Submitted:
01 April 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
Sea cucumber peptides are small molecular peptides with biological function obtained from fresh sea cucumber as feedstock, which were hydrolyzed by protease, separated ,and purified. As the main product of sea cucumber refinement, it more accurately expresses the nutritional effect of sea cucumber and has a wide range of potential applications. It has been demonstrated that sea cucumber has a certain value-added effect on bone mineral density regulation, but the differences in bone mineral density regulation of extracts from different parts of sea cucumber and the techniques contributing to the efficient isolation of bone sea cucumber peptides have not been reported. Firstly, the aim of this study was to reveal the differences in osteogenic activities of enzymatic peptides from three major parts of sea cucumber: the body wall, the egg protein, and the intestinal protein. Then, the bone-enhancing sea cucumber peptides were isolated based on the characteristics of the dif-ferences in molecular weight distribution, hydrophilicity, and ionisation degree of sea cucumber, and to explore the possible mechanisms of their action. The findings indicated that sea cucumber peptides extracted from three distinct sites exhibited a dose-dependent enhancement of osteoblast proliferation and differentiation. Upon evaluating the three methods of sea cucumber peptide separation based on molecular weight distribution, hydrophilicity, and ionization, it was observed that the fraction isolated by hydrophobicity and
Keywords:
protein
; osteoporosis
; MC3T3-E1 cells
; peptide
1. Introduction
Osteoporosis is a skeletal disease that results from reduced bone density and microstructural changes in bone tissue [1]. It primarily affects the spine, pelvis, shoulder joints, and other parts of the body, leading to excessive bone decomposition and hindering necessary bone metabolism processes, such as bone synthesis and remodeling. Individuals aged 50 and above, particularly middle-aged and elderly groups, are the main incidence groups of osteoporosis. Approximately one-fifth of these individuals are men, while one-third are women [2]. Osteoporosis is classified into primary and secondary types. Primary osteoporosis can occur in all age groups and is caused by the lack of substances related to osteogenesis, which are necessary for the process of bone synthesis and metabolism. According to Zhang’s report, secondary osteoporosis is primarily caused by acquired habits, physiological stages, and diseases such as pregnancy, menopause, liver disease, protein deficiency, consumption of corticosteroid drugs, and bone-related tumors[3]. Osteoblasts and osteoclasts maintain the bone remodeling process in the body. Osteoblasts are responsible for synthesising, secreting, and mineralising the bone mechanism. Their osteogenic differentiation and extracellular matrix mineralisation are crucial processes for maintaining bone mass stability. These processes are influenced and regulated by multiple signalling pathways. Currently, the primary signalling pathways associated with bone metabolism are the Wnt/β-catenin, BMP/Smads, RANK/RANKL/OPG, and Notch pathways. These four osteogenic pathways regulate the signalling connection between osteoblasts and osteoclasts, and also directly impact the value-added differentiation of osteoblasts associated with the organism [4]. It is important to note that this information is objective and based on scientific evidence. To alleviate osteoporosis, there are mainly Chinese and Western medical treatments, exercise, and dietary supplements. The concept of dietary supplementation for bone health has existed since ancient times, referred to as 'medicine in food' or 'medicine and food in the same source'. As the name suggests, natural food and medicine share a common and compatible aspect. If combined correctly in one's daily diet, they can silently improve the consumer's physical well-being. Regarding the promotion of bone health and the alleviation of osteoporosis, it has been suggested in folklore since ancient times to consume protein. Sea cucumber, which is produced in coastal cities, is a traditional tonic in China with various nutritional benefits. The main way to consume it is by steaming fresh sea cucumber, which is ready to eat. With the development of the food industry, other forms of sea cucumber consumption have emerged, such as freeze-dried sea cucumber, instant sea cucumber, and sea cucumber extracts. These forms have deepened the study of the effectiveness of sea cucumber and its mechanisms. It is important to note that sea cucumber should not be considered a cure for any specific ailment or disease. Sea cucumber collagen peptide is a degradation product of collagen. It has small molecular weight, strong water solubility, and is easily digested and absorbed by the human body. Supplementation of bioactive peptides can enhance immunity, as oligopeptides can reduce competition for free amino acid transportation and improve absorption [5]. Sea cucumber collagen peptides have lower viscosity, good solubility, and stability, making them suitable for commercial production and consumption [6]. They are mainly extracted from the sea cucumber body wall, egg white, and intestine. At the present stage, the main research directions of proteolytic peptide bone-promoting activity at home and abroad are divided into three aspects: verification of osteoblasts on bone-promoting activity, in vivo characterization of bone-promoting activity in mice, and investigation of the mechanism of bone-promoting activity [7]. The commercial production of sea cucumber still relies on relatively traditional separation technology, lacking efficient and directional purification techniques. This results in unsatisfactory purity of the separation effect and a wasteful production process. Research has shown that the protein peptide extracted from whole sea cucumber has a certain promoting effect on mouse embryonic osteoblasts and alkaline phosphatase activity [8]. Sea cucumber ovalbumin peptide has been shown to enhance the bone density of mice in vivo. The bone-promoting activity was characterized through cycle distribution, cell migration, immunoblotting analysis, and immunofluorescence analysis. However, there are still research gaps that need to be addressed. There are currently few bone-promoting functional foods on the market that are based on sea cucumber as a raw material, despite research and development in this area. Additionally, there is a lack of sufficient scientific basis and technical support for these products. This highlights the need for further investigation and development in this field. The published articles on sea cucumber peptides with bone-promoting activity mainly use enzyme-digested powder from a single part or a mixture of multiple parts of the whole sea cucumber as experimental raw materials. Subjective evaluations have been excluded, and technical term abbreviations have been explained when first used. The language is clear, objective, and value-neutral, with a formal register and precise word choice. The text adheres to style guides, uses consistent citation, and follows a consistent footnote style and formatting features. The structure is clear with logical progression and causal connections between statements. The text is free from grammatical errors, spelling mistakes, and punctuation errors. No new content has been added. While it has been verified that sea cucumber has osteogenic activity, it is unclear whether there are specific differences in the enzymatic peptides of various parts of the sea cucumber that contribute to this activity, or if there are differences in the pathways of osteogenic activity. In commercial production, the processor has a low utilization rate of recovered sea cucumber intestinal eggs, with the main part of the sea cucumber body wall being consumed by consumers. There is a dearth of research on efficient purification techniques for protein peptide fractions at the industrial level[9]. In summary, there is a need to optimise bone-promoting functional products made from sea cucumber peptides. This includes comparing the bone-promoting efficacy of sea cucumber peptides extracted from different parts of the sea cucumber and researching and developing highly efficient isolation methods for bone-promoting active peptides.
