
Submitted:
31 March 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
Aquaculture is an important source of food for the growing global population. To meet the increasing demand for fish, intensive aquaculture methods are commonly used. However, these methods can lead to stress and disease in fish, which can result in significant economic losses. Gamma-aminobutyric acid (GABA) is a non-protein amino acid that is found in the brain and central nervous system of animals as inhibitory neurotransmitter. It has been shown to have a variety of physiological functions, including stress reduction and immune enhancement. This study investigated the effects of dietary supplementation of GABA on growth, serum biochemistry, innate immunity and disease resistance in juvenile olive flounder (Paralichthys olivaceus) challenged with Edwardsiella tarda under high stocking density. A control diet and three experimental diets were prepared, with 150 mg/kg (GABA150), 200 mg/kg (GABA200), and 250 mg/kg (GABA250) of GABA added to each diet, respectively. The actual GABA content of the experimental diets was analyzed and found to be 63.9 mg/kg (CON), 231.3 mg/kg (GABA150), 291.6 mg/kg (GABA200), and 323.9 mg/kg (GABA250). Each experimental diet was fed to olive flounder with an initial weight of 12.75 g ± 0.3 g in 40 L tanks at two stocking densities: normal density (20 fish/tank) and high density (40 fish/tank). The experiment was conducted in triplicate for each diet and stocking density. The water temperature was maintained at 17.5 ± 1℃ and the salinity at 33 ± 1 ppt. After 8 weeks of feeding trial, growth, feed utilization, whole-body proximate composition, blood analysis, non-specific immune response, and challenge test were performed. There were no significant difference in weight gain and specific growth rate among the GABA concentrations and density × GABA concentration interaction groups. However, the low-density groups showed significantly higher values than the high-density groups (P < 0.05). There was no significant difference in feed efficiency and protein efficiency ratio among all groups. Moreover, there was no significant difference in the whole-body general composition analysis (P > 0.05). In the hematological analysis, there was no significant difference in cortisol in terms of diet and density × GABA concentration interaction, but the high-density experimental group showed a significantly higher value. Blood GABA significantly increased in proportion to the addition amount regardless of the density group (P<0.05). Superoxide dismutase activity in the non-specific immune response showed that the GABA-added experimental group had a significantly higher value than the control group, but there was no significant effect of the density densities or density × GABA concentration interaction (P<0.05). Myeloperoxidase activities in fish fed the GABA200 and GABA250 diets showed significantly higher values, and a density × GABA concentration interaction effect was observed (P<0.05). Lysozyme activity was significantly higher in the GABA150 group than in the CON, GABA200, and GABA250 groups (P < 0.05). After 15 days of challenge test by intraperitoneal injection of Edwardsiella tarda, the cumulative survival rates of the GABA150, GABA200, and GABA250 groups were significantly higher than that of the CON group (P < 0.05). The results suggested that the optimal dietary GABA level for juvenile olive flounder culture is 150 mg/kg, regardless of rearing density, to enhance growth, immune function and disease resistance.
Keywords:
γ-aminobutyric acid
; crowding effect
; growth performance
; innate immunity
; disease resistance
; olive flounder
1. Introduction
The world population is exploding, and food crisis continue to emerge. Accordingly, many scientists and researchers in all fields such as agriculture, livestock, and fisheries are working hard to solve the food crisis that will come. Among them, aquaculture is an emerging industry for food security and plays a very important role as a protein source. In fact, aquaculture production has increased by a 7 percent per year over the past 30 years (CAGR: compound annual growth rate), and in 2018, it represents 115 million tons, and this trend is expected to continue in the future. On the other hand, fishery production has stagnated at about 98 million tons from 2004 to the present [1]. The aquaculture industry is developing radically due to the standstill of fishery production and increasing demand for protein and aquatic products in modern society. The aquaculture industry has been changing from a labor-intensive primary industry to a knowledge-intensive fourth industry that utilizes Big Data, AI, and ICT [2,3]. What is important in this change is the stocking density of fish. As the aquaculture industry develops, high density culture naturally occurs, and the stress of fish increases due to high-density culture, which increases the fish's disease susceptibility and eventually leads to mass death [4]. For a successful aquaculture industry, the development of ways that can reduce the stress level of fish is essential. However, in the current aquaculture industry, diseases are suppressed by using large amounts of antibiotics, not substances that lower the stress level of fish. Due to the excessive use of these antibiotics, many pathogens have recently become resistant to antibiotics, and in particular, bacteria resistant to multiple antibiotics have emerged [5]. In addition, when antibiotics are added to feed and feeding environment, eventually aquatic products, they remain or accumulate in the water, damaging the health of end-users, humans, and polluting the aquatic environment due to antibiotics flowing into the natural environment. Therefore, research on antibiotic substitutes and alternatives that can lower the stress level is indispensable.
Gamma-Aminobutyric acid (GABA) used in this experiment is a natural substance present in bacteria, plants, and animals, and is synthesized from glutamic acid inside the synaptic presynaptic of inhibitory nerve cells [6,7,8]. Glutamic acid decarboxylase (GAD) makes GABA by removing carboxyl groups from glutamic acid. Although it is a kind of amino acid in terms of molecular structure, it is not used for protein synthesis [9,10,11]. Its chemical formula is C4H9NO2 and its mass is 103.120 g / M. The synthesized GABA is stored in synaptic vesicles, released to the synaptic cleft, and binds to the Gaba receptor (GABAA receptor & GABAB receptor) located on the posterior synaptic surface to activate the receptor [12]. GABAA receptor is a ligand-gated ion channel that passes chloride ions when GABA binds. In the developed brain, the concentration of chloride ions outside neurons is higher than inside, so when GABAA receptor opens, chloride ions enter the neurons, which induces hyperpolarization of neurons, suppressing excitability and suppressing the occurrence of action potentials [13,14]. GABAB receptor is a G protein-associated receptor that does not directly act as an ion channel, but it inhibits the excitability of nerve cells by indirectly controlling various ion channels such as potassium ion channel through G protein signaling. When there is a problem in the production of glutamic acid decarboxylase (GAD) and synthesizes GABA, the amount of GABA in the brain decreases, which causes a decrease in the activity of inhibitory synapses. Neurons that are not normally inhibited are overexcited, which in turn leads to abnormal neuronal activity [15]. In mammals, when there is a problem with GABA synthesis, it has been reported that schizophrenia and bipolar disorder occur [16,17,18,19].
