
Submitted:
01 April 2024
Posted:
02 April 2024
You are already at the latest version
Abstract
Histone deacetylases inhibitors (HDACi) have shown promising therapeutic outcomes in haematological malignancies such as leu-kaemia, multiple myeloma and lymphoma, with dis-appointing results in solid tumours when used as monotherapy. As a result, combination therapies either with radiation or other deoxyribonucleic acid (DNA) damaging agents have been suggested as ideal strategy to improve their efficacy in solid tumours. Numerous in vitro and in vivo studies have demonstrated that HDACi can sensitise malignant cells to both electromagnetic and particle types of radiation by inhibiting DNA damage repair. Although the radiosensitising ability of HDACi has been reported as early as 1990s, the mechanisms of radiosensitisation are yet to be fully understood. This review brings forth the various protocols used to sequence the administration of radiation and HDACi treatments in the different studies. The possible contribution of these various protocols on the ambiguity that surrounds the mechanisms of radiosensitisation is also highlighted.
Keywords:
histone deacetylase inhibitors
; radiosensitisation
; heterochromatin
; double strand break
; DNA repair
1. Introduction
Histone deacetylase inhibitors (HDACi) have attracted a lot of interest as potential radiosensitisers that have selective effects on malignant cells with little effect on healthy cells [1,2]. The radiosensitising capabilities of HDACi in combination with photon irradiation has been well studied [3,4,5,6,7,8,9,10,11,12] , while studies in combination with proton and carbon ion irradiation remain limited [13,14,15,16,17,18]. The exact mechanisms that underlie the radiosensitisation potential of these drugs for both photons and particle types of radiation also remain a matter of research. The effect of HDACi on the DNA damage repair (DDR) pathways as well as the effect they have on chromatin structure have been suggested as the main mechanisms [19,20,21,22,23,24]. However, the temporal sequence of HDACi in combination with radiation, as well as the optimal duration of HDACi treatment during a radiotherapy course remains to be an elusive subject. As a result, different administration times and sequences have been used in in vitro studies so far, which makes interpretation of results complex. Data on HDACi and radiation treatment from clinical studies and clinical trials is very limited [25,26,27,28]. In this review, the mechanisms that underlie cellular effects of HDACi as well mechanisms of radiosensitisation by HDACi are briefly reviewed, followed by the review of temporal sequences of radiation and duration of incubation with HDACi in combination with radiation that has been used in different in vitro studies using different types of radiation.
2. Epigenetic Modulation by HDACs and HDAC Inhibitors
During the process of malignant transformation in cells, genes that encode for histone acetyl transferases (HATs) can be amplified, translocated, or mutated leading to the inactivation of HATs. Consequently, histone deacetylase (HDACs) become overactive in malignant cells, resulting in the accumulation of deacetylated proteins. Overexpression of HDACs has been found in multiple human tumours such as lymphoma, prostate, gastric, leukaemia, colon and breast [3,4,6,7,8,9,10,11,12,29]. HDACs have thus been recognised as promising targets to modify and reverse the aberrant epigenetic control in cancer cells [7]. To this effect, (HDACi) have emerged as anti-cancer agents aimed at reversing the aberrant histone modification control in tumours [20,30,31] Inhibition of HDAC activity results in accumulation of acetylated proteins leading to cellular effects such as cell cycle arrest, differentiation, altered gene expression, inhibition of angiogenesis and metastasis and apoptosis, in a cell-type dependent manner, Figure 1 [30,32,33].
Mechanism of action of HDACi has been linked to the structure and class of HDACi. In brief, HDACi are classified into several classes according to either chemical structure or sequence homology to yeast. Chemical structure classification categorises HDCAi into benzamides (e.g., chidamide, entinostat), hydroxamic acids (e.g., vorinostat (SAHA), belinostat, panabinostat, CUDC-101), cyclic tetrapeptides (e.g., romidepsin) and aliphatic acids (e.g., butyrate, valproic acid) [6,35,36]. Of these classes, hydroxamic acids is the main class that has been used and continues to be used in most studies [37]. Hydroxamic acids are preferred as they inhibit a broad range of HDACs (HDACs1-11), and they can cause cellular effects at low (nM) concentrations [36]. Sequence homology to yeast classification groups HDACi into four classes, Class I (HDACs 1,2,3,8), Class II (HDACs 4,5,6,7,9,10 ), Class III (Sirtuins1-7 ) and Class IV (HDAC 11) [38].
