
Submitted:
26 March 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
Study of alterations in Osteoarthritis (OA) of Glypicans 1 to 6 (GPCs) and Notum in plasma, bone marrow mesenchymal stromal cells (BM-MSCs) and osteoblasts. Plasma levels of GPCs and Notum were measured of 25 patients and 24 healthy subjects. In addition, BM-MSCs from eight OA patients and eight healthy donors were cultured during 21 days with culture me-dium and osteogenic medium. Protein and gene expression levels of GPCs and Notum were de-termined by ELISA and qPCR at 0, 7 ,14 and 21 days. GPC5 and Notum have been found de-creased in OA patients’ plasma. OA BM-MSCs showed downexpression of GPC6 and upregula-tion of Notum. A decrease in GPC5 and Notum proteins and an increase in GPC3 were found. During osteogenic differentiation, elevated GPC2, 4, 5, 6 and Notum mRNA levels and decreased GPC3 were observed in OA. Furthermore, the protein levels of GPC2, GPC5 and Notum de-creased and GPC3 increased. Glypicans and Notum are altered in BM-MSCs and during osteo-genic differentiation from patients with OA. Furthermore, GPC5 and Notum can be proposed as new biomarkers of the pathology. The alterations found indicate that Notum could be a prom-ising therapeutic strategy for Osteoarthritis.
Keywords:
Osteoarthritis
; Bone Marrow Mesenchymal Stem Cells
; Osteoblast
; Glypican
; NOTUM
1. Introduction
In the global context, Osteoarthritis (OA) shows a prevalence with a growing dynamic related to the increase in risk factors such as obesity and greater aging of the population [1]. This joint disease affects the articular cartilage, subchondral bone, ligaments and synovial membrane [2]. Among the most characteristic symptoms of OA are pain, stiffness, reduced movement, swelling and joint crepitation [3].
Currently, there are no effective medications to treat Osteoarthritis. The most commonly used pharmacological option are analgesics to relieve pain and, in more advanced cases, joint replacement surgery [4]. Now a days, OA is one of the priority challenges for health systems. Given the high prevalence and disability of patients with osteoarthritis, it is essential to find drugs that can minimize the effects of OA.
Mesenchymal stem cells (MSCs) are cells that have the ability to differentiate into cells of the mesodermal lineage, such as chondrocyte, osteocytes and adipocytes. In adults, one of the areas where MSCs reside is in the bone marrow and can be obtained after in vitro proliferation [5]. In the joint, there are mainly two cell types that come from MSCs, chondrocytes and osteoblasts. In mesenchymal stem cells from bone marrow of patients with Osteoarthritis compared to healthy donors, it has been proven that the osteogenic potential is increased and the chondrogenic and adipogenic potential is attenuated. MSCs will differentiate preferentially into osteoblasts and a significant reduction into chondrocytes, causing a breakdown in homeostasis between osteogenesis and chondrogenesis. These facts implicate MSCs in the increase in bone mass and cartilage loss that characterizes OA [6]. Along these lines, several studies focus on alterations in bone metabolism as a direct contribution to OA and that influence cartilage injuries. Several signaling pathways, growth factors, cytokines and other components related to bone formation and maintenance have been found altered. This evidence, together with the fact that abnormalities in the activity of the subchondral bone are found in the early phase of OA, open the possibility that a pharmacological intervention aimed at regulating bone metabolism can improve or preserve the joint structure. In fact, new therapeutic proposals for Osteoarthritis focus on bone remodeling with promising results [7].
One of the important signaling pathways in bone formation and maintenance is the Wnt/β-Catenin pathway. Wnts are essential in the process of differentiation, proliferation and synthesis of bone matrix by osteoblasts [8]. The Wnt/β-catenin signalling pathway is a normal physiological response to mechanical loading in bone [7]. There is several evidence that the Wnt signaling pathway determines the cell fate of mesenchymal cells, as it favors the formation of osteoblasts and inhibits the formation of chondrocytes and adipocytes. In addition, it maintains the balance between bone formation and bone resorption [9].
Several evidence demonstrate that the Wnt/β-Catenin signaling pathway is involved in the pathology of Osteoarthritis. An accumulation of β-catenin has been found, which would point to excessive activation of this signaling pathway [10]. In in vitro assays, it has been proven that during osteogenesis there are alterations in the expression of components of the Wnt pathway [11]. Messenger RNA levels of WISP-1, a gene associated with the Wnt/β-Catenin pathway, are overexpressed during OA and are capable of inducing cartilage damage [12]. Blockade of canonical Wnt signaling prevents hypertrophic differentiation of chondrocytes [13]. All these findings show that alterations related to this signaling pathway contribute to the damage and loss of articular cartilage and the stimulation of bone formation in OA.
In the stimulation of the Wnt pathway, proteoglycans and, specifically, glypicans play a fundamental role. Glypicans are an important family of heparan sulfate proteoglycans that are attached to the outer surface of the plasma membrane via a glycosylphosphatidylinositol (GPI) linkage. This protein family contains six members (GPC1 to 6) in the human genome that share a characteristic domain of 14 conserved cysteine residues. Due to similarity, glypicans are divided into two subfamilies: GPC1, 2, 4 and 6 (with 35-63% homologous sequence) and GPC3 and GPC5 (54% homologous). The primary function of membrane-bound glypicans is to regulate the signaling of Wnts, Hedgehogs (Hhs), fibroblast growth factors (FGFs), and bone morphogenetic proteins (BMPs). The stimulatory mechanism is based on the ability of glypicans to facilitate and/or stabilize the interaction of Wnts with their signaling receptors. In addition to being anchored to the cell membrane, glypicans can be found in the extracellular environment. In this way they do not interact for the activation of the Wnt pathway, which is why they are considered inhibitory competitors [14,15]. In any case, more studies are needed to understand in depth the mechanisms carried out by each of the glypicans, both those bound to the membrane and those found in soluble form.