GABA induces stable metabolic processes and immune and antioxidant responses by inhibiting overactive neurons in stressful situations. Because of this effect, it is a feed additive used in the poultry industry and livestock industry for various purposes such as stress reduction, immunity enhancement, antioxidant status, and promotion of feeding [20,21]. Although it is a feed additive that has not been researched largely in fish, studies on feed with GABA are also being conducted in aquaculture diets. According to previous research [22,23,24,25,26]. GABA has also been proven the positive effect in fish. In juvenile grass carp, 87.5 mg/kg of GABA in feed had a positive effect on growth and antioxidant ability. Likewise, dietary GABA at 158.7 mg/kg in Nile tilapia showed an effect on growth, feed utilization ability, and homeostasis ability. In whiteleg shrimp, 150 mg/kg of GABA showed an effect on growth and antioxidant capacity. However, there were no or scarcity of research studies under any stress conditions.
Therefore, this study was conducted to evaluate the growth performance and feed utilization ability, hematological analysis, non-specific immune response and disease resistance that occurs during normal and high-density culture by adding different concentrations of GABA in feed for olive flounder, Paralichthys olivaceus as a major domestic species.
2. Materials and Methods
2.1. Experimental Design and Diets
Anchovy fish meal, soybean meal, squid liver meal, meat and bone meal, and poultry by-products were used as the main protein sources for the experimental feed, and marine fish oil was used as a lipid source, and flour and cellulose were used for the shape of the feed, and sufficient minerals and vitamins were supplied [27]. For this, a commercial premix was added. Choline, taurine, betaine, lecithin, and monocalcium phosphate were used as other additives, and the composition of the feed source is shown in Table.1. All feed sources were put in a mixer (HYVM-1214, Hanyoung Food Machinery, Rep. Korea), mixed well, and feed was prepared using a pellet machine (SMC-12, SUN Engineering, Rep. Korea), dried at room temperature for 48 hours, and stored in a freezer at -20°C. GABA was used in powder form obtained from Mirae Resources company (Seoul, Rep. Korea) and showed a purity of 76.5% GABA as a result of analysis with high performance liquid chromatography (HPLC). In order to evenly add GABA, which is a very small amount of the experimental feed, it was mixed with cellulose for 72 hours using a ball mill machine (PL-BM5L, POONG LIM Trading., co. Rep. Korea) to make cellulose containing 40,000 mg/kg of GABA. After that, four experimental diets were made by adding 0 (CON), 150 (GABA150), 200 (GABA200) and 250 (GABA250) mg of GABA per kg of diet. The HPLC analysis showed 63.92, 231.30, 291.59 and 323.90 mg/kg of actual GABA content in CON, GABA150, GABA200 and GABA250 diets, respectively Table. 2 and Table. 3.

Table 1.
Formulation of experimental diets.
| Diets % | ||||
|---|---|---|---|---|
| Ingredients | CON | GABA150 | GABA200 | GABA250 |
| Anchovy FM1 | 50.00 | 50.00 | 50.00 | 50.00 |
| Soybean meal1 | 15.00 | 15.00 | 15.00 | 15.00 |
| Wheat flour2 | 13.70 | 13.83 | 13.70 | 13.58 |
| Squid Liver Powder1 | 4.00 | 4.00 | 4.00 | 4.00 |
| Meat & Bone meal1 | 4.00 | 4.00 | 4.00 | 4.00 |
| Poultry by product1 | 4.00 | 4.00 | 4.00 | 4.00 |
| Fish oil1 | 4.00 | 4.00 | 4.00 | 4.00 |
| Lecithin1 | 0.50 | 0.50 | 0.50 | 0.50 |
| Betaine1 | 1.00 | 1.00 | 1.00 | 1.00 |
| Taurine1 | 0.50 | 0.50 | 0.50 | 0.50 |
| Mono calcium phosphate3 | 0.50 | 0.50 | 0.50 | 0.50 |
| Mineral mix4 | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin mix5 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline3 | 0.30 | 0.30 | 0.30 | 0.30 |
| Cellulose1 | 0.50 | |||
| Cellulose (included 40,000 ppm of GABA)6 | 0.000 | 0.375 | 0.500 | 0.625 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
1 The Feed Co., Seoul, Rep Korea. 2 Samhwa Flourmils Co., Seoul, Rep Korea. 3 Sigma-Aldrich Korea Co., Seoul, Rep Korea. 4 Mineral premix (as g/kg premix): Ferrous fumarate, 12.50; Manganese sulfate, 11.25; Dried ferrous sulfate, 20.0; Dried cupric sulfate, 1.25; Cobaltous sulfate, 0.75; Zinc sulfate KVP, 13.75; Cancium iodate, 0.75; Magnesium sulfate, 80.20; Aluminum Hydroxide, 0.75. 5 Vitamin premix (as mg/kg premix): A, 1000000 IU; D, 200000 IU; E, 10000; B1, 2000; B6, 1500; B12, 10; C, 10000; Calcium pantotenic acid, 5000; Nicotinic acid 4500; B-Biotin 10; Choline chloride, 30000; Inositol, 5000. 6 Milae Bioresources Co., Seoul, Rep Korea. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Table 2.
Proximate analysis (dry matter basis) of the experimental diets1.
| Diets (%) | CON | GABA150 | GABA200 | GABA250 |
|---|---|---|---|---|
| Moisture | 7.65ns | 7.16 | 7.90 | 7.95 |
| Protein | 52.90 ns | 52.10 | 52.30 | 52.00 |
| Lipid | 10.19 ns | 10.20 | 10.13 | 10.09 |
| Ash | 12.48 ns | 12.60 | 12.42 | 12.42 |
1 Values are means from triplicate groups where the values in each row with different superscripts are significantly different. (P<0.05). ns: no significant different. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Table 3.
