
Submitted:
29 March 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
In spite of prevalence gaps in several micronutrients supporting immune function, significance of their deficits/supplementation for efficacy of vaccines is underinvestigated. Thus, influence of supplementation combining vitamins C and D, zinc, selenium and manganese, and N-acetyl-cysteine on mouse IgG responses and underlying mechanisms to seasonal quadrivalent influenza vaccine (QIV) was investigated. QIV was given in two doses (28 days apart). The supplementation starting 5 days before the first injection increased the total serum IgG titre, through stimulation of germinal centre (GC) reaction in secondary lymphoid organs (SLOs), as indicated by increase in the frequency of GC B cells and follicular CD4+ T helper (Th) cells, and IL-21-producing Th cells. Additionally, the supplementation “optimized” IgG response following the second QIV injection by supporting more favorable (in respect to virus clearance) IgG2a response through favoring Th1 differentiation as indicating by Th1 (INF-γ) /Th2 (IL-4) signature cytokine level ratio upon QIV restimulation in SLO cell cultures. This most likely reflected partly their antioxidant action (as indicated by redox status analyses) on Th cell differentiation. Thus, the study forms solid base for further research aimed at repurposing use this safe and inexpensive micronutrient combination as an adjuvant for virus influenza vaccine.
Keywords:
antioxidant supplementation
; influenza vaccine
; antibody response
; germinal centre reaction
1. Introduction
The World Health Organization (WHO) estimates that there are 1 billion cases of seasonal influenza worldwide each year, of which 3–5 million are severe cases leading approximately to 650,000 deaths per year [1]. The most effective way to prevent influenza infection is through vaccination [2]. Most commonly used seasonal influenza vaccines are inactivated influenza virus vaccines administered through the intramuscular route [3]. Current seasonal influenza vaccines are prepared in line with the WHO recommendations and contain two influenza A viruses (H1N1 and H3N2) and one (in the trivalent influenza vaccine) or two [in quadrivalent influenza vaccine (QIV)] from each lineage (Victoria and Yamagata) influenza B viruses [4]. It is noteworthy that these vaccines have highly variable efficacy that ranges between 10% and 60% [5, 6], being particularly low in certain vulnerable groups, such as the elderly, whose participation in general population is constantly increasing [7]. This is consistent with data indicating that the elderly have diminished antibody responses to vaccines, including the seasonal influenza vaccine [8]. Considering that seasonal influenza vaccination campaigns are considered to be a major investment for countries and governments, it is absolutely clear that is important to improve/maximize the effectiveness of the vaccine not only from the global health point of view but also from economic reasons. So far much attention is paved to the virus-related factors contributing to inconsistency in the immune responses to this vaccine and therefore vaccine efficacy, while host-associated variables, such as genetic factors, biological sex, age, and hormonal and nutritional status-related present prior to vaccination are still underinvestigated [9]. In this context, it should be pointed out that although the vaccine-specific IgG antibody response can be used as an indicator of vaccine efficacy, cellular immune response also has an important role in the protection [10]. According to the statistical analysis of [11], the CD4+ cell response is crucial for helper B cell differentiation and cytokine production, so this response is sometimes more associated with protection than with antibody titres.
Having in mind all aforementioned, it is important to get insights into targetable mechanisms of influenza vaccine-induced immune response, and consequently to identify putative extrinsic factor that could positively impact/condition relevant immune mechanisms for the response even prior to the vaccination, and thereby “optimize” immune response to the vaccine. In this respect we focused on influence of supplementation with antioxidants/immunomodulators [vitamins, oligoelements and modified amino acids, such as N-acetyl-cysteine (NAC)], which is so far predominantly ordered and used to prevent viral infection and promote recovery from these infections [12-19]. The investigation of the impact of such a supplementation seems particularly justifiable as: i) variety of observational and some interventional studies report that adequate micronutrient status or micronutrient supplementation is associated with enhanced vaccine responses [20], ii) gaps in several micronutrients, including those aforementioned, are reported [20] and (iii) such an intervention could be implemented safely and inexpensively. Thus, in the present study the influence of a supplement containing vitamins D and C, oligoelements zinc, selenium and manganese, and modified amino acid N-acetyl-cysteine recommended for prophylaxis of viral infections and promotion of the recovery was attested for its capacity to quantitatively and qualitatively optimize antibody immune response to QIV in mice. Of note, to assess influence of this supplementation on the primary immune response and secondary (viz. the response to subsequent challenge with the same antigens, as upon subsequent injection of the vaccine or natural influenza infection) immune response, QIV was administered in two doses with 28-day interval between them. Consequently, the central goal of this study was to investigate possibility to repurpose use of such supplementation to include support to/optimization of immune response to influenza virus vaccine in the spectrum of indications for its use. To reach this central goal, apart from the total serum QIV-specific IgG titres, and titres of major IgG subclasses (IgG1 and IgG2a), germinal centre (GC) B cell and GC CD4+ T cell immune responses to the vaccine were investigated. Specifically, in draining lymph nodes (dLNs) and spleens the frequency of GC B cells and CD4+ (T helper, Th) cells and their proliferative responses to restimulation with QIV antigens in vitro were explored. Of note, in these secondary lymphoid organs (SLOs) the frequency of follicular helper (Tfh) cells, a specialized CD4+ T cell subset important for B cell differentiation into high-affinity antibody-secreting plasma cells, and follicular CD4+ T regulatory (Tfr) cells, acting directly and indirectly (in a Tfh-dependent manner) on GC B cells to restrict their response [21,22] were also investigated. Additionally, Th cells were examined for synthesis of IL-21, the cytokine with key role in GC biology through its independent actions on T and B cells [23]. Moreover, given that Th1 and Th2 cells have differential capabilities in stimulating B cells to secrete antibody isotypes, where Th1 type cells elicit IgG2a antibody production and Th2 type cells induce IgG1 antibody secretion [24], the ratio in production levels of Th1 and Th2 signature cytokines in QIV-restimulated dLN cell and splenocyte cultures was determined.
2. Materials and Methods
2.1. Experimental Animals
The study included 8-10-week-old female mice of BALB/c strain (6 per group) bred at the Institute of Virology, Vaccines and Sera "Torlak", Belgrade, Serbia. They were kept under the standard housing conditions (12/12 hour cycle of light and dark and controlled temperature and humidity environment) with ad libitum access to food and water. The animal care and all experimental procedures were performed in accordance with the Directive 2010/63/EU of the European Parliament and of the Council on the protection of animals used for scientific purposes and with the governmental regulations (Law on Animal Welfare, “Official Gazette of Republic of Serbia”, No. 41/2009). The study protocol was evaluated by the Animal Care and Use Committee of Institute of Virology, Vaccines and Sera “Torlak” and approved by The Ministry of Agriculture, Forestry and Water Economy of the Republic of Serbia-Veterinary Directorate (permition number: 323-07-12928/2022-05).
2.2. Immunization and Treatment
Experimental animals were immunised with QIV for season 2022/2023, which contained: A/Victoria/2570/2019 (H1N1)pdm09-like virus, A/Darwin/9/2021 (H3N2)-like virus, B/Austria/1359417/2021 (B/Victoria lineage)-like virus, and B/Phuket/3073/2013 (B/Yamagata lineage)-like virus (Sanofi Pasteur–Val de Reuil, Val de Reuil, France). Vaccine was applied intramuscularly 50 μl in each caudal tight, in a dose containing 3 μg of virus surface hemagglutinin (HA) of each virus strain/lineage, the human equivalent dose [25]. Immunization was carried out at two time points, with an interval of 28 days between them (Figure 1). Control animals were injected with saline in the same way. Animals were monitored daily by veterinarian and researchers and showed no signs of local or systemic reactions.
Supplementation with 3.78 mg/mouse, the human equivalent dose [25] of antioxidants/immunomodulators in saline or administration of same volume of saline alone started five days before the first injection of QIV or saline. The supplement contained combination of antioxidants/immunomodulators in the same ratio as in the commercially available combination of antioxidants/immunomodulators BiVits® ACTIVA Recovery (AbelaPharm, Belgrade, Serbia), viz. 500.0 mg vitamin C (L-ascorbic acid), 25.0 µg vitamin D3 (cholecalciferol), 10.0 mg zinc (zinc- citrate), 1.0 mg manganese (manganese gluconate), 27.5 µg selenium (L-selenomethionine) and 300.0 mg N-acetyl-cysteine (NAC) obtained by the process of natural fermentation in saline. The supplement or saline alone was administrated per os daily until the day of sacrifice (Figure 1).
2.3. Reagents and Antibodies
Ketamine (Ketamidor, Richter Pharma AG, Wels, Austria) and xylasine (Xylased, Bioveta, Ivanovice na Hané, Czech Republic) were used for making anaesthesia cocktail.
Horseradish peroxidase (HRP)-conjugated: goat anti-mouse IgG, IgG1, IgG2a antibodies (Jackson ImmunoResearch Laboratories Inc., WestGrove, PA, USA), and o-phenylenediamine (OPD) from Sigma were used for antibody titre determination in the sera of the experimental mice.
For flow cytometric analyses (FCA) are used the following antibodies: fluorescein isothiocyanate (FITC)-conjugated anti-B220 (CD45R, clone RA3-6B2), anti-FoxP3 (clone FJK-16s) and phycoerythrin (PE)-conjugated anti-CD95 (clone 15A7) obtained from eBiolegend (Carlsbad, CA, USA) and PE- or peridinin chlorophyll protein cyanine (PerCP-Cy™5.5)-conjugated anti-CD4 antibody (clone RM4-5) purchased from Biolegend (San Diego, CA, USA) and BD Biosciences Pharmingen (Mountain View, CA, USA), respectively. PerCP-Cy™5.5- or PE-conjugated anti-mouse CXCR5 (Clone 2G8) and Alexa Fluor (AF) 647 anti-human/mouse Bcl-6 (K112-91) acquired from BD Biosciences Pharmingen were also used. Additionally, rabbit polyclonal anti-IL-21 antibody (Merck KGaA, Darmstadt, Germany), and FITC-conjugated goat anti-rabbit Ig (BD Biosciences Pharmingen), as secondary antibody, were used. For analyses of cell proliferation FITC- and PE-conjugated anti-Ki67 (clone SolA15; eBioscience Inc. San Diego, CA, USA) were used. All antibodies used in the experiments specifically react with mouse antigens and have been tested for use in flow cytometry by the manufacturers. Target cell antigen specificity for all the antibodies used was also confirmed in our previous [26], and the present study, in single- or multi-colour analyses, and using an antigen-positive and -negative cell types.
Medium for cell cultivation consisted of RPMI 1640 (Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany) with 2 mM L-glutamine (Serva, Heidelberg, Germany), 1 mM sodium pyruvate (Serva), 100 unit/ml penicillin (ICN, Costa Mesa, CA, USA), 100 μg/ml streptomycin (ICN) and 10 % fetal calf serum (FCS) (Gibco, Grand Island, NY, USA). Bovine serum albumin (BSA) was obtained from Fluka AG (Chemie) GmbH (Buchs SG, Switzerland).
Phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich Chemie GmbH), ionomycin (Sigma-Aldrich Chemie GmbH) and brefeldin A (eBioscience) were applied for IL-21 expression.
Legend max™ mouse interferon (IFN)-γ and interleukin (IL)-4 ELISA kits (Biolegend) were used for determination of cytokines in cell culture supernatants.
2.4. Serum Collection
Animals were deeply anesthetised with ketamine [80 mg/kg body weight (BW)]/xylazine (8 mg/kg BW) cocktail and bled from retroorbital sinuses. Blood samples were left to coagulate for one hour at the room temperature (RT) and then centrifuged at 2000 x g for 10 min in order to collect sera. Sera were decomplemented by incubation for 30 minutes at 56°C and stored at -20°C until further analyses.
Blood samples were collected at two time points: 28 days after the injection of the first dose of QIV (when the titre reaches maximum according to preliminary experiment) or saline, and 14 days after the second injection of QIV or saline (Figure 1).
2.5. Antibody Titre Determination
For determination of the total IgG antibody titre and the IgG subclass titres (IgG1 and IgG2a) of virus strain specific antibodies, end point dilution enzyme-linked immunosorbent assay (ELISA) was used. As previously described [26], mice sera were serially diluted in PBS with 1% BSA, and added to Nunc MaxiSorp 96 well flat bottom plates pre-coated with 2.5 μg of HA/ml of QIV. After incubation for one hour at RT plates were washed firstly three times with 0.05% Tween 20/PBS and then once with PBS. Horseradish peroxidase-conjugated IgG and IgG subclass specific goat anti-mouse antibodies in appropriate dilutions: IgG (1:10000), IgG1 (1:10000), IgG2a (1:10000) were added and incubated for one hour at RT. After washing, OPD with H2O2 were added and reaction was stopped after 15 minutes with 2M H2SO4. Optical density was measured with Multiscan Ascent (Labsystems) at 490 nm and 620 nm. Titres were calculated as the reciprocal of the highest dilution of test sera that gave an absorbance reading value of 3 standard deviations above the control sera at an equivalent dilution.
2.6. Isolation of Cells from dLNs and Spleens
Cellular immune response was examined at two time points: two weeks after the first and two weeks after the second dose of QIV. Spleens and dLNs (inguinal) and were removed and grinded through 60 μm sieve screen in 10% FCS/RPMI 1640 under the sterile conditions. Lysis buffer (0.15 M NH4Cl, 1mM KHCO3, and 0.1 mM Na2EDTA in H2O, pH 7.2) was added to remove red blood cells from the splenic cell suspensions. After 10 minutes of incubation, cell suspensions were centrifuged for 10 min at 300 x g, and after washing, resuspended in 10% FCS/PBS and counted in 0.2% trypan blue dye using the improved Neubauer haemocytometer. Isolated cells were subjected to examination of phenotype, cytokine production and proliferation capability (Figure 1).