Many evidences suggest that the glypican family of proteins is involved in the regulation of bone metabolism [15]. GPC1 has been found to be highly expressed in bone marrow inducing fibroblast phenotype in Bone Marrow MSCs when it is inhibited [16]. On the other hand, GPC3 has been proposed along with Wnt/ β-Catenin signaling pathway to form Wnt/frizzled/β-catenin complexes, which inhibit β-catenin degradation, leading to osteosarcoma progression [17]. GPC1 and GPC3 have been described in bibliography as regulators of the response of primary human suture MSCs to bone morphogenetic protein [18]. Deletion in zebrafish of GPC4 homolog (knypek) results in craniofacial skeletal defects as well as pathogenic variants in GPC4 results in Keipert Syndrome in human people [19,20]. Finally, GPC6 has also been related with Wnt/ β-Catenin signaling pathway and specifically with osteoporosis. More than 150 specific loci have been found in GPC6 gene which could be responsible for bone mineral content. This found, proposed GPC6 as a novel drug target for osteoporosis care [21,22].
Until relatively few years ago, it was proposed that Glypicans were released from the membrane by Notum. Notum is primarily a negative feedback enzyme of Wnt signaling. For this reason, it was believed that Notum could act as a phospholipase enzyme and cleave the GPI anchor, which keeps Glypicans attached to the cell membrane [23]. However, a recent study by Kakugawa et al. studied the role of Notum in depth and attributed other functions to it [24]. As they demonstrated, Notum is a carboxylesterase that eliminates an essential palmitoleate fraction from Wnts and would deactivate the signaling pathway. For their part, Glypicans would help direct Notum to bind with Wnts since a glycosaminoglycan binding site was found in Notum.
All these facts reveal the importance that the Wnt/β-Catenin signaling pathway and the intermediates that are generated can have in the disease of Osteoarthritis. Pharmacological strategies to inhibit the Wnt pathway have been tested in mouse models of traumatic OA. Anticatabolic effects on chondrocytes and antifibrotic effects on synovial fibroblasts were achieved, mitigating the effects of the disease [25]. Another inhibitor that has been proposed as potentially disease-modifying OA drugs (DMOADs) is the molecule SM04690. In vitro and in vivo studies show that SM04690 manages to induce differentiation into chondrocytes [26]. Although the results are promising, pharmacologically altering the Wnt/b-catenin signaling pathway may entail several risks because it is involved in numerous processes. Therapies focused on modulating components associated with the Wnt pathway could be safer and have better results. However, in the context of Osteoarthritis, the possible alterations of the intermediates of the Wnt pathway have not been studied.
Our study aims to elucidate for the first time the dysregulations that may exist in glypicans 1 to 6 and Notum in Osteoarthritis patients at the cellular and circulating level. To do this, bone marrow mesenchymal stem cells from patients diagnosed with Osteoarthritis and healthy patients will be studied in vitro. In addition to studying the BM-MSCs, osteogenic differentiation will be induced to study if there are alterations in the osteoblasts. In this way we can evaluate whether the alterations are primarily intracellular, extracellular or both. Furthermore, it will be possible to verify whether these alterations come from mesenchymal stem cells or occur during osteogenic differentiation. With the dysregulations found intracellularly and, especially extracellularly, the aim is to propose strategies to return homeostasis to the joint. The altered extracellular levels of Glypicans 1 to 6 and Notum could be reflected in the circulation of patients, therefore, plasma levels of OA will be studied compared to controls
2. Materials and Methods
Patients
Written informed consent was obtained from all subject before sample collection. The study protocol was approved by the institutional ethics committee (Comité Ético de Investigacion Clínica Hospital Clinico San Carlos—Madrid, Spain) in concordance with the principles of the Declaration of Helsinki.
Plasma
Twenty-five patients with OA were recruited according to the American College of Rheumatology (ACR) criteria and Twenty-four control subjects belonging to the biobank of our institution. Blood samples were collected in K2-EDTA tubes, centrifuged at 2000g for 15 minutes and aliquoted plasma stored at -80°C.
Bone Marrow Mesenchymal Stromal Cells Isolation
BM-MSCs were prepared from bone marrow aspirates of eight Osteoarthritis patients undergoing joint replacement and eight healthy donors with subcapital hip fractures. BM-MSCs were isolated by density gradient (Ficoll-Paque, GE Healthcare). Subsequently, BM-MSCs were expanded in DMEM with Glutamax (Gibco, 21885), 10% FBS and 1% Pen/Strep (Gibco), from now on referred to as “culture medium”. Obtaining MSCs is confirmed by flow cytometry and histochemistry assays [11].
Cell Culture
Mesenchymal stem cells are cultured in 6-well plates. After 24 hours in culture medium, MSCs and the supernatant are collected from one of the wells as a basal value t=0. Part of the BM-MSCs were continued to be cultured in culture medium. The other part of the BM-MSCs, osteogenic inducing medium (StemMACS Osteodiff Media Human, 130-091-678, Mitenyi Biotec) with 1% Pen/Strep (Gibco) was added. Cells in both conditions were cultured for 21 days. The supernatant was collected at 7, 14 and 21 days. BM-MSCs and osteoblasts were harvested and preserved in RNAlater (Invitrogen) on the same days. The supernatant and cells were stored at -80ºC.
Determination of Glypicans and Notum in Plasma and Cell Culture Supernatants
Glypicans and Notum levels were determined using comercial kits and following the manufacturer´s instruction (Human Glypican 1 ELISA kit, ref. EH222RB, Invitrogen; Human Glypican 2 ELISA kit, ref. E-EL-H1711, Elabscience; Human Glypican 3 ELISA kit, ref. EH223RB, Invitrogen; Human Glypican 4 ELISA kit, ref. E-EL-H1713, Elabscience; Human Glypican 5 ELISA kit, ref. EH224RB, Invitrogen; Human Glypican 6 ELISA kit, Ref. CSB-EL009708HU, CUSABIO; Human NOTUM ELISA kit, ref. EH1816, FineTest). The absorbance was measured at 450nm (Heales MB-580, Shenzhen Heales Technology Development Co. Ltd.).
mRNA Analysis
RNA isolation cells was performed using the Speedtools Total RNA Extraction Kit (Biotools) and concentration was measured using a spectrophotometer (NanoDrop One, Thermo Scientific). To obtain cDNA, the Maxima First Strand cDNA Synthesis Kit for RT-qPCR (ThermoScientific) was used. The TaqMan® Universal PCR Master Mix kit (Applied Biosystems) was used to perform the qPCR on 7500 Fast Real-Time PCR System (Applied Biosystems).
