
Submitted:
26 March 2024
Posted:
27 March 2024
You are already at the latest version
Abstract
Calicioids form a research field that has encompassed ascomycetous fungi with stalked ascomata similar to those of the lichen genus Calicium. Early generic circumscriptions in calicioid lichens and fungi were mainly based on morphological and secondary chemistry information. After the introduction of molecular data, taxonomy in the group has been reconsidered. Here, based on a broad geographical sampling, Coniocybe Ach. was revised using molecular and morphological features. Three loci (ITS, LSU and rpb1) were compared to infer its phylogenetic position, and a total of 52 new sequences (14 ITS, 24 LSU and 14 rpb1) were produced. Coniocybe was revised and emended to include, apart for its type C. furfuracea, also C. brachypoda and C. confusa. In addition, a new species Coniocybe eufuracea, was described, and a key to the species of Coniocybe provided.
Keywords:
taxonomy
; molecular phylogeny
; nomenclature
; new species
1. Introduction
Calicioid fungi has since long been a research field of considerable interest. It includes the systematics of fungi having ascomata similar to those of Calicium Pers., i.e., stalked ascomata with a distinct head.
The investigation of calicioids dates back to the seminal papers of Acharius 1815-1817 [1] and has further been pursued by distinguished lichenologists such as Vainio 1927 [2], Keissler 1936-1938 [3] and Nádvorník 1942a, 1942b [4,5]. Only with the works of Schmidt 1970 [6] it became obvious that calicioids were quite a heterogenous assemblage, both in morphology and ecology, and that although the majority were lichenized and mazaediate (i.e., having prototuniate asci and passive spore dispersal) others were not. The diversity of calicioids was further exposed by Tibell 1984 [7]. Genetic data subsequently substantiated these observations and showed calicioids to have a variety of phylogenetic origins. In consequence a number of only distantly related higher taxa were recognized [7,8,9,10], Coniocybales being one of them. Coniocybaceae Rchb. in Coniocybales [9] comprise about 30 species in two genera, Chaenotheca (Th. Fr.) Th. Fr. and Sclerophora Chevall. [9,11]. Sclerophora was earlier treated in a separate family Sclerophoraceae [7]. Chaenotheca are crustose lichens with stalked apothecia [12] and (mostly) non-septate, spherical to ellipsoidal pale brown to brown spores.
Coniocybe Ach.: Fr. was described by Acharius [1]. The genus in Acharius’ circumscription included Mucor furfuraceum (=C. furfuracea (L.) Ach.) - in fact the only crustose calicioid described by Linnaeus - and also C. stilbea Ach. (an illegitimate name [13]), C. brachypoda Ach., and C. gracilenta (Ach.) Ach. Coniocybe was accepted by E. Fries [15] and also by Th. M. Fries and Zahlbruckner [16,17], as in contrast to Chaenotheca having a poorly developed excipulum. For many decades the genus was maintained until a major revision of the taxonomy of calicioids were undertaken by Tibell [7]. Taxonomy at that time relied solely on morphologic and chemistry data, and based on this Coniocybe brachypoda and C. furfuracea were transferred to Chaenotheca. This wider concept of Chaenotheca will henceforth be called Chaenotheca s. lat. Somewhat later, a species very similar to C. furfuracea, was described under the name Chaenotheca confusa Tibell [18].
Molecular studies have proven increasingly important in elucidating taxonomic relationships of calicioid lichens by inference of their phylogeny [9,19]. In a recent study based on Internal Transcribed Spacer (ITS) sequences it was shown that, within Chaenotheca s. lat., there are well characterized clades [19] which were also given preliminary nicknames. Thus C. brachypoda and C. furfuracea were shown to belong to a group referred to as ‘Coniocybe s. str.’. The species at that time included have slender, yellow-pruinose ascomata (Figure 3) and are associated with Stichococcus. That study was limited to comparisons of species mainly from Europe. The inclusion of C. brachypoda, C. furfuracea and C. confusa in a resurrected Coniocybe as proposed below is a first step towards re-evaluating the taxonomy of Chaenotheca s.lat., in a context of a wider sampling of material.
The main focus of this study is the emendation of Coniocybe utilizing a combination of molecular and morphological features based on a wide geographical sampling.
2. Materials and Methods
2.1. Taxon Sampling
This study is partly based on African materials collected jointly by the authors in the Kilimanjaro and Arusha regions in 2018, while some other material was collected in Australia (Tasmania), in addition to a wide geographical representation of the species, mainly vouchered by herbarium material kept in UPS.
2.2. Morphological Features
Ascomata anatomy was observed on freezing microtome sections 10 to 15 μm thick and on squash preparations under the light microscope. The sections were mounted in water. The spores of some specimens were investigated under the scanning electron microscope (SEM).
When statistical estimates of sizes are given the following format is applied: mean (X) minus one standard deviation/ (sd) / mean plus one standard deviation, number of observations (n), number of specimens observed (c).
2.3. Molecular Methods
Total DNA was extracted from freshly collected material, or material temporarily kept at -20 °C. by using the DNeasy Plant Mini Kit (Quiagen, Hilden, Germany), following the manufacturer’s instructions. About 10 to 30 apothecia were carefully selected under a dissecting microscope, cleaned from foreign material and used for DNA extraction. Total DNA was used for PCR amplifications with the following primers ITS1F [20], ITS4 [21] for ITS; LROR and LR5 [22] for the partial 28S large subunit rDNA (LSU), and gRPB1-A and gRPB1-C for partial RNA polymerase II largest subunit RPB1 (rpb1) [23]. The AccuPower PCR PreMix (Bioneer, Daejeon, Korea) was used, adding 3 µl diluted DNA, 1.5 ml of each primer (10 mM), and water to a total volume of 20 µl. The PCR conditions for ITS and LSU were: initial denaturation for 4 min at 95 ºC, followed by 35 cycles of 1min at 94 ºC, 1min at 54 ºC, 45 s. at 72 ºC, and final elongation for 5 min at 72 ºC. For rpb1, PCR amplifications were carried out using Illustra Hot Start PCR beads under the same PCR conditions. PCR products were visualized by electrophoresis on 1.5% agarose gels. Products were purified using Illustra™ ExoStar buffer diluted 10×, following the manufacturer’s protocol. Sequencing was conducted by Macrogen (www.macrogen.com).