2. Results
2.1. Compositional Analysis of Protein Peptides Extracted from Three Sites
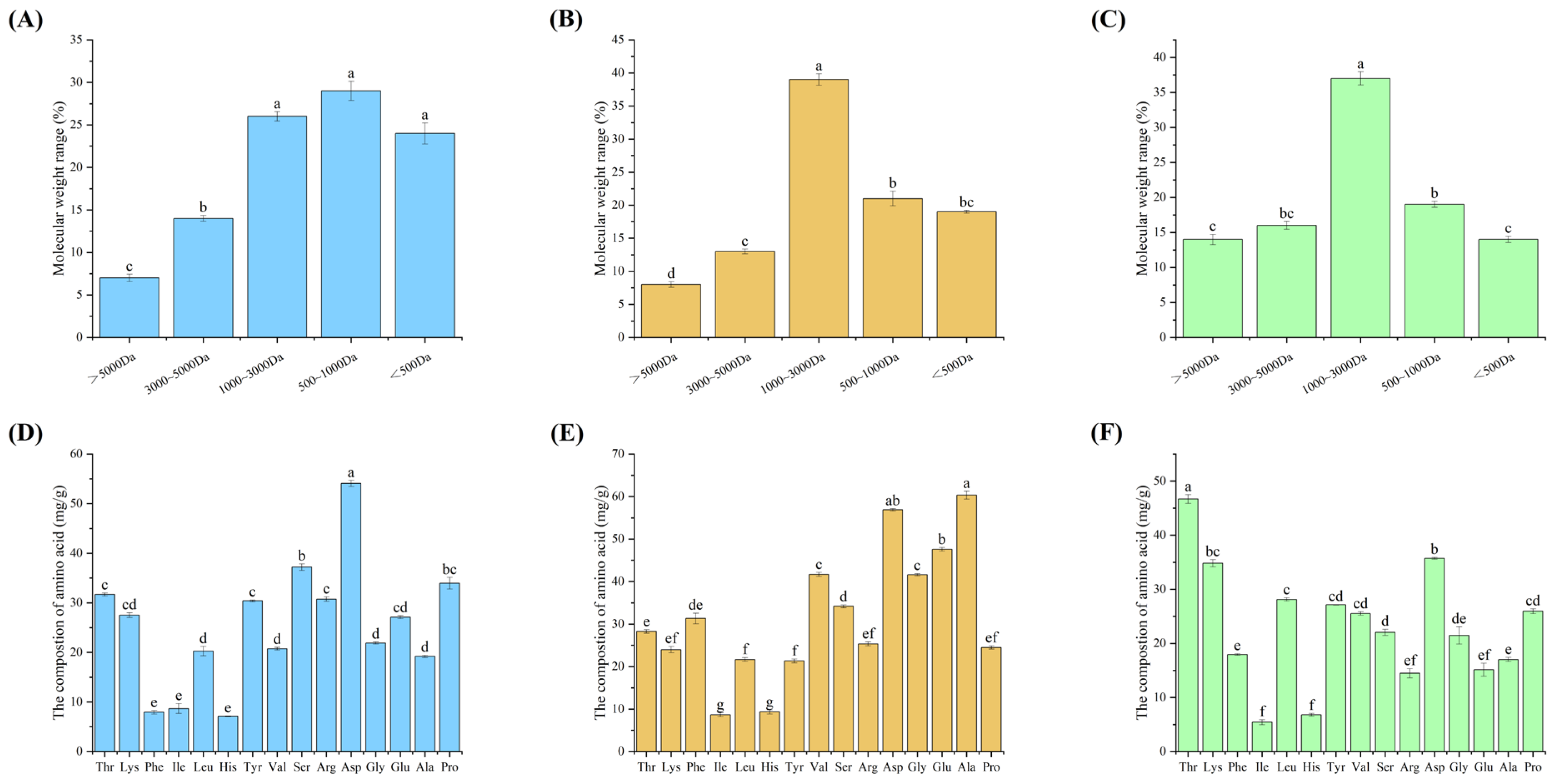
Sea cucumber is an ingredient and medicinal herb with a long history. Its medicinal value has attracted more attention from consumers in recent years [10]. Furthermore, it has been discovered that bioactive peptides, such as those found in sea cucumbers, exhibit greater efficacy when released from the protein sequence of the parent. Low molecular weight peptides produced by proteolytic enzymes also demonstrate higher bioactivity than high molecular weight peptides [25]. Before conducting an in-depth activity analysis and peptide purification, it is necessary to analyse the composition of the enzymatic products. Therefore, the three protein peptides were hydrolysed using neutral protease, and their main components are shown in Figure 1.
The extracted peptides from the body wall, egg, and intestine were predominantly small molecule peptides with a molecular weight of less than 3000 Da, accounting for 80%, 80%, and 73%, respectively. This suggests that neutral protease can effectively release biologically active peptides from proteins (see Figure 1A, 1B, and 1C). The molecular weight distribution of the three protein peptide enzymatic powders was compared. It was found that all three enzymatically extracted peptides had molecular weights mainly concentrated at <3000 Da. However, there was a difference in that protein ovalbumin and protein intestine were more concentrated at 1000~3000 Da, while protein body wall was mainly concentrated at <1000 Da. It is reported that Asp, Ala, Ser, and Lys all have good activity for osteoblasts [12]. The amino acid composition of the three peptides is relatively high in the proteasomal wall peptides, which are hypothesized to have the highest osteogenic activity.
Figure 1.
illustrates the composition analysis of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine). Panels (A, B, C) depict the molecular weight distribution of sea cucumber peptides extracted from different parts (body wall, coelomic fluid, intestine). Panels (D, E, F) show the amino acid composition of sea cucumber peptides extracted from different parts (body wall, coelomic fluid, intestine). Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
Figure 1.
illustrates the composition analysis of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine). Panels (A, B, C) depict the molecular weight distribution of sea cucumber peptides extracted from different parts (body wall, coelomic fluid, intestine). Panels (D, E, F) show the amino acid composition of sea cucumber peptides extracted from different parts (body wall, coelomic fluid, intestine). Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
2.2. Differential Value-Added Profile of the Three Proteolytic Peptides for Osteoblasts
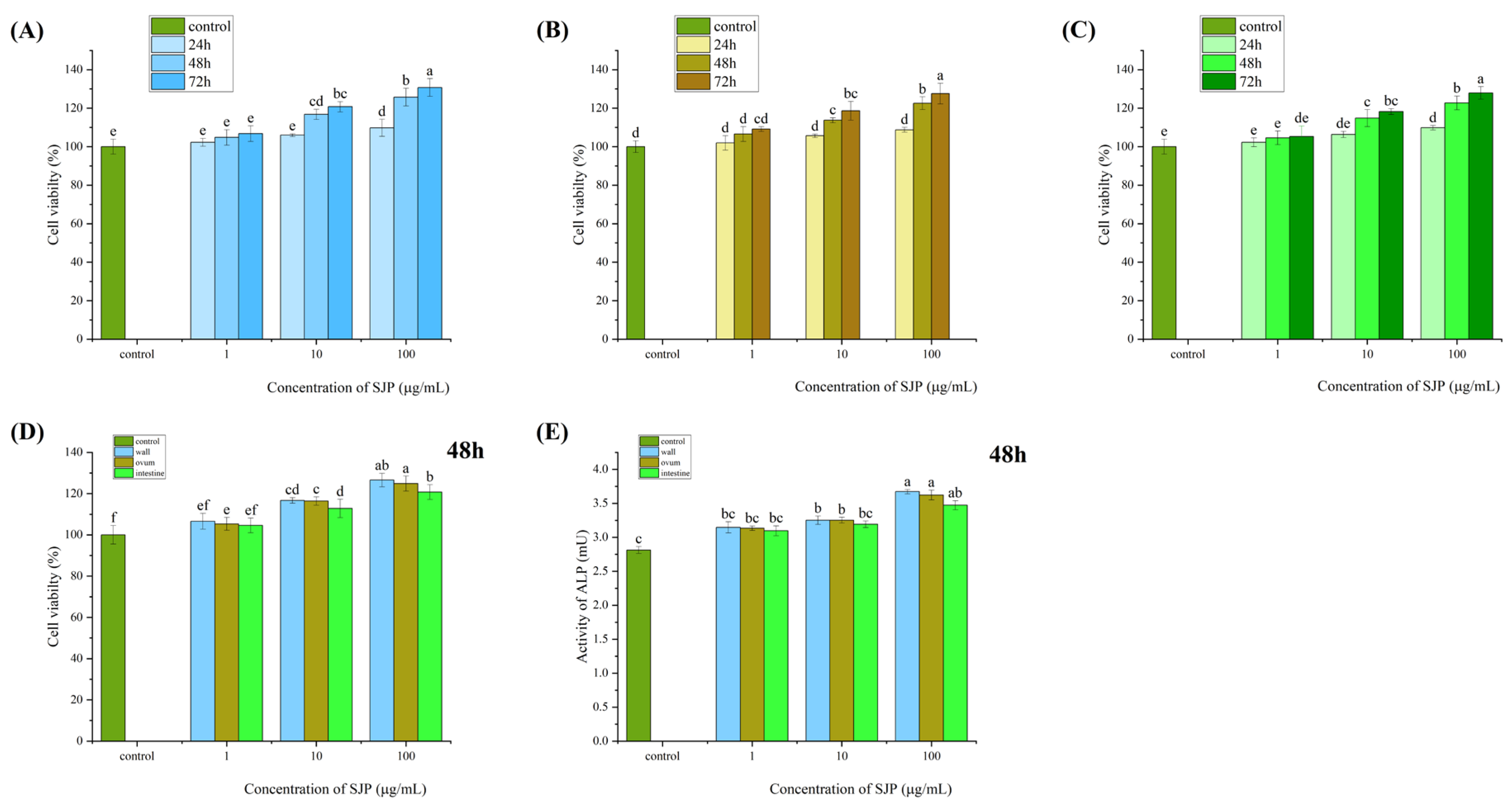
Enzymatic protein peptides at concentrations of 1, 10, and 100 μg/mL significantly promoted cell growth in MC3T3-E1 cells in a time- and concentration-dependent manner. The fastest growth rate was observed at 48 h, and the most prominent osteogenic advantage was observed at a concentration of 100 μg/mL (Figure 2A, 2B, and 2C).
