
Submitted:
27 March 2024
Posted:
28 March 2024
You are already at the latest version
Abstract
Berberis vulgaris L. (Berberidaceae) is a shrub that has been widely used in European folk medicine as an anti-inflammatory and antimicrobial agent. The purpose of our study was to elucidate the mechanisms of antiproliferative action of the plant’s methanolic root extract (BVR) against colon cancer cells. Cytotoxicity and pro-apoptotic activity were determined in human colon adenocarcinoma cell lines (LS180 and HT-29) and control colon epithelial CCD841 CoN cells. According to the MTT assay, after 48 h of cell exposure, the IC50 values were as follows: 4.3; 46.1 and 50.2 µg/mL for LS180, HT-29, and CCD841 CoN cells, respectively, showing the greater sensitivity of cancer cells to BVR. The Cell Death Detection ELISAPLUS kit demonstrated that BVR induced programmed cell death only against HT-29 cells. RT-qPCR with relative quantification method showed significant changes in the expression of genes related to apoptosis in both colon cancer cell lines. The genes BCL2L1 (126.84-366.66%), BCL2L2 (277.08-330.12%), CASP3 (180.36-261.36%) and CASP9 (158.08-213.46%) had significantly elevated expression, while BCL2 (23.08-52.06%) had reduced expression. Furthermore, in a panel of antioxidant tests, BVR showed positive effects (63.93±0.01, 122.92±0.01, and 220.29±0.02 mg Trolox equivalents (TE)/g in the DPPH•, ABTS•+ and ORAC assays, respectively). In the lipoxygenase (LOX) inhibition test, BVR revealed 62,60±0.87% of enzyme inhibition. The chemical composition of BVR was determined using UHPLC-UV-CAD-MS/MS analysis and confirmed the presence of several known alkaloids, including berberine, as well as other alkaloids and two derivatives of hydroxycinnamic acid (ferulic and sinapic acid hexosides). This is the first such extensive assessment of the biological activity of BVR. The results are very promising and encourage for the use of BVR as a comprehensive chemopreventive agent (anti-inflammatory, antioxidant, and pro-apoptotic) in colorectal cancer.
Keywords:
Berberis vulgaris L.
; antioxidants
; anti-inflammatory
; antiproliferative
; colon cancer
; inflammatory bowel disease
; phytopharmacology
1. Introduction
Colorectal cancer is a primary malignant tumor of the large intestine, originating in the mucosa of the epithelium of the large intestine [1,2]. According to the World Health Organisation, it is the most common cancer, along with breast, lung, rectum, and prostate cancers; in 2020, colon cancer was third in terms of occurrence (1.93 million cases) and second in terms of mortality (916,000 deaths) [3]. Lifestyle factors associated with an increased risk of colorectal cancer include diet, obesity and metabolic syndrome, physical inactivity, smoking, and moderate to heavy alcohol consumption [4]. However, non-modifiable risk factors in the development of the disease also exist. One of them is inflammatory bowel disease. Inflammatory bowel disease significantly increases the risk of developing colorectal cancer and is crucial in its pathogenesis; it may occur from the initial stage of tumorigenesis and facilitate the development of colorectal cancer. Inflammatory bowel disease, its spread and exacerbation, is also closely related to oxidative stress, which leads to damage to gastrointestinal cells, including DNA damage, protein aggregation, and membrane dysfunction. Several genetic risk loci for inflammatory bowel disease related to oxidative stress exist, and according to available research, oxidative stress is significantly involved in the development of colorectal cancer [5]. Another non-modifiable risk factor for colorectal cancer is the presence of polyps. Histological progression from polyp to cancer is the result of a series and/or accumulation of genetic changes. Over time, these mutations can cause loss of function of the TP53 gene, which is a master regulator of transcription and apoptosis, affecting a wide range of cellular functions and ultimately leading to carcinogenesis [4].
One of the most important directions of research in the fight against cancer diseases is chemoprevention. It involves the introduction of external factors, such as medications or supplements, to stop or delay the onset, progression, or recurrence of cancer. Single compounds or extracts from plants have been proven to interfere with a specific stage of the carcinogenic process. The US Food and Drug Administration (FDA) has approved several agents for clinical use for cancer prevention [6,7]. Currently, research attention is especially focused on intracellular signalling cascades as common molecular targets for various chemopreventive phytochemicals. In the case of a well-known mechanism of development of a given cancer, chemoprevention can be used at a specific stage of development and for a specific biochemical pathway. In patients at risk for colorectal cancer, the most important processes that require chemoprevention are oxidative stress, inflammation, and apoptosis disorders. Substances of plant origin are currently one of the most intensively researched groups of chemical substances in terms of their chemopreventive effects, and the most important direction in the chemoprevention of colorectal cancer is the search for new antioxidant and anti-inflammatory substances with pro-apoptotic effects.
Berberis vulgaris L. (Berberidaceae) is a medicinal plant of the genus Berberis with more than 3000 years of tradition in folk medicine – with other roots of the genus Berberis is listed in the Chinese Pharmacopoeia under the name “Sankezhen” [8], Ayurvedic medicine also uses the root and bark of Berberis vulgaris L. The species was also widely used in traditional medicine in Europe due to its beneficial effects on cardiovascular and liver diseases [9], also anti-inflammatory effect of the raw material was traditionally appreciated [10]. Despite its well-established traditional use for various inflammatory conditions, scientific research reveals little about the anti-inflammatory activity of barberry root and it focuses on the content of secondary metabolites with anti-inflammatory activity rather than the assessment of the root extract itself [11,12]. The anti-inflammatory effect of barberry is confirmed in research, but mainly in the range of action of fruit extracts [13,14,15,16,17].
Berberis vulgaris L. is currently appreciated in the Middle East as a medicinal and culinary plant. Whereas in the Western world, it is forgotten after it was devastated for economic and agricultural reasons in the twentieth century. Due to the intensification of scientific reports on the health-promoting effects of berberine, which is currently being intensively studied there is an increasing interest in plant raw materials in which it is present. Berberis vulgaris L. itself is currently becoming increasingly popular among scientists looking for new sources of chemopreventive and chemotherapeutic agents due to its anticancer effects [18,19,20,21,22], and some health benefits of the use of Berberis vulgaris L. have already been even assessed clinically [23].
Berberis vulgaris is a typically alkaloid plant material and the root of Berberis vulgaris is the most pharmacologically active part of the plant. Its pharmacological effect is probably related to its alkaloid composition. In the root of Berberis vulgaris isoquinoline alkaloids were determined: berberrubine, berlamibine, columbamine, lambertine, magnoflorine, acanthine, jathrorrhizine, palmatine, berbamine, bargustanine, and berberine [10]. However, the broader chemical composition of this part of the plant is still poorly understood.
In this study, the chemopreventive and antiproliferative potential of BVR against colorectal cancer was assessed for the first time. The antioxidant, anti-lipoxygenase as well as proapoptotic activity of BVR was evaluated against human colon cancer cell lines. Furthermore, the influence of BVR on the expression of pro-apoptotic genes was examined for the first time. Simultaneously, the chemical composition of BVR was determined to confirm and clarify the influence of the qualitative and quantitative composition of the metabolites on the biological activity of this extract.
2. Results
2.1. Biological Activity of BVR
2.1.1. Antiradical and Antilipooxigenase Activity of BVR
The study was carried out using three methods, each of which demonstrated the antioxidant character of BVR. DPPH• analysis showed that BVR is characterised by scavenging ability at the level of 63.93±0.006mg/g expressed as Trolox equivalents (mg Trolox / g of dry extract). The ORAC results revealed the oxygen radical absorbance capacity (ORAC) for BVR at a level of 220.29±0.02mg/g expressed as Trolox equivalents (mg Trolox / g of dry extract). ABTS•+ results showed that the antiradical capacity (ABTS•+) of BVR was at the level of 122.92±0.01 mg/g expressed as Trolox equivalents (mg Trolox / g of dry extract). To investigate the anti-inflammatory properties of BVR, an experiment was carried out with one of the enzymes involved in the development of inflammation – lipooxygenase (LOX). Therefore, the direct ability of BVR to LOX activity was tested. Table 1 shows the percentage of inhibition of LOX for concentrations of the 1 mg of dry extract/ml reaction mixture.
2.1.2. Influence of BVR and its Main Constituents on the Viability of CCD841 CoN, LS180, and HT-29 Cells Using the MTT Assay
The influence of BVR and its biologically active constituents: berberine, palmatine, and berbamine on the viability of both the human colon epithelial cell line CCD841 CoN, as well as human colon adenocarcinoma cell lines LS180 and HT-29, was examined using an MTT assay. Studies were carried out after 48 hours of treatment. As presented in Figure 1, BVR significantly decreased the metabolic activity of normal and cancer cells, and the observed effect was dose-dependent. Based on the results of the MTT test, the IC50 of BVR values were determined for individual cell lines: CCD841 CoN = 50.21 µg/mL, LS180 = 4.31 µg/mL, HT-29 = 46.06 µg/mL. Simultaneously, IC50 doses of berberine, palmatine and berbamine were calculated based on the results of the MTT assay and shown in Table 2.
In the next step, the pro-apoptotic properties of BVR were examined in the human colon adenocarcinoma cell lines LS180 and HT-29 using the commercial cell death detection ELISA kit. Studies were carried out after 48 hours of cell incubation with BVR at IC50 values specific for the investigated cells (IC50 LS180 = 4.31 µg/mL; IC50 HT-29 = 46.06 µg/mL). As presented in Figure 2, BVR at indicated concentration induced programmed cell death only in HT-29 cells, where the number of nucleosomes reached 549.0% of the control. In contrast, BVR at the tested concentration was unable to induce apoptosis in LS180 cells.
2.1.3. PCR Analysis
The analysis of gene expression considered five genes associated with apoptosis – BCL2 (B-cell CLL/lymphoma 2), BCL2L1 (BCL2 Like 1), BCL2L2 (BCL2 Like 2), CASP3 (caspase 3) and CASP9 (caspase 9) were carried out in human colon cancer cells exposed to the IC50 dose of BVR.
As presented in Figure 3 and Figure 4, BVR altered the expresssion og investigated genes in both cancer cell lines.
An increased level of expression of genes involved in apoptosis was observed in LS180 cells. The highest increase was determined for BCL2L1 (366.66%) and the expression of BCL2L2 was also elevated (277.08%). The increased expression of genes encoding caspases was observed: CASP3 (261.36%) and CASP9 (213.46%). The expression of BCL2 decreased by 52.06%(Figure 3). An increased level of expression of genes involved in apoptosis was also observed in HT-29 cells. The expression of BCL2L1 was enhanced, but only at a level of 126.84%. The expression of BCL2L2 was also elevated to the highest level of 330.12% . An increase in the expression of genes that encode caspases, CASP3 (180.36%) and CASP9 (158.08%) was also observed, but at a lower level than that recorded in LS180 cells. In HT-29 cells, BCL2 expression also decreases by 23.08% (Figure 4). The mean gene expression is shown in Table 3.