2.4. Sequence Alignments and Phylogenetic Analysis
Selected DNA sequences of ITS, LSU and rpb1 were downloaded from GenBank and, along with the newly produced sequences used in this study (Table 1; new sequences in bold), assembled and edited using AliView [24]. After assessment of their quality, the sequences were aligned by using MAFFT v.7 (on-line server: https//mafft.cbrc.jp/alignment/server/), with G-INS-1 Strategy (Slow; progressive method) and default parameters.
The study involved two datasets. The larger dataset consists of three marker region (ITS, LSU, rpb1) sequences representing 15 species of Chaenotheca s. lat. (with newly produced sequences in bold, Table 1). They represented the clades already demonstrated and nicknamed by Tibell et al. [19]. Sclerophora farinacea was chosen as outgroup for the analyses. The second dataset, a smaller dataset, only included sequences for the species of Coniocybe s. str., and here Chaenotheca biesboschii was chosen as outgroup.
For both datasets phylogenetic relationships and their posterior probabilities (PP) were inferred using a Bayesian approach, and additional support values were estimated using Maximum Likelihood Bootstrap Support (MLbs). For the Bayesian analyses, the most likely models of evolution were estimated using the Akaike Information Criterion (AIC) as implemented in Modeltest 3.7 [25]. For the first dataset, the GTR + I + G model of evolution was employed for ITS and LSU, and HKY + I + G was used for rpb1. For the second dataset, the GTR + G model was implemented for ITS. A conflict among single-locus data sets was considered significant if a well-supported monophyletic group (posterior probability [PP] ≥0.95) was found to be well supported as non-monophyletic when different loci were used. Further analyses were performed after concatenation using Sequence Matrix [26].
The Bayesian analysis was executed using MrBayes 3.2.6 [27], where two analyses of two parallel runs were carried out for 10 M generations. Each run included four chains, and trees were sampled every 1000 generations and 25% were discarded as burn-in. All runs converged on the same average likelihood score and topology. Maximum Likelihood (ML) estimates were carried out by RAxML version 8.2.10 using the GTR + G + I model of site substitution [28]. The branch support was acquired by maximum likelihood bootstrapping (MLbs) of 1000 replicates [29], and MLbs ≥ 70% were considered to be significant. The trees were visualized in FigTree version 1.3.1 [30].
3. Results
3.1. Phylogeny of Chaenotheca s. lat.
A phylogeny of Chaenotheca s. lat., based on concatenation of the three loci of species representing the different clades provisionally named in Tibell et al. [19], is presented in Figure 1. The analyses included 12 species of Chaenotheca s. lat. There was no conflict among the trees obtained for individual locus.
In this phylogeny, Coniocybe is distinct from the clades of Chaenotheca s. lat. and it has maximum support in Bayesian and ML analyses. It is close to the ‘gracillima group’ in agreement with the results of Tibell et al. [19].
S. coniophaea. The two support values associated with each internal branch corre- spond to posterior probability (PP) and bootstrap support (bs), respectively. Branches in bold indicate a support of PP ≥ 95% and a MLbs ≥ 70%. An asterisk on a bold branch indicates that this node has a support of 100 % for both support estimates. A dash instead of a MLbs value indicates that the node of the Bayesian tree was not recovered by ML bootstrapping. Coniocybe is highlighted by a shaded box.
3.2. Phylogeny of Coniocybe
A phylogeny of Coniocybe is presented in Figure 2. The analysis includes sequences representing four species of Coniocybe, two of which (C. brachypoda, C. furfuracea) were shown to belong in ‘Coniocybe s. str.’ in Tibell et al. [19]. In addition two further species belong to Coniocybe, viz. C. confusa that is here for the first time sequenced, and C. eufuracea here described as new. Coniocybe furfuracea is a species with a wide distribution occurring on several continents. It has a characteristic spore ornamentation of reticulate ridges as investigated by scanning electron microscopy (Figure 4D). The Tanzanian collections SGT 426 and SGT 431 of C. eufuracea are genetically very similar (Figure 2), but differ slightly from other collections of the species. However, a much wider sampling from all parts of the distribution area is required for resolving relationships within this species (or possibly species complex) and both molecular data, secondary chemistry and morphology need to be chartered in detail, which is beyond the scope of this investigation.
3.3. Taxonomy
ConiocybeAch. nom. sanct, emend.Temu & Tibell
Coniocybe Ach., K. Vetensk-Acad. Nya Handl. 4: 285 (1816).
Lectotype: C. brachypoda Ach. (Fink Cont. United States Nat. Herb. 14,1: 45 (1910)
Thallus crustaceous; ascomata with long stalks and rounded capitula with inconspicuous or missing excipulum; asci catenulate, with croziers; spores spherical, non-septate, small, pale brown, with an ornamentation of minute irregularly arranged ridges; mazaedium well developed; secondary metabolites vulpinic acid derivatives; photobiont Stichococcus.
Figure 3.
Ascomata of Coniocybe species; scales: 1 mm. A: C. eufuracea (Temu 422); B: C. brachypoda (Tibell 17062); C: C. confusa (Kantvilas 280/19); D: C. furfuracea (Temu 442). Pictures by George Hillman.
Figure 3.
Ascomata of Coniocybe species; scales: 1 mm. A: C. eufuracea (Temu 422); B: C. brachypoda (Tibell 17062); C: C. confusa (Kantvilas 280/19); D: C. furfuracea (Temu 442). Pictures by George Hillman.

