Submitted:
26 March 2024
Posted:
28 March 2024
You are already at the latest version
Abstract
Abstract
For the past 70 years, dopamine hypothesis has been the key working model in schizophrenia. This has contributed to the development of numerous inhibitors of dopaminergic signaling, antipsychotic drugs, which led to rapid symptom resolution but only marginal outcome improvement. Over the past decades, there was limited research on the quantifiable pathological changes in schizophrenia, including premature cellular/neuronal, senescence, brain volume loss, attenuation of gamma oscillations on electroencephalogram and oxidation of lipids in plasma and mitochondrial membranes.
We surmise that aberrant activation of aryl hydrocarbon receptor by toxins derived from the gut microbes or the environment drives premature cellular, including neuronal, senescence, a hallmark of schizophrenia. Early brain aging promotes secondary changes, including impairment and loss of mitochondria, gray matter depletion, decreased gamma oscillations, and a compensatory metabolic shift to lactate and lactylation.
The aim of this narrative review is twofold:
1. To summarize what is known about premature cellular/neuronal senescence in schizophrenia or schizophrenia-like disorders.
2. To discuss novel strategies for improving long-term outcome in severe mental illness with natural senotherapeutics, membrane lipid replacement, mitochondrial transplantation, microbial phenazines, novel antioxidant phenothiazines, inhibitors of glycogen synthase kinase-3 beta, and aryl hydrocarbon receptor.
Keywords:
aryl hydrocarbon receptor
; dopamine
; antipsychotic drugs
; naturally occurring antipsychotics
; senotherapeutics
Introduction
The discovery of chlorpromazine in the 1950s has revolutionized psychiatry and contributed to the deinstitutionalization of people with severe mental illness. Subsequently, homelessness and incarceration of individuals with schizophrenia (SCZ) and schizophrenia-like disorders (SLDs) increased dramatically, suggesting that symptomatic relief in these conditions rarely translates into sustained recovery [1,2].
Although most patients treated with antipsychotic drugs attain partial remission or amelioration of symptoms, few return to the premorbid level of functioning, measured by stable employment, attending school, raising a family, and being independent in all activities of daily living (ADLs,[3]). For this reason, large public institutions for the treatment of mental illness, such as State Hospitals, are still in existence, while sanatoria for tuberculosis or leprosy have been closed almost a century ago.
The early antipsychotic drugs were derived from methylene blue (MB) a phenothiazine synthesized in 1876 in Germany. The interest in this agent surged dramatically after the realization that it exerts antidepressant actions by inhibiting monoamine oxidase A (MAO-A), a surreptitious discovery which commenced the era of modern psychopharmacology [4]. Upon chlorpromazine approval in the US, over 40 dopamine blocking antipsychotic drugs were developed, aiming at restoring the premorbid functioning by targeting the major symptoms.
Although antipsychotic drugs are extremely efficacious for acute psychosis as the symptoms are often cleared within hours or days, sustained recovery is achieved by only 13.5% of patients after an initial psychotic episode [5]. Moreover, looking at the entire 20th century, during the early decades, long-term recovery was at 20%, not differing from the end of the century when antipsychotic drugs were widely utilized [6]. At present, 33% of patients with SCZ relapse within 12 months after an initial psychotic episode, 26% remain homeless at 2 years follow-up, and 5 years after the first psychotic outbreak, only 10% are employed [7,8,9]. Together, this data indicates that the blockade of dopamine (DA) receptors seldom improves the outcome of SCZ or SLDs. Moreover, some antipsychotic drugs, including clozapine and aripiprazole upregulate DA, suggesting that this neurotransmitter may play an indirect role in the etiopathogenesis of severe mental illness [10,11].
The aim of this narrative review is:
- To summarize what is known about the role of premature cellular/neuronal senescence in the pathogenesis of SCZ and SLDs.
- To discuss potential strategies for improving sustained recovery in SCZ and SLDs via natural senotherapeutics, microbial phenazines, aryl hydrocarbon receptor (AhR) antagonists, membrane lipid replacement (MLR), and mitochondrial transplantation.
Premature Cellular Senescence in Schizophrenia
Patients with SCZ and SLDs live on average 15-20 years shorter than the general population, exhibit shortened telomeres, and develop age-related diseases earlier in life, suggesting that premature cellular senescence plays an important role in this pathology [12,13,14,15]. Indeed, many researchers and clinicians refer to SCZ as a “segmental progeria” to highlight accelerated aging of tissues and organs, including the brain, in this disorder [12].
Cellular senescence is a program of permanent cell cycle arrest with an active metabolism, shortened telomeres, accumulation of macromolecular aggregates, increased level of senescence-associated β-galactosidase (SA-β-gal), and a toxic secretome, known as senescence-associated secretory phenotype (SASP), which can spread senescence to the neighboring healthy cells [16]. It is believed that cellular senescence defends against tumorigenesis by preventing oncogene-driven malignant transformation. However, the accumulation of aged cells and the subsequent inflammation may paradoxically promote cancer and disrupt biological barriers, facilitating the dissemination of metastases [16,17]. Inflammation and senescent cells increase the permeability of gut barrier, facilitating translocation of the gastrointestinal (GI) tact bacteria (or their molecules) into the systemic circulation, a phenomenon encountered in neuropsychiatric and neurodegenerative disorders [18]. Another example, microbiota-derived gallic acid converts p53, the key anticancer protein, into an oncogene that drives tumorigenesis [19,20]. As p53, a SCZ risk gene, also promotes cellular senescence, it likely connects microbial translocation to severe mental illness [22,23]. Indeed, bacterial molecules were demonstrated to induce cellular senescence in neurons and microglia, as documented in SCZ [24,25,26,27]. For example, Escherichia coli (E. coli)-induced psychosis was reported in epidemics as well as in urinary tract infections (UTI), connecting bacteria to SCZ and SLDs [28].
Recent studies have found that downregulated DA receptors and transporters induce premature neuronal senescence, suggesting that dopaminergic signaling is required to avert early brain aging [29]. These findings were further substantiated by virus-induced senescence (VIS), a phenotype documented during the COVID-19 pandemic, marked by brain aging due to infection of DA neurons [29,30]. Clostridium sp. are gut microbes also known for interfering with the dopaminergic signaling, further linking cellular senescence to insufficient DA [31,32]. This is significant since both treated and untreated SCZ patients were found to exhibit body-wide premature cellular/neuronal aging, linking this condition to abnormal intestinal permeability [33,34]. Indeed, SCZ was associated with increased microbial migration into the host systemic circulation [35,36,37]. Furthermore, premature cellular senescence may contribute to the other SCZ markers, including gray matter loss, decreased mitochondrial abundance, attenuation of gamma (γ) oscillations on EEG, and peroxidation of cell membrane lipids.
Ferrosenescence vs. Ferroptosis
Fe2+ is an essential nutrient for both the host and pathogens. It is also a cofactor for the biosynthesis of tyrosine hydroxylase and tryptophan hydroxylase, enzymes involved in the synthesis of DA and serotonin (5-HT) respectively. In the gut, DA acts as a microbial siderophore which clears Fe2+ from the microenvironment, lowering the risk of ferroptosis [38,39].
Senescent cells, including neurons, upregulate intracellular Fe2+ which in the vicinity of cytosolic lipids may increase the risk of peroxidation and neuronal death by ferroptosis. In addition, senescence-upregulated lactate may also increase intracellular Fe2+ further, predisposing to neuronal demise [40]. Moreover, Fe2+ and/or lactate are known for enhancing the biosynthesis of ceramide, a cell membrane lipid, which in excess can cause neurotoxicity [41,42]. Ferroptosis was documented in SCZ, however, senescent cells are often resistant to programmed cell death, suggesting that impaired autophagy may drive iron-mediated brain aging [43,44,45]. Conversely, inducing ferroptosis in senescent cells precipitates their clearance by the immune system, indicating that this mechanism may be compensatory [46].
In our previous work, we introduced the concept of ferrosenescence, senescent cells, including neurons, with damaged DNA and dysfunctional p53-mediated genomic repair as well as defective NKCs incapable of clearing senescent cells [47,48,49]. Ferrosenescent cells are resistant to ferroptosis due to the availability of glutathione peroxidase 4 (GPX-4) and apolipoprotein E (APOE) as well as impaired ferritin autophagy [50,51,52,53]. In SCZ, ferrosenescence likely accounts for the upregulated APOE, low ferritin, and increased intracellular Fe2+, characteristics of this condition [43,54]. Consequently, we believe that ferrosenescence may be more prevalent in SCZ then ferroptosis. Moreover, ferrosenescence may account for the other SCZ markers, including decreased brain volume, γ oscillations on EEG, and mitochondrial dysfunction [55,56,57,58].
Senescent Gut Barrier
The relationship between gut microbiome and senescent intestinal epithelial cells (IECs) is an emerging field which likely plays a major role in SCZ and SLDs [59,60]. For example, antibodies against translocated microbes Hafnei alvei, Pseudomonas aeruginosa, Pseudomonas putida, and Klebsiella pneumoniae, were demonstrated in SCZ with negative symptoms, connecting microbial translocation to this pathology [61]. Moreover, translocated gut microbes may trigger nutritional immunity, iron sequestration in macrophages to withhold it from microbes, decreasing total circulatory Fe2+ [62]. In conditions of low circulatory Fe2+, translocated bacteria may adopt a dormant state in human tissues, including the brain, awaiting increased Fe2+ availability to be reactivated [63,64].
Gut microbiota is immunologically tolerated in the gut lumen, however upon translocation into the systemic circulation, the immune system is activated, triggering inflammation and often antibodies against microbial molecules. Since the microorganisms populating the GI tract express receptors identical or structurally related to human proteins, antibodies against these molecules may be construed as autoantibodies. Moreover, translocated microbes elicit inflammation, promoting cellular senescence, engendering a vicious circle in which senescent IECs facilitate translocation of bacteria across the lamina propria, while inflammation triggered by these pathogens promotes further senescence [65,66,67].
Senescent cells were recently found to play a major role in the pathogenesis of inflammatory bowel disease (IBD), a condition marked by translocation of microbes from the GI tract into the host tissues, including the brain [68]. This is further enhanced by the increased prevalence of SCZ in patients with IBD, emphasizing the role of the GI tract in severe mental illness [68,69]. Microbial translocation has been studied extensively in infection with human immunodeficiency virus (HIV), a condition marked by massive exit of microbes from the GI tract due to deficient interleukin-22 (IL-22) [70]. For this reason, we believe that recombinant IL-22 may comprise a new SCZ treatment [72].
Aryl Hydrocarbon, the Master Regulator of Cellular Senescence
AhR is a ligand-activated transcription factor originally characterized as the receptor for dioxin (2,3,7,8-tetrachlorodibenzo-p-dioxin). Later it was revealed that AhR is also activated by various endogenous and exogenous ligands, driving multiple physiological processes and pathologies, likely including SCZ [73,74]. Aside from being the master regulator of cellular senescence, AhR negatively regulates lactate and by extension, the posttranslational modification, lactylation. As such, dysfunctional AhR may drive both premature neuronal and glial aging as well as the excessive lactylation documented in SCZ and SLDs [75,76].
Several AhR ligands are molecules of interest in neuropsychiatry, including DA, phenazines, phenothiazines, serotonin, melatonin, and clozapine, suggesting that this transcription factor may play a significant role in the etiopathogenesis of SCZ ([77,78,79], Figure 1). In prokaryotes, DA exerts iron-chelating properties, therefore, depletion of this neurotransmitter may upregulate intracellular Fe2+, leading to ferroptosis or ferrosenescence [39,43]. In addition, DA enhances phagocytic properties of NKCs, facilitating the elimination of senescent and damaged cells [80]. Moreover, DA, via DA 1 receptor (D1R), enhances the generation of acetylcholine (ACh), a neurotransmitter required for maintaining both the gray matter volume and rapid brain oscillations [81]. Conversely, lowering DA levels may promote inflammation by the accumulation of aged cells.
Compared to young neurons, senescent neuronal cells downregulate most surface receptors, including the dopaminergic and cholinergic ones, contain fewer mitochondria, and undergo metabolic and epigenetic reprograming via lactate and lactylation respectively. [82]. Indeed, lactylation of histone 3 (H3) lysine, opposes neuronal senescence and restores the pre-senescent cellular status, suggesting that harnessing lactylation may comprise a potential neuropsychiatric therapy [83]. However, dysfunctional lactylation was shown to induce cell cycle reentry in senescent neurons as demonstrated in Alzheimer’s disease (AD) and other neurodegenerative disorders [84]. Moreover, senescent microglia with histone 3 (H3K18) lactylation were demonstrated to adopt a neurotoxic phenotype, engaging in the elimination of healthy synapses and neurons, a phenomenon documented in SCZ and neurodegeneration [85,86,87,88,89]. Along this line, preclinical studies have revealed that neuronal excitation and social stress enhance lactylation of histones, further linking this posttranslational modification to the pathogenesis of SCZ and SLDs [90]. Furthermore, previous studies have associated SCZ to increased brain lactate and alteration of histone proteins, suggesting that lactylated neurotoxic microglia may be more detrimental for severe mental illness than previously thought [91,92,93,94].
Gray and White Matter Loss
The data on sustained remission in SCZ matches with neuroimaging studies, showing life-long gray matter loss, in both medicated and unmedicated patients. This is significant as preventing or restoring brain volume homeostasis likely improves the outcome in severe mental illness [95,96,97,98,99]. Indeed, preclinical studies have shown that AhR homeostasis is directly corelated with the brain volume, suggesting that DA/AhR binding is critical for averting parenchymal loss [100]. AhR also preserves gray matter volume by regulating the availability of ACh, which like DA, acts as a brain volume guardian [101]. Low brain DA levels also induce white matter loss but to a lesser extent in SCZ compared to demyelinating disorders [102,103]. This is significant as microbial toxins, including lipopolysaccharide (LPS) as well as environmental pollutants, such as plasticizers, are AhR ligands associated with SCZ. For example, the high comorbidity between SCZ and IBD may be the result of bacterial molecules “escaping” through the hyperpermeable intestinal barrier, a hallmark of IBD [104,105,106]. In addition, the higher prevalence of SCZ in Northern regions of the world compared to the equatorial countries may be explained by AhR binding vitamin D3 [107].
Taken together, DA is an indispensable neurotransmitter which prevents gray matter depletion by acting as an AhR ligand. Conversely, DA-blocking antipsychotic drugs may induce iatrogenic gray matter loss. This is further substantiated by the fact that clozapine, the most effective antipsychotic drug is an AhR ligand that also upregulates DA levels [77,78,108].
Dopamine-Sparing Antipsychotics
Like SCZ itself, treatment with first and second generation antipsychotic drugs was associated with gray matter loss, suggesting that brain DA levels are strictly regulated so that minimal fluctuation of this neurotransmitter may cause brain volume depletion [97,98,109,110,111,112,113,114].
Novel studies have found that antipsychotic drugs transfer positive or negative electrical charges to their substrate, donating or accepting electrons. For example, DA, lithium, clozapine, novel phenothiazines, and aripiprazole, give away electrons, preserving the gray matter volume, while most other antipsychotics are electron acceptors and associated with brain volume loss [115,116,117]. More studies are needed to identify and develop DA-sparing, electron donor, antipsychotics.