Earlier studies proposed histone hyper-acetylation and subsequent alterations in gene expression to be the main mechanism through which HDACi mediate their antiproliferative effect. Histone acetylation was reported to increase at 6 hours post treatment, reaching a maximum between 24 and 48 hours after treatment with class 1 HDACi MS-275 in prostate carcinoma (DU145) and glioma (U251) cell lines [3]. However, this hypothesis could not explain the high specificity of HDACi for tumour cells. Subsequently, hyper-acetylation of non-chromatin and non-histone proteins involved in cell death, proliferation, cell migration, inflammation, angiogenesis, cell cycle control and DNA repair was acknowledged [13,32,33]. HDACi-induced cell death is mediated by several mechanisms including apoptosis, autophagy, necrosis and cell cycle arrest in a cell-type dependent manner [30,32,33].
2.1. HDACi Induced Apoptosis and Autophagy
Induction of apoptosis was initially recognized as the predominant mode of HDACi induced cell death [39,40,41]. A number of studies reported HDACi induced apoptosis through both the intrinsic and extrinsic pathways [20,30,31,32,39,42,43,44]. In particular, the intrinsic (mitochondria-related) apoptotic pathway has been supported by many studies as the main pathway that is activated by HDACi [25,26,33,35,36,37,38,39]. In brief, HDACi increases production of reactive oxygen species (ROS) which leads to loss of membrane potential. Loss of membrane potential enables cytochrome c to be released from the mitochondria to the cytoplasm, leading to activation of caspase 9 and initiation of apoptosis [45]. Activation of both p53-dependent and independent apoptotic pathways post HDACi treatment have been reported, which would be beneficial for the treatment of p53 mutant tumours [2]. The role of the extrinsic apoptotic pathway and caspase-independent pathways in HDACi induced apoptosis have long been acknowledged but remain poorly understood [31,39]. However, a possible link between autophagy and the extrinsic apoptotic pathway has been reported [46]. It is possible that since autophagic cell death in cancer is also not well understood, some autophagic cell death may have previously been attributed to caspase-independent apoptotic death [47].
The role of autophagy in cancer is complex and remains controversial [48]. Traditionally, autophagy was regarded as a cell death mechanism which eliminates damaged organelles, proteins, macromolecules and breakdown products from cells, thereby suppressing tumour progression. Hence autophagy was referred to as cell death type II [2]. Later evidence suggested that autophagy can also act as cell survival mechanism to promote tumour growth [49]. A conceivable explanation of the dynamic nature of autophagy in cancer elucidated that the role of autophagy depends on the stage, type of tumour, and genetic pre-disposition of the tumour [2,49,50]. In the early tumour stages, autophagy plays a protective role by preventing accumulation of damaged organelles and macromolecules. In the late tumour stages, autophagy assumes the role of a survival mechanism by recycling degraded metabolites and counteracting the effect of chemotherapy treatment as well as oxygen and nutrient deprivation in hypoxic tumour areas, maintaining tumour growth [2,50]. It is appealing to associate the role of autophagy in late-stage tumours to the role of mammalian target of rapamycin (mTOR). mTOR plays a crucial role in metabolism and regulates autophagy by deactivating human autophagy initiation kinase ULK1, a component of upstream autophagic signalling pathway [2].