Apart from C. brachypoda Ach., and C. furfuracea (L.) Ach, C. confusa (Tibell) Temu & Tibell was found to belong here based on molecular information. One new species, C. eufuracea is here being described.
Key to the species of Coniocybe
- 1.1. Apothecia 0.4 – 1.4 mm high, mazaedium medium brown..............................................................................C. brachypoda
- 1.2. Apothecia 0.6 – 3.0 mm high high, mazedium pale brown..................................................................................................2
- 2.1. Spore surface with short, irregular ridges and cracks visible under the light microscope…………….……...C. confusa
- 2.2. Spores with reticulate ridges, but without cracks ……………………………...……………………………..…….………3
- 3.1. ITS1 diagnostic sequence: CTTCT; ITS2 diagnostic sequence: TGCAGC ……………………….….….……..C. eufuracea
- 3.2. ITS1 diagnostic sequence: TCGTGC; ITS2 diagnostic sequence: TGTAGT…………………...………….......C. furfuracea
Coniocybe brachypodaAch.
Coniocybe brachypoda Ach., K. Vetensk Acad. Handl. 1816: 287 (1816).
Type (H-Ach 535, lectotype, Tibell, Symb. Bot. Ups. 27(1): 71, 1987).
Thallus immersed; apothecia short, 0.4 – 1.4 mm high, covered by a dense greenish pruina; mazaedium dark to medium brown, ± pruinose; capitulum spherical, 0.1 – 0.2 mm diam., with poorly developed excipulum; stalk 0.04 – 0.08 mm wide, covered with pruina; spores medium brown, spherical to somewhat cuboid, 3.0 – 4.5 µm diam., with a very minute ornamentation of tiny ridges and conspicuous, larger irregular cracks (Figure 4A,B); photobiont: Stichococcus.
Figure 4.
Spore ornamentations of Coniocybe species, SEM; scales: 1 µm. A: minute irregularly arranged ridges of C. brachypoda; B: irregular cracks of C. brachypoda; C: short irregularly arranged ridges of C. confusa; D: reticulate arranged ridges in C. furfuracea; E-F: ornamentation of reticulate ridges of C. eufuracea.
Figure 4.
Spore ornamentations of Coniocybe species, SEM; scales: 1 µm. A: minute irregularly arranged ridges of C. brachypoda; B: irregular cracks of C. brachypoda; C: short irregularly arranged ridges of C. confusa; D: reticulate arranged ridges in C. furfuracea; E-F: ornamentation of reticulate ridges of C. eufuracea.

Figures 3B and 4A,B.
Note: Characterised by having rather short apothecia, an usually immersed thallus and a rather dark brown mazaedium with, at least in young stages, a yellowish green pruina covering the mazaedium. Capitulum 0.1 – 0.2 mm diam. The spores are spherical to cuboid, 3 – 4.5 μm diam. and have an ornamentation of minute, irregularly arranged ridges not visible under the light microscope and larger, irregular cracks (Figure 4B) that are well within the resolution of the light microscope.
The pictures of C. brachypoda in Tibell [14] Figure 44 agree well with this insofar that in the transmission electron microscopy picture; Figure 44A, shows gaps in the outermost spore wall corresponding to cracks visible in our SEM picture (Figure 4B), while the ridge ornamentation in Figure 44B is minute and only barely discernible. These figures then most likely represent C. brachypoda. However, for the New Zealand material the thallus was described as episubstratic and green [14], which might indicate that at least some of the material used for the description in fact refers to misidentified C. confusa. Coniocybe brachypoda grows on bark and wood in shaded and humid situations. A very widely distributed species in the Northern Hemisphere and also known from New Zealand, whereas Australian [14] and South American [18] reports have not yet been supported by sequence data.
Selected specimens examined: Sweden, Jämtland, Kall par., 2 km NW of Kall, Sandnäset, between Stortjärnen and Svarttjärnen, Tibell 17062 (UPS: GB: AF297962). Åre par. 10 km ESE of Handöl, 1 km from the mouth of River Järpån, Tibell 22193 (UPS; GB: AF297963). New Zealand, North Island, Tongariro National Park, 5.5 km NE of Ohakune Railway Station, Tibell 16627; UPSC2070 (UPS; GB: XXX).
Coniocybe confusa(Tibell) Tibell & Temu, comb. nov.
Chaenotheca confusa Tibell, Bibl. Lichenologica. 71: 46 (1998).
Holotype: Chile, Region XII, Isla Navarino, c 20 km E of Puerto Williams, c. 2 km SE of Puerto Eugenia, 1989, Tibell 17940 (UPS).
Thallus superficial and well developed, farinose to minutely granular, yellowish green; apothecia long and slender, 2.3 – 3.0 mm high, covered by a dense greenish pruina; mazaedium pale brown, ± pruinose; capitulum spherical, 0.3– 0.4 mm diam, with poorly developed excipulum forming a small collar at the basem when young covered by numerous hair-like crystals; stalk 0.10 – 0.15 mm wide, pruinose; Spores spherical, 2.5 – 3.5 µm diam. with an ornamentation of minute ridges and provided with distinct cracks (Figure 4C); photobiont: Stichococcus.
Figures 1C and 4C. See also Tibell [18] Figure 10.
Habitat. On tree trunks and decorticated stumps in dark and humid situations.
Distribution. Widely distributed in the Southern Hemisphere. Vouchered by molecular data from specimen from Australia.
Note. Characterized by having a farinose to minutely granular, greenish yellow thallus; long-stalked apothecia covered by a greenish-yellow pruina; a hemispherical to almost spherical capitulum with poorly developed excipulum; catenulate asci; and spherical to cuboid spores 2.5–3.5 µm diam. having a minutely fissured surface. Very similar to C. furfuracea. Known from temperate South America and Australasia.
Specimen examined: Australia, Tasmania, Eldon Road, alt. 300 m., 2019 Kantvilas 280/19, HO 598335; GB: XXX)
Coniocybe eufuracea Temu and Tibell sp. nov.