2.7. Cultivation of Cells
Splenic and dLN cell suspensions were adjusted to 5x105 cells/well and cells were cultivated in medium (10%FCS/RPMI 1640) in Nunc MaxiSorp 96 well U-bottom plates at 37°C in a 5% CO2 humidified air atmosphere. After 72 hours of incubation with or without stimuli (QIV), cells were subjected to immunofluorescent staining and FCA to determine proliferative cell response and IL-21 expression. Additionally, supernatants were collected for assessment of IFN-γ and IL-4 production by ELISA.
2.8. Cell Proliferation Assessment
Proliferation capacity of CD4+CXCR5+ T cells and B220+CD95+ B cells from spleen and dLNs were examined after in vitro (re)stimulation with QIV (5 μg of HA/ml). Briefly, after surface immunostaining of CD4 and CXCR5 or B220 and CD95 antigens, cells were fixed/permeabilized overnight at 4°C and then subjected to intracellular staining with anti-Ki67 and subsequently to FCA.
2.9. Intracellular Cytokine Production Assessment
IL-21 cytokine production was analysed in freshly isolated splenocytes and dLN cells, and following in vitro restimulation with QIV (5 μg of HA/ml) for 72 hours. Before IL-21 immunostaining, freshly isolated and in vitro restimulated cells were incubated in the presence of 200 ng/mL PMA, 400 ng/mL ionomycin, and 3 μg/mL of brefeldin A in a 5% CO2 humidified atmosphere at 37°C for 4 hours.
2.10. IFN-γ and IL-4 Production Assessment
After incubation of dLN cells and splenocytes in the presence or in the absence of QIV (5 μg of HA/ml) for 72 hours in a 5% CO2 humidified air atmosphere at 37°C supernatants were collected for IFN-γ and IL-4 production. The supernatants were stored at -20°C until determination of the cytokine concentration.
2.11. Immunostaining and FCA
2.11.1. Surface Antigens Staining
For surface immunostaining, cells were dispersed into BD tubes (5x105 cells) and washed in 2% FCS/0.1% NaN3/PBS. Following washing, cells were incubated with monoclonal antibodies against surface antigens (CD4, CXCR5, B220, CD95) for 30 minutes at 4°C. After incubation, cells were washed and then resuspended in 0.1% NaN3/ PBS for FCA or fixed and permeabilized overnight at 4°C for intracellular staining using fixation/permeabilization buffers (eBioscience) according to manufacturer’s instructions.
2.11.2. Intracellular Antigen Staining
For intracellular immunostaining, following staining of surface antigens, cells were subjected to overnight fixation/permeabilization, and then washed and incubated with fluorochrome-conjugated or unconjugated antibodies specific for intracellular antigens (FoxP3, Bcl6, Ki67, IL-21) for 30 minutes at RT. The cells incubated with unconjugated anti-IL-21 antibody were washed and then incubated with FITC-conjugated goat anti-rabbit Ig for further 30 min.
After PBS washing the cells immunolabeled for intracellular antigens were acquired on FACSVerse flow cytometer (Becton Dickinson, Mountain View, CA, USA).
2.11.3. FCA
To secure relevant number of event for statistical analysis in each gate 100,000 events per sample were acquired on flow cytometer as calculated by Poisson statistics. Frequency of a marker positive cells were determined using FlowJo software version 7.8. (TreeStar Inc., Ashland, OR, USA). To optimise analyses, unstained cells, fluorochrome-matched isotype staining and secondary antibody only controls were used. With the exception of markers exhibiting clear bimodal staining [27], fluorescence minus one (FMO) controls were used to determine the gating boundaries.
2.12. ELISA
Supernatants from splenocyte and dLN cell cultures were collected and IFN-γ and IL-4 concentrations were determined using Biolegend ELISA kits following manufacturer’s instructions. Briefly, precoated plates were washed with assay buffer and 50 μL/well of appropriately diluted standards or samples were added to the wells. After two-hour incubation at RT while shaking, plates were washed. Detection antibody was added and incubated for additional 1 hour at RT while shaking. Following washing, avidin-HRP solution was added and incubation continued for 30 minutes at RT while shaking. After washing, the substrate solution was added and incubated for following 15 min in the dark. The reaction was stopped with the stop solution and OD was measured with Multiscan Ascent was read at 450 nm and 570 nm. Standard curve to determine concentrations was calculated for each assay with the limit of detection of 0.5 pg/mL and 8 pg/mL for IL-4 and IFN-γ, respectively.
2.13. Redox Status Parameters Assessment
Frozen samples of spleens extirpated two weeks after the second dose of QIV were homogenized in 0.1 M phosphate buffer (pH = 7.4) in a 1:9 weight-to-volume ratio using the Tehtnica homogenizer (Železniki, Slovenia). After homogenization, the samples were centrifuged for 10 min at 800 x g, and then for 20 min at 9500 x g to obtain supernatants. All the steps were carried out at 0–4°C. In thereby obtained supernatants several pro-oxidant and antioxidant parameters were assessed. Specifically, (O2•−) and advanced oxidation protein product (AOPP) levels, total pro-oxidant status (TOC), and superoxide dismutase (SOD) activity and total SH-group (SHG) and reduced glutathione (GSH) levels were examined. All obtained results for redox parameters were normalized to the protein concentration of supernatants, which was determined using the Bradford method.
The level of superoxide anion radical O2•− was estimated according to the rate of yellow nitroblue tetrazolium reduction to blue diformazan [28]. Total pro-oxidant capacity (TOC) was determined using a slightly modified spectrophotometric method introduced by Erel [29]. The method is based on the ability of oxidants such as hydrogen peroxide and lipid hydroperoxide to oxidize the ferrous ion-o-dianisidine complex to ferric ion, which in reaction with xylenol orange in an acidic medium makes a coloured complex. The intensity of colour is proportional with the total amount of oxidant molecules in the sample. The assay is calibrated with hydrogen peroxide in concentration ranged from 10 to 200 μmol/L. The level of advanced oxidation protein products (AOPP) was estimated by measuring the absorbance at 340 nm of complex formed in their reaction with glacial acetic acid potassium-iodide [30]. Chloramine T, a compound exhibiting specific absorption maximum at 340 nm, was used as a standard with concentration range from 10 to 100 μmol/L. Cu/Zn superoxide dismutase (SOD) activity was measured using slightly modified method of [31]. This method is based on the ability of SOD to inhibit auto-oxidation of epinephrine in alkaline medium. The enzyme activity is calculated as percentage of inhibition of epinephrine auto-oxidation. The levels of SHG were measured by a modified Ellman's method [29], using 10 mmol/L dinitrodithiobenzoic acid as a reagent. This reagent reacts with aliphatic thiol compounds in a basic environment (pH 9.0) and this reaction generates 1 mol p-nitrophenol anion per mol of thiol. The absorbance was measured at 412 nm. The calibration of the method was achieved with the reduced glutathione in the concentration range from 0.1 to 1.0 mmol/L. The level of glutathione (GSH) was determined based on its reaction with 5,5′-dithiobis-2-nitrobenzoic acid leading to formation of a yellow complex 5-thio-2-nitrobenzoate, which is measured after the deproteinization with sulfosalicylic acid at 412 nm [32]. Reduced glutathione in a range from 0.5 to 2 μmol/L was used for the calibration.
The levels of O2•–, TOC, and SOD activity were determined on the ILAB 300 Plus analyser (Instrumentation Laboratory, Italy), whereas AOPP, SHG, GSH and protein concentrations were evaluated on a continuous spectrophotometer (Pharmacia LKB, UK).
2.14. Statistics
Statistically significant differences between the groups were assessed by Kruskal Wallis test followed by Mann-Whitney U test. All statistical analyses were performed using GraphPad Prism 9 software (GraphPad Software, Inc., La Jolla, CA, USA). Data are presented as median ± interquartile range, unless otherwise specified. Differences are considered significant when p<0.05.
3. Results
3.1. Antioxidant/Immunomodulatory Supplementation Increased the Total Titre of Influenza Virus-Specific IgG in Sera and the Frequency of GC B cells in dLNs and Spleens of Mice Injected with One Dose of QIV
Given that differently from immunised mice, the geo means of the total serum titre of influenza virus-specific IgG in mice who received one or two injections of saline and supplemented with antioxidants/immunomodulators (Controls) were equal to zero, it is obvious that all mice who received QIV developed influenza virus-specific antibody response (Figure 2a). In mice injected with one dose of QIV 28 days post immunization the total serum titre of influenza virus-specific IgG was higher (p≤0.05) in mice supplemented with the antioxidants/immunomodulators compared with those administered with saline (Figure 2a). As expected [33], the second injection of QIV on 28th day post the first injection boosted the total serum titre of influenza virus-specific IgG, so 14 days after the injection it was dramatically higher (p≤0.01) compared with that measured on the day when the second challenge was applied (Fig.2a). The supplementation did not significantly influence this response (Figure 2a).
In accordance with IgG antibody response to the first injection of QIV, 28 days post the first QIV injection the frequency of B220+CD95+GL7+ cells, presumably GC B cells [34], in dLNs and spleens from mice treated with supplement was higher (p≤0.01) compared with those administered with saline (Figure 2b). Furthermore, the second injection of QIV additionally increased the frequency of GC B cells in SLOs (p≤0.01), so 14 days post the second injection their frequency in dLNs (p≤0.01) and spleens (p≤0.05) was higher than at the time when the second injection was applied (Figure 2b). The treatment with the supplement did not statistically significantly influence frequency of B220+CD95+GL7+ cells either in dLNs or spleens from mice administered with two doses of QIV (Figure 2b).
Next, to enlighten the putative mechanisms underlying the increase in the frequency of B cells participating in GC reaction, their proliferative index in response to restimulation with QIV antigens (viz. the increase in the frequency of proliferating Ki67+ cells in the presence of QIV antigens over that in their absence) in vitro was investigated (Figure 2c). B cells participating in GC reaction were defined by surface co-expression of B220 and the pro-apoptotic receptor CD95 (also known as APO-1/Fas), the crucial regulator of B-cell–T-cell interactions in SLOs [35,36]. The proliferative index of GC B cells from cultures of dLN lymphocytes and splenocytes isolated from mice injected with one dose of QIV and treated with the supplement was greater (p≤0.01) when compared with proliferative index of corresponding cells isolated from mice injected with one dose of QIV and administered with saline (Figure 2c). As expected from previous findings, GC B cells from dLNs and spleens from mice injected with two doses of QIV and administered with saline exhibited greater (p≤0.01) proliferative index compared with mice injected with one dose of QIV and administered with saline (Figure 2c). Furthermore, supplementation with antioxidants/immunomodulators did not statistically significantly influence the proliferative response of GC B cells in cultures from either dLNs or spleens of mice receiving two doses of QIV (Figure 2c).
3.2. Supplementation with Anti-Oxidants/Immunomodulators Increased the Frequency of Tfh Cells Only in SLOs from Mice Injected with One Dose of QIV
Considering that the initiation of the GC requires both CD4+ T cells and B cells to be activated by cognate antigen [37], the frequency of GC T cells (defined by the surface expression of CXCR5 and Bcl6 molecules) [38] was investigated in SLOs of mice immunized with QIV. The frequency of CD4+CXCR5+Bcl6+ cells increased in dLNs (p≤0.05) and spleens (p≤0.01) of mice subjected to immunization with one dose of QIV and supplementation compared with mice treated with one injection of QIV and saline (Figure 3a). In mice administered with saline, their frequency in dLNs (p≤0.05) and spleens (p≤0.01) was also higher in mice injected with two doses of QIV in respect to those administered with only one dose of the vaccine (Figure 3a). The supplementation did not influence the frequency CD4+CXCR5+Bcl6+ cells in either dLNs or spleens from mice injected with two doses of QIV (Figure 3a). Their frequency was higher (p≤0.01) in spleens, but not in dLN of mice injected with two doses of QIV and supplemented with antioxidants/immunomodulators compared with their frequency in corresponding SLO of mice injected with one dose of QIV and treated with the supplement (Figure 3a).
To confirm significance of QIV antigen-specific expansion of GC CD4+ T cells for the changes in their frequency in SLOs, in cultures of dLNs and splenocytes the proliferative index of these cells upon in vitro restimulation with QIV (viz. the increase in the frequency of proliferating Ki67+ cells in the presence of QIV antigens over that in their absence) was calculated. Indeed, the greater (p≤0.01) proliferative index exhibited GC CD4+ cells from cultures of dLN lymphocytes and splenocytes of mice injected with one dose of QIV and supplemented with antioxidants/immunomodulators than those from cultures of their counterparts administered with saline (Figure 3b). Besides, in dLN lymphocyte and splenocyte cultures from mice administered with saline the greater (p≤0.01) GC CD4+ cell proliferative index was found than in cultures from mice injected with two doses of QIV than in those from mice injected with one dose of the vaccine (Figure 3b). Of note, GC CD4+ cells from cultures of dLN lymphocytes and splenocytes of mice injected with two doses of QIV and supplemented with antioxidants/immunomodulators and mice injected with two doses of the vaccine and administered with saline exhibited similar proliferative index (Figure 3b). Their proliferative index in splenocyte cultures of mice injected with two doses of QIV and administered with antioxidants/immunomodulators was greater (p≤0.01) than in splenocyte cultures from mice injected with one two doses of QIV and administered with antioxidants/immunomodulators (Figure 3b). However, in dLN cultures from mice supplemented with antioxidants/immunomodulators GC CD4+ cell proliferative index was similar in cultures from mice administered with one dose and two doses of QIV (Figure 3b).