Mitochondrial Dysfunction and Loss of Gamma Band
According to the endosymbiotic hypothesis, mitochondria are derived from ancestral bacteria and can communicate with the gut microbes via chemical messengers, such as ROS and sphingolipids [118,119,120]. Ceramide, one of the sphingolipids, is secreted by the microbiome and in excess can be toxic for mitochondria [121]. Senescent cells, including neurons, upregulate ceramide, probably accounting for the paucity of mitochondria in the aging brain [122,123,124]. Moreover, senescence-upregulated intracellular Fe2+ disrupts the ceramide metabolism, contributing further to mitochondrial loss [125]. Indeed, ceramide-induced mitochondrial damage has been associated with atherosclerosis and SCZ, disorders associated with premature cellular senescence [126,127].
Acid sphingomyelinase (ASM), the enzyme catalyzing the conversion of sphingomyelin into ceramide, has been identified as a SCZ target, and ASM inhibitors, such as fluvoxamine and rosuvastatin, appear to ameliorate the clinical outcome in this disorder [128]. In addition, the natural alkaloid, berberine, decreases ceramide as well as inflammation, indicating a potential beneficial effect for SCZ and SLDs [129,130].
Neurons have the capability of replacing defective mitochondria by importing them from microglia and astrocytes via tunneling nanotubules or extracellular vesicles (EVs) ([131,132], Figure 2). Indeed, preventing neuronal loss by supplying mitochondria to neurons is one of the main function of astrocytes.
Interestingly, antidepressant drugs from the class of serotonin reuptake inhibitors (SSRIs), including fluvoxamine, facilitate mitochondrial export to neuronal cells, emphasizing the neuroprotective role of these agents ([133] Figure 3). indeed, SSRIs were demonstrated to delay the conversion of mild cognitive impairment (MCI) into dementia, indicating that mitochondrial import contributes to neuronal rescue [134]. In addition, ferroptosis-blocking drugs and iron chelators, including halogenated phenazines, may delay or prevent neurodegenerative disorders, suggesting a novel therapeutic approach (see the section on phenazines and phenothiazines) [135,136].
Aside from supplying healthy mitochondria to neurons, astrocytes also export antioxidants, including GPX-4, an enzyme involved in rescuing neuronal cells from ferroptosis. Along this line, ASM inhibitors, such as fluvoxamine, facilitate GPX-4 biosynthesis, averting ferroptosis. Indeed, fluvoxamine and other SSRIs, preserve neuronal cells by both inhibiting ferroptosis and facilitating mitochondrial transfer [137,138]. GPX-4 is derived from cystine which enters astrocytes via cystine/glutamate antiporter (Xc-) and enable the biosynthesis of glutathione and GPX-4 (Figure 2).
Given that γ band frequencies are energy-consuming, mitochondria are essential for the generation of these rapid brain oscillations [139]. In this regard, preclinical studies have shown that lost γ oscillations can be restored by inhibiting GSK-3β, an enzyme previously implicated in SCZ and SLDs [140,141]. Several antipsychotic drugs, including lithium, and natural compound, kaempferol, are GSK-3β inhibitors therefore, capable of restoring the γ rhythm (Figure 3).
Entrainment of Gamma Band Oscillations in Schizophrenia
Oscillations in the γ range are rhythmic patterns of high frequency (25Hz to 100 Hz) EEG waves, playing a key role in cognition, attention, perception, and movement [142]. Under normal circumstances, this neuronal activity is synchronized across numerous brain regions, while in SCZ there are γ rhythm abnormalities, especially those elicited by auditory stimuli [143,144].
Interneurons, including parvalbumin (PV), vasoactive intestinal polypeptide (VIP), and somatostatin (SST), are the main drivers of γ rhythm, indicating that defective inhibition of pyramidal cells may trigger the loss of rapid rhythms [145,146]. From neurotransmitters, γ oscillations are dependent on ACh, an AhR-regulated biomolecule, controlled by acetylcholinesterase (AChE), a direct AhR ligand [147].
Several studies have connected γ oscillations to the microbiome-derived ACh, linking rapid EEG rhythms to intestinal microbes [148,149]. This is significant as decreased brain ACh in SCZ has been associated with gray matter loss, emphasizing the key role of AhR and AChE in maintaining both the brain volume and rapid oscillations [150,151]. Given the important role of cholinergic system in SCZ, it is not surprising that novel ACh-based antipsychotics have been developed [130,152].
Aside from ACh and mitochondrial import, the lost γ oscillations may be restored by entrainment with sensory stimuli, such as ultrasound or transcutaneous vagal nerve stimulation (tVNS) at 40 Hz, emphasizing a potential strategy for SCZ and SLDs [149,153]. Interestingly, tVNS improves not only the neuronal function but also optimizes gut permeability, decreasing microbial translocation [149,154].
Senotherapeutics
It is currently established that severe mental illness is associated with cellular/neuronal senescence, indicating that endogenous or exogenous toxins may play a key role in this pathology [155,156,157]. For example, viral or bacterial infections induce premature aging in brain cells, by eliciting immune responses, likely triggering psychosis [158,159]. Indeed, new onset psychosis was documented in infection with senescence-inducing viruses HIV and SARS-CoV-2 [160,161].
Senotherapeutics are natural or synthetic compounds that can delay, prevent, or reverse cellular/neuronal senescence. Senotherapeutics are comprised of senolytics agents that facilitate the elimination of senescent cells, and senomorphic compounds capable of deleting senescence markers, including SASP and SA-β-gal [162]. While in the past, it has been thought that cellular senescence cannot be reversed, newer studies found that inhibiting 3-phosphoinositide–dependent protein kinase 1 (PDK1) can revert cellular senescence in humans [163]. Interestingly, PDK1 is an upstream modulator of SCZ-linked GSK-3β.
Senolytic antibiotics belong to a distinct class of agents which include azithromycin, minocycline and roxithromycin and possess neuroprotective, anti-inflammatory and senolytic properties [164]. For example, it has been known for some time that minocycline may be beneficial for SCZ, suggesting that senolytics have a place in the treatment of SCZ and SLDs, probably by clearing neurotoxic glial cells [165].
Other senolytic agents relevant for SCZ and SLDs are summarized in Table 1.
A senolytic vaccine, recently tested in progeroid mice, may usher a new era in neuropsychiatry, raising the possibility of vaccination or serum treatment for SCZ and SLDs [174]. Another immunological intervention, an antibody–drug conjugate against a membrane senescence marker was demonstrated to clear senescent, damaged, or infected cells, emphasizing a new therapeutic strategy [175].
Membrane Lipid Replacement (MLR)
MLR refers to oral administration of natural, cell membrane glycerophospholipids along with kaempferol (3,4′,5,7-tetrahydroxyflavone), a flavonoid found in tea, broccoli, cabbage, kale, beans, endive, leek, tomato, strawberries, and grapes [176]. Like lithium and some antipsychotics, kaempferol is an inhibitor of GSK-3β, suggesting that that it may exert antipsychotic properties without the typical adverse effects of psychotropic drugs [177,178].
The administration of MLR + kaempferol gradually replace the damaged phospholipids, ceramides, and oxysterols from neuronal and/or mitochondrial membranes with natural glycerophospholipids and a polyphenol.
Oxidized membrane lipids are AhR ligands which have previously been implicated in the pathogenesis of SCZ (Figure 1). MLR and kaempferol exert a dual mechanism of action: elimination of lipid peroxides and GSK-3β inhibition [179]. Replacing oxidized plasma and/or mitochondrial membrane fats with healthy natural lipids, averts ferroptosis and optimizes neurotransmission by correcting membrane distortion. Conversely, oxidized membrane lipids can trigger neuronal demise by ferroptosis [180]. Indeed, MLR reverses biophysical changes in plasma and mitochondrial membrane induced by the oxidized lipids. This action is not different from phenothiazines which insert themselves into the lipid bilayer, lowering the curvature of cell membranes (Figure 4). In contrast, oxidized lipids form looped structures, generate membrane curvatures and pores which lead to cell death [181].
Phenazines and Antioxidant Phenothiazines
Phenazines are microbial metabolites produced by the various soil and water microorganisms which exert antibacterial, anticancer, antimalarial, and antipsychotic properties [182].
Natural phenazines are synthesized by bacteria, including Streptococcus species and Pseudomonas aeruginosa, the latter known for generating pyocyanin (5-N-methyl-1-hydroxyphenazine), a compound with electron shuttling properties [183,184].
The neuroprotective natural phenazines such as, geranyl-phenazine, an AChE inhibitor, upregulates ACh, exerting antipsychotic effects via muscarinic receptors [185,186]. Another natural phenazine with neuroprotective functions, bara-phenazines A–G are fused molecules with antipsychotic properties derived from Streptomyces sp. PU-10A [187].
Synthetic phenazine derivatives consist of over 6,000 compounds, exerting antimicrobial, antiparasitic, neuroprotective, anti-inflammatory, and anticancer activities. Pontemazines A and B are neuroprotective phenazine derived from Streptomyces sp. UT1123 which in animal studies have rescued hippocampal neurons from glutamate cytotoxicity, highlighting their pro-cognitive properties which could benefit patients with negative SCZ symptoms [188]. Pontemazines exert antioxidant, radical-scavenging properties, and inhibit lipid peroxidation, suggesting beneficial effects in SCZ [189]. Halogenated phenazines act as iron chelators and are probably helpful against ferroptosis [190,191]. We believe that Pontemazines and halogenated phenazines should be assessed for antipsychotic and anti-neurodegenerative properties.
From the biochemical standpoint, phenazines are almost identical to the phenothiazine antipsychotics and likely possess similar properties (Figure 5). Phenothiazines are typical antipsychotic drugs utilized primarily for SCZ and SLDs which block dopaminergic transmission at the level of postsynaptic neuron. They also correct the curvature and receptor alignment on neuronal/mitochondrial surfaces, restoring signaling homeostasis ([192], Figure 4). In contrast, oxidized lipids, toxic ceramides, and 7-ketocholesterol (7KCl), form looped structures, generating membrane curvatures and pores, that may lead to neuronal death [193].
Antioxidant phenothiazine and their derivatives have recently been developed for cancer, cardiovascular disease (CVD), Mycobacterium leprae, and other antibiotic-resistant microbes [194].
Phenothiazine derivatives exert anti-peroxidation properties and protect against lipid pathology and ferroptosis, suggesting efficacy as antipsychotic drugs [195]. Phenothiazine nucleus possesses hydrophobic properties which facilitates the insertion into plasma or mitochondrial membranes [196].
Propenyl-phenothiazine is a potent antioxidant with electron-donor capabilities that likely prevents gray matter loss in patients with SCZ or SLDs. Moreover, a new category of tetracyclic and pentacyclic phenothiazines with antioxidant properties have recently been developed, suggesting likely efficacy for cognitive impairment and negative SCZ symptoms [197,198]. Furthermore, N10-carbonyl-substituted phenothiazines were demonstrated to inhibit lipid peroxidation, suggesting enhanced antipsychotic efficacy [199].
Mitochondrial Transfer and Transplantation
Mitochondrial transplantation experiments started in the 1980s when naked organelles were co-incubated with various cell types, attempting to facilitate internalization [200]. Using HeLa cells and mesenchymal stem cells as mitochondrial sources, this transplantation technique takes only 1-2 hours to supply organelles to mitochondria-depleted cells [201,202,203]. At present, mitochondrial transplantation in cardiomyocytes is possible and can be confirmed by the presence of mitochondrial DNA (mtDNA) in the heart [204,205].
Mitochondrial transplantation to rescue neurons from ferroptosis is currently possible and was performed successfully in both animals and humans, however, to the best of our knowledge, it has not been attempted as treatment for mental illness [206].
Rescuing the mitochondrion with MLR, Kaempferol, and Berberine (Figure 6) is a strategy for averting GSK-3β overactivation by toxic ceramides, oxysterols, or oxidized phospholipids [130,207]. In addition, SSRIs, GJA1-20K, and CD38 signaling were demonstrated to facilitate mitochondrial transfer, emphasizing potential strategies for restoring the neurometabolic homeostasis in severe mental illness, and neurodegeneration [208].
AhR Antagonists as Antipsychotics
Aberrant AhR overactivation has been associated with psychosis, while several antagonists of this receptor exert antipsychotic properties. The following natural and synthetic AhR inhibitors were found therapeutic in SCZ:
- Quercetin is a natural flavonoid and plant pigment which exerts antioxidant and anticancer properties. In the CNS, quercetin is a negative allosteric modulator of GABARs as well as an enhancer of glutamatergic neurotransmission, a signaling pathway deficient in SCZ [210]. In addition, quercetin inhibits apoptosis of cortical neurons, likely preventing gray matter loss.
- Apigenin is a plant-based remedy extract from Elsholtzia rugulosa used by traditional practitioners from Africa for treating mental illness. Aside from antagonizing AhR, apigenin exhibits vasorelaxant, antioxidant, and antipsychotic properties [210].
- Alstonine is an indole alkaloid with antipsychotic effects which increases serotonergic, but not dopaminergic, signaling, possibly facilitating mitochondrial transfer [210].
- Luteolin is a natural antipsychotic that exerts its beneficial actions by reducing microglial inflammation [203]. Luteolin is currently in clinical trials for SCZ (NCT05204407)
Synthetic AhR Antagonists
Synthetic AhR antagonists are anti-inflammatory and anticancer compounds which are likely to exert antipsychotic properties.
IK-175, structure undisclosed, was shown to block ligand-mediated AhR activation in preclinical studies. IK-175 was recently approved for cancer and it may possess antipsychotic properties [212].
HBU651 is a novel synthetic AhR antagonist developed primarily for cancer which appears to be a suitable candidate for SCZ [213].
Interleukin-22
Interleukin 22 (IL-22) is a pleiotropic cytokine known for facilitating tissue regeneration and for protecting the GI tract barrier. Recombinant IL-22, comprised of two molecules connected by a fusion protein, exerts better efficacy with reduced systemic side effects [214].
In our previous work, we have hypothesized that SCZ and SLDs may be initiated by aberrant AhR hyperactivation by endogenous or exogenous ligands, including intestinal or environmental toxins, such as LPS or plasticizers respectively [215].
This hypothesis is supported by the following:
- SCZ is often comorbid with IBD, conditions associated with increased gut barrier permeability and microbial translocation from the GI tract into host tissues, including the brain.
- Translocation markers, including soluble CD14 (sCD14) and lipopolysaccharide binding protein (LBP), are elevated in SCZ, suggesting bacterial translocation.
- Increased BBB permeability in SCZ enables translocated gut microbes to reach the brain.
Examples of pathogens triggering psychosis:
- The Escherichia coli (E. coli) outbrake in 2011 in Germany has been associated with cases of new onset psychosis.
- Exacerbation or new onset psychosis in E. coli-associated UTIs.
IL-22 has been successfully used for restoring the integrity of gut barrier in various condtions, including IBD, HIV, and nonalcoholic fatty liver disease [216,217]. We construe that Recombinant IL-22 would be effective for SCZ by limiting the translocation of bacteria and/or their molecules.
Table 2 summarizes the major representatives from the categories discussed above with potential therapeutic properties for SCZ and SLDs
Vehicles: Lipid Nanoparticles
The COVID-19 pandemic has accelerated the development of lipid nanoparticles (LNPs), vehicles for drug delivery. As LNPs are liposoluble, they can access specific body niches, including the brain [218].