Holotype: Tanzania, Arusha, Mt. Meru, 3°16’58.35”S 36°42’09.41”E, alt. 2096 m, on Aguru salicifolia, Temu 422 (UDSM; GB: XXX).
Thallus superficial and well developed, yellowish green; apothecia middle sized, 0.6 – 1.5 mm high (X=1.05 mm, sd=0.45 mm, n=40, c=4), covered by a dense greenish pruina; mazaedium pale brown, ± pruinose; capitulum spherical, 0.16–0.22 mm diam., (X=0.16 mm, sd=0,03 mm, n=40, c=4) with poorly developed excipulum; stalk 0.04 – 0.08 mm wide (X=0.06 mm, sd=0.02 mm,n=40, c=4), pruinose; spores pale brown spherical, 2.3 – 2.6 um diam. (X=2.42 μm, sd=0.13 μm, n=40, c=4) with a minute ornamentation of reticulate ridges (Figures 4E,F); photobiont: Stichococcus.
Figures 3A and 4E,F.
Habitat. On tree trunks and decorticated stumps in dark and humid situations.
Distribution. Widely distributed in cool temperate to temperate areas of the Northern Hemisphere. Vouchered by molecular data from specimens from Canada, India, Japan, Sweden and high altitude in Tanzania.
Note: Together with Coniocybe confusa and C, furfuracea forming a complex of (macro-) morphologically cryptic species that differ in the DNA of the ITS and LSU regions.
Additional specimens examined: Canada, Kouchibouguad National Park, S bank of Black river N of the Biodiversity monitoring site, 46:50N 65:00:33W, 005m, on decayed wood of Betula alleghaniensis, Koffman 387 (UPS; GB; TBS). India, Uttaranchal, 25.5 km NNE of Ghuttu, above Kharsoli, on theW valley slope, in mixed Picea-Quercus semecarpifolia forest, on decorticated stump of Q. semecarpifolia, 30 44’N, 78°53’E, 2003, Tibell 23224 (UPS; GB TBS), 20 km NNE of Uttarkashi, Dodital, 2008 Tibell 25024 (UPS; GB XXX); Ghangaria just S of the village on W facing slope, 2008 Tibell 25106 (UPS; GB: TBS). Japan, Honshu, Kanagawa Pref. (Sagami Prov.), Odawara-city, 80 km SW Tokyo, 4 km ESE the town Odawara, 100-400 m N of the 300 years old cherry tree Shidare-zakura 1 km NW of Iryuda railway station, deciduous forest along small path up in the mountains, on deciduous tree, 35°15’N, 139°07’E, 200 m, Thor 15698 (hb. Thor; GB: AF298124). Sweden, Uppsala, Fiby Urskog, 59°53′ N 17°20′ E, 46 m, (Temu 443, UPS; GB: XXX); Jämtland, Åre par., 10 km ESE of Handöl, 1 km from the mouth of River Järpån, Tibell 22190 (UPS; GB; AF298125). Tanzania, Kilimanjaro Region, Kilimanjaro, Moshi, Mweka Route, 03◦10′ S, 37◦21′E, 2700–2900 m, at base of old Podocarpus in podocarp mountain forest, Temu 431 (UDSM; GB: XXX); Kilimanjaro National Park, Marangu route, 3°05′ S 37°10′ E, 2718 m., Temu 426 (UDSM; TBS). Location unknown: Wedin 6366 (S, GB: NR120128).
Coniocybe furfuracea(L.) Ach.
Kongelige Svenska Vetensk. Akad. Handl.: 1816: 286.
Mucor furfuraceus L., Sp. pl. 2: 1185 (1753). Epitype proposed here: Uppland, Dannemora par., 0.5 km S of Ruddu, 2000, Tibell 22364 (UPS; GB AF445357).
Nomenclatural note: There is no material of Mucor furfuraceus in the Linnaean herbarium [31]. In this paper there also is a claim that a neotype was designated. However, no identification information was given for this alleged neotype. In the lichenological tradition C. furfuracea has since long been recognized as a widely distributed and in many areas fairly common and easily recognized species. Coniocybe furfuracea was included in Chaenotheca as Chaenotheca furfuracea (L.) Tibell [4], although the inclusion of Coniocybe in Chaenotheca was described as provisional. As shown here, Coniocybe in a three-marker phylogeny is clearly within Chaenotheca s. lat. sensu Tibell [4], but also monophyletic and distinct both in the DNA regions applied and in morphology. Here we have shown that in an emended and resurrected Coniocybe, a complex of three morphologically cryptic species occur, two of them in Europe, viz. C. furfuracea and the newly described C. eufuracea, its sister species. To resolve the nomenclatural situation of C. furfuracea an epitypification is suggested. This is not without complication since this species in Acharius’sense might just as well have been C. eufuracea, but until our suggestion has been proven wrong we find the suggested epitypification a reasonable tentative solution. In the protologue, Solander was given as the collector and we find it suitable to epitypify on a recent Swedish collection for which some molecular information is available.
Thallus superficial, farinaceous, intensely yellowish green, occasionally almost completely immersed; apothecia tall, 1.6 – 2.6 mm high, mazaedium pale brown, ± pruinose; capitulum spherical, 0.1 – 0.2 mm in diam., with a poorly developed or lacking excipulum; stalk 0.06 – 0.10 mm diam., covered by a dense yellowish green pruina; spores pale brown. spherical, 2.3 – 3.0 µm diam., with an ornamentation of reticulate ridges just discernable under the light microscope. Photobiont Stichococcus.
Chemistry. Thallus K-, C-, KC-, PD-. The thallus contains vulpinic acid, pulvinic acid and pulvinic dilactone, substances which also form the pruina of the ascomata.
Habitat. In dark and humid situations, particularly on rootlets and soil of uprooted trees and decorticated stumps in coniferous forests, more rarely on deciduous trees.