Considering that CD4+CXCR5+Bcl-6+ T cells consist of Tfh cells and Tfr cells [38], dLN lymphocytes and splenocytes were simultaneously immunolabeled for CD4, CXCR5, Bcl6 and FoxP3 to delineate CD4+CXCR5+Bcl6+FoxP3- The cells with a critical role in promoting GC B cell proliferation [39,40]. The supplementation with antioxidants/immunomodulators increased the frequency of Tfh cells in dLNs and spleens of mice injected with one dose of QIV (Figure 4a). In mice administered with saline, 14 days post immunisation with the second dose of QIV the frequency of Tfh cells was higher in dLNs and spleens compared with their frequency at the time of the vaccine injection (28 days post the first injection), but this increase reached statistical significance (p≤0.01) only in spleens (Figure 4a). Supplementation with antioxidants/immunomodulators did not influence their frequency statistically significantly in either dLNs or spleens from mice administered with two doses of QIV (Figure 4b). However, it is noteworthy that in mice supplemented with antioxidants/immunomodulators the frequency of Tfh cells was higher (p≤0.01) in spleens from mice administered with two injections of QIV compared with those administered with one injection of the vaccine (Figure 4a).
Besides, the frequency of CD4+CXCR5+Bcl6+FoxP3+ Tfr cells shown to inhibit Tfh cell proliferation and the production of IL-21 [21,41] which, in turn, regulates their development and function, and thereby maintenance of an optimal Tfr/Tfh cell ratio [42], was also determined. The supplementation with antioxidants/immunomodulators did not influence on their frequency in SLOs of mice injected with either one or two doses of QIV (Figure 4b). In mice immunized with two doses of QIV and administered with saline the frequency of Tfr cells was higher in dLNs and spleens compared with their counterparts immunized with one dose of the vaccine, but this increase reached statistical significance (p≤0.01) only in spleens (Figure 4b). Thus, their frequency was higher (p≤0.01) in spleen from mice immunized with two doses of QIV and supplemented with antioxidants/immunomodulators compared with their counterparts administered with one dose of the vaccine (Figure 4b).
Next, given that Tfr/Tfh cell ratio is more important for maintaining Tfh cell proliferation and function than the frequency of any of them alone [21,41], this ratio was also investigated. Supplementation with anti-oxidants/immunomodulators shifted (p≤0.01) Tfr/Tfh cell ratio towards Tfh cell in SLOs from mice immunized with one dose of QIV (Figure 4c). Administration of the second dose of QIV increased (p≤0.01) Tfr/Tfh splenocyte ratio 14 days following the second injection of the vaccine, suggesting a formation of a new balance between these cells (Figure 4c). Of note, in mice supplemented with antioxidants/immunomodulators this ratio was also greater (p≤0.01) in mice injected with two doses of QIV than in those injected with one dose of vaccine (Figure 4c).
Considering that Tfr cells are suggested to exert direct suppressive effects on B cell differentiation [43], Tfr/GC B cell ratio was also determined (Figure 4d). In dLNs and spleens of mice injected with one dose of QIV supplementation with antioxidants/immunomodulators shifted (p≤0.01) this ratio towards GC B cells (Figure 4d). In spleens from mice injected with two doses of QIV and administered with saline Tfr/GC B cell ratio was greater (p≤0.01) compared with those injected with one dose of the vaccine and administered with saline (Fig 4d). This suggested a new balance between these two cell subpopulation following the second QIV injection (Fig 4d). The supplementation with antioxidants/immunomodulators did not influence Tfr/GC B cell ratio in SLOs (Figure 4e). Thus, in spleens from mice supplemented with antioxidants/immunomodulators this ratio was greater (p≤0.01) in spleens from mice injected with two doses of QIV compared with dose injected with one dose of the vaccine (Figure 4e).
3.3. Different Effects of Supplementation with Antioxidants/Immunomodulators on the Frequency of IL-21+CD4+ Cells in SLOs from Mice Injected with One and Two Doses of QIV
Considering that during CD4+ T cell-B cell collaboration IL-21 apart from an essential role in B-cell activation, expansion, and plasma cell generation [44, 45], also plays an important role in Tfh cell differentiation trough up-regulation of Bcl6 expression in naive CD4+ T cells [46,47], its expression in CD4+ dLN lymphocytes and splenocytes was examined. The supplementation with antioxidants/immunomodulators increased (p≤0.01) the frequency of IL-21+ cells among CD4+ cells from SLOs (Figure 5a). Fourteen days post the second QIV injection (42 days after the first QIV injection) the frequency of IL-21+ cells among CD4+ cells from SLOs increased (p≤0.01) compared with their frequency at time of the second QIV injection (28 days after the first QIV injection) (Figure 5a). In mice who received two doses of QIV their frequency was comparable in SLOs from those treated with supplement and saline (Figure 5a). However, the frequency of IL-21+ cells was higher (p≤0.01) among CD4+ cells from SLOs of mice injected with two doses of QIV compared with those injected with one dose of the vaccine (Figure 5a).
To confirm that the changes in the frequency of IL-21+ cells among CD4+ lymphocytes in SLOs were related to the response to QIV immunisation, their frequency was investigated in dLN lymphocyte and splenocyte cultures in the presence of QIV antigens and in their absence (under basal condition) and fold increase was calculated. Indeed, in cultures of SLO cells from mice injected with one dose of QIV and supplemented with antioxidants/immunomodulators the increase in the frequency of IL-21+ cells upon restimulated with QIV (over that in the absence of QIV) was greater (p≤0.01) than in the corresponding cultures from mice injected with one dose of QIV and administered with saline (Figure 5b). The increase in the frequency of these cells upon restimulation with QIV was also greater (p≤0.01) in culture of SLO cells from mice immunized with two doses of QIV and supplemented with saline compared with their counterpart administered with only one dose of the vaccine (Figure 5b). However, in cultures of SLO cells from mice immunized with two doses of QIV, the increase in the frequency of IL-21+ cells upon restimulation with QIV was comparable in cultures from mice supplemented with antioxidants/immunomodulators and in those from mice treated with saline (Figure 5b). However, it is noteworthy, that this increase was more prominent (p≤0.01) in cultures from mice immunized with two doses of QIV than in cultures from mice immunized only with one dose of the vaccine (Figure 5b).
3.4. Antioxidant/Immunomodulatory Supplementation Affected IgG Subclass Profile Only in Mice Injected with Two Doses of QIV
Considering CD4+ T cells providing help to B cells, apart from influencing B-cell survival, proliferation and differentiation into plasma cells, directly shape isotype switching [48], IgG subclass profile was examined. The supplementation with antioxidant/immunomodulators did not influence serum titre of QIV antigen-specific either IgG2a or IgG1 response (Figure 6a). In mice injected with two doses of QIV and administered with saline the serum titre of QIV antigen-specific IgG1 and IgG2a antibody was greater (p≤0.05) than in their counterparts injected with one dose of the vaccine (Figure 6a). The supplementation with anti-oxidants/immunomodulators augmented (p≤0.01) only the serum titre of QIV antigen-specific IgG2a antibody (Figure 6a). Accordingly, the serum titre of QIV antigen-specific IgG2a antibody was greater (p≤0.01) in mice injected with two doses of QIV and supplemented with antioxidants/immunomodulators than in mice injected with one dose of QIV and supplemented with antioxidants/immunomodulators (Figure 6a). Consequently, differently from mice injected with one dose of QIV, in mice injected with two doses of the vaccine IgG2a/IgG1 response ratio was shifted (p≤0.05) towards IgG2a response in those supplemented with antioxidants/immunomodulators compared with those administered with saline (Figure 6a). Besides, in supplemented mice this ratio was shifted (p≤0.01) towards IgG2a response in those injected with two doses of QIV compared with mice injected with one dose of the vaccine (Figure 6a).
To unveil the type of cell-mediated immune response as crucial for IgG subclass profile [24], in SLO lymphocyte cultures stimulated with QIV antigens the levels of INF-γ and IL-4 as Th1 and Th2 cell signature cytokine, respectively, were measured [49]. Upon restimulation with QIV antigens the levels of INF-γ and IL-4 increased (p≤0.01) in dLN cell cultures and splenocyte cultures over basal level (level in cultures without QIV antigens), which served as negative controls (Figure 6b). In SLO cultures from mice injected with one dose of QIV the level of INF-γ did not statistically significantly different between cultures from mice supplemented with antioxidant/immunomodulators and from mice treated with saline (Figure 6b). On the other hand, in dLN and splenocyte cultures from mice injected with two doses of QIV the level of INF-γ was higher (p≤0.05 and p≤0.01 in dLN lymphocyte cultures and splenocyte cultures, respectively) in cultures from mice supplemented with antioxidants/immunomodulators than in those from mice treated with saline (Figure 6). Additionally, it was also higher (p≤0.01) in SLO cultures from mice injected with two doses of QIV and either supplemented with antioxidants/immunomodulators or treated with saline compared with corresponding cultures from mice injected with one dose of the vaccine (Figure 6). In SLO cultures from mice injected with either one dose of QIV or two doses of QIV, the level was comparable between cultures from mice supplemented with antioxidants/immunomodulators and treated with saline (Figure 6b). However, IL-4 level was higher in SLO cultures from mice injected with two doses of QIV compared with those from mice injected with one dose of the vaccine, but this increase reached statistical significance (p≤0.01) only in dLN lymphocyte cultures (Fig.6b).
Next, considering that INF-γ/IL-4 secretion ratio rather than absolute secretory level of any of two cytokines may be considered as surrogate marker for IgG2a/IgG1 response ratio [49], their ratio was also examined. In cultures of cells from dLNs and spleens of mice injected with two doses of QIV, this ratio was shifted (p≤0.05) to the side of INF-γ (Figure 6c). Besides, it was higher (p≤0.01) as it was shifted towards INF-γ in SLO lymphocyte culture from supplemented mice injected with two doses of QIV compared with supplemented mice injected with one dose of the vaccine (Figure 6c).
Next, given that redox status is shown to influence Th cell polarization [50], to enlighten putative mechanism standing behind the shift in Th1/Th2 response in mice injected with two doses of QIV and supplemented with antioxidants/immunomodulators, the parameters of redox status were investigated in their spleens. Redox status was determined by measuring O2•−, AOPP, TOC and several other antioxidant markers: SOD, SHG and GSH, which is shown to have a central role in the maintenance of the thiol-disulfide redox state in mammalian cells [51]. Supplementation markedly diminished O2•− level in spleens from mice injected with doses of QIV, but this decrease did not reach statistical significance (Figure 7a). However, TOC was statistically significantly elevated (p≤0.01) in spleens from mice supplemented with antioxidants/immunomodulators compared with mice treated with saline (Figure 7a). Consistently, AOPP level was markedly (p≤0.01) lower in spleens from mice supplemented with antioxidants/immunomodulators compared with those treated with saline (Figure 7a). In accordance with these findings, compared with mice injected with two doses of QIV and administered with saline, in spleens from supplemented mice injected with two doses of QIV were elevated the levels of all antioxidants, but only the changes in SOD and GSH reached statistical significance (p≤0.01) (Figure 7b).
4. Discussion
The study revealed that the supplementation combining the micronutrients with antioxidant/immunomodulatory properties (vitamins C and D, oligoelements zinc, manganese and selenium, and N-acetyl-cysteine), which are at high risk of deficit, leads to different effects in mice who received one dose and two doses (28 days apart) of QIV. The supplementation in mice injected with one dose of QIV increased the frequency of GC B cells in dLNs and spleens enhancing their proliferative response to QIV antigens, most likely, at least partly due to QIV-specific expansion of Tfh cells and augmented production of T cell-derived IL-21, and thereby increased the total serum QIV-specific IgG titre. On the other hand, the supplementation in mice received two doses of QIV, without any significant affection of the total serum QIV-specific IgG titre (which was substantially increased 14 days following the second dose of QIV), led to clear increase in serum QIV-specific IgG2a titre, as a consequence of the shift in Th1/Th2 signature cytokine response ratio towards Th1 (INF-γ); the phenomenon which could be at least partly associated with significant increase in anti-oxidant status in their SLOs.
To explain hereby reported increase in total serum influenza virus-specific IgG titres 28 days post the first dose of QIV, data on influence of each component on antibody response to influenza vaccine/antiviral vaccines were analyzed. The analysis showed that although all components used for supplementation, except for NAC, may affect antibody synthesis and consequently serum IgG titres, data on significance of their supplementation/deficiency for humoral immune response to influenza vaccine (and thereby the vaccine efficacy), are inconclusive and inconsistent or lacking. Specifically, although zinc supplementation is shown to stimulate antibody synthesis [52,53], data from rather limited number of studies investigating antibody responses to influenza vaccine in relation to zinc status are controversial [54], as they indicate that zinc supplementation may increase humoral immune response to influenza vaccine [55], but also that this supplementation is ineffective in this respect [56]. On the other hand, although it has been suggested that supplementation with vitamin D may diminish antibody response [57], there are data indicating that supplementation with this vitamin may be ineffective in modulating antibody response, but also those indicating that it may enhance this response in humans and mice [58-60] or at least this response to some influenza virus strains [61]. The discrepancies in studies of influence of adjuvant capacities of zinc and vitamin D in influenza vaccination could be reconciled by considering differences in many factors that influence outcome of the vaccination and thereby interfere with the comparison of response rates. Thus, it is noteworthy that these studies differ regarding characteristics of study participants, including serum/plasma status of the supplement under investigation, and, in humans, pre-vaccinated influenza virus-specific antibody levels [54,62]. Additionally, in vitamin D studies polymorphisms of gene for vitamin D receptor, may also contribute to the outcome of the vaccination [54,62]. Moreover, supplement dose, its bioavailability and regime of administration may also be a contributing factor [54,62]. It is also of note that, to the best of our knowledge, despite stimulatory influence of vitamin C [63,64], selenium [65] and manganese [66] on antibody synthesis in humans and/or experimental animals, there is no studies on their influence on serological response to influenza virus vaccine. However, there are data indicating that they exert stimulatory influence on antibody response to some other vaccine, including (when selenium is in question) avian influenza virus in humans and/or experimental animals [67-71].
