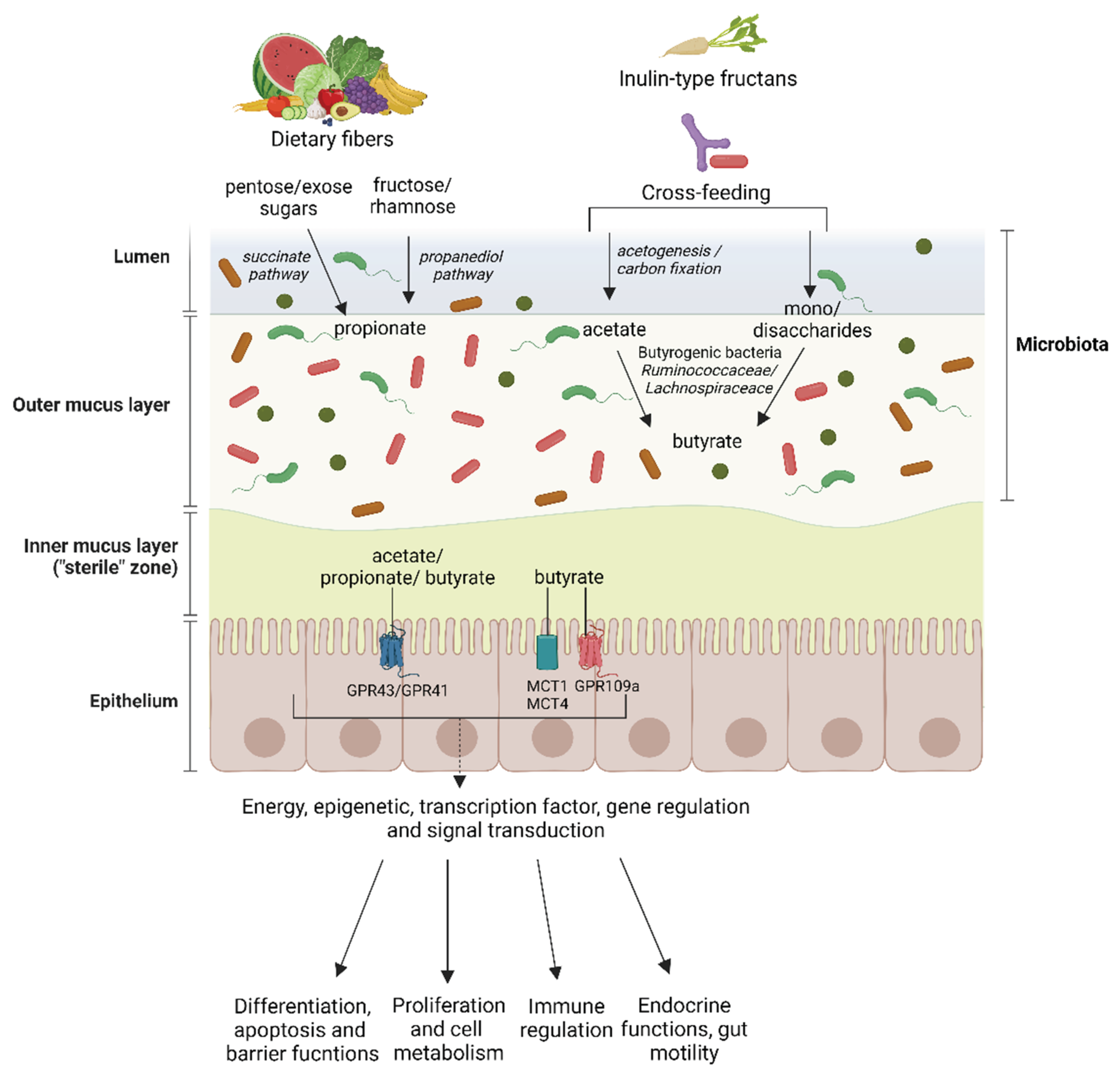
Figure 2.