It comes as no surprise that HDACi induced autophagy is also a highly debated topic. Proposed working mechanisms includes acetylation and upregulation of numerous autophagy related proteins such as p53, p21, ATG3, ATG 7, ULK1 and Nuclear Factor kappa B (NF-ĸB) genes. Inhibition of mTOR, transcription of FOX 01, inactivation of apoptosomes, upregulation of death-associated protein kinase (DAPK) and accumulation of reactive oxygen species (ROS) have also been suggested as possible mechanisms [2]. Of interest is the reported paradoxical relationship between HDACi induced autophagy and apoptosis. Several studies have reported a molecular shift between autophagy and apoptosis following HDACi treatment [2,38,47]. In this regard, tumour suppressor gene p53 was identified as the molecular switch that either promotes or inhibit autophagy in response to HDACi treatment in a cell line dependant manner [2,47]. Captivatingly, Garufi et al. reported autophagy as the main route in HDACi-mediated reduction of mutant p53 in human colon cancer (RKO), breast cancer (SKBR3), lung cancer (H1299), human colon cancer (HCT116) and pancreatic cell line [2]. As an example, HDACi induced autophagy was diminished in chronic leukocytic leukemia cells following treatment with HDACis mocetinostat and MGCD0103. On the other hand, HDACi mocetinostat induced autophagy in breast MCF-7 cell line [2,51]. In addition, HDACi-induced autophagy was reported in uterine sarcoma cells ESS-1, while apoptosis was reported in MES-SA cells following a 24 hour treatment with HDCAi SAHA [52]. To further support the molecular shift between autophagy and apoptosis, autophagy was also reported in apoptosis resistant uterine sarcoma and hepatocellular carcinoma cells after treatment with HDACi SAHA and OSU-HDAC42 [53,54]. Clearly, the molecular shift between autophagy and apoptosis depends on the HDACi, cell line, and possibly p53 status. This could in part explain the diverse results observed in different studies as well as the uncertainty that surrounds the mechanisms of HDACi-induced cell death. In accordance with this review, if the mechanisms remain to be unclear, then determining the optimal mechanistic sequence proves to be a challenge.
2.2. HDACi-Induced Upregulation of p21 and Cell Cycle Arrest
Cell cycle arrest is a prerequisite for the induction of programmed cell death [41]. HDACi induced apoptosis has been associated with upregulation of cyclin-dependent kinase (CDK) inhibitor p21 and cell cycle arrest [2]. Transcriptional re-activation of p53 and subsequent upregulation of p21 by HDACi has been reported in different cell lines [2,31,55]. Activation of p53 induces expression of p21 to induce cell cycle arrest mainly in the gap 1 (G1) phase. An earlier report by Richon et al. alluded that HDACi are gene specific after having observed only upregulation of p21 and no alteration in the expression of neither p27, also a CDK inhibitor, nor γ-actin genes [56]. Cell cycle arrest in G2/M phase of the cell cycle has also been reported and is accomplished by downregulating the expression of cyclin A by HDACi [30,57,58,59].
2.3. HDACi-Induced Inhibition of Angiogenesis
Studies on the effect of HDACi on angiogenesis remain limited. Inhibition of angiogenesis by HDACi was reported in nucleus polposus cells of interverbal discs, endothelial progenitor cells, Human Embryonic Kidney (HEK) 293 and epithelial fibrosarcoma (HT1080) cells [60,61,62,63]. Inhibition of angiogenesis was evidenced by attenuation of vascular endothelial growth factor (VEGF), hyper-acetylation of hypoxia inducible factor 1 (HIF-1α) and degradation of hypoxia-induced transcription factor [60,61,64]. Altered expression of pro- and anti-angiogenic genes following HDACi treatment has also been reported [63,65].
3. Radiosensitisation by HDAC Inhibitors
Evidence from pre-clinical studies has revealed that the combination of radiation and HDACi results in increased cell kill in a number of cell lines including lung, melanoma, prostate, glioma, colon, non-small cell lung cancer (NSCLC), osteosarcoma and breast to name a few [3,4,5,6,7,8,9,10,11,12,14,15,17,18,66]. When used as monotherapy, HDACi have shown promising therapeutic outcomes in haematological malignancies such as leukaemia, multiple myeloma and lymphoma, with disappointing results in solid tumours [20,30,36,67,68]. The molecular basis for the poor clinical outcomes in solid tumours are still unclear, but are thought to be due to the short drug half-life of HDACi which leads to poor drug distribution, poor HDAC isoform selectivity, and poor patient selection [13,38]. HDACi induced radiosensitisation is mainly attributed to their role in DNA damage response (DDR) and their effect on chromatin structure [12]. As a result, combination therapies either with radiation or other DNA damaging agents have been suggested as ideal strategy to improve their efficacy in solid tumours [5,14,36].