When comparing the value-added of the three proteolytic peptides to osteoblasts and alkaline phosphatase, it was found that treating MC3T3 IE1 cells with the three proteolytic peptide proteins at different concentrations (1, 10, and 100 μg/mL) significantly promoted osteoblast growth and enhanced alkaline phosphatase activity in a time- and concentration-dependent manner. The onset of effects was observed at 48 h. At 48 h, the impact of the three proteolytic peptides on osteoblast growth and alkaline phosphatase activity was observed. The enzymatic peptide extracted from the protein body wall was found to have the most significant utility when compared to the other two protein peptides. The order of overall osteogenesis was protein body wall > protein ovalbumin > protein intestine (Figure 2D and 2E).
Figure 2.
illustrates the analysis and comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level.Panels (A, B, C) show the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation of mouse embryonic osteoblasts.Panels (D, E) depict the comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation and alkaline phosphatase activity of mouse embryonic osteoblasts.Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
Figure 2.
illustrates the analysis and comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level.Panels (A, B, C) show the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation of mouse embryonic osteoblasts.Panels (D, E) depict the comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation and alkaline phosphatase activity of mouse embryonic osteoblasts.Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
2.3. Three Purifications of Sea Cucumber Protein Peptides for Osteoblasts Value-Added Situation
The protein purification system was presented in terms of the differences in sample ionization and anion exchange at 220 nm for elution separation [13]. Four fractions were harvested per sample. Different concentrations (1, 10, and 100 g/mL) of nine sea cucumber peptide-isolated samples were used to treat MC3T3 I E1 cells for 48 h. The two isolated peptides with hydrophobicity and a molecular weight of less than 1kDa showed more significant pro-osteoporotic activity, with the peptide with a molecular weight of less than 1kDa having the most significant effect. Different concentrations (1, 10, and 100 g/mL) of three sea cucumber isolated peptides, classified as <1kDa and <1kDa hydrophobic, were used to treat MC3T3 I E1 cells. The results showed that the proteins significantly promoted cell growth in a time- and concentration-dependent manner. The hydrophobically isolated peptides and those with a molecular weight of less than 1kDa demonstrated a relative advantage over the comparison group consisting of peptides from the same species of sea cucumber across all three time gradients. This advantage was observed in the values for sea cucumber body wall (130%), sea cucumber ovalbumin (117%), and sea cucumber intestine (126%).
2.4. The Enhancement of Each Index after Efficient Purification of Sea Cucumber Protein Peptides
The peptides extracted from sea cucumbers at three different sites were separated and collected based on their hydrophobicity and size (<1kDa). The purified peptide yield was 31% for sea cucumber intestines, 39% for sea cucumber egg, and 37% for sea cucumber body walls. The enhancement rates of cell viability and protein content were determined for the three purified peptides using mouse embryonic osteoblast culture and the Kjeldahl method. The purified peptides from sea cucumber intestine showed a cell viability enhancement rate of 129.3% and a protein content enhancement rate of 112.0%. Similarly, the purified peptide from sea cucumber egg white exhibited a cell viability enhancement rate of 131.2% and a protein content enhancement rate of 109.1%. Finally, the protein content of the purified peptide from sea cucumber body wall increased by 111.5%. All three purified sea cucumber peptides demonstrated a significant enhancement effect, meeting industrial requirements [14].
2.5. Prediction and Screening of Sea Cucumber Peptides' Bone-Enhancing Function Activity
Thyroid hormone PTH is a commonly used peptide composed drug for bone-related conditions such as osteoporosis [15].RANK/RANKL is a relevant pathway of bone metabolism has a direct role in relation to bone synthesis [30]. Integrin AVB3 and integrin α5β are commonly used integrin receptors associated with osteoblast proliferation. Based on the above information, the receptors related to the docking of sea cucumber peptide molecules were selected [16]. And according to the difficulty of enrichment, whether it contains osteogenesis-related amino acids to coarse screening of sea cucumber peptides to select the screening peptide and four receptor proteins were molecularly docked, and the docking results were evaluated by the CDOCKER docking algorithm [17], so as to screen the advantageous osteogenic peptides.
