
Submitted:
30 March 2024
Posted:
01 April 2024
You are already at the latest version
Abstract
Stem cell-based regeneration strategies have shown therapeutic efficacy in various fields of regenerative medicine. These include bone healing after bone augmentation, often complicated by pain, that is managed by using non-steroidal anti-inflammatory drugs (NSAIDSs). Information is limited about how NSAIDSs affect the therapeutic potential of stem cells. Therefore, we investigated the effects of ibuprofen and diclofenac in low and high therapeutic doses on the properties of human mesenchymal stromal cells isolated from the dental pulp (DPSCs) and cultured in vitro. Ibuprofen and diclofenac significantly reduced the viability of DPSCs, while the expression of mesenchymal stem cell surface markers was unaffected. Both ibuprofen and diclofenac treatment significantly upregulated the expression of HGF, while the expression of VEGFA remained unchanged. Ibuprofen significantly altered the expression of several apoptosis-related genes, including upregulation of CASP9 and BCL2, with decreased CASP3 expression. BAK, CASP3, CASP9, and BCL2 expressions were significantly increased in the diclofenac-treated DPSCs, while no difference was demonstrated in BAX expression. Our results suggest that concomitant use of the NSAIDSs ibuprofen or diclofenac with stem cell therapy may negatively impact cell viability and alter the expression of apoptosis-related genes, which can affect the efficacy of stem cell therapy.
Keywords:
dental pulp stem cells
; nonsteroidal anti-inflammatory drugs
; ibuprofen
; diclofenac
; angiogenesis
; apoptosis
1. Introduction
Stem cell-based tissue engineering approaches offer important therapeutic modalities in numerous medical disciplines. These include bone augmentation in oral/maxillofacial surgery for bone defect treatment and periodontal reconstruction [1,2]. The most common treatments to restore extensive bone loss and defects are autogenous bone grafts, allograft, xenograft, isograft or alloplastic material, which remain a standard procedure for significant defects [3]. However, these approaches have many limitations and shortcomings in restoring morphological and functional reconstruction of defects [4,5].
To overcome the difficulties associated with grafting procedures, intensive research and development of bone graft alternatives employ different combinations of osteoconductive materials, growth factors, and stem cells [6]. The goal of regenerative medicine is to replace the damaged area with tissue specific to the patient at cellular and immunological levels. Therefore, a promising alternative to standard therapy is using collagen scaffolds in combination with cells possessing osteogenic potential, i.e., stem cells [7,8]. Mesenchymal stem cells (MSCs) are multipotent stromal stem cells that can be harvested from various sources and differentiated into multiple cell types, such as osteogenic chondroblasts and osteoblasts [9]. The beneficial therapeutic effects of MSCs are due to their ability to support a regenerative microenvironment through immunomodulatory effects, stimulation of angiogenesis and antiapoptotic effects, as well as recruitment of host stem/ progenitor cells into the site of bone repair [10,11]. The efficacy of regenerative therapy using stem cells depends on several factors, including the delivery method, the concentration of stem cells per injection, the vehicle used and the extent of injury [12,13]. Moreover, the outcome of regenerative therapy depends on a combination of the interactions between transplanted MSCs and the recipient's cellular and molecular components, as well as any current pharmacotherapy of the patient with effects on MSC and bone microenvironment [14].
Clinical recommendations for the postsurgical pain after bone augmentations in oral surgery include non-steroidal anti-inflammatory drugs (NSAIDSs), such as ibuprofen (IBU) and diclofenac (DIC) [15]. Despite their extensive use, a clear understanding of their mechanism is still lacking. The exact mechanism of action is not entirely known. Still, the primary mechanism responsible for anti-inflammatory, antipyretic, and analgesic action is the inhibition of prostaglandin synthesis by the inhibition of cyclooxygenase (COX) [16]. Some studies suggested that NSAIDSs have inhibitory effects on MSCs, specifically the secretion of regenerative factors [17,18]. Therefore, this study aimed to investigate the effects of ibuprofen and diclofenac on the characteristics and gene expression of human mesenchymal stromal cells isolated from the dental pulp (DPSCs) from patients and cultured in vitro. We focused on the expression of MSC markers, viability, and gene expression of apoptosis-related genes in ibuprofen and diclofenac pre-treated DPSCs.
2. Results
2.1. Characterisation of Isolated DPSCs populations
All cell populations isolated from human dental pulps used in the experiments displayed mesenchymal-like morphology involving spindle-shaped cell types (Figure 1a). The immunophenotype evaluated using flow cytometry showed specific surface antigen expression typical for MSCs (CD90, CD 73, CD 105), while more than 98% of cells were negative for the non-MSC markers, CD14, CD 20, CD34 and CD45 (Figure 1b,c).
2.2. Effect of ibuprofen and diclofenac pre-treatment on characteristics, morphology and immunophenotype of DPSCs
Immunofluorescence staining of untreated and pre-treated DPSCs was used to investigate changes in the expression of cytoskeletal intermediate filament vimentin and nuclei of cells. Treatment with NSAIDSs showed no significant differences in expression between DPSCs groups (Figure 2a). Regarding cellular morphology, images showed elongated cells with no significant morphological difference between groups. Surface markers were analysed with flow cytometry, with no differences in expression observed between NSAIDSs-treated and untreated DPSCs (Figure 2b).
2.2. Effect of ibuprofen and diclofenac pre-treatment on viability of DPSCs
Viability was evaluated by flow cytometry using propidium iodide staining to determine the effect of ibuprofen and diclofenac pre-treatment on DPSCs. Cell viability analysis showed decreasing patterns in both treatments of DPSCs (Figure 2a,b). DPSCs cultivated with low-dose ibuprofen for 72 hours resulted in significantly reduced cell viability by 2.5% (mean 94.9±1.4%) compared to the control (mean 97.4± 0.16%, p ≤ 0.0061). Similarly, high-dose diclofenac supplementation for 72 hours led to a 2.58% decrease in viability (mean 95.6±1.4%) compared to the control (mean 98.1±0.19%, p ≤ 0.0242). The reduction in the survival of hDPSCs after ibuprofen treatment was not observed in the cytotoxicity assay. The decrease in the viability of hDPSCs was observed with the increase in the concentration of diclofenac in 48 hour intervals (mean 83.6±7.7%) compared to control (100±1.4%, p ≤ 0.043).
An MTT assay was performed to assess potential differences in the proliferation rate between DPSCs groups. The results revealed no statistically significant differences between the control and treated groups. In addition, no proportional relation was observed between the concentrations of the tested drugs.
2.3. Effect of ibuprofen and diclofenac on angiogenic growth factors expression
One of the essential components of bone repair is angiogenesis. Therefore, the mRNA expression level of vascular endothelial growth factor α (VEGFA) and hepatocyte growth factor (HGF) were measured by qRT-PCR in cells pre-treated with lower or higher concentrations of ibuprofen and diclofenac for 24, 48 and 72 hours (Figure 3). There were no statistically significant differences between untreated or treated DPSCs in VEGFA expression. Analysis of HGF expression revealed significant changes after pre-treatment with ibuprofen and diclofenac. Specifically, HGF transcript was significantly upregulated in DPSCs cultured with high-dose ibuprofen (p ≤ 0.0040) at 72-hour intervals and in DPSCs treated with low-dose (p ≤ 0.0392) and high-dose diclofenac (p ≤ 0.0040) for 72 hours (Figure 3a,b).