Mechanism by which SCFAs exert their effects on target cells. SCFAs enter cells via MCT and transporters located on the cell membrane. Once inside the cell nucleus, they inhibit HDAC and activate HAT, facilitating histone acetylation. This process gradually relaxes compacted chromosomes, ultimately resulting in increased gene expression. Additionally, upon entering colonocytes, SCFAs may undergo beta-oxidation and enter the mitochondria, where the citric acid cycle (also known as the Krebs cycle) generates energy for the cell. Another mechanism involves SCFAs binding to GPCR, such as GPR43, GPR41, or GPR109A, on the cell membrane of both colonocytes and immune cells. This interaction inhibits downstream signaling pathways, including NF-κB, Akt, MAPK, and mTOR, while activating the AMPK pathway. Consequently, this regulates gene transcription and translation, leading to inflammation mitigation, oxidative stress reduction, and autophagy enhancement. AKT refers to the activation of a serine/threonine kinase; NF-κB to nuclear factor-κB; AMPK to adenosine 5’-monophosphate (AMP)-activated protein kinase; MAPK to mitogen-activated protein kinase; NLRP3 to nucleotide-binding oligomerization domain (NOD), leucine-rich repeat (LRR)-containing proteins (NLR); mTOR to mammalian target of rapamycin. The figure was created using BioRender.com.
Figure 2.
Mechanism by which SCFAs exert their effects on target cells. SCFAs enter cells via MCT and transporters located on the cell membrane. Once inside the cell nucleus, they inhibit HDAC and activate HAT, facilitating histone acetylation. This process gradually relaxes compacted chromosomes, ultimately resulting in increased gene expression. Additionally, upon entering colonocytes, SCFAs may undergo beta-oxidation and enter the mitochondria, where the citric acid cycle (also known as the Krebs cycle) generates energy for the cell. Another mechanism involves SCFAs binding to GPCR, such as GPR43, GPR41, or GPR109A, on the cell membrane of both colonocytes and immune cells. This interaction inhibits downstream signaling pathways, including