Actual percentage of GABA according to HPLC results for the experimental diets.
| Diet | CON | GABA150 | GABA200 | GABA250 |
|---|---|---|---|---|
| GABA (%) | 0.006392 | 0.02313 | 0.029159 | 0.03239 |
| GABA (mg/kg) | 63.92 | 231.30 | 291.59 | 323.90 |
GABA: Gamma aminobutyric acid. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
2.2. Experimental Fish and Feeding Trial
The olive flounder used in this experiment was transported from SAMWOO Aqua hatchery (Boryeong, Chungcheongnam-do, Rep. Korea) to the Laboratory of Feeds and Foods Nutrition Research Center (FFNRC) at Pukyong National University. Before starting the experiment, acclimation phase was carried out by mixing and supplying commercial feed and CON for 3 weeks in a 2000 L tank. After the acclimation phase, olive flounder with an average weight of 12.75 ± 0.5 g were reared in a 40 L square tank and 20 fish at the normal density and 40 fish at the high density. Each experimental tank was controlled so that the amount of water flow was 2.0 L/min by semi-recirculating system, and 150 % water was exchanged every day. The water temperature was set to 17.5 ± 1℃, and the salinity of the water was maintained at 33 ± 1 ppt. For sufficient oxygen supply, an air stone was installed in each tank to keep the dissolved oxygen content at 6~7 ppm, and feed was supplied by dividing 3~4% of the fish's body weight twice a day.
2.3. Sample Collection and Analysis
2.3.1. Growth Performance
After the 8 weeks culture experiment was over, the total weight of the fish in each tank was measured after fasting for 24 hours in order to investigate the growth performance and feed efficiencies. The equations for WG, FE, SGR and PER analysis are as follows.
Weight gain (WG, %) = [final weight - initial weight] × 100 / initial weight
Feed Efficiency (FE, %) = [wet weight gain / dry feed intake] × 100
Specific growth rate (SGR, % / day) = [loge final weight - loge initial weight] × 100 / days
Protein efficiency Ratio (PER) = Wet weight gain / Protein intake
2.3.2. Proximate Analysis
Proximate analysis was performed according to the standard AOAC [28] method, Moisture and ash were measured by a sample of constant weight for 24 hours in a dry oven at 105°C and burning it in a muffle furnace at 500°C for 3 hours. For the analysis of crude lipid and crude protein of feed and whole fish, the sample was frozen at -20°C for 24 hours, and then made into a powder form using a freeze-drying machine (Advantage 2.0, VirTis, New York, USA). Afterwards, crude protein was analyzed using the Auto Kjeldahl system (Buchi B-324/435/412, Switzerland; Metrohm 8-719/806, Switzerland) by Kjeldahl nitrogen determination (Nitrogen × 6.25). (Tecator AB, Sweden). In addition, the actual GABA content of the experimental feed was analyzed using HPLC by requesting the National Instrumentation Center for Environmental Management College of Agriculture and Life science at Seoul National University (Seoul 151-742, Rep. Korea).
2.3.3. Hematological Parameters
After eight weeks of feeding trial, 6 experimental fish were randomly selected for each tank, and blood was collected from the caudal vein using a non-heparinized 1mL syringe. In order to analyze the amount of GABA remaining in the blood, blood was collected 24 hours after feeding. Blood was centrifuged (5000 × g) for 10 minutes, and the serum was retaken and immediately put at -70°C. Glutamic oxaloacetic transaminase (GOT), Glutamic pyruvic transaminase (GPT), and Glucose were Fuji DRI-CHEM 3500i (Fuji Photo Film Ltd, Tokyo, Japan.) was used.
Cortisol ELISA kit (ENZO Life Sciences Inc., New York, USA; ADI-900-071) was used to analyze cortisol in blood, and carried out according to the manufacturer’s standard protocol. The kit for the quantitative measurement of cortisol uses a monoclonal antibody to cortisol to bind, in a competitive manner, cortisol in a sample or an alkaline phosphatase molecule which has cortisol covalently attached to it. After a simultaneous incubation at room temperature the excess reagents are washed away and substrate is added. After a short incubation time the enzyme reaction is stopped and read on a microplate reader at 405 nm. The measured optical density is used to calculate the concentration of cortisol.
To analyze GABA in blood, QuickDetect™ GABA ELISA Kit (Biovision Inc., San Francisco, USA; E4455-100) was used, and the analysis method of use was carried out according to the manufacturer’s standard protocol. In addition, blood was collected and analyzed exactly 24 hours after feeding to determine the amount of GABA in the blood.
2.3.4. Non-Specific Immune Response
Lysozyme activity was measured using the absorbance of the microplate technique. For this measurement, a bacterial suspension was prepared by mixing 0.2 mg of Micrococcus lysodeikticus (Sigma) for each sample in 1 mL sodium citrate buffer (0.02M, pH 5.52). Prepare serum dilution by mixing 30 μL of serum and 270 μL of sodium citrate buffer (0.02M, pH 5.52). A 180 μL of bacterial suspension was placed into each 96-well plate, and 20 μL serum dilution was subsequently added and is directly incubated thereafter at room temperature for 30 min. The decrease in absorbance was then recorded at 450 nm at 0, 30 and 60 min in a microplate reader (Sunrise TECAN, Männedorf, Switzerland). The active unit of lysozyme was defined as the amount of enzyme showing a decrease in absorbance of 0.001 per minute.
Serum isolated from fish in each experimental section. Using the SOD assay kit (Sigma-Aldrich, 191600), according to the manufacturer's instructions, the inhibition rate of the enzyme was calculated as a percentile with WST-1 (Water soluble tetrazolium dye) and xanthine oxidase. Each sample was reacted in a 37°C incubator for 20 minutes, and then absorbance was measured at a wavelength of 450 nm (absorbance of the wavelength to measure the color displayed by the reaction of WST-1 and reactive oxygen) in microplate reader (Sunrise TECAN, Männedorf, Switzerland). The inhibition rate was expressed in units of SOD activity per mg protein.
Measurement of myeloperoxidase (MPO) activity was determined broadly with 20 μL of serum was diluted with 80 μL of HBSS (Hanks Balanced Salt Solution) without Ca2+ or Mg2+ in 96-well plates and was consequently added with 35 μL of 3, 3’, 5, 5’ tetramethylbenzidine hydrochloride (TMB, 20 mM) (Sigma Aldrich) and H2O2 (5 mM). After addition of 35 μL of 4 M sulfuric acid, absorbance was measured at 450 nm wavelength after 2 min incubation in a 37°C incubator.