The COVID-19 messenger RNA (mRNA) vaccines, Pfizer-BioNTech and Moderna, are incorporated in LNPs comprised of four lipids: 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), PEG, an alternative cholesterol, and ionizable lipids SM-102 or ALC-0315 [219]. The SM-102, ALC-0315 and the alternative cholesterol are proprietary molecules and have not been revealed. However, looking at the previous LNP research, ionizable lipids likely resemble DLin-MC3-DMA which were approved by the Food and Drug Administration (FDA) for transthyretin-mediated amyloidosis [220,221]. Like phenothiazines, LNPs enter the cells, including neurons, through the endocytic pathway (EP). Subsequently, LNPs travel from the early to late endosomes, but not to lysosomes because the organelle pH of 4.5–5.0 could degrade the nanoparticles. Therefore, “lysosomal escape” into the cytosol must occur from the late endosomes without interfering with autophagy as most antipsychotic drugs do.
Utilizing LNPs not for vaccination, but as vehicle for transporting psychotropic drugs directly to the neuronal networks could revolutionize psychopharmacology. As nano-doses of antipsychotics or mood stabilizers would be utilized for the treatment of psychotic symptoms or affective disorders, systemic adverse effects would be avoided. We surmise that LNPs would be extremely efficacious as vehicles for psychotropic drugs.
Conclusions
At present, neuropsychiatric treatments have reached an impasse: although antipsychotic medications are extremely efficacious for treating the acute symptoms of SCZ or SLDs, sustained recovery (measured by the ability to hold a job, go to school, raise a family, or be independent in all ADLs) is rare. For this reason, the next era in neuropsychiatry will have to address functionality, rather than symptom resolution.
Over the past 70 years, it has become obvious that lowering dopaminergic transmission does not restore the premorbid function in patients with SCZ and SLDs. DA is an indispensable neurotransmitter which maintains the integrity of brain parenchyma, a physiological function disrupted by both SCZ and several antipsychotic drugs.
The study of neuronal senescence-induced neuropsychiatric disorders is still in its infancy. However, abnormal AhR, the master regulator of cellular senescence, can explain not only how gut microbes and/or their molecules can trigger psychosis but also how environmental pollutants precipitate SCZ or SLDs. Aberrant AhR activation may account for gray matter loss, impaired rapid brain oscillations, as well as oxidized lipid bilayer in plasma and mitochondrial membranes. The recent discovery that DA, serotonin, vitamin D3, clozapine, and melatonin are AhR ligands opens new horizons in the treatment of chronic psychosis. Novel strategies, such as natural senotherapeutics, MLR, GSK-3β and ceramide inhibitors, IL-22, mitochondrial transplantation, or transfer, and AhR antagonists, could improve sustained recovery in SCZ or SLDs. Senescence-associated downregulation of ACh and DA receptors drives the brain volume reduction and loss of rapid oscillations.
Utilization of natural compounds, such as kaempferol or berberine, alone or in conjunction with LNP-delivered DA-sparing antipsychotic drugs may improve the sustained recovery in patients with severe mental illness.
References
- Lamb HR. Deinstitutionalization and the homeless mentally ill. Hosp Community Psychiatry. 1984 Sep;35(9):899-907. [CrossRef]
- Scott J. Homelessness and mental illness. Br J Psychiatry. 1993 Mar;162:314-24. PMID: 8453425. [CrossRef]
- Insel, T. Rethinking schizophrenia. Nature 468, 187–193 (2010). [CrossRef]
- Delport A, Harvey BH, Petzer A, Petzer JP. The monoamine oxidase inhibition properties of selected structural analogues of methylene blue. Toxicol Appl Pharmacol. 2017 Jun 15;325:1-8. [CrossRef]
- Jääskeläinen, E.; Juola, P.; Hirvonen, N.; McGrath, J.J.; Saha, S.; Isohanni, M.; Veijola, J.; Miettunen, J. A Systematic Review and Meta-Analysis of Recovery in Schizophrenia. Schizophr. Bull. 2012, 39, 1296–1306.
- Warner, R. Recovery from Schizophrenia Psychiatry and Political Economy, 3rd ed.; Brunner-Routledge: Hove, UK; New York, NY, USA, 1997; p. 74.
- Üçok, A.; Polat, A.; Çakır, S.; Genç, A. One year outcome in first episode schizophrenia: Predictors of relapse. Eur. Arch. Psychiatry Clin. Neurosci. 2005, 256, 37–43.
- Holm, M.; Taipale, H.; Tanskanen, A.; Tiihonen, J.; Mitterdorfer-Rutz, E. Employment among people with schizophrenia or bipolar disorder: A population-based study using nationwide registers. Acta Psychiatr. Scand. 2020, 143, 61–71.
- Lévesque, I.S.; Abdel-Baki, A. Homeless youth with first-episode psychosis: A 2-year outcome study. Schizophr. Res. 2019, 216, 460–469.
- Lai CH, Wu YT, Chen CY, Hou YC. Gray matter increases in fronto-parietal regions of depression patients with aripiprazole monotherapy: An exploratory study. Medicine (Baltimore). 2016 Aug;95(34):e4654. [CrossRef]
- Rollema H, Lu Y, Schmidt AW, Zorn SH. Clozapine increases dopamine release in prefrontal cortex by 5-HT1A receptor activation. Eur J Pharmacol. 1997 Nov 5;338(2):R3-5. [CrossRef]
- Papanastasiou E, Gaughran F, Smith S. Schizophrenia as segmental progeria. J R Soc Med. 2011 Nov;104(11):475-84. [CrossRef]
- Russo P, Prinzi G, Proietti S, Lamonaca P, Frustaci A, Boccia S, et al. Shorter telomere length in schizophrenia: evidence from a real-world population and meta-analysis of Most recent literature. Schizophr Res. (2018) 202:37–45. Epub 2018/07/14, PMID. [CrossRef]
- Dada O, Adanty C, Dai N, Jeremian R, Alli S, Gerretsen P, Graff A, Strauss J, De Luca V. Biological aging in schizophrenia and psychosis severity: DNA methylation analysis. Psychiatry Res. 2021 Feb;296:113646. [CrossRef]
- Schnack HG, van Haren NE, Nieuwenhuis M, Hulshoff Pol HE, Cahn W, Kahn RS. Accelerated Brain Aging in Schizophrenia: A Longitudinal Pattern Recognition Study. Am J Psychiatry. 2016 Jun 1;173(6):607-16. [CrossRef]
- Langhi Prata LGP, Tchkonia T, Kirkland JL. Cell senescence, the senescence-associated secretory phenotype, and cancers. PLoS Biol. 2023 Sep 21;21(9):e3002326. [CrossRef]
- Huang, W., Hickson, L.J., Eirin, A. et al. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol 18, 611–627 (2022). [CrossRef]
- Stec A, Maciejewska M, Zaremba M, Paralusz-Stec K, Michalska M, Rudnicka L, Sikora M. The Clinical Significance of Serum Biomarkers of the Intestinal Barrier in Systemic Sclerosis: A Cross-Sectional Study. J Pers Med. 2023 Apr 18;13(4):678. [CrossRef]
- Sepich-Poore GD, Zitvogel L, Straussman R, Hasty J, Wargo JA, Knight R. The microbiome and human cancer. Science. 2021 Mar 26;371(6536):eabc4552. [CrossRef]
- White MG, Wargo JA. Gut Microbes' Impact on Oncogenic Drivers: Location Matters. Mol Cell. 2020 Sep 17;79(6):878-880. PMID: 32946762; PMCID: PMC7717102. [CrossRef]
- Ağagündüz D, Cocozza E, Cemali Ö, Bayazıt AD, Nanì MF, Cerqua I, Morgillo F, Saygılı SK, Berni Canani R, Amero P, Capasso R. Understanding the role of the gut microbiome in gastrointestinal cancer: A review. Front Pharmacol. 2023 Jan 24;14:1130562. [CrossRef]
- Mijit M, Caracciolo V, Melillo A, Amicarelli F, Giordano A. Role of p53 in the Regulation of Cellular Senescence. Biomolecules. 2020 Mar 8;10(3):420. PMID: 32182711; PMCID: PMC7175209. [CrossRef]
- Ni X, Trakalo J, Valente J, Azevedo MH, Pato MT, Pato CN, Kennedy JL. Human p53 tumor suppressor gene (TP53) and schizophrenia: case-control and family studies. Neurosci Lett. 2005 Nov 18;388(3):173-8. [CrossRef]
- Dono A, Nickles J, Rodriguez-Armendariz AG, McFarland BC, Ajami NJ, Ballester LY, Wargo JA, Esquenazi Y. Glioma and the gut-brain axis: opportunities and future perspectives. Neurooncol Adv. 2022 Apr 14;4(1):vdac054. [CrossRef]
- Palacios E, Lobos-González L, Guerrero S, Kogan MJ, Shao B, Heinecke JW, Quest AFG, Leyton L, Valenzuela-Valderrama M. Helicobacter pylori outer membrane vesicles induce astrocyte reactivity through nuclear factor-κappa B activation and cause neuronal damage in vivo in a murine model. J Neuroinflammation. 2023 Mar 9;20(1):66. [CrossRef]
- Zhuo, C., Tian, H., Song, X. et al. Microglia and cognitive impairment in schizophrenia: translating scientific progress into novel therapeutic interventions. Schizophr 9, 42 (2023). [CrossRef]
- Niraula A., Sheridan J.F., Godbout J.P. Microglia Priming with Aging and Stress. Neuropsychopharmacol. Rev. 2017;42:318–333. [CrossRef]
- Wiwanitkit V. Hemolysis in E. coli O104:H4 Infection. Indian J Hematol Blood Transfus. 2012 Jun;28(2):127. Epub 2011 Sep 4. PMID: 23730024; PMCID: PMC3332272. [CrossRef]
- Karrer TM, Josef AK, Mata R, Morris ED, Samanez-Larkin GR. Reduced dopamine receptors and transporters but not synthesis capacity in normal aging adults: a meta-analysis. Neurobiol Aging. 2017 Sep;57:36-46. [CrossRef]
- Yang L, Kim TW, Han Y, Nair MS, Harschnitz O, Zhu J, Wang P, Koo SY, Lacko LA, Chandar V, et al. SARS-CoV-2 infection causes dopaminergic neuron senescence. Cell Stem Cell. 2024 Feb 1;31(2):196-211.e6. [CrossRef]
- Fettucciari K, Fruganti A, Stracci F, Spaterna A, Marconi P, Bassotti G. Clostridioides difficile Toxin B Induced Senescence: A New Pathologic Player for Colorectal Cancer? Int J Mol Sci. 2023 May 2;24(9):8155. [CrossRef]
- Vinithakumari AA, Padhi P, Hernandez B, Lin SJ, Dunkerson-Kurzhumov A, Showman L, Breitzman M, Stokes C, Sulaiman Y, Tangudu C, et al. Clostridioides difficile Infection Dysregulates Brain Dopamine Metabolism. Microbiol Spectr. 2022 Apr 27;10(2):e0007322. [CrossRef]
- Feng Y, Shen J, He J, Lu M. Schizophrenia and cell senescence candidate genes screening, machine learning, diagnostic models, and drug prediction. Front Psychiatry. 2023 Apr 11;14:1105987. [CrossRef]
- Ling, E., Nemesh, J., Goldman, M. et al. A concerted neuron–astrocyte program declines in ageing and schizophrenia. Nature (2024). [CrossRef]
- Ishida I, Ogura J, Aizawa E, Ota M, Hidese S, Yomogida Y, Matsuo J, Yoshida S, Kunugi H. Gut permeability and its clinical relevance in schizophrenia. Neuropsychopharmacol Rep. 2022 Mar;42(1):70-76. [CrossRef]
- Scheurink TAW, Borkent J, Gangadin SS, El Aidy S, Mandl R, Sommer IEC. Association between gut permeability, brain volume, and cognition in healthy participants and patients with schizophrenia spectrum disorder. Brain Behav. 2023 Jun;13(6):e3011. [CrossRef]
- Wasiak J, Gawlik-Kotelnicka O. Intestinal permeability and its significance in psychiatric disorders - A narrative review and future perspectives. Behav Brain Res. 2023 Jun 25;448:114459. [CrossRef]
- Lotan, A., Luza, S., Opazo, C.M. et al. Perturbed iron biology in the prefrontal cortex of people with schizophrenia. Mol Psychiatry 28, 2058–2070 (2023). [CrossRef]
- Dichtl S, Demetz E, Haschka D, Tymoszuk P, Petzer V, Nairz M, Seifert M, Hoffmann A, Brigo N, Würzner R, Theurl I, Karlinsey JE, Fang FC, Weiss G. Dopamine Is a Siderophore-Like Iron Chelator That Promotes Salmonella enterica Serovar Typhimurium Virulence in Mice. mBio. 2019 Feb 5;10(1):e02624-18. [CrossRef]
- Gregory J Anderson, David M Frazer, Lactate as a regulator of iron homeostasis, Life Metabolism, Volume 2, Issue 5, October 2023, load033. [CrossRef]
- Cai, Z.; Deng, L.; Fan, Y.; Ren, Y.; Ling, Y.; Tu, J.; Cai, Y.; Xu, X.; Chen, M. Dysregulation of Ceramide Metabolism Is Linked to Iron Deposition and Activation of Related Pathways in the Aorta of Atherosclerotic Miniature Pigs. Antioxidants 2024, 13, 4. [CrossRef]
- de la Monte SM. Triangulated mal-signaling in Alzheimer's disease: roles of neurotoxic ceramides, ER stress, and insulin resistance reviewed. J Alzheimers Dis. 2012;30 Suppl 2(0 2):S231-49. [CrossRef]
- Ravanfar P, Syeda WT, Jayaram M, Rushmore RJ, Moffat B, Lin AP, Lyall AE, Merritt AH, Yaghmaie N, Laskaris L, Luza S, Opazo CM, Liberg B, Chakravarty MM, Devenyi GA, Desmond P, Cropley VL, Makris N, Shenton ME, Bush AI, Velakoulis D, Pantelis C. In Vivo 7-Tesla MRI Investigation of Brain Iron and Its Metabolic Correlates in Chronic Schizophrenia. Schizophrenia (Heidelb). 2022 Oct 26;8(1):86. [CrossRef]
- Feng S, Chen J, Qu C, Yang L, Wu X, Wang S, Yang T, Liu H, Fang Y, Sun P. Identification of Ferroptosis-Related Genes in Schizophrenia Based on Bioinformatic Analysis. Genes (Basel). 2022 Nov 20;13(11):2168. [CrossRef]
- Lian K, Li Y, Yang W, Ye J, Liu H, Wang T, Yang G, Cheng Y, Xu X. Hub genes, a diagnostic model, and immune infiltration based on ferroptosis-linked genes in schizophrenia. IBRO Neurosci Rep. 2024 Feb 3;16:317-328. [CrossRef]
- Go S, Kang M, Kwon SP, Jung M, Jeon OH, Kim BS. The Senolytic Drug JQ1 Removes Senescent Cells via Ferroptosis. Tissue Eng Regen Med. 2021 Oct;18(5):841-850. [CrossRef]
- Masaldan S, Clatworthy SAS, Gamell C, Meggyesy PM, Rigopoulos AT, Haupt S, Haupt Y, Denoyer D, Adlard PA, Bush AI, Cater MA. Iron accumulation in senescent cells is coupled with impaired ferritinophagy and inhibition of ferroptosis. Redox Biol. 2018 Apr;14:100-115. [CrossRef]
- Sfera A, Osorio C, Maguire G, Rahman L, Afzaal J, Cummings M, Maldonado JC. COVID-19, ferrosenescence and neurodegeneration, a mini-review. Prog Neuropsychopharmacol Biol Psychiatry. 2021 Jul 13;109:110230. [CrossRef]
- Sfera A, Bullock K, Price A, Inderias L, Osorio C. Ferrosenescence: The iron age of neurodegeneration? Mech Ageing Dev. 2018 Sep;174:63-75. [CrossRef]
- Dean DC 3rd, Sojkova J, Hurley S, Kecskemeti S, Okonkwo O, Bendlin BB, Theisen F, Johnson SC, Alexander AL, Gallagher CL. Alterations of Myelin Content in Parkinson's Disease: A Cross-Sectional Neuroimaging Study. PLoS One. 2016 Oct 5;11(10):e0163774. [CrossRef]
- Belaidi, A.A., Masaldan, S., Southon, A. et al. Apolipoprotein E potently inhibits ferroptosis by blocking ferritinophagy. Mol Psychiatry (2022). [CrossRef]
- Arnold SE, Joo E, Martinoli MG, et al. Apolipoprotein E genotype in schizophrenia: frequency, age of onset, and neuropathologic features. Neuroreport. 1997;8:1523–6.