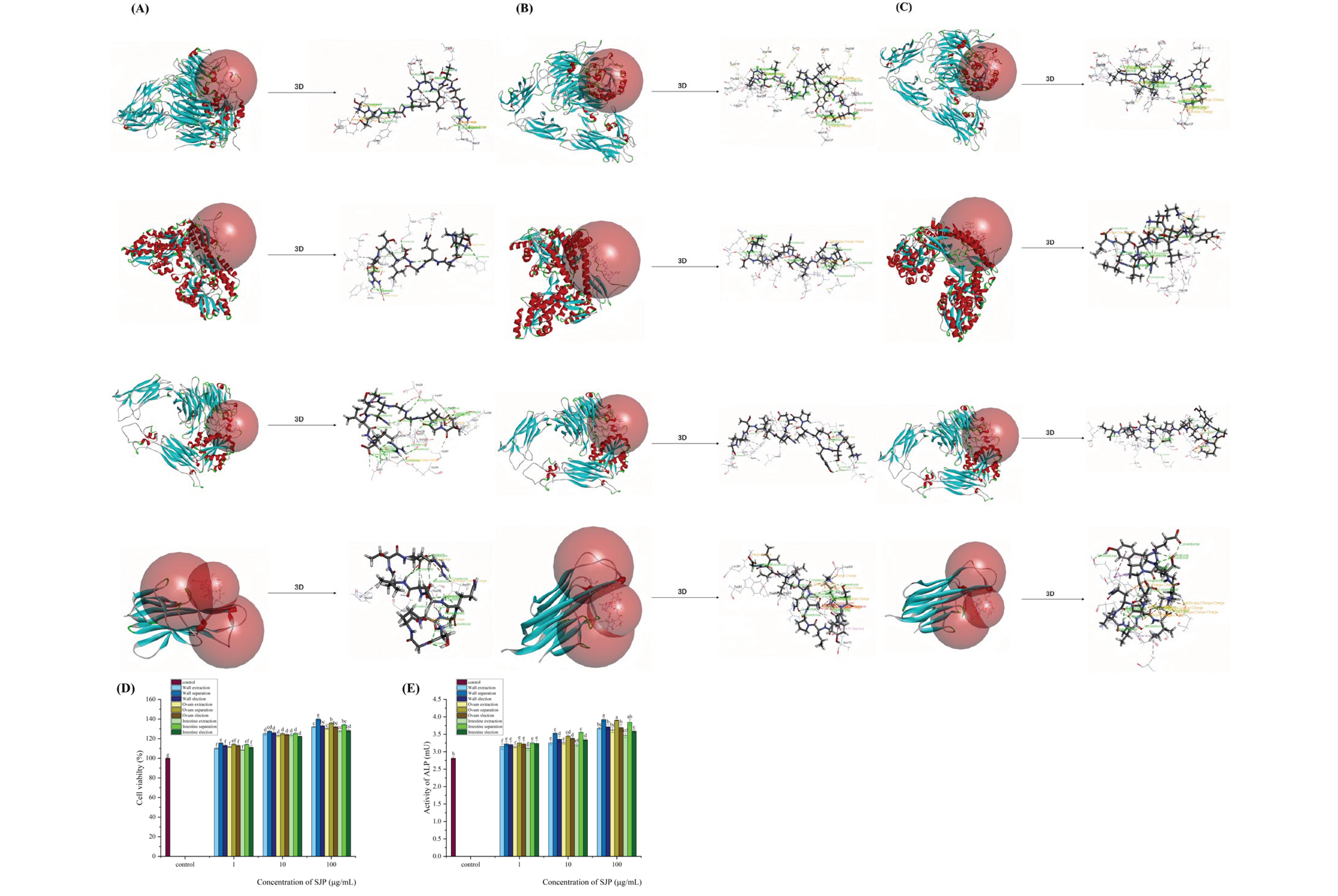
Comparing the docking of each receptor protein with the sea cucumber peptides, five peptide chains were screened for each sea cucumber peptide that successfully docked to all four receptor proteins [18]. and scored according to the CDOCKER docking algorithm (Table 2). Based on the docking scores, sea cucumber body wall KSYELPDGQVITIG, sea cucumber egg white KSYELPDGQVI and sea cucumber intestinal SGEGGQGSLTR were selected as the screened peptides with better molecular docking predictions for osteogenesis.
2.6. Interaction between Bone-Enhaning Sea Cucumber Peptides and Osteogenic Receptor Protein
During the rough screening and molecular docking process of three types of sea cucumber peptides with four osteogenic receptor proteins, peptides that successfully interacted with all four receptor proteins were selected based on their CDOCKER docking scores. Among these, KSYELPDGQVITIG from the body wall, KSYELPDGQVI from the coelomic fluid and SGEGGQGSLTR from the intestine were selected. Notably, the docking score of the body wall peptide KSYELPDGQVITIG was the highest in the CDOCKER algorithm, with scores of 274.357 for integrin AVB3, 236.992 for parathyroid hormone-related protein (PTHrP), 231.575 for integrin α5β1 and 148.756 for mouse RANK/RANKL binding receptor. These conformations provided a theoretical basis for further screening of peptides with osteogenic activity [19].
A comparative analysis of the effect of extracted peptides, separated peptides and screened peptides on osteoblasts and alkaline phosphatase activity was performed. It was observed that the separated peptides from all three parts of the sea cucumber showed the highest osteogenic and alkaline phosphatase activity. Specifically, the order of effectiveness was body wall > coelomic fluid > intestine in terms of separation methods, and separated peptides > screened peptides > extracted peptides in terms of isolation methods. Taking into account the yield and industrial cost of the three separation methods, proteins extracted from the sea cucumber body wall, particularly those with hydrophilicity and molecular weight less than 1 kDa, showed the most significant osteogenic effects.
Figure 3.
presents the analysis and comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level, based on differences in molecular weight distribution, hydrophilic properties, and ionization.Panels (A, B, C) show the purified profiles of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) based on differences in ionization obtained through protein purification systems.Panels (D, E, F) depict the comparison of the effects of various components of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation of mouse embryonic osteoblasts, based on differences in molecular weight distribution, hydrophilic properties, and ionization.Panel (G) analyzes the synergistic effects of protein peptide separation methods from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteoblast proliferation.Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
Figure 3.
presents the analysis and comparison of the effects of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level, based on differences in molecular weight distribution, hydrophilic properties, and ionization.Panels (A, B, C) show the purified profiles of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) based on differences in ionization obtained through protein purification systems.Panels (D, E, F) depict the comparison of the effects of various components of protein peptides extracted from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on the proliferation of mouse embryonic osteoblasts, based on differences in molecular weight distribution, hydrophilic properties, and ionization.Panel (G) analyzes the synergistic effects of protein peptide separation methods from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteoblast proliferation.Data are presented as mean ± SD (n=3), and different superscript letters within different numbers indicate significant differences (p < 0.05).
