
Submitted:
29 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
Walnut trees are grown worldwide for their edible fruits of high nutritional value. To address climate change, researchers have studied walnut phenology to create cultivars adapted to warmer climates. The standardisation of walnut phenological observations is needed and the Biologische Bundesanstalt, Bundessortenamt und CHemische Industrie (BBCH) scale is the most accurate and internationally accepted for this purpose. Here, the principal growth stages (PGS) of Persian walnut (Juglans regia L.) are described using stages from a previously available alphanumerical scale. This standardised phenological scale describes Persian walnut growth from the dormant vegetative state through reproductive budding to senescence. This phenological scale is expected to increase the efficiency of walnut phenological monitoring. Fifty-seven stages were used to describe the life cycle of Persian walnut in this BBCH scale. Of these 57 stages, 3 stages are dedicated to seed germination (PGS-0), 4 stages to bud development (PGS-0), 7 stages to leaf development (PGS-1), 4 stages to stem elongation (PGS-3), 8 stages to inflorescence emergence (PGS-5), 5 stages to male flowering (PGS-6), 5 stages to female flowering (PGS-6), 5 stages to fruit development (PGS-7), 12 stages to fruit ripening (PGS-8) and 4 stages to leaf senescence (PGS-9).
Keywords:
English walnut
; phenology
; phenophase
; growth stages
; climate change
; global warming
1. Introduction
Persian walnut (Juglans regia L.), also known as English walnut, is a widespread tree of the Juglandaceae family. Juglandaceae comprises more than 50 species in 11 genera such as Carya (hickories including pecans), Pterocarya (wingnuts) and Juglans (walnuts) [1,2,3]. The Juglans genus includes more than 20 species, all of which are diploid (2n = 2x = 32), such as J. cinerea (white walnut), J. nigra (black walnut) and J. regia (Persian walnut) [4,5,6]. Persian walnut is disseminated worldwide in temperate regions (Europe, North and South America, South Africa, Asia, Australia and New Zealand), and was domesticated in central Asia (western Himalayas). It first spread to the west (northern Iran, the Caucasus and eastern Türkiye) and then spread to the east (northern India and western China) [7,8]. Persian walnut was present in refugia in the Balkans and western Europe during the last glacial period [9,10] and human-mediated dispersal is thought to have occurred in the Early Bronze Age.
Persian walnut is a wind-pollinated, deciduous, monoecious and dichogamous tree; its dichogamy limits self-fertilisation [2]. Usually, more than one hundred flowers form the male catkin, whereas two or three flowers form the female inflorescence.
The study of the changes in the timing of seasonal events such as plant flowering is called phenology [11]. There is a scientific consensus that phenology is dependent on environmental conditions and impacted by climate change, as is the case for most deciduous tree species [12,13]. In Persian walnut, Charrier et al. [14] have shown that the winter dormancy from September through January in the northern hemisphere is mainly under environmental control. Climate change leads to phenological shifts that disrupt the whole growth cycle of the trees. During the spring, a warming climate increases the fulfilment of heat requirements and promotes earlier flowering and leaf unfolding [15,16], making trees more susceptible to late frost. However, during the winter, a warming climate counteracts phenological advancement because it decreases fulfilment of chilling requirements [17,18]. Fu et al. [19] have shown that higher temperatures also delay autumn leaf senescence, and the consequence of the phenological shift is the amplification of drought stress because of the extended growing season [20].
In Persian walnut, these phenological shifts have been observed in Slovenia [21] and in Romania [22]; moreover Luedeling [23] has confirmed using partial least square regression that the phenology of Persian walnut in California is advanced by high temperatures in spring and delayed by high temperatures in winter. Therefore, the phenology of dormancy in Persian walnut is impacted by both spring and winter temperatures, which makes its response to climate change difficult to model [24,25,26]. For instance, fruit winter chill modelling approaches are numerous and may perform poorly, so that the quantification of chilling requirements may be imprecise [23,27].
Differences in chilling and heat requirements are observable between genotypes of a species: genotypes with low chilling requirements can flower earlier and may suffer frost damages, whereas genotypes with high chilling requirements flower later and may suffer insufficient chilling fulfilment, leading to abnormal growth and harvest loss [28,29,30,31]. In Persian walnut, a strong variability in chilling and heat requirement traits is observed in orchards and within herbarium accessions. In 2019 in France, Bernard et al. [32] observed a 71-day gap in bud break date between the earliest and the latest accession in the INRAE (Institut National de la Recherche Agronomique et Environnementale) walnut germplasm collection. In California, the germplasm collection of the Walnut Improvement Program of the University of California-Davis showed a 44-day gap in the leafing date in 2018-2019 [33]. This research has led to the identification of SNP-phenological trait associations [33]. It is crucial to create a precise and globally accepted scale of phenological observations in Persian walnut to ensure an efficient characterisation of genotypes whose data could be reused to feed models and statistical analyses and to better manage orchards.
Based on a decimal code used in plant breeding by Zadoks et al. [34], the Biologische Bundesanstalt, Bundessortenamt und CHemische Industrie (BBCH) scale was first developed for cereal crops to help standardisation [35]. Later, Hack et al. [36] developed an extended BBCH scale for other species such as pome and stone fruits. BBCH scales are already available for other nut species, such as almond [37], cashew [38], chestnut [39], hazelnut [40,41] and pecan [42]. However, a BBCH scale for Persian walnut is still missing. For Persian walnuts, a BBCH codification for flowering, fruit development and ripening stages only was recently presented during the IX International Symposium on Walnut and Pecan organized by the International Society of Horticultural Science (ISHS) [43].