2.4. Ibuprofen and diclofenac significantly affect mRNA expression of selected genes in apoptosis signalling pathways
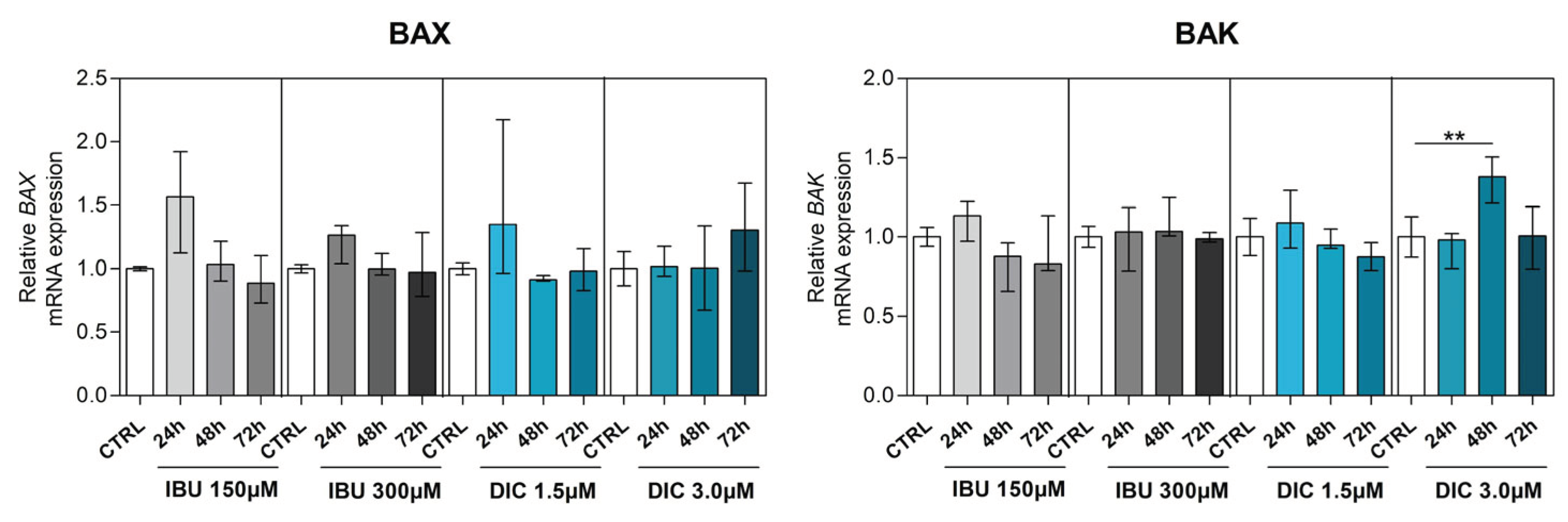
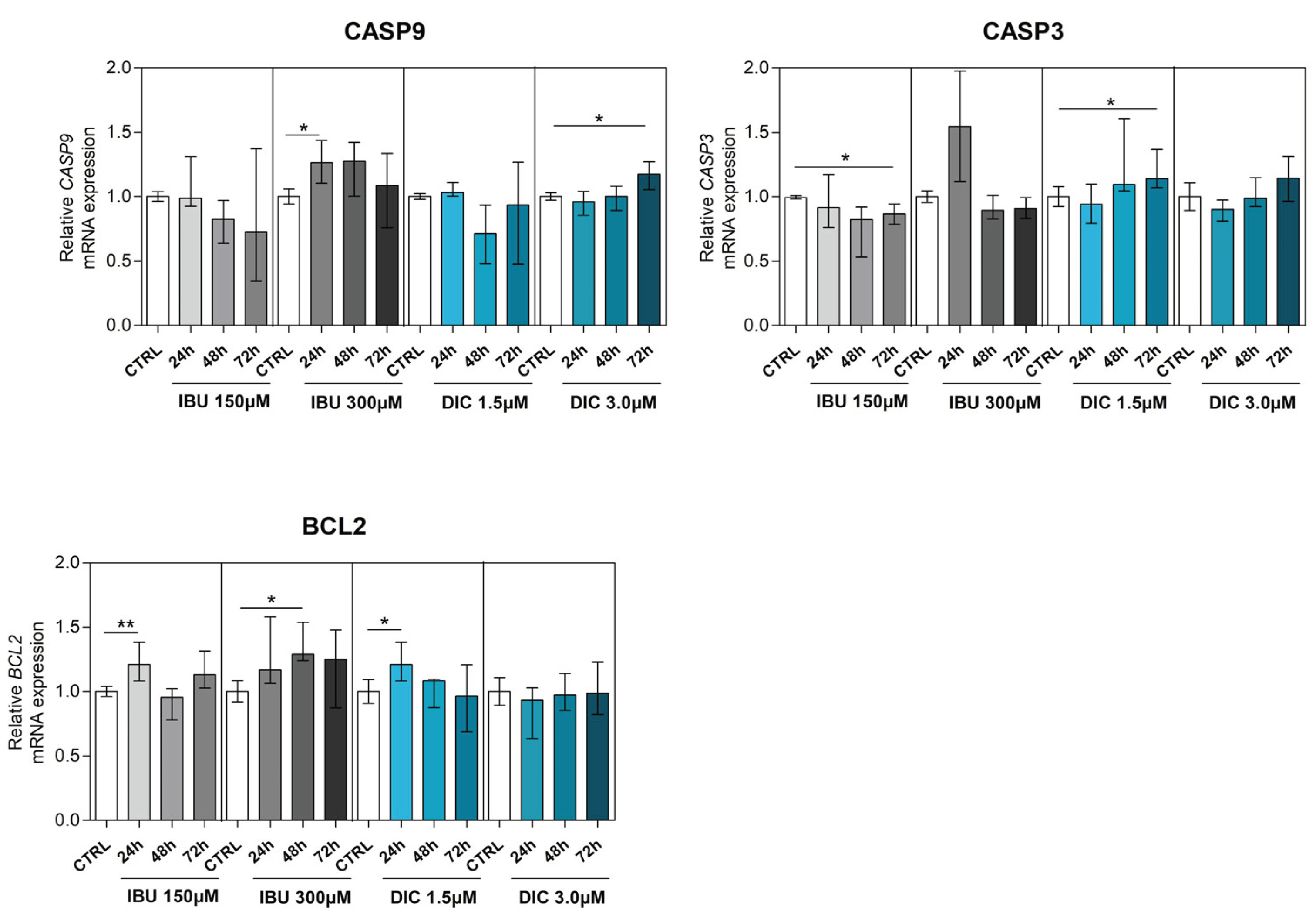
To determine the effect of NSAIDSs on genes associated with apoptosis, BCL2 associated X (BAX), BCL2 antagonist (BAK), caspase 3 (CASP3), caspase 9 (CASP9) and B-cell lymphoma 2 (BCL2) were analysed in DPSCs. BAX and BAK, which are members of the Bcl-2 family and regulators of the intrinsic apoptosis pathway, were slightly changed compared to the control groups. While there was no statistically significant difference in BAX gene expression after pre-treatment, a significant upregulation of BAK expression was shown in DPSCs treated with DIC for 48 hours (p ≤ 0.0040) (Figure 4a,b). Regarding CASP9 expression, low-dose NSAIDSs pre-treatment did not cause changes compared to control groups. However, CASP9 expression was significantly upregulated in DPSCs cultivated with 300µM ibuprofen for 24 hours (p ≤ 0.0162), similarly in 72 hour intervals with 3.0µM diclofenac treatment (p ≤ 0.0172) (Figure 4c). By evaluating the expression of CASP3 in DPSCs, we found that ibuprofen and diclofenac supplementation led to diverse effects. While low-dose ibuprofen led to a significant decrease in CASP3 expression in 72 hour intervals compared to control (p ≤ 0.0121), high-dose ibuprofen had no significant effect. In contrast, low-dose diclofenac significantly upregulated CASP3 expression in 72 hours (p ≤ 0.0162), whereas its high-dose treatment had no effect compared to control in none of the time intervals (Figure 4d).
BCL2, a gene associated with preventing apoptosis and reducing oxidative stress, was significantly upregulated in DPSCs in almost all treatment regimens. Low-dose supplementations with ibuprofen led to a significant increase in BCL2 expression in 24-hour intervals (p ≤ 0.0081). In contrast, high-dose ibuprofen treatment caused significant upregulation in BCL2 expression in 48 hour intervals (p ≤ 0.0107) compared to control. DPSCs treated with low-dose diclofenac had significantly increased BCL2 expression in 24-hour time intervals (p ≤ 0.0283) compared to the control group (Figure 4e). (a) (b) (c) (d) (e)
3. Discussion
Bone healing after bone augmentation is a complex and coordinated process involving many different cell types and multiple cascades of mechanisms and signalling pathways [19,20]. Over the past several years, stem cell-based regeneration strategies have shown great promise for bone healing through endogenous restoration or exogenous transplantation of stem cells [21,22,23]. However, the therapeutic efficacy of stem cell-mediated regeneration is under solid control of the recipient microenvironment, which regulates resident MSCs and the regenerative efficacy of transplanted MSCs [24,25]. Therefore, there is a great effort to improve the endogenous microenvironment or to enhance exogenous MSC resistance, thus benefiting from transplanted MSCs. Besides mentioned, bone healing can be affected by various other factors, such as the extent of damage, age or nutrition, and the administration of several pharmacological agents [26].
