Submitted:
29 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
The long-term use of fertilizers and pesticides in conventional cultivation has resulted in a decrease in soil productivity and vegetable yields in greenhouses. However, only a limited number of studies have explored the impacts of organic cultivation on greenhouse soil. Specifically, there is a scarcity of research examining the alterations in soil aggregate mediated organic carbon and microbial communities, which are known to enhance soil productivity. This study investigated the properties of soil aggregates, including the levels of organic carbon fractions, microbial community, and enzyme activity with the three aggregate classes microaggregates (<0.25 mm), small macroaggregates (2-0.25 mm) and large macroaggregates (>2 mm). These investigations were conducted under three different cultivation methods in a greenhouse: conventional cultivation (CC), integrated cultivation (IC), and organic cultivation (OC) in greenhouse. The results showed that (1) OC and IC promoted the formation of small macro-aggregates and enhanced aggregate stability compared to CC, (2) SOC in the three size fractions of OC increased by 98.99%-92.06% compared to CC; EOC increased by 98.47%-117.59%; POC increased by 138.59%-208.70%; MBC increased by 104.71%-230.61%; DOC increased by 21.93%-40.90%, respectively; (3) Organic cultivation significantly increased enzyme activity in all three particle size aggregates and increased the relative abundance of bacteria in micro-aggregates as well as the relative abundance of fungi in small macro-aggregates. Structural equation model (SEM) analysis revealed that organic farming practices fostered the development of smaller macro-aggregates, elevated microbial and enzyme activities within soil aggregates, and facilitated the conversion of soil nutrients and carbon sequestration. consequently, this led to an increase in soil organic carbon content and improved vegetable yield in greenhouse environments. In conclusion, the soil carbon content and vegetable yield are enhanced through long-term organic cultivation in greenhouses, primarily by increasing the proportion of small macro-aggregates.
Keywords:
organic cultivation
; soil aggregates
; organic carbon fraction
; microbial community
; enzyme activity
1. Introduction
The utilization of greenhouse production system (protected cropping) is crucial in addressing the growing global food demand, considering the challenges posed by global climate change. These systems offer sustainability and high productivity, making them essential. Greenhouse vegetable production has made significant contributions to China agricultural. However it relies heavily on fertilizer and water input, which places a considerable burden on the environment and ecology [1,2]. China, as the top vegetable-producing nation, achieved a staggering production of million tons of vegetables in 2017, representing for 51.2% of the global vegetables yield [3,4]. In comparison to open-field production systems, greenhouses have demonstrated a remarkable capacity to enhance vegetable yields. By artificially manipulating temperature and employing higher levels of fertilizer and water inputs, greenhouses can successfully produce off-season vegetables regardless of the specific climate type or region in China.
Nevertheless, the extensive use of chemical fertilizers and pesticides in greenhouses has led to significant soil and ecological issues [1,2]. Applying substantial quantities of chemical fertilizers and pesticides is deemed crucial in achieving high yields of greenhouse vegetables. Nonetheless, this approach fails to provide a sustainable means of consistently improving soil fertility or effectively managing pests and diseases. Numerous studies have demonstrated that prolonged utilization of chemical fertilizer and pesticide application in greenhouses leads to soil acidification, salinization and degradation [5]. Additionally, it leads to the accumulation of heavy metals [6], and severe pollution of groundwater and irrigation water [7], Furthermore, it results in a decline in soil microbial diversity and enzyme activity [8]. A multitude of studies have substantiated the excessive application of pesticides in greenhouses as a response to diseases and insect pests influenced by high temperature and humidity [9,10,11].
Organic cultivation serves as a sustainable agricultural production system that enhances soil fertility, biodiversity and human health. It achieves this by prohibiting the utilization of chemical synthetics and instead emphasizes the application of organic waste or crop rotation [12,13]. Hence, organic cultivation is regarded as an advantageous and viable alternative to intensive agriculture, playing a significant role in the pursuit of sustainable agricultural development.
Soil aggregates, consisting of the coupling of mineral particles and organic substances, serve as fundamental units of soil structure and function [14]. Aggregates of varying sizes exhibit distinct properties and functions. For instance, macro-aggregates (>0.25 mm) contain a higher abundance of labile organic substances and fresh litter, which are more readily decomposed by microorganisms. In contrast, stable organic carbon within micro-aggregates (<0.25 mm) is characterized by greater stability and reduced accessibility for biological degradation. Nevertheless, the prolonged use of chemical fertilizers and pesticides can detrimentally impact soil structure, disrupt the distribution of soil aggregates, and diminish their stability [15]. SOC plays a crucial role in their formation and stability of soil aggregates by acting as a binding agent, with its properties influencing the process of aggregate formation and evolution.
Organic cultivation practices, including the application of organic fertilizer, crop rotation, and integrated management, exhibit a favorable impact on the content of SOC. Maintaining the dynamic equilibrium of SOC is of paramount importance in enhancing soil structure, soil quality, and mitigating global climate change [16,17]. Organic farms demonstrated significantly higher concentrations, stocks, and rates of soil organic carbon (SOC) sequestration in comparison to conventional farms [12]. This highlights organic cultivation as a crucial approach in reducing greenhouse gas emissions. The labile organic carbon (LOC) fractions exhibit greater sensitivity to changes in agricultural practices compared to SOC, and they demonstrate a stronger correlation with soil fertility, soil microbial activity, and crop yield [18]. LOC pool comprises easily oxidizable carbon (EOC), microbial biomass carbon (MBC), light fraction organic carbon (LFOC), and dissolved organic carbon (DOC) [19,20]. Research studies have demonstrated that organic cultivation practices not only increase the content of SOC, but also enhances the levels of labile organic carbon fractions [19,21]. In addition, organic farming practices have been shown to sustain long-term higher fertility, enhances soil microbial diversity, provide protection against pathogen infections, and facilitates the degradation of pollutants [12,22,23].
Soil microorganisms play a critical role as the principal agents in the synthesis of soil organic matter. They actively contribute to the formation of soil aggregates, which in turn promotes the accumulation and long-term stability of SOC. Soil aggregates of varying particle sizes exhibit diverse physical and chemical properties, leading to a heterogeneous distribution of microbial communities and substrate availability [24]. Soil microbial communities residing within soil aggregates play a pivotal role in energy transfer and nutrient cycling processes, with their mycelia facilitating the formation and stability soil aggregation. The microbial community structure in soil aggregates serves as a significant indicator for assessing soil carbon sequestration and ecological functionality [25,26]. Enzyme activity is a sensitive parameter that accurately reflects carbon turnover dynamics within the soil and serves as a valuable biological indicator for assessing soil fertility [27]. The activity of enzyme is subject to the influence microbial communities, thereby impacting the conversion of various organic carbon forms within aggregates [28]. Consequently, significant variations in both the organic carbon content and availability exist among soil aggregates of different particle sizes, thereby governing the distribution of enzyme activity within these aggregates [29].
Agricultural fertilization stands as the foremost influential factor in promoting the enhancement of soil aggregates [30]. Organic fertilizers possess a substantial quantity of organic matter, and the increment in soil organic carbon content facilitates the formation of macro-aggregates while augmenting the physical safeguarding of organic matter [31]. However, limited research has been conducted to explore the impacts of organic cultivation on the intricate interplay among organic carbon fractions, microbial community composition, and enzyme activity at the aggregate level.
In this study, we employed a16-year long-term field experiment in greenhouses in Quzhou County, Hebei Province, China. The experiment involved organic, integrated, and conventional cultivation management. The objectives of this study were to: (i) examine the distribution patterns of organic carbon fractions and microbial community structure across various particle sizes, (ii) investigate how organic cultivation practices can enhance soil function through alterations in soil aggregates, and (iii) elucidate the mechanisms underlying carbon sequestration in organic cultivation systems. This study primarily emphasizes the impact of organic cultivation on both microbial community composition and SOC at the aggregate level, as well as the sequestration of SOC at the larger-scale bulk soil level. Additionally, it explores the relationships between these factors and vegetable yield within the greenhouse setting.
2. Materials and Methods
2.1. Study Site
In March 2002, a long-term experiment was commenced at the Quzhou Experimental Station of China Agricultural University (latitude: 36°52'N, longitude: 115°01'E), situated in the northern region of Quzhou County, Hebei Province, China. This region exhibits a temperate, semi-humid continental monsoon climate, with an average temperature of 13.1℃. The average annual rainfall in this region amounts to 556.2 mm, with the majority of it occurring between the months of July and September. This period accounts for approximately one-third of the total annual rainfall. The solar greenhouse utilized in this study had dimensions of 7 × 52 m (Figure 1). The soil under analysis was salinized cinnamon soil. Prior to the long-term experiment, the soil at the experimental site consisted of 54.11% sand, 28.45% silt, and 17.44% clay. The physicochemical properties of the topsoil (0-20 cm) under various cultivation practices in 2002 are presented in Table 1.
The experiment investigating greenhouse vegetable cultivation entailed three methods: conventional cultivation (CC), integrated cultivation (IC), and organic cultivation (OC). A two-season vegetable rotation was implemented within the greenhouses, with the specific types of vegetables planted each year as indicated in Table S1. Conventional cultivation (CC) followed local traditional management practices. The primary fertilizer utilized is chemical fertilizer, with a small amount of organic fertilizer used as supplementation. Insecticides (such as triazophos, imidacloprid, and abamectin) and fungicides (such as azoxystrobin, propineb, carbendazim, and mancozeb) are employed for pest and disease control.