To describe Persian walnut phenological events, Germain et al. [44] have released the first scale that provides the basis for the International Plant Genetic Resources Institute (IPGRI) descriptors of walnut phenology-related traits [45]. This scale has been used by breeders and genetic resource curators across the world to characterise Persian walnut genotypes; for instance, the IPGRI scale has been used in Iran and India to describe the phenological characteristics of promising genotypes [46,47,48]. Here, we propose an extended BBCH scale for Persian walnut, adapted from the scales of Germain et al. and IPGRI [44,45] and we provide pictures of the main stages useful for genotype characterisation. We expect this work to contribute to meeting findable, accessible, interoperable and reusable (FAIR) data principles and to contribute to better genetic resource management and crop practices.
2. Materials and Methods
2.1. Study Area
The phenological phases of Persian walnut were monitored in a varietal collection orchard composed of 10 cultivars (Table 1) and located in Agmé (59 m above sea level, N44.495210, E0.357202) in southwestern France in the department of Lot-et-Garonne. The trees are 11 years old. The soil type is a clay loam (20.5% sand, 41.4% silt and 38.1% clay). This soil has a pH of 8.3, an organic matter content of 1.8 g · kg-1 and a C/N ratio of 9.2.
2.2. Monitoring of Phenophases and Fruit Growth Measurements
The phenological stages of flowering and leaf development were monitored weekly from April to September on 16 trees per cultivar. These observations were repeated in 2019, 2020, 2021, 2022 and 2023.
Fruit development was monitored weekly from early May to late August 2022 on 40 fruits on trees of the ‘Lara’ cv. Fruits were randomly chosen in the orchard. On each fruit, the length (from the basal scar to the apical scar) and the width were measured with an electronic calliper (MarCal 16 EWRi digital calliper IP66). Then the fruits were weighed with a precision balance (Mettler Toledo model MS-S/MS-L). Means (n = 40) are presented with their standard deviations. For lignification, the method described by Paradinas et al. [40] was used.
2.3. BBCH Scale Characteristics
The development of Persian walnut was described using main growth stages numbered from 0 to 9 according to the extended BBCH reference scale [36]. The numerical order of the main growth stages was respected in the first digit of this scale. The second digit of this scale describes the plant or organ stage in further detail. To avoid errors during phenology monitoring, male flowering was described using a two-digit code while female flowering was described using a three-digit code. As for hazelnuts [40], once the walnut husk and the shell were fully developed, a third digit was added to describe kernel development. For fruit development, a third digit was added to the two-digit scale of fruit ripening to detail the percentage of falling fruits at maturity.
3. Results
3.1. Principal Growth Stage 0: Seed Germination
The principal growth stage 0 for seed germination is subdivided into 3 developmental stages and presented with three-digit codes to avoid ambiguity between seed germination and bud development, which is expressed with a two-digit code.
Stage 000, which corresponds to stage 89 of fruit ripening in this scale, represents the dry seed. When the radicle emerges from the seed, stage 050 is reached. Finally, stage 090 represents the beginning of root growth when the rootlet elongates, absorbent hairs appear and secondary root development begins, and the shoot emerges from the seed and pierces the soil surface.
3.2. Principal Growth Stage 0: Bud Development
The principal growth stage 0 for bud development is subdivided into 4 developmental stages. Bud development stage 00, which was ‘Af’ in the Germain scale is reached when buds are dormant and covered by scales. When the hard scales of the first order fall and buds are still enveloped by poorly differentiated semi-membranous scales, stage 03 is reached, historically described as stage Af2. Once the bud swells (stage Bf), the outer envelopes loosen and the ends of the underlying bracts covered with whitish down appear. This is the so-called ‘woolly’ or ‘white bud’ stage described in this scale as stage 07. The final stage, initially described as stage Cf, represents bud elongation, when the extremity of the outermost leaves can be distinguished, also known as bud burst. Here, bud burst is described as stage 09.
3.3. Principal Growth Stage 1: Leaf Development
The principal growth stage 1 for leaf development is subdivided into 7 developmental stages. Historically, 3 stages were used to describe leaf development. Stage Cf2, which represents the separation of the scales and bracts and the beginning of separation of the first leaf, is here described as stage 10. When the bud is open, the first leaves are separated and their leaflets are well individualised, stage 11 is reached, initially described as stage Df. Finally, stage 12 represents the point at which the first leaves are completely unfolded and erect and have taken on an oblique habit, revealing the female flower in their centre. Four new stages are here added to the historical description. These are stage 13, which represents the point at which the first leaf is fully developed with a loss of red colour, stage 14, representing the point at which more than two leaves are fully developed with a green colour, stage 17, corresponding to the point at which all the leaves are fully expanded and have a dark-green colour and stage 19, which represents the complete development of leaves on the tree.
3.4. Principal Growth Stage 3: Stem Elongation
The principal growth stage 3 for stem elongation is subdivided into 4 developmental stages. The beginning of stem elongation is described as stage 30. This stage is reached when the stem has elongated to less than 10% of its final length. Stage 31 is reached when the stem has elongated to between 10% and 50% of its final length. When the stem has elongated to between 50% and 90% of its final length, stage 35 is reached. Finally, when the stem has elongated to more than 90% of its final length, stage 39 is reached.