Distribution. Wide distribution in cool temperate to temperate areas of the Northern Hemisphere (Eurasia, North America). Vouchered by molecular data from specimen from India, Sweden and Switzerland.
Figures 3D and 4D.
Note: Characterized by having a farinose to minutely granular, greenish yellow thallus; long-stalked apothecia covered by a greenish-yellow pruina; a hemispherical to almost spherical capitulum with poorly developed excipulum; catenulate asci; spherical to cuboid spores 2.5-3.0 um diam. with a minutely verrucose surface (Figure 4D). Very similar to C. confusa.
Selected specimens examined: India, Uttarkhand, 16 km NNE of Uttarkashi, between Manji and Dodi Tal, 1999, Tibell 21874 (UPS; T092 GB: XXX); Sangam chatti, Tibell 21829 (T046, UPS, GB: XXX). Sweden, Jämtland. Åre par., 2.9 km WSW of Åre church, Kvarnån, 2000, Tibell 22299 (UPS, T199, GB: XXX), Tibell 22307b (UPS; T198, GB: XXX). Uppland, Dannemora par., 0.5 km S of Ruddu, 2000, Tibell 22364 (UPS, epitype; GB: AF445357); Vänge par., Fiby urskog, 2020, Temu 442 (SGT 442; GB: XXX). Switzerland, no further locality data (GB: KX098351).
4. Discussion
Here we have emended Acharius’ description of Coniocybe also to include the occurrence of catenulate asci and a very unusual type of spore micro-ornamentation consisting of short irregularly arranged ridges. Along with molecular data, this has led to the exclusion of some species originally included in the genus and we have also been able to add two species unknown to Acharius, one of them, C. eufuracea, a new species.
It is interesting that Acharius’ recognition of Coniocybe stands up quite well to scrutinizing by genetic investigations some 200 years later, insofar that both C. brachypoda and ‘C. furfuracea’ (although this species in Acharius’ sense might just as well have been C. eufuracea) were included, and the features of these species very much put their mark on the generic description. This emphasized the farinose thallus, the long, thin and flexuous stalks, the small, spherical capitulum with its knob-like central part, a rather pale mazaedium, and the occurrence of a pruina. He also commented that the species occur in dark and humid sites. In Acharius’ own words (in Swedish): ‘Ehuru vid första påseende en visss formal likhet visar sig imellan detta och nyss förut beskrifna Slägtes (i. e. Calicium) arter, så upptäckas dock snart vid en nogare uppmärksamhet den väsendtliga skillnad, som är dem imellan...’ – in short, careful attention convinced Acharius about the considerable difference between Coniocybe as compared to Calicium. Yes, we have to acknowledge Acharius’ careful attention! But apart from C. brachypoda and ‘C. furfuracea’, Acharius included in Coniocybe also ‘C. stilbea’ = Sclerophora pallida (Pers.) Y.J. Yao & Spooner and Calicium aciculare Ach. = Chaenotheca hispidula (Ach.) Zahlbr. Both these latter species, although different from Calicium in having a pale brown mazaedium, rather poorly agree with Acharius’ description of Coniocybe. The inclusion of C. gracilenta Ach. is, in contrast, more excusable since in its ascoma morphology is quite similar to that of Coniocybe, but DNA information is at odds and has shown that although it belongs in Chaenotheca s. lat., it is outside Coniocybe (Figure 1).
Author Contributions
Conceptualization, L.T.; methodology, LT. and S.T; software, S.-G.T; validation, S.T., L.T. and S.-G.T.; formal analysis, S.-G.T.; investigation, S.-G.T., L.T.; resources, S.T., L.T., S.-G.T. and D.-D.T.; data curation, S.-G.T.; writing—original draft preparation, S.-G.T. and L.T.; writing—review and editing, S.-G.T., L.T., D.-D.T. and S.T.; visualization, S.-G.T. and S.T.; supervision, L.T., S.T., D.-D.T.; project administration, L.T., D.-D.T. and S.T.; funding acquisition, D.-D.T. and S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Swedish International Development Cooperation Agency (SIDA) – SIDA-UDSM, Project No. 2221.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All new sequence data in the manuscript marked with GenBank accession numbers in bold are available in NCBI.
Acknowledgments
SIDA is endorsed for providing funding to Dr. Stella G. Temu and supporting the Swedish-Tanzanian bilateral project. We are grateful to Mr. Frank Mbago (UDSM) for his help in identifying host plants, and to local field guides and the local community for their cooperation throughout the field study in Tanzania. Dr. George Hillman is thanked for providing macro photographs.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Acharius, E. Afhandling om de cryptogamiske vexter, som komma under namn af Calicioidea. –1. K. Vetensk. -Akad. Handl. 1815, 246–271. 2. K. Vetensk. -Akad. Handl. 1816, 260–291. 3. K. Vetensk. -Akad. Handl. 1817, 220–244.
- Vainio, E. Lichenographia fennica. III. Acta Soc. Fauna Fl. Fenn. 1927, 57: 1: 1–138.
- Keissler, K. von. Pyrenulaceae bis Mycoporaceae. Coniocarpineae. In Rabenhorst’s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz. 9. 1:2: 1-846 Leipzig. 1936–1938.
- Nádvorník, J. Übersicht der mitteleuropäischen Arten der Flechtenfamilie Caliciaceae. Stud. Bot. Čech. 1942a, 5: 6–46.
- Nádvorník, J. Kurze Übersicht der Flechtenfamilie Caliciaceae. Stud. Bot. Čech. 1942a, 5: 121–128.
- Schmidt, A. Anatomisch-taxonomische Untersuchungen an europäischen Arten der Flechtenfamilie Caliciaceae. Mitt. Staatsinst. Allg. Bot. Hamburg 1970, 13: 111–166.
- Tibell, L. A reappraisal of the taxonomy of Caliciales. Nova Hedwig. 1984, Beiheft 79: 597–713.