- Kampman O, Anttila S, Illi A, et al. Apolipoprotein E polymorphism is associated with age of onset in schizophrenia. J Hum Genet. 2004;49:355–9.
- Xu M, Guo Y, Cheng J, Xue K, Yang M, Song X, Feng Y, Cheng J. Brain iron assessment in patients with First-episode schizophrenia using quantitative susceptibility mapping. Neuroimage Clin. 2021;31:102736. [CrossRef]
- Sabbatinelli J, Prattichizzo F, Olivieri F, Procopio AD, Rippo MR, Giuliani A. Where Metabolism Meets Senescence: Focus on Endothelial Cells. Front Physiol. 2019 Dec 18;10:1523. [CrossRef]
- Pruett B.S., Meador-Woodruff J.H. Evidence for altered energy metabolism, increased lactate, and decreased pH in schizophrenia brain: A focused review and meta-analysis of human postmortem and magnetic resonance spectroscopy studies. Schizophr. Res. 2020;223:29–42. [CrossRef]
- Chou SM, Yen YH, Yuan F, Zhang SC, Chong CM. Neuronal Senescence in the Aged Brain. Aging Dis. 2023 Oct 1;14(5):1618-1632. PMID: 37196117; PMCID: PMC10529744 . [CrossRef]
- Murty DVPS, Manikandan K, Kumar WS, Ramesh RG, Purokayastha S, Javali M, Rao NP, Ray S. Gamma oscillations weaken with age in healthy elderly in human EEG. Neuroimage. 2020 Jul 15;215:116826. [CrossRef]
- Sharma R. Emerging Interrelationship Between the Gut Microbiome and Cellular Senescence in the Context of Aging and Disease: Perspectives and Therapeutic Opportunities. Probiotics Antimicrob Proteins. 2022 Aug;14(4):648-663. Epub 2022 Jan 5. PMID: 34985682. [CrossRef]
- Frey, N., Venturelli, S., Zender, L. et al. Cellular senescence in gastrointestinal diseases: from pathogenesis to therapeutics. Nat Rev Gastroenterol Hepatol 15, 81–95 (2018). [CrossRef]
- Maes M, Kanchanatawan B, Sirivichayakul S, Carvalho AF. In Schizophrenia, Increased Plasma IgM/IgA Responses to Gut Commensal Bacteria Are Associated with Negative Symptoms, Neurocognitive Impairments, and the Deficit Phenotype. Neurotox Res. 2019 Apr;35(3):684-698. [CrossRef]
- Hennigar SR, McClung JP. Nutritional Immunity: Starving Pathogens of Trace Minerals. Am J Lifestyle Med. 2016 Feb 4;10(3):170-173. [CrossRef]
- Pretorius L, Kell DB, Pretorius E. Iron Dysregulation and Dormant Microbes as Causative Agents for Impaired Blood Rheology and Pathological Clotting in Alzheimer's Type Dementia. Front Neurosci. 2018 Nov 16;12:851. [CrossRef]
- Link CD. Is There a Brain Microbiome? Neurosci Insights. 2021 May 27;16:26331055211018709. PMID: 34104888; PMCID: PMC8165828. [CrossRef]
- Dworkin J, Shah IM. Exit from dormancy in microbial organisms. Nat Rev Microbiol. 2010 Dec;8(12):890-6. [CrossRef]
- Peyrusson F, Nguyen TK, Najdovski T, Van Bambeke F. Host Cell Oxidative Stress Induces Dormant Staphylococcus aureus Persisters. Microbiol Spectr. 2022 Feb 23;10(1):e0231321. [CrossRef]
- Berthelot JM, de la Cochetière MF, Potel G, Le Goff B, Maugars Y. Evidence supporting a role for dormant bacteria in the pathogenesis of spondylarthritis. Joint Bone Spine. 2013 Mar;80(2):135-40. [CrossRef]
- Sienkiewicz M, Sroka K, Binienda A, Jurk D, Fichna J. A new face of old cells: An overview about the role of senescence and telomeres in inflammatory bowel diseases. Ageing Res Rev. 2023 Nov;91:102083. [CrossRef]
- Qian, L., He, X., Gao, F. et al. Estimation of the bidirectional relationship between schizophrenia and inflammatory bowel disease using the mendelian randomization approach. Schizophr 8, 31 (2022). [CrossRef]
- Bartocci, B.; Dal Buono, A.; Gabbiadini, R.; Busacca, A.; Quadarella, A.; Repici, A.; Mencaglia, E.; Gasparini, L.; Armuzzi, A. Mental Illnesses in Inflammatory Bowel Diseases: mens sana in corpore sano. Medicina 2023, 59, 682. [CrossRef]
- Zevin AS, McKinnon L, Burgener A, Klatt NR. Microbial translocation and microbiome dysbiosis in HIV-associated immune activation. Curr Opin HIV AIDS. 2016 Mar;11(2):182-90. [CrossRef]
- Adonis Sfera, Nyla Jafri and Leah Rahman. F-652 (Recombinant Human Interleukin-22) For Schizophrenia. Arch Phar &Pharmacol Res. 3(3): 2023. APPR.MS.ID.000564. [CrossRef]
- Schubert KO, Föcking M, Cotter DR. Proteomic pathway analysis of the hippocampus in schizophrenia and bipolar affective disorder implicates 14-3-3 signaling, aryl hydrocarbon receptor signaling, and glucose metabolism: potential roles in GABAergic interneuron pathology. Schizophr Res. 2015 Sep;167(1-3):64-72. [CrossRef]
- Ludmila Juricek, Xavier Coumoul. The Aryl Hydrocarbon Receptor and the Nervous System. International Journal of Molecular Sciences, 2018, 19 (9), pp.2504. ff10.3390/ijms19092504ff. ffhal-01955.
- Lopez-Otin C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153:1194–1217. [CrossRef]
- Murray, I., Nichols, R., Zhang, L. et al. Expression of the aryl hydrocarbon receptor contributes to the establishment of intestinal microbial community structure in mice. Sci Rep 6, 33969 (2016). [CrossRef]
- Park H, Jin UH, Karki K, Jayaraman A, Allred C, Michelhaugh SK, Mittal S, Chapkin RS, Safe S. Dopamine is an aryl hydrocarbon receptor agonist. Biochem J. 2020 Oct 16;477(19):3899-3910. [CrossRef]
- Fehsel K, Schwanke K, Kappel BA, Fahimi E, Meisenzahl-Lechner E, Esser C, Hemmrich K, Haarmann-Stemmann T, Kojda G, Lange-Asschenfeldt C. Activation of the aryl hydrocarbon receptor by clozapine induces preadipocyte differentiation and contributes to endothelial dysfunction. J Psychopharmacol. 2022 Feb;36(2):191-201. [CrossRef]
- Moura-Alves P, Faé K, Houthuys E, Dorhoi A, Kreuchwig A, Furkert J, Barison N, Diehl A, Munder A, Constant P, Skrahina T, Guhlich-Bornhof U, Klemm M, Koehler AB, Bandermann S, Goosmann C, Mollenkopf HJ, Hurwitz R, Brinkmann V, Fillatreau S, Daffe M, Tümmler B, Kolbe M, Oschkinat H, Krause G, Kaufmann SH. AhR sensing of bacterial pigments regulates antibacterial defence. Nature. 2014 Aug 28;512(7515):387-92. [CrossRef]
- Bai, Z., Yang, P., Yu, F. et al. Combining adoptive NK cell infusion with a dopamine-releasing peptide reduces senescent cells in aged mice. Cell Death Dis 13, 305 (2022). [CrossRef]
- Yurchenko OP, Grigoriev NG, Turpaev TM, Konjević D, Rakić L. Intracellular injection of dopamine enhances acetylcholine responses of neuron R2 in the Aplysia abdominal ganglion. Comparative Biochemistry and physiology. C, Comparative Pharmacology and Toxicology. 1987 ;87(2):389-391. PMID: 2888583. [CrossRef]
- Nakao M, Tanaka H, Koga T. Cellular Senescence Variation by Metabolic and Epigenomic Remodeling. Trends Cell Biol. 2020 Dec;30(12):919-922. [CrossRef]
- Xuebin Zhang, Fanju Meng, Wencong Lyu, Jianuo He, Ran Wei, Zhehao Du, Chao Zhang. Histone lactylation antagonizes senescence and skeletal muscle aging via facilitating gene expression reprogramming. bioRxiv preprint; this version posted May 26, 2023. [CrossRef]
- Xie Y, Hu H, Liu M, Zhou T, Cheng X, Huang W, Cao L. The role and mechanism of histone lactylation in health and diseases. Front Genet. 2022 Aug 23;13:949252. [CrossRef]
- Wei L, Yang X, Wang J, Wang Z, Wang Q, Ding Y, Yu A. H3K18 lactylation of senescent microglia potentiates brain aging and Alzheimer's disease through the NFκB signaling pathway. J Neuroinflammation. 2023 Sep 11;20(1):208. [CrossRef]
- Ng PY, McNeely TL, Baker DJ. Untangling senescent and damage-associated microglia in the aging and diseased brain. FEBS J. 2023 Mar;290(5):1326-1339. [CrossRef]
- Tay TL, Béchade C, D'Andrea I, St-Pierre MK, Henry MS, Roumier A, Tremblay ME. Microglia Gone Rogue: Impacts on Psychiatric Disorders across the Lifespan. Front Mol Neurosci. 2018 Jan 4;10:421. [CrossRef]
- Zhu, H., Guan, A., Liu, J. et al. Noteworthy perspectives on microglia in neuropsychiatric disorders. J Neuroinflammation 20, 223 (2023). [CrossRef]
- Galle E, Wong CW, Ghosh A, Desgeorges T, Melrose K, Hinte LC, Castellano-Castillo D, Engl M, de Sousa JA, Ruiz-Ojeda FJ, De Bock K, Ruiz JR, von Meyenn F. H3K18 lactylation marks tissue-specific active enhancers. Genome Biol. 2022 Oct 3;23(1):207. [CrossRef]
- Hagihara H, Shoji H, Otabi H, Toyoda A, Katoh K, Namihira M, Miyakawa T. Protein lactylation induced by neural excitation. Cell Rep. 2021 Oct 12;37(2):109820. [CrossRef]
- Melanie Föcking, Benjamin Doyle, Nayla Munawar, Eugene T. Dillon, David Cotter, Gerard Cagney; Epigenetic Factors in Schizophrenia: Mechanisms and Experimental Approaches. Molecular Neuropsychiatry 15 March 2019; 5 (1): 6–12. [CrossRef]
- Miwa S, Kashyap S, Chini E, von Zglinicki T. Mitochondrial dysfunction in cell senescence and aging. J Clin Invest. 2022 Jul 1;132(13):e158447. [CrossRef]
- Wiley CD, Campisi J. From Ancient Pathways to Aging Cells-Connecting Metabolism and Cellular Senescence. Cell Metab. 2016 Jun 14;23(6):1013-1021. [CrossRef]
- Chen AN, Luo Y, Yang YH, Fu JT, Geng XM, Shi JP, Yang J. Lactylation, a Novel Metabolic Reprogramming Code: Current Status and Prospects. Front Immunol. 2021 Jun 10;12:688910. [CrossRef]
- Howes, O.D.; Cummings, C.; Chapman, G.E.; Shatalina, E. Neuroimaging in schizophrenia: An overview of findings and their implications for synaptic changes. Neuropsychopharmacology 2022, 48, 151–167.
- Leung, M.; Cheung, C.; Yu, K.; Yip, B.; Sham, P.; Li, Q.; Chua, S.; McAlonan, G. Gray Matter in First-Episode Schizophrenia Before and After Antipsychotic Drug Treatment. Anatomical Likelihood Estimation Meta-analyses With Sample Size Weighting. Schizophr. Bull. 2009, 37, 199–211.
- Fusar-Poli, P.; Smieskova, R.; Kempton, M.; Ho, B.; Andreasen, N.; Borgwardt, S. Progressive brain changes in schizophrenia related to antipsychotic treatment? A meta-analysis of longitudinal MRI studies. Neurosci. Biobehav. Rev. 2013, 37, 1680–1691. [Google Scholar] [CrossRef] [Green Version].
- Ho, B.C.; Andreasen, N.C.; Ziebell, S.; Pierson, R.; Magnotta, V. Long-term antipsychotic treatment and brain volumes: A longitudinal study of first-episode schizophrenia. Arch. Gen. Psychiatry 2011, 68, 128–137.
- Cahn, W.; Pol HE, H.; Lems, E.B.; van Haren, N.E.; Schnack, H.G.; van der Linden, J.A.; Schothorst, P.F.; van Engeland, H.; Kahn, R.S. Brain volume changes in first-episode schizophrenia: A 1-year follow-up study. Arch. Gen. Psychiatry 2002, 59, 1002–1010.