NF-κB, Akt, MAPK, and mTOR, while activating the AMPK pathway. Consequently, this regulates gene transcription and translation, leading to inflammation mitigation, oxidative stress reduction, and autophagy enhancement. AKT refers to the activation of a serine/threonine kinase; NF-κB to nuclear factor-κB; AMPK to adenosine 5’-monophosphate (AMP)-activated protein kinase; MAPK to mitogen-activated protein kinase; NLRP3 to nucleotide-binding oligomerization domain (NOD), leucine-rich repeat (LRR)-containing proteins (NLR); mTOR to mammalian target of rapamycin. The figure was created using BioRender.com.
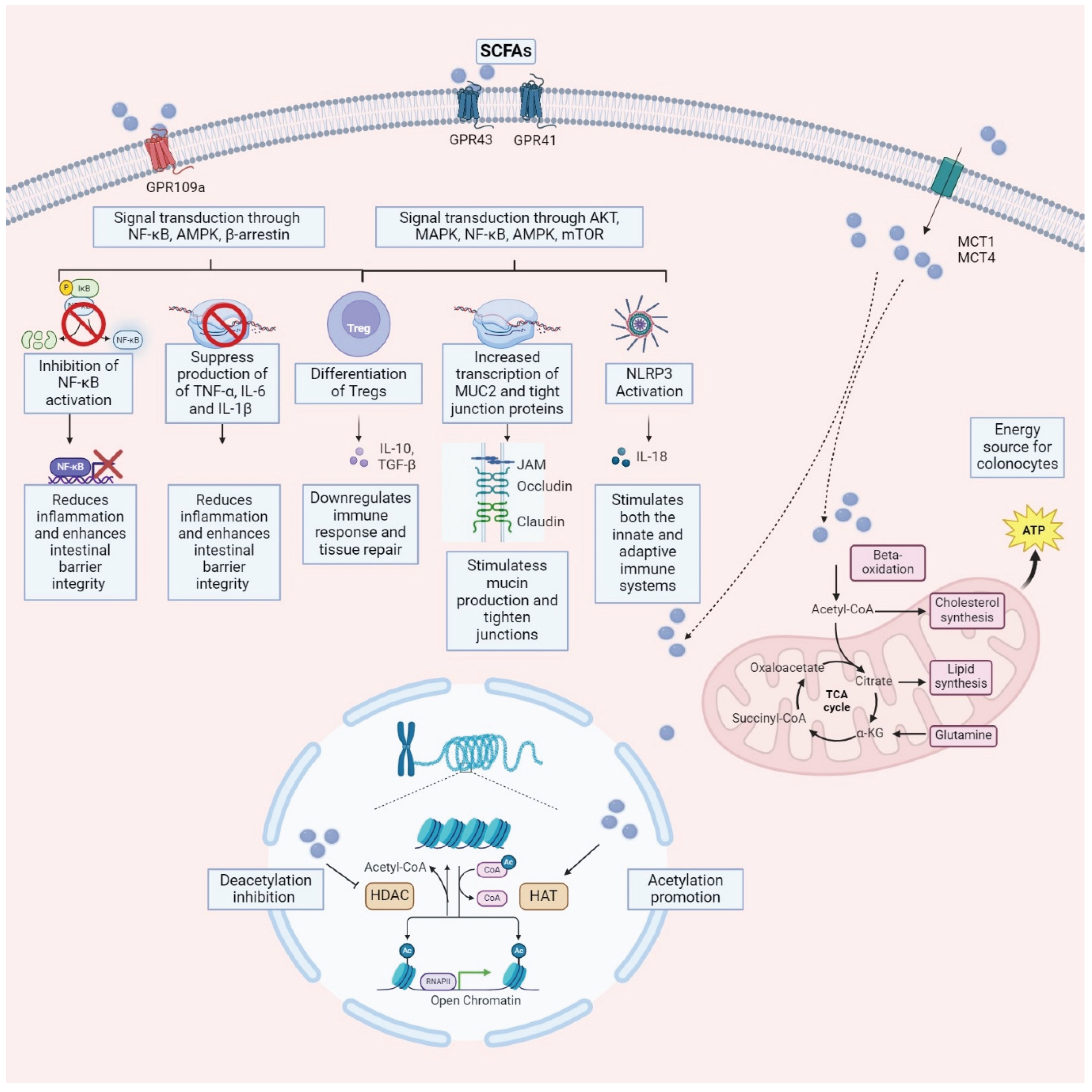

Table 1.
Production of the three different SCFA forms different pathways and by other intestinal bacteria.
Table 1.
Production of the three different SCFA forms different pathways and by other intestinal bacteria.
| SCFA | Metabolic pathway | Bacteria involved in the production |
| acetate | Acetogenesis | Acetobacterium, Acetoanaerobium, Acetogenium, Butyrbacterium, Clostridium, Eubacterium, Pelobacter |
| Carbon fixation | Bacteroides succinogenes, Clostridium butyricum, Syntophomonas sp. | |
| Propionate | Succinate | Firmicutes (Bacteroidetes and Negativicutes) |
| Propanediol | Lachnospiraceae (Roseburia inlinivorans, Balutia sp.) | |
| Butyrate | Butyryl-CoA : acetate-CoA transferase | Eubacterium, Roseburia, Anaerostipes, Faecalibacterium prausnitzii |
| Butyrate kinase | specific Coprococcus species and Clostridium species |
Table 2.
Effects of butyrate interventions in IBD and non-IBD conditions. Abbreviations: s= significative improvement, nr= Information not reported, std = Standard therapy, ns=Not significative improvement, ps=Partial significative effect DC=Diverting Colitis, DB=Double Blind, SB=Single Blind; UC=Ulcerative Colitis; CD= Crohn disease, A-S= Mesalamine+Sulfalazine, CRP=Cronic Radiation Proctitis, ARP=Acute Radiation Proctitis, DM=Diabetes mellitus, DT1=Type 1 diabetes, DT2=14Type 2 diabetes , Ob ped=Obese pediatrics, COPD=Chronic obstructive pulmonary disease, not impr.= not improvement.
Table 2.