Expression gene was evaluated of GPC1 (Hs00892476_m1), GPC2 (Hs00415099_m1), GPC3 (Hs01018936_m1), GPC4 (Hs00155059_m1), GPC5 (Hs00942154_m1), GPC6 (HS01569271_m1), NOTUM (Hs00394510_m1). Relative gene expression was normalized to housekeeping genes β-actin (Hs99999903_m1) and 18S (Hs99999901_s1). The data were calculated with the method of fold change (2-ΔΔCt).
Statistical Analysis
All experiments were analyzed with the GraphPad Prism 8.0. For the detection of outliers, the ROUT method (Q = 1%) was used. If the data follow a normal distribution, they were analyzed with t- test; when the data did not follow a normal distribution, the non-parametric Mann-Whitney test was performed. Significance level p<0.05 were considered significant. Biochemical data are expressed as mean±SEM.
3. Results
3.1. Circulating Levels of Glypicans and Notum
3.1.1. Demographics Characteristics of the Study Population
To study the levels of Glypicans and Notum in plasma, 25 patients with Osteoarthritis and 24 healthy donors were recruited. The mean age (± SD) ranged between 61.56 (± 7.70) in OA patients and 58.21 (± 4.40) in healthy donors. The age between groups was not statistically different when evaluated with T-test (p= 0.0692).
Among OA patients, 22 were women and 3 men and in healthy donors they were 19 and 5, respectively. There was no difference between the groups regarding gender when the Chi-square test was applied (p= 0.4635).
3.1.2. Plasma Levels in Controls and Patients with Osteoarthritis
Patients with OA showed a significant decreased in plasma levels of GPC5 (p= 0.0208) and Notum (p= 0.0260) compared to healthy controls. No differences were found between the two groups in the rest of the Glypicans. (Figure 1).
3.2. BM-MSCs and Osteoblasts
Five women and three men were recruited from each group, with a mean age of 72,50 ± 8,04 in patients with OA and 69,50 ± 10,30 in controls. The two groups did not differ in age when evaluated with t-test (p= 0.5264).
3.2.1. Expression Gene MSCs and during Osteogenic Differentiation
Our results showed a decrease in the OA group compared to the control group of GPC6 (p= 0.0471) and an increase of NOTUM (p= 0.0156) at 7 days of BM-MSC culture. When inducing osteogenic differentiation, upregulation was observed in the mRNA levels of OA patients in GPC2 (t= 14d; p= 0.034), GPC4 (t= 14d, p= 0.025; t= 21d, p= 0.017), GPC5 (t= 14d, p= 0.007), GPC6 (t= 14d, p= 0.013) and Notum (t= 14d, p= 0.009). During the 21 days of differentiation, GPC3 showed a downregulation in patients with OA compared to the control group (t= 7d, p= 0.009; t=14d, p= 0.045; t= 21d, p= 0.036) (Figure 2).
3.2.2. Protein Levels in Supernatant
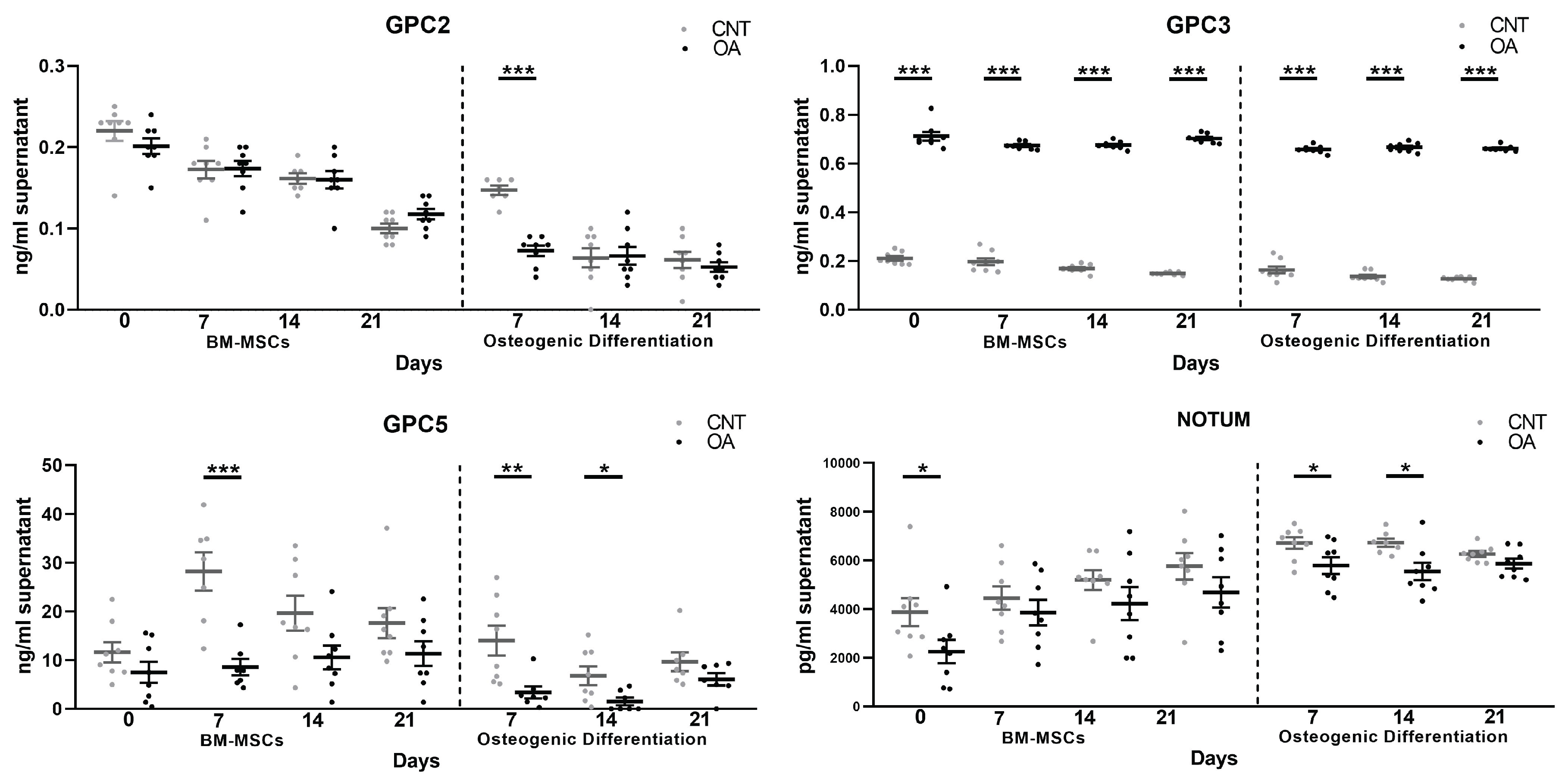
In the BM-MSCs of the OA group compared to the control group, GPC3 protein levels were higher during the 21 days of culture (t= 0, 7, 14, 21, p<0.0001). In addition, lower protein levels were found in GPC5 (t= 7d, p= 0.0006) and Notum (t= 0, p= 0.0497).
During all osteogenic differentiation, an increase in GPC3 was found (p<0.0001). Furthermore, low levels of GPC2 (t= 7d, p<0.0001), GPC5 (t=7d, p=0.0089; t=14d, p=0.0311) and NOTUM (t=7, p=0.0432; t=14d, p=0.0136) were observed. Soluble form of GPC1, GPC4 and GPC6 were not found in any of the cell cultures. (Figure 3)
4. Discussion
Globally, Osteoarthritis is a leading cause of disability, with a dynamically increasing prevalence due to an aging population and risk factors increase. This degenerative joint disease is characterized by the loss of articular cartilage and a progressive thickening of the subchondral bone. Current treatments are mainly based on anti-inflammatory drugs to relieve symptoms and joint replacement surgery in cases where it disables patients' daily lives [4]. Therefore, to find an effective treatment is one of the main objectives in the health system today.