Figure 1.
The phenological growth stages of Persian walnut according to the BBCH codification.

Figure 1.
The phenological growth stages of Persian walnut according to the BBCH codification.


Table 2.
Description of BBCH stages applied for Persian walnut.
| BBCH codification | Germain et al., 1999 scale | IPGRI scale | Description |
|---|---|---|---|
| Principal growth stage 0: seed germination | |||
| 000 | Dry seed | ||
| 050 | The radicle emerges from the seed | ||
| 090 | Rootlet elongation, appearance of absorbent hairs and secondary root development, and soil emergence of the beetle | ||
| Principal growth stage 0: bud development | |||
| 00 | Af | Dormant buds: scale-covered bud | |
| 03 | Af2 | Fall of the hard scales of the first order. Bud still enveloped by other poorly differentiated semi-membranous scales | |
| 07 | Bf | The bud swells: the outer envelopes loosen, and the ends of the underlying bracts covered with a whitish down appear this is the so-called "woolly" or "white bud" stage. | |
| 09 | Cf | The bud elongates; the extremity of the terminal leaves of the outermost leaves can be distinguished; it's the bud burst | |
| Principal growth stage 1: leaf development | |||
| 10 | Cf2 | The scales and bracts move apart, the first leaves begin to separate | |
| 11 | Df | The bud is open, the first leaves separate, and their leaflets are well individualised | |
| 12 | Df2 | The first leaves are completely unfolded, first erect, then they take on a more or less oblique habit, revealing the female flowers in their centre. | |
| 13 | First leaf fully developed, loss of red foliage colour | ||
| 15 | More than two leaves are fully developed with a green foliage | ||
| 17 | All the leaves are fully expanded, are growing and turn dark green | ||
| 19 | All the leaves are mature and have their final length | ||
| Principal growth stage 3: stem elongation | |||
| 30 | Starting stem elongation | ||
| 31 | <10% of final stem length | ||
| 33 | >10% and <50% of final stem length | ||
| 35 | >50% and <90% of final stem length | ||
| 39 | >90% of final stem length | ||
| Principal growth stage 5: inflorescence emergence | |||
| 50 | Amr | In early summer, the differentiated male catkin, globose in shape, has a pinkish hue while the buds remain green | |
| 51 | Amv | During the summer, the catkin grows slightly, becomes conical, reaches about 0.5 cm length, and takes on a green colour | |
| 53 | Amg | At the beginning of October, the catkin stops growing, it measures 0.5-0.8 cm and takes on a gray colour that it will keep all winter. | |
| 55 | Bm | About 3 weeks before the bud break, growth resumes, the catkin swells and lengthens to reach 1.3-2 cm length | |
| 57 | Cm | Catkin, stiff and oblique, reaches the size of a pencil and measures 3-4 cm. Its colour gradually changes from green-brown to light green; the flower clusters are distinct. | |
| 58 | Dm | Catkin loses its rigidity, becomes semi-drooping; the glomeruli separate | |
| 59 | Dm2 | The glomeruli space out and begin to open; the catkin hangs | |
| 590 | Ef | Appearance of female flowers | |
| Principal growth stage 6: flowering | |||
| 60 | Em | First male bloom date | Complete opening of the glomeruli and separation of the anthers which begin to turn yellow |
| 61 | Fm | Peak male bloom date | Beginning of anther dehiscence from base of catkin |
| 65 | Fm2 | Peak male bloom date | Complete anther dehiscence, full pollen emission |
| 67 | Gm | Last male bloom date | The anthers emptied of their pollen turn black |
| 69 | Hm | The catkin falls to the ground and dries up | |
| 610 | Ff | First female bloom date | Appearance of stigmata |
| 630 | Ff1 | Peak female bloom date | The orange-yellow stigmata are divergent. Their receptivity is optimal: it is the full female flowering. |
| 650 | FF2 | Peak female bloom date | Stigmata take on a pale green-yellow colour and are completely recurved |
| 670 | FF3 | Last female bloom date | The stigmata begin to become necrotic; these are streaked with fine brown threads. |
| 690 | Gf | Drying and blackening of the stigmata | |
| Principal growth stage 7: fruit development | |||
| 71 | Beginning of fruit-husk growth | ||
| 75 | 50% of the final size in fruit husk | ||
| 79 | 100% of the final size in husk and beginning of lignification: beginning of resistance to the knife | ||
| 790 | 100% of the shell is lignified and the beginning of kernel filling | ||
| 799 | 100% lignified shell and kernel filling completed | ||
| Principal growth stage 8: fruit ripening | |||
| 81 | White septum in kernel | ||
| 82 | Beginning of browning with some brown pitting on the septum | ||
| 83 | Brown internal septum on 1/3 of its surface | ||
| 84 | Packing tissue brown date | Brown internal septum on 3/3 of its surface | |
| 85 | In allover brown septum but damp and matt | ||
| 86 | Shiny dry brown septum | ||
| 87 | Cracking of the husk | ||
| 88 | Opening of the husk: the nut remains trapped in the husk | ||
| 89 | Husk opens enough for the nut to freely fall to the ground | ||
| 891 | Harvest date | < 10% of the nuts have fallen to the ground | |
| 895 | Harvest date | > 50% of the nuts have fallen to the ground | |
| 899 | Harvest date | 100% of the nuts have fallen to the ground | |
| Principal growth stage 9: leaf senescence | |||
| 93 | Beginning of leaf colour change | ||
| 95 | 50% of the leaves have changed colour | ||
| 97 | All the leaves have changed colour and leaf fall begins | ||
| 99 | Defoliation date | All the leaves have fallen | |
3.5. Principal Growth Stage 5: Inflorescence Emergence