2.3.5. Challenge Test
In the Challenge test with Edwardsiella tardar was incubated at 27°C for 24 to 48 hours in BHI broth medium and then suspended in sterile distilled water at 1×107 CFU/mL. After injecting 0.1 mL of suspension into the peritoneal cavity of 15 fish randomly selected for each experimental feed, the survival rate of olive flounder was investigated according to elapsed time without feeding, and the experimental group and the control group were compared and analyzed.
2.4. Statistical Analysis
The arrangement of the experimental tank was completely random, and the growth and analysis results were statistically analyzed by two-way ANOVA using the IBM SPSS 26 statics program. When a significant difference was observed, a Least Significant Difference (LSD) test was used to compare means. The significance differential level was set to P<0.05.
3. Results and Discussion
3.1. Growth Performance
After 8 weeks of feeding trial, weight gain (WG, %), specific growth rate (SGR, %/day), feed efficiency (FE, %) and protein efficiency ratio (PER) are measured. As a result, there was no significant difference according to the GABA level. In addition, there was no significant interaction effect between GABA level and Density in all experimental groups. However, the normal-density group showed significantly higher values than the high-density group in WG and SGR (P<0.05). In the results of this experiment, dietary GABA did not reduce the stocking density stress, so the result of WG and SGR was significantly higher at the normal density group. Compared with previous experiment conducted on olive flounder in our laboratory, there was a significant difference in growth performance at a specific GABA level, and the same results were obtained for other fish species (Nile tilapia, grass carp and whiteleg shrimp). This showed contrary results to our experiment. [24,25,26]. For the reason, the stocking density that is too high compared to the carrying capacity that the culture system in the laboratory can handle is considered to have influenced the normal-density group as well. Therefore, if designing such an experiment in the future, another culture system should be introduced to minimize the influence between density groups. But there was no significant difference in feed utilization abilities such as FE and PER. This is because GABA is converted through GABA transaminase and glutamine synthetase into glutamate and glutamine, which affects the TCA cycle, and affects the glial cells that provide energy to neurons, thereby reducing glucose oxidation in oxidative metabolism. Because it reduces, it affects energy metabolism [29,30,31,32]. Therefore, it is considered that there was no significant difference in feed utilization. In addition, we could see the effect of GABA in tilapia, white shrimp, and grass carp with the same results. However, the exact pathways for inhibitory neurotransmitters for energy metabolism have not yet been identified [24,25,26]. The results refer to the Table 4.
3.2. Proximate Analysis
After 8 weeks of feeding trial, there was no significant difference in GABA level and Density in all experimental group. There was also no significant interaction effect of GABA level and Density (P<0.05). The results refer to the Table 5.
3.3. Hematological Parameters
Hematological analysis is an important indicator of the health and physiological condition of fish [33]. After 8 weeks of feeding trial, glutamic oxalacetic transaminase (GOT), glutamic pyruvate transaminase (GPT) and glucose (GLU) were no significant difference in GABA level and Density in all experimental group. Also, there was no significant interaction effect of GABA level and density (P>0.05). The GOT (=aspartate aminotransferase) and GPT (=alanine aminotransferase) are enzymes present in hepatocytes and are mainly released into the blood when hepatocytes are damaged. As a result of GOT and GPT analysis, there was no significant difference in all experimental groups, which is a good indicator of the safety of GABA as a feed additive [34,35]. In addition, blood cortisol level was no significant difference in the GABA level and interaction effect, however high-density group showed significantly higher value than normal-density group (P<0.05). Blood GABA content was significantly increased according to GABA intake without density and interaction effects (P<0.05). Analysis of GABA content in blood to determine the amount of GABA actually digested after ingestion showed a result that significantly increased with the amount consumed without the density group. This means dietary GABA is well absorbed in olive flounder [36]. The content of cortisol in blood is a representative stress indicator. In case of chronic stress, the interrenal cells of the head kidney are stimulated through the HPI axis to promote cortisol secretion, and metabolic reactions are promoted, thereby increasing the concentration of glucose in the blood. As a result of the analysis, the high-density group showed significantly higher cortisol value compared to the normal-density group, which is judged that dietary GABA did not show the direct effect of reducing stocking density stress. However, broiler chicken, pig and cow showed the effect of reducing temperature stress [21,37,38]. This means dietary GABA plays an important role in temperature stress rather than density stress, and it is necessary to proceed as a follow-up study. In addition, glucose was no significant difference in all experimental group, which is believed to be maintained at a similar level of blood glucose by insulin, thyroid hormone and other hormones, which control blood glucose level in the body [39,40]. The results refer to the Table 6.
3.4. Non-Specific Immune Response
A non-specific immune response is the innate immune system that reacts primarily in the body when exposed to dangerous substances. The reaction to external substances or abnormal substances (infectious substances, chemical / physical damage) occurs immediately and non-selectively, thus acting as an important indicator of the immune response [41,42] After 8 weeks of feeding trial, there were no significant differences according to density in lysozyme activity, myeloperoxidase activity (MPO, absorbance) and superoxide dismutase (SOD, % inhibition) (P>0.05). However, there was a significant difference in the GABA level. Lysozyme activity of fish fed GABA150 was significantly higher values than those of fish fed CON, GABA200 and GABA250, and there was no interaction effect (P>0.05). MPO of fish fed GABA200 and GABA250 were significantly higher values than those of fish fed CON and GABA150, and there were also interactions effect (P<0.05). SOD of fish fed GABA250 and GABA150 were significantly higher values than those of fish fed CON and there was no significant difference compare to fish fed GABA200, and there was no interaction effect (P>0.05). SOD is an enzyme that acts as an antioxidant defense mechanism in almost all of cell exposed to oxygen. As a result of the analysis, significantly higher values were obtained in the experimental group to which GABA was added. It was found that GABA decrease antioxidant stress and increases the activity of antioxidant enzymes in fish as found in humans, rats, and broilers, similar to those found in whiteleg shrimp and Nile tilapia. MPO is an enzyme mainly contained in neutrophils. It catalyzes the reaction to produce hypochlorous acid (HOCl) from hydrogen peroxide (H2O2) and chloride ion (Cl-), and has antibacterial and antiviral effects. As a result of the analysis, significantly higher values were found in fish fed GABA200 and GABA250. Lysozyme activity is an enzyme that destroys the cell wall of bacteria and plays an important part in the innate immune system. As a result of the analysis, fish fed GABA150 has a significantly higher value. This confirmed that the treatment with GABA or GABAergic drugs is related to the results of recent studies that improve macrophage maturation, autophagy activation, antibacterial response to bacterial infection, and enhance the host's innate immune response (Keshav, S., et al., 1991). In summary, it could be seen that the innate immune response was enhanced in the experimental group to which GABA was added without the rearing density. However, different trends were shown depending on the amount of GABA added to the feed, so the amount of GABA added according to the fish species and size should be identified. The results refer to the Table 7.