2.2. Phytochemical Profiling and Quantification of Major Specialised Metabolites
The estimated phytochemical profile of the methanolic extract of the root of B. vulgaris (BVR) is presented in Figure 5 and a description of the detected metabolites can be found in Table 4. The main components determined in this study are alkaloids with a total content of 131.89 mg equivalents of berberine per 1g of dry extract [mg BE/g BVR], specifying main metabolite– berberine (70.27mg/g BVR).
3. Discussion
In this study, we evaluated the chemopreventive activity of BVR and the antioxidant effect of the extract obtained from the root of Berberis vulgaris L. was the firt step of this evaluation. There is little available in the literature on the antioxidant activity of Berberis vulgaris L., so far, research in this area has focused on fruits or other parts of this species [28]. Gorizpa et al. 2022 determined antioxidant activity in root extract of Berberis vulgaris subsp. Asperma at the IC50 level of 89.70±0.92 μg/mL and the values for root extract of Berberis vulgaris subsp. Orientalis were at the level of 167.24±1.65 μg/mL [29]. El Khalki et al. 2018 reported an EC50 value of 69.65 μg/mL for the root ethanolic extract and 77.75 μg/mL for the root acetone extract [22]. Based on the chromatographical results obtained, it can be concluded that the antioxidant potential is caused by alkaloids with established antioxidant activity and determined in this study in BVR: berberine [30], palmatine [31], jatrorrhizine [25], magnoflorine [32]. However, probable there are other metabolites with antioxidant activity that have not yet been identified in BVR, which requires further research.
In addition to antioxidant activity, effective chemopreventive agents should also have anti-inflammatory activity. In traditional Iranian medicine, B. vulgaris is a known anti-inflammatory agent. Traditional European medicine has also used these properties in the past centuries to treat inflammation, especially diseases of the digestive system. Recent studies have shown that the main mechanisms of the anti-inflammatory effects of barberry are due to the content of berberine and include changes in the cellular immune response to Th2, Treg induction, inhibition of inflammatory cytokines (IL-1, TNF and IFN-γ), and stimulation of IL-4 and IL-10 [24]. However, the anti-inflammatory effects of individual parts of the plant have not been studied so far. In this study, the anti-lipoxygenase activity of BVR was established at the level of 62±0.87%. It is higher than the determined activity of fruit (inhibition 45.24±2.45%) but lower than the activity of stem (83.57±0.13%) and leaves (79.78±2.19%) [28]. Currently, certain alkaloids marked in BVR are known to have anti-inflammatory properties and are probably responsible for its anti-inflammatory activity. The alkaloid with the best documented anti-inflammatory effect is berberine [33], which is determined in our study at a level of 70.27 ± 0.48 mg / g BVR. New reports also mention the anti-inflammatory activity of palmatine [25] and magnoflorine [32,34], which are determined in our study. The available data indicate the need for more detailed information on the composition of barberry root extracts.
The next step of evaluation was the assssment of cytotoxic activity of BVR towards colon cancer cells. As presented in Figure 2, t BVR significantly decreased the metabolic activity of normal and cancer colon cells, and the observed effect was dose dependent. The most significant changes were observed in LS180 cells, where BVR in the concentration range (5-250 µg/mL) caused a decrease in cell viability of 62.76 and 92.90%, respectively. Surprisingly, BVR at the lowest concentration tested (5 µg/mL) reduced the viability of CCD841 CoN by 35.02%, while metabolic activity of HT-29 was lowered by 7.10%. Nevertheless, in the remaining concentration range, cancer cells were more sensitive to the effect of the extract than normal cells.
Based on the results of the MTT test, the BVR IC50 values were determined for individual cell lines. CCD841 CoN = 50.21 µg/mL, LS180 = 4.31 µg/mL, HT-29 = 46.06 µg/mL. Comparison of IC50 values revealed greater sensitivity of colon cancer cells than colon epithelial cells to BVR. Furthermore, differences in the sensitivity of colon cancer cells to proapoptotic effect of BVR were also observed. Simultaneously, the IC50 doses of berberine, palmatine, and berbamine against investigated human colon cell lines were determined based on the results of the MTT assay and the data are shown in Table 3. A comparison of IC50doses revealed that BVR is significantly more cytotoxic to colon epithelial CCD841 CoN cells than berberine and palmatine. In contrast, it turned out to be the most toxic to normal cells. Significantly lower doses of both extract and alkaloids reduced the metabolic activity of colon cancer cell lines. In the case of LS180 l cells, berbamine showed the relatively lowest cytotoxicity (IC50= 37.62 µg/mL; 61.8µM) and berberine the highest (IC50 = 0.45 µg/mL; 1.33µM). At the same time, berbamine turned out to be the most cytotoxic towards HT-29 cells with an IC50 of 8.77 µg/mL (14.4µM). There are not many studies that evaluated the cytotoxicity of BVR; most researches concern single compounds present in the root of B. vulgaris. Abd El-Wahab et al. 2013 showed that barberry root ethanolic extract inhibited the growth of breast, liver and colon cancer cell lines (MCF-7, HepG2, and Caco-2, respectively) in a time and dose-dependent manner. They also compared the effect of the extract and berberine on the proliferation of these cell lines. They described that barberry ethanolic extract was cytotoxic at concentrations ranging from 1 μg/mL up to 100 μg/mL [19]. El Khalki et al. 2018 showed that ethanolic extract of Berberis vulgaris root bark is cytotoxic towards human breast adenocarcinoma cell line (MCF-7) with the IC50 value of 3.54 μg/mL with no cytotoxicity to normal human peripheral blood mononuclear cells [22].
As regards the presence of individual compounds in the extract the main component determined in this study was berberine (70.27 mg/gBVR). Other metabolites determined in BVR are jatrorrhizine, magnoflorine palmatine, aromoline, columbamine, berbamine and oxycanthine. Our results confirm the literature data [25,26,27]. Furthemore, berberrubine, berlamibine, lambertine, acanthine, bargustanine, were also recorded in the root of Berberis vulgaris L, but the data are mainly qualitative [10]. However, due to the variation in plant material caused, for example, by soil and climatic factors, there are also publications with slightly different levels of metabolites, as well as a varied phytochemical profile. Furthermore, two glycosidic derivatives of hydroxycinnamic acid (ferulic and sinapic) were identified and quantified in the BVR (in total about 8 FAE mg/g of BVR). This confirms our preliminary investigation, which showed that the BVR is rather poor in phenolic compounds (not published data).
Among alkaloids determined in this study, berberine is currently the widest investigated compound from the extract. It was described to be cytotoxic to HT-29 after 48 hours of exposure in a dose of 52.37 ±3.45 μM and these cells occurred to be much more sensitive than other cell lines tested: Tca8113 – 218.52 μM; CNE2 – 249.18 μM; MCF-7 – 272.15 μM; HeLa 245.18 μM [35]. In the study of haematopoietic cell lines, berberine was cytotoxic in range of 80.00 μM for CCRF/CEM cells to 250 μM for HL-60/MX2 cells [36]. Anticancer activity against several different cancer lines, including HT-29 examined in this study was also proven for an alkaloid palmatine, which presence has been reported in the investigated extract [37]. In addition, berbamine determined in the investigated root extract exerts an anticancer effect by induction of apoptosis. The results of Mou et al. 2019 research conducted in normal and cancer colon cells revealed the following IC50 values: HT-29 = 14 µM and IC50 CCD18 Co = 50 µM, wchat indicates lower toxicity of berbamine against normal cells. Moreover, they described nuclear fragmentation after berbamine exposure, which indicates apoptosis induction in HT-29 cells after exposure to berbamine [38]. The anticancer activity of jatrorrhizine, which occurs in the root, stem, and bark of Berberis vulgaris L, has also been described. Zhang et al. 2014 proved that jatrorrhizine in the dose range of 100–500 µg/mL inhibited cell growth and induced apoptosis of HepG2 human hepatoma cells in a time- and concentration-dependent manner [39]. The available scientific data clearly suggested that alkaloids contained in the BVR may influence the extract cytotoxicity Nevertheless, , it seems that the final beneficial effect may be influenced by yet undetermined secondary metabolites of the root. Thus, there is a great need to continue research on the content and influence of other chemical groups on the cytotoxic and pro-apoptotic activity of the root of Berberis vulgaris L., including for example tannins or terpene derivatives.
The PCR analysis of the influence of BVR on apoptotic gene expression in colon cancer cells is also consistent with those given in the literature regarding individual alkaloids determined in BVR. They may be individually responsible for influencing the expression of the examined genes. However next to alkaloids other yet unidentified secondary metabolites may play role.
Currently, the best described compound of BVR also in the field of pro-apoptotic activity, is berberine. Described induction of several biochemical events, that is, reduction of the mitochondrial membrane potential, release of cytochrome c, Bcl2 family proteins, and activation of caspases or degradation of PARP after exposure to berberine, confirms the pro-apoptotic abilities of berberine. It is described that berberine induces apoptosis in cancer cells, mainly by upregulating pro-apoptotic genes and downregulating anti-apoptotic genes [36,40,41]. The pro-apoptotic effect of berberine has been demonstrated in colon in HCT-116 cells, where caspase 3-dependent apoptosis was demonstrated [42], while in the case of other colon cancer SW480 cells, apoptosis involving caspases 3 and 9 was proven [43]. Dai et al. 2019 described that long noncoding RNA cancer susceptibility candidate 2 (lncRNA CASC2) mediates the berberine-induced pro-apoptotic effect in colorectal cancer HT-29 cells by inhibiting Bcl-2 expression at the post-transcriptional level. Caspases 3 and 9 are also targets regulated by CASC2-regulated lncRNA that are related to berberine-induced apoptosis [44].