- Nishijo M., Tawara K., Nakagawa H., Honda R., Kido T., Nishijo H., Saito S. 2,3,7,8-Tetrachlorodibenzo-p-dioxin in maternal breast milk and newborn head circumference. J. Expo. Sci. Environ. Epidemiol. 2008;18:246–251. [CrossRef]
- Keo A, Dzyubachyk O, van der Grond J, Hafkemeijer A, van de Berg WDJ, van Hilten JJ, Reinders MJT, Mahfouz A. Cingulate networks associated with gray matter loss in Parkinson's disease show high expression of cholinergic genes in the healthy brain. Eur J Neurosci. 2021 Jun;53(11):3727-3739. Epub 2021 May 4. PMID: 33792979. [CrossRef]
- Chung SJ, Kim YJ, Kim YJ, Lee HS, Jeong SH, Hong JM, Sohn YH, Yun M, Jeong Y, Lee PH. Association Between White Matter Networks and the Pattern of Striatal Dopamine Depletion in Patients With Parkinson Disease. Neurology. 2022 Dec 12;99(24):e2672-e2682. [CrossRef]
- Dean DC 3rd, Sojkova J, Hurley S, Kecskemeti S, Okonkwo O, Bendlin BB, Theisen F, Johnson SC, Alexander AL, Gallagher CL. Alterations of Myelin Content in Parkinson's Disease: A Cross-Sectional Neuroimaging Study. PLoS One. 2016 Oct 5;11(10):e0163774. [CrossRef]
- Brown JS Jr. Effects of bisphenol-A and other endocrine disruptors compared with abnormalities of schizophrenia: an endocrine-disruption theory of schizophrenia. Schizophr Bull. 2009 Jan;35(1):256-78. [CrossRef]
- Newbury JB, Stewart R, Fisher HL, Beevers S, Dajnak D, Broadbent M, Pritchard M, Shiode N, Heslin M, Hammoud R, Hotopf M, Hatch SL, Mudway IS, Bakolis I. Association between air pollution exposure and mental health service use among individuals with first presentations of psychotic and mood disorders: retrospective cohort study. Br J Psychiatry. 2021 Dec;219(6):678-685. [CrossRef]
- Domínguez-Acosta O, Vega L, Estrada-Muñiz E, Rodríguez MS, Gonzalez FJ, Elizondo G. Activation of aryl hydrocarbon receptor regulates the LPS/IFNγ-induced inflammatory response by inducing ubiquitin-proteosomal and lysosomal degradation of RelA/p65. Biochem Pharmacol. 2018 Sep;155:141-149. [CrossRef]
- Kinney DK, Teixeira P, Hsu D, Napoleon SC, Crowley DJ, Miller A, Hyman W, Huang E. Relation of schizophrenia prevalence to latitude, climate, fish consumption, infant mortality, and skin color: a role for prenatal vitamin d deficiency and infections? Schizophr Bull. 2009 May;35(3):582-95. [CrossRef]
- Youngren, K., Inglis, F., Pivirotto, P. et al. Clozapine Preferentially Increases Dopamine Release in the Rhesus Monkey Prefrontal Cortex Compared with the Caudate Nucleus. Neuropsychopharmacol 20, 403–412 (1999). [CrossRef]
- Tronchin G, Akudjedu TN, Ahmed M, Holleran L, Hallahan B, Cannon DM, McDonald C. Progressive subcortical volume loss in treatment-resistant schizophrenia patients after commencing clozapine treatment. Neuropsychopharmacology. 2020 Jul;45(8):1353-1361. Epub 2020 Apr 8. Erratum in: Neuropsychopharmacology. 2021 Sep;46(10):1857-1858 . [CrossRef]
- van Erp TGM, Walton E, Hibar DP, et al. Cortical brain abnormalities in 4474 individuals with schizophrenia and 5098 control subjects via the enhancing neuro imaging genetics through meta analysis (ENIGMA) consortium. Biol Psychiatry 2018; 84: 644–654.
- Zhang X, Zhang Y, Liao J, Jiang S, Yan J, Yue W, Zhang D, Yan H. Progressive Grey Matter Volume Changes in Patients with Schizophrenia over 6 Weeks of Antipsychotic Treatment and Their Relationship to Clinical Improvement. Neurosci Bull. 2018 Oct;34(5):816-826. [CrossRef]
- Liu, N., Xiao, Y., Zhang, W. et al. Characteristics of gray matter alterations in never-treated and treated chronic schizophrenia patients. Transl Psychiatry 10, 136 (2020). [CrossRef]
- Voineskos AN, Mulsant BH, Dickie EW, Neufeld NH, Rothschild AJ, Whyte EM, Meyers BS, Alexopoulos GS, Hoptman MJ, Lerch JP, Flint AJ. Effects of Antipsychotic Medication on Brain Structure in Patients With Major Depressive Disorder and Psychotic Features: Neuroimaging Findings in the Context of a Randomized Placebo-Controlled Clinical Trial. JAMA Psychiatry. 2020 Jul 1;77(7):674-683. [CrossRef]
- Vita A, De Peri L, Deste G, et al. The effect of antipsychotic treatment on cortical gray matter changes in schizophrenia: does the class matter? A meta-analysis and meta-regression of longitudinal magnetic resonance imaging studies. Biol Psychiatry 2015; 78: 403–412.
- Hulshoff Pol HE, Schnack HG, Mandl RC, van Haren NE, Koning H, Collins DL, Evans AC, Kahn RS. Focal gray matter density changes in schizophrenia. Arch Gen Psychiatry. 2001 Dec;58(12):1118-25. [CrossRef]
- Winkler TE, Lederer SL, Kim E, Ben-Yoav H, Kelly DL, Payne GF, Ghodssi R. Molecular processes in an electrochemical clozapine sensor. Biointerphases. 2017 May 1;12(2):02B401. PMID: 28460529. [CrossRef]
- Abdel Majid A. Adam, Hosam A. Saad, Moamen S. Refat, Mohamed S. Hegab .Charge-transfer complexes of antipsychotic drug sulpiride with inorganic and organic acceptors generated through two different approaches: Spectral characterization jJournal: Journal of Molecular Liquids, 2022, p. 118819. [CrossRef]
- Chartoumpekis DV, Zaravinos A, Apidianakis Y, Lagoumintzis G. Editorial: Microbiota and mitochondria: Impact on cell signaling, physiology, and disease. Front Microbiol. 2022 Oct 18;13:1056499. [CrossRef]
- Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The Similarities between Human Mitochondria and Bacteria in the Context of Structure, Genome, and Base Excision Repair System. Molecules 2020, 25, 2857. [CrossRef]
- Liesa M, Shirihai OS. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. 2013 Apr 2;17(4):491-506. [CrossRef]
- Johnson, E.L., Heaver, S.L., Waters, J.L. et al. Sphingolipids produced by gut bacteria enter host metabolic pathways impacting ceramide levels. Nat Commun 11, 2471 (2020). [CrossRef]
- Venable ME, Lee JY, Smyth MJ, Bielawska A, Obeid LM. Role of ceramide in cellular senescence. J Biol Chem. 1995 Dec 22;270(51):30701-8. [CrossRef]
- Dadsena, S., Bockelmann, S., Mina, J.G.M. et al. Ceramides bind VDAC2 to trigger mitochondrial apoptosis. Nat Commun 10, 1832 (2019). [CrossRef]
- Colombini M. Ceramide channels and their role in mitochondria-mediated apoptosis. Biochim Biophys Acta. 2010 Jun-Jul;1797(6-7):1239-44. [CrossRef]
- Debdeep Dutta et al, A defect in mitochondrial fatty acid synthesis impairs iron metabolism and causes elevated ceramide levels, Nature Metabolism (2023). [CrossRef]
- Zietzer A, Düsing P, Reese L, Nickenig G, Jansen F. Ceramide Metabolism in Cardiovascular Disease: A Network With High Therapeutic Potential. Arterioscler Thromb Vasc Biol. 2022 Oct;42(10):1220-1228. [CrossRef]
- Smesny S, Schmelzer CE, Hinder A, Köhler A, Schneider C, Rudzok M, Schmidt U, Milleit B, Milleit C, Nenadic I, Sauer H, Neubert RH, Fluhr JW. Skin ceramide alterations in first-episode schizophrenia indicate abnormal sphingolipid metabolism. Schizophr Bull. 2013 Jul;39(4):933-41. [CrossRef]
- Zhuo C, Zhao F, Tian H, Chen J, Li Q, Yang L, Ping J, Li R, Wang L, Xu Y, Cai Z, Song X. Acid sphingomyelinase/ceramide system in schizophrenia: implications for therapeutic intervention as a potential novel target. Transl Psychiatry. 2022 Jun 23;12(1):260. [CrossRef]
- Yuan X, Bhat OM, Zou Y, Li X, Zhang Y, Li PL. Endothelial Acid Sphingomyelinase Promotes NLRP3 Inflammasome and Neointima Formation During Hypercholesterolemia. J Lipid Res. 2022 Dec;63(12):100298. [CrossRef]
- Xia QS, Wu F, Wu WB, Dong H, Huang ZY, Xu L, Lu FE, Gong J. Berberine reduces hepatic ceramide levels to improve insulin resistance in HFD-fed mice by inhibiting HIF-2α. Biomed Pharmacother. 2022 Jun;150:112955. [CrossRef]
- D. Torralba, F. Baixauli, F. Sanchez-Madrid. Mitochondria know no boundaries: mechanisms and functions of intercellular mitochondrial transfer. Front. Cell. Dev. Biol., 4 (107 (2016), 10.3389/fcell.2016.00107.
- L.H. Fairley, A. Grimm, A. Eckert. Mitochondria transfer in brain injury and disease.Cells., 11 (22) (2022), 10.3390/cells11223603.
- Jackson JG, Robinson MB. Regulation of mitochondrial dynamics in astrocytes: Mechanisms, consequences, and unknowns. Glia. 2018 Jun;66(6):1213-1234. [CrossRef]
- Hogan DB, Jetté N, Fiest KM, Roberts JI, Pearson D, Smith EE, Roach P, Kirk A, Pringsheim T, Maxwell CJ. The Prevalence and Incidence of Frontotemporal Dementia: a Systematic Review. Can J Neurol Sci. 2016 Apr;43 Suppl 1:S96-S109. [CrossRef]
- Course MM, Wang X. Transporting mitochondria in neurons. F1000Res. 2016 Jul 18;5:F1000 Faculty Rev-1735. PMID: 27508065; PMCID: PMC4955021. [CrossRef]
- Nuñez MT, Chana-Cuevas P. New Perspectives in Iron Chelation Therapy for the Treatment of Neurodegenerative Diseases. Pharmaceuticals (Basel). 2018 Oct 19;11(4):109. [CrossRef]
- Thayyullathil, F., Cheratta, A.R., Alakkal, A. et al. Acid sphingomyelinase-dependent autophagic degradation of GPX4 is critical for the execution of ferroptosis. Cell Death Dis 12, 26 (2021). [CrossRef]
- Abdel-Salam OME, Morsy SMY, Sleem AA. The effect of different antidepressant drugs on oxidative stress after lipopolysaccharide administration in mice. EXCLI J. 2011 Dec 8;10:290-302.
- Kann O. The energy demand of fast neuronal network oscillations: insights from brain slice preparations. Front Pharmacol. 2012 Jan 10;2:90. [CrossRef]
- Nakao, K., Singh, M., Sapkota, K. et al. GSK3β inhibition restores cortical gamma oscillation and cognitive behavior in a mouse model of NMDA receptor hypofunction relevant to schizophrenia. Neuropsychopharmacol. 45, 2207–2218 (2020). [CrossRef]
- Linseman DA, Butts BD, Precht TA, Phelps RA, Le SS, Laessig TA, Bouchard RJ, Florez-McClure ML, Heidenreich KA. Glycogen synthase kinase-3beta phosphorylates Bax and promotes its mitochondrial localization during neuronal apoptosis. J Neurosci. 2004 Nov 3;24(44):9993-10002. [CrossRef]
- McNally JM, McCarley RW. Gamma band oscillations: a key to understanding schizophrenia symptoms and neural circuit abnormalities. Curr Opin Psychiatry. 2016 May;29(3):202-10. [CrossRef]
- Tada, M., Kirihara, K., Koshiyama, D. et al. Alterations of auditory-evoked gamma oscillations are more pronounced than alterations of spontaneous power of gamma oscillation in early stages of schizophrenia. Transl Psychiatry 13, 218 (2023). [CrossRef]
- Williams S, Boksa P. Gamma oscillations and schizophrenia. J Psychiatry Neurosci. 2010 Mar;35(2):75-7. [CrossRef]
- Veit J, Handy G, Mossing DP, Doiron B, Adesnik H. Cortical VIP neurons locally control the gain but globally control the coherence of gamma band rhythms. Neuron. 2023 Feb 1;111(3):405-417.e5. [CrossRef]
- Antonoudiou P, Tan YL, Kontou G, Upton AL, Mann EO. Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations. J Neurosci. 2020 Sep 30;40(40):7668-7687. [CrossRef]
- Betterton, R., Mellor, J. & Tsaneva-Atanasova, K. Modulation of hippocampal gamma oscillations by acetylcholine: insights from mathematical and in vitro optogenetic models. BMC Neurosci 16 (Suppl 1), P267 (2015). [CrossRef]
- Çalışkan G, French T, Enrile Lacalle S, Del Angel M, Steffen J, Heimesaat MM, Rita Dunay I, Stork O. Antibiotic-induced gut dysbiosis leads to activation of microglia and impairment of cholinergic gamma oscillations in the hippocampus. Brain Behav Immun. 2022 Jan;99:203-217. [CrossRef]
- Chen Y, Xu J, Chen Y. Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients. 2021 Jun 19;13(6):2099. [CrossRef]
- Klaver R, De Vries HE, Schenk GJ, Geurts JJ. Grey matter damage in multiple sclerosis: a pathology perspective. Prion. 2013 Jan-Feb;7(1):66-75. [CrossRef]
- Keo A, Dzyubachyk O, van der Grond J, Hafkemeijer A, van de Berg WDJ, van Hilten JJ, Reinders MJT, Mahfouz A. Cingulate networks associated with gray matter loss in Parkinson's disease show high expression of cholinergic genes in the healthy brain. Eur J Neurosci. 2021 Jun;53(11):3727-3739. [CrossRef]
- Yohn SE, Weiden PJ, Felder CC, Stahl SM. Muscarinic acetylcholine receptors for psychotic disorders: bench-side to clinic. Trends Pharmacol Sci. 2022 Dec;43(12):1098-1112. [CrossRef]
- Sahu PP, Tseng P. Gamma sensory entrainment for cognitive improvement in neurodegenerative diseases: opportunities and challenges ahead. Front Integr Neurosci. 2023 Apr 17;17:1146687. [CrossRef]
- Yan L, Li H, Qian Y, Zhang J, Cong S, Zhang X, Wu L, Wang Y, Wang M, Yu T. Transcutaneous vagus nerve stimulation: a new strategy for Alzheimer's disease intervention through the brain-gut-microbiota axis? Front Aging Neurosci. 2024 Feb 27;16:1334887. [CrossRef]
- Karpiński P, Żebrowska-Różańska P, Kujawa D, Łaczmański Ł, Samochowiec J, Jabłoński M, Plichta P, Piotrowski P, Bielawski T, Misiak B. Gut microbiota alterations in schizophrenia might be related to stress exposure: Findings from the machine learning analysis. Psychoneuroendocrinology. 2023 Sep;155:106335. [CrossRef]
- Attademo L, Bernardini F, Garinella R, Compton MT. Environmental pollution and risk of psychotic disorders: A review of the science to date. Schizophr Res. 2017 Mar;181:55-59. [CrossRef]
- Breno S. Diniz BS, Seitz-Holland J, Sehgal R, Kasamoto J, Higgins-Chen AT, Lenze E. Geroscience-Centric Perspective for Geriatric Psychiatry: Integrating Aging Biology With Geriatric Mental Health Research. Geriatric Psychiatry (2024). VOLUME 32, ISSUE 1, P1-16, JANUARY 2024.
- Seeman, M.V. Subjective Overview of Accelerated Aging in Schizophrenia. Int. J. Environ. Res. Public Health 2023, 20, 737. [CrossRef]
- Marin, I., Serrano, M. & Pietrocola, F. Recent insights into the crosstalk between senescent cells and CD8 T lymphocytes. npj Aging 9, 8 (2023). [CrossRef]
- Harris MJ, Jeste DV, Gleghorn A, Sewell DD. New-onset psychosis in HIV-infected patients. J Clin Psychiatry. 1991 Sep;52(9):369-76.