- Tibell, L.; Wedin, M. Mycocaliciales, a new order for nonlichenized calicioid fungi. Mycologia 2000, 92: 577–581. [CrossRef]
- Prieto, M.; Baloch, E.; Tehler, A.; Wedin, M. Mazaedium evolution in the Ascomycota (Fungi) and the classification of mazaediate groups of unclear relationship. Cladistics 2013, 29: 1–13. [CrossRef]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota – Approaching one thousand genera. Bryologist 2016, 119: 361−416. [CrossRef]
- Lumbsch, H.T.; Huhndorf, S.M. Myconet Volume 14. Part One. Outline of Ascomycota—2009. Part Two. Notes on Ascomycete Systematics. Nos. 4751– 5113. Fieldiana Life and Earth Sciences 2010; 1−64.
- Tibell, L. The lichen genus Chaenotheca in the Northern Hemisphere. Symb. Bot. Upsal. 1980, 23(1): 1−65.
- Tibell, L. Typification of names of infrageneric taxa described by Acharius and placed by him in Caliciales. Ann. Bot. Fenn. 1987, 24: 257−280.
- Tibell, L. Australasian Caliciales., Symb. Bot. Upsal. 1987, 27(1): 1–279.
- Fries, E. Lichenographia Europaea reformata. Praemittuntur lichenologiae fundamenta... Conscripsit Elias Fries... Typis Berlingianis. 1831.
- Fries, T.M. Lichenes arctoi Europae Groenlandiaeque hactenus cogniti (Vol. 3). 1860, Leffler.
- Zahlbruckner, A.V. (1926). Lichenes (Flechten). B. Spezieller Teil. Die naturlichen Pflanzenfamilien. Engelmann, Leipzig 61−263.
- Tibell, L. Crustose mazaediate lichens and the Mycocaliciaceae in temperate-South America. Bibl. Lichenol. 1998, 71: 1−107. [CrossRef]
- Tibell, L.; Tibell, S.; Van Der Pluijm, A. Chaenotheca biesboschii a new calicioid lichen from willow forests in the Netherlands. Lichenologist 2019, 51: 123–135. [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2: 113–118. [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322.
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172: 4238–4246. [CrossRef]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89: 688–698. [CrossRef]
- Larsson, A. AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30: 3276–3278. [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: testing the model of DNA substitution. Bioinformatics 1998, 14: 817–818. [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27: 171−180. [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 2012, 61: 539– 542. [CrossRef]
- Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post- analysis of large phylogenies. Bioinformatics 2014, 30: 1312–1313. [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42: 182–192. [CrossRef]
- Rambaut, A.; Drummond, A.J. FigTree v1. 3.1. Institute of Evolutionary Biology; University of Edinburgh: Edinburgh, UK, 2010.
- Jørgensen, P.M.; James, P.W.; Jarvis, C.E. Linnaean lichen names and their typification. Biol. J. Linn. Soc. 1994, 115: 261−405. [CrossRef]
Figure 1.
Consensus tree based on a Bayesian and Maximum Likelihood (ML) analyses of concatenated ITS, nuLSU and rpb1 of Chaenotheca s. lat. showing the phylogenetic position of Coniocybe. The tree was rooted using Sclerophora farinacea and.
Figure 1.
Consensus tree based on a Bayesian and Maximum Likelihood (ML) analyses of concatenated ITS, nuLSU and rpb1 of Chaenotheca s. lat. showing the phylogenetic position of Coniocybe. The tree was rooted using Sclerophora farinacea and.
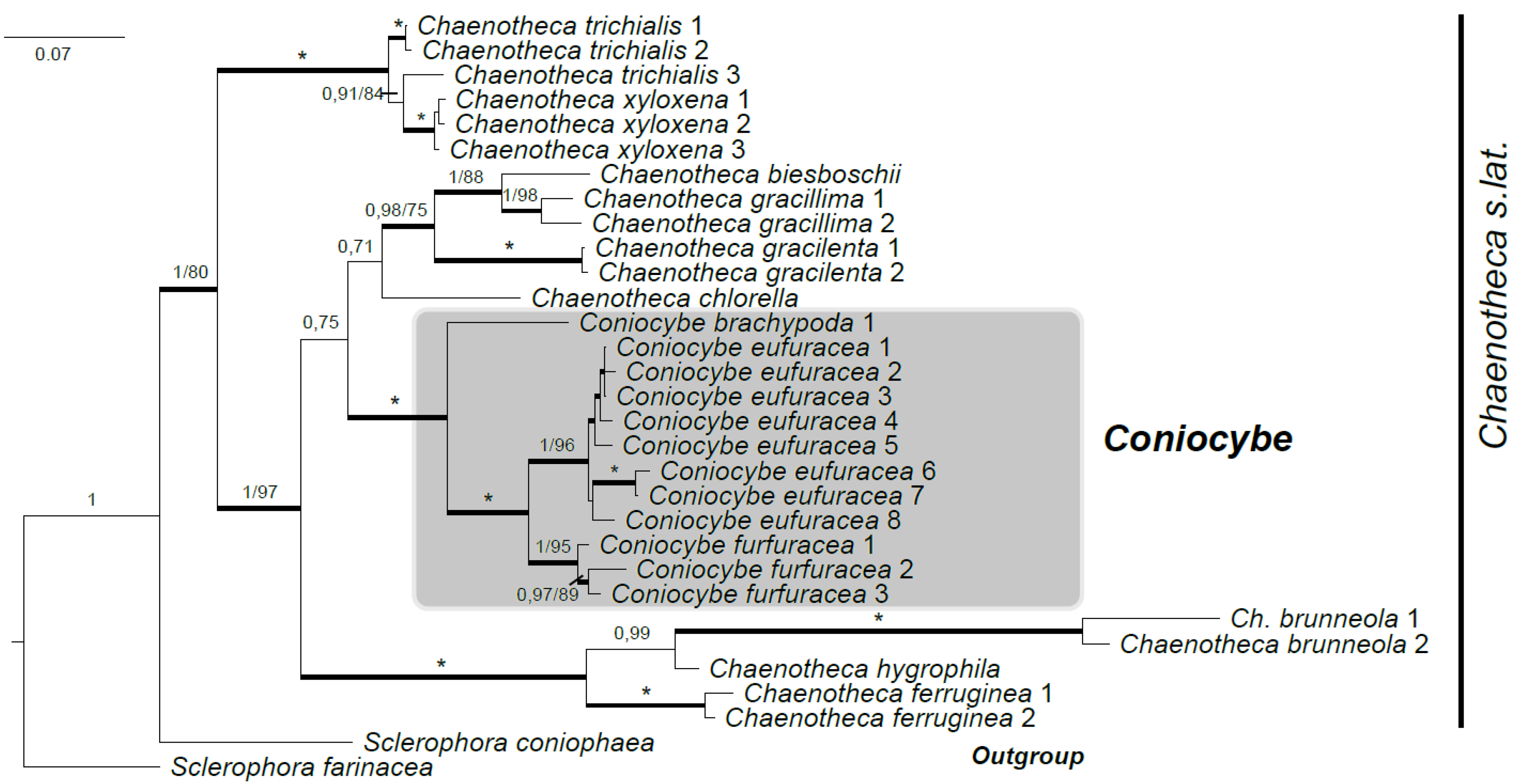
Figure 2.