As mentioned above, the anticancer activity jatrorrhizine, which occurs in the root, stem, and bark of B. vulgaris has been also described. Sun et al. (2019) revealed a reduction in the levels of protein Bcl-2, procaspase-3, procaspase-8, procaspase-9, and PARP and an increase in the level of pro-apoptotic proteins BAX after exposure to jatrorrhizine in MDA-MB-231, MCF-7 and 4T1 cell lines [46]. Wang et al. (2019) examined the effect of jatrorrhizine treatment on HCT-116 and HT-29 cells and also reported down-regulation of Procaspase-9 in HT-29 cells but increased in HCT-116 cells. Further examination showed a slight reduction in procaspase-8 levels in HT-29 cells and no significant changes in HCT-116 cells. A slight reduction in procaspase-3 level was observed in HCT-116 cells without significant changes in HT-29 cells [47]. The results indicated that the mechanism underlying its anticancer effect is the induction of apoptosis through caspase, including through the induction of ROS depletion [48]; but the caspase-independent mechanism is also described [47]. The effect of jatrorrhizine on proteins in the Wnt signalling pathway has also been described, which are important regulators of cell proliferation and differentiation, whose signalling pathway is closely related to proteins that initiate apoptosis, [49] gene transcription and cell adhesion [50]. Metastasis-inhibiting activity of jatrorrhizine has also been shown through the influence on N-cadherin and E-cadherin in human HCT-116 and HT-29 colon cells [47]. Another alkaloid determined in the barberry extract with anticancer activity is palmatine. Using the example of a human skin epithelial cancer cell line A431, palmatine was shown to induce apoptosis depending on concentration and exposure time through severe DNA damage and inhibition of the activity of the Bcl-2 protein [51,52]. Wu et al. 2016 showed, also in HT-29 cells, that this compound induces early and late apoptosis, acting photodynamically [53]. Furthermore, Inbaraj et al. and Hirakawa et al. reported that palmatine can bind to DNA and destroy DNA through photooxidation, and then kill human keratinocyte line HaCaT [54,55]. Furthermore, palmatine inhibited the proliferation of human colorectal cancer cell lines by reduction of the expression of the inflammatory cytokines IL-1a, IL1-b, IL-8, granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor [56]. The columbamine also identified in the extract, inhibited the proliferation, migration and invasion of colon cancer cells line HCT-116 and increased the rate of their apoptosis. Data on the mechanism of apoptosis in the case of this compound indicate that both signal transduction and expression of key factors of the Wnt/β-catenin signaling pathway are suppressed in a dose-dependent manner [57]. Berbamine also identified in the BVR extract demonstrated pro-apoptotic activity. According to Mou et al. 2019 berbamine activated caspase-3 and 9 and increasedthe Bax/Bcl-2 ratio in the colon cancer cell line HT-29. Additionally, it triggered the development of autophagic vesicles in HT-29 cells, which was concomitant with an increase in protein levels of LC3B-I, ATG-5, ATG-12, and Beclin-1. Furthermore, in the wound healing assay, this compound decreased the migration potential of mentioned cancer cells and also blocks their MEK / ERK signaling pathway [38].
It has also been shown that magnoflorine exerts anticancer effects in gastric cancer cells by regulating autophagic cell death, apoptosis, and cell cycle arrest in the S/G2 phase. Furthermore, magnoflorine inhibited AKT and activated JNK signaling, ROS accumulation-dependent pathways that are associated with autophagy, apoptosis, and cell cycle arrest in various cancer cells [58]. It has been shown that magnoflorine increases the expression of caspase 3, responsible for apoptosis induction [32]. Some anticancer properties of other plant extracts containing magnoflorine have been also demonstrated, eg. inhibition of the expression of vascular endothelial growth factor (VEGF), expression and stimulation of the angiogenesis process in hepatocellular carcinoma by aqueous extract of the Coptidis rhizome [39], inhibition of the development of cervical cancer cells of the HeLa line, and hepatocellular carcinoma cells of HepG2 and brain tumor line U251 by methanolic extract of Magnolia grandiflora leaves [59], also Ziziphus jujuba fruit extract, containing magnoflorine, shows cytotoxic effect by inhibiting the proliferation of human breast cancer cell line MCF-7, human liver cancer cell line HepG2 and human colorectal adenocarcinoma cell line HT-29 [60].
Among the alkaloids determined in this study, oxycanthin was previously presented as a compound with no impact on viability of different cancer cell lines [82]. Literature data mentioned above indicate that alkaloids whose presence in BVR was confirmed in this study are largely responsible for the pro-apoptotic effects of Berberis vulgaris L. and the results of biological activity of BVR are strongly supported by literature data on their anticancer activity.
At the same time, the quantitative content of compounds determined in this study allows us to conclude that when used in the extract, they have a more beneficial effect than when used separately in higher concentrations. This shows the advantage of a potential plant drug and the possibility of lowering the drug dose, which is one of the goals of phytopharmacology.
Furthermore, recent studies also show that, in addition to apoptosis, the isoquinoline alkaloids determined in BVR are promising in targeting angiogenesis and metastasis in colon cancer [88,91], what further broadens the prospects for the use of BVR as a new phytopharmaceutical drug. Subsequently, berberine and palmatine are also proven radiosensitizing agents, what also expands the possibilities of BVR being developed as a new phytopharmaceutical anticancer drug or adjuvant in cancer therapy.
4. Materials and Methods
4.1. Plant Material and Preparation of Extract
The root of Berberis vulgaris L. was obtained from Botanical Garden of the Maria Curie-Skłodowska University of Lublin in September 2020 (the voucher specimen AO2020091 is deposited at the Department of Pharmaceutical Botany). The material was washed and dried in the shade with ventilation. Subsequently, roots were ground in an electric mill to a homogeneous size and sieved through a 0.5 mm sieve. The powdered plant material was vacuum packed and stored at -30 °C until the extraction.
2 g of powdered root was extracted with the Dionex ASE 150 accelerated solvent extractor (Sunnyvale, CA, USA) using 80% MeOH (v/v), at 80 ° C and 1500 psi (solvent pressure), conducting three repeated extraction cycles. Subsequently, the barberry root extract was evaporated to dryness under reduced pressure and lyophilised in a FreeZone 1 apparatus (Labconco, Kansas City, KS, USA). The estimated extraction efficiency was 10.29%.
For phytochemical analyses, the freeze dried extract was re-dissolved in 80% MeOH using an ultrasonic bath and stored at -20 °C until tests.
For bioassays tests, a 50 mg / ml stock solution was prepared with DMSO and it was stored at -20 ° C until tests. Working solutions of BVR were prepared prior to analysis by diluting with culture medium, the final concentration of DMSO in test samples was <0.05% (v / v), and its effects were determined in experiments.
4.2. Chemicals and Reagents
A 2,2-diphenyl-1-picrylhydrazyl (DPPH•), 2,20-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS•+), Folin–Ciocalteu reagent, 2,20 -azobis (2-methylpropionamide) dihydrochloride (AAPH), Trolox, acetonitrile and formic acid (both MS grade), berberine (95% purity) and ferulic acid were purchased from Sigma–Aldrich (Stenheim, Germany). Ascorbic acid was purchased from Stanlab (Poland), and analytical grade methanol and aluminium chloride hexahydrate were purchased from POCH (Gliwice, Poland). Ultrapure water was prepared with a Milli-Q purification system (Millipore, Burlington, MA, USA).
The SuperScript Vilo cDNA Synthesis kit was used according to the manufacturer’s recommendation. cDNA was stored at -20 ° C until subsequent analysis. PCR reactions were performed using SG onTaq qPCR Master Mix (Eurx, Poland) on the QuantStudio 5 Real Time PCR System (Applied Biosystems).
4.3. Antiradical Activity Analyses
4.3.1. Determination of Antiradical Potential with the DPPH• Assay
The assay was carried out according to Brand-Williams et al. [61], with some modifications [62]. Absorbance was measured after 60 min at 517 nm using an Infinite Pro 200F microplate reader (Tecan Group). The results were expressed as milligrams of Trolox equivalents per gramme of dry extract [mg TE/g].
4.3.2. Determination of Antiradical Capacity with the ABTS•+ Assay
The assay was carried out according to Pellegrini et al. [63], with some modifications [64]. The absorbance was measured at 734 nm after a 6 min incubation. The level (%) was calculated as follows: [(Abscontrol – Abssample)/ Abscontrol] × 100. The results were expressed as milligrams of Trolox equivalents per gramme of dry extract [mg TE/g].
4.3.3. Oxygen Radical Absorbance Capacity (ORAC) Assay
4.4. Lipoxygenase (LOX) Inhibitor Screening Assay
Anti-lipoxygenase activity of BVR was determined according to Baraniak and Szymanowska [66]. The absorbance at 234 nm was measured immediately. The absorbance inhibition (%) was calculated as follows: [(Abscontrol − Abssample)/ Abscontrol] × 100.
4.5. Cell Cultures
The human colon epithelial cell line CCD841 CoN was purchased from the American Type Culture Collection (ATCC, Menassas, VA, USA). Human colon adenocarcinoma cell lines LS180 and HT-29 were obtained from the European Collection of Cell Cultures (ECACC, Centre for Applied Microbiology and Research, Salisbury, UK). CCD841 CoN cells were grown in Dulbecco’s modified Eagle’s Medium (DMEM). Both LS180 and HT-29 cells were grown in Dulbecco’s modified eagle medium / nutrient mix F-12 Ham (DMEM/F12). All cell culture mediums were supplemented with 10% foetal bovine serum (FBS), penicillin (100 U / ml), and streptomycin (100 μg/mL). The cells were maintained in a humidified atmosphere of 95% air and 5% CO2 at 37 ° C.
4.6. Cell Viability Assessment – MTT Assay
The cells were seeded on 96-well microplates at a density of 5 × 104 cells / ml. The next day, the culture medium was removed and cells were exposed to BVR at concentrations of 5, 50, 100 and 250 μg/mL. After 48 hours of incubation, under standard conditions (5% CO2, 37 °C), the MTT solution (5 mg / ml in PBS) was added to the cells for 3 h. The resulting crystals were solubilised overnight in SDS buffer pH 7.4 (10% SDS in 0.01 N HCl), and the product was quantified spectrophotometrically by measuring the absorbance at 570 nm wavelength using the microplate reader (BioTek ELx800, Highland Park, Winooski, Vermont, USA). The results were presented as a percentage of the metabolic activity of cells treated with the extract versus cells grown in the control medium (indicated as 100%).
4.7. Cell Death Assessment - ELISA
The cells were seeded on 96-well microplates at a density of 5 × 104 cells / ml. The next day, the culture medium was removed and cells were exposed to BVR at concentrations equal to the IC50 values calculated on the basis of the results of the MTT assay performed on the cell lines. After 48 hours of treatment, cell death was assessed using the Cell Death Detection ELISAPLUS kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer’s instructions. Absorbance was measured at a wavelength of 405 nm using BioTek ELx800 microplate reader. Results were presented as a percentage of cells underwent apoptosis in response to the investigated compound versus the number of apoptotic cells presented in control – untreated cells (indicated as 100%).
4.8. Gene Expression Analysis
Cells were seeded in 6-well plates at a density of 5 × 104 cells / ml. The next day, the culture medium was removed and cells were exposed to BVR at concentrations equal to the IC50 values calculated on the basis of the results of the MTT assay performed on the cell lines. After 48 hours of treatment, cells were washed with ice cold PBS and harvested. Then, the total RNA was extracted using a HighPure RNA Isolation Kit (Roche Diagnostics GmbH) according to the manufacturer’s instruction. The amount and purity of isolated nucleic acids were determined spectrophotometrically using NanoDrop (Thermo Scientific).