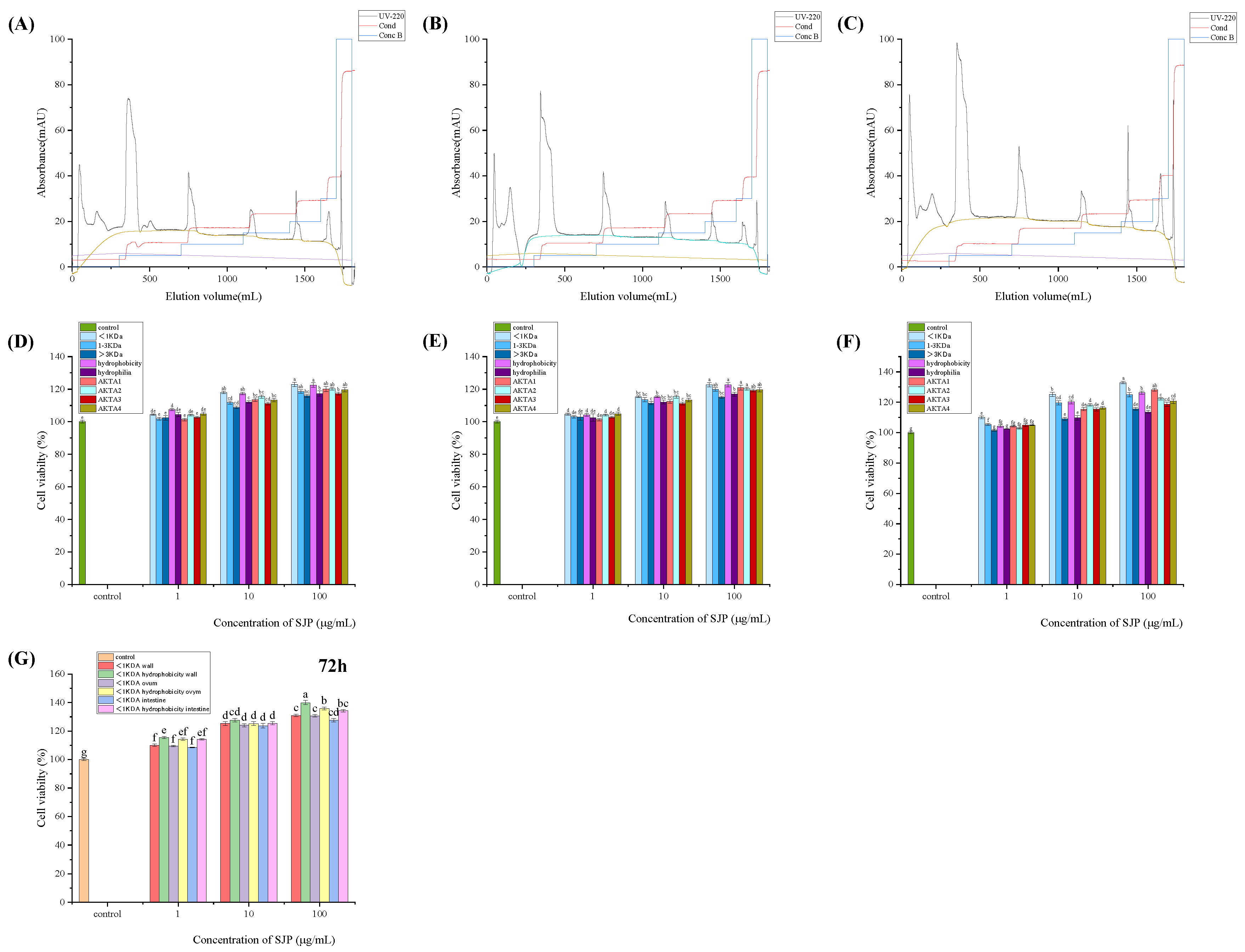

Table 1.
presents the analysis and comparison of the effects of optimized protein peptide extraction from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level, based on differences in molecular weight distribution, hydrophilic properties, and ionization.
Table 1.
presents the analysis and comparison of the effects of optimized protein peptide extraction from three different parts of sea cucumber (body wall, coelomic fluid, intestine) on osteogenic indicators at the cellular level, based on differences in molecular weight distribution, hydrophilic properties, and ionization.
| Component | Cell Viability Enhancement Rate | Protein Content Increase Rate | Yield |
|---|---|---|---|
| Intestine separation | 129.30% | 112.00% | 31.20% |
| Ovum separation | 131.20% | 109.10% | 39.30% |
| Wall separation | 133.70% | 111.50% | 37.80% |
Table 2.
Prediction of Osteogenic Effects of Protein Peptides Extracted from Three Different Parts of Sea Cucumber (Body Wall, Coelomic Fluid, Intestine) by Screening Peptides with Osteogenic Receptor Proteins.
Table 2.
Prediction of Osteogenic Effects of Protein Peptides Extracted from Three Different Parts of Sea Cucumber (Body Wall, Coelomic Fluid, Intestine) by Screening Peptides with Osteogenic Receptor Proteins.
| Specimen | Ligand | Receptor | Arithmetic | Score |
|---|---|---|---|---|
| Intestine extraction | SGEGGQGSLTR | 1L5G | CDOCKER | 274.357 |
| 3C4M | CDOCKER | 236.992 | ||
| 3VI4 | CDOCKER | 232.065 | ||
| 4GIQ | CDOCKER | 148.756 | ||
| AYLGKDVY | 1L5G | CDOCKER | 175.322 | |
| 3C4M | CDOCKER | 135.32 | ||
| 3VI4 | CDOCKER | 143.67 | ||
| 4GIQ | CDOCKER | 139.043 | ||
| TGSKLVVS | 1L5G | CDOCKER | 146.116 | |
| 3C4M | CDOCKER | 100.055 | ||
| 3VI4 | CDOCKER | 153.105 | ||
| 4GIQ | CDOCKER | 127.719 | ||
| KRSIEGGNLVY | 1L5G | CDOCKER | 205.241 | |
| 3C4M | CDOCKER | 160.821 | ||
| 3VI4 | CDOCKER | 147.637 | ||
| 4GIQ | CDOCKER | 160.906 | ||
| GEGGQGSLTR | 1L5G | CDOCKER | 171.331 | |
| 3C4M | CDOCKER | 131.638 | ||
| 3VI4 | CDOCKER | 146.215 | ||
| 4GIQ | CDOCKER | 136.137 | ||
| Ovum extraction | KSYELPDGQVI | 1L5G | CDOCKER | 202.511 |
| 3C4M | CDOCKER | 152.484 | ||
| 3VI4 | CDOCKER | 192.802 | ||
| 4GIQ | CDOCKER | 169.882 | ||
| AGRDLTDYLM | 1L5G | CDOCKER | 195.779 | |
| 3C4M | CDOCKER | 146.028 | ||
| 3VI4 | CDOCKER | 177.333 | ||
| 4GIQ | CDOCKER | 106.331 | ||
| NAPAMYVAIQ | 1L5G | CDOCKER | 166.736 | |
| 3C4M | CDOCKER | 115.185 | ||
| 3VI4 | CDOCKER | 134.672 | ||
| 4GIQ | CDOCKER | 111.114 | ||
| DLAGRDLTDYLMK | 1L5G | CDOCKER | 233.578 | |
| 3C4M | CDOCKER | 167.606 | ||
| 3VI4 | CDOCKER | 198.733 | ||
| 4GIQ | CDOCKER | 184.698 | ||
| VPIYEGY | 1L5G | CDOCKER | 141.316 | |
| 3C4M | CDOCKER | 120.023 | ||
| 3VI4 | CDOCKER | 105.006 | ||
| 4GIQ | CDOCKER | 84.0958 | ||
| Wall extraction | KSYELPDGQVITIG | 1L5G | CDOCKER | 285.931 |
| 3C4M | CDOCKER | 272.861 | ||
| 3VI4 | CDOCKER | 199.712 | ||
| 4GIQ | CDOCKER | 199.459 | ||
| VPIYEGYALPHAILRL | 1L5G | CDOCKER | 177.91 | |
| 3C4M | CDOCKER | 145.818 | ||
| 3VI4 | CDOCKER | 115.815 | ||
| 4GIQ | CDOCKER | 127.528 | ||
| DLAGRDLTDY | 1L5G | CDOCKER | 214.347 | |
| 3C4M | CDOCKER | 164.821 | ||
| 3VI4 | CDOCKER | 203.027 | ||
| 4GIQ | CDOCKER | 189.014 | ||
| IVRDIKEKLNYVAL | 1L5G | CDOCKER | 223.277 | |
| 3C4M | CDOCKER | 214.109 | ||
| 3VI4 | CDOCKER | 204.98 | ||
| 4GIQ | CDOCKER | 241.457 | ||
| AVLSLYASGRTTGIVLDSGDGVTH | 1L5G | CDOCKER | 256.585 | |
| 3C4M | CDOCKER | 214.712 | ||
| 3VI4 | CDOCKER | 151.5 | ||
| 4GIQ | CDOCKER | 189.974 |
Figure 4.
illustrates the predicted osteogenic effects of protein-peptide screening and molecular docking with osteogenic receptor proteins for sea cucumber extracted from three different parts (body wall, coelomic fluid, intestine), as well as the comparative impact of extracted peptides, separated peptides (purified based on hydrophilicity, molecular weight, and ionization conditions), and screened peptides (selected based on molecular docking with osteogenic receptor proteins) on osteoblasts. Panels (A, B, C) depict the 3D schematic representations of the predicted osteogenic activity of screening protein peptides extracted from three parts of the sea cucumber (body wall, coelomic fluid, intestine) interacting with integrin AVB3 (PDB: 1L5G), parathyroid hormone-related protein PTHrP (PDB: 3C4M), integrin α5β1 (PDB: 3VI4), and the mouse RANK/RANKL binding receptor (PDB: 4GIQ). Panels (D, E) show the impact of extracted peptides, separated peptides, and screened peptides from three parts of the sea cucumber on osteoblasts and alkaline phosphatase activity.