Postsurgical pain is mainly driven by inflammation, with the production of prostaglandins in the periphery and central nervous system, which are the principal components of initiation and propagation of pain [15]. Non-steroidal anti-inflammatory drugs (NSAIDS), due to their anti-inflammatory, analgesic and antipyretic properties, are widely used in the treatment of post-operative pain [27,28]. Dental postsurgical pain is also mainly caused by inflammation, with cyclooxygenase-derived prostaglandins (PG) being the significant sensitisers of free nerve ending, compared to other mediators of pain such as histamine, bradykinin, adenosine triphosphate or low pH [29]. NSAIDS exert anti-inflammatory properties by inhibiting the synthesis of cyclooxygenases (COX) enzymes with a reduction of PGs production [30]. These lipid mediators play an important role in bone repair. Moreover, they influence the regulation of inflammation, increase osteoblast proliferation and differentiation and enhance osteoclast activity and bone resorption [18]. Accumulating evidence suggests that the general therapeutic effects of MSCs in bone repair are due to their abilities to promote a regenerative microenvironment and not because of their capabilities to differentiate and incorporate into the host tissue. The beneficial effects of MSCs involve immunomodulatory effects, stimulation of angiogenesis and antiapoptotic effects, as well as recruitment of host stem/ progenitor cells into the site of bone repair [10,11]. However, to date, little is known about the effect of NSAIDSs on the therapeutic potential of mesenchymal stem cells used in stem cell-based tissue engineering approaches. Therefore, the main research objective of the present study was to evaluate the in vitro effects of therapeutic doses of non-selective NSAIDSs, ibuprofen and diclofenac on the properties of human mesenchymal stromal cells isolated from the dental pulp (DPSCs). Ibuprofen and diclofenac were chosen because of their popularity and long history of use as anti-inflammatory drugs and pain relievers in patients.
By previous studies [31,32,33], our analysis revealed that isolated cells from dental pulp were positive for the mesenchymal stem cell markers CD73, CD90 and CD105 and did not express markers typical for hematopoietic and endothelial cells, CD14, CD20, CD34 and CD45, which characterise mesenchymal stem cells according to Dominici et al. [34]. Ibuprofen or diclofenac pre-treatment did not cause any significant alterations in the expression of the mentioned markers during the therapeutic dosage range. Meanwhile, doses of ibuprofen used in the study of Salkin et al. (2022) were several times higher than therapeutic plasma concentration. In their research, high-dose ibuprofen (3mmol/l) significantly increased CD73 expression in DPSCs [35], which indicates dosage-dependent results.
Regarding cellular viability, there is conflicting evidence in the literature in the case of NSAIDS effect on stem cell viability. While doses of ibuprofen used in the study by Salkin et al. (2022) were 0.1mmol/l and 3mmol/l, DPSCs viability increased significantly in the ibuprofen-applied groups [35]. However, different results were revealed with therapeutic concentrations of ibuprofen pre-treatment (25µg/ml, 50µg/ml) on human bone marrow mesenchymal stem cells. In the study of Kulesza et al. (2022), the authors demonstrated that higher ibuprofen doses negatively affected MSCs' viability. However, the maximum decrease in cell viability was observed after ten days of treatment for 50µg/ml of ibuprofen (92% relative viability) [17]. The conclusion of Müller et al. study (2011) demonstrated that the effects of NSAIDSs on MSCs depend mainly on the concentrations used [36]. Ibuprofen and diclofenac concentrations chosen in our study did not differ from the maximum serum concentrations during standard pharmacotherapy. We found that NSAIDSs have unfavourable effects on DPSCs viability. Ibuprofen in low concentration (150µM) significantly suppressed the viability of DPSCs in the 72 hour interval. Similarly, diclofenac in high doses (3µM) significantly decreased cell viability in the 72 hour interval of DPSCs pre-treatment. Comparable results were observed in the study of Kudo et al. (2003), who demonstrated that diclofenac (10µM) induced the death of neural stem cells after 24-hour treatment [37]. These differences between studies may result from the NSAIDS concentrations used, different pre-treatment times, and the cell type and species used.
Over the past several years, cell-based therapies for bone regeneration have been extensively investigated [25]. A growing body of literature suggests that mesenchymal stromal cells may secrete factors that support angiogenesis at the site of injury, which presents an essential component of bone repair [38,39]. Angiogenesis is regulated by various growth factors, hormones, cytokines and low molecular-mass mediators [40]. A significant role plays vascular endothelial growth factor α (VEGFA) produced mainly by inflammatory cells and stromal cells to induce blood vessel growth. The various cellular functions of VEGF result from its ability to initiate a diverse, complex network of signalling pathways [41]. Indeed, in bone healing, VEGF not only increases endothelial cell differentiation and proliferation but also increases tube formation together with mobilisation and recruitment of endothelial progenitor cells [39]. Paracrine VEGF signalling is mediated by the tyrosine kinase receptors VEGFR1 and VEGFR2. All VEGF isoforms can bind to both receptors. However, VEGFR2 has strong tyrosine kinase activity and thus is the main receptor involved in cell signalling, including the activation of mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K)/Akt, Src and Rac signalling [42]. Another important pleiotropic cytokine involved in numerous complex biological processes in tissue regeneration, tumour growth and angiogenesis is hepatocyte growth factor (HGF). Binding and activating its c-met receptor, expressed in several cell types, triggers several signalling pathways, such as PI3K/Akt, MAPK and others, most of which are expected to VEGFA signalling [43]. Regarding angiogenesis, HGF stimulates endothelial cells directly through the c-Met receptor and indirectly by facilitating the expression of other angiogenic factors represented by VEGF [44].
To investigate the impact of ibuprofen and diclofenac on the secretory activity of DPSCs, we performed qRT-PCR analysis of selected gene expression. DPSCs are known to secrete numerous bioactive factors, and this activity is crucial for their immunomodulatory and pro-regenerative as angiogenic abilities. First, we evaluated the effect of low and high-dose ibuprofen and diclofenac on the mRNA expression level of VEGFA. The results indicated that neither ibuprofen nor diclofenac significantly changed the expression of the most important angiogenic factor, VEGFA. However, the analysis of the HGF transcript revealed the significant effect of NSAIDSs on DPSCs. While low-dose ibuprofen did not affect HGF expression, high-dose ibuprofen and both used diclofenac concentrations caused a significant increase in HGF transcript level in DPSCs after 72 hour intervals of pre-treatment. The effects of ibuprofen preconditioning have already been reported regarding MSCs isolated from bone marrow. Kulesza and colleagues (2022) evaluated the consequence of ibuprofen on MSCs secretome by Proteome Profiler and Luminex immunoassays. Ibuprofen (25µg/mL for 72 hours) significantly decreased the mean secretion of VEGF (by 20%) and HGF (by 31%) compared to secretion of control MSCs [17]. Similarly, reduced VEGFA expression was observed in osteoblast cells treated with 10μM doses of diclofenac and ibuprofen at 24h treatment, evaluated by RT-PCR and compared with the expression of untreated osteoblast [45]. These results indicate that NSAIDSs modulate the expression of angiogenic factors.
Previous studies have shown that NSAIDSs are implicated in the apoptosis and death of cells and tissues and can have anti-cancer effects. However, the mechanism of this effect is not well known in molecular and cellular terms [46]. The reduction of viability of cancer cells, as well as activation of caspase pathways, were confirmed in ibuprofen pre-treated cervical cancer cells [47]. A study by Akrami et al. (2014) evaluated the impact of ibuprofen on the expression of set genes involved in apoptosis. The results revealed that ibuprofen at 500µM downregulated transcription of the BCL2 gene in gastric cancer cells [48]. Another study revealed that neural stem cells treated with diclofenac (60µM) for 24 hours showed nuclear condensation, and western blot analysis reported that the activation of caspase 3 was increased by treatment with diclofenac in a concentration-dependent manner (10µM, 30µM, 60µM) [37].
Regarding the effects on genes involved in apoptosis, we found that NSAIDSs used in our study alter the transcription of selected genes, except for BAX, where no changes after pre-treatment were observed. Firstly, we evaluated the effect of low-dose ibuprofen, which significantly downregulated expression of caspase 3 in a 72 hour interval, while high-dose ibuprofen increased transcription of caspase 9 after 24 hours. Both used concentrations caused upregulation of the BCL2 gene in DPSCs in early intervals. BAK, CASP3, and CASP9 expressions were significantly increased in the diclofenac pre-treatment group compared to the untreated group. Low-dose diclofenac significantly increases BCL2 expression in a 24 hour interval. However, in the high-dose diclofenac group, this effect has not been reported. The difference between ibuprofen and diclofenac's effects on the induction of cell death may be explained by the selectivity of COX-2 inhibition. Although diclofenac is generally accepted as a traditional NSAIDS in the published literature, it was proven to have a higher selectivity for COX-2 than for COX-1, in contrast with most traditional NSAIDSs. The degree of COX-2 selectivity demonstrated for diclofenac can be compared to that of celecoxib, a selective COX-2 inhibitor [49,50,51]. Many experimental and clinical studies have suggested that COX-2 inhibitors may reduce cancer risk through induction of apoptosis [46,52]. The changes in HGF, BAK, CASP3, CASP9 and BCL2 expression, in combination with the reduced viability of NSAIDS-treated DPSCs, indicate a variety of effects on stem cell properties that might affect the therapeutic outcome. Unfavourable effects of both ibuprofen and diclofenac on viability and apoptosis-related genes in stem cells may help in designing better pharmacotherapy strategies and highlight the need for increased caution in use of NSAIDSs in postsurgical pain therapy after regenerative treatment with DPSCs.