One µg of total RNA was reverse transcripted using the SuperScript Vilo cDNA Synthesis kit according to the manufacturer’s recommendation. Subsequently, the cDNA was stored at -20 ° C until later analysis. Before polymerase chain reaction (PCR), cDNA was diluted 20 times and reactions were performed using SG onTaq qPCR Master Mix (Eurx, Poland) in the QuantStudio 5 real-time PCR system (Applied Biosystems) under the thermal cycling conditions given in the mix manual, in 40 amplification cycles and with an annealing/elongation step at 60 ° C for 1 min. The melt curve analysis was performed after each reaction plate. All samples were evaluated in triplicate. The expression of the following genes has been examined: BCL2 apoptosis regulator (BCL2), BCL2 like 1 (BCL2L1), BCL2 like 2 (BCL2L2), caspase 3 (CASP3) and caspase 9 (CASP9). Both the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and hypoxanthine phosphoribosyltransferase 1 (HPRT1) genes were used for expression normalisation. All primers, except those previously used for GAPDH [67], were designed by the Primer-BLAST online tool [68]. Detailed characterisation of primers is presented in the Supplementary Table S1. The specificity of the PCR products was evaluated in 2% agarose gel electrophoresis. Data were collected and analysed using the comparative Ct method (ΔΔCt method; QuantStudio Design and Analysis Software v1.5.2, Applied Biosystems). Expression results were presented as a relative quantification (RQ).
4.9. Phytochemical Profiling and Quantification of Major Specialised Metabolites Using LC-UV- MS/MS Technique
High-resolution MS analysis of BVR extract was performed using a Bruker Impact II HD (Bruker, Billerica, MA, USA) quadrupole time-of-flight mass spectrometer (Q-TOF-MS) coupled with a Ultimate 3000 RS chromatographic system (Thermo Fischer Scientific, Waltham, MS, USA). The sample was chromatographed on a Cortecs T3 column (2.1 × 150 mm, 2.7 μm, Waters, Milford, MA, USA) equipped with a precolumn. A 25 min linear gradient (5→50%) of acetonitrile-water mixture (both acidified with 0.1% formic acid), with a flow rate of 0.5 mL/min was applied. The column was kept at 40 °C. The injection volume was 2 µL. The MS operated in electrospray ionisation and both polarity modes, with the following settings: mass scan range 50–1200 m/z; capillary voltage 4.0 kV (ESI+) or 3.0 kV (ESI–); nebuliser and drying gas (N2) 2.0 bar and 10 L/min, respectively; dry gas temperature 220 ° C. The MS/MS spectra were registered using a collision energy of 35 eV with stepping 50% and 125% of CE. The acquired data was calibrated internally with sodium formate (10 mM solution in 50% 2-propanol) which was injected into the ion source before the sample analysis. The MS acquisition was accompanied by a charged aerosol detector (CAD, Corona Veo RS, Thermo) that collected data at a frequency of 10 Hz. Data were processed using DataAnalysis 4.4 software (Bruker).
Quantitative analysis of alkaloids and phenolic acids was carried out using a UPLC-PDA system (ACQUITY class, Waters, Milford, MA, USA).
Chromatographic conditions were as follows: BEH C18 column (2.1 × 100 mm, 1.7 μm, Waters) maintained at 45 °C, a linear gradient of 28 min (5→40%) of methanol-water mixture (both acidified with 0.1% formic acid), the flow rate of 0.25 ml / min and a 2.5 µL of injection volume.
Alkaloids were detected based on UV 270 nm (3.6 nm resolution) and a calibration curve for berberine (y = 882.3 x - 287.85; R2 = 0.9995; linear range 0.5–30 µg/mL; LOD = 58.85 ng/ml; LOQ = 176.55 ng/ml). Quantitative results were expressed as milligrams of berberine equivalents (BE) per gramme of extract (BVR).
Quantitative analysis of the content of phenolic acid derivatives was performed using system and chromatographic conditions as previously described, although UV 320 nm detection and a calibration curve for ferulic acid (y = 2716.55x – 67.37; R2 = 0.9999; range 0.5–19.8 µg/mL; LOD = 18.65 ng/mL; LOQ = 55.95 ng/mL) were used. Quantitative results were expressed as milligrams of ferulic acid equivalents (FAE) per gram of extract (BVR).
Data were collected and processed using MassLynx 4.1 software (Waters).
4.10. Statistical Analysis
Phytochemical experiments were carried out in triplicate at a minimum and the results are expressed as means ± standard deviations (SD) or standard errors (SEM). Differences between two independent groups were compared using the Student’s t test and the comparison between more than two groups was performed using one-way ANOVA, followed by Dunnett’s comparison test. Data were analysed using Statistica 13.0 (Statsoft Inc, Tulsa, OK, USA) and Microsoft Excel software. The IC50 value (concentration leading to 50% inhibition of cell viability compared to the control) was calculated using GraphPad PRISM.
5. Conclusions
The root of Berberis vulgaris L. is a raw material with a high chemopreventive potential to prevent the development of colorectal cancer. The extract is noteworthy because its action is closely related to the impact on processes related to the occurrence of non modifiable risk factors for colorectal cancer (inflammatory bowel disease, polyps), which, combined with the modification of modifiable factors (lifestyle, diet, obesity), can significantly overall reduce the risk of developing colon cancer. Due to its antioxidant, anti-inflammatory, and antiproliferative effects, it can be treated as a comprehensive chemopreventive agent in patients at risk of developing colorectal cancer. It concerns especially those suffering from inflammatory bowel disease or polyps, which are non modifiable risk factors for colorectal cancer, characterized by inflammation, increased oxidative stress, and impaired apoptosis. BVR is cytotoxic to colorectal cancer cells but has less cytotoxicity to normal cells. The extract increases the expression of genes related to apoptosis of colorectal cancer cells, inhibiting tumor proliferation and development.
Barberry root is the raw material is rich in alkaloids with proven anticancer effectiveness. The quantitative content of these compounds determined in this study allows us to conclude that they act in BVR at much lower doses than in the case of the independent action of a single compound, what can provide an opportunity to reduce the dose of the simple drug by using BVR. Synergistic action or the influence of not determined metabolites are the probable reason of BVR activity. However as shown in this study, barberry root has an appropriate quantity of pharmacologically active alkaloids responsible for pro-apoptotic activity. Nonetheless further research to better understand the chemical composition of BVR seems to be very important.
An additional advantage is the safety of using the raw material due to the centuries-old tradition of use, which is why it can be treated as a dietary supplement or a traditional medicine - the French Pharmacopoeia contains a monograph on the bark of the root of Berberis vulgaris L.
Supplementary Materials
Table S1
Author Contributions
Conceptualization, A.O. and R.N.; methodology, A.O.,R.N.,M.L.,D.J. and M.C.; software, A.O.,R.N.,M.L. D.J. and M.C.; validation, A.O.,R.N.,M.L.,D.J. and M.C..; formal analysis, A.O.,R.N.,M.L.,D.J.,A.K.,S.P. and M.C.; investigation, A.O.,R.N.,M.L.,D.J. and M.C.; resources, A.O.,R.N.,M.L. D.J.,A.K.,S.P and M.C.; data curation, A.O.,R.N.,M.L. D.J.,A.K.,S.P and M.C.; writing—original draft preparation, A.O.; writing— A.O. and R.N.; visualization, A.O.,M.L.,D.J. and M.C.; supervision, A.O. and R.N.; project administration, A.O. and R.N.; funding acquisition, R.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- World Cancer Research Fund International. Available online: https://www.wcrf-uk.org/. (accessed on 12 Mar 2023).
- Labianca, R.; Nordlinger, B.; Beretta, G.D.; Mosconi, S.; Mandalà, M.; Cervantes, A.; Arnold, D. Early colon cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24, vi64–vi72. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer. (accessed on 19 January 2023).
- Simon, K.; Balchen, V. Colorectal cancer development and advances in screening. Clin. Interv. Aging 2016, 11, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Bardelčíková, A.; Šoltys, J.; Mojžiš, J. Oxidative Stress, Inflammation and Colorectal Cancer: An Overview. Antioxidants 2023, 12, 901. [Google Scholar] [CrossRef]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Maresso, K.C.; Tsai, K.Y.; Brown, P.H.; Szabo, E.; Lippman, S.; Hawk, E.T. Molecular cancer prevention: Current status and future directions. CA: A Cancer J. Clin. 2015, 65, 345–383. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yan-ze, L.; Yong, P.E.; Zhong-zhi, Q.I; . Pei-gen, X.I. New collection of crude drugs in Chinese pharmacopoeia 2010 II. Sankezhen (Berberis spp.). Chin. Herbal Med. 2011, 3, 272–88. [Google Scholar]
- Henriette’s Herbal Homepage. Available online: https://www.henriettes-herb.com. 2020.
- Imanshahidi, M.; Hosseinzadeh, H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytotherapy Res. 2008, 22, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, P.; Chai, M.-J.; Yang, F.; Li, H.-S.; Zhao, J.; Wang, H.; Lu, D.-D. [Effects of berberine on serum inflammatory factors and carotid atherosclerotic plaques in patients with acute cerebral ischemic stroke]. China J. Chin. Mater. Medica 2016, 41, 4066–4071. [Google Scholar] [CrossRef]
- Meng, S.; Wang, L.; Huang, Z.; Zhou, Q.; Sun, Y.; Cao, J.; Li, Y.; Wang, C. Berberine ameliorates inflammation in patients with acute coronary syndrome following percutaneous coronary intervention. Clin. Exp. Pharmacol. Physiol. 2012, 39, 406–411. [Google Scholar] [CrossRef]
- Kiasalari, Z.; Khalili, M.; Ahmadi, P. Effect of alcoholic extract of berberis vulgaris fruit on acute and chronic inflammation in male rats. Journal of Babol University of Medical Sciences 2011, 13(1).
- Minaiyan, M.; Ghannadi, A.; Mahzouni, P.; Jaffari-Shirazi, E. Comparative Study of Berberis vulgaris Fruit Extract and Berberine Chloride Effects on Acetic Acid-Induced Colitis in Rats. 2011, 10, 97–104.
- Majeed, W.; Aslam, B.; Javed, I.; Khaliq, T.; Muhammad, F.; Ali, A.; Raza, A. Histopathological evaluation of gastro protective effect of Berberis vulgaris (Zereshk) seeds against aspirin induced ulcer in albino mice. . 2015, 28, 1953–8. [Google Scholar]
- Mohebali, S.; Nasri, S.; Kamalinejhad, M.; Noori, A.S. Antinociceptive & anti-inflammatory effects of Berberis vulgaris L. root’s hydroalcoholic extract and determination of it’s possible antinociceptive mechanism in male mice. Journal of Paramedical Sciences (JPS) 2011, 2(4), 12-18.