The increase in the total serum QIV-specific IgG titres in immunized mice supplemented with antioxidants/immunomodulators compared with their counterparts administered with saline 28 days post immunization was fully in accordance with higher frequency of GC B cells in their SLOs, which could be associated with their greater proliferation in SLO cell cultures in response to QIV antigen stimulation. The latter could be linked with the higher frequency of Tfh cells, which provide CD40L and IL-21 signals required for B cell proliferation and differentiation [72] and, accordingly lower Tfr/Tfh cell ratio as the frequency of Tfr cells was comparable in SLOs from QIV-immunized mice who were supplemented with antioxidants/immunomodulators and those from their counterparts administered with saline. The higher frequency of Tfh cells in SLOs from QIV-immunized mice supplemented with antioxidants/immunomodulators is consistent with greater proliferative response of GC CD4+CXCR5+ cells to stimulation with QIV antigens in cultures from these animals compared with cultures from their counterparts administered with saline. In favor of these findings are data indicating that vitamin C [73-75], zinc [52,53,76] and selenium [65,77] stimulate B-cell and T-cell differentiation and proliferation resulting in enhanced antibody generation. Given that the promotion of T cell expansion and Tfh cell differentiation by IL-21 is shown to set the GC response magnitude [23], herein presented findings indicating the increased frequency of IL-21-producing T-cells in SLOs from mice supplemented with antioxidants/immunimodulators and injected with one dose of QIV ex vivo and in vitro upon restimulation with QIV compared with their saline-administered counterparts additionally corroborate the augmented proliferation/differentiation of GC T and GC B cells and consequently antibody production in mice supplemented with antioxidants/immunimodulators.
To explain at least partly the lack of stimulatory effect of antioxidant/immunomodulatory supplementation on GC T cell and GC B cell proliferation/differentiation and consequently antibody production in mice administered with two doses of QIV are data indicating that proliferative response of T cells to supplementation with antioxidants/immunomodulators, such as zinc, depends on starting activational status of the lymphocytes, so in low responders this was enhanced whereas in high responders it tended to be reduced [53], most likely implying differential response of QIV-specific naïve and memory cells predominantly responding upon primary and secondary QIV immunization, respectively. Although in mice injected with two doses of QIV the total serum IgG level was comparable between animals supplemented with antioxidants/immunomodulators and saline, herein presented data revealed that the supplementation potentiated the shift in the ratio of IgG2a/IgG1 serum titres evident, as expected [78] upon the second dose of QIV towards IgG2a. Given that ratio in the production levels of IFN-γ (Th1 signature cytokine) and IL-4 (Th2 signature cytokine) is shown to be critical in shapes IgG2a/IgG1 ratio [49,79], herein presented findings showing supplementation-induced changes in their production level ratio in SLO cell cultures restimulated with QIV corroborate the shift in serum QIV-specific IgG2a/IgG1 ratio. To additionally strengthen these findings are data indicating that oxidative stress promotes Th1/Th2 cell ratio toward the latter [80], as a clear shift in prooxidant/antioxidant parameter towards the latter leading to diminished generation of AOPP in spleens from mice injected with two doses of QIV supplemented with antioxidants/immunomodulators compared with their counterparts administered with saline. In the same line are data indicating that components used for the supplementation in the present study known to improve redox imbalance, viz. vitamin C [81,82], zinc [81, 83-85] and selenium [86,87] increase isotype switching to IgG2a in mice and or humans in a redox status independent manner. Given that vitamin D, vitamin with antioxidant properties [88], is acknowledged to shift the Th response profile from a Th1- to a Th2-mediated response [89,90], it may be assumed that herein reported supplementation-induced changes in Th1/Th2 signature cytokine production ratio represent the net effect of antioxidant action of antioxidants used for supplementation and direct action on Th cell differentiation of its components. To fully appreciate the significance of herein presented findings are data indicating that IgG1 and IgG2a have differential functional characteristics [91,92,93]. Specifically, IgG2a antibodies are shown to be more effective at clearing infections than antibodies of the IgG1 isotype, even when each was specific for the same epitope [92]. Thus, it is clear that the efficacy of IgG antibody response does not depend only on magnitude, but also on IgG subclass profile [94].
5. Conclusions
In conclusion, briefly, supplementation with vitamins D and C, oligoelements zinc, selenium and manganese and modified amino acid cysteine NAC (i) augmented the magnitude of the primary IgG antibody response by stimulating GC reaction and optimized the secondary antibody IgG response by favoring more favorable (viz. more effective response in the context of virus infection clearance) IgG2a response, by shifting Th1/Th2 cell ratio on the side of Th1 cells, acting not only indirectly, through modulation of prooxidant/antioxidant balance in SLO, but also directly on Th cell differentiation in SLOs. Thus, the study firms solid base for further studies aimed at repurposing use this preparation in order to be used not only to prevent virus infection and stimulate recovery, but also as adjuvant for virus influenza and possible some other virus vaccines. To add additional weight to this notion are data indicating that nutritional gaps in several micronutrients reported to support immune function, including vitamins A, D, and E, as well as iron, zinc, and selenium are relatively higher prevalent [20]. Thus, supplementation combining the micronutrients at highest risk of deficit should be considered a safe and effective way to prevent or correct inadequacies and reduce the incidence and severity of infective diseases.
Author Contributions
Conceptualization, G.L. and N.A.R.; Methodology, B.B, I.Ž, N.A.R., I.Ć., B.V. and J.K.S.; Validation, B.B. and N.A.R.; Formal Analysis, G.L, B.B, I.Ž. and J.K.S.; Investigation, B.B, N.A.R, I.Ž., J.K.S., I.Ć. and V.B.; Data Curation, G.L., N.A.R; Writing – Original Draft Preparation, G.L. and B.B.; Writing – Review & Editing, G.L, N.A.R, B.B. I.Ž. and J.K.S; Visualization, B.B; Supervision, G.L.; Project Administration, B.B. and G.L.; Funding Acquisition, N.A.R, B.B, I.Ž. and J.K.S.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation, Republic of Serbia through two Grant Agreements with University of Belgrade-Faculty of Pharmacy No 451-03-65/2024-03/ 200161 and No 451-03-66/2024-03/ 200161 and through Grant Agreement with Institute of Virology, Vaccines and Sera "Torlak", Belgrade No 451-03-66/2024-03.
Institutional Review Board Statement
The animal study protocol was evaluated by the Animal Care and Use Committee of Institute of Virology, Vaccines and Sera “Torlak” and approved by The Ministry of Agriculture, Forestry and Water Economy of the Republic of Serbia - Veterinary Directorate (permition number: 323-07-12928/2022-05).
Data Availability Statement
Data is contained within the article.
Acknowledgments
Authors would like to thank AbelaPharm, Belgrade, Serbia for the donation of components for supplementations used in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization (WHO). Influenza (seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal)#:~:text=There%20are%20around%20a%20billion,650%20000%20respiratory%20deaths%20annually (accessed on 18. 3. 2024).
- Buchy, P.; Badur, S. Who and When to Vaccinate against Influenza. Int. J. Infect. Dis. 2020, 93, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; Garcia-Sastre, A. Influenza. Nat. Rev. Dis. Primers. 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.; Palese, P. Overcoming barriers in the path to a universal influenza virus vaccine. Cell. Host. Microbe. 2018, 24, 18–24. [Google Scholar] [CrossRef]
- Dhakal, S.; Klein, S.L. Host Factors Impact Vaccine Efficacy: Implications for Seasonal and Universal Influenza Vaccine Programs. J. Virol. 2019 Nov 1; 93(21), e00797-19. [CrossRef]
- Erbelding, E.J.; Post, D.; Stemmy, E.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A universal influenza vaccine: the strategic plan for the National Institute of Allergy and Infectious Diseases. J. Infect. Dis. 2018, 218(3), 347–354. [Google Scholar] [CrossRef] [PubMed]
- WHO. Ageing and Health Unit. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed 18.3.2024.).
- Goodwin, K.; Viboud, C.; Simonsen, L. Antibody Response to Influenza Vaccination in the Elderly: A Quantitative Review. Vaccine 2006, 24, 1159–1169. [Google Scholar] [CrossRef]
- Jackson, M.L.; Phillips, C.H.; Benoit, J.; Kiniry, E.; Madziwa, L.; Nelson, J.C.; Jackson, L.A. The Impact of Selection Bias on Vaccine. Vaccine 2018, 36(5), 751–757. [Google Scholar] [CrossRef]
- Muller, L.; Andree, M.; Moskorz, W.; Drexler, I.; Walotka, L.; Grothmann, R.; Ptok, J.; Hillebrandt, J.; Ritchie, A.; Rabl, D.; Ostermann, P.N.; Robitzsch, R.; Hauka, S.; Walker, A.; Menne, C.; Grutza, R.; Timm, J.; Adams, O.; Schaal, H. Age-dependent immune response to the Biontech/Pfizer BNT162b2 COVID-19 vaccination. Clin. Infect. Dis. 2021, 73(11), 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef]
- Wouters-Wesseling, W.; Rozendaal, M.; Snijder, M.; Graus, Y.; Rimmelzwaan, G.; De Groot, L.; Bindels, J. Effect of a Complete Nutritional Supplement on Antibody Response to Influenza Vaccine in Elderly People. J. Gerontol. A Biol. Sci. Med. Sci. 2002, M563–M566. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9(11), 1211. [Google Scholar] [CrossRef]
- Hemilä, H.; Fitzgerald, J.T.; Petrus, E.J.; Prasad, A. Zinc acetate lozenges may improve the recovery rate of common cold patients: an individual patient data meta-analysis. Open. Forum. Infect. Dis. 2017, 4(2), ofx059. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.; Sooriyaarachchi, P.; Chourdakis, M.; Jeewandara, C.; Ranasinghe, P. Enhancing immunity in viral infections, with special emphasis on COVID-19: A review. Diabetes Metab. Syndr. 2020, 14(4), 367–382. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary Selenium in Adjuvant Therapy of Viral and Bacterial Infections. Adv. Nutr. 2015, 6(1), 73–82. [Google Scholar] [CrossRef]
- Schloss, J.; Leach, M.; Brown, D.; Hannan, N.; Kendall-Reed, P.; Steelc, A. The effects of N-acetyl cysteine on acute viral respiratory infections in humans: A rapid review. Adv. Integr. Med. 2020, 7(4): 232–239. [CrossRef]