The principal growth stage 5 for inflorescence emergence is subdivided into 7 developmental stages for male inflorescence emergence and 1 stage for female inflorescence emergence. These stages are an adaptation of the Germain et al. and IPGRI scale into the BBCH codes. The first stage (stage 50), initially described as stage Amr, occurs in early summer. The differentiated male catkin, globose in shape, has a pinkish hue while the buds remain green. Then, during the summer, the catkin grows slightly, becomes conical, reaches about 0.5 cm in length, takes on a green colour and reaches stage 51 (Amv). Stage 53 occurs at the beginning of October, when the catkin stops growing, measures 0.5–0.8 cm in length and takes on a grey colour that will be kept during the winter. About 3 weeks before bud break, growth resumes, the catkin swells and lengthens to reach 1.3–2 cm in length and reaches stage 55 (Bm). When the catkin remains stiff and oblique, reaches the length of a pencil and measures 3–4 cm, stage 57 is reached (Cm). Its colour gradually changes from green-brown to light green and the flower clusters become distinct. Stage 58 (Dm) is reached when the catkin loses its rigidity, becomes semi-drooping and the glomeruli separate. When the glomeruli spread out and begin to open, the catkin hangs and reaches stage 59 (Dm2).
The appearance of the first female flowers is here described as stage 590. Generally, this stage occurs just after stage 12, when the first leaf is completely unfolded. This stage was historically named Ef.
3.6. Principal Growth Stage 6: Flowering
As Persian walnut develops male and female reproductive organs on separate flowers, two codifications of developmental stages are here developed, a two-digit code for male flowering and a three-digit code for female flowering.
3.6.1. Male Flowering
The principal growth stage 6 for male flowering is subdivided into 5 developmental stages. The first male bloom, historically named Em, is described here as stage 60 and represents the complete opening of the glomeruli and separation of the anthers, which begin to turn yellow. When male blooming peaks, the dehiscence of anthers begins from the base of the catkin. This stage is stage 61 (Fm). Stage 65 represents full pollen emission and complete anther dehiscence, initially described as Fm2. When the anthers that are emptied of their pollen turn black, stage 67 is reached. Stage 67 is also known as the late male bloom date or Gm. Finally, when the catkin falls to the ground and dries up, stage 69 is reached (Hm).
3.6.2. Female Flowering
The principal growth stage 6 for female flowering is subdivided into 5 developmental stages. The appearance of stigmata, initially described as stage Ff or ‘first female bloom date,’ is described here as stage 610. When the orange-yellow stigmata are divergent, their receptivity is optimal: this stage is known as full female flowering. Full female flowering was previously described as stage Ff1 and is here described as stage 630. The next stage, here described as stage 650, occurs when stigmata take on a pale green-yellow colour and are completely recurved (Ff2). When the stigmata begin to become necrotic, they are streaked with fine brown threads and stage 670 is reached (Ff3). This stage is known as the last female bloom date. The last stage of female flowering, stage 690, occurs when the stigmata dry and blacken (Gf).
3.7. Principal Growth Stage 7: Fruit Development
The principal growth stage 7 for fruit development is subdivided into 5 developmental stages. The beginning of fruit husk growth is here described as stage 71. When the fruit husk has reached 50% of its final size, stage 75 is reached. Complete growth, that is, when the fruit husk has reached 100% of its final size and lignification begins, manifested by a low resistance to the knife when the nut is cut, is here described as stage 79. When 100% of the shell is lignified and the kernel begins to fill, stage 790 is reached. When kernel filling is complete, stage 799 is reached. Persian walnut fruit development is illustrated in Figure 2. Fruit size (length and width) and weight change over time follow similar curves. These growth curves can be divided into two phases, a first phase that represents rapid fruit husk and shell growth between BBCH stages 71 to 79 and a second phase with slower fruit growth, during which the kernel grows, between BBCH stages 790 to 799.
3.8. Principal Growth Stage 8: Fruit Ripening
The principal growth stage 8 for fruit ripening is subdivided into 12 developmental stages. The first stage, here described as stage 81, is reached when the white septum is observable. When the septum begins to brown and develops some brown pitting, stage 82 is reached. Stage 83 occurs when one-third of the internal septum is brown. When the entire surface of the internal septum is brown, stage 84 is reached. This stage is known as the septum browning date. Stage 85 describes the point at which the septum is completely brown but still damp and matte. When the septum is shiny, dry and brown, stage 86 is reached. Then, when the husk cracks, stage 87 is reached. The next stage, 88, is reached when the husk begins to open but the nuts remain trapped in the husk. When nuts fall freely to the ground, stage 89 occurs. This stage can also be described as stage 000 if the BBCH code for seed germination is used.
The harvest date is subdivided into three stages: stage 891, which represents the point at which fewer than 10% of the nuts have fallen to the ground, stage 895, the point at which more than 50% of the nuts have fallen to the ground and stage 899, the point at which 100% of the nuts have fallen to the ground.
Figure 2.