- Ivanovska, N.; Philipov, S. Study on the anti-inflammatory action of Berberis vulgaris root extract, alkaloid fractions and pure alkaloids. Int. J. Immunopharmacol. 1996, 18, 553–561. [Google Scholar] [CrossRef] [PubMed]
- NIH. Clinical Trials.gov. Available online: https://clinicaltrials.gov/ (accessed on 26 April 2023).
- El-Wahab, A.E.A.; A Ghareeb, D.; Sarhan, E.E.; Abu-Serie, M.M.; A El Demellawy, M. In vitro biological assessment of berberis vulgaris and its active constituent, berberine: antioxidants, anti-acetylcholinesterase, anti-diabetic and anticancer effects. BMC Complement. Altern. Med. 2013, 13, 218–218. [Google Scholar] [CrossRef] [PubMed]
- Gird, C.E.; Ligiaelena, D.U. ; Costea,T. ; Nencu,I.; Popescu, M.; Balaci, T.; Olaru, O. Research regarding obtaining herbal extracts with antitumor activity. note ii. phytochemical analysis, antioxidant activity and cytotoxic effects of Chelidonium majus L., Medicago sativa L. and Berberis vulgaris L. dry extracts. Medicago sativa L. and Berberis vulgaris L. dry extracts. Farmacia 2017, 65, 703–708. [Google Scholar]
- Ghafourian, E.; Sadeghifard, N.; Pakzad, I.; Valizadeh, N.; Maleki, A.; Jafari, F.; Ghiasvand, N.; Abdi, J.; Shokoohinia, Y.; Ghafourian, S. Ethanolic Extract of Berberis Vulgaris Fruits Inhibits the Proliferation of MCF-7 Breast Cancer Cell Line Through Induction of Apoptosis. Infect. Disord. - Drug Targets 2017, 17, 192–198. [Google Scholar] [CrossRef] [PubMed]
- El Khalki, L.; Tilaoui, M.; Jaafari, A.; Mouse, H.A.; Zyad, A. Studies on the Dual Cytotoxicity and Antioxidant Properties of Berberis vulgaris Extracts and Its Main Constituent Berberine. Adv. Pharmacol. Sci. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Ng, L.T.; Hsu, F.-F.; Shieh, D.-E.; Chiang, L.-C. Cytotoxic effects of Coptis chinensis and Epimedium sagittatum extracts and their major constituents (berberine, coptisine and icariin) on hepatoma and leukaemia cell growth. Clin. Exp. Pharmacol. Physiol. 2004, 31, 65–69. [Google Scholar] [CrossRef]
- Kalmarzi, R.N.; Naleini, S.N.; Ashtary-Larky, D.; Peluso, I.; Jouybari, L.; Rafi, A.; Ghorat, F.; Heidari, N.; Sharifian, F.; Mardaneh, J.; et al. Anti-Inflammatory and Immunomodulatory Effects of Barberry (Berberis vulgaris) and Its Main Compounds. Oxidative Med. Cell. Longev. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Villinski, J.; Dumas, E.; Chai, H.B.; Pezzuto, J.; Angerhofer, C.; Gafner, S. Antibacterial activity and alkaloid content of Berberis thunbergii, Berberis vulgaris and Hydrastis canadensis. Pharm. Biol. 2003, 41, 551–557. [Google Scholar] [CrossRef]
- Saeidnia, S.; Gohari, A.; Kurepaz-Mahmoodabadi, M.; Mokhber-Dezfuli, N. Phytochemistry and Pharmacology of Berberis Species. Pharmacogn. Rev. 2014, 8, 8–15. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Kaushik, N. Phytochemical and pharmacological studies in genus Berberis. Phytochem. Rev. 2012, 11, 523–542. [Google Scholar] [CrossRef]
- Och, A.; Olech, M.; Bąk, K.; Kanak, S.; Cwener, A.; Cieśla, M.; Nowak, R. Evaluation of the Antioxidant and Anti-Lipoxygenase Activity of Berberis vulgaris L. Leaves, Fruits, and Stem and Their LC MS/MS Polyphenolic Profile. Antioxidants 2023, 12, 1467. [Google Scholar] [CrossRef] [PubMed]
- Gorizpa, M.; Bahmanyar, F.; Mirmoghtadaie, L.; Shafaei, F. Evaluation of Antioxidant and Antimicrobial Properties of Root and Stem Bark Extracts of Three Species of Barberry in Bread. Research and Innovation in Food Science and Technology 2022, 10(4), 413–426. [Google Scholar] [CrossRef]
- Luo, A.; Fan, Y. Antioxidant activities of berberine hydrochloride. J Med Plants Res 2011, 5(16), 3702–7. [Google Scholar]
- Chaves, S.K.M.; Afzal, M.I.; Islam, M.T.; Hameed, A.; Da Mata, A.M.O.F.; Araújo, L.D.S.; Ali, S.W.; Rolim, H.M.L.; Medeiros, M.d.G.F.D.; Costa, E.V.; et al. Palmatine antioxidant and anti-acetylcholinesterase activities: A pre-clinical assessment. Cell. Mol. Biol. 2020, 66, 54–59. [Google Scholar] [CrossRef]
- Xu, T.; Kuang, T.; Du, H.; Li, Q.; Feng, T.; Zhang, Y.; Fan, G. Magnoflorine: A review of its pharmacology, pharmacokinetics and toxicity. Pharmacol. Res. 2020, 152, 104632. [Google Scholar] [CrossRef] [PubMed]
- Sarraf, M.; Babaei, A.B.; Naji-Tabasi, S. Investigating functional properties of barberry species: an overview. J. Sci. Food Agric. 2019, 99, 5255–5269. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Jiang, K.; Wu, H.; Yang, C.; Zhao, G.; Deng, G. Magnoflorine ameliorates lipopolysaccharide-induced acute lung injury via suppressing NF-κB and MAPK activation. Frontiers in pharmacology 2018, 30, 9:393441. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, H.; Chen, W.; Huang, P. Berberine induces apoptosis and arrests the cell cycle in multiple cancer cell lines. Arch. Med Sci. 2021, 19, 1530–1537. [Google Scholar] [CrossRef] [PubMed]
- Och, A.; Zalewski, D.; Komsta. ; Kołodziej, P.; Kocki, J.; Bogucka-Kocka, A. Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines. Toxins 2019, 11, 485. [Google Scholar] [CrossRef]
- Bala, M.; Pratap, K.; Verma, P.K.; Singh, B.; Padwad, Y. Validation of ethnomedicinal potential of Tinospora cordifolia for anticancer and immunomodulatory activities and quantification of bioactive molecules by HPTLC. J. Ethnopharmacol. 2015, 175, 131–137. [Google Scholar] [CrossRef]
- Mou, L.; Liang, B.; Liu, G.; Jiang, J.; Liu, J.; Zhou, B.; Huang, J.; Zang, N.; Liao, Y.; Ye, L.; Liang, H. ; Berbamine exerts anticancer effects on human colon cancer cells via induction of autophagy and apoptosis.; inhibition of cell migration and MEK/ERK signalling pathway. J BUON 2019, 24(5), 1870-1875.
- Zhang, L.-L.; Ma, L.-N.; Yan, D.; Zhang, C.-E.; Gao, D.; Xiong, Y.; Sheng, F.-Y.; Dong, X.-P.; Xiao, X.-H. Dynamic monitoring of the cytotoxic effects of protoberberine alkaloids from Rhizoma Coptidis on HepG2 cells using the xCELLigence system. Chin. J. Nat. Med. 2014, 12, 428–435. [Google Scholar] [CrossRef]
- Kalaiarasi, A.; Anusha, C.; Sankar, R.; Rajasekaran, S.; John Marshal, J.; Muthusamy, K.; Ravikumar, V. Plant Isoquinoline Alkaloid Berberine Exhibits Chromatin Remodeling by Modulation of Histone Deacetylase To Induce Growth Arrest and Apoptosis in the A549 Cell Line. J. Agric. Food Chem. 2016, 64, 9542–9550. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-H.; Tang, W.-C.; Cheng, Y.-W.; Sia, P.; Huang, C.-C.; Lee, Y.-C.; Jiang, H.-Y.; Wu, M.-H.; Lai, I.-L.; Lee, J.-W.; et al. Targeting of multiple oncogenic signaling pathways by Hsp90 inhibitor alone or in combination with berberine for treatment of colorectal cancer. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2015, 1853, 2261–2272. [Google Scholar] [CrossRef]
- He, B.; Wu, K.; Yang, Q.; Mu, Y.; Zhou, L.; Liu, Y.; Zhou, Q. Berberine inhibits the proliferation of colon cancer cells by inactivating Wnt/β-catenin signaling. Int. J. Oncol. 2012, 41, 292–298. [Google Scholar] [CrossRef]