3.1. DNA DSB Induction and DNA Damage Repair (DDR)
Following the induction of DNA double strand breaks by radiation pathways that sense, respond and repair the damage are activated [69]. DNA double strand breaks are repaired using two basic mechanisms, homologous recombination (HR) or non-homologous end joining (NHEJ). During the initial stages of both HR and NHEJ, ATM promotes the processing of the broken DNA ends by the MRE11/NBS/Rad50 (MRN) complex to resects the broken ends into 3’ DNA single strand tails [70]. The choice of repair pathway is dictated in part by the presence or absence of p53 binding protein 1 (53BP1). In the presence of 53BP1, HR is inhibited and NHEJ is initiated. During NHEJ, Ku70 and Ku80 heterodimer binds the DNA ends and blocks resection of the 5’ end. Ku70/80 recruits DNA PK’s to the broken. In the final steps, DNA PK’ complex recruits ligase complex, X-ray repair complementing defective in Chinese Hamster 4 (XRCC4), XRCC4 like factor-DNA ligase 4(XLF-LIGIV), and polynucleotide kinase (PNK) to perform ligation of broken ends [21,71]. NHEJ is an error-prone mechanism which active throughout the cell cycle but mainly in the G1 phase [21,72].
During HR, processing of the damaged DNA ends is carried by Mre11-Rad50-Nbs1(MRN) complex and CTIp followed by RPA coating to form a nucleoprotein filament to which the HR protein can bind HR proteins, breast cancer tumour suppressor (BRCA1), RAD51 and RAD52 are the recruited to the nucleofilament [69]. RAD51 is key a protein in HR that facilitates strand exchange with the complementary undamaged DNA strand to form the Holiday junction. Resolution of the Holiday junction into two DNA duplexes is carried out by MMS4 and MUS81 [71]. HR requires the presence of an undamaged DNA strand (sister chromatid or chromosomes) to use as a template for repair. Sister chromatids are only available during the late S-and G2 phases after DNA replication, thus HR is active during these phases. The use of a DNA template for repair facilitates accurate repair which makes HR an error-free pathway [71,72]. HDACi have been observed to repress DNA repair proteins such as MRE11/Rad50/NBS1 (MRN) complex and Rad51 involved in HR and ku70, ku80, DNA-PK’s involved in NHEJ [12,19,22,73].
3.2. Role of HDACs and HDACi in the DNA Damage Response (DDR)
A growing body of evidence indicates that HDACs play an important role in the DDR [74,75]. During the earliest steps of the DDR, ataxia-telengiectasia mutated (ATM), a sensor of DNA damage, is autophosphorylated and initiates the DNA damage signalling cascade by recognising the DSB and phosphorylating several proteins that regulate the cell cycle as well as DNA damage repair. ATM phosphorylates tumour suppressor p53, which in turn activates cyclin-depended kinase (CDK) inhibitors p21 and p27. p21 binds the cyclin-dependent kinase inhibitors CDK4/6-cyclin D complex leading to its deactivation [39]. Deactivation of CDK4/6-cyclin D complex keeps the Rb gene in a hypo-phosphorylated state and represses the function of the E2F1 transcription factor, resulting in induction and maintenance of G1/S cell cycle arrest [39,40]. Repression of E2F1 by pRb is mediated by SIN3-HDAC corepressor complex (HDAC1,2,3, RBP1) as well as directly by deacetylation of p300 and CBP residues of the E2F1 transcription factor [41]. The role of HDACs in the DDR is illustrated in Figure 2.