Figure 4.
illustrates the predicted osteogenic effects of protein-peptide screening and molecular docking with osteogenic receptor proteins for sea cucumber extracted from three different parts (body wall, coelomic fluid, intestine), as well as the comparative impact of extracted peptides, separated peptides (purified based on hydrophilicity, molecular weight, and ionization conditions), and screened peptides (selected based on molecular docking with osteogenic receptor proteins) on osteoblasts. Panels (A, B, C) depict the 3D schematic representations of the predicted osteogenic activity of screening protein peptides extracted from three parts of the sea cucumber (body wall, coelomic fluid, intestine) interacting with integrin AVB3 (PDB: 1L5G), parathyroid hormone-related protein PTHrP (PDB: 3C4M), integrin α5β1 (PDB: 3VI4), and the mouse RANK/RANKL binding receptor (PDB: 4GIQ). Panels (D, E) show the impact of extracted peptides, separated peptides, and screened peptides from three parts of the sea cucumber on osteoblasts and alkaline phosphatase activity.
3. Discussion
The amino acid composi-tion of the three peptides is relatively high in the proteasomal wall peptides, which are hypothesized to have the highest osteogenic activity.The impact of the three proteolytic peptides on osteoblast growth and alkaline phosphatase activity was observed. The enzymatic peptide extracted from the protein body wall was found to have the most significant utility when compared to the other two protein peptides. The order of overall osteogenesis was protein body wall > protein ovalbumin > protein intestine (Figure 2D and 2E). Proteins significantly promoted cell growth in a time- and concentra-tion-dependent manner. The hydrophobically isolated peptides and those with a mo-lecular weight of less than 1kDa demonstrated a relative advantage over the compari-son group consisting of peptides from the same species of sea cucumber across all three time gradients. Based on the docking scores, sea cucumber body wall KSY-ELPDGQVITIG, sea cucumber egg white KSYELPDGQVI and sea cucumber intestinal SGEGGQGSLTR were selected as the screened peptides with better molecular docking predictions for osteogenesis.. A comparative analysis of the effect of extracted peptides, separated peptides and screened peptides on osteoblasts and alkaline phosphatase activity was performed. It was observed that the separated peptides from all three parts of the sea cucumber showed the highest osteogenic and alkaline phosphatase activity. Specifically, the or-der of effectiveness was body wall > coelomic fluid > intestine in terms of separation methods, and separated peptides > screened peptides > extracted peptides in terms of isolation methods. Taking into account the yield and industrial cost of the three sepa-ration methods, proteins extracted from the sea cucumber body wall, particularly those with hydrophilicity and molecular weight less than 1 kDa, showed the most sig-nificant osteogenic effects.4. Materials and Methods
4. Materials and Chemicals
The MC3T3-E1 cells were acquired from the Cell Bank of the Chinese Academy of Sciences (Beijing, China). The fetal bovine serum (FBS), phosphate-buffered saline (PBS), α-minimal essential medium (α-MEM), penicillin, and streptomycin were purchased from HyClone. The neutral proteinase (Bacillus subtilis) was obtained from Novozymes (China) Biotechnology Co., Ltd. (Tianjin, China). 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) and estradiol were purchased from Sigma-Aldrich (St Louis, MO, USA). C18 solid phase microextraction columns with ultrafiltration centrifuge tubes of 1k and 3kDa were purchased from Beyotime (Tianjin, China). Alkaline phosphatase (ALP), tartrate-resistant acid phosphatase (TRAP) assay kits, and a cell cycle and apoptosis kit were purchased from Beyotime (Shanghai, China).
4.1. Preparation of Sea Cucumber Enzymatic Powder
Fresh wild sea cucumbers caught on the same day were cleaned and the excess internal organs were removed. The three parts of the sea cucumber were then separated into body wall, egg, and intestine. The separated tissues were mixed with distilled water at a solid-liquid ratio of 1:10 (w/v) and homogenized. The homogenate was hydrolyzed with 0.3% neutral protease (SARE peptidase) at 50°C and pH 6.8 [20]. Stirring was continued at 60 rpm during hydrolysis. Two hours later, the supernatant of the enzymatic hydrolysis products was collected. The salt was removed from the enzymatic hydrolysis product by dialysis using a Cato500Da membrane. The dialyzed enzymatic products were freeze-dried to obtain proteolytic powders from the three sites.
4.2. Amino Acid Composition and Molecular Weight Distribution of Sea Cucumber Enzyme Powder Extracted from Three Sites
The amino acid assays for three sea cucumber peptides were filtered through a 0.22 μm membrane with buffer prior to the assay. The amino acid standards (40 μl) were dissolved in 0.02 M hydrochloric acid. Similarly, the samples (at a concentration of 3 mg/ml) were dissolved in 0.02 M hydrochloric acid and injected (20 μl) for detection at a wavelength of 570 nm. The amino acid content was calculated from the corresponding integrated area of each amino acid in the analyzer detector[20]. The peptides' relative molecular weights were determined using a high-performance liquid chromatograph (Waters, USA). The samples were dissolved in ultrapure water at a concentration of 2 mg/ml and passed through a 0.22 μm filter membrane[21]. The analysis was performed on a semi-preparative liquid-phase piggyback column TSKgel G2000SWXL (7.8 × 300 mm) with an absorption wavelength of 280 nm. The mobile phase consisted of 45% pure acetonitrile (with 0.1% trifluoroacetic acid) and eluted at a flow rate of 0.5 ml/min. Standards with varying molecular weights were used, including glycine (MW: 75), glutathione (MW: 307), vitamin B12 (MW: 1355), peptidase (MW: 6512), and cytochrome C (MW: 12500). The relative molecular weight distributions were obtained by plotting the logarithmic molecular weights of the standards against the retention time of the fitted curves. The distribution of molecular weight was determined by integrating the area of the molecular weight segments from the size curve obtained.