3.5. Challenge Test
After 8 weeks of feeding trial, challenge test with pathogenic bacteria (Edwardsiella tarda 1×107 CFU/mL) for 15 days showed significantly higher cumulative survival rate for those of fish fed GABA150, GABA200, and GABA250 diets compared to these of fish fed CON diet (P<0.05). But there is no fully established mechanism for the interaction of the nervous and immune systems. In particular, it has not been studied mainly in fish. However, according to a recently published paper, the effect of enhancing immunity through macrophages and autophagy through in vivo GABA and GABAergic is gradually revealed. In the end, it is judged that it showed a high cumulative survival rate [44,45,46]. The results refer to the Figure 1.
4. Conclusions
Rearing density has always been an important topic in the aquaculture industry. Efforts to create high production per area are ongoing and various aquaculture technologies are developing. However, the high stocking density increases the risk of mass mortality as the stress on fish increases, low growth performance, and disease susceptibility increases. In order to prevent this, it is necessary to develop feed additives to reduce stress, promote immune response and disease resistance. The gamma aminobutyric acid used in this experiment is an inhibitory neurotransmitter and is a feed additive already used in livestock industries such as poultry, pig and cow farming. Therefore, this study was conducted to see the effect of enhancing fish growth and immunity by adding dietary GABA under density stress. These results indicated that dietary gamma aminobutyric acid (GABA) did not affect olive flounder growth performance but could have beneficial effect on immune responses and disease resistance. However, in the non-specific immune response, it showed a positive effect according to the different GABA level. It is necessary to experiment by dividing the GABA level more accurately in future experiments. Lastly, as mentioned in the body of this paper, the Gut-brain axis and microbiota is a model that helps to establish explanations for the beneficial effects found not only in GABA, but other microbial metabolites in the gut of fish. Though this system provides an excellent basis upon which to assert a mechanism for GABA’s systemic effects, the scope of the current trial focuses primarily on GABA’s general effects on growth and immunity that can be assessed by using tests common to short term feeding trials with juvenile fish. Therefore, in future studies, it is necessary to observe the exact activity of neurons or immune cells, and also to study other environmental stresses such as temperature stress.
Author Contributions
J.B.: planned the experiment; formulated the feed; determined the growth, somatic indices, non-specific immune enzyme activities, and gene expressions; and drafted the manuscript. M.M.: performed the statistical analyses, drafted and finalized the manuscript. H.J., W.C. and S.L.: performed the investigation and statistical analyses and drafted the manuscript. T.M. and K.K.: reviewed the manuscript. S.C.B.: planned the experiment, supervised the project and gave final approval of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the National Institute of Fisheries Science (NIFS), Republic of Korea (R2024038) and National Research Foundation (NRF) of Korea (2018K1A3A1A39088080).
Institutional Review Board Statement
This experiment was conducted under the guidelines of Institutional Animal Care and Use Committee Regulations, No. 554, issued by Pukyong National University, Busan, Republic of Korea. Every effort was taken to minimize the number of fish sacrificed.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO (2020) FISHSTAT Plus, universal software for fishery statistical time series. Food and Agriculture Organization, United Nations, Rome. Electronic webpage.
- Gorbunova, A. V. , Kostin, V. E., Pashkevich, I. L., Rybanov, A. A., Savchits, A. V., Silaev, A. A.,... & Judaev, Y. V. (2020, January). Prospects and opportunities for the introduction of digital technologies into aquaculture governance system. In IOP Conference Series: Earth and Environmental Science (Vol. 422, No. 1, p. 012125). IOP Publishing. [CrossRef]
- Li, P. , Mai, K., Trushenski, J., & Wu, G. (2009). New developments in fish amino acid nutrition: towards functional and environmentally oriented aquafeeds. Amino acids, 37(1), 43-53. [CrossRef]
- Noble, A. C. , & Summerfelt, S. T. Diseases encountered in rainbow trout cultured in recirculating systems. Annual Review of Fish Diseases 1996, 6, 65–92. [Google Scholar]
- Serrano, P. H. (2005). Responsible Use of Antibiotics in Aquaculture. FAO Fisheries Technical Paper No. 469.