Therapeutic interventions for Osteoarthritis have focused especially on cartilage degradation. However, in recent years there is increasing evidence that alterations in the subchondral bone in OA have an integral role in the disease process [7]. Among the molecular alterations involved in OA and related to bone formation, an excessive activation of the Wnt/β-catenin signaling pathway has been found [27]. Activation of this pathway promotes osteogenic differentiation preferentially, inhibiting chondrogenic differentiation [9]. This provides an explanation why increased bone mass and cartilage loss are found in OA. Mediators related to the canonical Wnt/β-catenin signaling pathway promote activation and deactivation of the pathway [15]. Despite their importance, alterations in some mediators related to the Wnt signaling pathway in the context of Osteoarthritis are still unknown. By expanding the knowledge of deregulations in bone metabolism in OA, treatments can be proposed to restore homeostasis in this disease, modulating the active osteogenic metabolism and, as a possible consequence, the regulation of chondrogenic metabolism. Therefore, the objective of this study was to explore for the first time the levels of Glypicans and Notum, modulators of the Wnt pathway, in patients with OA compared to healthy patients. These molecules will be studied in patient plasma, BM-MSCs and during osteogenic differentiation.
Our results show lower circulating levels of Glypican 5 and Notum in patients with OA. These results agree with the protein levels studied in cells. In BM-MSCs, both Glypican 5 and Notum are found in lower amounts in OA compared to cells from healthy patients. During osteogenic differentiation, it is found that the levels are also decreased. This demonstrates that the deregulation of these two components at the cellular level can be confirmed in patient plasma. For this reason, Glypican 5 and Notum could be promising candidates as biomarkers of osteoarthritis.
On the other hand, glypicans in the extracellular matrix are negative regulators of the Wnt β-Catenin signaling pathway. Notum also contributes as a negative regulator of the Wnt pathway [23]. Our findings on the soluble form of Glypican 5 and Notum are very important in this sense. The alterations found are intrinsic to the BM-MSCs of patients with osteoarthritis, since these two molecules are decreased. It is possible that this alteration contributes to the activation of the Wnt β-Catenin signaling pathway found in OA.
Interestingly, Glypican 3, homologous to Glypican 5, showed an increase at the protein level throughout the culture of the mesenchymal cells. In a study with GPC3 knockout mice it led to inhibition of the non-canonical Wnt/JNK signaling pathway. However, it caused the activation of canonical Wnt/β-Catenin signaling [28]. Although it has been proposed that membrane-bound GPC3 is closely related to the activation of the canonical Wnt/β-Catenin signaling pathway [29], it is likely that the complete lack of Glypican 3 activates other processes so that the canonical pathway remains active. The excessive increase in soluble Glypican 3 found in our study, together with a decreased production of mRNA, indicates that Glypican 3 may not be practically membrane-bound and is contributing to the activation of the canonical Wnt pathway.
There is some evidence that Glypican 5 inhibits the Wnt/β-catenin pathway. An example would be the research they carried out on prostate cancer. Glypican 5 is poorly expressed in the cell lines of this type of cancer. Upregulation in vitro inhibited cell proliferation and invasion and in vivo attenuated tumor growth. Furthermore, they found that Glypican 5 overexpression inhibited epithelial-mesenchymal transition (EMT) and Wnt/β-catenin signaling [30]. In our study, during osteogenesis we found an overexpression of Glypican 5, perhaps as a negative regulatory element of the Wnt pathway.