- Kozato N, Mishra M, Firdosi M. New-onset psychosis due to COVID-19. BMJ Case Rep. 2021 Apr 26;14(4):e242538. [CrossRef]
- Luís C, Maduro AT, Pereira P, Mendes JJ, Soares R, Ramalho R. Nutritional senolytics and senomorphics: Implications to immune cells metabolism and aging - from theory to practice. Front Nutr. 2022 Sep 8;9:958563. [CrossRef]
- An S, Cho SY, Kang J, Lee S, Kim HS, Min DJ, Son E, Cho KH. Inhibition of 3-phosphoinositide-dependent protein kinase 1 (PDK1) can revert cellular senescence in human dermal fibroblasts. Proc Natl Acad Sci U S A. 2020 Dec 8;117(49):31535-31546. [CrossRef]
- Gregory M SolisRozina KardakarisElizabeth R ValentineLiron Bar-PeledAlice L ChenMegan M BlewettMark A McCormickJames R WilliamsonBrian KennedyBenjamin F CravattMichael Petrascheck (2018) Translation attenuation by minocycline enhances longevity and proteostasis in old post-stress-responsive organisms eLife 7:e40314. [CrossRef]
- Deakin B, Suckling J, Dazzan P, Joyce E, Lawrie SM, Upthegrove R, Husain N, Chaudhry IB, Dunn G, Jones PB, Lisiecka-Ford D, Lewis S, Barnes TRE, Williams SCR, Pariante CM, Knox E, Drake RJ, Smallman R, Barnes NM. Minocycline for negative symptoms of schizophrenia and possible mechanistic actions: the BeneMin RCT. Southampton (UK): NIHR Journals Library;
- Abir MH, Mahamud AGMSU, Tonny SH, Anu MS, Hossain KHS, Protic IA, Khan MSU, Baroi A, Moni A, Uddin MJ. Pharmacological potentials of lycopene against aging and aging-related disorders: A review. Food Sci Nutr. 2023 Jun 27;11(10):5701-5735. [CrossRef]
- Perrott KM, Wiley CD, Desprez PY, Campisi J. Apigenin suppresses the senescence-associated secretory phenotype and paracrine effects on breast cancer cells. Geroscience. 2017 Apr;39(2):161-173. [CrossRef]
- Elsallabi O, Patruno A, Pesce M, Cataldi A, Carradori S, Gallorini M. Fisetin as a Senotherapeutic Agent: Biopharmaceutical Properties and Crosstalk between Cell Senescence and Neuroprotection. Molecules. 2022 Jan 23;27(3):738. [CrossRef]
- Kumar R, Sharma A, Kumari A, Gulati A, Padwad Y, Sharma R. Epigallocatechin gallate suppresses premature senescence of preadipocytes by inhibition of PI3K/Akt/mTOR pathway and induces senescent cell death by regulation of Bax/Bcl-2 pathway. Biogerontology. 2019 Apr;20(2):171-189. [CrossRef]
- Li W, He Y, Zhang R, Zheng G, Zhou D. The curcumin analog EF24 is a novel senolytic agent. Aging (Albany NY). 2019 Jan 28;11(2):771-782. [CrossRef]
- Dang Y, An Y, He J, Huang B, Zhu J, Gao M, Zhang S, Wang X, Yang B, Xie Z. Berberine ameliorates cellular senescence and extends the lifespan of mice via regulating p16 and cyclin protein expression. Aging Cell. 2020 Jan;19(1):e13060. [CrossRef]
- Islam MT, Tuday E, Allen S, Kim J, Trott DW, Holland WL, Donato AJ, Lesniewski LA. Senolytic drugs, dasatinib and quercetin, attenuate adipose tissue inflammation, and ameliorate metabolic function in old age. Aging Cell. 2023 Feb;22(2):e13767. [CrossRef]
- von Kobbe C. Targeting senescent cells: approaches, opportunities, challenges. Aging (Albany NY). 2019 Nov 30;11(24):12844-12861. [CrossRef]
- Suda M, Shimizu I, Katsuumi G, Yoshida Y, Hayashi Y, Ikegami R, Matsumoto N, Yoshida Y, Mikawa R, Katayama A, Wada J, Seki M, Suzuki Y, Iwama A, Nakagami H, Nagasawa A, Morishita R, Sugimoto M, Okuda S, Tsuchida M, Ozaki K, Nakanishi-Matsui M, Minamino T. Senolytic vaccination improves normal and pathological age-related phenotypes and increases lifespan in progeroid mice. Nat Aging. 2021 Dec;1(12):1117-1126. [CrossRef]
- Poblocka, M., Bassey, A.L., Smith, V.M. et al. Targeted clearance of senescent cells using an antibody-drug conjugate against a specific membrane marker. Sci Rep 11, 20358 (2021). [CrossRef]
- Nicolson GL, Ferreira de Mattos G, Ash M, Settineri R, Escribá PV. Fundamentals of Membrane Lipid Replacement: A Natural Medicine Approach to Repairing Cellular Membranes and Reducing Fatigue, Pain, and Other Symptoms While Restoring Function in Chronic Illnesses and Aging. Membranes (Basel). 2021 Nov 29;11(12):944. [CrossRef]
- Zhou M, Ren H, Han J, Wang W, Zheng Q, Wang D. Protective Effects of Kaempferol against Myocardial Ischemia/Reperfusion Injury in Isolated Rat Heart via Antioxidant Activity and Inhibition of Glycogen Synthase Kinase-3β. Oxid Med Cell Longev. 2015;2015:481405. [CrossRef]
- Jope RS, Roh MS. Glycogen synthase kinase-3 (GSK3) in psychiatric diseases and therapeutic interventions. Curr Drug Targets. 2006 Nov;7(11):1421-34. [CrossRef]
- Jin S, Zhang L, Wang L. Kaempferol, a potential neuroprotective agent in neurodegenerative diseases: From chemistry to medicine. Biomed Pharmacother. 2023 Sep;165:115215. [CrossRef]
- Nicolson GL, Ash ME. Membrane Lipid Replacement for chronic illnesses, aging and cancer using oral glycerolphospholipid formulations with fructooligosaccharides to restore phospholipid function in cellular membranes, organelles, cells and tissues. Biochim Biophys Acta Biomembr. 2017 Sep;1859(9 Pt B):1704-1724. [CrossRef]
- Schmidt NW; Mishra A; Lai GH; Davis M; Sanders LK ; Dat T; Garcia A; Tai KP; McCray J; Paul B; Ouellette AJ; Selsted ME; Wong GCL Criterion for amino acid composition of defensins and antimicrobial peptides based on geometry of membrane destabilization. J. Am. Chem. Soc 2011, 133, 6720–6727.
- Blankenfeldt W, Parsons JF. The structural biology of phenazine biosynthesis. Curr Opin Struct Biol. 2014 Dec;29:26-33. Epub 2014 Sep 15. PMID: 25215885; PMCID: PMC4268259. [CrossRef]
- Pierson LS 3rd, Pierson EA. Metabolism and function of phenazines in bacteria: impacts on the behavior of bacteria in the environment and biotechnological processes. Appl Microbiol Biotechnol. 2010 May;86(6):1659-70. [CrossRef]
- Abdelaziz, A.A., Kamer, A.M.A., Al-Monofy, K.B. et al. Pseudomonas aeruginosa’s greenish-blue pigment pyocyanin: its production and biological activities. Microb Cell Fact 22, 110 (2023). [CrossRef]
- Ohlendorf B, Schulz D, Erhard A, Nagel K, Imhoff JF. Geranylphenazinediol, an acetylcholinesterase inhibitor produced by a Streptomyces species. J Nat Prod. 2012 Jul 27;75(7):1400-4. [CrossRef]
- Paul SM, Yohn SE, Popiolek M, Miller AC, Felder CC. Muscarinic Acetylcholine Receptor Agonists as Novel Treatments for Schizophrenia. Am J Psychiatry. 2022 Sep;179(9):611-627. [CrossRef]
- Wang X, Abbas M, Zhang Y, Elshahawi SI, Ponomareva LV, Cui Z, Van Lanen SG, Sajid I, Voss SR, Shaaban KA, Thorson JS. Baraphenazines A-G, Divergent Fused Phenazine-Based Metabolites from a Himalayan Streptomyces. J Nat Prod. 2019 Jun 28;82(6):1686-1693. [CrossRef]
- Cha JW, Lee S, Kim MC, Thida M, Lee JW, Park JS, et al. Pontemazines a and B, phenazine derivatives containing a methylamine linkage from Streptomyces sp. UT1123 and their protective effect to HT-22 neuronal cells. Bioorganic & Medicinal Chemistry Letters. 2015;25:5083-5086. [CrossRef]
- Kim WG, Ryoo IJ, Yun BS, Shin-ya K, Seto H, Yoo ID. Phenazostatin C, a new diphenazine with neuronal cell protecting activity from Streptomyces sp. J Antibiot (Tokyo). 1999 Aug;52(8):758-61. [CrossRef]
- Kato S., Shindo K., Yamagishi Y., Matsuoka M., Kawai H., Mochizuki J. Phenazoviridin, a novel free radical scavenger from Streptomyces sp. taxonomy, fermentation, isolation, structure elucidation and biological properties. J. Antibiot. 1993;46:1485–1493. doi:.
- Nadeem A., Meijler MM. Unraveling the Antibacterial and Iron Chelating Activity of N-Oxide Hydroxy-Phenazine natural Products and Synthetic Analogs against Staphylococcus aureus. Israel Journal of Chemistry (2023) Volume63, Issue 5-6. [CrossRef]
- Heitmann ASB, Zanjani AAH, Klenow MB, Mularski A, Sønder SL, Lund FW, Boye TL, Dias C, Bendix PM, Simonsen AC, Khandelia H, Nylandsted J. Phenothiazines alter plasma membrane properties and sensitize cancer cells to injury by inhibiting annexin-mediated repair. J Biol Chem. 2021 Aug;297(2):101012. [CrossRef]
- Boonnoy, P., Jarerattanachat, V., Karttunen, M. & Wong-ekkabut, J. Bilayer Deformation, Pores, and Micellation Induced by Oxidized Lipids. J. Phys. Chem. Lett. 6, 4884–4888 (2015).
- Voronova O, Zhuravkov S, Korotkova E, Artamonov A, Plotnikov E. Antioxidant Properties of New Phenothiazine Derivatives. Antioxidants (Basel). 2022 Jul 14;11(7):1371. [CrossRef]
- Keynes RG, Karchevskaya A, Riddall D, Griffiths CH, Bellamy TC, Chan AWE, Selwood DL, Garthwaite J. N10 -carbonyl-substituted phenothiazines inhibiting lipid peroxidation and associated nitric oxide consumption powerfully protect brain tissue against oxidative stress. Chem Biol Drug Des. 2019 Sep;94(3):1680-1693. [CrossRef]
- Philot EA, da Mata Lopes D, de Souza AT, Braz AS, Nantes IL, Rodrigues T, Perahia D, Miteva MA, Scott LP. Binding of phenothiazines into allosteric hydrophobic pocket of human thioredoxin 1. Eur Biophys J. 2016 Apr;45(3):279-86. [CrossRef]
- Ho BC, Andreasen NC, Ziebell S, et al. Long-term antipsychotic treatment and brain volumes: a longitudinal study of first-episode schizophrenia. Arch Gen Psychiatry 2011; 68: 128–137.
- Veijola J, Guo JY, Moilanen JS, et al. Longitudinal changes in total brain volume in schizophrenia: relation to symptom severity, cognition antipsychotic medication. PLoS One 2014; 9: 101689.
- Engwa G.A., Ayuk E.L., Igbojekwe B.U., Unaegbu M. Potential Antioxidant Activity of New Tetracyclic and Pentacyclic Nonlinear Phenothiazine Derivatives. Biochem. Res. Int. 2016;2016:9896575. [CrossRef]
- Clark MA, Shay JW. Mitochondrial transformation of mammalian cells. Nature. (1982) 295:605–7. [CrossRef]
- Katrangi E, D’Souza G, Boddapati SV, Kulawiec M, Singh KK, Bigger B, et al. Xenogenic transfer of isolated murine mitochondria into human rho0 cells can improve respiratory function. Rejuvenation Res. (2007) 10:561–70. [CrossRef]
- Pacak AP, Preble JM, Kondo H, Seibel P, Levitsky S, del Nido PJ, et al. Actin-dependent mitochondrial internalization in cardiomyocytes: evidence for rescue of mitochondrial function. Biol Open. (2015) 4:622–6. [CrossRef]
- Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, et al. Transfer of mitochondria from astrocytes to neurons after stroke. Nature. (2016) 535:551–5. [CrossRef]
- Ali Pour P, Hosseinian S, Kheradvar A. Mitochondrial transplantation in cardiomyocytes: foundation, methods, and outcomes. Am J Physiol Cell Physiol. (2021) 321:C489–503. [CrossRef]
- Sasaki, D., Abe, J., Takeda, A. et al. Transplantation of MITO cells, mitochondria activated cardiac progenitor cells, to the ischemic myocardium of mouse enhances the therapeutic effect. Sci Rep 12, 4344 (2022). [CrossRef]
- Chen T, Majerníková N, Marmolejo-Garza A, Trombetta-Lima M, Sabogal-Guáqueta AM, Zhang Y, Ten Kate R, Zuidema M, Mulder PPMFA, den Dunnen W, Gosens R, Verpoorte E, Culmsee C, Eisel ULM, Dolga AM. Mitochondrial transplantation rescues neuronal cells from ferroptosis. Free Radic Biol Med. 2023 Nov 1;208:62-72. [CrossRef]
- Um, J.-H.; Lee, K.-M.; Kim, Y.-Y.; Lee, D.-Y.; Kim, E.; Kim, D.-H.; Yun, J. Berberine Induces Mitophagy through Adenosine Monophosphate-Activated Protein Kinase and Ameliorates Mitochondrial Dysfunction in PINK1 Knockout Mouse Embryonic Fibroblasts. Int. J. Mol. Sci. 2024, 25, 219. [CrossRef]
- Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, Ji X, Lo EH. Transfer of mitochondria from astrocytes to neurons after stroke. Nature. 2016 Jul 28;535(7613):551-5. Erratum in: Nature. 2016 Sep 14;539(7627):123. [CrossRef]
- Zhang S, Qin C, Safe SH. Flavonoids as aryl hydrocarbon receptor agonists/antagonists: effects of structure and cell context. Environ Health Perspect. 2003 Dec;111(16):1877-82. PMID: 14644660; PMCID: PMC1241760. [CrossRef]
- Linck VM, Ganzella M, Herrmann AP, Okunji CO, Souza DO, Antonelli MC, Elisabetsky E. Original mechanisms of antipsychotic action by the indole alkaloid alstonine (Picralima nitida). Phytomedicine. 2015 Jan 15;22(1):52-5. [CrossRef]
- Cordaro, M.; Cuzzocrea, S.; Crupi, R. An Update of Palmitoylethanolamide and Luteolin Effects in Preclinical and Clinical Studies of Neuroinflammatory Events. Antioxidants 2020, 9, 216. [CrossRef]
- McGovern K, Castro AC, Cavanaugh J, Coma S, Walsh M, Tchaicha J, Syed S, Natarajan P, Manfredi M, Zhang XM, Ecsedy J. Discovery and Characterization of a Novel Aryl Hydrocarbon Receptor Inhibitor, IK-175, and Its Inhibitory Activity on Tumor Immune Suppression. Mol Cancer Ther. 2022 Aug 2;21(8):1261-1272. [CrossRef]
- Kang S, Lee AG, Im S, Oh SJ, Yoon HJ, Park JH, Pak YK. A Novel Aryl Hydrocarbon Receptor Antagonist HBU651 Ameliorates Peripheral and Hypothalamic Inflammation in High-Fat Diet-Induced Obese Mice. Int J Mol Sci. 2022 Nov 28;23(23):14871. [CrossRef]
- Zai W, Chen W, Liu H, Ju D. Therapeutic Opportunities of IL-22 in Non-Alcoholic Fatty Liver Disease: From Molecular Mechanisms to Clinical Applications. Biomedicines. 2021 Dec 14;9(12):1912. [CrossRef]
- Sfera, A. Six Decades of Dopamine Hypothesis: Is Aryl Hydrocarbon Receptor the New D2? Reports 2023, 6, 36. [CrossRef]
- Shi C, Su C, Cen L, Han L, Tang J, Wang Z, Shi X, Ju D, Cao Y, Zhu H. Vunakizumab-IL22, a Novel Fusion Protein, Promotes Intestinal Epithelial Repair and Protects against Gut Injury Induced by the Influenza Virus. Biomedicines. 2023 Apr 12;11(4):1160. [CrossRef]
- Kim CJ, Nazli A, Rojas OL, Chege D, Alidina Z, Huibner S, Mujib S, Benko E, Kovacs C, Shin LY, Grin A, Kandel G, Loutfy M, Ostrowski M, Gommerman JL, Kaushic C, Kaul R. A role for mucosal IL-22 production and Th22 cells in HIV-associated mucosal immunopathogenesis. Mucosal Immunol. 2012 Nov;5(6):670-80. [CrossRef]
- Nsairat H, Khater D, Odeh F, Al-Adaileh F, Al-Taher S, Jaber AM, Alshaer W, Al Bawab A, Mubarak MS. Lipid nanostructures for targeting brain cancer. Heliyon. 2021 Sep 16;7(9):e07994. [CrossRef]
- Aldosari BN, Alfagih IM, Almurshedi AS. Lipid Nanoparticles as Delivery Systems for RNA-Based Vaccines. Pharmaceutics. 2021 Feb 2;13(2):206. [CrossRef]
- Sun D, Lu ZR. Structure and Function of Cationic and Ionizable Lipids for Nucleic Acid Delivery. Pharm Res. 2023 Jan;40(1):27-46. [CrossRef]
- Brannagan TH 3rd, Berk JL, Gillmore JD, Maurer MS, Waddington-Cruz M, Fontana M, Masri A, Obici L, Brambatti M, Baker BF, Hannan LA, Buchele G, Viney NJ, Coelho T, Nativi-Nicolau J. Liver-directed drugs for transthyretin-mediated amyloidosis. J Peripher Nerv Syst. 2022 Dec;27(4):228-237. [CrossRef]
Figure 1.