Nut growth dynamic of nut length (black line  ), width (dash line
), width (dash line  ) and weight (dash-point line
) and weight (dash-point line  ) of ‘Lara’ cultivar associated with their standard deviation of mean.
) of ‘Lara’ cultivar associated with their standard deviation of mean.
), width (dash line ) and weight (dash-point line ) of ‘Lara’ cultivar associated with their standard deviation of mean.
Figure 2.
Nut growth dynamic of nut length (black line ), width (dash line ) and weight (dash-point line ) of ‘Lara’ cultivar associated with their standard deviation of mean.
), width (dash line ) and weight (dash-point line ) of ‘Lara’ cultivar associated with their standard deviation of mean.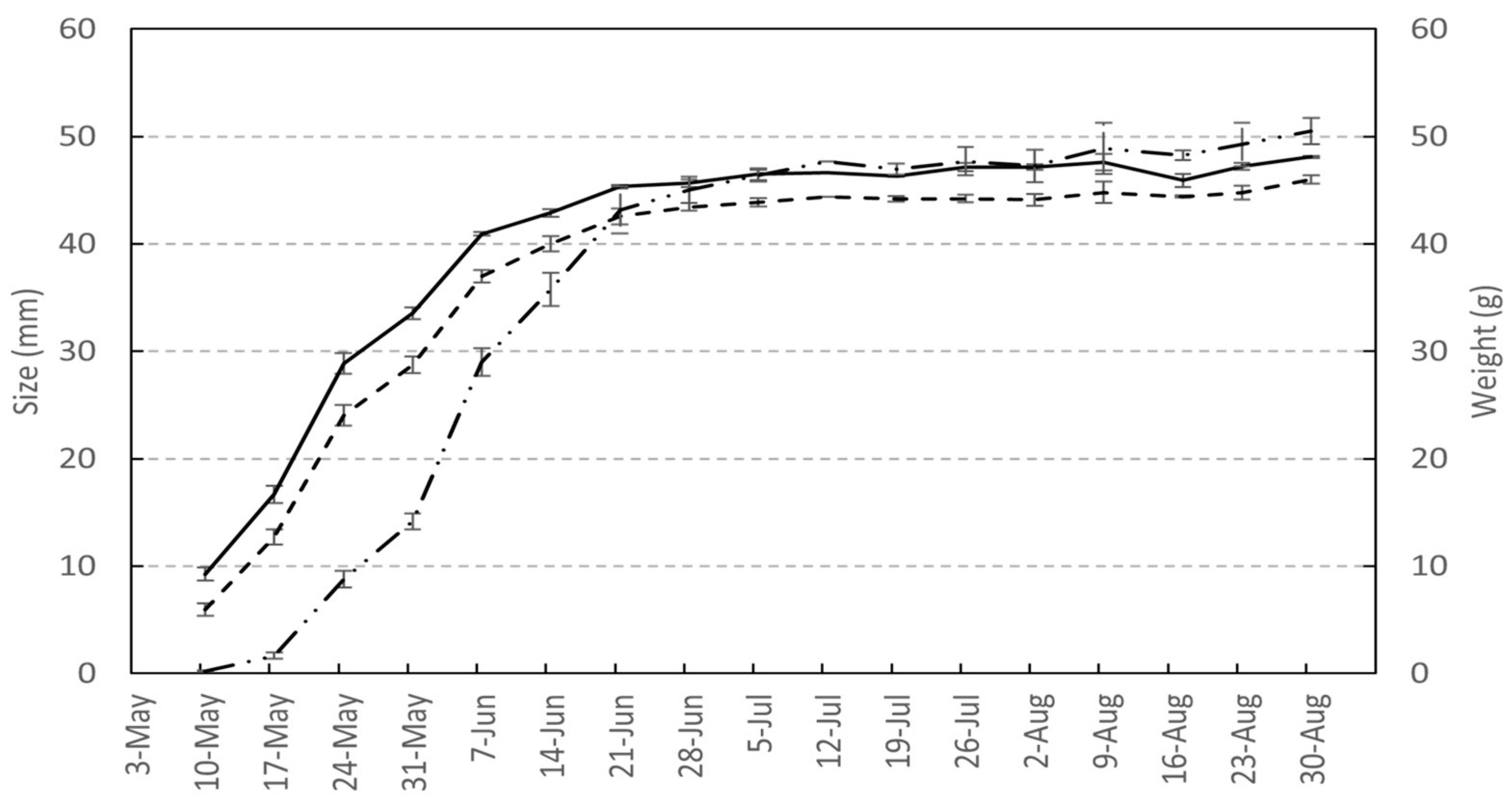
3.9. Principal Growth Stage 9: Leaf Senescence
The principal growth stage 9 for leaf senescence is subdivided into 4 developmental stages. Stage 93 occurs when leaf colour change begins. When more than 50% of the leaves have changed colour, stage 95 is reached. Stage 97 is reached when all the leaves have changed colour and leaf fall begins. Finally, when all the leaves have fallen, the defoliation date is reached, here described as stage 99.
4. Discussion
The Persian walnut BBCH scale that is outlined in this paper uses 9 principal growth stages and 57 detailed stages to describe the developmental phenophases of Persian walnut. This level of detail is in agreement with other BBCH scales of nut tree fruits that have been developed, such as the scale for hazelnut with 53 stages [41], chestnut with 48 stages [39], almond with 39 stages [37] and pecan with 51 stages [42]. The BBCH scale outlined in Table 2 allows walnut phenological monitoring data that have been recorded in one of the three historically used scales to be translated into BBCH codes.