The chemical fertilizer application rate is 4 t ha-1 year-1, while the organic fertilizer application rate is 13.4 t ha-1 year-1. The chemical fertilizers used comprised of urea (46% N), calcium superphosphate (12% P2O5), potassium sulfate (50% K2O), and diammonium phosphate (18% N and 46% P2O5), with a mixing ratio of 3:4:5. The fertilization approach for integrated cultivation (IC) combines organic and chemical fertilizers, while biological control is primarily employed for pest and disease prevention, supplemented with the application of insecticides and fungicides in severe cases (similar to conventional farming practices). In this system, the utilization of chemical fertilizers and pesticides is reduced by 50% compared to conventional cultivation (CC). The types and mixing ratios of chemical fertilizers remain consistent with conventional cultivation (CC). The application rate of organic fertilizer is reduced by 50% compared to organic cultivation (OC). Strict adherence to organic management principles in OC prohibits the use of chemical fertilizers, pesticides, and any hormonal substances. Only organic fertilizers are applied, in conjunction with artificial control, sulfur fumigation, high temperature enclosed structures, and biological pesticides, to prevent and control pests and diseases.
Organic fertilizers are produced through a high-temperature composting process and primarily comprise local organic waste, such as chicken manure, cow manure, and straw. The application rate of organic fertilizer is 59.2 t ha-1 year-1. The nutrient content of organic fertilizer on a dry basis is 1.21% N, 0.60% P2O5, and 1.58% K2O. The crops were fertilized three times per season, with one application of basal manure and two applications of top dressing. The irrigation in the greenhouses was centrally managed. The management of the three greenhouses was conducted simultaneously, utilizing flood irrigation as the irrigation method. Details regarding nutrient input, pesticide application, and irrigation under various cultivation methods are provided in Table S2.
2.2 Soil Sampling and Aggregate Size Fractionation
Undisturbed soil samples were collected from three cultivation types (CC, IC, OC) at a depth of 0-20 cm on May 11, 2018. Sampling was conducted in each greenhouse following an "S" shape pattern, with a composite soil sample being created by combining every five points sampled. Finally, the average samples obtained using the quarter method were homogenized, carefully disassembled along the natural fracture points, and sieved through a 5-mm mesh. Plant and organic residues in the sieved soil were meticulously extracted using forceps. The soil samples needed for enzyme activity analysis were stored in a refrigerator at a temperature of 4℃. Subsamples weighing 200 g were subsequently subjected to shaking through sieving devices with pore diameters of 2 and 0.25 mm for a duration of 5 minutes, resulting in the separation of three size fractions: <0.25 mm (micro-aggregates), 2-0.25 mm (small macro-aggregates), and >2 mm (large macro-aggregates). The sieved soil was divided into two portions: one portion was stored at 4 ℃ for the analysis of soil enzyme activity and microbial community composition, while the other portion was utilized for the analysis of organic carbon fractions. The mean weight diameter (MWD in mm) was employed to assess aggregate stability, which was calculated using Eq (1) as described by Fattet [32].
Where Xi represents the mean diameter of the size fractions (mm) and represents the proportion of the size fraction in the entire soil sample (%).
2.3 Soil Organic Carbon Fractions Analyses
The soil organic carbon (SOC) content was determined using the K2Cr2O7 oxidation method. Specifically, 5 mL of 0.8 M K2Cr2O7 solution and 5 mL of concentrated H2SO4 were added to the pre-weighed soil sample. The mixture was subsequently heated to boiling at a temperature range of 170℃-180℃ for a duration of 5 minutes. The residual K2Cr2O7 solution was titrated using 0.2 M FeSO4. The soil particulate organic carbon (POC) content was determined using the method outlined by Cambardella and Elliott in their publication from May-June 1992[33]. In summary, 20 g of each soil aggregate fraction was dispersed in 100 mL of sodium hexametaphosphate by agitation with a reciprocating shaker for a duration of 18 hours. The resulting soil suspension was passed through a 53-μm sieve under distilled water to facilitate separation. All the material retained on the sieve was transferred onto a dry dish, subjected to oven-drying at 60℃ for 12 hours, and subsequently ground to determine the carbon (C) content of the dried particulate. The organic carbon (C) concentrations within the POC samples were determined using the potassium dichromate oxidation method.
The extractable organic carbon (EOC) content was estimated using the method described by Blair et al. in their publication from 1995. Furthermore, 1.5 g of each soil aggregate fraction was accurately weighed and transferred into plastic screw-top centrifuge tubes. Subsequently, 25 mL of 1/3 mol L-1 KMnO4 solution was added to each tube. All the tubes were securely sealed, agitated for a duration of 1 hour, and then centrifuged for 5 minutes at a speed of 3000 rpm. The supernatant was then diluted with deionized water, and the carbon (C) content was measured through colorimetry at a wavelength of 565 nm. Finally, the change in KMnO4 concentration was utilized to quantify the amount of carbon dioxide, based on the assumption that 1 mM of KMnO4 was consumed during the oxidation of 9 g of carbon (C).
2.4 Soil Enzyme Activities Analyses
The methods employed for measuring the activities of invertase, catalase, urease, and cellulase were as follows:
Urease: 5 g of dry soil was subjected to incubation with 20 mL of a 10% urea solution, 20 mL of citric acid buffer at pH 6.7, and 1 mL of toluene for a duration of 24 hours at 37℃ in the absence of light. Following the incubation, the soil suspension was filtered, and 3 mL of the filtrate was employed to determine the concentration of ammonium ions using the indophenol-blue colorimetric method. The activity of urease was expressed by the unit of mg kg-1 24 h-1.
Catalase: 2 g of fresh soil, 40 mL distilled water, and 5 mL of 0.3% hydrogen peroxide solution were shaken for 20 minutes. The activity of urease was expressed in units of mg kg-1 24 h-1. For catalase assay, 2 g of fresh soil, 40 mL of distilled water, and 5 mL of a 0.3% hydrogen peroxide solution were vigorously shaken for a duration of 20 minutes. Following the shaking step, 5 mL of 3 M sulfuric acid was introduced to the mixture, and the resulting soil suspension was subsequently filtered. Moreover, a 25-mL portion of the filtrate was utilized to determine the quantity of hydrogen peroxide through titration with 0.1 M potassium permanganate. The activity of catalase was expressed in units of mg g-1. Invertase: A total of 5 g of fresh soil was subjected to incubation with 15 mL of an 8% sucrose solution, 5 mL of phosphate buffer at pH 5.5, and five drops of toluene for a duration of 24 hours at 37℃ in the absence of light. Following the incubation, the soil suspension was filtered, and 1 mL of the filtrate was utilized to quantify the amount of glucose produced using the 3,5-dinitrosalicylic acid colorimetric method. The activity of invertase was expressed in units of mg g-1 24 h-1.
Cellulase: A total of 10 g of fresh soil was subjected to incubation with 20 mL of a 1% carboxymethyl cellulose solution, 5 mL of phosphate buffer at pH 5.5, and a toluene solution of carboxymethyl cellulose for a duration of 72 hours at 37℃ in the absence of light. Immediately following the incubation, the soil suspension was filtered, and 1 mL of the filtrate was employed to quantify the amount of glucose produced using the 3,5-dinitrosalicylic acid colorimetric method. The activity of invertase was expressed in units of mg kg-1 72 h-1.
2.5 High-Throughput Sequencing
The soil microbial communities at different aggregate levels were analyzed for high-throughput sequencing in different cultivations. The Fast DNA Spin Kit (MP Biomedicals, USA) was employed for soil DNA extraction, and the purity and concentration of the extracted DNA were assessed using 1% agarose gel electrophoresis. The DNA was diluted with sterile water in centrifuge tubes to a concentration of 1 ng μl-1. The diluted DNA was used to amplify the hypervariable regions V3-V4 of the bacterial 16S rRNA gene using primers 341F and 806R. Additionally, the fungal ITS gene was amplified using primers ITS1-1F-F and ITS1-1F-R. The primer sequences for bacteria and fungi were provided in Table S3. Moreover, the PCR reactions were presented in Table S4, while the reaction program was displayed in Table S5. Subsequently, the PCR products were combined and purified using a GeneJET purification kit (Thermo Fisher Scientific, USA). The raw sequences underwent quality screening and trimming using Cut adapt. The chimeric sequences were identified and removed using the UCHIME algorithm [34]. Bacterial and fungal operational taxonomic units (OTUs) were clustered at a sequence similarity threshold of 97% using UPARSE. The SSUrRNA database and the Unit database were utilized for bacterial and fungal OTU annotation analysis, respectively [35].
2.6 Statistical Analysis
All data were analyzed using Microsoft Excel (2016) (Microsoft, USA) and GraphPad Prism 8.0.2. Furthermore, a one-way analysis of variance (ANOVA) was utilized to evaluate the impact of various cultivations on soil aggregate distribution, as well as the content of organic carbon fractions mediated by aggregates, enzyme activity, and microbial activity. Multiple comparisons were conducted using a one-way ANOVA with a Tukey's HSD post hoc test in SPSS 20.0. Structural Equation Modeling (SEM) was employed to analyze the internal relationships between soil organic carbon fractions, enzyme activities, microbial community, and vegetable yield.