Effects of butyrate interventions in IBD and non-IBD conditions. Abbreviations: s= significative improvement, nr= Information not reported, std = Standard therapy, ns=Not significative improvement, ps=Partial significative effect DC=Diverting Colitis, DB=Double Blind, SB=Single Blind; UC=Ulcerative Colitis; CD= Crohn disease, A-S= Mesalamine+Sulfalazine, CRP=Cronic Radiation Proctitis, ARP=Acute Radiation Proctitis, DM=Diabetes mellitus, DT1=Type 1 diabetes, DT2=14Type 2 diabetes , Ob ped=Obese pediatrics, COPD=Chronic obstructive pulmonary disease, not impr.= not improvement.
| Ref. | Delivery | Year | Groups(n) | Design | Duration | Dosage Butyrate | Drugs(ad) | Improvement |
|---|---|---|---|---|---|---|---|---|
| [72] | enema | 1991 | DC (13) | DB | 2w | 40mmol/Lit | nr | ns |
| [73] | enema | 1992 | UC (10) | SB crossover | 2w | 100mmol/Lit | A-S | s |
| [74] | enema | 1994 | UC (10) | open label | 6w | 80mmol/Lit | A-S | 60% |
| [75] | enema | 1995 | UC (40) | DB-RCT | 6w | 200ml/d mix | A-S | s |
| [76] | enema | 1996 | UC (38) | RCT | 6w | 80mmol/d | A/S | not improve. |
| [77] | enema | 1996 | UC (47) | DB-RCT | 6w | 80mmol/d | nr | not impr. |
| [78] | enema | 1999 | CRP (17) | DB-RCT | 5w | 80mmol/d | nr | s |
| [79] | enema | 2000 | UC (30) | RCT | 6w | 4gr/d | A | ns |
| [80] | enema | 2000 | APR (20) | RCT crossover | 3w | 80mmol/Lit | nr | s |
| [81] | enema | 2002 | UC (11) | RCT | 8w | 100mM | A/S/steroid | s |
| [82] | enema | 2003 | UC (51) | DB-RCT | 6w | 80mmol/Lit | M/steroid | S |
| [83] | oral | 2005 | CD (13) | open label | 8w | 4gr/d | A/S | 69% |
| [84] | oral | 2008 | UC (216) | open label | 24w | 921mg/d | A+S | 82,4% |
| [85] | enema | 2009 | IBS (11) | DB-RCT | 1w | 50/100mmol/Lit/d | nr | s |
| [86] | enema | 2010 | UC (35) | DB-RCT crossover | 20d | 100mmol/d | nr | s |
| [87] | oral | 2013 | IBS (66) | RCT | 12w | 300mg/d | std9 | s |
| [88] | oral | 2014 | DC (63) | RCT | 12month | 300mg/d | nr | s |
| [89] | oral | 2014 | TD (42) | RCT | 3d+trip | 1500mg/d | various | s |
| [90] | enema | 2014 | APR (166) | RCT | 3w | 1-2-4gr/d | nr | ns |
| [91] | enema | 2016 | Mix (20) | DB-RCT | 4w | 600mmol/Lit | nr | s |
| [92] | oral | 2017 | DM (40) | DB-RCT | 45d | 600mg/d | +inulin | s(+ inulin) |
| [93] | oral | 2020 | UC (39) | Prospective | 12months | 1g/d | std | s |
| [94] | oral | 2020 | IBD (49) | DB-RCT | 8w | 600mg/d | std | ps |
| [95] | oral | 2020 | DT1 (30) | DB-RCT | 4w | 4g/d | nr | no |
| [96] | oral | 2022 | Ob ped (54) | QB-RCT | 13 months | 20mg/kg | std | s |
| [97] | oral | 2022 | IBD ped (80) | RCT | 12w | 150mg/d | std | no |
| [98] | oral | 2022 | DT2 (42) | TB-RCT | 6w | 600mg/d | nr | s |
| [99] | oral | 2024 | COPD (121) | RCT | 12w | 300mg/d | nr | s |
Table 3.