- Abe, T. , & Takeya, K. (2005). Difference in gamma-aminobutyric acid content in vegetable soybean seeds. Journal of the Japanese Society for Food Science and Technology (Japan). [CrossRef]
- Arumugam, K. , Tung, P., Chinnappa, C. C., & Reid, D. M. γ-Aminobutyric acid stimulates ethylene biosynthesis in sunflower. Plant Physiology 1997, 115, 129–135. [Google Scholar]
- Bown, A. W. , & Shelp, B. J. (). The metabolism and functions of [gamma]-aminobutyric acid. PlantPhysiology 1997, 115, 1. [Google Scholar] [CrossRef]
- Dhakal, R. , Bajpai, V. K., & Baek, K. H. Production of GABA (gamma - Aminobutyric acid) by microorganisms: A review. Brazilian Journal of Microbiology 2012, 43, 1230–1241. [Google Scholar] [PubMed]
- Fenalti, G. , Law, R. H., Buckle, A. M., Langendorf, C., Tuck, K., Rosado, C. J.,... & Wilce, M. (2007). GABA production by glutamic acid decarboxylase is regulated by a dynamic catalytic loop. Nature structural & molecular biology, 14(4), 280. [CrossRef]
- Walls, A. B. , Waagepetersen, H. S., Bak, L. K., Schousboe, A., & Sonnewald, U. (2015). The glutamine–glutamate/GABA cycle: Function, regional differences in glutamate and GABA production and effects of interference with GABA metabolism. Neurochemical research, 40 (2), 402-409. [CrossRef]
- Watanabe, M. , Maemura, K., Kanbara, K., Tamayama, T., & Hayasaki, H. (2002). GABA and GABA receptors in the central nervous system and other organs. In International review of cytology (Vol. 213, pp. 1–47). Academic Press. [CrossRef]
- Ong, J. , & Kerr, D. I. (1990). GABA-receptors in peripheral tissues. Life sciences, 46(21), 1489-1501. [CrossRef]
- McCormick, D. A. (1989). GABA as an inhibitory neurotransmitter in human cerebral cortex. Journal of neurophysiology, 62(5), 1018-1027. [CrossRef]
- Abdou, A. M. , Higashiguchi, S., Horie, K., Kim, M., Hatta, H., & Yokogoshi, H. (2006). Relaxation and immunity enhancement effects of γ Aminobutyric acid (GABA) administration - in humans. Biofactors, 26(3), 201-208. [CrossRef]
- Bach, B. , Meudec, E., Lepoutre, J. P., Rossignol, T., Blondin,B., Dequin, S., & Camarasa, C. New insights into gamma-aminobutyric acid catabolism: Evidence for gamma-hydroxybutyric acid and polyhydroxybutyrate synthesis in Saccharomyces cerevisiae. Applied and Environmental Microbiology 2009, 75, 4231–4239. [Google Scholar] [PubMed]
- Bongianni, F. , Mutolo, D, Nardone, F., & Pantaleo, T. GABAergic and glycinergic inhibitory mechanisms in the lamprey respiratory control. Brain research. 2006, 1090, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, E. , de Kleijn, R., Colzato, L. S., Alkemade, A., Forstmann, B. U., & Nieuwenhuis, S. (2015). Neurotransmitters as food supplements: the effects of GABA on brain and behavior. Frontiers in psychology, 6, 1520. [CrossRef]
- Hsu, Y. T. , Chang, Y. G., & Chern, Y. Insights into GABAergic system alteration in Huntington's disease. Royal Society Open Biology 2018, 8, 180165. [Google Scholar]
- Al Wakeel, R. A. , Shukry, M., Abdel Azeez, A., Mahmoud, S., & Saad, M. F. (2017). Alleviation by gamma amino butyric acid supplementation of chronic heat stress-induced degenerative changes in jejunum in commercial broiler chickens. Stress, 20(6), 562-572. [CrossRef]
- Li, Y. H. , Li, F., Liu, M., Yin, J. J., Cheng, B. J., Shi, B. M., & Shan, A. S. Effect of γ-aminobutyric acid on growth performance, behavior and plasma hormones in weaned pigs. Canadian Journal of Animal Science 2015, 95, 165–171. [Google Scholar]
- Chand, N. , Muhammad, S., Khan, R. U., Alhidary, I. A., & ur Rehman, Z. Ameliorative effect of synthetic γ- aminobutyric acid (GABA) on performance traits, antioxidant status and immune response in broiler exposed to cyclic heat stress. Environmental Science and Pollution Research 2016, 23, 23930–23935. [Google Scholar] [PubMed]
- Kim, S. K. , Takeuchi, T., Yokoyama, M., & Murata, Y. (). Effect of dietary supplementation with taurine, β-alanine and GABA on the growth of juvenile and fingerling Japanese flounder Paralichthys olivaceus. Fisheries science 2003, 69, 242–248. [Google Scholar]
- Wu, F. , Liu, M., Chen, C., Chen, J., & Tan, Q. Effects of Dietary Gamma Aminobutyric Acid on Growth Performance, Antioxidant Status, and Feeding related Gene Expression of Juvenile - Grass Carp, Ctenopharyngodon idellus. Journal of the World Aquaculture Society 2016, 47, 820–829. [Google Scholar] [CrossRef]
- Xie, S. W. , Li, Y. T., Zhou, W. W., Tian, L. X., Li, Y. M., Zeng, S. L., & Liu, Y. J. Effect of γ-aminobutyric acid supplementation on growth performance, endocrine hormone and stress tolerance of juvenile Pacific white shrimp, Litopenaeus vannamei, fed low fishmeal diet. Aquaculture Nutrition 2017, 23, 54–62. [Google Scholar] [CrossRef]
- Temu, V. , Kim, H., Hamidoghli, A., Park, M., Won, S., Oh, M.,... & Bai, S. C. Effects of dietary gamma aminobutyric acid in juvenile Nile tilapia, Orechromis niloticus. Aquaculture 2019, 507, 475–480. [Google Scholar]
- NRC. 2011. Nutrient requirements of fish and shrimp. National Academy Press, Washington, DC, USA.
- AOAC. 2005. Official methods of analysis of the association of official analytical chemists, 12th edn. Association of Official Analytical Chemists, Washington, DC, USA.
- Patel, A. B. , de Graaf, R. A., Mason, G. F., Rothman, D. L., Shulman, R. G., & Behar, K. L. The contribution of GABA to glutamate/glutamine cycling and energy metabolism in the rat cortex in vivo. Proceedings of the National Academy of Sciences 2005, 102, 5588–5593. [Google Scholar]
- Carabotti, M. , Scirocco, A., Maselli, M. A., & Severi, C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Annals of gastroenterology: quarterly publication of the Hellenic Society of Gastroenterology 2015, 28, 203. [Google Scholar]
- Strandwitz, P. (2018). Neurotransmitter modulation by the gut microbiota. Brain research, 1693, 128-133. [CrossRef]
- Strandwitz, P., Kim, K. H., Terekhova, D., Liu, J. K., Sharma, A., Levering, J., ... & Mroue, N. (2019). GABA modulating bacteria of the human gut microbiota. Nature microbiology, 4(3), 396. [CrossRef]
- Magnadóttir, B. (2006). Innate immunity of fish (overview). Fish & shellfish immunology, 20(2), 137-151. [CrossRef]
- Bucher, F., & Hofer, R. (1990). Effects of domestic wastewater on serum enzyme activities of brown trout (Salmo trutta). Comparative Biochemistry and physiology. C, Comparative Pharmacology and Toxicology, 97(2), 381-385. [CrossRef]
- Vaglio, A., & Landriscina, C. (1999). Changes in liver enzyme activity in the Teleost Sparus auratain response to cadmium intoxication. Ecotoxicology and Environmental Safety, 43(1), 111-116. [CrossRef]
- Fugelli, K., Storm-Mathisen, J., & Fonnum, F. Synthesis of γ-aminobutyric acid in fish erythrocytes. Nature 1970, 228, 1001–1001. [CrossRef]
- Tang, J. , & Chen, Z. The protective effect of γ-aminobutyric acid on the development of immune function in chickens under heat stress. Journal of animal physiology and animal nutrition 2016, 100, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. M. , Wang, C., Liu, H. Y., Liu, J. X., & Ferguson, J. D. Effects of rumen-protected γ-aminobutyric acid on feed intake, lactation performance, and antioxidative status in early lactating dairy cows. Journal of Dairy Science 2013, 96, 3222–3227. [Google Scholar] [PubMed]
- Peter, M. S. The role of thyroid hormones in stress response of fish. General and Comparative Endocrinology 2011, 172, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Tort, L. Stress and immune modulation in fish. Developmental & Comparative Immunology 2011, 35, 1366–1375. [Google Scholar]
- Abbas, A. K. , Lichtman, A. H., & Pillai, S. (2014). Cellular and molecular immunology E-book. Elsevier Health Sciences.