Phylogenetic relationships of 4 species of Coniocybe based on a Bayesian and Maximum Likelihood (ML) analyses of an ITS dataset. The tree was rooted using Chaenotheca biesboschii and Chaenotheca gracillima. The two support values associated with each internal branch correspond to posterior probabilities (PP) and maximum likelihood bootstrap support (MLbs) proportions, respectively. Branches in bold indicate a support of PP ≥ 95% and MLbs ≥ 70%. An asterisk on a bold branch indicates that this node has a support of 100% for both support estimates.
Figure 2.
Phylogenetic relationships of 4 species of Coniocybe based on a Bayesian and Maximum Likelihood (ML) analyses of an ITS dataset. The tree was rooted using Chaenotheca biesboschii and Chaenotheca gracillima. The two support values associated with each internal branch correspond to posterior probabilities (PP) and maximum likelihood bootstrap support (MLbs) proportions, respectively. Branches in bold indicate a support of PP ≥ 95% and MLbs ≥ 70%. An asterisk on a bold branch indicates that this node has a support of 100% for both support estimates.
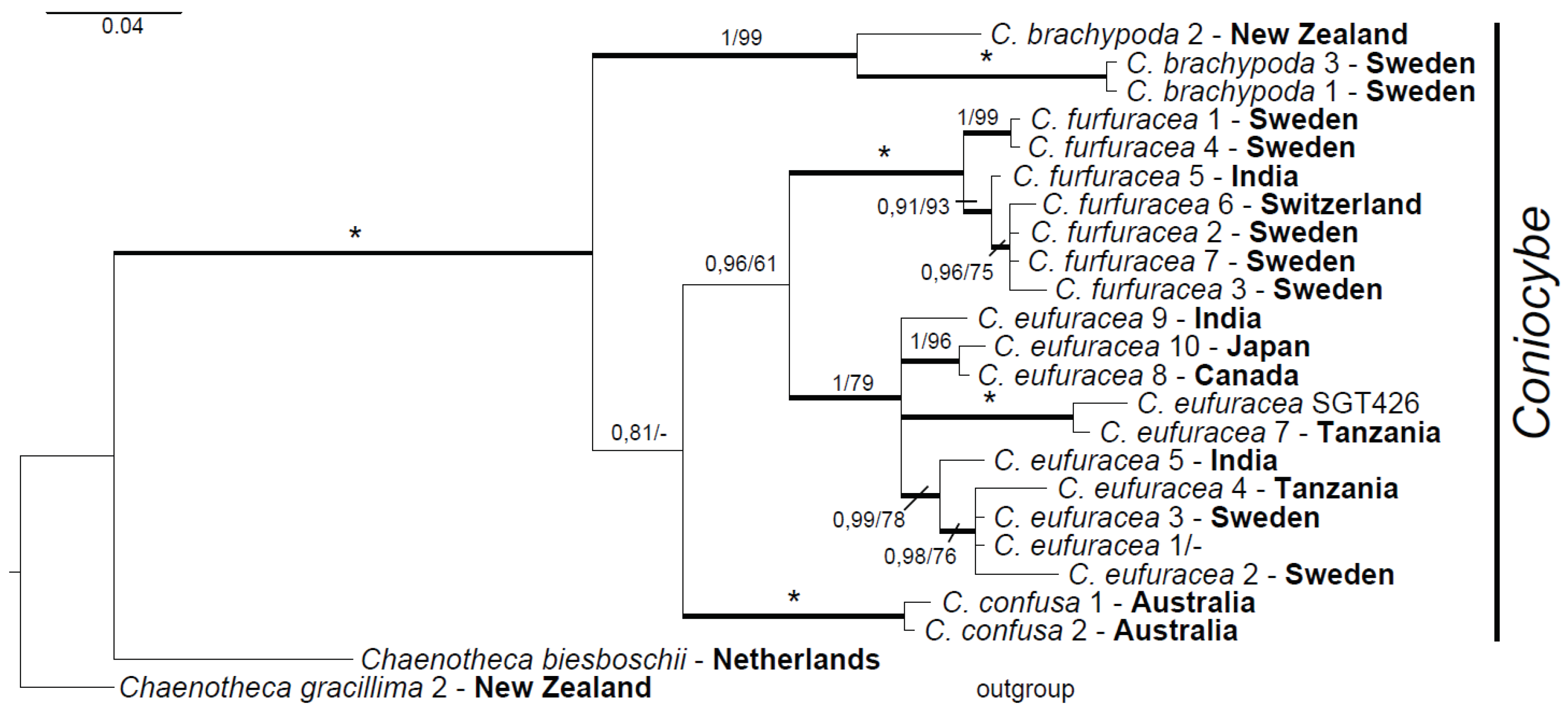

Table 1.