4.3. Identification of Enzymatic Peptide Sequences of Sea Cucumber Extracted from Three Sites by UPLC-Q-TOF Coupled with CESI-Q-TOF
The peptides were separated using ultra-high performance liquid phase (UPLC) on an Ultimate3000 column packed with C18 (3 μm pore size, 110 Å packing particle size). The mobile phase A consisted of an aqueous solution of formic acid with 0%. The mobile phase A consisted of 0.1% formic acid in water (0.1% FA-H2O), while the mobile phase B was an acetonitrile solution (0.1% FA-ACN) containing 0.1% formic acid in acetonitrile solution (0.1% FA-ACN). The total chromatographic separation time was 70 min at a flow rate of 0.3 mL/min, using gradient elution under the following conditions: The mobile phase B composition was 5-12% from 0-12 min, 10-30% from 12-46 min, 30-50% from 46-50 min, 50-85% from 50-62 min, and 85-5% from 62-70 min. The injection volume was 10 μL. The electrospray ion source (ESI source) was used to carry out the separation, which was connected to the UPLC system and Q-TOF (impact II, Bruker Daltonic GmbH, Bremen, Germany). The source parameters used for UPLC-Q-TOF-MS were: dry temperature of 200°C, dry gas rate of 9.0 L/min, and nebulizer gas of 1.5 Bar. Peptide capillary electrophoretic separation was performed using a CESI8000Plus system (ABSciex, Inc., Redwood City, CA, USA) with data acquisition parameters set to Automatic MS/MS mode. A fused silica capillary with an inner diameter of 30 μm and a total length of 91 cm, without any coating, was inserted into the interface [22]. The background electrolyte (BGE) buffer used was a 10% acetic acid solution with a pH of approximately 2.3, while the conductive solution was a 100 mmol/L ammonium acetate solution with a pH of 4.0. Samples were injected using a positive air pressure method after rinsing. Prior to loading each specimen, the capillary was rinsed successively with 0.1 M NaOH solution for 3 min, 0.1 M HCl solution for 3 min, ultrapure water for 4 min, and BGE for 4 min [23]. The peptide samples were then loaded into the sample at 20psi pressure for 60 s and concentrated and enriched with BGE at 20psi pressure for 30 s.
4.4. Purification and Separation of Sea Cucumber Peptides based on Differences in Molecular Weight Distribution, Hydrophilicity and Ionization
The peptides from sea cucumber were separated using a C18 solid phase extraction column to obtain both hydrophilic and hydrophobic fractions [24]. The extraction column was activated with 5 mL of 100% methanol and equilibrated with 5 mL of 0.1% formic acid. The hydrophilic peptide isolation samples were eluted three times with 3 mL of 0.1% formic acid, while the hydrophobic peptide isolated samples were eluted three times with 3 mL of 80% methanol[25]. The peptides from sea cucumber were separated using two ultrafiltration centrifuge tubes with molecular weight cutoffs of 1kDa and 3kDa to obtain fractions with molecular weight distribution in the intervals of >3kDa, 1-3kDa, and <1kDa. Prior to passing through the aqueous membrane, the ultrafiltration centrifuge tubes were activated by pre-cooling them in a refrigerator for five minutes with de-ionized water. The tubes were equilibrated to ensure a balanced sample mass and center of gravity. The sea cucumber peptide samples were finally centrifuged at 10000c×g for 20 min. The peptides were then separated into protein peptides of the target three intervals using ultrafiltration centrifuge tubes with 3kDa and 1kDa. Subsequently, the sea cucumber peptides were separated by ionization difference using the AKTA protein purification system. A single injection of 2 mL (5 mg/mL) of sea cucumber peptides was loaded into the AKTA protein purification system [26]. The anion exchange column was eluted with water at a flow rate of 0.2 mL/min and equilibrated. The elution was checked at 220 nm at 4°C. The eluate was collected into a 96-well plate at a concentration of 1mL per well [27].
4.5. Molecular Docking Predicts the Analysis of Osteoactive Peptides in Sea Cucumbers
Molecular docking was performed using Discovery Studio 2017 software (Biovia). The lectins (PDB: 3VI4, 1L5G, 3C4M, 4GIQ) were docked to peptides with potential bone-promoting activity [28]. The co-crystal structures of the RGD peptide ligands and the lectins can be downloaded from the Protein Data Bank (PDB: 3VI4, 1L5G, 3C4M, 4GIQ). The crystal structures of the integrally linked proteins (PDB: 3VI4, 1L5G, 3C4M, 4GIQ) were used as template molecules for the target peptides. The peptides were then screened using a counting method [29]. Computational analyses were carried out using CDOCKER and LibDOCKER in Discovery Studio software.
4.6. Cell Culture and MTT Assay
MC3T3-E1 cells were cultured in α-MEM medium supplemented with 1% antibiotics (100 U/mL penicillin and 100 g/mL streptomycin) and 10% fetal bovine serum at 37°C in a humidified environment. The MTT assay was used for cell detection. The cells were inoculated into 96-well culture plates (5 × 104 cells per mL) containing 100 μL of α-MEM [30]. MC3T3-E1 cells were incubated for 24 h and then treated with protein peptides at concentrations of 0, 1, 10, and 100 μg/mL at 37°C for 24, 48, and 72 h. After treatment, 10 μL of MTT solution (5 mg/mL in PBS) was added to each well and incubated for 4 h. The wells were then replaced with 150 μL DMSO and measured at 570 nm using a Microplate Reader (Infinite™ M200, TECAN, Switzerland) [31].
4.7. Alkaline Phosphatase (ALP) Activity Assay
MC3T3-E1 cells were treated with proteolytic peptides at concentrations of 0, 1, 10, and 100 μg/mL for 72 h. The culture fluid was collected and alkaline phosphatase (ALP) activity was evaluated following the instructions provided in reference [32].5. Conclusions
The sea cucumber peptides extracted from three distinct sites exhibited a dose-dependent enhancement of osteoblast proliferation and differentiation. Upon evaluating the three methods of sea cucumber peptide separation based on molecular weight distribution, hydrophilicity, and ionization, it was observed that the fraction isolated by hydrophobicity and <1KDa demonstrated the most pronounced osteogenic effect. The molecular docking of the three sea cucumber peptides with osteogenesis-related receptor proteins, namely integrin AVB3, parathyroid hormone PTHrP, integrin α5β1, and mouse RANK/RANKL binding-related receptor, resulted in the selection of three osteogenic peptides based on their docking scores. These peptides are sea cucumber wall KSYELPDGQVITIG, sea cucumber ovalbumin KSYELPDGQVI, and sea cucumber intestine SGEGGQGSLTR. It was also found that the three isolated peptides showed the most significant osteogenic effects relative to each other.
Acknowledgments
This study was financially supported by the National Key Research and Development Plan Project (2023YFD2401404).
References
- Jingwei J ,Shan G ,Zelong Z , et al.A novel short-type peptidoglycan recognition protein with unique polysaccharide recognition specificity in sea cucumber, Apostichopus japonicus.J].Fish shellfish immunology,2023,144109263-109263.
- Yingqiu Z ,Huachen L ,Xiao C , et al.A potential feeding regulation strategy during aestivation: Relaxation of intestine mediated by pedal peptide/orcokinin-type neuropeptides in sea cucumber Apostichopus japonicusJ].Aquaculture,2024,579.
- Cheng, X.; Zhao, K.; Zha, X.; Du, X.; Li, Y.; Chen, S.; Wu, Y.; Li, S.; Lu, Y.;Zhang, Y. Opportunistic Screening Using Low-Dose CT and the Prevalence of Osteoporosis in China: A Nationwide, Multicenter Study. J. Bone Miner. Res.2021, 36 (3), 427−435Wei Q ,Holle A ,Li J , et al.BMP-2 Signaling and Mechanotransduction Synergize to Drive Osteogenic Differentiation via YAP/TAZ.J].Advanced science (Weinheim, Baden-Wurttemberg,Germany),2020,7(15):1902931-1902931.