- Murthy, K.N.C.; Jayaprakasha, G.K.; Patil, B.S. The natural alkaloid berberine targets multiple pathways to induce cell death in cultured human colon cancer cells. Eur. J. Pharmacol. 2012, 688, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Mu, L.; Cui, Y.; Li, Y.; Chen, P.; Xie, H.; Wang, X. Berberine Promotes Apoptosis of Colorectal Cancer via Regulation of the Long Non-Coding RNA (lncRNA) Cancer Susceptibility Candidate 2 (CASC2)/AU-Binding Factor 1 (AUF1)/B-Cell CLL/Lymphoma 2 (Bcl-2) Axis. J. Pharmacol. Exp. Ther. 2019, 25, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Vuddanda, P.R.; Chakraborty, S.; Singh, S. Berberine: a potential phytochemical with multispectrum therapeutic activities. Expert Opin. Investig. Drugs 2010, 19, 1297–1307. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, X.; Wu, P.; Wink, M.; Li, J.; Dian, L.; Liang, Z. Jatrorrhizine inhibits mammary carcinoma cells by targeting TNIK mediated Wnt/β-catenin signalling and epithelial-mesenchymal transition (EMT). Phytomedicine 2019, 63, 153015. [Google Scholar] [CrossRef]
- Wang, P.; Gao, X.-Y.; Yang, S.-Q.; Sun, Z.-X.; Dian, L.-L.; Qasim, M.; Phyo, A.T.; Liang, Z.-S.; Sun, Y.-F. Jatrorrhizine inhibits colorectal carcinoma proliferation and metastasis through Wnt/β-catenin signaling pathway and epithelial–mesenchymal transition. Drug Des. Dev. Ther. 2019, ume 13, 2235–2247. [Google Scholar] [CrossRef]
- Luo, T.; Zhang, H.; Zhang, W.W.; Huang, J.T.; Song, E.L.; Chen, S.G.; He, F.; Xu, J.; Wang, H.Q. Neuroprotective effect of Jatrorrhizine on hydrogen peroxide-induced cell injury and its potential mechanisms in PC12 cells. Neuroscience Letters 2011, 498, 227–231. [CrossRef]
- Kumari, S.; Kaladhar, D.; Solmon, K.S.; Malla, R.; Kishore, G. Anti-proliferative and metastatic protease inhibitory activities of protoberberines: An in silico and in vitro approaches. Process. Biochem. 2013, 48, 1565–1571. [Google Scholar] [CrossRef]
- Rolle, J.; O Asante, D.; Kok-Fong, L.L.; Boucetta, H.; A Seidu, T.; Tai, L.L.K.; Alolga, R.N. Jatrorrhizine: a review of its pharmacological effects. J. Pharm. Pharmacol. 2021, 73, 709–719. [Google Scholar] [CrossRef]
- Ali, D.; Ali, H. Assessment of DNA damage and cytotoxicity of palmatine on human skin epithelial carcinoma cells. Toxicol. Environ. Chem. 2014, 96, 941–950. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Zhang, X.F.; Tang, Y.L.; Xiang, J.F.; Tian, M.Y. Studies of the interactions between three protoberberine alkaloids and Bcl-2 by fluorescence spectroscopy. Acta Chimica Sinica 2011, 69(02), 247. [Google Scholar]
- Wu, J.; Xiao, O.; Zhang, N. ; Xue. , C.; Leung, A.W.; Zhang, H.; Xu, C.; Tang, Q. Photodynamic action of palmatine hydrochlo-ride on colon adenocarcinoma HT-29 cells. Photodiagnosis and Photodynamic Therapy 2016, 15, 53–58. [Google Scholar] [CrossRef]
- Inbaraj, J.J.; Kukielczak, B.M.; Bilski, P.; He, Y.-Y.; Sik, R.H.; Chignell, C.F. Photochemistry and Photocytotoxicity of Alkaloids from Goldenseal (Hydrastis canadensisL.). 2. Palmatine, Hydrastine, Canadine, and Hydrastinine. Chem. Res. Toxicol. 2006, 19, 739–744. [Google Scholar] [CrossRef]
- Hirakawa, K.; Kawanishi, S.; Hirano, T. The Mechanism of Guanine Specific Photooxidation in the Presence of Berberine and Palmatine: Activation of Photosensitized Singlet Oxygen Generation through DNA-Binding Interaction. Chem. Res. Toxicol. 2005, 18, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.-K.; Li, H.; Dong, C.-L.; He, X.; Guo, C.-R.; Zhang, C.-F.; Yu, C.-H.; Wang, C.-Z.; Yuan, C.-S. Palmatine from Mahonia bealei attenuates gut tumorigenesis in ApcMin/+ mice via inhibition of inflammatory cytokines. Mol. Med. Rep. 2016, 14, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yao, Y.; Shen, B.; Liu, J.; Pan, Q.; Liu, N.; Li, L.; Huang, J.; Long, Z.; Shao, L. Columbamine suppresses the proliferation and malignization of colon cancer cells via abolishing Wnt/β-catenin signaling pathway. Cancer Manag. Res. 2019, ume 11, 8635–8645. [Google Scholar] [CrossRef]
- Sun, X.-L.; Zhang, X.-W.; Zhai, H.-J.; Zhang, D.; Ma, S.-Y. Magnoflorine inhibits human gastric cancer progression by inducing autophagy, apoptosis and cell cycle arrest by JNK activation regulated by ROS. Biomed. Pharmacother. 2020, 125, 109118. [Google Scholar] [CrossRef]
- Mohamed, S.; Hassan, E.M.; Ibrahim, N.; Mohamed, S.; Hassan, E.M.; Ibrahim, N. Cytotoxic and antiviral activities of aporphine alkaloids of Magnolia grandiflora L. Nat. Prod. Res. 2010, 24, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Zhang, H.; Liu, Q.; Zhao, Y.; Cui, X.; Guo, S.; Zhang, L.; Ho, C.-T.; Bai, N. Chemical characterization of the main bioactive constituents from fruits of Ziziphus jujuba. Food Funct. 2016, 7, 2870–2877. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Olech, M.; Łyko, L.; Nowak, R. Influence of Accelerated Solvent Extraction Conditions on the LC-ESI-MS/MS Polyphenolic Profile, Triterpenoid Content, and Antioxidant and Anti-lipoxygenase Activity of Rhododendron luteum Sweet Leaves. Antioxidants 2020, 9, 822. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pieczykolan, A.; Pietrzak, W.; Nowak, R.; Pielczyk, J.; Łamacz, K. Optimization of Extraction Conditions for Determination of Tiliroside in Tilia L. Flowers Using an LC-ESI-MS/MS Method. J. Anal. Methods Chem. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Baraniak, B.; Szymanowska, U. Lipooxygenase in food of plant origin. Żywność Nauka Technologia Jakość 2006, 2, 29–45. [Google Scholar]
- Podgórski, R.; Cieśla, M.; Podgórska, D.; Bajorek, W.; Płonka, A.; Czarny, W.; Trybulski, R.; Król, P. Plasma microRNA-320a as a Potential Biomarker of Physiological Changes during Training in Professional Volleyball Players. J. Clin. Med. 2022, 11, 263. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134–134. [Google Scholar] [CrossRef]
Figure 1.
The effect of the BVR on the viability of human colon epithelial cell line CCD841 CoN and the human colon adenocarcinoma cell lines LS180 and HT-29. The results are presented as mean ± SEM (n = 5). Data were analysed using one-way ANOVA followed by post hoc Dunnett’s test: ** p < 0.01; *** p < 0.001 versus control.
Figure 1.
The effect of the BVR on the viability of human colon epithelial cell line CCD841 CoN and the human colon adenocarcinoma cell lines LS180 and HT-29. The results are presented as mean ± SEM (n = 5). Data were analysed using one-way ANOVA followed by post hoc Dunnett’s test: ** p < 0.01; *** p < 0.001 versus control.
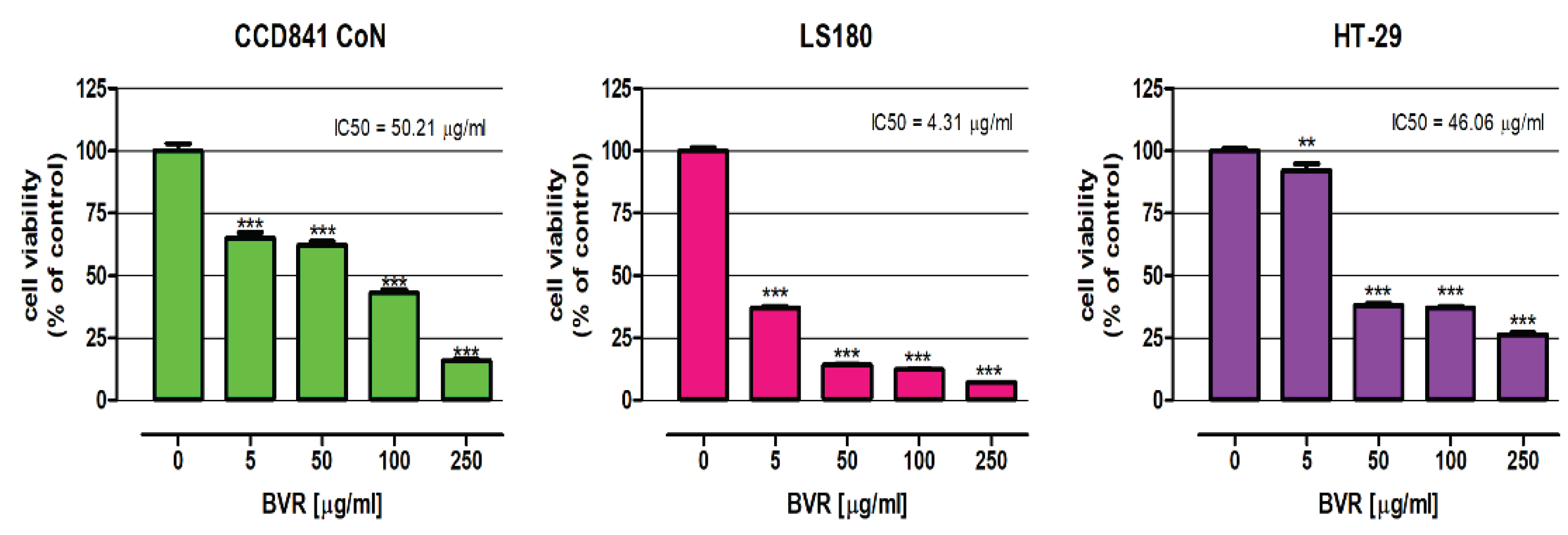
Figure 2.
Pro-apoptotic effect of BVR in human colon adenocarcinoma cell lines LS180 and HT-29. In test samples, cells were exposed to BVR at the IC50 level (4.31 and 46.06 µg/mL for LS180 and HT-29 cells, respectively). The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): *** p < 0.001.
Figure 2.
Pro-apoptotic effect of BVR in human colon adenocarcinoma cell lines LS180 and HT-29. In test samples, cells were exposed to BVR at the IC50 level (4.31 and 46.06 µg/mL for LS180 and HT-29 cells, respectively). The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): *** p < 0.001.