4. Materials and Methods
4.1. Cell isolation
Cells were isolated from the dental pulp of extracted third molars from healthy donors (age 18-20) according to the Helsinki Declaration after the donor's informed consent. The teeth were removed due to orthodontic therapy and did not show any pathological alterations. Pulp tissues were washed thoroughly in PBS containing antibiotics and cut into 1-2mm2 pieces. Small pieces were placed in 60mm culture dishes in a random pattern. Drops of fetal bovine serum (FBS, Sigma Aldrich, Germany) were applied on the tissues sufficient to cover them entirely and maintained at 37°C in a humidified incubator with 5% CO2. After 2-hours incubation, explants were maintained in low glucose DMEM (Dulbecco' modified Eagle medium, Sigma Aldrich, Germany) enriched with 10% FBS, penicillin (100IU/ml; Sigma Aldrich, Germany) and streptomycin (100µg/ml; (Sigma Aldrich, Germany). The culture medium was changed every 2-3 days, and the cell outgrowth was monitored regularly with an M-795 inverted microscope (OPTIKA S.R.L., Italy). The outgrown cells at 70-80% confluence were detached using 0.25% Trypsin-EDTA solution and transferred to a T-75 flask. All experiments were performed between 4-6 passages.
4.2. hDPSCs Characterisation
To confirm phenotype of isolated cells, every hDPSCs population at the 3rd passage was identified by flow cytometry (MACS Quant Analyzer, Miltenyi Biotec, Germany) according to the phenotypic signature described by the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy (ISCT) using the MSC Phenotyping kit (Miltenyi Biotec, Germany) according to manufacturer's instructions. Cell viability was assessed with propidium iodide (PI, Miltenyi Biotec, Germany). The morphology of DPSCs was evaluated with microscopic observations using the M-795 inverted microscope (OPTIKA S.R.L-, Italy). For the experiment, the required passage was transferred to a cell culture plate in an amount of 0.5x106 cells. After pharmacological treatment, DPSCs were evaluated by flow cytometry, as described above. Medians with 25-75% percentiles were calculated using two experiments with cells from different donors (n=4).
4.3. Ibuprofen and Diclofenac Treatment
For all experiments presented in the study, an ibuprofen stock (Sigma Aldrich, Germany) at a concentration of 50 mg/ml (240mM) in ethanol and a diclofenac stock (Sigma Aldrich, Germany) at a concentration of 50 mg/ml (157mM) in methanol were used. The stock solutions were prepared freshly before each experimental set-up. According to literature data, the selected concentrations of ibuprofen were 150µM (low-dose) and 300µM (high-dose) of IBU, which refers to serum concentrations of single-dose ibuprofen use during pharmacotherapy [53,54]. Low-dose (1.5µM) and high-dose (3µM) diclofenac were administered to DPSCs based on the blood concentrations [49,55]. The duration of ibuprofen and diclofenac treatment was 24, 48 and 72 hours. The control group (CTRL) in all ibuprofen experiments were hDPSCs treated with equal amounts of solvent (ethanol) in the medium that was in the tested samples. The final solvent concentration in samples achieved 0,06% (v/v) in samples treated with 150µM of ibuprofen and 0,125% (v/v) in samples with 300µM of ibuprofen.
The control group (CTRL) in all diclofenac experiments were hDPSCs treated with equal amounts of solvent (methanol) in the medium that was in the tested samples. The final solvent concentration in samples achieved 0,0008% (v/v) in samples treated with 1.5µM of diclofenac and 0,0019% (v/v) in samples with 3.0µM of diclofenac.
4.4. Immunofluorescence staining
DPSCs (1x104 cells/well) were seeded on coverslips in 12-well plates and cultured as described above to evaluate morphological changes induced by different treatments. After seven days, cells were fixed with 4% paraformaldehyde and permeabilised with 0.2% Triton X/0.1% Tween/1xPBS (30min). The cells were then blocked with 5% goat serum (Sigma Aldrich, Germany) and incubated with rabbit vimentin antibody (1:100 dilution; cat. No D21H3 XP, Cell Signalling Technology, USA). Alexa Fluor 488-conjugated Goat anti-mouse IgG (H+L) (1:500 dilution; Cell Signalling Technology, USA) was used to incubate the cells at room temperature for 2 hours. DAPI (10 min) was used to stain cell nuclei. Fluorescent images were captured on a Ti-E microscope (Nikon instrument, USA) at 40X magnification.
4.5. MTT assay
To determine the cytotoxicity effect and proliferation rate of hDPSCs in culture after ibuprofen and diclofenac treatment, the tetrazolium salt (MTT) reduction test was carried out with a Cell proliferation kit (Sigma Aldrich, Germany). Cells were seeded in 96-well plates at 3x104 cells per well in a standard culture medium. Cells were incubated for 24 hours, and after the incubation period, were cells treated with ibuprofen and diclofenac as described above. All conditions were performed in quadruplicate. After incubation (24, 48 and 72 hours), 10µl of the MTT labelling reagent (final concentration 0.5mg/ml) was added to each well. Following 4 hours of incubation, 100µl of the Solubilization solution was added to each well with a 24 hour incubation period. The absorbance was measured at a wavelength of 570nm with a Varioskan LUX microplate reader (Thermo Fisher Scientific, USA).
4.6. Gene Expression
Total RNA was isolated from DPSCs using the Tri-Reagent® (Sigma Aldrich, Germany) with phenol-chloroform extraction and quantified using the Qubit RNA XR Assay Kit (Thermo Fisher Scientific, USA) according to the manufacturer's instruction. Then, 3000ng of nucleic acid was transcribed into cDNA using the High-Capacity cDNA Reverse KIT with RNAse inhibitor (Thermo Fisher Scientific, USA). Quantification of mRNA expression was performed using TaqMan Universal PCR Master Mix kit on QuantStudio 5 Real-Time PCR system (Thermo Fisher Scientific, USA) and probes for the following genes: vascular endothelial growth factor alpha (VEGFA; Hs00900055_m1), hepatocyte growth factor (HGF; Hs00300159_m1), BCL2 associated X (BAX; Hs00180269_m1), BCL2 antagonist 1 (BAK; Hs00832876_g1 ), caspase 9 (CASP9; Hs00962278_m1), caspase 3 (CASP3; Hs00234387_m1), B-cell lymphoma 2 (BCL2; Hs00608023_m1). The Pfaffl method was used to calculate the relative expression [56]. Results were normalised to the geometric mean of the two most suitable reference genes, beta-2-microglobulin (B2M; Hs99999907) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Hs99999905) [57]. Calculated normalised quantities were calibrated to appropriate control groups. Medians with 25-75% percentiles were calculated based on two experiments with cells from different donors (n=4) performed in triplicates.
4.7. Statistical analysis
Statistical analysis was performed using GraphPad Prism version 5.00 (GraphPad Software, San Diego, California, USA). Values in text are presented as mean ± SEM, all values in graphs are presented as medians with 25-75% percentiles and statistical analysis was performed comparing treated DPSCs with appropriate control. The Shapiro-Wilk test was used to determine data distribution. We used the Kruskal-Wallis test followed by the Dunn's test to compare more than two groups. The Student t-test determined the statistical significance between the two groups. P-value ≤ 0.05 was considered statistically significant.
5. Conclusions
The beneficial effects of MSCs in bone repair involve immunomodulatory effects, stimulation of angiogenesis and antiapoptotic effects, as well as recruitment of host stem/ progenitor cells into the site of bone repair. Our results showed that ibuprofen and diclofenac at concentrations that are achieved in the serum of patients during standard pharmacotherapy do not significantly affect the expression of mesenchymal stem cell markers. However, we demonstrated that both ibuprofen and diclofenac significantly decrease the viability of DPSCs, yet these significant decreases were approximately 2.5%, and therefore additional studies are needed. Moreover, our results revealed that used NSAIDSs do have an impact on the gene expression of DPSCs. The observed changes indicate that ibuprofen and diclofenac increase HGF transcript levels and, in addition, alter the expression of several genes involved in apoptosis (BAK, CASP3, CASP9, BCL2). These results suggest that the concomitant use of ibuprofen or diclofenac with stem cell-based tissue engineering approaches might impact the therapeutic outcome of the procedure by decreasing the viability and altering the expression of apoptosis-related genes in stem cells. Thus, this knowledge may help design better pharmacotherapy strategies and highlight the need for increased caution in their use after regenerative treatment with DPSCs. However, further more extensive studies are required for the verification of these hypotheses.