In the context of Osteoarthritis and Glypican 3, a recent study evaluated GPC3 levels in synovial fluid in patients with knee OA compared to controls. An accumulation of GPC3 was found in the synovial fluid [31]. These findings, together with the excessive amount of Glypican 3 in the extracellular medium that we found in BMCs and osteoblasts, could indicate that in OA there is a localized accumulation of GPC3 throughout the joint. In this same study, a decrease in GPC3 in the plasma of patients with the disease was associated and inversely associated with radiographic severity [31]. Our data were not significant regarding Glypican 3 in the blood circulation. Studying a larger number of participants that included Glypicans 1 to 6 and Notum, would be necessary to relate it to the severity of the disease.
During osteogenic differentiation, our results in the soluble form of Glypicans continue with the same existing alterations as in mesenchymal stem cells regarding GPC3, GPC5 and Notum. On days 7 and 14 of differentiation, the levels of Glypican 5 in the extracellular medium are decreased. Added to these deregulations is a decrease in Glypican 2. In the study of the gene expression of these components, the results show an overexpression of GPC2 and 5. For its part, in Glypican 3 there is an excessive accumulation throughout the culture in medium extracellular, as found in MSCs. In addition, the mRNA data showed an up-regulation of GPC3. It is proven that the results are inversely related between gene expression and protein levels in the extracellular medium during differentiation towards osteoblasts. It could be that during osteogenic differentiation an attempt will be made to regulate existing alterations.
Regarding Notum, in MSCs and during osteogenesis it shows an increase in its production and is decreased in the extracellular matrix. Wnt activation stimulates Notum production as a regulatory mechanism of this Wnt/β-Catenin signaling pathway [32]. Furthermore, this enzyme is related to bone formation since Notum expression is elevated in cultured osteoblasts and not in osteoclasts [33]. Our results are consistent with the evidence, as they show a high production of Notum at the cellular level in patients with OA, an indirect indication that the Wnt pathway is active and is related to the bone growth characteristic of this disease.
GPC1, 4 and 6 were not present in soluble form in the cultures under either condition or in patients and healthy donors. However, we have found deregulations in the gene expression of Glypicans 4 and 6. The specific function of each of them is not exactly known, so their involvement remains to be elucidated. However, due to the great interest in the involvement of Glypicans in different diseases, there is more evidence of their role. In the case of Glypican 6, in a study was determined that it binds to Wnt3a, even more specifically than Glypican 3, and inhibits canonical Wnt signaling [34], despite what might be expected, since membrane-bound glypicans are supposed to facilitate the activation of the pathway. In our study, we found in OA mesenchymal cells a low production of mRNA in Glypican 6, so it would not contribute to deactivating the pathway. In a possible attempt at regulation, during osteogenic differentiation the data showed an up-regulation in Glypican 6 gene expression.
In the normal maintenance of the joint in adults, mesenchymal cells differentiate into osteoblasts, chondrocytes and adipocytes. One of the most important pathways in bone metabolism is the canonical Wnt/β-catenin pathway. When this signaling pathway is activated thanks to the different mediators involved, mesenchymal stem cells differentiate into osteoblasts, thereby inhibiting chondrogenic and adipogenic differentiation [5,6,7]. Among the mediators involved in the activation and deactivation of the canonical Wnt pathway are Glypicans [12,13]. Another important mediator in this pathway is Notum, which acts as a negative extracellular mediator of the signaling pathway [22]. In osteoarthritis it has been proven that the Wnt/β-catenin pathway is abnormally active [33], with an excessive accumulation of intracellular β-catenin [8]. This would produce an increase in bone mass in osteoarthritis and non-regeneration of cartilage, characteristic of this disease. Taken together, the dysregulations exposed in our results of Glypicans and Notum in mesenchymal stem cells from patients with OA show that these mediators could contribute and/or being a consequence to the Wnt/β-catenin signaling pathway activation in this disease. Furthermore, these dysregulations continue during osteogenic differentiation, adding to other alterations found in the expression of components of the Wnt pathway in osteoblast formation [9]. Probably, as physiologically really occurs, these dysregulations are also implied in the inhibition of chondrogenic and adipogenic differentiation characteristics of osteoarthritis but we have not addressed these particular issues.
In our findings there appears to be a functional relationship between GPC3 and NOTUM. In both MSCs and osteoblasts, we found an upregulation of gene expression in Notum accompanied by a decrease in the extracellular medium and the opposite occurs in Glypican 3. In a study on colorectal cancer, they observed a relationship between Notum and GPC3 in which the production of mRNA and protein is inversely correlated [36]. In fact, this relationship was observed in another previous investigation that proposed that Notum could act as a phospholipase enzyme and cleave the GPI anchor, which keeps Glypicans attached to the cell membrane [23]. However, a recent investigation demonstrated the role Notum played extracellularly and what its interactions were. They verified that Notum is not a glypican-specific phospholipase as had been deduced but that it is a Wnt-specific deacylated with a preference for long-chain cis-unsaturated lipids. Therefore, it eliminates an essential palmitoleate fraction from Wnts and would deactivate the signaling pathway. Notum would not act on Glypicans enzymatically but would bind to them and its sulfated GAG chains of glypicans probably mediate the interaction of Notum with Wnts [24]. Therefore, although there is evidence in different studies of the relationship between Notum and Glypican 3, at the moment it is not known what the specific mechanism by which they may be related is.
On the other hand, returning homeostasis can be a good therapeutic strategy in Osteoarthritis. Notum seems like a very promising molecule for reestablishing increased bone metabolism in OA. In fact, it is beginning to arouse interest as a therapy in diseases where the Wnt/β-Catenin pathway is altered. In a study they verified the effect of Notum on human periodontal ligament stem cells (hPDLSC). Their results revealed that Notum inhibited cell differentiation, probably by regulating the Wnt/β-Catenin signaling pathway [37]. It will be interesting in future studies to know if Notum modulation is also capable of restoring existing alterations in Glypicans.