4.1. Flowering Phenophases
To develop a strictly numerical scale, male flowering is here presented using a two-digit code and female flowering is presented using a three-digit code using the same method that has previously been used for hazelnuts [40,41]. This method has the advantage of preventing confusion between male and female flowering because Persian walnut can be protogynous, protandrous or homogamous depending on the cultivar.
4.2. Fruit Development
Development of the fruit, that is, the nut, is described using an outline view. Fruit development is divided into two steps. The first step, concerning husk development, is described by stages 71 to 79 in this scale and the second step, concerning shell development and lignification, is described by stages 790 to 799. Kernel development and ripening are described using stages 81 to 89. Nut fall is detailed using three-digit codes from the beginning of fall (stage 891) to the point at which all nuts have fallen to the ground (stage 899). Nut growth shows a sigmoidal curve, with a rapid growth phase from May to mid-June, a slowdown in growth from mid-June to late June and finally a halt in growth in the beginning of July. The nut weight shows a slight increase of 5 g during fruit ripening from the gelatinous phase of the kernel (stage 79) to the time the nut is dry and rich in lipids and proteins (stage 899) [56].
4.3. Orchard and Research Implications of BBCH Codification
The BBCH scale that is outlined here will allow the standardisation of phenological stage description between research teams and advisors. The use of numerical stages will simplify the use of phenological stages in models using phenological characteristics. In commercial orchards, the application of phytosanitary products is generally recommended for a specific BBCH stage that has not been described for Persian walnut before 2023. Indeed, the first communication presenting a BBCH scale for Persian walnuts was presented by Papillon et al. during the XI International Symposium on Walnut & Pecan in June 2023 hosted by the ISHS [43].
5. Conclusions
The scale described here is the first to detail Persian walnut phenophases in 57 stages from seed germination to leaf fall. The two-digit and three-digit codes used for presenting male and female flowering provides an advantage to avoid ambiguity during phenology monitoring. Finally, the use of strictly numerical codes provides an advantage when data are stored in computer files, including tables. This proposed phenological scale is expected to increase the efficiency of both worldwide germplasm characterisation and breeding and provide help for future comprehensive studies of the adaptation of Persian walnuts to climate change.
Author Contributions
Conceptualization, J.T.; methodology, J.T., J.R. and S.P.; validation, J.R., A.B., L.A., S.P., E.T., M.-N.H., J.-B.P., M.B., S.S., B.W., T.B., M.D., F.L. and J.T.; formal analysis, J.T. and L.A.; investigation, J.T., J.R., S.P. and L.A.; resources, J.T.; data curation, J.T. and L.A.; writing—original draft preparation, J.R., A.B. and J.T.; writing—review and editing, A.B. and J.T.; supervision, J.T.; project administration, J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to auto financial support of the members of the French National Association of Hazelnut Growers (ANPN) and the cooperative Unicoque.
Acknowledgments
The authors thank the technical staffs of the Association Nationale des Producteurs de Noisettes (ANPN), Station Expérimentale de Creysse, Station Expérimentale Nucicole Rhône-Alpe (SENuRA), Centre Technique Interprofessionnel des Fruits et Légumes (CTIFL), INRAE, and of the cooperative Unicoque Noisette et Noix de France. The authors specifically grateful Charlène Thiriet for the providing of some pictures illustrated the scale. The authors thank the editor and the reviewers for their useful comments on the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rehder, A. Manual of Cultivated Trees and Shrubs. Man. Cultiv. Trees Shrubs 1949. [Google Scholar]
- Germain, É.; Prunet, J.-P.; Garcin, A. Le noyer; Monographie; CTIFL: Paris, 1999; ISBN 978-2-87911-104-9. [Google Scholar]
- Bernard, A.; Lheureux, F.; Dirlewanger, E. Walnut: Past and Future of Genetic Improvement. Tree Genet. Genomes 2018, 14, 1. [Google Scholar] [CrossRef]
- Woodworth, R.H. Meiosis of microsporogenesis in the Juglandaceae. Am. J. Bot. 1930, 17, 863–869. [Google Scholar] [CrossRef]
- Manning, W.E. The Classification Within the Juglandaceae. Ann. Mo. Bot. Gard. 1978, 65, 1058. [Google Scholar] [CrossRef]