Funding
This article was created thanks to support under the Operational Program Integrated Infrastructure for the project National Infrastructure for supporting technology transfer in Slovakia II – NITT SK II, co-financed by the European Regional Development Fund.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets used and analysed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest
References
- Xue, N.; Ding, X.; Huang, R.; Jiang, R.; Huang, H.; Pan, X.; Min, W.; Chen, J.; Duan, J.-A.; Liu, P.; et al. Bone Tissue Engineering in the Treatment of Bone Defects. Pharmaceuticals 2022, 15, 879. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Dong, Z.; Wang, W.; Li, B.; Jin, Y. Dental Stem Cell and Dental Tissue Regeneration. Front. Med. 2019, 13, 152–159. [Google Scholar] [CrossRef]
- Janjua, O.S.; Qureshi, S.M.; Shaikh, M.S.; Alnazzawi, A.; Rodriguez-Lozano, F.J.; Pecci-Lloret, M.P.; Zafar, M.S. Autogenous Tooth Bone Grafts for Repair and Regeneration of Maxillofacial Defects: A Narrative Review. IJERPH 2022, 19, 3690. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.T.; Lee, S.; Tideman, H.; Stoelinga, P.J.W. Mandibular Reconstruction in Adults: A Review. International Journal of Oral and Maxillofacial Surgery 2008, 37, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.H. Autologous Bone Graft: Is It Still the Gold Standard? Injury 2021, 52, S18–S22. [Google Scholar] [CrossRef]
- Chamieh, F.; Collignon, A.-M.; Coyac, B.R.; Lesieur, J.; Ribes, S.; Sadoine, J.; Llorens, A.; Nicoletti, A.; Letourneur, D.; Colombier, M.-L.; et al. Accelerated Craniofacial Bone Regeneration through Dense Collagen Gel Scaffolds Seeded with Dental Pulp Stem Cells. Sci Rep 2016, 6, 38814. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nakamura-Yamada, S.; Konoki, R.; Baba, S. Promising Advances in Clinical Trials of Dental Tissue-Derived Cell-Based Regenerative Medicine. Stem Cell Res Ther 2020, 11, 175. [Google Scholar] [CrossRef]
- Hernández-Monjaraz, B.; Santiago-Osorio, E.; Ledesma-Martínez, E.; Alcauter-Zavala, A.; Mendoza-Núñez, V.M. Retrieval of a Periodontally Compromised Tooth by Allogeneic Grafting of Mesenchymal Stem Cells from Dental Pulp: A Case Report. J Int Med Res 2018, 46, 2983–2993. [Google Scholar] [CrossRef] [PubMed]
- Meesuk, L.; Suwanprateeb, J.; Thammarakcharoen, F.; Tantrawatpan, C.; Kheolamai, P.; Palang, I.; Tantikanlayaporn, D.; Manochantr, S. Osteogenic Differentiation and Proliferation Potentials of Human Bone Marrow and Umbilical Cord-Derived Mesenchymal Stem Cells on the 3D-Printed Hydroxyapatite Scaffolds. Sci Rep 2022, 12, 19509. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, G.D.; Carthew, J.; Lim, R.; Frith, J.E. Effect of the Microenvironment on Mesenchymal Stem Cell Paracrine Signaling: Opportunities to Engineer the Therapeutic Effect. Stem Cells and Development 2017, 26, 617–631. [Google Scholar] [CrossRef]
- Oryan, A.; Kamali, A.; Moshiri, A.; Baghaban Eslaminejad, M. Role of Mesenchymal Stem Cells in Bone Regenerative Medicine: What Is the Evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.M.; Pham, P.T.; Bach, T.Q.; Ngo, A.T.L.; Nguyen, Q.T.; Phan, T.T.K.; Nguyen, G.H.; Le, P.T.T.; Hoang, V.T.; Forsyth, N.R.; et al. Stem Cell-Based Therapy for Human Diseases. Sig Transduct Target Ther 2022, 7, 272. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.-D.; Hu, C.-H.; Liu, A.-Q.; Zheng, C.-X.; Xuan, K.; Jin, Y. Stem Cell-Based Bone Regeneration in Diseased Microenvironments: Challenges and Solutions. Biomaterials 2019, 196, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Musiał-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transplant 2019, 28, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Hersh, E.V.; Moore, P.A.; Grosser, T.; Polomano, R.C.; Farrar, J.T.; Saraghi, M.; Juska, S.A.; Mitchell, C.H.; Theken, K.N. Non-steroidal Anti-Inflammatory Drugs and Opioids in Postsurgical Dental Pain. J Dent Res 2020, 99, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Wongrakpanich, S.; Wongrakpanich, A.; Melhado, K.; Rangaswami, J. A Comprehensive Review of Non-Steroidal Anti-Inflammatory Drug Use in The Elderly. Aging Dis 2018, 9, 143–150. [Google Scholar] [CrossRef]
- Kulesza, A.; Zielniok, K.; Hawryluk, J.; Paczek, L.; Burdzinska, A. Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity. Biomolecules 2022, 12, 287. [Google Scholar] [CrossRef]
- Pountos, I.; Georgouli, T.; Calori, G.M.; Giannoudis, P.V. Do Non-steroidal Anti-Inflammatory Drugs Affect Bone Healing? A Critical Analysis. ScientificWorldJournal 2012, 2012, 606404. [Google Scholar] [CrossRef]
- Cottrell, J.; O'Connor, J.P. Effect of Non-Steroidal Anti-Inflammatory Drugs on Bone Healing. Pharmaceuticals 2010, 3, 1668–1693. [Google Scholar] [CrossRef]
- Moshiri, A.; Shahrezaee, M.; Shekarchi, B.; Oryan, A.; Azma, K. Three-Dimensional Porous Gelapin–Simvastatin Scaffolds Promoted Bone Defect Healing in Rabbits. Calcif Tissue Int 2015, 96, 552–564. [Google Scholar] [CrossRef]
- Aimetti, M.; Ferrarotti, F.; Gamba, M.N.; Giraudi, M.; Romano, F. Regenerative Treatment of Periodontal Intrabony Defects Using Autologous Dental Pulp Stem Cells: A 1-Year Follow-Up Case Series. Restorative Dentistry 2018, 38, 9. [Google Scholar] [CrossRef] [PubMed]
- d'Aquino, R.; De Rosa, A.; Lanza, V.; Tirino, V.; Laino, L.; Graziano, A.; Desiderio, V.; Laino, G.; Papaccio, G. Human Mandible Bone Defect Repair by the Grafting of Dental Pulp Stem/Progenitor Cells and Collagen Sponge Biocomplexes. European Cells and Materials 2009, 18, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Xuan, K.; Li, B.; Guo, H.; Sun, W.; Kou, X.; He, X.; Zhang, Y.; Sun, J.; Liu, A.; Liao, L.; et al. Deciduous Autologous Tooth Stem Cells Regenerate Dental Pulp after Implantation into Injured Teeth. Sci. Transl. Med. 2018, 10, eaaf3227. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.-D.; Hu, C.-H.; Zheng, C.-X.; Shuai, Y.; He, X.-N.; Gao, P.-P.; Zhao, P.; Li, M.; Zhang, X.-Y.; He, T.; et al. Recipient Glycemic Micro-Environments Govern Therapeutic Effects of Mesenchymal Stem Cell Infusion on Osteopenia. Theranostics 2017, 7, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Chen, J.; Liu, S.; Jin, Y. Stem Cell-Based Bone and Dental Regeneration: A View of Microenvironmental Modulation. Int J Oral Sci 2019, 11, 23. [Google Scholar] [CrossRef]
- Hadjicharalambous, C.; Alpantaki, K.; Chatzinikolaidou, M. Effects of NSAIDSs on Pre-osteoblast Viability and Osteogenic Differentiation. Exp Ther Med 2021, 22, 740. [Google Scholar] [CrossRef]
- Bailey, E.; Worthington, H.V.; van Wijk, A.; Yates, J.M.; Coulthard, P.; Afzal, Z. Ibuprofen and/or Paracetamol (Acetaminophen) for Pain Relief after Surgical Removal of Lower Wisdom Teeth. Cochrane Database of Systematic Reviews 2013. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.K.-S.; Lirk, P.; Seymour, R.A.; Jenkins, B.J. The Efficacy of Preemptive Analgesia for Acute Post-operative Pain Management: A Meta-Analysis: Anesthesia & Analgesia 2005, 100, 757–773. [CrossRef]