- Tianming W ,Zheng C ,Zhangfei S , et al.Existence and functions of a kisspeptin neuropeptide signaling system in a non-chordate deuterostome species.J].eLife,2020,9.
- Qiufang B ,Ming Z ,Jizhi Z , et al.NGF mediates protection of mesenchymal stem cells-conditioned medium against 2,5-hexanedione-induced apoptosis of VSC4.1 cells via Akt/Bad pathway.J].Molecular and cellular biochemistry,2020,469(1-2):53-64.
- Li X ,Dong Y ,Yin H , et al.Mesenchymal stem cells induced regulatory dendritic cells from hemopoietic progenitor cells through Notch pathway and TGF-β synergisticallyJ].Immunology Letters,2020,222(prepublish):49-57.
- Xingwei S ,Weiwei Z ,Chen Q , et al.Focal adhesion kinase promotes BMP2-induced osteogenic differentiation of human urinary stem cells via AMPK and Wnt signaling pathways.J].Journal of cellular physiology,2020,235(5):4954-4964.
- Bowen M ,Dongle W ,Yangfan C , et al.Interleukin-20 differentially regulates bone mesenchymal stem cell activities in RANKL-induced osteoclastogenesis through the OPG/RANKL/RANK axis and the NF-κB, MAPK and AKT signalling pathways.J].Scandinavian journal of immunology,2020,91(5):e12874.
- Hu Z ,Cao X ,Guo M , et al.Identification and characterization of a novel short-type peptidoglycan recognition protein in Apostichopus japonicusJ].Fish and Shellfish Immunology,2020,99257-266.
- Chaiyamoon A ,Tinikul Y ,Chaichotranunt S , et al.Existence of two mature sequences of cubifrin neuropeptide and their effects on spawning in the sea cucumber, Holothuria scabraJ].Aquaculture,2020,519734753-734753.
- Chaiyamoon A ,Tinikul R ,Nontunha N , et al.Characterization of TRH/GnRH-like peptides in the sea cucumber, Holothuria scabra , and their effects on oocyte maturationJ].Aquaculture,2020,518734814-734814.
- Chieu D H ,Suwansa-ard S ,Wang T , et al.Identification of neuropeptides in the sea cucumber Holothuria leucospilotaJ].General and Comparative Endocrinology,2019,283113229.
- Dinh H C ,Luke T ,K M S , et al.Aquaculture Breeding Enhancement: Maturation and Spawning in Sea Cucumbers Using a Recombinant Relaxin-Like Gonad-Stimulating Peptide.J].Frontiers in genetics,2019,1077.
- Chen M ,Talarovicova A ,Zheng Y , et al.Neuropeptide precursors and neuropeptides in the sea cucumber Apostichopus japonicus: a genomic, transcriptomic and proteomic analysisJ].Scientific Reports,2019,9(1):1-22.
- Cusimano G M ,Spinello A ,Barone G , et al.A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenesJ].Marine Drugs,2019,17(3):159.
- Xianliang L ,Wangxin L ,Baodong Z , et al.Sea cucumber peptides positively regulate sexual hormones in male mice with acute exhaustive swimming: possibly through the Casup2+/sup/PKA signaling pathway.J].Food function,2023,.
- Cong X ,Liu H ,Zheng Y , et al.A Putative Role of Vasopressin/Oxytocin-Type Neuropeptide in Osmoregulation and Feeding Inhibition ofApostichopus japonicusJ].International Journal of Molecular Sciences,2023,24(18):.
- Xueying G ,Kui D ,Libin Z .The effects of kisspeptin-type neuropeptides on feeding behavior and intestinal metabolism in the sea cucumber, Apostichopus japonicusJ].Aquaculture,2023,571.
- Xueying G ,Libin Z ,Kang X .Effect of Kisspeptin-Type Neuropeptide on Locomotor Behavior and Muscle Physiology in the Sea Cucumber Apostichopus japonicusJ].Animals,2023,13(4):705-705.
- Jiahui X ,G S V .Critical role of the mTOR pathway in poultry skeletal muscle physiology and meat quality: an opinion paper.J].Frontiers in physiology,2023,141228318-1228318.
- Ziqian W ,Xudong W ,Jingcun S , et al.Identification of Functional Modules and Key Pathways Associated with Innervation in Graft Bone-CGRP Regulates the Differentiation of Bone Marrow Mesenchymal Stem Cells via p38 MAPK and Wnt6/β-Catenin.J].Stem cells international,2023,20231154808-1154808.
- Retraction: MNK1-Induced eIF-4E Phosphorylation in Myeloma Cells: A Pathway Mediating IL-6-Induced Expansion and Expression of Genes Involved in Metabolic and Proteotoxic Responses.J].PloS one,2023,18(9):e0291491-e0291491.
- Hongxu L ,Zhaoxia Z ,Zhaoying W , et al.Human Mesenchymal Stem Cells-Derived Exosome Mimetic Vesicles Regulation of the MAPK Pathway and ROS Levels Inhibits Glucocorticoid-Induced Apoptosis in Osteoblasts.J].Stem cells international,2023,20235537610-5537610.
- W F F ,R R M ,B P M , et al.Effects of HSP70 chaperones Ssa1 and Ssa2 on Ste5 scaffold and the mating mitogen-activated protein kinase (MAPK) pathway in Saccharomyces cerevisiae.J].PloS one,2023,18(10):e0289339-e0289339.
- Mathematical A C M I M .Retracted: miR-138 Reduces the Dysfunction of T Follicular Helper Cells in Osteosarcoma via the PI3K/Akt/mTOR Pathway by Targeting PDK1.J].Computational and mathematical methods in medicine,2023,20239765985-9765985.
- Bo C ,Weidong S ,Song C , et al.Bone Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Ameliorated Lipopolysaccharide-Induced Lung Injury Via the miR-21-5p/PCSK6 Pathway.J].Journal of immunology research,2023,20233291137-3291137.
- Yuko K ,Miho K ,Christella W , et al.Regulation of Syk activity by antiviral adaptor MAVS in FcεRI signaling pathway.J].Frontiers in allergy,2023,41098474-1098474.
- Kyungsoo K ,Gyeom M K ,Min G L .Improving bone morphogenetic protein (BMP) production in CHO cells through understanding of BMP synthesis, signaling and endocytosisJ].Biotechnology Advances,2023,62108080-108080.
- Xi T ,Danning W ,Pei L , et al.Bone marrow mesenchymal stem cells alleviate stress-induced hyperalgesia via restoring gut microbiota and inhibiting neuroinflammation in the spinal cord by targeting the AMPK/NF-κB signaling pathway.J].Life sciences,2022,314121318-121318.
- Yingqiu Z ,Xiao C ,Huachen L , et al.Nervous System Development and Neuropeptides Characterization in Embryo and Larva: Insights from a Non-Chordate Deuterostome, the Sea Cucumber Apostichopus japonicusJ].Biology,2022,11(10):1538-1538.
- Hao Y ,Yingying T ,Xiaoxuan F , et al.Novel Peptides Derived from Sea Cucumber Intestine Promotes Osteogenesis by Upregulating Integrin-Mediated Transdifferentiation of Growth Plate Chondrocytes to Osteoblasts.J].Journal of agricultural and food chemistry,2022,70(41):.
- Y. C Z ,Qi Z ,Q. Q P , et al.Analysis of lncRNA in the skeletal muscle of rabbits at different developmental stages#13;J].Frontiers in Veterinary Science,2022,9948929-948929.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