Species and GenBank accession numbers of sequences used in the DNA analyses. Newly produced sequences in bold.
Table 1.
Species and GenBank accession numbers of sequences used in the DNA analyses. Newly produced sequences in bold.
| Species | Isolation | Country | Voucher | GB Acc. No | ||
| ITS | LSU | rpb1 | ||||
|
Chaenotheca biesboschii |
L380 | Netherlands | A.v.d.Pluijm3244 (UPS) | MK514539 | XXX | XXX |
| Ch. brunneola 1 | T076 | Sweden | Tibell22202 (UPS) | AF297964 | XXX | ⸺ |
| Ch. brunneola 2 | T193 | Estonia | TU<EST>: 76415 | KX348127 | XXX | ⸺ |
| Ch. chlorella | T061 | Sweden | Tibell22186 | AF297965 | XXX | ⸺ |
| Ch. ferruginea 1 | T099 | Sweden | Tibell22276 (UPS) | MK514541 | XXX | ⸺ |
| Ch. ferruginea 2 | DF82/ T835 | Switzerland | WSL: DF82 | KX098349 | XXX | ⸺ |
| Ch. gracilenta 1 | T055 | Sweden | Tibell22197 (UPS) | AF410675 | XXX | XXX |
| Ch. gracilenta 2 | T135/ T310 | Sweden | Thor (hb. Thor) | AF410676 | XXX | ⸺ |
| Ch. gracillima 1 | T037 | Sweden | Tibell17052 (UPS) | AF298127 | XXX | XXX |
| Ch. gracillima 2 | T107 | New Zealand | Tibell16725 (UPS) | AF408682 | XXX | XXX |
| Ch. hygrophila | T024 | Japan | Thor 15612 (UPS) | AF298129 | XXX | ⸺ |
| Ch. trichialis 2 | UPSC: 2297/ T038 |
Sweden | Tibell16878 (UPS) | AF298139 | KF157985 | XXX |
| Ch. trichialis 1 | ⸺ | ⸺ | Prieto3028 (S) | JX000102 | JX000085 | JX000136 |
| Ch. trichialis 3 | T129 | Sweden | Tibell22300 (UPS) | AF421203 | XXX | XXX |
| Ch. xyloxena 1 | T066 | Sweden | Tibell22188 (UPS) | AF298140 | XXX | XXX |
| Ch. xyloxena 3 | T181/T131 | Sweden | Tibell22329 (UPS | AF421212 | XXX | XXX |
| Ch. xyloxena 2 | T103 | Sweden | Tibell22171 (UPS) | AF421210 | XXX | XXX |
|
Coniocybe brachypoda 3 |
T030 | Sweden | Tibell17062 (UPS) /UPSC2446 |
AF297962 | ⸺ | ⸺ |
| C. brachypoda 1 | T060/ Prieto3023 |
Sweden | Tibell22193(UPS) /Prieto3023 (S) |
AF297963 | JX000086.1 | JX000135 |
| C. brachypoda 2 | T027 | New Zealand | UPSC2070; Tibell16627 | XXX | ⸺ | ⸺ |
| C. confusa 1 | C1 | Australia | Kantvilas280/19(Ho) | XXX | ⸺ | ⸺ |
| C. confusa 2 | C2 | Australia | Kantvilas280 /19 (Ho) |
XXX | ⸺ | ⸺ |
| C. eufuracea 10 | T036 | Japan | Thor15698 | AF298124 | ⸺ | ⸺ |
| C. eufuracea 3 | T081/T062 | Sweden | Tibell22190 (UPS) | AF298125 | XXX | XXX |
| C. eufuracea 1 | ⸺ | ⸺ | Wedin6366 (UPS) | NR120128_1 | JX000087 | JX000137 |
| C. eufuracea 8 | T439 | Canada | Coffman387 | XXX | XXX | ⸺ |
| C. eufuracea 4 | SGT422 | Tanzania | Temu422 | XXX | XXX | XXX |
| C. eufuracea 2 | SGT443 | Sweden | Temu443 | XXX | XXX | XXX |
| C. eufuracea 5 | T901 | India | Tibell25106 | XXX | XXX | XXX |
| C. eufuracea 9 | T355 | India | Tibell23224 | XXX | ⸺ | ⸺ |
| C. eufuracea 6 | SGT426 | Tanzania | Temu426 | XXX | XXX | ⸺ |
| C. eufuracea 7 | SGT431 | Tanzania | Temu431 | XXX | XXX | XXX |
| C. furfuracea 1 | SGT442 | Sweden | Temu442 | XXX | XXX | ⸺ |
| C. furfuracea 2 | T046 | Sweden | Tibell21829 | XXX | XXX | ⸺ |
| C. furfuracea 3 | T198 | Sweden | Tibell22299 | XXX | XXX | ⸺ |
| C. furfuracea 4 | T155 | Sweden | Tibell22364 (UPS) | AF445357 | ⸺ | ⸺ |
| C. furfuracea 5 | T092 | India | Tibell21874 | XXX | ⸺ | ⸺ |
| C. furfuracea 6 | WSL:DF252 | Switzerland | WSL:DF252 | KX098351_1 | ⸺ | ⸺ |
| C. furfuracea 7 | T199 | Sweden | Tibell22307b | XXX | ⸺ | ⸺ |
| Sclerophora coniophaea | ⸺ | ⸺ | Wedin6367 (UPS) | ⸺ | JX000094 | JX000145 |
| S. farinacea | ⸺ | Estonia | Wedin6414 (UPS) | JX000113 | JX000095 | JX000144 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