- Ji, R.-R.; Chamessian, A.; Zhang, Y.-Q. Pain Regulation by Non-Neuronal Cells and Inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Nagi, R.; Yashoda Devi, B.K.; Rakesh, N.; Reddy, S.S.; Patil, D.J. Clinical Implications of Prescribing Non-steroidal Anti-Inflammatory Drugs in Oral Health Care—a Review. Oral Surgery, Oral Medicine, Oral Pathology and Oral Radiology 2015, 119, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Rajan, T.S.; Gatta, V.; D’Aurora, M.; Merciaro, I.; Marchisio, M.; Muttini, A.; Caputi, S.; Bramanti, P.; Mazzon, E.; et al. Stemness Maintenance Properties in Human Oral Stem Cells after Long-Term Passage. Stem Cells International 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhong, Q.; Yang, T.; Qi, Y.; Fu, M.; Yang, X.; Qiao, L.; Ling, Q.; Liu, S.; Zhao, Y. Comparative Characterization of SHED and DPSCs during Extended Cultivation In�vitro. Molecular Medicine Reports 2018. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem Cell Properties of Human Dental Pulp Stem Cells. J Dent Res 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Salkın, H.; Basaran, K.E. Effects of Non-Steroidal Anti-Inflammatory Drug (Ibuprofen) in Low and High Dose on Stemness and Biological Characteristics of Human Dental Pulp-Derived Mesenchymal Stem Cells. Connective Tissue Research 2022, 0, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Raabe, O.; Addicks, K.; Wenisch, S.; Arnhold, S. Effects of Non-Steroidal Anti-Inflammatory Drugs on Proliferation, Differentiation and Migration in Equine Mesenchymal Stem Cells. Cell. Biol. Int. 2011, 35, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Kudo, C.; Kori, M.; Matsuzaki, K.; Yamai, K.; Nakajima, A.; Shibuya, A.; Niwa, H.; Kamisaki, Y.; Wada, K. Diclofenac Inhibits Proliferation and Differentiation of Neural Stem Cells. Biochemical Pharmacology 2003, 66, 289–295. [Google Scholar] [CrossRef] [PubMed]
- De Luca, A.; Gallo, M.; Aldinucci, D.; Ribatti, D.; Lamura, L.; D'Alessio, A.; De Filippi, R.; Pinto, A.; Normanno, N. Role of the EGFR Ligand/Receptor System in the Secretion of Angiogenic Factors in Mesenchymal Stem Cells. J. Cell. Physiol. 2011, 226, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Hankenson, K.D.; Dishowitz, M.; Gray, C.; Schenker, M. Angiogenesis in Bone Regeneration. Injury 2011, 42, 556–561. [Google Scholar] [CrossRef]
- Tao, H.; Han, Z.; Han, Z.C.; Li, Z. Proangiogenic Features of Mesenchymal Stem Cells and Their Therapeutic Applications. Stem Cells International 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Zhang, H.; Hou, J.; Wan, L.; Cheng, W.; Wang, X.; Dong, D.; Chen, C.; Xia, J.; Guo, J.; et al. VEGF Secreted by Mesenchymal Stem Cells Mediates the Differentiation of Endothelial Progenitor Cells into Endothelial Cells via Paracrine Mechanisms. Molecular Medicine Reports 2017. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, A.D.; Olsen, B.R. How Vascular Endothelial Growth Factor-A (VEGF) Regulates Differentiation of Mesenchymal Stem Cells. Journal of Histochemistry & Cytochemistry 2014, 62, 103–108. [Google Scholar]
- Sulpice, E.; Ding, S.; Muscatelli-Groux, B.; Bergé, M.; Han, Z.C.; Plouet, J.; Tobelem, G.; Merkulova-Rainon, T. Cross-Talk between the VEGF-A and HGF Signalling Pathways in Endothelial Cells. Biology of the Cell 2009, 101, 525–539. [Google Scholar] [CrossRef]
- Matsumura, A.; Kubota, T.; Taiyoh, H.; Fujiwara, H.; Okamoto, K.; Ichikawa, D.; Shiozaki, A.; Komatsu, S.; Nakanishi, M.; Kuriu, Y.; et al. HGF Regulates VEGF Expression via the C-Met Receptor Downstream Pathways, PI3K/Akt, MAPK and STAT3, in CT26 Murine Cells. International Journal of Oncology 2013, 42, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Moreno, F.J.; Costela-Ruiz, V.J.; Melguizo-Rodríguez, L.; Illescas-Montes, R.; García-Martínez, O.; Ruiz, C.; Ramos-Torrecillas, J. Inhibition of VEGF Gene Expression in Osteoblast Cells by Different NSAIDSs. Archives of Oral Biology 2018, 92, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Tan, T.; Liu, P. Regulation of Autophagy by Non-Steroidal Anti-Inflammatory Drugs in Cancer. CMAR 2020, Volume 12, 4595–4604. [Google Scholar] [CrossRef]
- Ghadiri, E.; Ahmadi, R.; Rostami, E. The Effects of Ibuprofen Cytoxic Dose on Caspase-3, -8 and -9 Activity Level in Cervical Cancer (Hela) Cells. JBUMS 2019, 21. [Google Scholar] [CrossRef]
- Akrami, H.; Aminzadeh, S.; Fallahi, H. Inhibitory Effect of Ibuprofen on Tumor Survival and Angiogenesis in Gastric Cancer Cell. Tumor Biol. 2015, 36, 3237–3243. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Bosch, B.; Brune, K.; Patrignani, P.; Young, C. Advances in NSAIDS Development: Evolution of Diclofenac Products Using Pharmaceutical Technology. Drugs 2015, 75, 859–877. [Google Scholar] [CrossRef] [PubMed]
- Gan, TJ. Diclofenac: An Update on Its Mechanism of Action and Safety Profile. Current medical research and opinion 2010, 26. [Google Scholar] [CrossRef] [PubMed]
- Grosser, T.; S, F.; Ga, F. Biological Basis for the Cardiovascular Consequences of COX-2 Inhibition: Therapeutic Challenges and Opportunities. The Journal of clinical investigation 2006, 116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, F.; Shang, L. Advances in Antitumor Effects of NSAIDSs. Cancer Manag Res 2018, 10, 4631–4640. [Google Scholar] [CrossRef]
- Leidgens, V.; Seliger, C.; Jachnik, B.; Welz, T.; Leukel, P.; Vollmann-Zwerenz, A.; Bogdahn, U.; Kreutz, M.; Grauer, O.M.; Hau, P. Ibuprofen and Diclofenac Restrict Migration and Proliferation of Human Glioma Cells by Distinct Molecular Mechanisms. PLoS ONE 2015, 10, e0140613. [Google Scholar] [CrossRef] [PubMed]
- Weiser, T.; Schepers, C.; Mück, T.; Lange, R. Pharmacokinetic Properties of Ibuprofen (IBU) From the Fixed-Dose Combination IBU/Caffeine (400/100 Mg; FDC) in Comparison With 400 Mg IBU as Acid or Lysinate Under Fasted and Fed Conditions—Data From 2 Single-Center, Single-Dose, Randomized Crossover Studies in Healthy Volunteers. Clinical Pharmacology in Drug Development 2019, 8, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Lissy, M.; Stiff, D.D.; Kowalski, M.M.; Moore, K.A. Single-Dose Pharmacokinetic Study of Rapidly Dispersing Diclofenac Potassium Formulations in Healthy Volunteers. Current Medical Research and Opinion 2009, 25, 2423–2428. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
Figure 1.
Characterisation of DPSCs. (a) Primary cell colonies growing from dental pulp were observed 20 days after initial seeding (arrows marked spindle-shaped cells). (b) DPSCs expressed characteristic mesenchymal stem cell markers (CD73, CD90, CD105), while the non-MSC markers (CD14, CD20, CD34 and CD45) were not detected. (c) Representative overlay histograms of DPSCs show the control populations (grey) and the specifically stained cells (red).
Figure 1.
Characterisation of DPSCs. (a) Primary cell colonies growing from dental pulp were observed 20 days after initial seeding (arrows marked spindle-shaped cells). (b) DPSCs expressed characteristic mesenchymal stem cell markers (CD73, CD90, CD105), while the non-MSC markers (CD14, CD20, CD34 and CD45) were not detected. (c) Representative overlay histograms of DPSCs show the control populations (grey) and the specifically stained cells (red).