- Stanford, A.M.; Harden, R.; Parks, C.R. Phylogeny and Biogeography of Juglans (Juglandaceae) Based on matK and ITS Sequence Data. Am. J. Bot. 2000, 87, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Zeven, A.C.; de Wet, J.M. Dictionary of Cultivated Plants and Their Regions of Diversity: Excluding Most Ornamentals, Forest Trees and Lower Plants, 2nd ed.; rev.; Centre for Agricultural Publishing and Documentation: Wageningen, 1982; ISBN 978-90-220-0785-3. [Google Scholar]
- Leslie, C.; McGranahan, G. Native Populations of Juglans Regia—A Draft. 1988, 19, 111–124. [Google Scholar]
- Carrion, J.S.; Sanchez-Gomez, P. Palynological Data in Support of the Survival of Walnut (Juglans regia L.) in the Western Mediterranean Area During Last Glacial Times. J. Biogeogr. 1992, 19, 623. [Google Scholar] [CrossRef]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Del Lungo, S.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; et al. Rethinking the History of Common Walnut (Juglans regia L.) in Europe: Its Origins and Human Interactions. PLoS ONE 2017, 12, e0172541. [Google Scholar] [CrossRef] [PubMed]
- Lieth, H. Phenology and Seasonality Modeling. Springer Science & Business Media, 2013; Volume 8, ISBN 3-642-51863-X. [Google Scholar] [CrossRef]
- Chmielewski, F.-M.; Rötzer, T. Response of Tree Phenology to Climate Change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Telling Time with Trees. Nat. Clim. Change 2022, 12, 299–299. [CrossRef]
- Charrier, G.; Bonhomme, M.; Lacointe, A.; Améglio, T. Are Budburst Dates, Dormancy and Cold Acclimation in Walnut Trees (Juglans regia L.) under Mainly Genotypic or Environmental Control? Int. J. Biometeorol. 2011, 55, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European Phenological Response to Climate Change Matches the Warming Pattern. Glob. Change Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Chuine, I.; Morin, X.; Bugmann, H. Warming, Photoperiods, and Tree Phenology. Science 2010, 329, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Piao, S.; Vitasse, Y.; Zhao, H.; De Boeck, H.J.; Liu, Q.; Yang, H.; Weber, U.; Hänninen, H.; Janssens, I.A. Increased Heat Requirement for Leaf Flushing in Temperate Woody Species over 1980–2012: Effects of Chilling, Precipitation and Insolation. Glob. Change Biol. 2015, 21, 2687–2697. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining Global Warming Effects on the Phenology of Spring Leaf Unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hänninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger Temperature Response of Autumn Leaf Senescence than Spring Leaf-out Phenology. Glob. Change Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Vitasse, Y.; Bugmann, H.; Bigler, C. Phenological Shifts Induced by Climate Change Amplify Drought for Broad-Leaved Trees at Low Elevations in Switzerland. Agric. For. Meteorol. 2021, 307, 108485. [Google Scholar] [CrossRef]
- Črepinšek, Z.; Solar, M.; Štampar, F.; Solar, A. Shifts in Walnut (Juglans regia L.) Phenology Due to Increasing Temperatures in Slovenia. J. Hortic. Sci. Biotechnol. 2009, 84, 59–64. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Baciu, A.; Botu, M.; Achim, Gh. Environmental factors' influence on walnut flowering. Acta Hortic. 2010, 83–88. [Google Scholar] [CrossRef]
- Luedeling, E. Climate Change Impacts on Winter Chill for Temperate Fruit and Nut Production: A Review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef]
- Cook, B.I.; Wolkovich, E.M.; Davies, T.J.; Ault, T.R.; Betancourt, J.L.; Allen, J.M.; Bolmgren, K.; Cleland, E.E.; Crimmins, T.M.; Kraft, N.J.B.; et al. Sensitivity of Spring Phenology to Warming Across Temporal and Spatial Climate Gradients in Two Independent Databases. Ecosystems 2012, 15, 1283–1294. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant Phenology and Global Climate Change: Current Progresses and Challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, A.K.; Chamberlain, C.J.; Morales-Castilla, I.; Buonaiuto, D.M.; Flynn, D.F.B.; Savas, T.; Samaha, J.A.; Wolkovich, E.M. Winter Temperatures Predominate in Spring Phenological Responses to Warming. Nat. Clim. Change 2020, 10, 1137–1142. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; McGranahan, G.; Leslie, C. Validation of Winter Chill Models Using Historic Records of Walnut Phenology. Agric. For. Meteorol. 2009, 149, 1854–1864. [Google Scholar] [CrossRef]
- Arora, R.; Rowland, L.J.; Tanino, K. Induction and Release of Bud Dormancy in Woody Perennials: A Science Comes of Age. HortScience 2003, 38, 911–921. [Google Scholar] [CrossRef]
- Petri, J.L.; Leite, G.B. Consequences of insufficient winter chilling on apple tree bud-break. Acta Hortic. 2004, 53–60. [Google Scholar] [CrossRef]
- Man, R.; Lu, P.; Dang, Q.-L. Insufficient Chilling Effects Vary among Boreal Tree Species and Chilling Duration. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Man, R.; Lu, P.; Dang, Q.-L. Effects of Insufficient Chilling on Budburst and Growth of Six Temperate Forest Tree Species in Ontario. New For. 2021, 52, 303–315. [Google Scholar] [CrossRef]