- Weiss, G.; Carver, P.L. Role of divalent metals in infectious disease susceptibility and outcome. Clin. Microbiol. Infect. 2018, 24, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System—Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12(1), 236. [Google Scholar] [CrossRef]
- Calder, P.C.; Berger, M.M.; Gombart, A.F.; McComsey, G.A.; Martineau, A.R.; Eggersdorfer, M. Micronutrients to Support Vaccine Immunogenicity and Efficacy. Vaccines 2022, 10(4), 568. [Google Scholar] [CrossRef]
- Sage, P.T.; Alvarez, D.; Godec, J.; von Andrian, U.H.; Sharpe, A.H. Circulating T follicular regulatory and helper cells have memory-like properties. J. Clin. Invest. 2014, 124(12), 5191–5204. [Google Scholar] [CrossRef]
- Wing, J.B.; Tekgüç, M.; Sakaguchi, S. Control of germinal center responses by Tfollicular regulatory cells. Front. Immunol. 2018, 9, 1910. [Google Scholar] [CrossRef]
- Quast, I.; Dvorscek, A.R.; Pattaroni, C.; Steiner, T. M.; McKenzie, C. I.; Pitt, C.; O’Donnel, K.; Ding, Z.; Hill, D. L.; Brink, R.; Robinson, M.J.; Zotos, D.; Tarlinton, D.M. Interleukin-21, acting beyond the immunological synapse, independently controls T follicular helper and germinal center B cells. Immunity, 2022; 55, 1414–1430.e5. [Google Scholar] [CrossRef]
- Stevens, T.L.; Bossie, A.; Sanders, V.L.; Fernandez-Botran, R. , Coffman, R.L.; Mosmann, T.R.; Vitetta, E.S. Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature, 1988; 334, 255–258. [Google Scholar] [CrossRef]
- Jacob, S.; Nair, A.B.; Morsy, M.A. Dose Conversion Between Animals and Humans: A Practical Solution. Indian J. Pharm. Educ. Res. 2002, 56, 600–607. [Google Scholar] [CrossRef]
- Petrović, R.; Bufan, B.; Arsenović-Ranin, N.; Živković, I.; Minić, R.; Radojević, K.; Leposavić, G. Mouse strain and sex as determinants of immune response to trivalent influenza vaccine. Life Sci. 2018, 207, 117–126. [Google Scholar] [CrossRef]
- Alvarez, D.F.; Helm, K.; DeGregori, J.; Roederer, M.; Majka, S. Publishing flow cytometry data. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2010, 298, L127–L130. [Google Scholar] [CrossRef]
- Auclair, C.; Voisin, E. Nitroblue tetrazolium reduction. In Handbook of methods for oxygen radical research, 1st ed.; Greenwald, R.A., Ed.; Publisher: CRC Press, Boca Raton, USA, 1985; pp. 123–132. [Google Scholar] [CrossRef]
- Kotur-Stevuljevic, J.; Bogavac-Stanojevic, N.; Jelic-Ivanovic, Z.; Stefanovic, A.; Gojkovic, T.; Joksic, J.; Sopic, M.; Gulan, B.; Janac, J.; Milosevic, S. Oxidative stress and paraoxonase 1 status in acute ischemic stroke patients. Atherosclerosis 2015, 241, 192–198. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney. Int. 1996, 49(5), 1304–1313. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in thautoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247(10), 3170–3175. [Google Scholar] [CrossRef]
- Dimitrijević, M.; Kotur-Stevuljević, J.; Stojić-Vukanić, Z.; Vujnović, I.; Pilipović, I.; Nacka-Aleksić, M.; Leposavić, G. Sex differencein oxidative stress parameters in spinal cord of rats with experimental autoimmune encephalomyelitis: relation to neurological deficit. Neurochem. Res. 2017, 42, 481–492. [Google Scholar] [CrossRef]
- Hauge, S.; Madhun, A.; Cox, R.G.; Haaheim, L.R. Quality and Kinetics of the Antibody Response in Mice after Three Different Low-Dose Influenza Virus Vaccination Strategies. Clin. Vaccine. Immunol. 2007, 14(8), 978–983. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Yan, J.; Xiao, Z.; Hou, X.; Lu, P.; Hou, S.; Mao, T.; Liu, W.; Ma, Y.; Zhang, L.; Yang, X.; Qi, H. Germinal-center development of memory B cells driven by IL-9 from follicular helper T cells. Nat. Immunol. 2017, 18, 921–930. [Google Scholar] [CrossRef]
- Shinall, S.M.; Gonzalez-Fernandez, M.; Noelle, R.J; Waldschmid, T.J. Identification of Murine Germinal Center B Cell Subsets Defined by the Expression of Surface Isotypes and Differentiation Antigens. J. Immunol. 2000, 164(11), 5729–5738. [Google Scholar] [CrossRef]
- Minton, K. Germinal-centre B cells take control. Nat. Rev. Immunol. 2008, 8, 826–827. [Google Scholar] [CrossRef]
- Stebegg, M.; Kumar, S.D.; Silva-Cayetano, A.; Fonseca, V.R.; Linterman, M.A.; Graca, L. The initiation of the GC requires both CD4+ T cells and B cells to be activated by cognate antigen. Front. Immunol. 2018, 9, 2469. [Google Scholar] [CrossRef]
- Yeh, C-H.; Finney, J.; Okada, T.; Kurosaki, T.; Kelsoe, G. Primary germinal center-resident T follicular helper cells are a physiologically distinct subset of CXCR5hiPD-1hi T follicular helper cells. Immunity, 2022; 55, 272–289.e7. [CrossRef]
- Kaneko, N.; Kuo, H-H.; Boucau, J.; Farmer, J.R.; Allard-Chamard, H.; Mahajan, V.S., Alicj Piechocka-Trocha, A.; Lefteri, K.; Osborn, M.; Bals, J.; Bartsch, Y.S.; Bonheur, N.; Caradonna, T.M.; Chevalier, J.; Chowdhury, F.; Diefenbach, T.J.; Einkauf, K.; Fallon, J.; Feldman, J.; Finn, K.K.; Pillai, S. Loss of Bcl-6-Expressing T Follicular Helper Cells and Germinal Centers in COVID-19. Cell 2020, 183, 143-157.e13. [CrossRef]
- Krishnaswamy, J.K.; Alsén, S.; Yrlid, U.; Eisenbarth, S.C.; Williams, A. Determination of T Follicular Helper Cell Fate by Dendritic Cells. Front. Immunol. 2018, 9, 2169. [Google Scholar] [CrossRef]
- Sage, P.T.; Ron-Harel, N.; Juneja, V.R.; Sen, D.R.; Maleri, S.; Sungnak, W.; Kuchroo, V.K.; Haining, W.N.; Chevrier, N.; Haigis, M.; Sharpe, A.H. Suppression by TFR cells leads to durable and selective inhibition of B cell effector function, Nat. Immunol. 2016, 17, 1436–1446. [Google Scholar] [CrossRef]
- Ding, T.; Niu, H.; Zhao, X.; Gao, C.; Li, X.; Wang, C. T Follicular Regulatory Cells: Potential Therapeutic Targets in Rheumatoid Arthritis. Front. Immunol. 2019, 10, 2709. [Google Scholar] [CrossRef]
- Lopez-Ocasio, M.; Buszko, M.; Blain, M.; Wang, K.; Shevach, E.M. T Follicular Regulatory Cell Suppression of T Follicular Helper Cell Function Is Context-Dependent in vitro. Front. Immunol. 2020, 17, 637. [Google Scholar] [CrossRef]
- Kuchen, S.; Robbins, R.; Sims, G.P.; Sheng, C.; Phillips, T.M.; Lipsky, P.E.; Ettinger, R. Essential role of IL-21 in B cell activation, expansion, and plasma cell generation during CD4+ T cell-B cell collaboration. J. Immunol. 2007, 179, 5886–5896. [Google Scholar] [CrossRef]
- Linterman, M.A.; Beaton, L.; Yu, D.; Ramiscal, R.R.; Srivastava, M.; Hogan, J.J.; Verma, N.K.; Smyth, M.J.; Rigby, R.J.; Vinuesa, C.G. IL-21 acts directly on B cells to regulate Bcl-6 expression and germinal center responses, J. Exp. Med. 2010, 207, 353–363. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Hwang, D.; Yang, X.O.; Kang, H.S.; Ma, L.; Wang, Y.H.; Watowich, S.S.; Jetten, A.M.; Tian, Q.; Dong, C. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 2008, 29, 138–149. [Google Scholar] [CrossRef]
- Vogelzang, A.; McGuire, H.M.; Yu, D.; Sprent, J.; Mackay, C.R.; King, C. A fundamental role for interleukin-21 in the generation of T follicular helper cells. Immunity 2008, 29, 127–137. [Google Scholar] [CrossRef]
- Grodeland, G.; Fossum, E.; Bogen, B. Polarizing T and B Cell Responses by APC-Targeted Subunit Vaccines. Front. Immunol. 2015, 6, 367. [Google Scholar] [CrossRef]
- Daifalla, N.S.; Bayih, A.G.; Gedamu, L. Differential Immune Response against Recombinant Leishmania donovani Peroxidoxin 1 and Peroxidoxin 2 Proteins in BALB/c Mice. J. Immunol. Res. 2015, 348401. [Google Scholar] [CrossRef]
- King, M.R.; Ismail, A.S.; Davis, L.S.; Karp, D.R. Oxidative stress promotes polarization of human T cell differentiation toward a T helper 2 phenotype. J. Immunol. 2006, 176(5), 2765–2772. [Google Scholar] [CrossRef]
- Miller, L.T.; Watson, W.H.; Kirlin, W.G.; Ziegler, T.R.; Jones, D.P. Oxidation of the glutathione/glutathione disulfide redox state is induced by cysteine deficiency in human colon carcinoma HT29 cells. J. Nutr. 2002, 132(8), 2303–2306. [Google Scholar] [CrossRef] [PubMed]
- Cunningham-Rundles, S.; Cunningham-Rundles, C.; Dupont, B. Zinc-induced activation of human B lymphocytes. Clin. Immunol. Immunopathol. 1980, 16, 115–22. [Google Scholar] [CrossRef]
- Duchateau, J.; Delespesse, G.; Vereecke, P. Influence of oral zinc supplementation on lymphocyte response to mitogensnormal subjects. Am. J. Clin. Nutr. 1981, 34, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Sadeghsoltani, F.; Mohammadzadeh, I.; Safari, M-M.; Hassanpour, P.; Izadpanah, M.; Qujeq, D.; Moein, S.; Vaghari-Tabar, M. Zinc and Respiratory Viral Infections: Important Trace Element in Anti-viral Response and Immune Regulation. Biol. Trace. Elem. Res. 2022, 200, 2556–2571. [CrossRef]
- Provinciali, M.; Montenovo, A.; Di Stefano, G.; Colombo, M.; Daghetta, L.; Cairati, M.; Veroni, C.; Cassino, R.; Della Torre, F.; Fabris, N. Effect of zinc or zinc plus arginine supplementation on antibody titre and lymphocyte subsets after influenza vaccination in elderly subjects: a randomized controlled trial. Age Ageing 1998, 27(6), 715–722. [Google Scholar] [CrossRef]
- Türk, S.; Bozfakioğlu, S.; Ecder, S.T.; Kahraman, T.; Gürel, N.; Erkoç, R.; Aysuna, N.; Türkmen, A.; Bekiroğlu, N.; Ark, E. Effects of zinc supplementation on the immune system and on antibody response to multivalent influenza vaccine in hemodialysis patients. Int. J. Artif. Organs. 1998, 21(5), 274–278. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J. M.; Adams, J. S. ; R. Sakai, R.; Jordan, S.C. 1α,25-Dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J. Clin. Invest. 1984; 74, 657–661. [Google Scholar] [CrossRef]
- Govaert, M.T.; Thijs, C.T.; Masurel, N.; Sprenger, M.J.; Dinant, G. J.; Knottnerus, J.A. The Efficacy of Influenza Vaccination in Elderly Individuals: A Randomized Double-blind Placebo-Controlled Trial. JAMA 1994, 272, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- Principi, N.; Marchisio, P.; Terranova, L.; Zampiero, A.; Baggi, E.; Daleno, C.; Tirelli, S.; Pelucchi, C.; Esposito, S. Impact of vitamin D administration on immunogenicity of trivalent inactivated influenza vaccine in previously unvaccinated children. Hum. Vaccin. Immunother. 2013, 9, 969–974. [Google Scholar] [CrossRef]
- Daynes, R.A.; Araneo, B.A.; Hennebold, J.; Enioutina, E.; Mu, H.H. Steroids as regulators of the mammalian immune response. J. Invest. Dermatol. 1995, 105(1 Suppl), 14S–19S. [Google Scholar] [CrossRef]
- Lee, M-D.; Lin, C-H.; Lei, W-T.; Chang, H-Y.; Lee, H-C.; Yeung, C-Y.; Chiu, N-C.; Chi, H.; Liu, J-M.; Hsu, R.J.; Cheng, Y-J.; Yeh, T-L., Lin, C-J. Does Vitamin D Deficiency Affect the Immunogenic Responses to Influenza Vaccination? A Systematic Review and Meta-Analysis. Nutrients 2018, 10(4), 409. [CrossRef]
- Overbeck, S.; Rink, L.; Haase, H. Modulating the immune response by oral zinc supplementation: a single approach for multiple diseases. Arch. Immunol. Ther. Exp. 2008, 56, 15–30. [Google Scholar] [CrossRef]
- Prinz, W.; Bloch, J.; Gilich, G.; Mitchell, G. A systematic study of the effect of vitamin C supplementation on the humoral immune response in ascorbate-dependent mammals. I. The antibody response to sheep red blood cells (a T-dependent antigen) in guinea pigs. Int. J. Vitam. Nutr. Res. 1980, 50, 294–300.