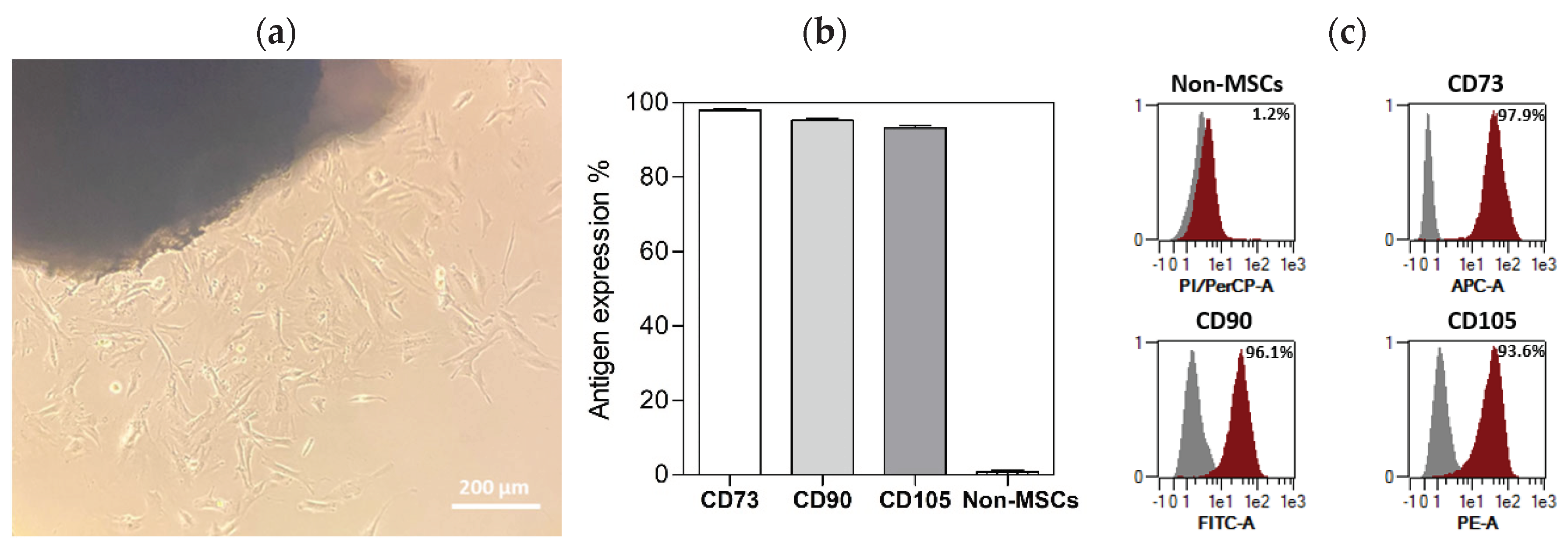
Figure 2.
Fluorescence microscopy of DPSCs cultured with IBU and DIC after 48 hours and stained for vimentin (green). Cell nuclei were counterstained with DAPI (blue). (a) Morphology of DPSCs at light microscopic levels and representative overlay histograms of DPSCs treated with IBU or DIC show the control populations (grey) and the specifically stained cells (red) (b).
Figure 2.
Fluorescence microscopy of DPSCs cultured with IBU and DIC after 48 hours and stained for vimentin (green). Cell nuclei were counterstained with DAPI (blue). (a) Morphology of DPSCs at light microscopic levels and representative overlay histograms of DPSCs treated with IBU or DIC show the control populations (grey) and the specifically stained cells (red) (b).
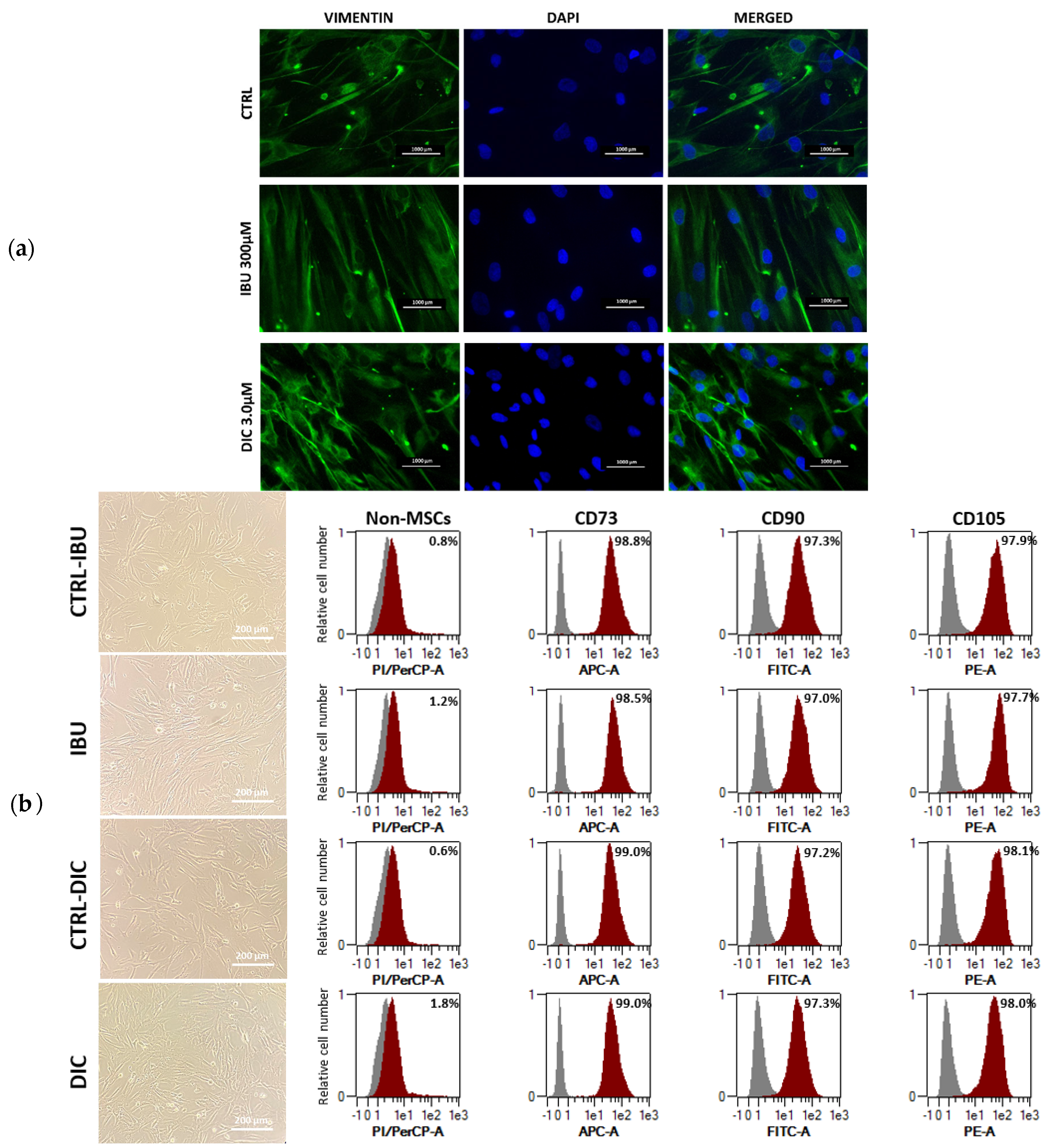
Figure 2.
Viability and morphology of DPSCs. Cell viability of DPSCs treated with (a) IBU and (b) DIC after 24, 48 and 72 hours. Cell viability was assessed by flow cytometry. The effect of ibuprofen (c) and diclofenac (d) on the cytotoxicity of hDPSCs was evaluated with MTT assay. The proliferation rate of hDPSCs after NSAIDSs treatment (e, f) was assessed by MTT assay. Data are presented as means ± SEM (n=4). The Mann-Whitney test performed statistical analysis. P* ≤ 0.05 compared to the CTRL group.
Figure 2.
Viability and morphology of DPSCs. Cell viability of DPSCs treated with (a) IBU and (b) DIC after 24, 48 and 72 hours. Cell viability was assessed by flow cytometry. The effect of ibuprofen (c) and diclofenac (d) on the cytotoxicity of hDPSCs was evaluated with MTT assay. The proliferation rate of hDPSCs after NSAIDSs treatment (e, f) was assessed by MTT assay. Data are presented as means ± SEM (n=4). The Mann-Whitney test performed statistical analysis. P* ≤ 0.05 compared to the CTRL group.