- Bernard, A.; Marrano, A.; Donkpegan, A.; Brown, P.J.; Leslie, C.A.; Neale, D.B.; Lheureux, F.; Dirlewanger, E. Association and Linkage Mapping to Unravel Genetic Architecture of Phenological Traits and Lateral Bearing in Persian Walnut (Juglans regia L.). BMC Genomics 2020, 21, 203. [Google Scholar] [CrossRef]
- Marrano, A.; Sideli, G.M.; Leslie, C.A.; Cheng, H.; Neale, D.B. Deciphering of the Genetic Control of Phenology, Yield, and Pellicle Color in Persian Walnut (Juglans regia L.). Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Bleiholder, H.; Van Den Boom, J.; Langelüddeke, P.; Stauss, R. Einkeitliche Codierung Der Phänologischen Stadien Bei Kultur-Und Schadpflanzen. Gesunde Pflanz. 1989, 41, 381–384. [Google Scholar]
- Hack, H.; Bleiholder, H.; Buhr, L.; Meier, U.; Schnock-Fricke, U.; Weber, E.; Witzenberger, A. Einheitliche Codierung der phänologischen Entwicklungsstadien mono-und dikotyler Pflanzen—Erweiterte BBCH-Skala, Allgemein. Nachrichtenblatt Dtsch. Pflanzenschutzdienstes 1992, 44, 265–270. [Google Scholar]
- Sakar, E.H.; El Yamani, M.; Boussakouran, A.; Rharrabti, Y. Codification and Description of Almond (Prunus dulcis) Vegetative and Reproductive Phenology According to the Extended BBCH Scale. Sci. Hortic. 2019, 247, 224–234. [Google Scholar] [CrossRef]
- Adiga, J.D.; Muralidhara, B.M.; Preethi, P.; Savadi, S. Phenological Growth Stages of the Cashew Tree (Anacardium occidentale L.) According to the Extended BBCH Scale. Ann. Appl. Biol. 2019, 175, 246–252. [Google Scholar] [CrossRef]
- Larue, C.; Barreneche, T.; Petit, R.J. Efficient Monitoring of Phenology in Chestnuts. Sci. Hortic. 2021, 281, 109958. [Google Scholar] [CrossRef]
- Paradinas, A.; Ramade, L.; Mulot-Greffeuille, C.; Hamidi, R.; Thomas, M.; Toillon, J. Phenological Growth Stages of ‘Barcelona’ Hazelnut (Corylus Avellana L.) Described Using an Extended BBCH Scale. Sci. Hortic. 2022, 296, 110902. [Google Scholar] [CrossRef]
- Toillon, J.; Hamidi, R.; Paradinas, A.; Ramade, L.; Thomas, M.; Suarez Huerta, E. Consolidated BBCH Scale for Hazelnut Phenotyping. Acta Hortic. 2023, 159–168. [Google Scholar] [CrossRef]
- Han, M.; Peng, F.; Marshall, P. Pecan Phenology in Southeastern China: Pecan Phenology in Southeastern China. Ann. Appl. Biol. 2018, 172, 160–169. [Google Scholar] [CrossRef]
- Papillon, S.; Robin, J.; Tranchand, E.; Hebrard, M.-N.; Philibert, J.; Barbedette, M.; Lheureux, F.; Toillon, J. Application of BBCH Codification to Walnut (Juglans regia L.) Phenophase. Presented at the IX International Symposium on Walnut & Pecan, Grenoble, France, 2023. [CrossRef]
- Germain, E.; Jalinat, J.; Marchou, M. Divers Aspects de La Biologie Florale Du Noyer. 1975. [Google Scholar]
- International Plant Genetic Resources Institute. Descriptors for Walnut (Juglans spp.); Bioversity International, 1994; ISBN 92-9043-211-X. [Google Scholar]
- Mahmoodi, R.; Hassani, D.; Amiri, M.E.; Jaffaraghaei, M. Phenological and Pomological Characteristics of Five Promised Walnut Genotypes in Karaj, Iran. J. Nuts 2016, 7. [Google Scholar] [CrossRef]
- Soleimani, A.; Rabiei, V.; Hassani, D.; Mozaffari, M.R. Effect of Genetic-Environment Interaction on Phenology of Some Persian Walnut (Juglans regia L.) Genotypes. Crop Breed. J. 2020. [Google Scholar] [CrossRef]
- Shah, R.A.; Bakshi, P.; Sharma, N.; Jasrotia, A.; Itoo, H.; Gupta, R.; Singh, A. Diversity Assessment and Selection of Superior Persian Walnut (Juglans regia L.) Trees of Seedling Origin from North-Western Himalayan Region. Resour. Environ. Sustain. 2021, 3, 100015. [Google Scholar] [CrossRef]
- McGranahan, G.; Leslie, C. ‘Robert Livermore’, a Persian Walnut Cultivar with a Red Seedcoat. HortScience 2004, 39, 1772. [Google Scholar] [CrossRef]
- McGranahan, G.; Leslie, C. Walnut Tree Named ‘Forde’ 2006. USPP16495P3. https://patents.google.com/patent/USPP16495P3/en.
- McGranahan, G.; Leslie, C. Walnut Tree Named ‘Gillet’ 2006. USPP17135P3. https://patents.google.com/patent/USPP17135P3/en.
- McGranahan, G.; Leslie, C. Walnut Variety Named “DURHAM” 2017. US20170215313P1. https://patents.google.com/patent/US20170215313P1/en.
- McGranahan, G.H.; Forde, H.I.; Snyder, R.G.; Sibbett, G.S.; Reil, W.; Hasey, J.; Ramos, D.E. ‘Tulare’ Persian Walnut. HortScience 1992, 27, 186–187. [Google Scholar] [CrossRef]
- Leslie, C.A.; McGranahan, G.H. The Origin of the Walnut. Walnut Prod. Man. 1998, 3–7. [Google Scholar]
- Germain, E.; Jalinat, J.; Marchou, M. Divers Aspects de La Biologie Florale Du Noyer. In Le noyer; Invuflec, 1975; p. 187. [Google Scholar]
- Martínez, M.L.; Labuckas, D.O.; Lamarque, A.L.; Maestri, D.M. Walnut (Juglans regia L.): Genetic Resources, Chemistry, by-Products. J. Sci. Food Agric. 2010, 90, 1959–1967. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