- Anderson, R.; Oosthuizen, R. , Maritz, R., Theron, A.; Van Rensburg, A.J. The effects of increasing weekly doses of ascorbate on certain cellular and humoral immune functions in normal volunteers. Am. J. Clin. Nutr. 1980, 33, 71–76. [Google Scholar] [CrossRef]
- Carlson, B.A.; Yoo, M.-H.; Shrimali, R.K.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Role of selenium-containing proteins in T-cell and macrophage function. Proc. Nutr. Soc. 2010, 69, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, C.; Guan, Y.; Wei X.; Sha, M.; Yi, M.; Jing, M.; Lv, M.; Guo, W.; Xu, J.; Wan, Y.; Jia X-M.; Jiang, Z. Manganese salts function as potent adjuvants. Cell. Mol. Immunol. 2021, 18, 1222–1234. [Google Scholar] [CrossRef]
- Otomaru, K.; Oishi, S.; Fujimura, Y.; Iwamoto, Y.; Nagai, K.; Ijiri, M. Effects of vitamin C supplementation on the blood oxidative stress and antibody titre against Histophilus somni vaccination in calves. J. Vet. Res. 2021, 65, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; He, M.; Kang, Y. Vitamin C supplementation improved the efficacy of foot-and-mouth disease vaccine. Food Agricul. Immunol. 2018, 29:1, 470-483. [CrossRef]
- Demircan, K.; Chillon, T.S.; Sun, Q.; Heller, A.R.; Klingenberg, G.J.; Hirschbil-Bremer, I.M.; Seemann, P.; Diegmann, J.; Bachmann, M.; Moghaddam, A.; Schomburg, L. Humoral immune response to COVID-19 mRNA vaccination in relation to selenium status. Redox Biol. 2022, 50, 102242. [Google Scholar] [CrossRef]
- Shojadoost, B.; Taha-Abdelaziz, K.; Alkie, T.N.; Bekele-Yitbarek, A.; Barjesteh, N.; Laursen, A.; Smith, T.K.; Shojadoost, J.; Shari, S. Supplemental dietary selenium enhances immune responses conferred by a vaccine against low pathogenicity avian influenza virus. Vet. Immunol. Immunopathol. 2020, 227, 110089. [Google Scholar] [CrossRef]
- Cui, C.; Wang, S.; Lu, W.; Wang, Y.; Li, J.; Qu, K.; Yang, M.; Wang, L.; Yu, Y. The adjuvanticity of manganese for microbial vaccines via activating the IRF5 signaling pathway. Biochem. Pharmacol. 2021, 192, 114720. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef] [PubMed]
- Huijskens, M.J.; Walczak, M.; Koller, N., Briede, J.J.; Senden-Gijsbers, B.L.; Schnijderberg, M.C.; Bos, G.M.; Germeraad, W.T. Technical advance: Ascorbic acid induces development of double-positive T cells from human hematopoietic stem cells in the absence of stromal cells. J. Leukoc. Biol. 2014, 96, 1165–1175. [CrossRef]
- Molina, N.; Morandi, A.C.; Bolin, A.P.; Otton, R. Comparative effect of fucoxanthin and vitamin C on oxidative and functional parameters of human lymphocytes. Int. Immunopharmacol. 2014, 22, 41–50. [Google Scholar] [CrossRef]
- Manning, J.; Mitchell, B.; Appadurai, D.A.; Shakya, A.; Pierce, L.J.; Wang, H.; Nganga, V.; Swanson, P.C.; May, J.M.; Tantin, D., Spangrude, G.J. Vitamin C promotes maturation of T-cells. Antioxid. Redox Signal. 2013, 19, 2054–2067. [CrossRef]
- Winchurch, R.A.; Togo, J.; Adler, W.H. Supplemental zinc restores antibody formation in cultures of aged spleen cells. Effects of mediator production. Eur. J. Immunol. 1987, 17, 127–131. [Google Scholar] [CrossRef]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary selenium modulates activation and differentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Coutelier, J.P.; van der Logt, J.T.; Heessen, F.W. IgG subclass distribution of primary and secondary immune responses concomitant with viral infection. J. Immunol. 1991, 147, 1383–1386. [Google Scholar] [CrossRef]
- Reinhardt, R.L.; Liang, H.E.; Locksley, R.M. Cytokine-secreting follicular T cells shape the antibody repertoire. Nat. Immunol. 2009, 10, 385–393. [Google Scholar] [CrossRef]
- King, M.R.; Ismail, A.S.; Davis, L.S.; Karp, D.R. Oxidative stress promotes polarization of human T cell differentiation toward a T helper 2 phenotype. J. Immunol. 2006, 176(5), 2765–2772. [Google Scholar] [CrossRef] [PubMed]
- Woo, A.; Kim, J. H.; Jeong, Y. J.; Maeng, H. G.; Lee, Y. T.; Kang, J.S. Vitamin C acts indirectly to modulate isotype switching in mouse B cells. Anat. Cell Biol. 2010, 43, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, H-Y.; Almonte-Loya, A.; Lay, F-Y.; Hsu, M.; Johnson, E.; González-Avalos, E.; Yin, J.; Bruno, R.S.; Ma, Q.; Ghoneim, H.E.; Wozniak, D.J.; Harrison, F.E.; Lio, C-W.J. Epigenetic remodeling by vitamin C potentiates plasma cell differentiation. eLife 2022, 11, e73754. [CrossRef]
- Salas, M.; Kirchner, H. Induction of interferon-γ in human leukocyte cultures stimulated by Zn2+. Clin. Immunol. Immunopathol. 1987, 45, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Effects of zinc deficiency on Th1 and Th2 cytokine shifts. J. Infect. Dis. 2000, 182(1), S62–S68. [Google Scholar] [CrossRef]
- Bao, B.; Prasad, A.S.; Beck, F.W.; Godmere, M. Zinc modulates mRNA levels of cytokines. Am. J. Physiol. Endocrinol. Metab. 2003, 285, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.A.; Nelson, H.K.; Shi, Q.; Van Dael, P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A. Selenium deficiency increases the pathology of an influenza virus infection. FASEB J. 2001, 15, 1481–1483. [Google Scholar] [CrossRef]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary selenium modulates activation and differentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar] [CrossRef]
- Sepidarkish, M.; Farsi, F.; Akbari-Fakhrabadi, M.; Namazi, N.; Almasi-Hashiani, A.; Hagiagha, A.M.; Heshmati, J. The effect of vitamin D supplementation on oxidative stress parameters: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2019, 139, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Aranow, C. Vitamin D and the Immune System. J. Investig. Med. 2011, 59(6), 881–886. [Google Scholar] [CrossRef]
- Sundaram, M.E.; Coleman, L.A. Vitamin D and Influenza. Adv. Nutr. 2012, 3, 517–525. [Google Scholar] [CrossRef] [PubMed]
- DiLillo, D.J.; Tan, G.S.; Palese, P.; Ravetch, J.V. ; Broadly neutralizing hemagglutinin stalk-specific antibodies require FcγR interactions for protection against influenza virus in vivo. Nat. Med. 2014, 20(2), 143–151. [Google Scholar] [CrossRef]
- Huber, V.C.; McKeon, R.M.; Brackin, M.N.; Miller, L.A.; Keating, R.; Brown, S.A.; Makarova, N.; Perez, D.R.; Macdonald, G.H.; McCullers, J.A. Distinct contributions of vaccine-induced immunoglobulin G1 (IgG1) and IgG2a antibodies to protective immunity against influenza. Clin. Vaccine Immunol. 2006, 13(9), 981–990. [Google Scholar] [CrossRef]
- Schmitz, N.; Beerli, R.R.; Bauer, M.; Jegerlehner, A.; Dietmeier, K.; Maudrich, M., Pumpens, P.; Saudan, P.; Bachmann, M.F. Universal vaccine against influenzavirus: linking TLR signaling to anti-viral protection. Eur. J. Immunol. 2012, 42(4), 863–869. [CrossRef]
- Potluri, T.; Fink, A.L.; Sylvia, K.E.; Dhakal, S.; Vermillion, M.S.; Vom Steeg, L.; Deshpande, S.; Narasimhan, H.; Klein, S.L. Age-associated changes in theimpact of sex steroids on influenza vaccine responses in males and females. NPJ Vaccines 2019, 4, 29. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Immunization, treatment, blood/organs collection and analysis timeline. The mice were immunized intramuscularly with two doses (D) of QIV (Vx) or saline (Sal) on days 0 (1D) and 28 (2D). Per os daily treatment with antioxidants/immunomodulators in saline (Suppl) or administration of same volume of saline alone (Sal) started five days before the first injection of QIV or saline and lasted until the day of sacrifice (mice were sacrificed on the days 28 and 42 days post the first injection of QIV). Blood was collected on the days 28 and 42 after the first injection (14 days after the second injection) of QIV and analyzed using ELISA. Draining lymph nodes (dLNs) and spleens were harvested on the days 14 and 42 after the first injection (14 days after the second injection) of QIV, and dLN lymphocytes and splenocytes were separated and analyzed by flow cytometry. Redox status was analyzed in spleens on the day 42 (14 days after the second injection of QIV).
Figure 1.
Immunization, treatment, blood/organs collection and analysis timeline. The mice were immunized intramuscularly with two doses (D) of QIV (Vx) or saline (Sal) on days 0 (1D) and 28 (2D). Per os daily treatment with antioxidants/immunomodulators in saline (Suppl) or administration of same volume of saline alone (Sal) started five days before the first injection of QIV or saline and lasted until the day of sacrifice (mice were sacrificed on the days 28 and 42 days post the first injection of QIV). Blood was collected on the days 28 and 42 after the first injection (14 days after the second injection) of QIV and analyzed using ELISA. Draining lymph nodes (dLNs) and spleens were harvested on the days 14 and 42 after the first injection (14 days after the second injection) of QIV, and dLN lymphocytes and splenocytes were separated and analyzed by flow cytometry. Redox status was analyzed in spleens on the day 42 (14 days after the second injection of QIV).
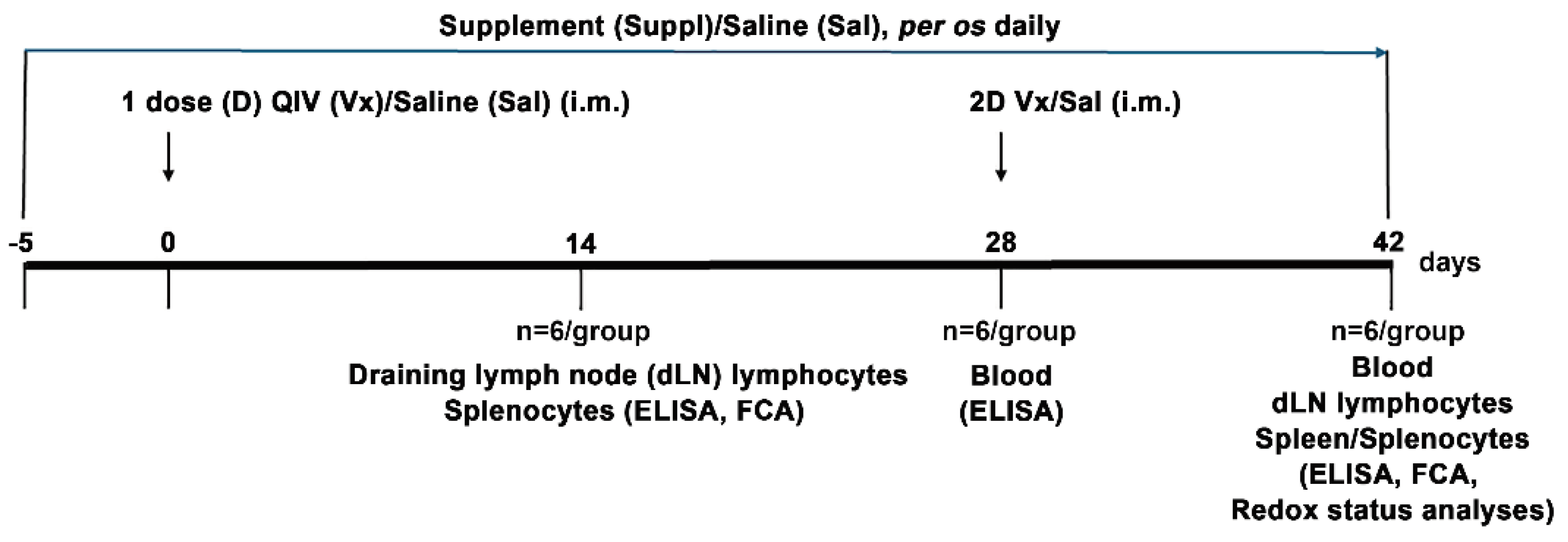
Figure 2.
Supplementation with antioxidants/immunomodulators increased influenza virus-specific IgG response in mice injected with one dose of QIV, the frequency of GC B cells in dLNs and spleens. a) Graph bar indicates serum geometric mean titres of the influenza virus-specific IgG in mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and treated with antioxidant/immunomodulatory supplement (Suppl) or saline (Sal), and in mice injected intramuscularly with Sal and treated per os with Suppl (control group, Ctr). Titres were determined 28 days after the first injection of QIV/Sal (1D) and 14 days after the second injection of QIV/Sal (2D). Error bars indicate the 95% confidence interval (CI). n=6 mice/group. b) Graph bars display the percentages of B220+CD95+GL7+ cells (GC B cells) among (upper row) draining lymph nodes (dLN) lymphocytes and (lower row) splenocytes from mice 14 days after 1D or 2D of QIV (Vx) in mice administered with Suppl or Sal. Flow cytometry dot plots indicate gating strategy for flow cytometry analyses of GC B cells among (upper row) dLN lymphocytes and (lower row) splenocytes. Flow cytometry dot plots indicate (right) GL7 vs CD95 staining of B220+ cells gated as shown in middle flow cytometry dot plots from (left) “lymphocyte” gate. c) The graph bars indicate proliferative index of GC B cells, i.e the increase in the frequency of proliferating Ki67+ cells among B220+CD95+ cells in the presence of QIV antigens over that in their absence in cultures of (upper row) dLN lymphocytes and (lower row) splenocytes from mice injected with 1D and 2D of QIV (Vx) and administered with Suppl or Sal. Flow cytometry dot plots indicate gating strategy for flow cytometry analysis of the frequency of Ki67+ cells among (upper row) B220+CD95+ dLN lymphocyte and (lower row) splenocyte cultures. Flow cytometry dot plots indicate (right) Ki67 vs CD95 staining of B220+ cells gated as shown middle dot plots. B220+ cells were gated within “lymphocytes” as shown in left flow cytometry dot plots. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
Figure 2.
Supplementation with antioxidants/immunomodulators increased influenza virus-specific IgG response in mice injected with one dose of QIV, the frequency of GC B cells in dLNs and spleens. a) Graph bar indicates serum geometric mean titres of the influenza virus-specific IgG in mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and treated with antioxidant/immunomodulatory supplement (Suppl) or saline (Sal), and in mice injected intramuscularly with Sal and treated per os with Suppl (control group, Ctr). Titres were determined 28 days after the first injection of QIV/Sal (1D) and 14 days after the second injection of QIV/Sal (2D). Error bars indicate the 95% confidence interval (CI). n=6 mice/group. b) Graph bars display the percentages of B220+CD95+GL7+ cells (GC B cells) among (upper row) draining lymph nodes (dLN) lymphocytes and (lower row) splenocytes from mice 14 days after 1D or 2D of QIV (Vx) in mice administered with Suppl or Sal. Flow cytometry dot plots indicate gating strategy for flow cytometry analyses of GC B cells among (upper row) dLN lymphocytes and (lower row) splenocytes. Flow cytometry dot plots indicate (right) GL7 vs CD95 staining of B220+ cells gated as shown in middle flow cytometry dot plots from (left) “lymphocyte” gate. c) The graph bars indicate proliferative index of GC B cells, i.e the increase in the frequency of proliferating Ki67+ cells among B220+CD95+ cells in the presence of QIV antigens over that in their absence in cultures of (upper row) dLN lymphocytes and (lower row) splenocytes from mice injected with 1D and 2D of QIV (Vx) and administered with Suppl or Sal. Flow cytometry dot plots indicate gating strategy for flow cytometry analysis of the frequency of Ki67+ cells among (upper row) B220+CD95+ dLN lymphocyte and (lower row) splenocyte cultures. Flow cytometry dot plots indicate (right) Ki67 vs CD95 staining of B220+ cells gated as shown middle dot plots. B220+ cells were gated within “lymphocytes” as shown in left flow cytometry dot plots. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
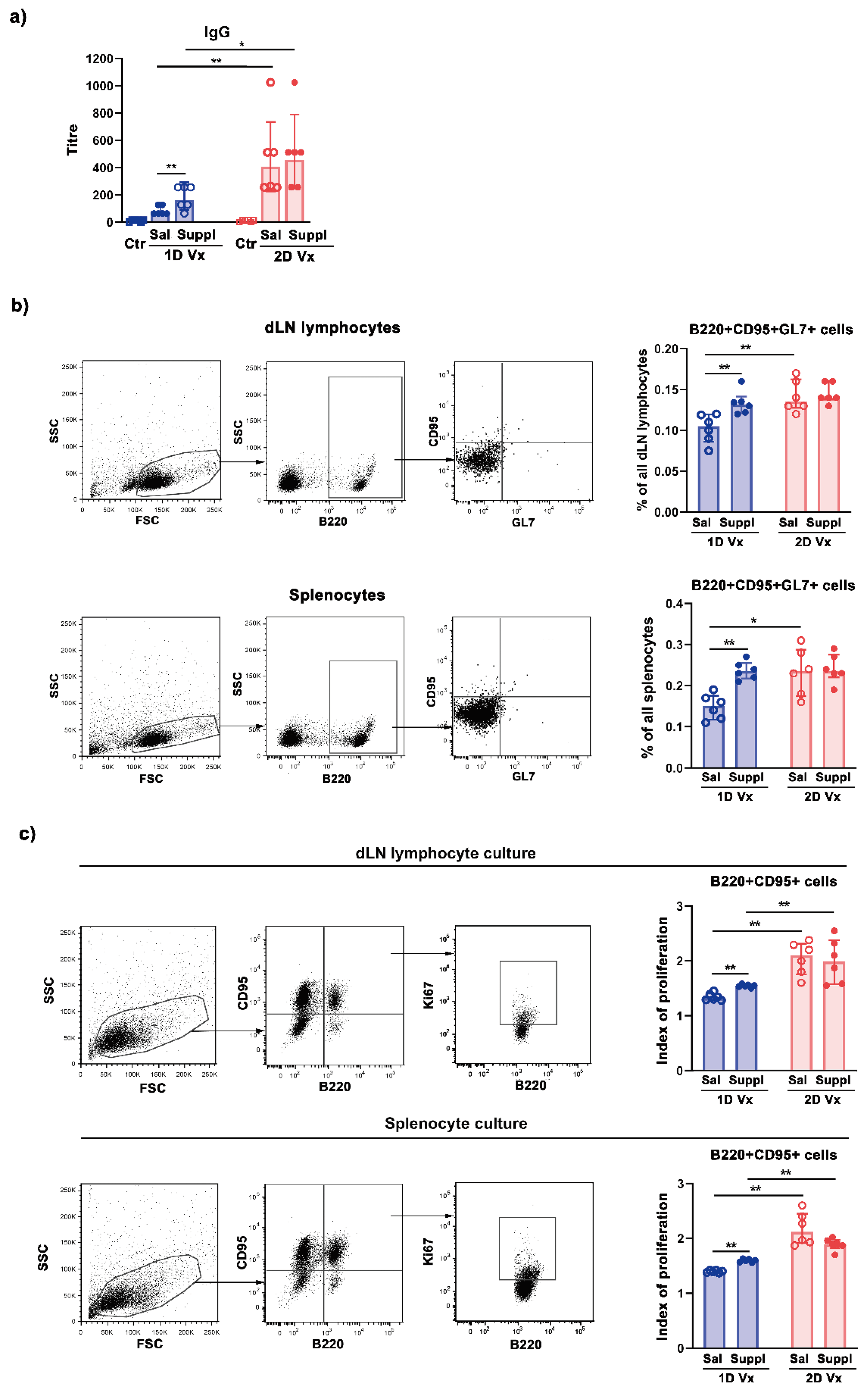
Figure 3.