In the cytosol, AhR is stabilized by two HSP90 molecules. DA, oxidized lipids (and toxic ceramide), clozapine, serotonin, melatonin, and vitamin D3 are AhR ligands. Pollutants, such as phthalate and bisphenol A (BPA) are also AhR ligands. In contrast, aripiprazole binds AhR chaperone, HSP90. HSP90 prevents AhR entry into the nucleus where it drives the transcription of genes, including those for cellular senescence.
Figure 1.
In the cytosol, AhR is stabilized by two HSP90 molecules. DA, oxidized lipids (and toxic ceramide), clozapine, serotonin, melatonin, and vitamin D3 are AhR ligands. Pollutants, such as phthalate and bisphenol A (BPA) are also AhR ligands. In contrast, aripiprazole binds AhR chaperone, HSP90. HSP90 prevents AhR entry into the nucleus where it drives the transcription of genes, including those for cellular senescence.
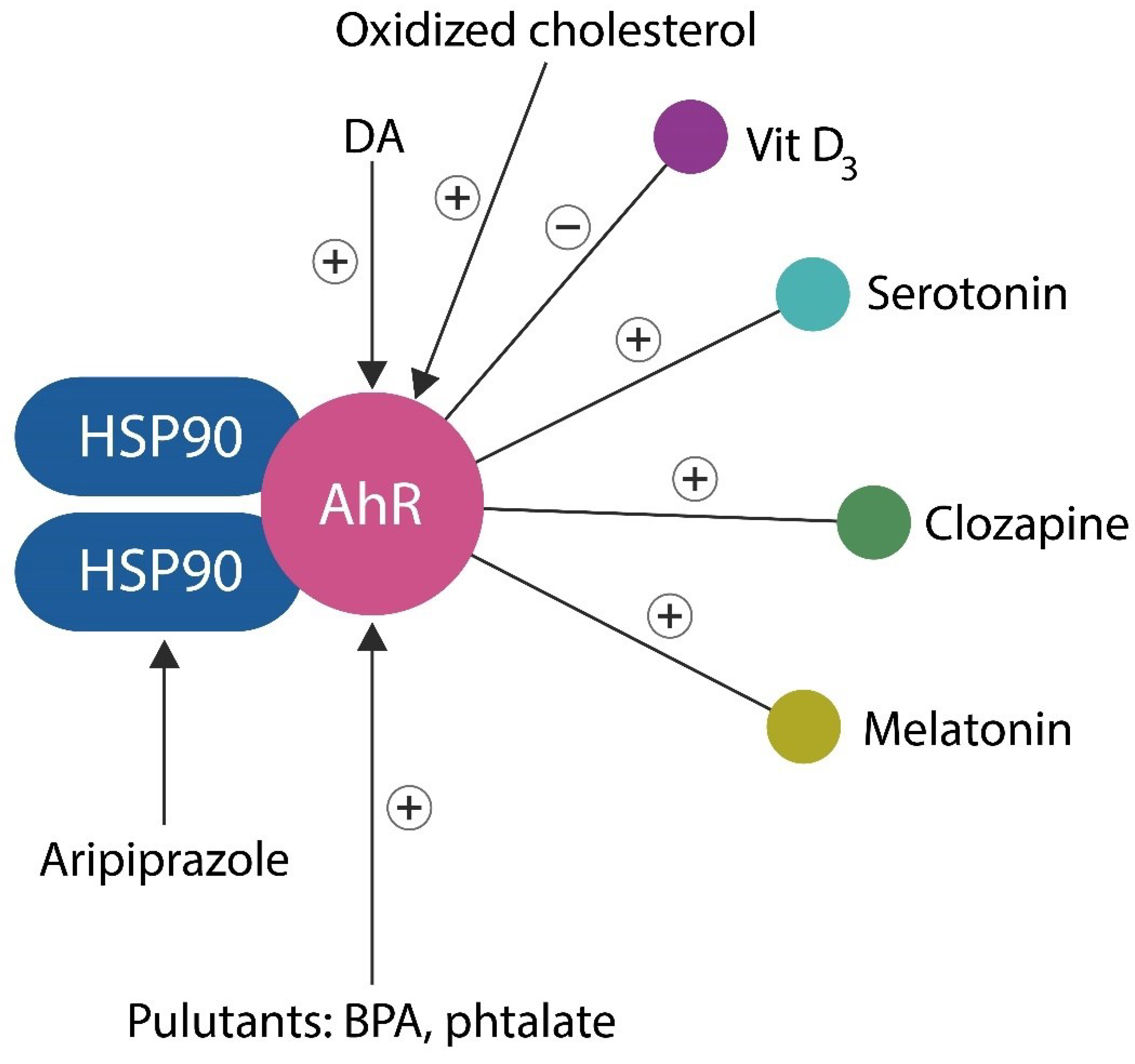
Figure 2.
Glial cells, including astrocytes supply the neuron with healthy mitochondria via tunneling nanotubules, preventing apoptosis. In addition, astrocytes, prevent neuronal ferroptosis by transferring antioxidants, including GPX-4. The astrocyte uptakes cystine via cystine/glutamate antiporter (Xc-). Cysteine can also be obtained from methionine via glutathione. Fe2+ enters the neuron through the transferrin receptor 1 (TRF-1), is stored in ferritin, and requires ferritinophagy to be released. Excess Fe2+exits the neuron via ferroportin (FPT) channels.
Figure 2.
Glial cells, including astrocytes supply the neuron with healthy mitochondria via tunneling nanotubules, preventing apoptosis. In addition, astrocytes, prevent neuronal ferroptosis by transferring antioxidants, including GPX-4. The astrocyte uptakes cystine via cystine/glutamate antiporter (Xc-). Cysteine can also be obtained from methionine via glutathione. Fe2+ enters the neuron through the transferrin receptor 1 (TRF-1), is stored in ferritin, and requires ferritinophagy to be released. Excess Fe2+exits the neuron via ferroportin (FPT) channels.
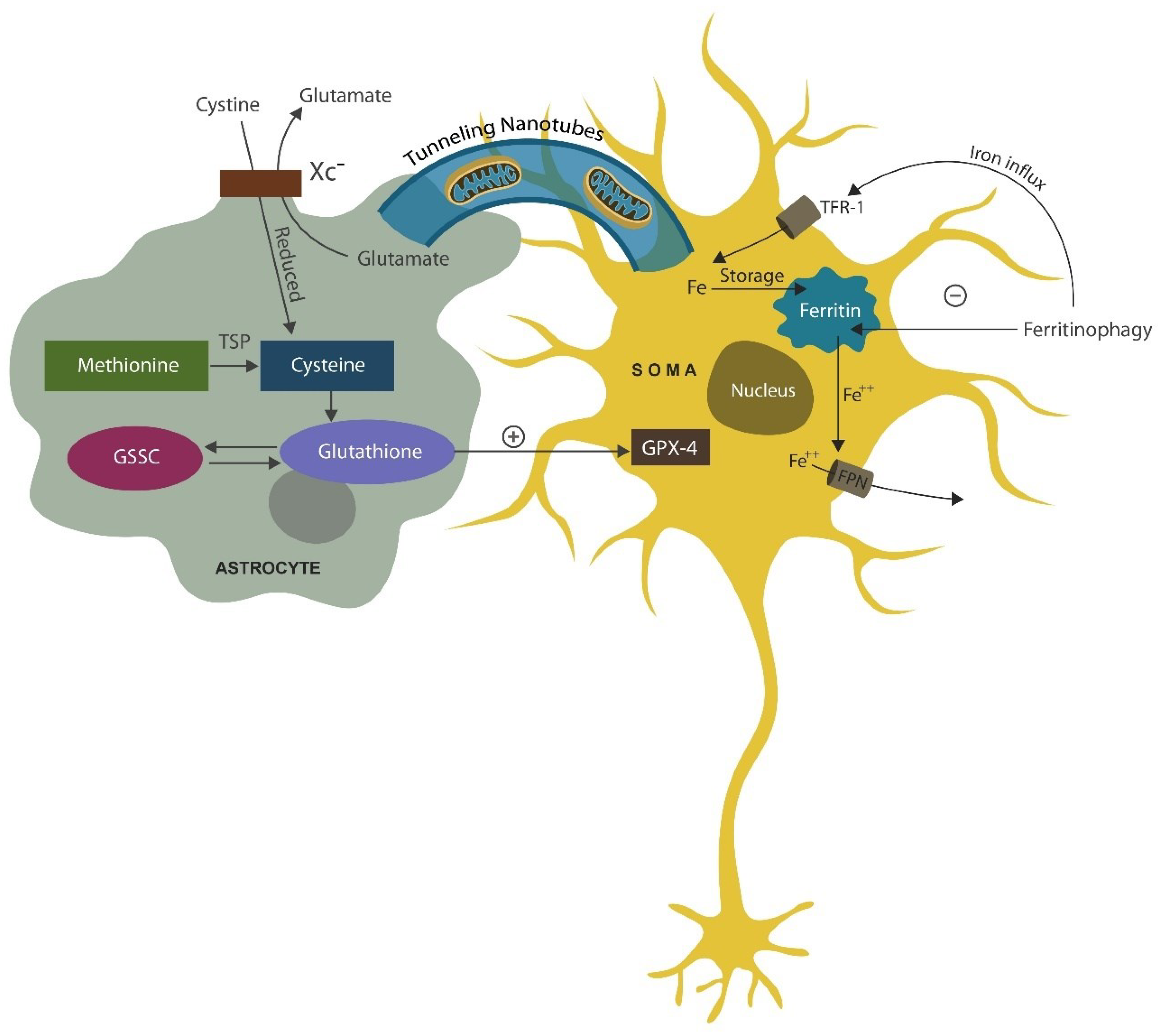
Figure 3.
Both AhR and GSK-3β are expressed in cytosol as well as in the mitochondria (mitoAhR). Akt negatively phosphorylates GSK-3β, inhibiting its function. Toxic ceramides and iron activate GSK-3β, resulting in excessive mitochondrial ROS (mtROS) which activate mitoAhR, triggering organelle death. mtROS can also cause mitochondrial demise by activating cytochrome C and caspase 3. Natural compounds, berberine and kaempferol inhibit GSK-3β, averting organelle death.
Figure 3.
Both AhR and GSK-3β are expressed in cytosol as well as in the mitochondria (mitoAhR). Akt negatively phosphorylates GSK-3β, inhibiting its function. Toxic ceramides and iron activate GSK-3β, resulting in excessive mitochondrial ROS (mtROS) which activate mitoAhR, triggering organelle death. mtROS can also cause mitochondrial demise by activating cytochrome C and caspase 3. Natural compounds, berberine and kaempferol inhibit GSK-3β, averting organelle death.
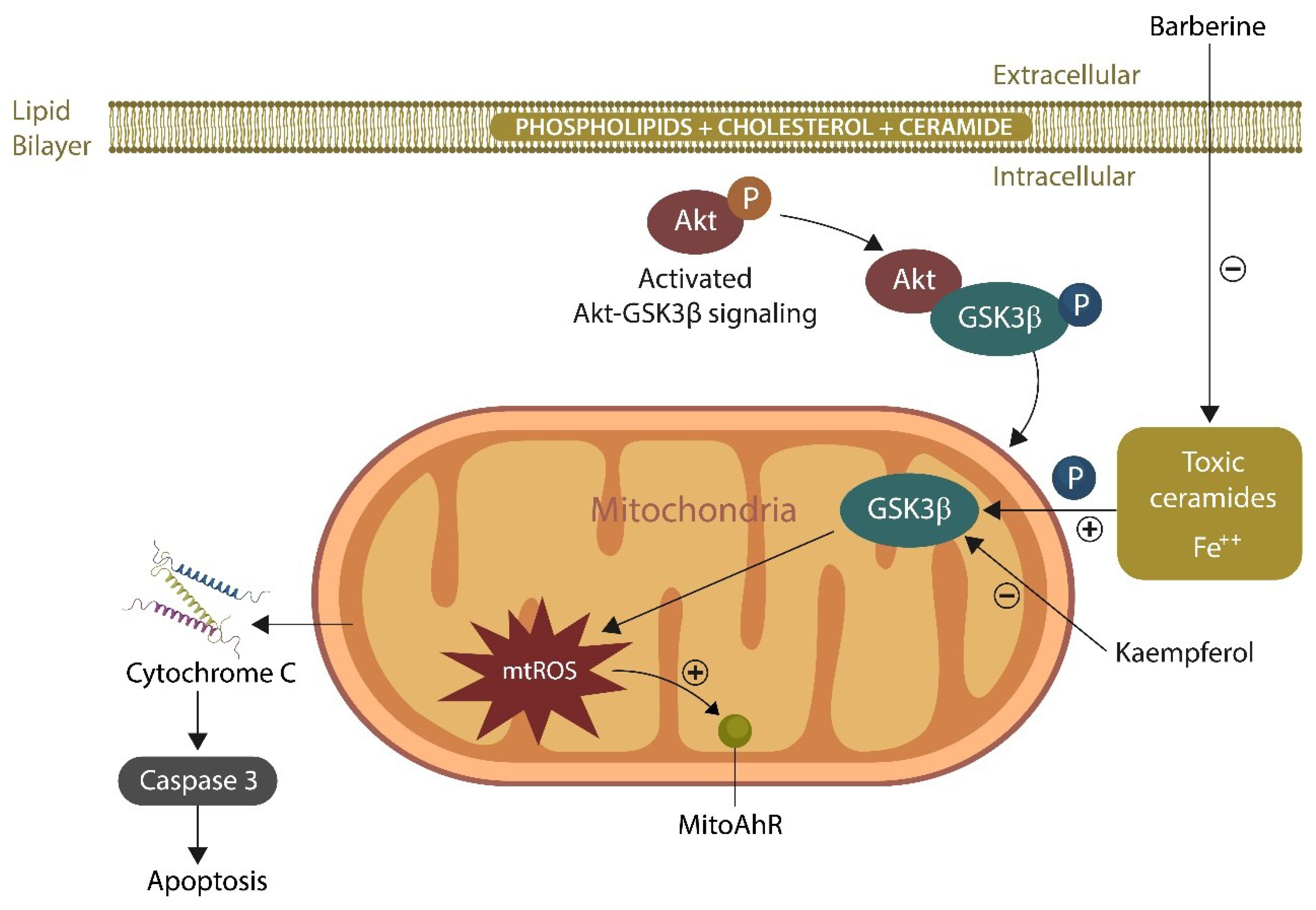
Figure 4.