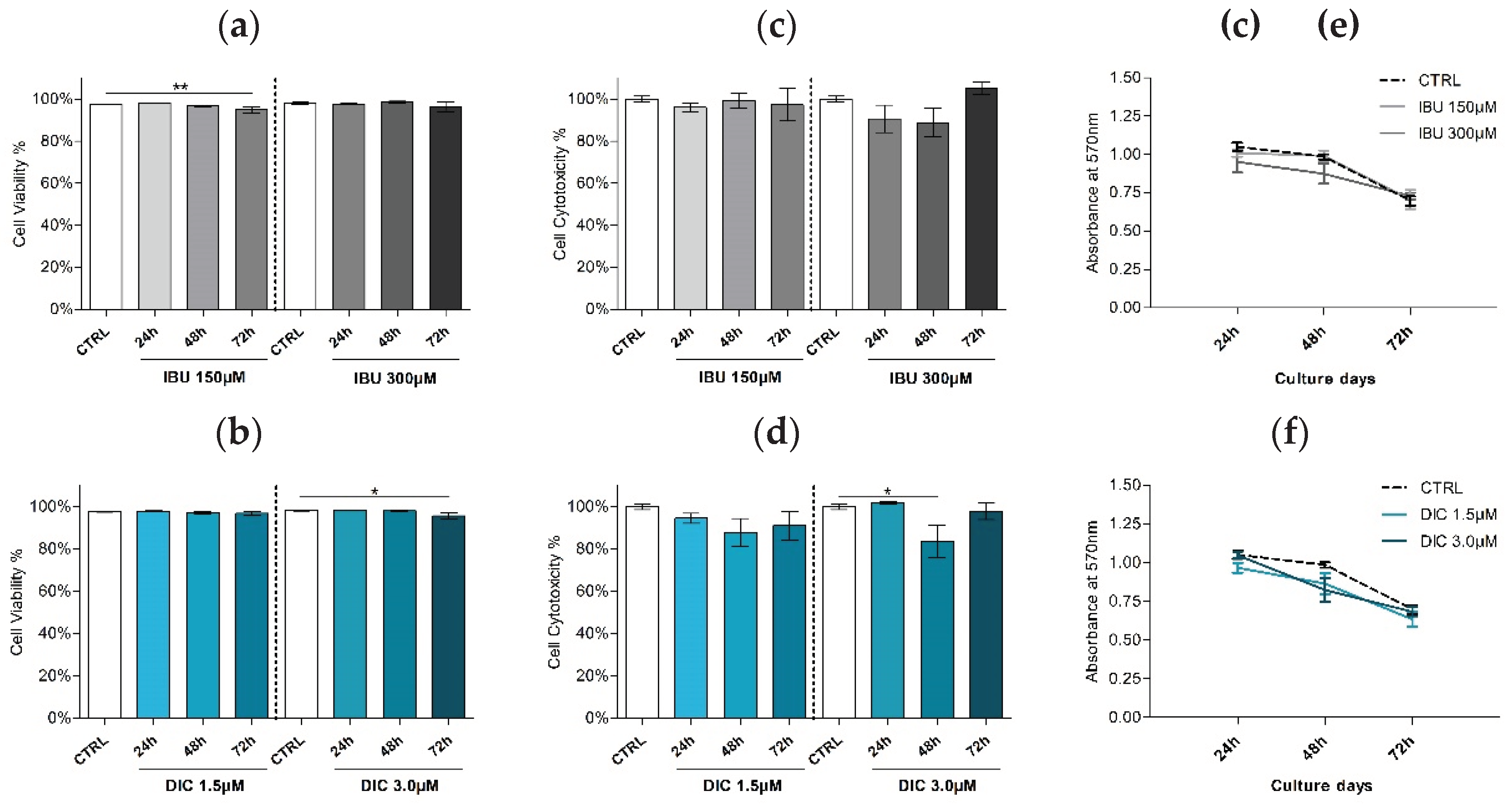
Figure 3.
Expression of angiogenesis-associated genes. Effects of different concentrations of IBU and DIC on (a) VEGFA and (b) HGF mRNA expression in DPSCs at 24, 48 and 72 hours. Target gene expressions are depicted relative to the control groups (DPSCs treated with equal solvents, ethanol in the medium for ibuprofen or methanol in the medium for diclofenac). Data are presented as median values with error bars of 25-75% percentiles of two experiments from different donors (n=4) performed in triplicate (p* ≤ 0.05 compared to the CTRL group).
Figure 3.
Expression of angiogenesis-associated genes. Effects of different concentrations of IBU and DIC on (a) VEGFA and (b) HGF mRNA expression in DPSCs at 24, 48 and 72 hours. Target gene expressions are depicted relative to the control groups (DPSCs treated with equal solvents, ethanol in the medium for ibuprofen or methanol in the medium for diclofenac). Data are presented as median values with error bars of 25-75% percentiles of two experiments from different donors (n=4) performed in triplicate (p* ≤ 0.05 compared to the CTRL group).
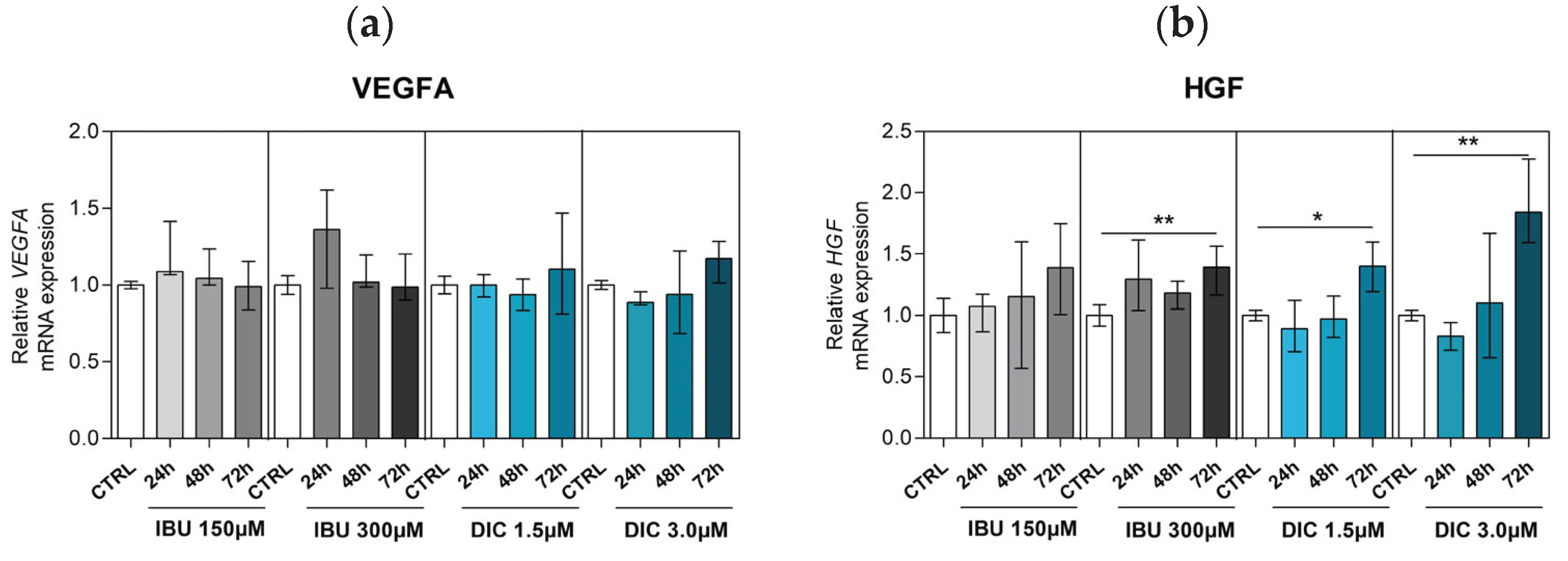
Figure 4.
Expression of apoptosis-associated genes. Effects of different concentrations of IBU and DIC on (a) BAX, (b) BAK, (c) CASP9, (d) CASP3 and (e) BCL2 mRNA expression in DPSCs at 24, 48 and 72 hours. Target gene expressions are depicted relative to the control groups (DPSCs treated with equal solvents, ethanol in the medium for ibuprofen or methanol in the medium for diclofenac). Data are presented as median values with error bars of 25-75% percentiles of two experiments from different donors (n=4) performed in triplicate (p* ≤ 0.05 compared to the CTRL group). Marker expression as demonstrated by flow cytometry (n=4).
Figure 4.
Expression of apoptosis-associated genes. Effects of different concentrations of IBU and DIC on (a) BAX, (b) BAK, (c) CASP9, (d) CASP3 and (e) BCL2 mRNA expression in DPSCs at 24, 48 and 72 hours. Target gene expressions are depicted relative to the control groups (DPSCs treated with equal solvents, ethanol in the medium for ibuprofen or methanol in the medium for diclofenac). Data are presented as median values with error bars of 25-75% percentiles of two experiments from different donors (n=4) performed in triplicate (p* ≤ 0.05 compared to the CTRL group). Marker expression as demonstrated by flow cytometry (n=4).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