The limitations of this study were mainly the low number of participants in the study of Glypicans and Notum in plasma. It could be interesting to relate the results to the severity of the disease, age, or gender. On the other hand, in vitro tests do not fully reflect the complexity of the joint, so some in vivo studies would be appropriate.
5. Conclusions
Our results show a generalized alteration in Glypicans 2 to 6 and Notum at the cellular level in Osteoarthritis. It is noteworthy that our findings show that the dysregulations would be intrinsic to mesenchymal stem cells from patients with OA, specifically Glypican 3, 5, 6 and in Notum. In mesenchymal stem cells, when the Wnt/β-catenin signaling pathway is active, MCS preferentially differentiate into osteoblasts and not chondrocytes [33]. Glypicans, both membrane-bound and in soluble form, are mediators of this pathway [12,13]. The alterations found in Glypicans in our study could contribute to the excessive activation of the Wnt/β-catenin signaling pathway that has been observed in Osteoarthritis. For its part, Notum is a negative extracellular regulator of the Wnt pathway [22], which in our study is found to be decreased in the extracellular environment, contributing to the signaling pathway remaining active. In addition, it has been proven that the alterations continue during osteogenic differentiation, also involving other Glypicans.
Another important fact in our research is the decrease found in the circulation of patients with Osteoarthritis of Glypican 5 and Notum. It is partly a reflection of what happens at the cellular level in the joint, so they could be good biomarkers of the disease.
Finally, with the results we have obtained, a future direction suggests that Notum may be an excellent therapeutic strategy for Osteoarthritis. The close relationship of this molecule with Glypicans and the canonical Wnt/β-catenin signaling pathway makes it a promising drug to return homeostasis to the joint in this disease by regulating osteogenic differentiation.
Author Contributions
Conceptualization, BENJAMIN FERNANDEZ-GUTIERREZ; Formal analysis, Luis Rodríguez-Rodriguez and Lydia Abasolo; Funding acquisition, BENJAMIN FERNANDEZ-GUTIERREZ; Investigation, IRENE GONZALEZ-GUEDE and MARIA LOPEZ-RAMOS; Methodology, Arkaitz Mucientes; Project administration, BENJAMIN FERNANDEZ-GUTIERREZ; Resources, BENJAMIN FERNANDEZ-GUTIERREZ; Supervision, BENJAMIN FERNANDEZ-GUTIERREZ; Validation, BENJAMIN FERNANDEZ-GUTIERREZ; Writing – original draft, IRENE GONZALEZ-GUEDE, MARIA LOPEZ-RAMOS and BENJAMIN FERNANDEZ-GUTIERREZ.
Funding
Grant ISCIII FIS PI18/00026
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by Hospital Clinico San Carlos Research Ethics Committee.
Informed Consent Statement
Written informed consent has been obtained from the patients to publish this paper.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Palazzo, C.; Nguyen, C.; Lefevre-Colau, M.M.; Rannou, F.; Poiraudeau, S. Risk factors and burden of osteoarthritis. Ann Phys Rehabil Med. 2016, 59, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; et al. Osteoarthritis. Nat Rev Dis Prim. 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; McDougall, J.J.; Keefe, F.J. The Symptoms of Osteoarthritis and the Genesis of Pain. Rheum Dis Clin North Am. 2008, 34, 623–643. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Felson, D.T. Osteoarthritis. BMJ 2006, 332, 639–642. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat Rev Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.M.; Dixon, K.; Beck, S.; Fabian, D.; Feldman, A.; Barry, F. Reduced chondrogenic and adipogenic activity of mesenchymal stem cells from patients with advanced osteoarthritis. Arthritis Rheum. 2002, 46, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Kwan Tat, S.; Lajeunesse, D.; Pelletier, J.P.; Martel-Pelletier, J. Targeting subchondral bone for treating osteoarthritis: what is the evidence? Best Pract Res Clin Rheumatol. 2010, 24, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.; Bonewald, L.F. The role of the wnt/β-catenin signaling pathway in formation and maintenance of bone and teeth. Int J Biochem Cell Biol. 2016, 77, 23–29. [Google Scholar] [CrossRef]
- Escobar-Gómez, F.; Jódar, E.; Hawkins, F. Receptor Wnt: fisiología, fisiopatología y potenciales nuevas dianas terapéuticas. Rev Española Enfermedades Metabólicas Óseas. 2009, 18, 39–44. [Google Scholar] [CrossRef]
- Corr, M. Wnt–β-catenin signaling in the pathogenesis of osteoarthritis. Nat Clin Pract Rheumatol. 2008, 4, 550–556. [Google Scholar] [CrossRef]
- Tornero-Esteban, P.; Peralta-Sastre, A.; Herranz, E.; et al. Altered Expression of Wnt Signaling Pathway Components in Osteogenesis of Mesenchymal Stem Cells in Osteoarthritis Patients. Shi XM, ed. PLoS One 2015, 10, e0137170. [Google Scholar] [CrossRef]
- Blom, A.B.; Brockbank, S.M.; van Lent, P.L.; et al. Involvement of the Wnt signaling pathway in experimental and human osteoarthritis: Prominent role of Wnt-induced signaling protein 1. Arthritis Rheum. 2009, 60, 501–512. [Google Scholar] [CrossRef]