3. Results
3.1 Soil Aggregate Distribution and Stability
Soil aggregates are fundamental functional units in soil structure and the soil ecosystem, playing a vital role in the cycling, transformation, and sequestration of soil organic carbon. The CC treatment exhibited the highest proportion of large macroaggregates, accounting for 38.90% (Figure 2). After 15 years, organic and integrated cultivation practices have been observed to elevate the content of small macroaggregates and microaggregates, while decreasing the content of large macroaggregates. In comparison to CC, OC and IC cultivation methods stimulated the formation of small macroaggregates, leading to a significant increase. In OC, the proportions of small macroaggregates and microaggregates increased by 5.95% and 10.05%, respectively, while the proportion of large macroaggregates decreased by 13.41%. Similarly, in IC, the proportions of small macroaggregates and microaggregates increased by 13.83% and 7.98%, respectively, while the proportion of large macroaggregates decreased by 23.00% compared to CC (Figure 2). MWD (mean weight diameter) serves as an indicator of the size distribution of soil aggregates, where a larger MWD value signifies greater stability of the soil aggregates. In this study, there was no significant difference observed in MWD among the three cultivation methods.
3.2 Soil Organic Carbon Fractions and Enzyme Activities within Aggregates
Organic and integrated cultivation practices resulted in a significant increase in the concentration of organic carbon fractions across all sizes of aggregates, as compared to conventional cultivation. No significant difference was observed in the concentration of organic carbon fractions among different particle sizes of aggregates in both conventional and integrated cultivation methods (Figure 3). However, the distribution of organic carbon fractions in organic cultivation exhibited significant variations depending on aggregate size fractions, with an increase observed as the aggregate size decreased. The concentrations of organic carbon fractions in the bulk soil and three aggregate classes were found to be highest in OC, followed by IC, while CC exhibited the lowest concentrations (Figure 3and Figure S1). In comparison to CC, the soil organic carbon (SOC) content in the three size fractions of OC increased by 92.06% to 98.99%, EOC increased by 98.47% to 117.59%, POC increased by 138.59% to 208.70%, MBC increased by 104.71% to 230.61%, and DOC increased by 21.93% to 40.90% (Figure 3). The results indicate that organic cultivation significantly enhanced the concentration of organic carbon fractions in all aggregate sizes, with a notable increase observed in small macroaggregates.
The enzyme activity is highly sensitive and related to carbon conversion. Organic cultivation resulted in a significant increase in enzyme activity in both the bulk soil and three particle size aggregates (Figure 4 and Figure S2). In the OC treatment, the urease activity in all aggregates exhibited a significant increase compared to CP, while the catalase activity showed a significant increase in macroaggregates but not in microaggregates (Figure 4). Compared to CC, the activities of invertase showed a significant increase in large macroaggregates and microaggregates in OC, while the activities of cellulase exhibited a significant increase in small macroaggregates and microaggregates (Figure 4).
3.4 Microbial Community
The relative abundance of soil microbial communities varies across different particle size aggregates and cultivation systems (Figure 5). Organic cultivation resulted in an increased relative abundance of bacteria in microaggregates and an increased relative abundance of fungi in small macroaggregates. Proteobacteria, Bacteroidetes, Acidobacteria, and Actinobacteria were identified as the dominant bacterial phyla in all aggregates, with their relative abundance accounting for 79.45% to 86.25%. Organic cultivation resulted in an increased relative abundance of Proteobacteria and Bacteroidetes in the microaggregates (<0.25 mm). Ascomycota, Mortierellomycota, Blastocladiomycota, and Basidiomycota were identified as the predominant fungal phyla in all aggregates. Across all cultivation treatments, the relative abundance of Mortierellomycota, Blastocladiomycota, and Basidiomycota was found to be highest in small macroaggregates (2-0.25 mm) in the organic cultivation treatment.
3.5 Vegetable Yield
Long-term organic cultivation had a significant impact on vegetable yields in greenhouses. The results demonstrated that organic cultivation in greenhouse exhibits comparable vegetable yields in the early stages when compared to conventional cultivation. However, as the years progress, organic cultivation yields surpass those of conventional cultivation. As illustrated in Figure 6, the organic cultivation (OC) of vegetables in both spring and autumn surpasses that of integrated cultivation (IC) and conventional cultivation (CC) after 16 years. Compared to conventional cultivation (CC), the spring vegetable production increased by a range of 3.21% to 62.28%, while the autumn vegetable production saw an increase of 4.52% to 34.84%. Over the course of 15 years of cultivation, there was significant fluctuation observed in vegetable output (Figure 6). This fluctuation can be attributed to the variations in vegetable types planted across different years. Significant differences in vegetable output were observed due to the notable variation in the quality of root vegetables and leafy vegetables.
Nevertheless, organic cultivation demonstrated clear advantages in terms of yield, regardless of the number of planting years or the types of vegetables, and these advantages became even more pronounced as the cultivation period increased. This indicates that the increase in vegetable yields in organic greenhouses can be attributed to the positive impact of organic cultivation on biodiversity, which in turn may contribute to higher yields.
3.5 Quantitative Examination of the Effects of Different Cultivations and Aggregates
Various cultivation methods exert a discernible influence on the composition of soil aggregates, as well as the organic carbon and microorganisms they contain (Figure 7). As depicted in Figure 7a, there was a negative correlation observed between soil aggregates and soil organic carbon fractions, microbial activity, enzyme activity, and vegetable yield in the conventional cultivation (CC) group. In the integrated cultivation (IC) treatment group, soil aggregates exhibited a negative correlation with soil organic carbon fractions, microbial activity, and enzyme activity, while showing a positive correlation with vegetable yield (Figure 7b). In contrast, the organic cultivation (OC) treatment group demonstrated a positive correlation between soil aggregates and microbial activity, enzyme activities, and vegetable yield, while exhibiting a negative correlation with soil organic carbon fractions (Figure 7c).
These findings suggest that organic cultivation promotes an increase in soil aggregate particle size and stability, leading to enhanced microbial activity and enzyme activity, ultimately resulting in increased vegetable yield. Furthermore, soil aggregates were found to exert a detrimental influence on soil organic carbon fractions, microbial activity, enzyme activity, and vegetable yield in both conventional and integrated management approaches. This indicates that the soil nutrient content, biological activity, and vegetable yield are comparatively lower.
4. Discussion
4.1 Organic Cultivation Effects on Soil Aggregation
Soil aggregates serve as fundamental units for soil structure and ecological function [36]. However, conventional cultivation practices have been found to disrupt soil structure through excessive tillage or the excessive application of nitrogen fertilizer. Organic cultivation practices have demonstrated a significant improvement in soil structure by enhancing soil porosity and water retention [12]. Organic cultivation practices, including the utilization of organic fertilizer, have been shown to facilitate the formation of soil aggregates. These aggregates play a crucial role as binding agents, promoting the cohesion of soil particles and aiding in the formation of macro-aggregates. Organic matter present in organic fertilizers acts as a vital binding agent for soil structure. Consequently, increasing the content of soil organic matter serves as a crucial strategy for enhancing soil structure.
Organic cultivation induces an increase in the content of organic matter and biologically active substances in the soil, thereby promoting the development of soil aggregates. This is primarily accomplished through the utilization of organic fertilizers and the exclusion of synthetic chemical fertilizers and pesticides [37,38]. The study conducted by Pimentel et al. revealed that organic cultivation practices facilitate the formation of aggregates exceeding a particle size of 0.25 mm and enhance their stability [39].
This study demonstrated that both organic and integrated cultivation approaches resulted in an increased proportion of small macroaggregates and microaggregates, while reducing the proportion of large macroaggregates in greenhouse soils when compared to conventional cultivation practices. Small macroaggregates constitute the predominant component of the soil, comprising 43.11% to 49.07% of the total bulk soil volume. When compared to conventional cultivation, organic cultivation practices have resulted in a 5.94% increase in the proportion of small macroaggregates, while integrated cultivation has exhibited a more significant increase of 13.83% in the same category. Furthermore, organic cultivation has exhibited a reduction of 13.83% in the proportion of large macroaggregates, whereas integrated cultivation has achieved a more substantial reduction of 23.00% in the same category. Comparative analysis reveals that integrated cultivation presents the most notable enhancement in soil structure, exhibiting the highest content of small macroaggregates. However, no significant difference is observed between organic cultivation and integrated cultivation in terms of their impact on soil structure.
Nevertheless, after 16 years of management, organic cultivation exhibited the lowest soil bulk density compared to integrated and conventional cultivation practices (Figure S3). This implies that organic cultivation significantly enhanced the soil structure, surpassing the impact achieved by integrated cultivation. Consequently, no significant disparity was observed in the particle size distribution of soil aggregates between organic and integrated cultivation methods. This finding can be attributed primarily to the employed method for isolating soil aggregates. The soil aggregate isolation method comprises both dry sieving and wet sieving techniques. The utilization of the dry sieving method introduces noticeable variations between replicated soil samples, as it involves the sieving of air-dried soil. This process hampers the identification of potential disparities in the aggregate distribution among different cultivated soils.