- Delves, P. J. , Martin, S. J., Burton, D. R., & Roitt, I. M. (2017). Roitt's essential immunology. John Wiley & Sons.
- Keshav, S., Chung, P., Milon, G., & Gordon, S. (1991). Lysozyme is an inducible marker of macrophage activation in murine tissues as demonstrated by in situ hybridization. Journal of Experimental Medicine, 174(5), 1049-1058. [CrossRef]
- Kim, J. K., Kim, Y. S., Lee, H. M., Jin, H. S., Neupane, C., Kim, S., & Choe, S. K. (2018). GABAergic signaling linked to autophagy enhances host protection against intracellular bacterial infections. Nature communications, 9(1), 4184. [CrossRef]
- Sasai, M. , Sakaguchi, N., Ma, J. S., Nakamura, S., Kawabata, T., Bando, H., & Standley, D. M. (2017). Essential role for GABARAP autophagy proteins in interferon-inducible GTPase-mediated host defense. Nature immunology, 18(8), 899. [CrossRef]
- Auteri, M. , Zizzo, M. G., & Serio, R. GABA and GABAreceptors in the gastrointestinal tract: From motility to inflammation. Pharmacological Research 2015, 93, 11–21. [Google Scholar] [PubMed]
Figure 1.
Challenge Test with Edwardsiella tarda (1×107 CFU/mL)1. 1 Cumulative survival percentage of olive flounder fed four different experimental diet and two different stocking density for 8 weeks and challenged with Edwardsiella tarda (1×107 CFU/mL) for 15 days. NCON: Normal Density + CON diet. HCON: High Density + CON diet. NGABA150: Normal Density + GABA150. HGABA150: High Density + GABA150. NGABA200: Normal Density + GABA200. HGABA200: High Density + GABA200. NGABA250: Normal Density + GABA250. HGABA250: High Density + GABA250.
Figure 1.
Challenge Test with Edwardsiella tarda (1×107 CFU/mL)1. 1 Cumulative survival percentage of olive flounder fed four different experimental diet and two different stocking density for 8 weeks and challenged with Edwardsiella tarda (1×107 CFU/mL) for 15 days. NCON: Normal Density + CON diet. HCON: High Density + CON diet. NGABA150: Normal Density + GABA150. HGABA150: High Density + GABA150. NGABA200: Normal Density + GABA200. HGABA200: High Density + GABA200. NGABA250: Normal Density + GABA250. HGABA250: High Density + GABA250.
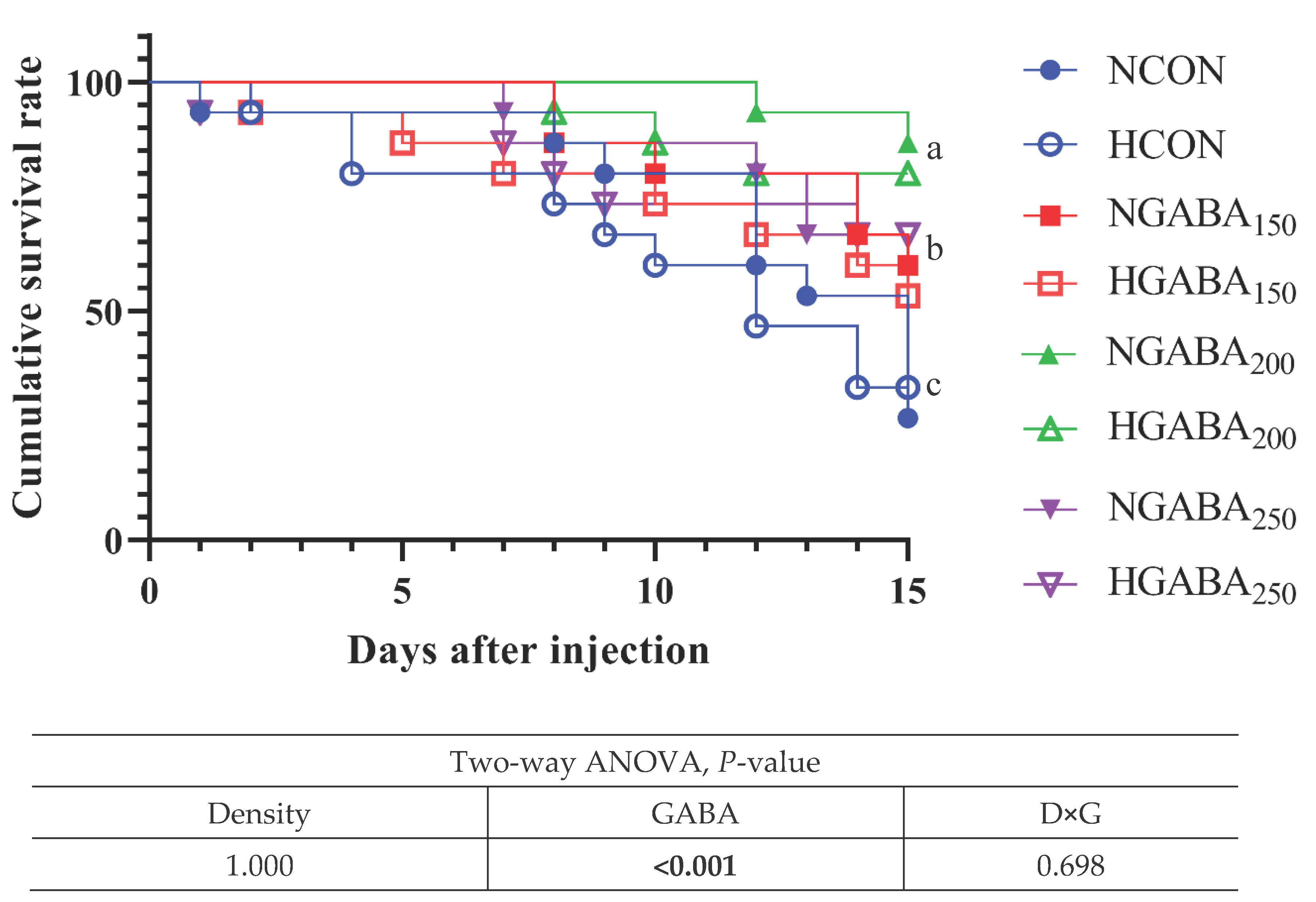
Table 4.