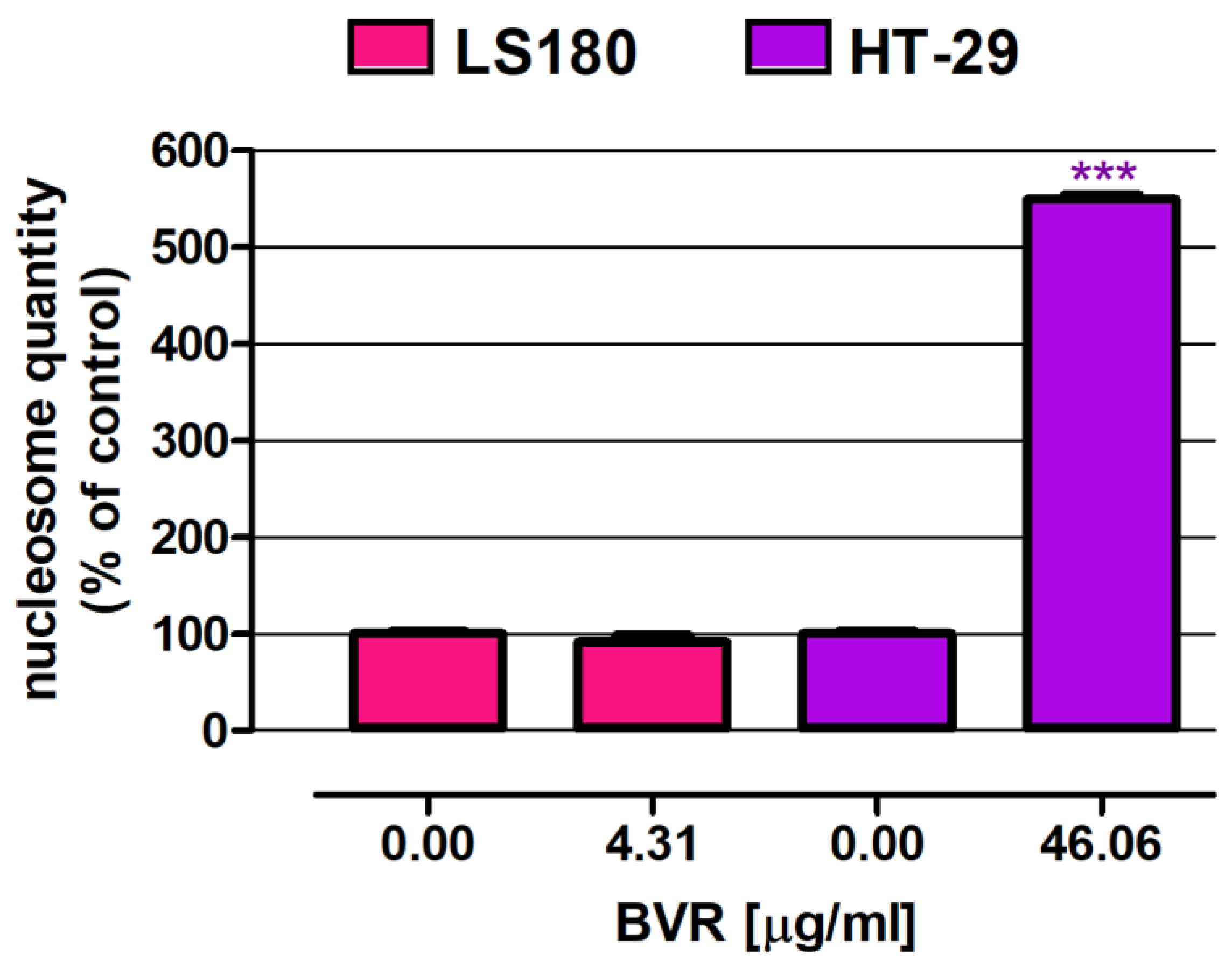
Figure 3.
Differences in relative expression of five apoptotic genes (CASP3, CASP9, BCL2, BCL2L1, and BCL2L2) between LS180 cells treated with BVR and controls. The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 3.
Differences in relative expression of five apoptotic genes (CASP3, CASP9, BCL2, BCL2L1, and BCL2L2) between LS180 cells treated with BVR and controls. The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): * p < 0.05, ** p < 0.01, *** p < 0.001.
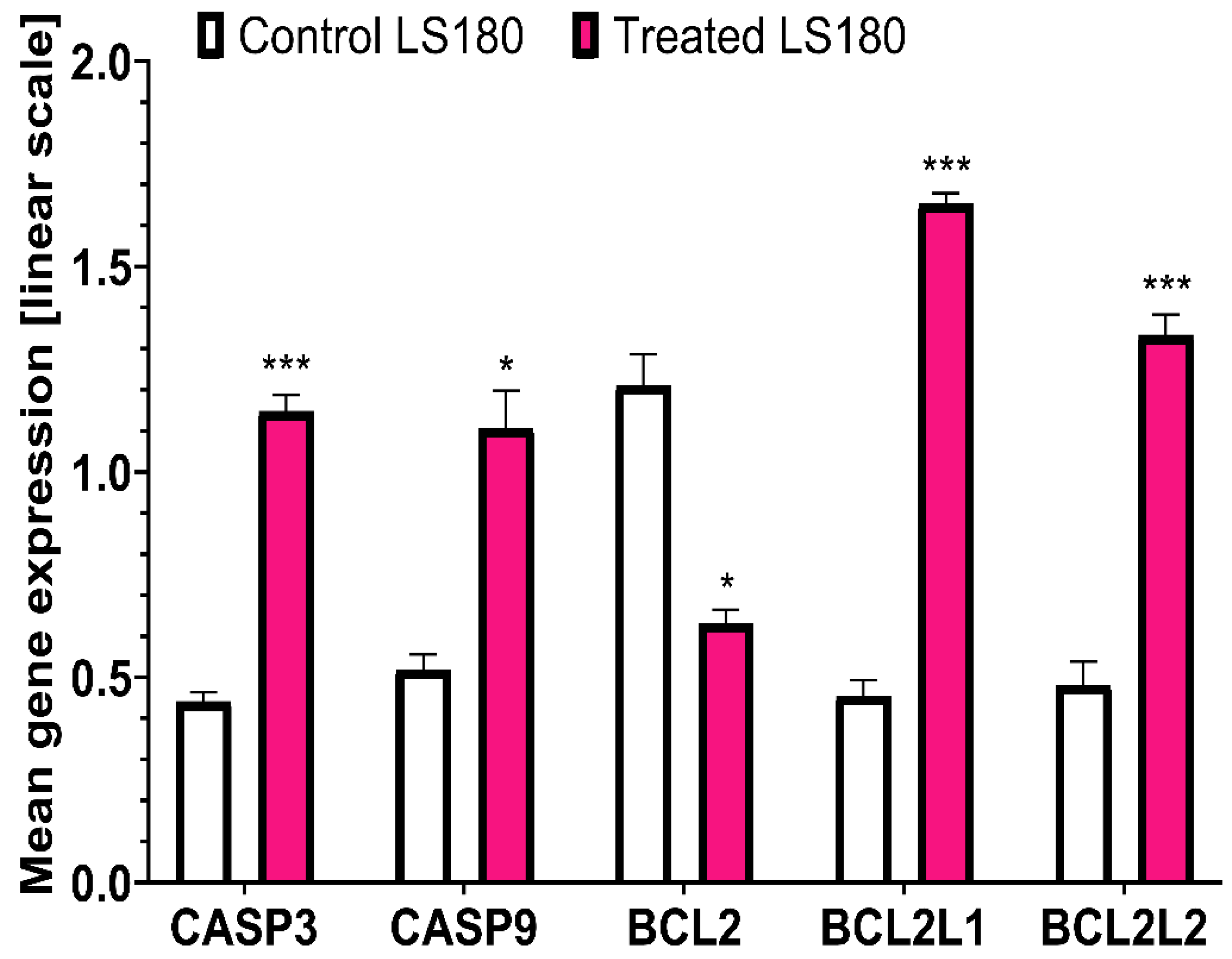
Figure 4.
Differences in relative expression of five apoptotic genes (CASP3, CASP9, BCL2, BCL2L1, and BCL2L2) between HT-29 cells treated with BVR and controls. The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): * p < 0.05, ** p < 0.01.
Figure 4.
Differences in relative expression of five apoptotic genes (CASP3, CASP9, BCL2, BCL2L1, and BCL2L2) between HT-29 cells treated with BVR and controls. The results are presented as mean ± SEM (n = 3). Data were analysed using Student’s t-test (control vs. treated): * p < 0.05, ** p < 0.01.
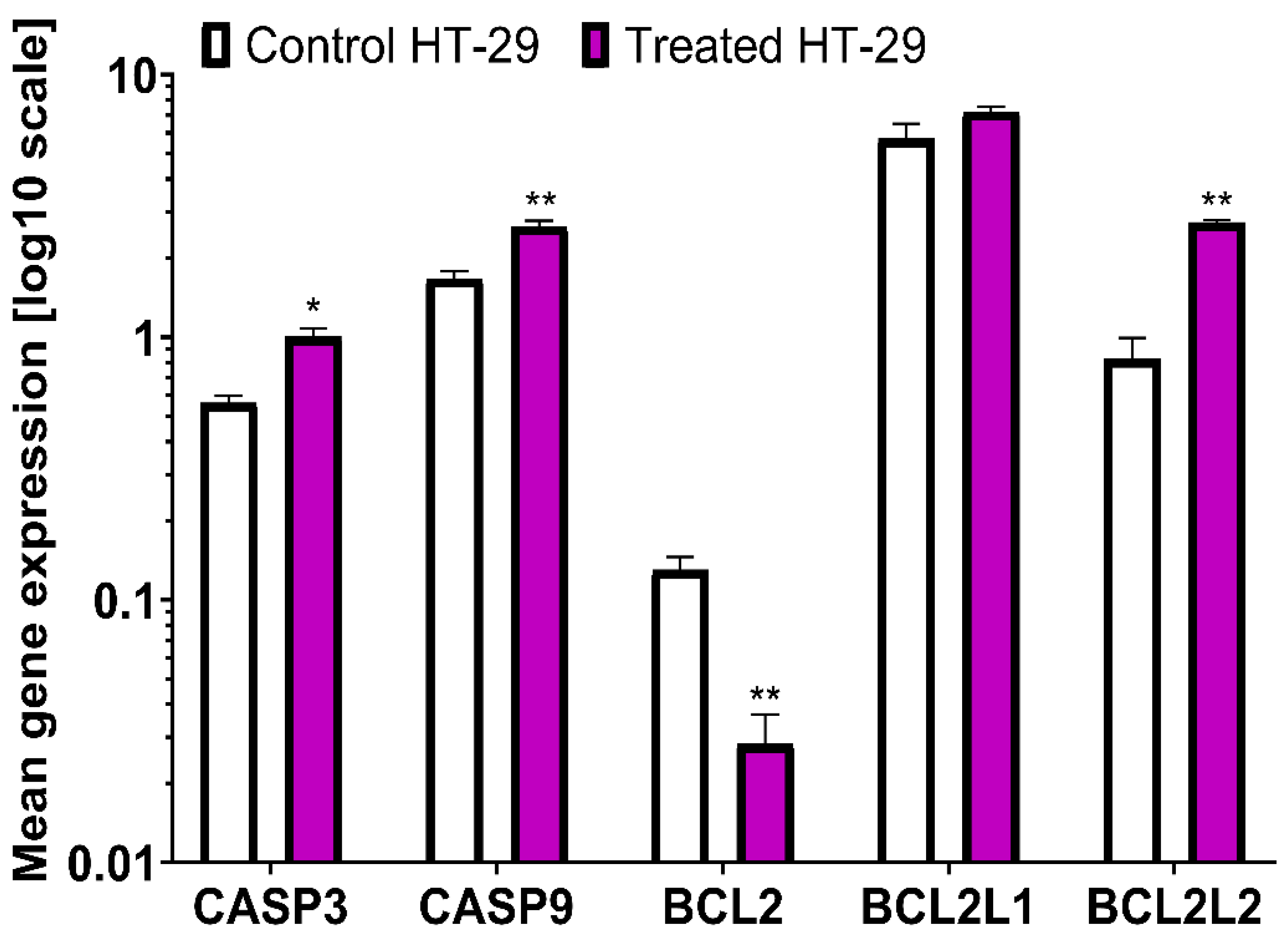
Figure 5.
UHPLC-CAD-QTOF-MS analysischromatograms of BVR. The peak numbers correspond to Table 3.
Figure 5.
UHPLC-CAD-QTOF-MS analysischromatograms of BVR. The peak numbers correspond to Table 3.