- Held, A.; Glas, A.; Dietrich, L.; et al. Targeting β-catenin dependent Wnt signaling via peptidomimetic inhibitors in murine chondrocytes and OA cartilage. Osteoarthr Cartil. 2018, 26, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Filmus, J.; Capurro, M.; Rast, J. Glypicans. Genome Biol. 2008, 9, 224. [Google Scholar] [CrossRef] [PubMed]
- Boudin, E.; Fijalkowski, I.; Piters, E.; Van Hul, W. The role of extracellular modulators of canonical Wnt signaling in bone metabolism and diseases. Semin Arthritis Rheum. 2013, 43, 220–240. [Google Scholar] [CrossRef]
- Kaur, S.P.; Verma, A.; Lee, H.K.; Barnett, L.M.; Somanath, P.R.; Cummings, B.S. Inhibition of glypican-1 expression induces an activated fibroblast phenotype in a human bone marrow-derived stromal cell-line. Sci Rep. 2021, 11, 9262. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Yang, T.; Li, H.; et al. Frequently expressed glypican-3 as a promising novel therapeutic target for osteosarcomas. Cancer Sci. 2022, 113, 3618–3632. [Google Scholar] [CrossRef]
- Dwivedi, P.P.; Grose, R.H.; Filmus, J.; et al. Regulation of bone morphogenetic protein signalling and cranial osteogenesis by Gpc1 and Gpc3. Bone 2013, 55, 367–376. [Google Scholar] [CrossRef]
- LeClair, E.E.; Mui, S.R.; Huang, A.; Topczewska, J.M.; Topczewski, J. Craniofacial skeletal defects of adult zebrafish Glypican 4 (knypek) mutants. Dev Dyn. 2009, 238, 2550–2563. [Google Scholar] [CrossRef]
- Amor, D.J.; Stephenson, S.E.M.; Mustapha, M.; et al. Pathogenic Variants in GPC4 Cause Keipert Syndrome. Am J Hum Genet. 2019, 104, 914–924. [Google Scholar] [CrossRef]
- Trajanoska, K.; Rivadeneira, F. The genetic architecture of osteoporosis and fracture risk. Bone 2019, 126, 2–10. [Google Scholar] [CrossRef]
- Kemp, J.P.; Morris, J.A.; Medina-Gomez, C.; et al. Identification of 153 new loci associated with heel bone mineral density and functional involvement of GPC6 in osteoporosis. Nat Genet. 2017, 49, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Traister, A.; Shi, W.; Filmus, J. Mammalian Notum induces the release of glypicans and other GPI-anchored proteins from the cell surface. Biochem J. 2008, 410, 503–511. [Google Scholar] [CrossRef]
- Kakugawa, S.; Langton, P.F.; Zebisch, M.; et al. Notum deacylates Wnt proteins to suppress signalling activity. Nature 2015, 519, 187–192. [Google Scholar] [CrossRef]
- Lietman, C.; Wu, B.; Lechner, S.; et al. Inhibition of Wnt/β-catenin signaling ameliorates osteoarthritis in a murine model of experimental osteoarthritis. JCI Insight 2018, 3(3). [Google Scholar] [CrossRef]
- Deshmukh, V.; Hu, H.; Barroga, C.; et al. A small-molecule inhibitor of the Wnt pathway (SM04690) as a potential disease modifying agent for the treatment of osteoarthritis of the knee. Osteoarthr Cartil. 2018, 26, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, X.; Xing, L.; Tian, F. Wnt signaling: a promising target for osteoarthritis therapy. Cell Commun Signal. 2019, 17, 97. [Google Scholar] [CrossRef] [PubMed]
- Song, H.H.; Shi, W.; Xiang, Y.Y.; Filmus, J. The Loss of Glypican-3 Induces Alterations in Wnt Signaling. J Biol Chem. 2005, 280, 2116–2125. [Google Scholar] [CrossRef]
- Capurro, M.; Martin, T.; Shi, W.; Filmus, J. Glypican-3 binds to frizzled and plays a direct role in the stimulation of canonical Wnt signaling. J Cell Sci. Published online January 1. 2014. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, K.; He, M.; Fan, G.; Lu, H. Overexpression of Glypican 5 (GPC5) Inhibits Prostate Cancer Cell Proliferation and Invasion via Suppressing Sp1-Mediated EMT and Activation of Wnt/β-Catenin Signaling. Oncol Res Featur Preclin Clin Cancer Ther. 2018, 26, 565–572. [Google Scholar] [CrossRef]
- Udomsinprasert, W.; McConachie, E.; Ngarmukos, S.; Theerawattanapong, N.; Tanavalee, A.; Honsawek, S. Plasma and Joint Fluid Glypican-3 Are Inversely Correlated with the Severity of Knee Osteoarthritis. Cartilage 2021, 12, 505–511. [Google Scholar] [CrossRef]
- Torisu, Y.; Watanabe, A.; Nonaka, A.; et al. Human homolog of NOTUM, overexpressed in hepatocellular carcinoma, is regulated transcriptionally by β-catenin/TCF. Cancer Sci. 2008, 99, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Movérare-Skrtic, S.; Nilsson, K.H.; Henning, P.; et al. Osteoblast-derived NOTUM reduces cortical bone mass in mice and the NOTUM locus is associated with bone mineral density in humans. FASEB J. 2019, 33, 11163–11179. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.; Martin, T.; Shi, W.; Filmus, J. Glypican-3 binds to frizzled and plays a direct role in the stimulation of canonical Wnt signaling. J Cell Sci. Published online January 1. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.P.; Später, D.; Taketo, M.M.; Birchmeier, W.; Hartmann, C. Canonical Wnt/β-Catenin Signaling Prevents Osteoblasts from Differentiating into Chondrocytes. Dev Cell. 2005, 8, 727–738. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, M.; Arigoni, M.; Loiacono, L.; et al. Novel insights into Notum and glypicans regulation in colorectal cancer. Oncotarget 2015, 6, 41237–41257. [Google Scholar] [CrossRef]
- Yang, P.; Li, C.; Kou, Y.; et al. Notum suppresses the osteogenic differentiation of periodontal ligament stem cells through the Wnt/Beta catenin signaling pathway. Arch Oral Biol. 2021, 130, 105211. [Google Scholar] [CrossRef]
Figure 1.