The supplementation with antioxidants/immunomodulators increased the frequency of GC CD4+ T cells in dLNs and spleens from mice injected with one dose of QIV. a) The graph bars display the percentages of GC CD4+CXCR5+Bcl6+ T cells among (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) determined by flow cytometry. The analyses were performed 14 days after injection of the first and second dose of QIV. The flow cytometry dot plots indicate gating strategy for GC CD4+CXCR5+Bcl6+ T cells among (upper row) dLN lymphocytes and (lower row) splenocytes. (Right) CXCR5 vs Bcl6 staining of CD4+ cells gated as indicated in (left) flow cytometry dot plot. CD4+ cells were gated within lymphocyte gates shown in Fig 2b. b) The graph bars display the proliferative index of GC CD4+CXCR5+ T cells, i.e the increase in frequency of proliferating Ki67+ cells among GC CD4+CXCR5+ T cells in the presence of QIV antigens over that in their absence in cultures of (upper row) dLN lymphocytes and (lower row) splenocytes from mice injected with 1D and 2D of QIV (Vx) and administered with Supp or Sal in response to restimulation with QIV antigens. Flow cytometry dot plots indicate Ki67+ staining of CD4+ CXCR5+ (upper row) dLN lymphocytes and (lower row) splenocytes from lymphocyte gates shown in Fig 2c. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
Figure 3.
The supplementation with antioxidants/immunomodulators increased the frequency of GC CD4+ T cells in dLNs and spleens from mice injected with one dose of QIV. a) The graph bars display the percentages of GC CD4+CXCR5+Bcl6+ T cells among (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) determined by flow cytometry. The analyses were performed 14 days after injection of the first and second dose of QIV. The flow cytometry dot plots indicate gating strategy for GC CD4+CXCR5+Bcl6+ T cells among (upper row) dLN lymphocytes and (lower row) splenocytes. (Right) CXCR5 vs Bcl6 staining of CD4+ cells gated as indicated in (left) flow cytometry dot plot. CD4+ cells were gated within lymphocyte gates shown in Fig 2b. b) The graph bars display the proliferative index of GC CD4+CXCR5+ T cells, i.e the increase in frequency of proliferating Ki67+ cells among GC CD4+CXCR5+ T cells in the presence of QIV antigens over that in their absence in cultures of (upper row) dLN lymphocytes and (lower row) splenocytes from mice injected with 1D and 2D of QIV (Vx) and administered with Supp or Sal in response to restimulation with QIV antigens. Flow cytometry dot plots indicate Ki67+ staining of CD4+ CXCR5+ (upper row) dLN lymphocytes and (lower row) splenocytes from lymphocyte gates shown in Fig 2c. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
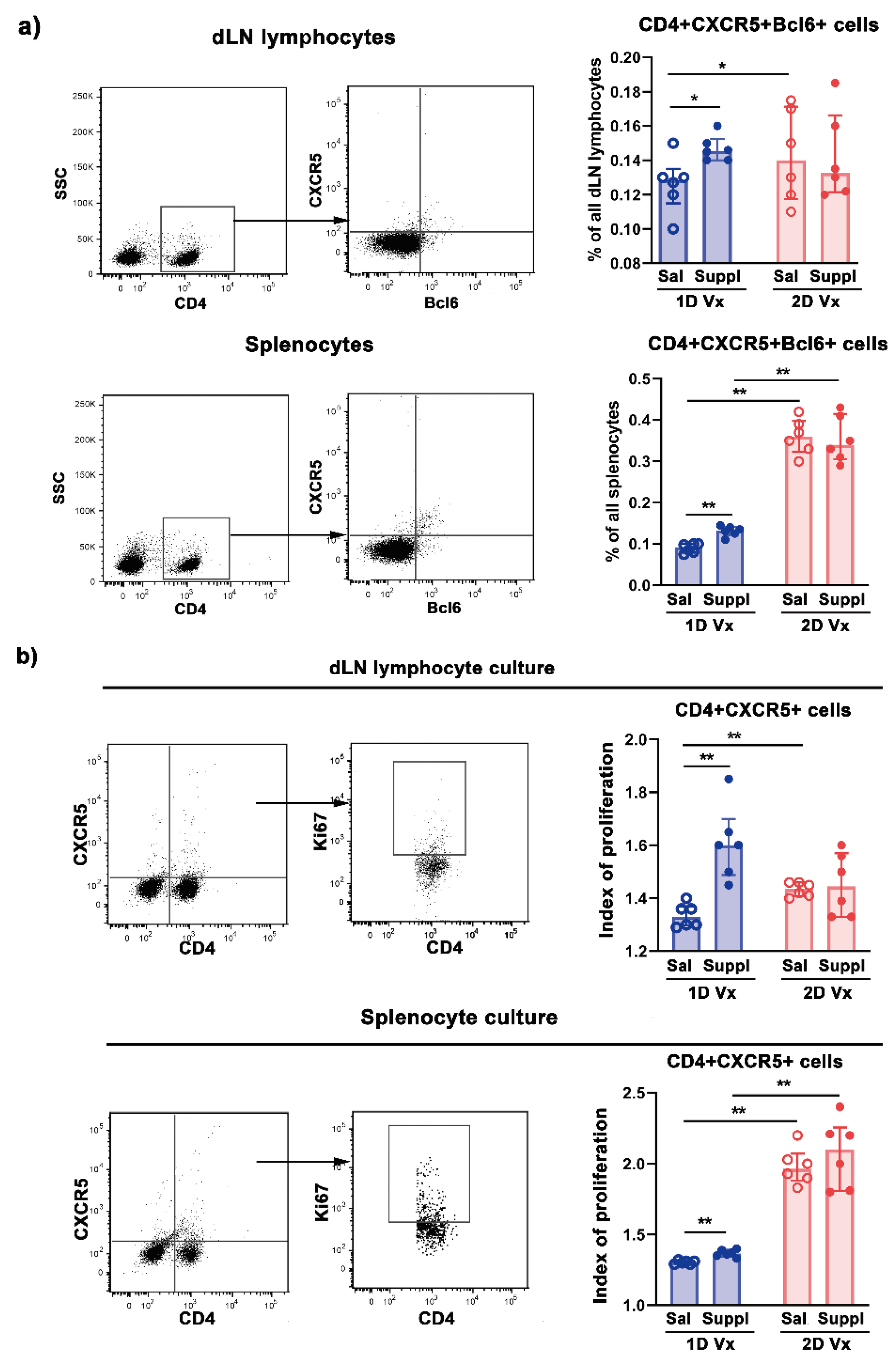
Figure 4.
Supplementation with antioxidants/immunomodulators increased the frequency of Tfh cells and affected Tfr/Tfh and Tfr/GC B cell ratios in dLNs and spleens from mice injected with one dose of QIV. a) Bar graphs indicate the frequency of CD4+CXCR5+Bcl6+FoxP3- (Tfh) cells and CD4+CXCR5+Bcl6+FoxP3+ cells (Tfr) cells among (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and treated with saline (Sal) or supplement (Suppl). The analyses were performed 14 days after the first and second QIV injection. Flow cytometry dot plots indicate FoxP3 staining (R1= FoxP3- cells; R2 = FoxP3+ cells) of CD4+CXCR5+ Bcl6+ (upper row) dLN lymphocytes and (lower row) splenocytes gated as shown in Fig.3a. b) Bar graphs indicate Tfr/Tfh cell ratio and c) Tfr/GC B cell ratio in dLN lymphocytes and splenocytes from mice injected with 1D and 2D of QIV (Vx) and administred with saline (Sal) or supplement (Suppl). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
Figure 4.
Supplementation with antioxidants/immunomodulators increased the frequency of Tfh cells and affected Tfr/Tfh and Tfr/GC B cell ratios in dLNs and spleens from mice injected with one dose of QIV. a) Bar graphs indicate the frequency of CD4+CXCR5+Bcl6+FoxP3- (Tfh) cells and CD4+CXCR5+Bcl6+FoxP3+ cells (Tfr) cells among (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and treated with saline (Sal) or supplement (Suppl). The analyses were performed 14 days after the first and second QIV injection. Flow cytometry dot plots indicate FoxP3 staining (R1= FoxP3- cells; R2 = FoxP3+ cells) of CD4+CXCR5+ Bcl6+ (upper row) dLN lymphocytes and (lower row) splenocytes gated as shown in Fig.3a. b) Bar graphs indicate Tfr/Tfh cell ratio and c) Tfr/GC B cell ratio in dLN lymphocytes and splenocytes from mice injected with 1D and 2D of QIV (Vx) and administred with saline (Sal) or supplement (Suppl). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01.
Figure 5.
The supplementation with antioxidants/immunomodulators increased the frequency of IL-21+ cells among CD4+ cells from dLNs and spleens of mice injected with one dose of QIV. a) The bar graphs display the percentage of IL-21+ cells among freshly isolated CD4+ (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) on the day14 after the first and the second dose of QIV. Flow cytometry dot plots indicate gating strategy for flow cytometry analysis of IL-21+ cells among CD4+ cells in freshly isolated (upper row) dLN lymphocytes and (lower row) splenocytes from lymphocytes gated as shown in Fig 2b. b) The bar graphs indicate the percentage of increase in the frequency of IL-21+ cells among CD4+ lymphocytes in QIV-supplemented over QIV-free (upper row) dLN lymphocyte and (lower row) splenocyte cultures from mice injected with 1D and 2D of QIV (Vx) and administered with Sal or Suppl. Flow cytometry dot plots indicate IL-21 staining of CD4+ cells from lymphocytes gated as presented in Fig 2c. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. **p<0.01.
Figure 5.
The supplementation with antioxidants/immunomodulators increased the frequency of IL-21+ cells among CD4+ cells from dLNs and spleens of mice injected with one dose of QIV. a) The bar graphs display the percentage of IL-21+ cells among freshly isolated CD4+ (upper row) draining lymph node (dLN) lymphocytes and (lower row) splenocytes from mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) on the day14 after the first and the second dose of QIV. Flow cytometry dot plots indicate gating strategy for flow cytometry analysis of IL-21+ cells among CD4+ cells in freshly isolated (upper row) dLN lymphocytes and (lower row) splenocytes from lymphocytes gated as shown in Fig 2b. b) The bar graphs indicate the percentage of increase in the frequency of IL-21+ cells among CD4+ lymphocytes in QIV-supplemented over QIV-free (upper row) dLN lymphocyte and (lower row) splenocyte cultures from mice injected with 1D and 2D of QIV (Vx) and administered with Sal or Suppl. Flow cytometry dot plots indicate IL-21 staining of CD4+ cells from lymphocytes gated as presented in Fig 2c. Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. **p<0.01.