4.2 Organic Cultivation Effects the Distribution of Organic Carbon Fractions and Enzyme Activity within Aggregate
Organic cultivation not only significantly enhanced the distribution of organic carbon fractions in three particle sizes of aggregates and bulk soil, but also had a pronounced impact on the distribution of organic carbon fractions across all aggregates (Figure 3 and Figure S1). There were no significant differences observed in the concentrations of organic carbon fractions within different particle aggregates between conventional and integrated cultivation. However, organic cultivation resulted in a significant increase in the content of organic carbon fractions within both small macro-aggregates and micro-aggregates. Specifically, organic cultivation resulted in a significant increase in the concentrations of SOC, EOC, and POC within small macro-aggregates, while MBC and EOC showed a notable increase within micro-aggregates (Figure 3). In organic cultivation, the content of soil organic carbon fractions is influenced by both the application of organic fertilizers and the implementation of specific management practices.
However, the application of organic fertilizers has been identified as the most significant factor [37,38]. This can be mainly attributed to two factors: (i) the increased crop yields achieved through the application of organic fertilizer, which led to a significant addition of crop residues incorporated into the soil, and (ii) the direct input of organic substances derived from the use of organic manure. Xie et al. demonstrated that the application of organic fertilizer significantly increased the concentrations of SOC in all size fractions of soil aggregates [40]. Therefore, organic cultivation led to a significant increase in the concentration of soil carbon fractions across all size fractions of aggregates and resulted in alterations in the distribution of SOC through the process of aggregation.
The Microbial Efficiency-Matrix Stabilization (MEMS) model proposed that soil organic matter (SOM) is generated via the decomposition of plant litter and subsequent microbial transformations [41]. The interplay between SOC and the mineral soil substrate plays a crucial role in the long-term retention of carbon in the soil. As mineral particles and SOC combine to form various sizes of soil aggregates, these soil aggregates have significant implications in enhancing soil fertility, improving soil structure, and increasing the reservoir of soil carbon pools. Agricultural fertilization has been established as the primary driver for augmenting soil aggregates [30].
Organic fertilizers are rich in organic matter, and the enhancement of soil organic carbon content can facilitate the formation of macro-aggregates while bolstering the physical safeguarding of organic matter [31]. The augmentation of aggregates not only serves to enhance soil structure but also contributes to the preservation of soil organic carbon [42]. Furthermore, the substantial application of organic fertilizer in organic cultivation not only resulted in an elevation of organic carbon concentration within aggregates but also stimulated the interaction between carbon and soil particles, consequently fostering the sequestration of carbon in the soil.
The involvement of soil enzymes is essential for facilitating all biochemical reactions in the soil, primarily originating from soil microorganisms, plants, soil animals, and the introduction of organic carbon into the soil. The sizable presence of labile organic carbon (LOC) and microorganisms in organic fertilizer substantially augmented enzyme activity within the soil. This study revealed a noteworthy enhancement in enzyme activity within all aggregates and bulk soil as a result of organic cultivation. However, it did not lead to any alterations in the distribution of enzyme activities among the aggregates (Figure 4 and Figure S2). The concentration of soil enzyme activity did not exhibit significant changes among aggregates with varying particle sizes in different cultivation practices. Given that small macro-aggregates constitute the major component of soil, organic cultivation exhibited a substantial increase in the proportions of small macro-aggregates and micro-aggregates. Consequently, organic cultivation led to a significant elevation in enzyme activities within the small macro-aggregates and micro-aggregates.
As catalysts for biochemical reactions, soil enzymes play a crucial role in facilitating the cycling and transformation of nutrients within the soil. The level of their activity serves as an indicator of soil fertility. In particular, small macro-aggregates exhibit a greater abundance of active nutrients and higher nutrient supply capacity. Conversely, micro-aggregates comprise a higher concentration of stabilized organic carbon. Consequently, organic cultivation enhances soil nutrient content through the promotion of small macro-aggregates, while concurrently fostering the sequestration of soil organic carbon (SOC) through the facilitation of micro-aggregates. Nevertheless, a study conducted by Jastrow et al. indicated that substantial quantities of labile organic carbon (LOC) tend to accumulate within macro-aggregates [43]. This phenomenon subsequently leads to heightened soil enzyme activity in macro-aggregates when compared to micro-aggregates. In contrast, studies conducted by Yu et al. have demonstrated that organic fertilizers exhibit a reduction in invertase activity specifically within macro-aggregates and the silt+clay fraction [44].
In the case of manured treatments, microaggregates demonstrated higher activities of β-glucosidase and phosphatase compared to macro-aggregates, as observed in the study conducted by Wang et al. [27]. The distribution of enzyme activity within aggregates is dependent on the specific type of enzyme. Carbohydrases were found to be more predominant in macro-aggregates, while phosphatase and leucine aminopeptidases were more prevalent in micro-aggregates [45]. This study revealed that the distribution of soil enzyme activity (urease, catalase, invertase, and cellulase) did not exhibit significant changes across aggregates of diverse particle sizes. This observation may be attributed to the influence of soil characteristics and the type of enzymes involved.
4.3 Organic Cultivation Effects Microbial Community Composition within Aggregate
Organic cultivation brings about alterations in the microbial community structure found within aggregates. In organic cultivation, there was an increase in the relative abundance of bacteria (Proteobacteria and Bacteroidetes) in micro-aggregates, along with an increase in the relative abundance of fungi (Mortierellomycota, Blastocladiomycota, and Basidiomycota) in small macro-aggregates (refer to Figure 5). The primary reason behind this observation is attributed to the higher nutrient levels present in organic fertilizers used in organic cultivation, thus providing increased support for microbial biomass. This notion is supported by the findings of Börjesson et al. The addition of exogenous organic fertilizers introduces a larger population of microbial communities, leading to alterations in the microbial composition within aggregates [46]. Furthermore, this introduction also induces changes in the competition and interaction among native microorganisms in the soil, ultimately resulting in transformations within the structure of the microbial community [1,2]. The shifts observed in soil microbial communities could potentially indicate variations in the chemical composition of aggregates, which can be attributed to the diverse microbial preferences for the utilized substrates [47].
Fungi demonstrate a preference for colonizing macro-aggregates and bulk soils that have been fertilized with organic matter, contributing to a greater relative abundance of fungi within macro-aggregates [48]. Furthermore, fungal hyphae play a physical role in binding smaller aggregates together, resulting in the formation of larger aggregates [49]. In contrast, bacterial communities are smaller in size and exhibit a preference for colonization within micro-aggregates. The results of this study are in accordance with previous findings, demonstrating that organic cultivation led to an increase in the relative abundance of fungi within macro-aggregates and bacteria within micro-aggregates. On one hand, the utilization of exogenous organic fertilizers resulted in an increased relative abundance of fungi within macro-aggregates and bacteria within micro-aggregates in organic cultivation. On the other hand, organic management practices such as biological control and manual weed control serve to sustain and enhance the amplified presence of microbial communities within soil aggregates.
These findings suggest that organic management practices are associated with an increased relative abundance of microorganisms within aggregates and have the potential to reconfigure microbial communities. Consequently, this can have implications for soil fertility, vegetable yield, and carbon sequestration capacity.
5. Conclusions
Long-term organic cultivation fosters soil aggregation, resulting in a noticeable elevation of small macro-aggregates and micro-aggregates by 5.95% and 10.05%, respectively, in comparison to conventional cultivation. Organic cultivation not only exhibited a significant increase in organic carbon fractions and enzyme activity within three particle size aggregates but also induced a shift in the distribution of organic carbon fractions within soil aggregates. Moreover, organic cultivation resulted in an enhanced relative abundance of bacteria within micro-aggregates and fungi within small macro-aggregates. SEM analysis revealed that organic cultivation led to an augmentation in both soil aggregate particle size and stability, thereby promoting enhanced microbial activity and enzyme activity. In conclusion, long-term organic cultivation contributes to the enhancement of soil structure through soil aggregation, leading to increased soil productivity and vegetable yield in greenhouse environments.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: The concentration of SOC, EOC, and POC in bulk soil under different cultivation treatments. Capital letters indicate significant differences under different cultivation treatments (p<0.05); Figure S2: The sucrase, cellulase, urease and catalase activities in the bulk soil under different cultivation treatments. Capital letters indicate significant differences under different cultivation treatments (p<0.05); Fig.S3: Soil bulk density (0-60 cm) in different cultivations; Table S1: Types of vegetables grown in different years in greenhouses; Table S2: Nutrient input, pesticide application and irrigation under different cultivation; Table S3: Primer sequence; Table S4: PCR reaction system (30 μL); Table S5: Reaction procedure.