Growth performance and feed utilization1.
| Density | ND | HD | Two-way ANOVA, P-value |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON | GABA150 | GABA200 | GABA250 | CON | GABA150 | GABA200 | GABA250 | Density | GABA | D×G |
| WG2 | 145.21aa | 149.03a | 135.66a | 139.48a | 129.07b | 120.93b | 122.97b | 123.26b | <0.001 | 0.292 | 0.325 |
| SGR3 | 1.87aa | 1.90a | 1.78a | 1.82a | 1.73b | 1.65b | 1.67b | 1.67b | <0.001 | 0.302 | 0.348 |
| FE4 | 84.10ns | 82.27a | 73.40a | 81.80a | 77.40a | 73.30a | 70.70a | 77.13a | 0.070 | 0.213 | 0.891 |
| PER5 | 1.59ns | 1.58a | 1.42a | 1.57a | 1.46a | 1.41a | 1.37a | 1.48a | 0.067 | 0.324 | 0.896 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different. (P<0.05). 2 Weight gain (WG, %) = [final weight - initial weight] × 100 / initial weight. 3 Specific growth rate (SGR, % / day) = [loge final weight - loge initial weight] × 100 / days. 4 Feed Efficiency (FE, %) = [wet weight gain / dry feed intake] × 100. 5 Protein efficiency Ratio (PER) = Wet weight gain / Protein intake. ns: no significant different. ND: Normal density; HD: High density. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Table 5.
Whole-body proximate composition (as is %) of olive flounder1.
| Density | ND | HD | Two-way ANOVA, P-value |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON | GABA150 | GABA200 | GABA250 | CON | GABA150 | GABA200 | GABA250 | Density | GABA | D×G |
| Moisture | 79.6ns | 80.3 | 79.7 | 79.0 | 79.3 | 79.1 | 79.4 | 79.7 | 0.154 | 0.623 | 0.620 |
| Protein | 15.2ns | 14.6 | 15.2 | 15.4 | 15.2 | 15.6 | 15.3 | 14.9 | 0.064 | 0.406 | 0.898 |
| Lipid | 1.17ns | 1.18 | 1.00 | 1.06 | 1.16 | 1.16 | 1.19 | 1.13 | 0.370 | 0.729 | 0.177 |
| Ash | 3.54ns | 3.84 | 3.87 | 3.81 | 4.04 | 3.71 | 3.89 | 3.89 | 0.109 | 0.643 | 0.623 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different. (P<0.05). ND: Normal density; HD: High density. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Table 6.
Blood Chemistry of olive flounder (biochemical characteristics)1.
| Density | ND | HD | Two-way ANOVA, P-value |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON | GABA150 | GABA200 | GABA250 | CON | GABA150 | GABA200 | GABA250 | Density | GABA | D×G |
| GOT2 | 17.7 ns | 18.3 | 19.0 | 18.7 | 16.7a | 16.0a | 16.7a | 16.0 | 0.053 | 0.959 | 0.936 |
| GPT3 | 16.7ns | 17.0 | 16.0 | 16.3 | 16.7 | 15.0a | 16.3a | 16.0 | 0.188 | 0.605 | 0.149 |
| GLU4 | 12.7ns | 13.0 | 11.7 | 13.3 | 12.0 | 14.3 | 16.0a | 11.7 | 0.379 | 0.560 | 0.145 |
| Cortisol5 | 12.6bb | 51.7b | 51.8b | 51.7b | 54.2a | 55.9a | 55.8a | 55.0a | 0.002 | 0.980 | 0.739 |
| GABA6 | 166a | 220b | 239c | 302d | 175a | 214b | 247c | 293d | 0.790 | <0.001 | 0.059 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different. (P<0.05). 2 GOT: Glutamic oxaloacetic transaminase (U L-1). 3 GPT: Glutamic pyruvic transaminase (U L-1). 4 GLU: Glucose (mg dL-1). 5 Cortisol: Cortisol in blood (ng mL-1). 6 GABA: gamma aminobutyric acid in blood (pg mL-1). ns: no significant different. ND: Normal density; HD: High density. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Table 7.
Non-specific immune responses of olive flounder1.
| Density | ND | HD | Two-way ANOVA, P-value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON | GABA150 | GABA200 | GABA250 | CON | GABA150 | GABA200 | GABA250 | Density | GABA | D×G |
| SOD2 | 16.24b | 23.15a | 18.78ab | 21.79a | 12.66b | 21.09a | 18.22ab | 22.05a | 0.462 | 0.046 | 0.906 |
| MPO3 | 1.45b | 1.20b | 1.87a | 1.44a | 1.23b | 1.44b | 1.40ab | 1.81a | 0.818 | 0.015 | 0.006 |
| Lysozyme4 | 0.15b | 0.39a | 0.10b | 0.25b | 0.17b | 0.39a | 0.26bb | 0.14b | 0.542 | <0.001 | 0.520 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different. (P<0.05). 2 SOD: Superoxide dismutase activity (% inhibition). 3 MPO: Myeloperoxidase (OD at 450nm). 4 Lysozyme: Lysozyme activity (U mL-1). ND: Normal density; HD: High density. CON: Control; GABA150: GABA 150 mg/kg added; GABA200: GABA 200 mg/kg added; GABA250: GABA 250 mg/kg added.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