Plasma levels of Glypicans and Notum in patients with Osteoarthritis (n= 25) and controls (n=24). Non-parametric Mann-Whitney test (GPC2, GPC4, GPC6 and Notum) and t-test (GPC1, GPC5, GPC3) were used to compare the groups: * p < 0.05, **p <0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.
Figure 1.
Plasma levels of Glypicans and Notum in patients with Osteoarthritis (n= 25) and controls (n=24). Non-parametric Mann-Whitney test (GPC2, GPC4, GPC6 and Notum) and t-test (GPC1, GPC5, GPC3) were used to compare the groups: * p < 0.05, **p <0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.
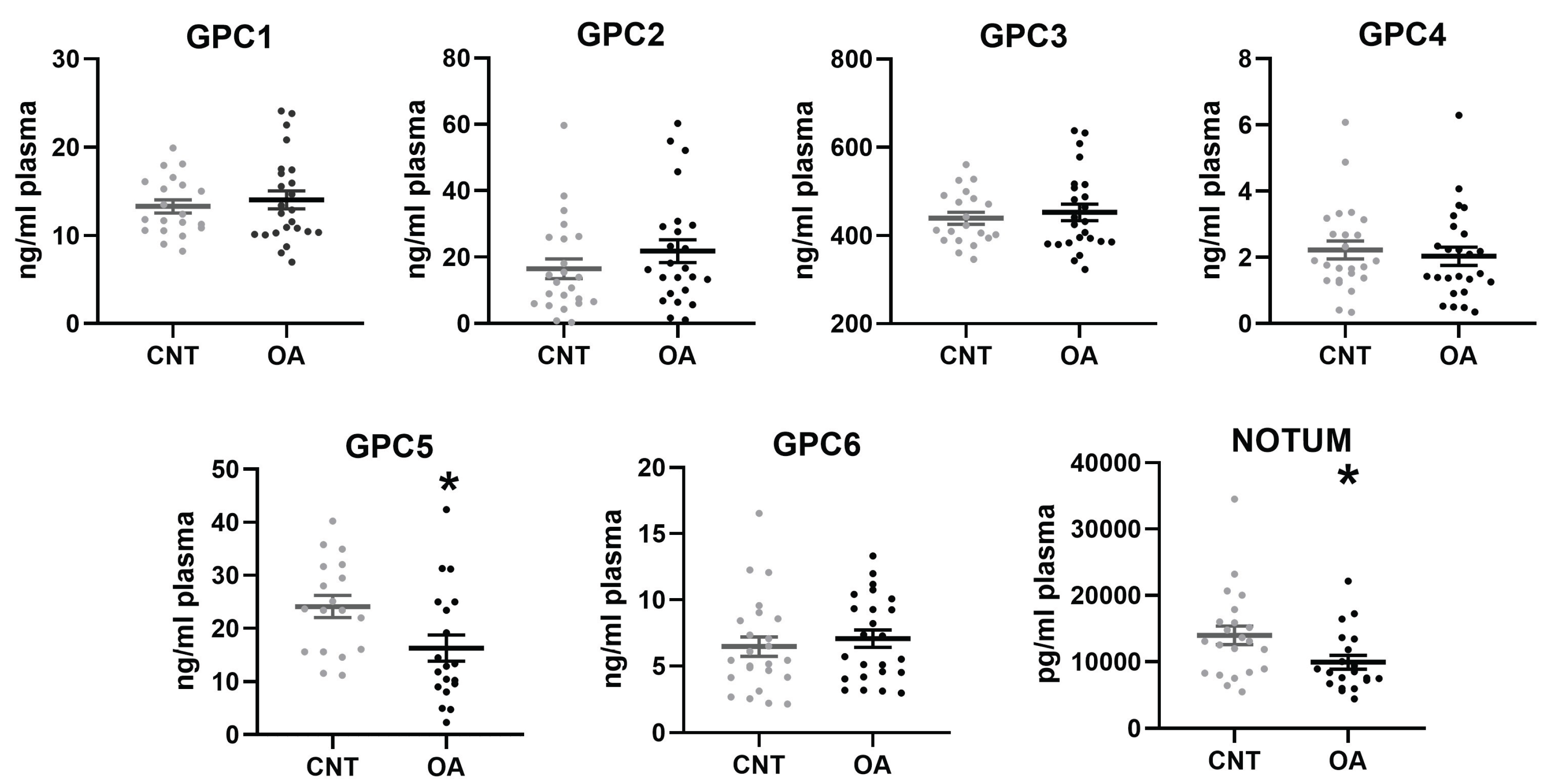
Figure 2.
Expression gene of Glypicans and Notum measured by qPCR at 7, 14 and 21 days of culture in patients with Osteoarthritis (n= 8) and Controls (n= 8). Relative gene expression was normalized to housekeeping genes β-actin y RNA18S. The data were calculated with the method of fold change (2-ΔΔCt) relative t=0. T-test were used to compare the groups: *p < 0.05, **p<0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.
Figure 2.
Expression gene of Glypicans and Notum measured by qPCR at 7, 14 and 21 days of culture in patients with Osteoarthritis (n= 8) and Controls (n= 8). Relative gene expression was normalized to housekeeping genes β-actin y RNA18S. The data were calculated with the method of fold change (2-ΔΔCt) relative t=0. T-test were used to compare the groups: *p < 0.05, **p<0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.

Figure 3.
Protein levels of Glypicans and Notum determined by ELISA kit at 0, 7, 14 and 21 days of BM-MSC and Osteoblast culture in patients with Osteoarthritis (n= 8) and Controls (n= 8).T-test were used to compare the groups: *p < 0.05, **p<0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.
Figure 3.
Protein levels of Glypicans and Notum determined by ELISA kit at 0, 7, 14 and 21 days of BM-MSC and Osteoblast culture in patients with Osteoarthritis (n= 8) and Controls (n= 8).T-test were used to compare the groups: *p < 0.05, **p<0.01, ***p < 0.001. All bar graphs are presented as mean ± SEM.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