Author Contributions
L.H.T. and Y.J.L.; methodology, formal analysis, Y.J.L. and X.Y.L.; investigation, Y.J.L. and X.Y.L.; writing—original draft preparation, L.H.T. and Y.J.L.; writing—review and editing, L.H.T. and T.L.; visualization, L.C.Z.; supervision, A. E.; project administration, Y.J.W.; funding acquisition, T.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Inner Mongolia Autonomous Region Department of Human Resources and Social Security, 2022 Annual Research Support Fund for the Introduction of High-level Talents in Institutions Affiliated to the Inner Mongolia Autonomous Region (DC2300001259).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tei, F.; De Neve, S.; de Haan, J.; Kristensen, H.L. Nitrogen management of vegetable crops. Agric. Water Manage. 2020, 240, 106316. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, X. Comparison of agrochemicals allocation efficiency between greenhouse and open-field vegetables in China. Sci. Rep. 2021, 11, 12807. [Google Scholar] [CrossRef] [PubMed]
- Ti, C.; Luo, Y.; Yan, X. Characteristics of nitrogen balance in open-air and greenhouse vegetable cropping systems of China. Environ. Sci. Pollut. Res. 2015, 22, 18508–18518. [Google Scholar] [CrossRef] [PubMed]
- FAO, Food and Agriculture Organization of the United Nations. 2017.
- Tripathi, S.; Srivastava, P.; Devi, R. S. Chapter 2 - Influence of synthetic fertilizers and pesticides on soil health and soil microbiology, in Agrochemicals Detection, Treatment and Remediation, M.N.V. Prasad, Editor. 2020, Butterworth-Heinemann, 25-54.
- Chen, Z.K.; Muhammad, I.; Zhang, Y.X.; Hu, W.Y.; Lu, Q.Q.; Wang, W.X.; Huang, B.; Hao, M.D. Transfer of heavy metals in fruits and vegetables grown in greenhouse cultivation systems and their health risks in Northwest China. Sci. Total Environ. 2021, 766, 142663. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Jing, Y.; Feng, Y. Vegetable cultivation under greenhouse conditions leads to rapid accumulation of nutrients, acidification and salinity of soils and groundwater contamination in South-Eastern China. Nutr. Cycling Agroecosyst. 2009, 83, 73–84. [Google Scholar] [CrossRef]
- Wang, C.N.; Wu, R.L.; Li, Y.Y.; Qin, Y.F.; Li, Y.L.; Meng, F.Q.; Wang, L.G.; Xu, F.L. Effects of pesticide residues on bacterial community diversity and structure in typical greenhouse soils with increasing cultivation years in Northern China. Sci. Total Environ. 2020, 710, 136321. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhao, X.; Meng, T. What are the driving factors of pesticide overuse in vegetable production? Evidence from Chinese farmers. China Agr Econ Rev. 2019, 11, 672–687. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Xi, X.C.; Tang, X.; Luo, D.M.; Gu, B.J.; Lam, S.K.; Vitousek, P.M.; Chen, D.L. Policy distortions, farm size, and the overuse of agricultural chemicals in China. PNAS. 2018, 115, 201806645. [Google Scholar] [CrossRef]
- Min, J.; Zhang, H.L.; Shi, W.M. Optimizing nitrogen input to reduce nitrate leaching loss in greenhouse vegetable production. Agric. Water Manage. 2012, 111, 53–59. [Google Scholar] [CrossRef]
- Gomiero, T.; Pimentel, D.; Paoletti, M.G. Environmental Impact of Different Agricultural Management Practices: Conventional vs. Organic Agriculture. Crit. Rev. Plant Sci. 2011, 30, 95–124. [Google Scholar] [CrossRef]
- Tuomisto, H.L.; Hodge, I.D.; Riordan, P.; Macdonald, D.W. Does organic farming reduce environmental impacts? – A meta-analysis of European research. J. Environ. Manage. 2012, 112, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Alagöz, E.; Yilmaz, Z. Effects of different sources of organic matter on soil aggregate formation and stability: A laboratory study on a Lithic Rhodoxeralf from Turkey. Soil Till Res. 2009, 103, 419–424. [Google Scholar] [CrossRef]
- Kamran, M.; Nie, J.; Geng, M.J.; Lu, Y.H.; Liao, Y.L.; Zhou, F.L.; Xu, Y.H. Effect of reduced mineral fertilization (NPK) combined with green manure on aggregate stability and soil organic carbon fractions in a fluvo-aquic paddy soil. Soil Till Res. 2021, 211, 105005. [Google Scholar] [CrossRef]
- King, G.M. Enhancing soil carbon storage for carbon remediation: potential contributions and constraints by microbes. Trends Microbiol. 2011, 19, 75–84. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; Papale, D.; Rammig, A.; Smith, P.; Thonicke, K.; van der Velde, M.; Vicca, S.; Walz, A.; Wattenbach, M. Climate extremes and the carbon cycle. Nature. 2013, 500, 287. [Google Scholar] [CrossRef] [PubMed]
- Riestra, D.; Noellemeyer, E.; Quiroga, A. Soil Texture and Forest Species Condition the Effect of Afforestation on Soil Quality Parameters. Soil Sci. 2012, 177, 279–287. [Google Scholar] [CrossRef]
- Purakayastha, T.J.; Rudrappa, L.; Singh, D.; Swarup, A.; Bhadraray, S. Long-term impact of fertilizers on soil organic carbon pools and sequestration rates in maize–wheat–cowpea cropping system. Geoderma. 2008, 144, 370–378. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Álvaro-Fuentes, J.; Cantero-Martínez, C. Identifying soil organic carbon fractions sensitive to agricultural management practices. Soil Till Res. 2014, 139, 19–22. [Google Scholar] [CrossRef]
- Thorburn, P.J. , et al., Changes in soil carbon sequestration, fractionation and soil fertility in response to sugarcane residue retention are site-specific. Soil Till Res. 2012, 120, 99–111. [Google Scholar] [CrossRef]
- Aulakh, C.S.; Sharma, S.; Thakur, M.; Kaur, P. A review of the influences of organic farming on soil quality, crop productivity and produce quality. J. Plant Nutr. 2022, 45, 1–22. [Google Scholar] [CrossRef]
- Gömöryová, E.; Strelcová, K.; Fleischer, P.; Gömöry, D. Soil microbial characteristics at the monitoring plots on windthrow areas of the Tatra National Park (Slovakia): their assessment as environmental indicators. Environ. Monit. Assess. 2011, 174, 31–45. [Google Scholar] [CrossRef]
- Young, I.M.; Crawford, J.W.; Nunan, N.; Otten, W.; Spiers, A. Chapter 4- Microbial Distribution in Soils: Physics and Scaling, in Advances in Agronomy, Academic Press, 2008, 81-121.
- Rillig, M.C.; Muller, L.A.H.; Lehmann, A. Soil aggregates as massively concurrent evolutionary incubators. ISME J. 2017, 11, 1943–1948. [Google Scholar] [CrossRef]