The lipid bilayer of neuronal membrane is easily oxidated when intracellular iron is upregulated. Oxysterols, including 7-Ketocholesterol (a toxic oxide), and oxidated phospholipids alter the biophysical properties of cell membranes, disrupting neurotransmission. In addition, oxidized lipids activate AhR, triggering premature neuronal senescence. Phenazines, Phenothiazines, and their derivatives, intercalate themselves into the lipid bilayer, repairing the lipids in cellular and/or mitochondrial membranes.
Figure 4.
The lipid bilayer of neuronal membrane is easily oxidated when intracellular iron is upregulated. Oxysterols, including 7-Ketocholesterol (a toxic oxide), and oxidated phospholipids alter the biophysical properties of cell membranes, disrupting neurotransmission. In addition, oxidized lipids activate AhR, triggering premature neuronal senescence. Phenazines, Phenothiazines, and their derivatives, intercalate themselves into the lipid bilayer, repairing the lipids in cellular and/or mitochondrial membranes.
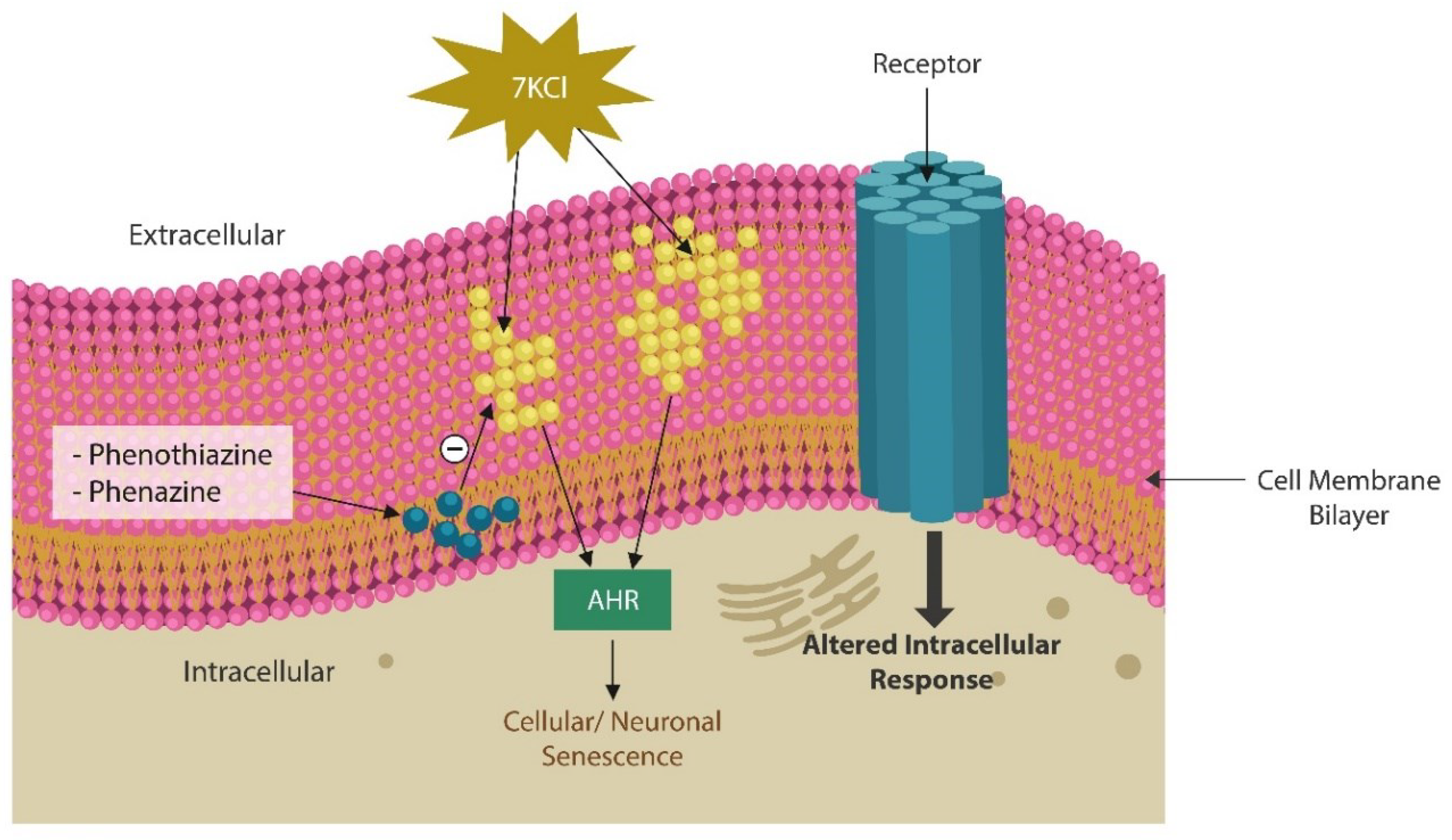
Figure 5.
Phenazine vs. Phenothiazine: similarities.
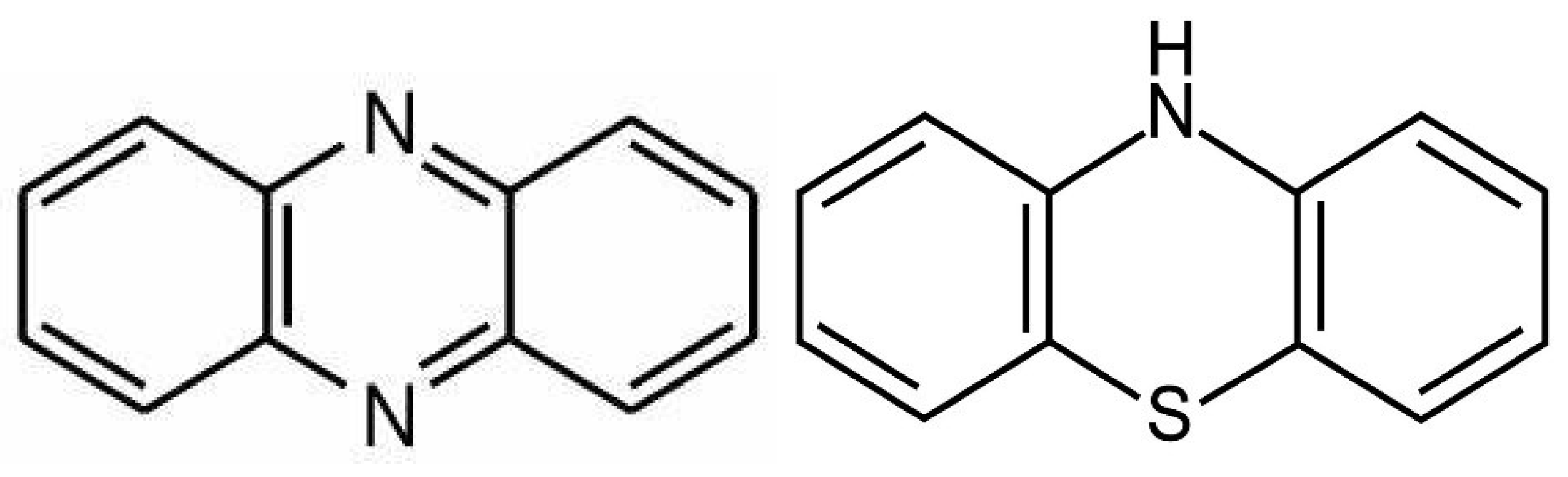
Figure 6.
Membrane lipid replacement (MLR) replenishes oxidized lipids from the plasma and mitochondrial membrane, such as oxysterols, ceramide, and oxidized phospholipids (OxPL) with natural glycerophospholipids. The oxidized lipids inhibit AKT (by serine 9 phosphorylation), activating GSK-3β, an enzyme associated with SCZ, SLDs, and cancer (by p53 inhibition). Berberine and Kaempferol inhibit GSK-3β activation by different mechanisms. Ceramide activates GSK-3β by dephosphorylation at serine 9.
Figure 6.
Membrane lipid replacement (MLR) replenishes oxidized lipids from the plasma and mitochondrial membrane, such as oxysterols, ceramide, and oxidized phospholipids (OxPL) with natural glycerophospholipids. The oxidized lipids inhibit AKT (by serine 9 phosphorylation), activating GSK-3β, an enzyme associated with SCZ, SLDs, and cancer (by p53 inhibition). Berberine and Kaempferol inhibit GSK-3β activation by different mechanisms. Ceramide activates GSK-3β by dephosphorylation at serine 9.
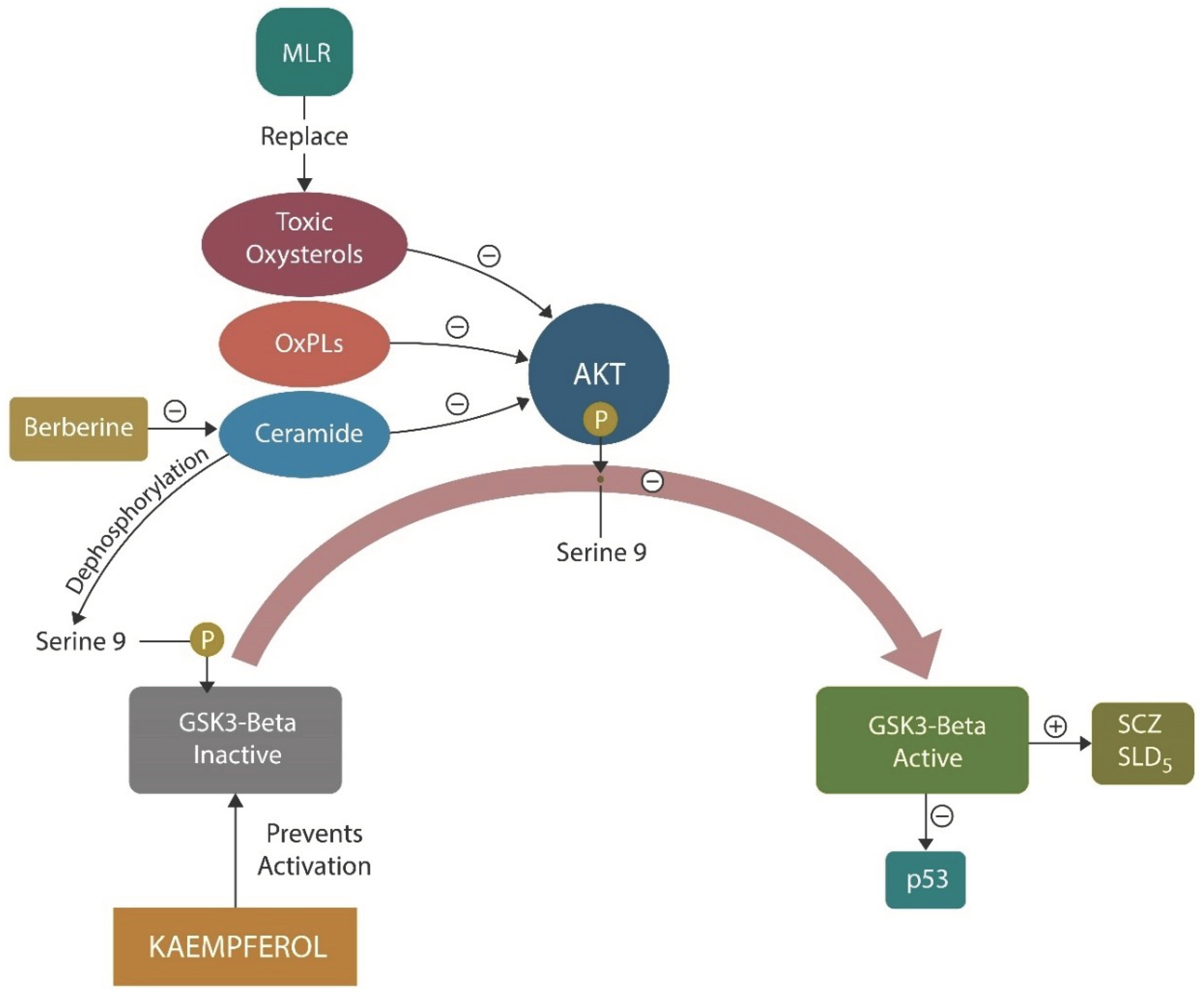

Table 1.
Natural senolytics and their source.
| Senolytics | Source | References |
| Lycopene | Grape skin, guava, grapefruit, blueberries, tomatoes | [166] |
| Apigenin | Cabbage, blueberries, acai berries | [167] |
| Fisetin | Strawberries, onions, apples, mangoes, persimmons, kiwis. | [168] |
| Curcumin and analog EF24 | chicken, beef, tofu, vegetables | [169] |
| Epigallocatechin gallate | Apples, blackberries, broad beans, cherries, black grapes, pears, raspberries, and chocolate | [170] |
| Berberine | Oregon grape, phellodendron, and tree turmeric. | [171] |
| Quercetin | Fruits, apples, onions, parsley, sage, tea, and red wine | [172] |
| Kaempferol | Fruits and vegetables. | [173] |
Table 2.
Naturally occurring and synthetic compounds with potential benefit in SCZ and SLDs.
| Compound | Naturally occurring | Synthetic |
|---|---|---|
| Phenazines | geranyl-phenazine, bara-phenazines A–G | Pontemazines A and B; Halogenated phenazines |
| Phenothiazines | Propenyl-phenothiazine; N10-carbonyl-substituted phenothiazines |
|
| GSK-3β inhibitors | Kaempferol; Curcumin | Lithium Valproic acid Clozapine; Olanzapine |
| AhR inhibitors | quercetin, apigenin, alstonine, luteolin | IK-175; HBU651 |
| Acid sphingomyelinase (ASM) inhibitors | fluvoxamine rosuvastatin tricyclic antidepressants |
|
| Dopamine D1R agonists | A68930; A77636; Dihydrexidine |
|
| Mitochondrial export | SSRIs | |
| ACh agonists |
Catharanthus roseus; Salvia spp. (Lamiaceae) |
Cholinesterase inhibitors: donepezil, galantamine, rivastigmine |
| Senotherapeutics | Please see Table 1 | Senotherapeutic antibiotics |
| Ferroptosis inhibitors | Natural flavonoids; Berberine |
Fluvoxamine; SSRI; N acetyl cysteine (NAC) |
| Recombinant IL-22 | ||
| Membrane lipid replacement | ||
| Mitochondrial transplantation | ||
| 40 HZ entrainment with sensory stimuli | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