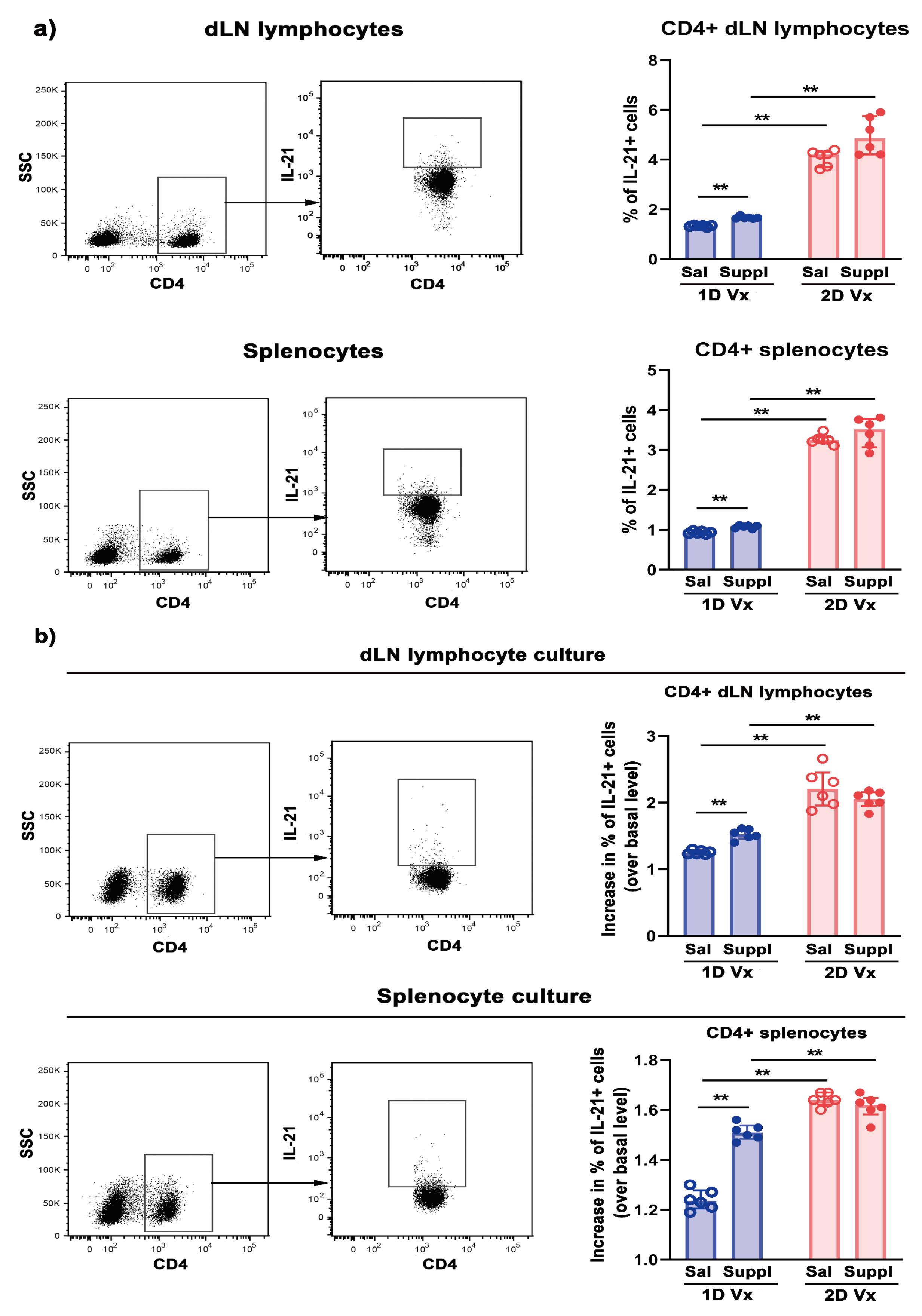
Figure 6.
Supplementation with antioxidants/immunomodulators shifted serum IgG2a/IgG1 titre ratio towards IgG2a and IFN-γ/IL-4 secretion ratio towards IFN-γ in dLN and splenocyte cultures from mice injected with two doses of QIV. a) Serum geometric mean titres of influenza virus-specific IgG1 and IgG2a, and IgG2a/IgG1 ratio in mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) determined 28 days after the first QIV dose and 14 days after the second QIV dose. Error bars indicate the 95% CI (n = 6 mice/group). Bar graphs indicate: b) concentrations of IFN-ɣ and IL-4 and c) IFN-ɣ/IL-4 ratio determined in cultures of drain lymph node (dLN) lymphocytes and splenocytes in the presence of QIV antigens (QIV antigens) and in medium alone (Medium). dLN lymphocytes and splenocytes were retrieved from mice administered with Sal or Suppl 14 days after the first (1D) and the second (2D) dose of QIV (Vx). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01, # p<0.05 vs Medium.
Figure 6.
Supplementation with antioxidants/immunomodulators shifted serum IgG2a/IgG1 titre ratio towards IgG2a and IFN-γ/IL-4 secretion ratio towards IFN-γ in dLN and splenocyte cultures from mice injected with two doses of QIV. a) Serum geometric mean titres of influenza virus-specific IgG1 and IgG2a, and IgG2a/IgG1 ratio in mice injected with one dose (1D) and two doses (2D) of QIV (Vx) and administered with saline (Sal) or supplement (Suppl) determined 28 days after the first QIV dose and 14 days after the second QIV dose. Error bars indicate the 95% CI (n = 6 mice/group). Bar graphs indicate: b) concentrations of IFN-ɣ and IL-4 and c) IFN-ɣ/IL-4 ratio determined in cultures of drain lymph node (dLN) lymphocytes and splenocytes in the presence of QIV antigens (QIV antigens) and in medium alone (Medium). dLN lymphocytes and splenocytes were retrieved from mice administered with Sal or Suppl 14 days after the first (1D) and the second (2D) dose of QIV (Vx). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. *p<0.05, **p<0.01, # p<0.05 vs Medium.
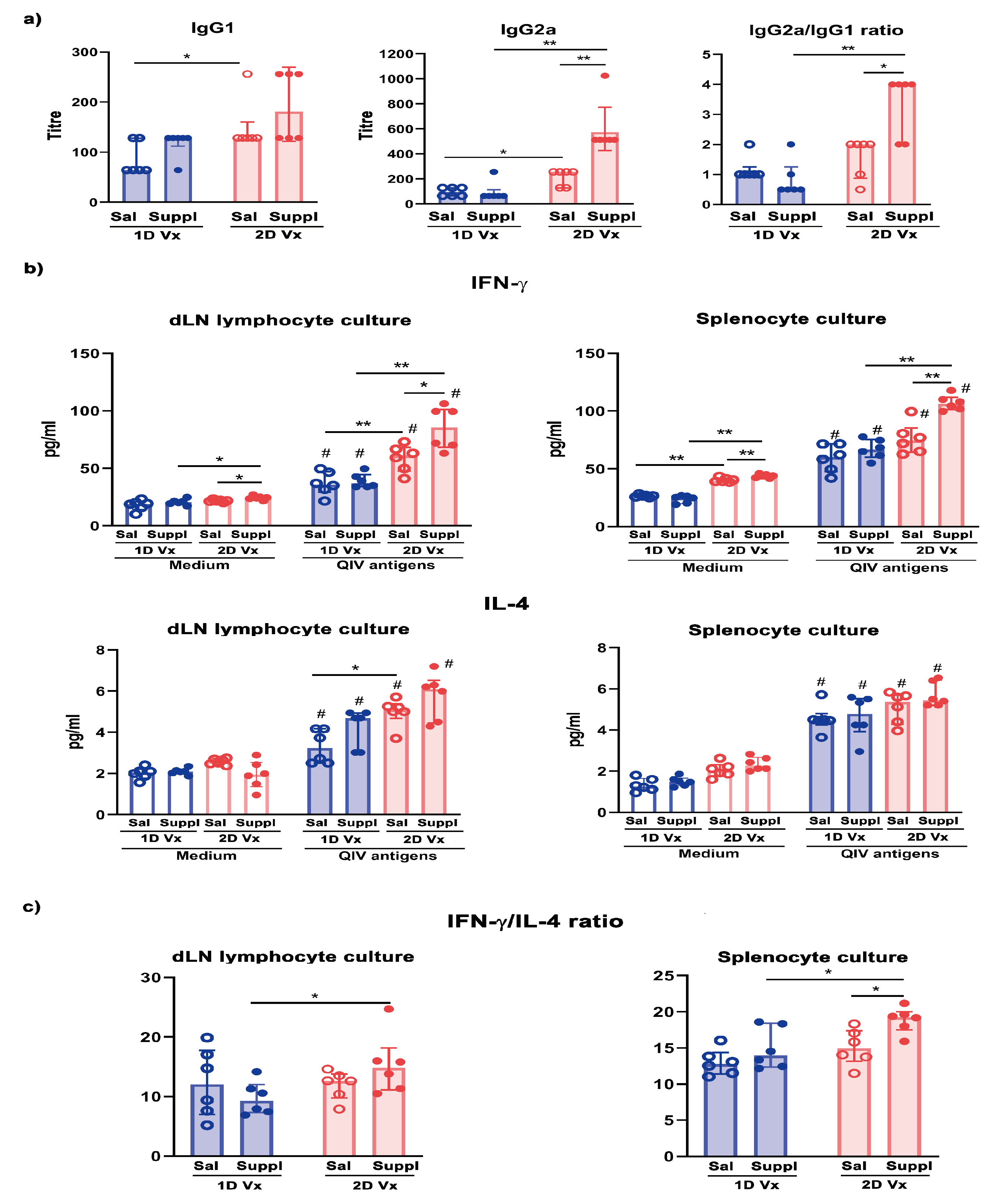
Figure 7.
Supplementation with antioxidants/immunomodulators shifted the balance between prooxidant and antioxidant parameters towards the latter parameters in spleens from mice injected with two doses of QIV. The bar graphs indicate the levels of: a) prooxidant parameter superoxide anion radical (O2• −), total oxidant capacity (TOC) and parameter of tissue damage i.e. advanced oxidation protein products (AOPP) and b) antioxidant parameters i.e. glutathione (GSH), Cu/Zn superoxide dismutase (SOD) and sulfhydryl groups (SHG) on the day 14 after the second QIV dose (2D Vx) in spleens of mice administered with saline (Sal) or supplement (Suppl). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. **p<0.01.
Figure 7.
Supplementation with antioxidants/immunomodulators shifted the balance between prooxidant and antioxidant parameters towards the latter parameters in spleens from mice injected with two doses of QIV. The bar graphs indicate the levels of: a) prooxidant parameter superoxide anion radical (O2• −), total oxidant capacity (TOC) and parameter of tissue damage i.e. advanced oxidation protein products (AOPP) and b) antioxidant parameters i.e. glutathione (GSH), Cu/Zn superoxide dismutase (SOD) and sulfhydryl groups (SHG) on the day 14 after the second QIV dose (2D Vx) in spleens of mice administered with saline (Sal) or supplement (Suppl). Data are presented as median and interquartile range (IQR) (individual data points are incorporated into bars). n=6 mice/group. **p<0.01.
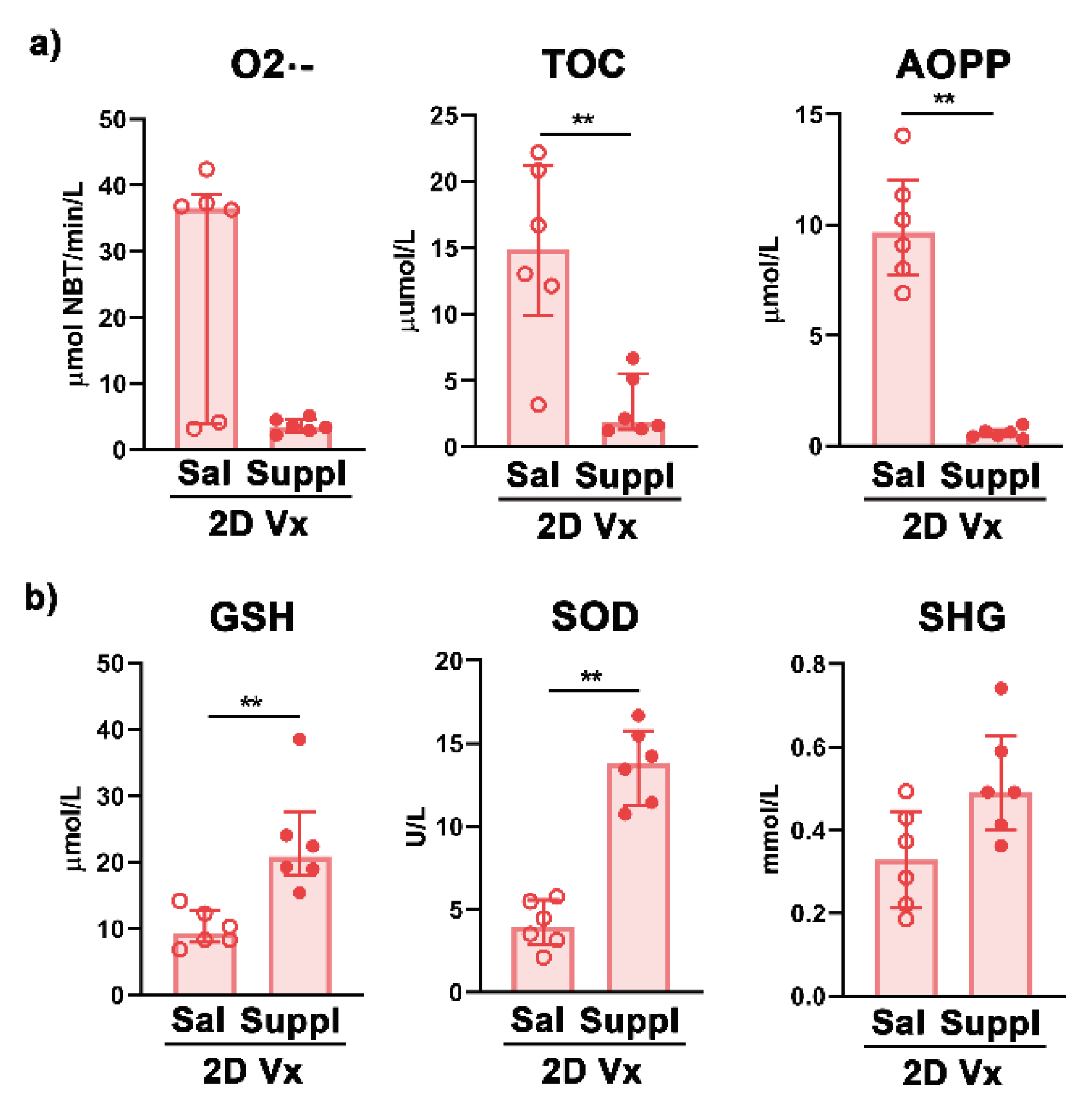
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