- Tong, L.H.; Zhu, L.; Lv, Y.H.; Zhu, K.; Liu, X.Y.; Zhao, R. Response of organic carbon fractions and microbial community composition of soil aggregates to long-term fertilizations in an intensive greenhouse system. J. Soils Sediments. 2020, 20, 641–652. [Google Scholar] [CrossRef]
- Wang, R.; Liu, J.; Xu, J. ChemInform Abstract: Organocatalytic Enantioselective Sulfur-Michael Addition of Thioacetic Acid to Arylmethylidenemalonates. Adv Synth Catal. 2014, 357, 1. [Google Scholar] [CrossRef]
- Chu, H.J.; Wang, S.P.; Yue, H.W.; Lin, Q.Y.; Hu, Y.G.; Li, X.Z.; Zhou, J.Z.; Yang, Y.F. Contrasting soil microbial community functional structures in two major landscapes of the Tibetan alpine meadow. MicrobiologyOpen. 2014, 3, 585–594. [Google Scholar] [CrossRef]
- Allison, S.D.; Jastrow, J.D. Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol. Biochem. 2006, 38, 3245–3256. [Google Scholar] [CrossRef]
- Abiven, S.; Menasseri, S.; Chenu, C. The effect of organic inputs over time on soil aggregate stability-A literature analysis. Soil Biol. Biochem. 2009, 41, 1–12. [Google Scholar] [CrossRef]
- Jiang, Y.; Sun, B.; Jin, C.; Wang, F. Soil aggregate stratification of nematodes and microbial communities affects the metabolic quotient in an acid soil. Soil Biol. Biochem. 2013, 60, 1–9. [Google Scholar] [CrossRef]
- Fattet, M.; Fu, Y.; Ghestem, M.; Ma, W.; Foulonneau, M.; Nespoulous, J.; Le Bissonnais, Y.; Stokes, A. Effects of vegetation type on soil resistance to erosion: Relationship between aggregate stability and shear strength. Catena. 2011, 87, 60–69. [Google Scholar] [CrossRef]
- Cambardella, C.A.N.S.T.L.; Ames, I.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci Soc Am J. 1992, 56, 3. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics. 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.; Bahram, M.; Bates, S.; Bruns, T.; Bengtsson-Palme, J.; Callaghan, T.; Douglas, B.; Drenkhan, T.; Eberhardt, U.; Dueñas, M.; Grebenc, T.; Griffith, G.W.; Hartmann, M.; Kirk, P.; Kohout, P.; Larsson, K.H. Towards a unified paradigm for sequence-based identification of Fungi. Mol Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Li, S.S.; Wang, M.; Zhao, Z.Q.; Li, X.Y.; Han, Y.; Chen, S.B. Alleviation of cadmium phytotoxicity to wheat is associated with Cd re-distribution in soil aggregates as affected by amendments. RSC Adv. 2018, 8, 17426–17434. [Google Scholar] [CrossRef]
- Maillard, É.; Angers, D.A. Animal manure application and soil organic carbon stocks: a meta-analysis. Global Change Biol. 2014, 20, 666–679. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, N.; Ge, T.; Kuzyakov, Y.; Wang, Z. L.; Li, Z.; Tang, Z.; Chen, Y.; Wu, C.; Lou, Y. Soil aggregation regulates distributions of carbon, microbial community and enzyme activities after 23-year manure amendment. Appl Soil Ecol. 2017, 111, 65–72. [Google Scholar] [CrossRef]
- Pimentel, D.; Hepperly, P.; Hanson, J.; Douds, D.; Seidel, R. Environmental, Energetic, and Economic Comparisons of Organic and Conventional Farming Systems. J. Biosci. 2005, 55, 573–582. [Google Scholar] [CrossRef]
- Xie, J.; Hou, M.; Zhou, Y.; Wang, R.; Zhang, S.; Yang, X.; Sun, B. Carbon sequestration and mineralization of aggregate-associated carbon in an intensively cultivated Anthrosol in north China as affected by long term fertilization. Geoderma. 2017, 296, 1–9. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Global Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science. 2004, 304(5677), 1623–1627. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim. Change. 2007, 80, 5–23. [Google Scholar] [CrossRef]
- Yu, H.; Ding, W.; Luo, J.; Geng, R.; Cai, Z. Long-term application of organic manure and mineral fertilizers on aggregation and aggregate-associated carbon in a sandy loam soil. Soil Till Res. 2012, 124, 170–177. [Google Scholar] [CrossRef]
- Marx, M.C.; Kandeler, E.; Wood, M.; Wermbter, N.; Jarvis, S.C. Exploring the enzymatic landscape: distribution and kinetics of hydrolytic enzymes in soil particle-size fractions. Soil Biol. Biochem. 2005, 37, 35–48. [Google Scholar] [CrossRef]
- Börjesson, G.; Menichetti, L.; Kirchmann, H.; Kätterer, T. Soil microbial community structure affected by 53 years of nitrogen fertilisation and different organic amendments. Biol. Fertil. 2012, 48, 245–257. [Google Scholar] [CrossRef]
- Fanin, N.; Hättenschwiler, S.; Fromin, N. Litter fingerprint on microbial biomass, activity, and community structure in the underlying soil. Plant Soil. 2014, 379, 79–91. [Google Scholar] [CrossRef]
- Huygens, D.; Denef, K.; Vandeweyer, R.; Godoy, R.; Van Cleemput, O.; Boeckx, P. Do nitrogen isotope patterns reflect microbial colonization of soil organic matter fractions? Biol. Fertil. 2008, 44, 955–964. [Google Scholar] [CrossRef]
- Duchicela, J.; Sullivan, T.S.; Bontti, E.; Bever, J.D. Soil aggregate stability increase is strongly related to fungal community succession along an abandoned agricultural field chronosequence in the Bolivian Altiplano. J Appl Ecol. 2013, 50, 1266–1273. [Google Scholar] [CrossRef]
Figure 1.
Location of the experimental site and the picture of a greenhouse for organic cultivation.
Figure 1.
Location of the experimental site and the picture of a greenhouse for organic cultivation.

Figure 2.
Soil aggregate distribution, mean weight diameter of aggregates under different cultivation treatments. Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
Figure 2.
Soil aggregate distribution, mean weight diameter of aggregates under different cultivation treatments. Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
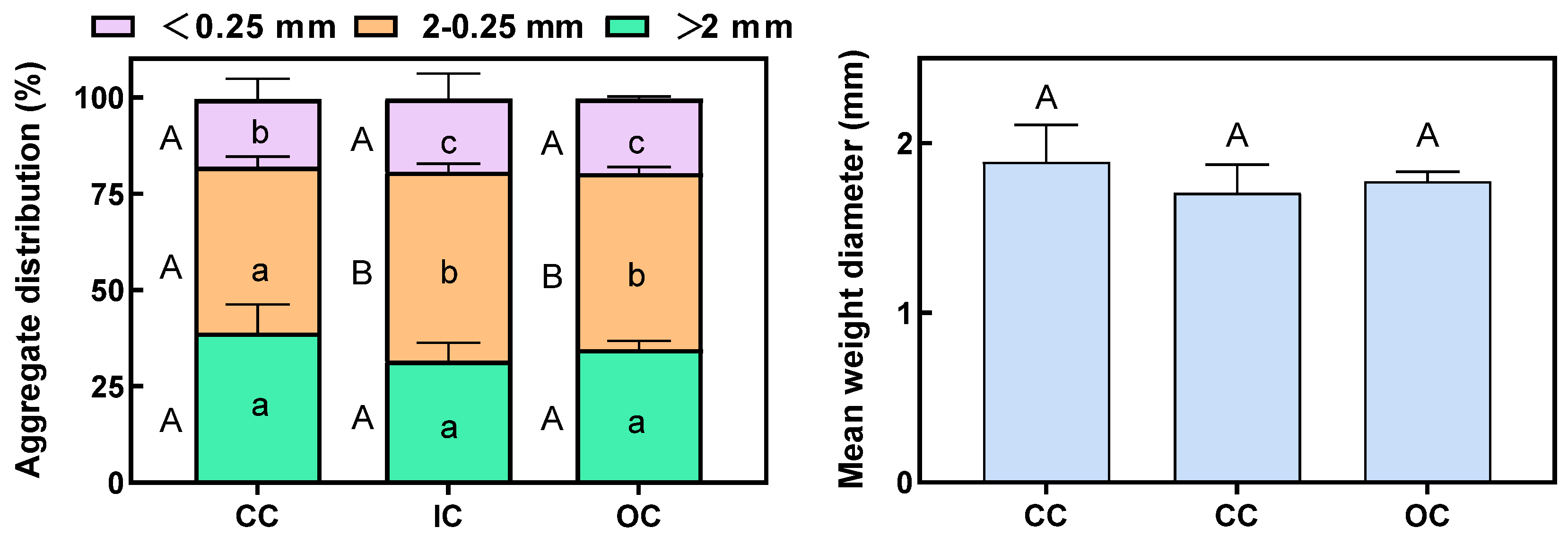
Figure 3.
The concentration of SOC, EOC, and POC in three size classes of soil aggregate under different cultivation treatments. The values denote the means of three replicates (±SE). Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
Figure 3.
The concentration of SOC, EOC, and POC in three size classes of soil aggregate under different cultivation treatments. The values denote the means of three replicates (±SE). Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
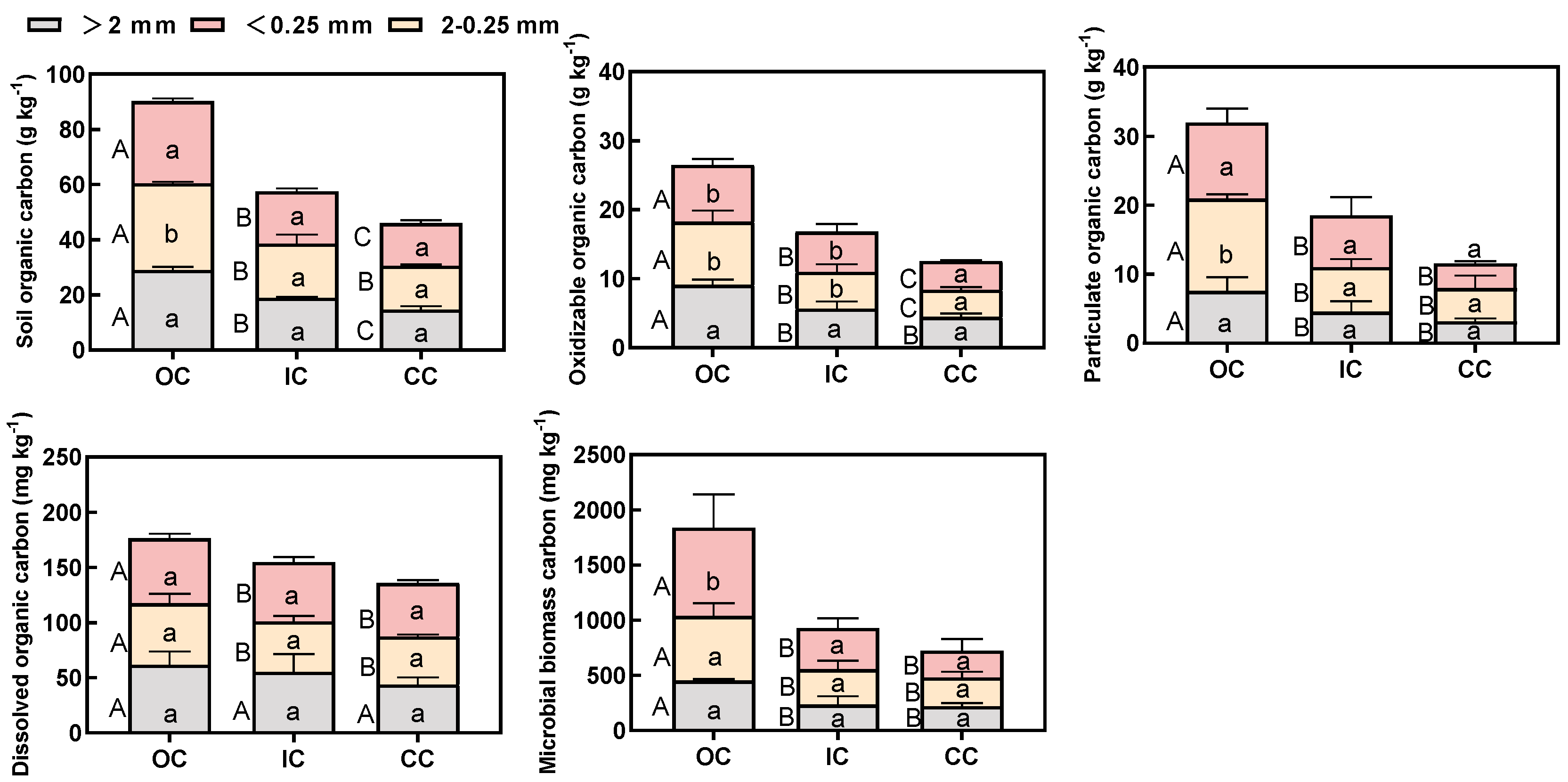
Figure 4.