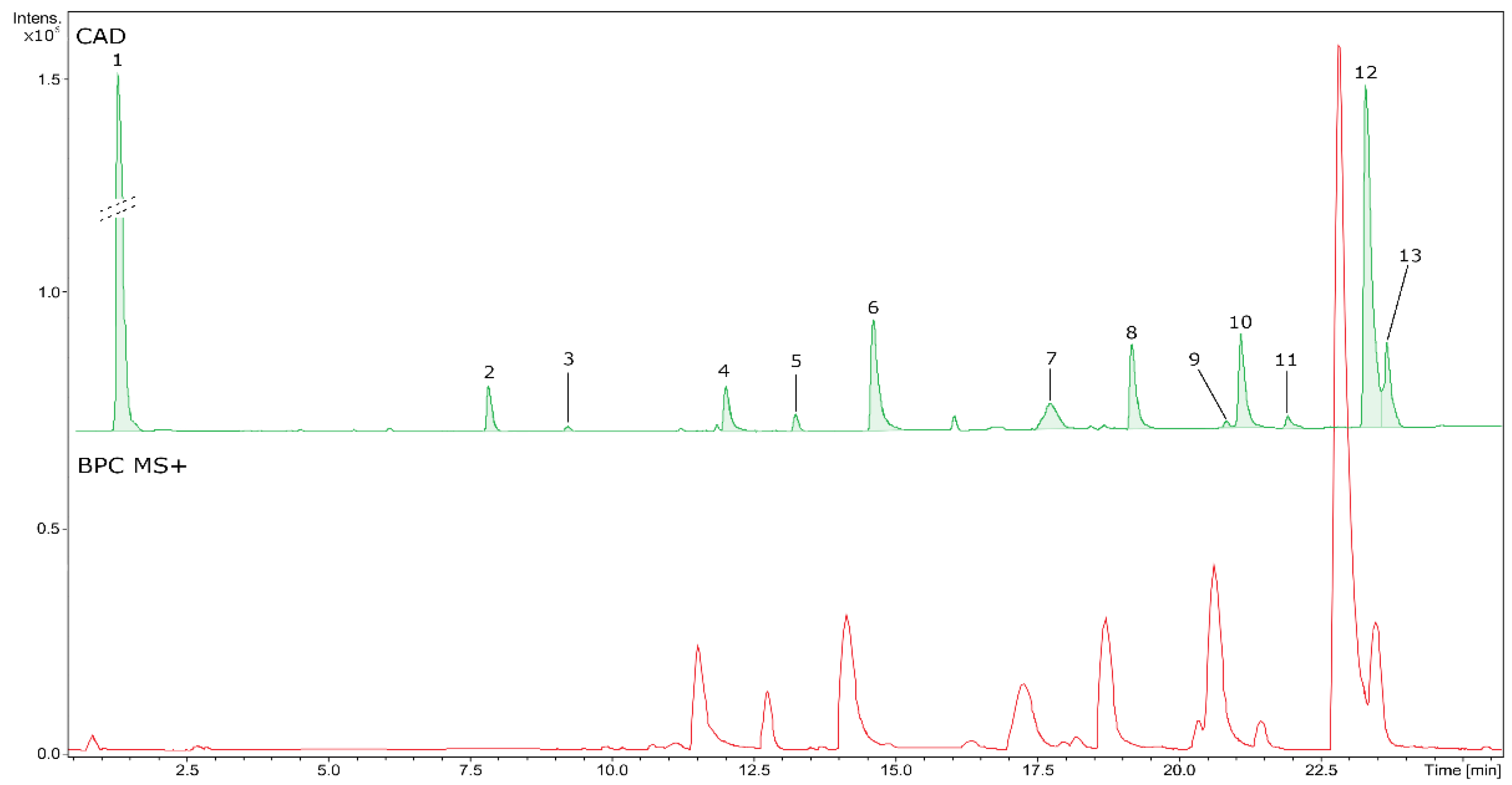

Table 1.
Antioxidant and anti-lipoxygenase activity of BVR. Antiradical effects were determined using ABTS•+, DPPH• and ORAC assays, and the results are expressed Trolox equivalents (mg Trolox / g of dry extract). The results are expressed as % of lipoxygenase inhibition. Data are presented as means ± standard deviations of triplicate measurements.
Table 1.
Antioxidant and anti-lipoxygenase activity of BVR. Antiradical effects were determined using ABTS•+, DPPH• and ORAC assays, and the results are expressed Trolox equivalents (mg Trolox / g of dry extract). The results are expressed as % of lipoxygenase inhibition. Data are presented as means ± standard deviations of triplicate measurements.
| ABTS•+ [mgTE/g] |
ORAC [mgTE/g] |
DPPH• [mgTE/g] |
LOX inhibition [%] |
|---|---|---|---|
| 122.92±0.01 | 220.29±0.02 | 63.93±0.01 | 62.60±0.87 |
ABTS•+: 2,2′-azinobis-3-ethylbenzthiazoline-6-sulfonic acid; ORAC: Oxygen Radical Absorbance Capacity; DPPH•: 2,2-diphenyl-1-picrylhydrazyl radical; LOX: lipoxygenase.
Table 2.
The IC50 values of BVR and berberine, palmatine, and berbamine estimated using the MTT assay. IC50 data are presented as means ± standard deviations of triplicate measurements.
Table 2.
The IC50 values of BVR and berberine, palmatine, and berbamine estimated using the MTT assay. IC50 data are presented as means ± standard deviations of triplicate measurements.
| CCD841 CoN | LS180 | HT-29 | |
| µg/mL | µg/mL | µg/mL | |
| BVR | 50.21 ± 1.22 | 4.31 ± 1.19 | 46.06 ± 1.11 |
| Berberine | 254.8 ± 2.05 | 0.45 ± 1.25 | 15.92 ± 1.08 |
| Palmatine | 179.8 ± 2.24 | 12.92 ± 1.10 | 29.34 ± 1.08 |
| Berbamine | 13.63 ± 1.02 | 37.62 ± 1.02 | 8.77 ± 1.07 |
Table 3.
Expression of genes associated with apoptosis in LS180 and HT29 cells exposed to IC50 levels of BVR (4.31 and 46.06 µg/mL for LS180 and HT-29 cells, respectively). Values are expressed as means RQ ± standard deviations.
Table 3.
Expression of genes associated with apoptosis in LS180 and HT29 cells exposed to IC50 levels of BVR (4.31 and 46.06 µg/mL for LS180 and HT-29 cells, respectively). Values are expressed as means RQ ± standard deviations.
| Gene | LS180 | HT29 | |||||
|---|---|---|---|---|---|---|---|
| Control | Exposed | Change of expression (%) | Control | Exposed | Change of expression (%) | ||
| CASP3 | 0.44±0.04 | 1.15±0.07 | 261.36 ↑ | 0.56±0.04 | 1.01±0.08 | 180.36 ↑ | |
| CASP9 | 0.52±0.06 | 1.11±0.16 | 213.46 ↑ | 1.67±0.12 | 2.64±0.14 | 158.08 ↑ | |
| BCL2 | 1.21±0.13 | 0.63±0.06 | 52.06 ↓ | 0.13±0.02 | 0.03±0.01 | 23.08 ↓ | |
| BCL2L1 | 0.45±0.07 | 1.65±0.05 | 366.66 ↑ | 5.7±0.81 | 7.23±0.31 | 126.84 ↑ | |
| BCL2L2 | 0.48±0.1 | 1.33±0.09 | 277.08 ↑ | 0.83±0.17 | 2.74±0.06 | 330.12 ↑ | |
Table 4.
Phytochemicals detected and quantified in BVR based on UHPLC-UV-CAD-MS/MS analysis. Compound numbers correspond to Figure 5.
Table 4.
Phytochemicals detected and quantified in BVR based on UHPLC-UV-CAD-MS/MS analysis. Compound numbers correspond to Figure 5.
| No | RT (min) | Formula | Error (ppm) | Measured m/z | MS/MS fragments | CAD area (%) | Identity | Content [mg eq/g of dry BVR] ± SD |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.81 | - | 0.0 | 341.1089a | 179.0580,119.0366 | 54.62 | - | uic |
| 2 | 7.28 | C16H19O9 | -0.4 | 355.1036a | 193.0514, 78.0270, 134.0379 | 1.81 | Feruloyl-hexoside | 7.10d ± 0.03 |
| 3 | 8.67 | C17H22O10 | -1.3 | 385.1145a | 223.0624, 79.0699, 164.0471 | 0.22 | Sinapoyl-hexoside | 0.66d ± 0.00 |
| 4 | 11.46 | C20H24NO4 | 1.1 | 342.1695b | 265.0854, 97.1117, 282.0882, 65.0694 | 1.89 | Magnoflorine* | 11.63e ± 0.12 |
| 5 | 12.69 | C19H24NO3 | 0.4 | 314.1749b | 269.1169,209.0957, 237.0907, 165.0697 | 0.64 | Unidentified alkaloid | < LLOQ |
| 6 | 14.06 | C36H39N2O6 | 0.6 | 595.2799b | 595.2798,564.2381, 552.2376, 367.1648 | 6.04 | Aromoline | 7.11e ± 0.22 |
| 7 | 17.16 | C37H40N2O6 | 0.9 | 609.2954b | 609.2955,578.2539, 566.2538, 381.1804 | 2.46 | Berbamine* | 3.19e ± 0.04 |
| 8 | 18.61 | C37H40N2O6 | 0.4 | 609.2957b | 609.2955,381.1807, 174.0913, 578.2532 | 3.85 | Oxycanthine | 5.48e ± 0.10 |
| 9 | 20.27 | C20H20NO4 | -0.2 | 338.1381b | 322.1077,308.0919, 294.1126, 236.0709 | 0.44 | Columbamine | 3.32e ± 0.08 |
| 10 | 20.53 | C20H20NO4 | -0.5 | 338.1389b | 322.1079,308.0920, 294.1128, 236.0708 | 4.4 | Jatrorrhizine* | 23.32e ± 0.11 |
| 11 | 21.35 | C38H43N2O6 | 0.9 | 623.3110b | 623.3108,381.1810, 174.0912, 592.2681 | 0.22 | Rodiasine | < LLOQ |
| 12 | 22.73 | C20H18NO4 | 0.3 | 336.1229b | 320.0916,292.0966, 278.0809, 306.0760 | 19.34 | Berberine* | 70.27 ± 0.48 |
| 13 | 22.97 | C21H22NO4 | 1.4 | 352.1537b | 336.1226,278.0807, 322.1073, 308.1278 | 4.07 | Palmatine* | 7.57e ± 0.30 |
a adduct [M-H]–, b adduct [M+H]+, c ui – unidetyfied, d ferulic acid equivalents (FAE), e berberine equivalents (BE) in mg BE per g BVR [mg BE/g BVR], * – the identity of metabolite was confirmed with the standards.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