Effects of propionate interventions. Abbreviations: s=significative improvement, ACVD=Atherosclerotic cardiovascular disease, HovF=Healthy overweight females, MS= multiple sclerosis IPE= inulin-propionate ester, DB=Double Blind, NaP= sodium-propionate.
Table 3.
Effects of propionate interventions. Abbreviations: s=significative improvement, ACVD=Atherosclerotic cardiovascular disease, HovF=Healthy overweight females, MS= multiple sclerosis IPE= inulin-propionate ester, DB=Double Blind, NaP= sodium-propionate.
| Ref. | Delivery | Year | Groups(n) | Design | Duration | Dosage Propionate | Formula | Improvement |
|---|---|---|---|---|---|---|---|---|
| [115] | oral | 2015 | Obese(60) | DB-RCT | 24w | 10g/d | IPE | s |
| [116] | oral | 2019 | Obese(12) | DB-RCT cross over | 42d | 20g/d | IPE | s |
| [117] | oral | 2019 | HovF(20) | RCT | 4w | 10g/d | IPE | s |
| [114] | oral | 2020 | MS(36)/Healthy(68) | proof-of-concept | 2w | 1g/d | NaP | s |
| [118] | oral | 2022 | ACVD(62) | DB-RCT | 8w | 1g/d | propionic acid | s |
Table 3.
a. Currently recruiting and upcoming clinical trials examining the effects of propionate.
| Name of trial | Type | Identifier/status | condition | intervention | Location | |
|---|---|---|---|---|---|---|
| Combination of Medium Cut-off Dialyzer Membrane and Diet Modification to Alleviate Residual Uremic Syndrome of Dialysis Patients | RCT | NCT04247867/recruiting | Uremic syndrome | Psyllium-inulin/ sodium propionate | University medical Centre Ljubljana, Ljubljana, Slovenia | |
| The Effect of Combining Medium Cut-Off Dialysis Membrane and Diet Modification on Reducing Inflammation Response | RCT | NCT04247867/recruiting | Uremic syndrome | Psyllium-inulin/ sodium propionate | University medical Centre Ljubljana, Ljubljana, Slovenia |
Table 4.
Effects of acetate interventions. Abbreviations: HinsF= Hyperinsulinemic females.
| Ref. | Delivery | Year | Groups(n) | Design | Duration | Dosage Propionate | Formula | Improvement |
|---|---|---|---|---|---|---|---|---|
| [124] | Rectally and intravenous | 2010 | HinsF(6) | open label | 4 times | 60mmol/lit rectal+20mmon/Lit intravenous | NaAcetate | s |
| [125] | Intravenous | 2012 | Overweight normoglycemic and hyperglycemic subjects (9) | open label | 90 | 140mmol/Lit | NaAcetate | no |
| [126] | Proximal and distal colonic | 2016 | Obese(6) | DB-RCT crossover | 3d | 100-180mmol/lit | Acetate | s |
| [127] | Colonic infusions | 2017 | Obese(12) | DB-RCT cross over | 4d | 200mmol/lit mix | (acetate, propionate, and butyrate) | s |
Table 5.
Mechanisms of SCFA in gastrointestinal diseases.