The sucrase, cellulase, urease and catalase activities of three soil aggregate-size classes under different cultivation treatments. Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
Figure 4.
The sucrase, cellulase, urease and catalase activities of three soil aggregate-size classes under different cultivation treatments. Capital letters indicate significant differences among different cultivation treatments for each of the aggregate size classes (p<0.05), while lower letters indicate significant differences among aggregate size fractions in the same cultivation treatments (p<0.05).
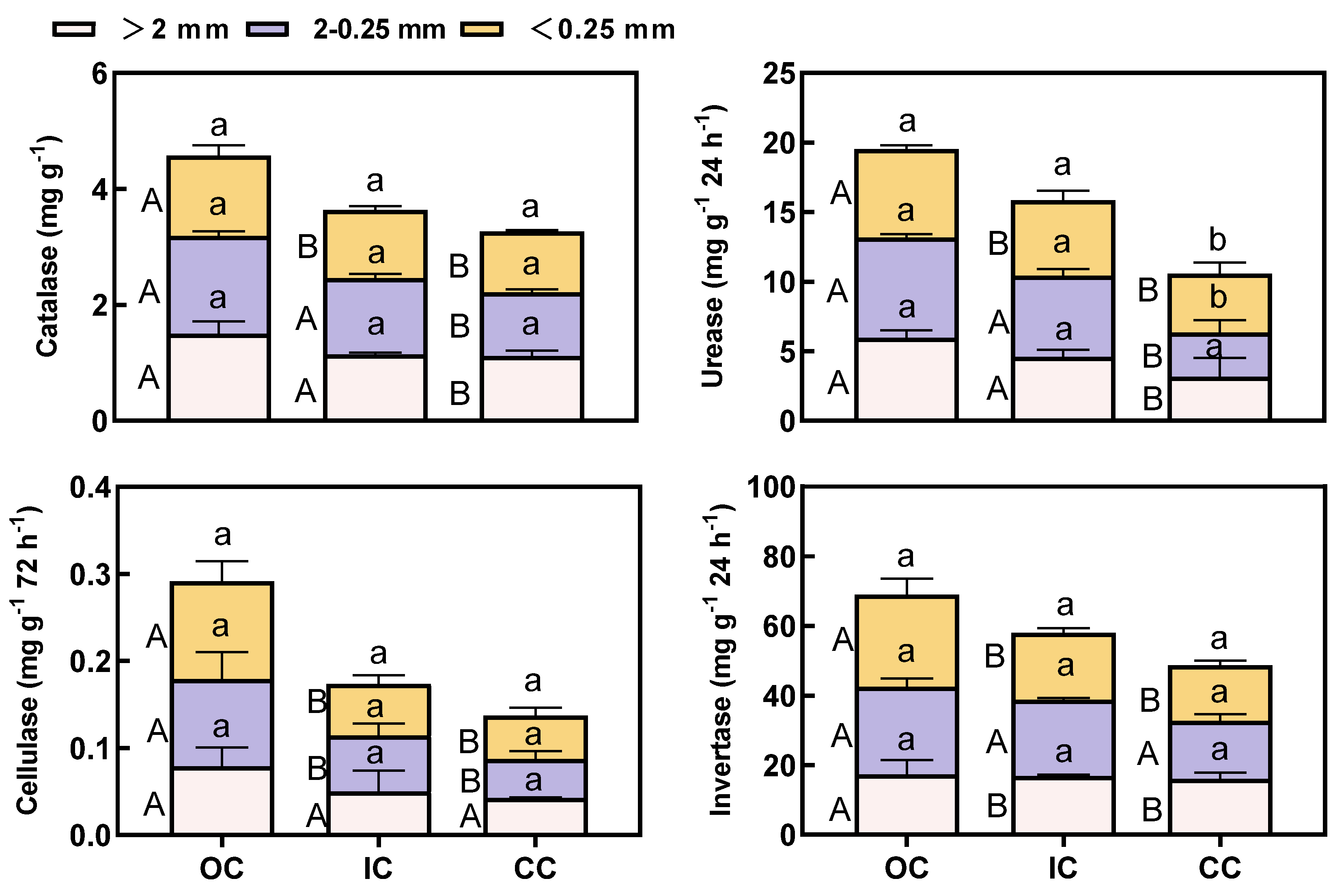
Figure 5.
Heatmap of bacteria (a) and fungi (b) at phylum level in different particle sizes of aggregates under different cultivation treatments.
Figure 5.
Heatmap of bacteria (a) and fungi (b) at phylum level in different particle sizes of aggregates under different cultivation treatments.
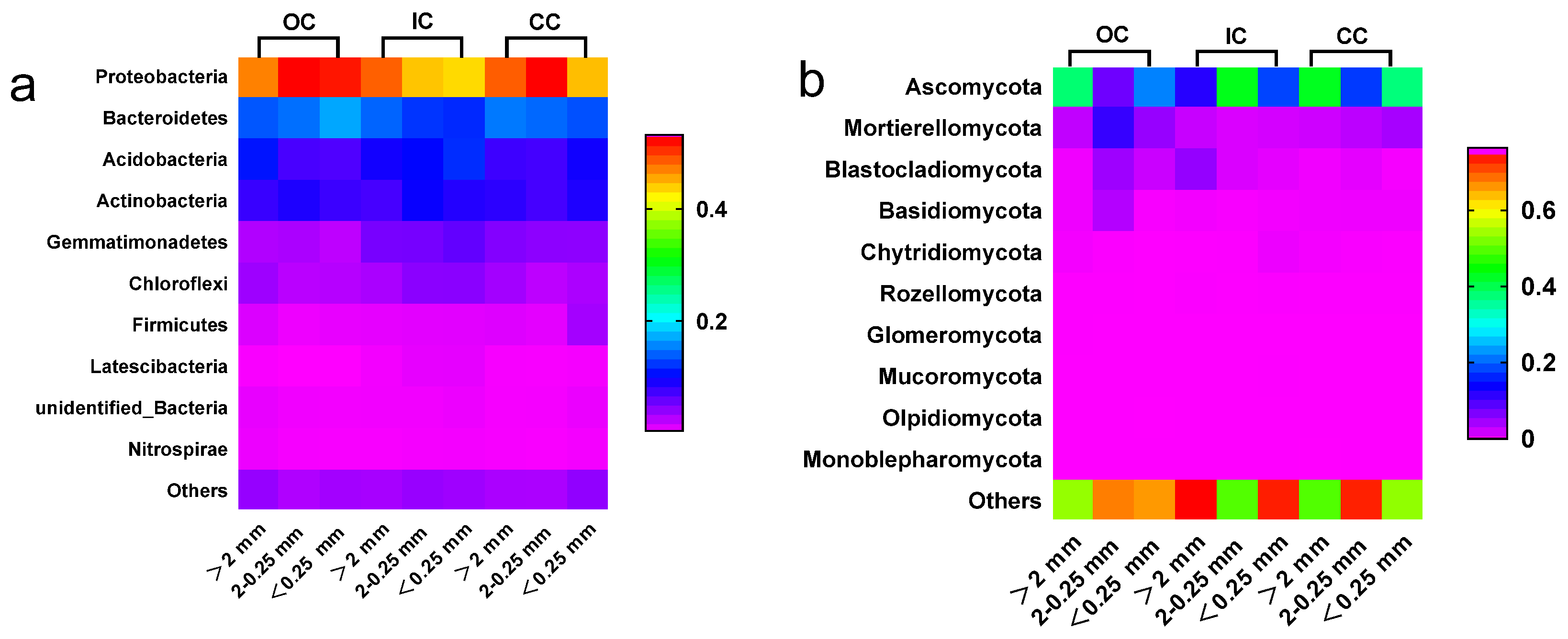
Figure 6.
Vegetable yield in spring and autumn during CC, IC and OC treatments. OC (Organic cultivation); IC (integrated cultivation); and CC (Conventional cultivation). The values denote the means of three replicates (±SE).
Figure 6.