| Disease | Supposed Mechanisms of SCFA Protection/Risk |
|---|---|
| Inflammatory bowel disease | 1. Anti-inflammatory effects: Butyrate, a primary energy source for colonocytes, inhibits NF-κB activation, reducing proinflammatory gene expression. 2. Maintenance of gut barrier integrity: SCFAs promote mucus production and tighten epithelial cell junctions, enhancing the intestinal epithelial barrier. 3. Modulation of immune responses: SCFAs influence the differentiation and function of regulatory T cells (Tregs), suppressing excessive immune reactions. They engage with receptors like GPR43 and GPR109A to stimulate Treg production 4. Tissue repair and healing: SCFAs promote the proliferation and differentiation of epithelial cells, facilitating tissue repair processes within the gut damaged by inflammation in IBD. |
| Colon cancer | 1. Protective effects against CRC development: SCFAs exert protective effects against colorectal cancer (CRC) by regulating gene expression, promoting apoptosis, and inhibiting CRC cell proliferation and metabolism. 2. Anti-inflammatory actions: SCFAs mitigate inflammation in CRC by inhibiting NF-κB activation, decreasing pro-inflammatory cytokine expression, and promoting anti-inflammatory cytokines and regulatory T-cell differentiation. 3. Potential DNA damage modulation: While SCFAs are anticipated to decrease DNA damage in CRC cells, reports suggest they may exacerbate DNA damage accumulation in some instances, possibly due to disruptions in DNA repair mechanisms. Further evidence is needed. |
| Disorders of the Gut-Brain Axis | 1. Neuroprotective effects: SCFAs exert neuroprotective effects by influencing brain function, regulating blood flow, and modulating neuroinflammation via interactions with specific receptors and epigenetic modulation. 2. Role in neurodegenerative diseases: Reduced SCFA levels are implicated in neurodegenerative diseases like Alzheimer's and Parkinson's. They contribute to intestinal barrier impairment, the release of pro-inflammatory molecules, and microglial activation, ultimately impacting disease progression. 3. Gut barrier function and motility: SCFAs promote mucus secretion and strengthen intestinal tight junctions, improving barrier integrity. SCFAs can influence nerve activity, neurotransmitters, and muscle contractions. |
Table 6.
Mechanisms of SCFA in metabolic diseases.
| Disease | Supposed Mechanisms of SCFA Protection/Risk |
| Obesity | 1. Appetite Regulation: SCFAs can stimulate the release of Peptide YY (PYY) and glucagon-like peptide-1 (GLP-1) from gut endocrine cells. These hormones act centrally in the hypothalamus to signal satiety and decrease appetite. 2. Fat Storage and Metabolism: Increased SCFA-mediated adipocyte activity might favor fat storage in subcutaneous adipose tissue. SCFAs might enhance brown adipose tissue (BAT) activity, promoting thermogenesis and potentially increasing energy expenditure. 3. Metabolic effects: SCFAs activate GPR41 and GPR43 receptors on fat and immune cells, potentially influencing insulin sensitivity, fat metabolism, inflammation, and, thus, weight regulation. |
| Type 2 diabetes | 1. Effects on glucose metabolism: SCFAsact as secretagogues for hormones like GLP-1 and PYY, which enhance satiety and decrease appetite. GLP-1 enhances insulin secretion from the pancreas and reduces glucagon secretion, lowering blood sugar levels. In the liver, SCFAs inhibit glycolysis and gluconeogenesis, promoting glycogen synthesis and fatty acid oxidation. In skeletal muscle and adipose tissue, they improve glucose uptake and glycogen synthesis. 2. Role in intestinal gluconeogenesis (IGN): SCFAs promote IGN production, which is crucial for glucose and energy homeostasis. 3. Gut Health: SCFAs promote a healthy gut environment, which may be linked to a lower risk of developing diabetes. |
| Metabolic dysfunction–associated steatotic liver disease | 1. Improved Insulin Sensitivity: SCFAs can activate GPR43 on adipocytes and hepatocytes. GPR43 activation can stimulate insulin signaling pathways, leading to increased glucose uptake by these cells and potentially improving overall insulin sensitivity. SCFAs might also suppress gluconeogenesis in the liver. 2. Anti-inflammatory Effects: SCFAs can modulate the activity of immune cells like macrophages in the liver. They might suppress pro-inflammatory cytokine production (e.g., TNF-α, IL-6) and promote the activity of regulatory T cells, creating an anti-inflammatory environment. SCFAs inhibit the NF-κB signaling pathway, a key player in inflammatory responses. 3. Gut-Liver Axis: SCFAs might also influence Fibroblast Growth Factor (FGF) signaling pathways in the gut-liver axis, potentially impacting bile acid metabolism and hepatocyte function. SCFAs might stimulate the enterohepatic circulation of bile acids. SCFA-mediated bile acid signaling can activate FXR, a nuclear receptor in the liver, potentially influencing hepatic lipid metabolism and reducing steatosis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