Vegetable yield in spring and autumn during CC, IC and OC treatments. OC (Organic cultivation); IC (integrated cultivation); and CC (Conventional cultivation). The values denote the means of three replicates (±SE).
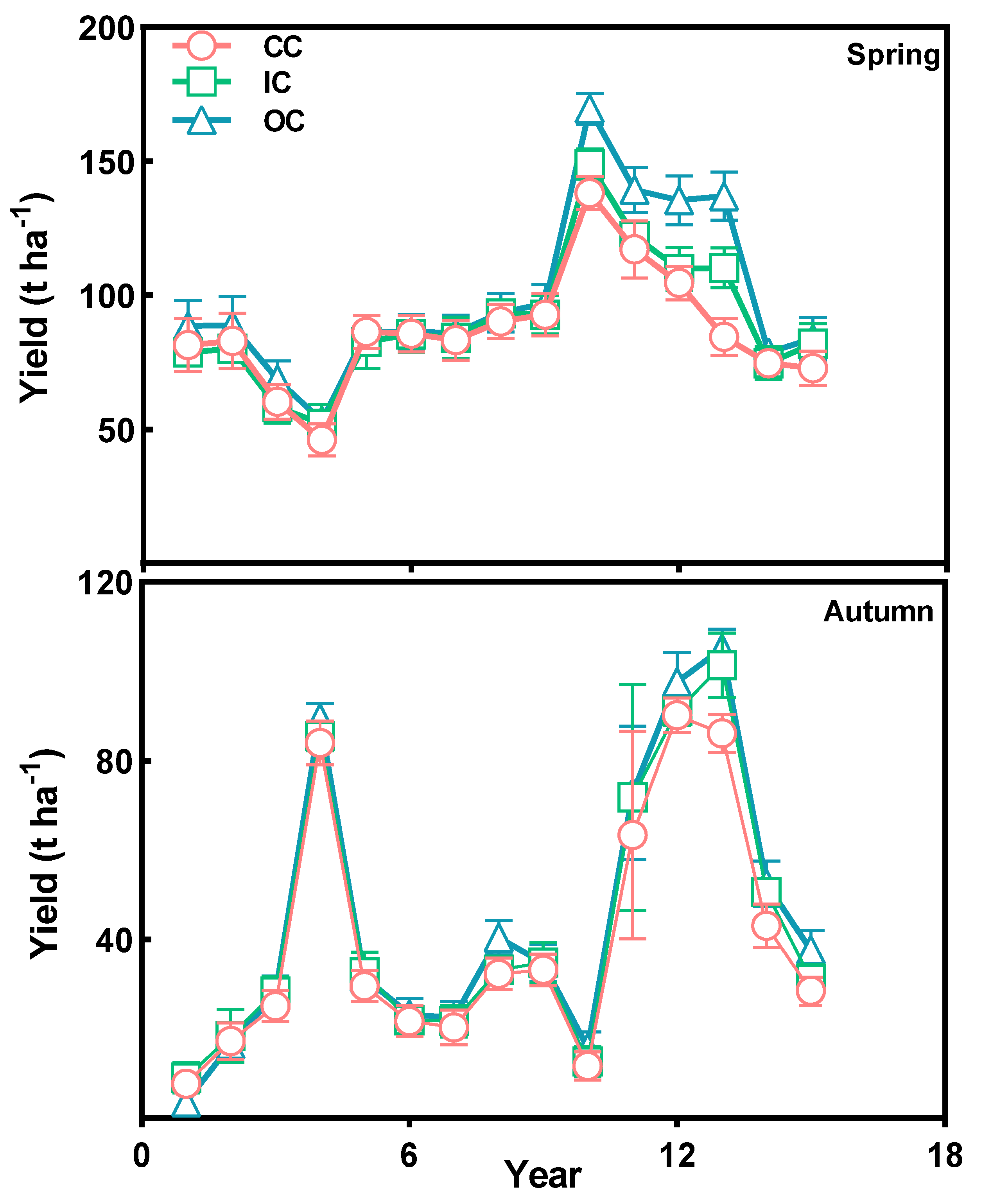
Figure 7.
SEM showing that different cultivation treatments affect soil aggregates, soil organic fractions, microbial diversity, enzyme activities and vegetable yield: (a) CC, (b) IC and (c) OC. Red lines indicate a negative association, and blue lines indicate a positive association (*, p < 0.05; **, p < 0.01; ***, p < 0.001). The numbers reflect the normalized path coefficients.
Figure 7.
SEM showing that different cultivation treatments affect soil aggregates, soil organic fractions, microbial diversity, enzyme activities and vegetable yield: (a) CC, (b) IC and (c) OC. Red lines indicate a negative association, and blue lines indicate a positive association (*, p < 0.05; **, p < 0.01; ***, p < 0.001). The numbers reflect the normalized path coefficients.
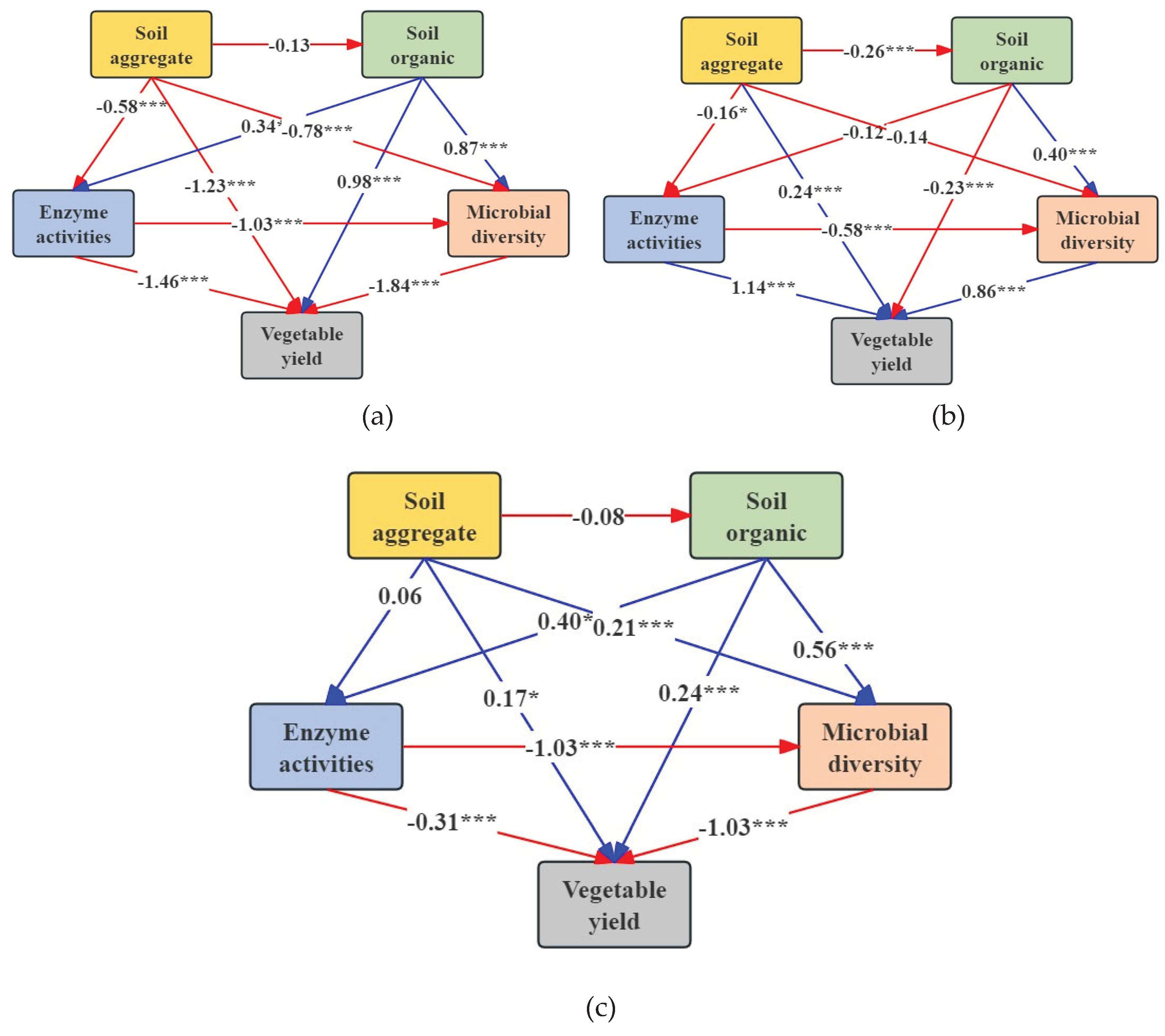

Table 1.
The physicochemical properties of top soil (0-20 cm) under different cultivation in 2002.
| Treatments | SOC g kg-1 |
Total N g kg-1 |
Total P g kg-1 |
Available N mg kg-1 |
Available P mg kg-1 |
Available K mg kg-1 |
|---|---|---|---|---|---|---|
| OC | 11.08 | 1.17 | 1.38 | 101.28 | 139.13 | 257.30 |
| IC | 9.60 | 1.19 | 1.24 | 95.35 | 81.68 | 364.28 |
| CC | 12.18 | 1.36 | 2.22 | 128.38 | 163.05 | 212.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