Evidence suggests that HDACs play a role in regulating ATM. HDAC1 was observed to interact with ATM particularly after exposure to gamma-radiation in fibroblast cells [75]. In support of this observation, reduced activation of ATM after treatment with HDACi SAHA was reported in breast (MCF-7, T-47D), melanoma (SK-MEL-28), human osteosarcoma (Saos-2) and A549 cell lines [75]. This observation was further corroborated by reports of accumulation of HDAC1 and 2 at the damaged sites within 5 minutes of DNA damage induction and dissociation at 30 minutes after radiation treatment [74]. The rapid accumulation and dispersal of HDAC1 and 2 was associated with rapid deacetylation of Histone3 lysine 56 (H3K56) and Histone4 lysine 16 (H4K16) which favours non-homologous end-joining (NHEJ), followed by histone acetylations that favours homologous repair (HR) [74]. The authors also observed that the acetylation levels of H3K56 were reduced upon induction of DNA damage without treatment with HDACi [74]. HDAC 1 and 2 bind CDK inhibitors p21 and p27 reducing their activity resulting in cell cycle progression from G1 to S-phase [39,40,42]. Inhibition of HDAC1 and HDAC2 therefore restores the activity of p21 leading to G1 cell cycle arrest [40]. Depletion of HDAC1 was also reported to partially contribute to G2/M arrest cell cycle arrest [40,43] HDAC4 was reported to co-localise with another DDR indicator, 53BP1 at DSB sites in fibroblast (FT169A, YZ5 cells). Furthermore, DNA damage induced G2 checkpoint was inactivated and levels of 53BP1 were observed to be reduced when HDAC4 was knocked down leading to a conclusion that HDAC4 is critical in maintaining G2 checkpoint [47,76]. The roles of the different HDACs in the DDR are summarised in Table 1.
4. Impact of Chromatin Structure on Radiation Response
The architecture of chromatin during DNA damage induction, recognition, signalling and repair is an important factor that can dictate successful repair or mis-repair of DNA double strand breaks [23]. However, the topic has received little attention over the years, partly due to lack of efficient in vitro systems for manipulation of long chromatin and quantitative detection for DSBs [66]. As early as 1991, Smerdon proposed the “access-repair-restore” model to describe the impact of chromatin structure on DNA repair. In brief, the model proposed that in order for DNA damage to be repaired, DNA damage in different chromatin structures should be detected, local chromatin structure needs to be re-modelled to allow access for repair proteins to damaged sites and to be restored after repair [84].
For purposes of this review, a brief description of chromatin structure is justified. Chromatin is organized in structures named nucleosomes. Each nucleosome consists of DNA wrapped around histone octamer which consists of histones, H2A, H2B, H3 and H4. The nucleosome is then connected to a linker DNA and histone H1 [85]. The amino terminus tails of the histones protrude from the histone core and are open to various histone post translational modifications such as acetylation, ubiquitination or methylation. The two forms of chromatin, heterochromatin and euchromatin, are regulated by post-translation modification (PMT) of histones. Some of the post translational modifications that occur on the histone tails play an important role in the DDR [85,86].
4.1. Influence of Chromatin Structure on DNA Damage Induction , Detection and Repair
It has long been accepted that heterochromatin has a protective effect on DNA against ionizing radiation [23,24,87]. Cowel et al. observed that γ-H2AX foci, a marker of DSB, were absent from areas which contained heterochromatin markers HP1α and H3K9Me3 in nuclei of MCF-7 cells [88]. Similarly, Kim et al. reported an increased number of γ-H2AX foci in areas of open chromatin [89]. A similar observation was made by Takata et al., they reported a 5-50 fold decrease in DSB induction by γ-rays in condensed chromatin as compared to decondensed chromatin [23]. Clearly, there is agreement that heterochromatin confers protection against DNA DSB induction by radiation. However, the underlying mechanisms around this protective effect remain a matter of debate. Warters et al. argued that the protective effect against radiation is dependent on the level of chromatin-associated non-histone proteins in heterochromatin rather than the concentration of chromatin [87]. In other words, the more proteins carried by the chromatin, the higher the protection of chromatin from ionizing radiation. The authors came to the conclusion after observing a 70 times higher yield of DSB in deproteinised DNA as compared to intact nuclei. Elia and Bradley resolved that chromatin domains that differ in tertiary structure and protein composition may also differ in their susceptibility to DNA double strand breaks induced by ionizing radiation [23]. However, in both studies, the protective effect offered by chromatin compaction was acknowledged.
Nygren et al. investigated the role of DNA bound proteins in the protective effect and they reported an increase in the protective effect of a factor of 14 in single strand breaks and a factor of 5 in double strand breaks when DNA bound proteins were removed [90]. They concluded that DNA bound proteins protects chromatin to a limited extent by acting as free radical scavengers. The greater part of the protection was attributed to the fact that DNA in the chromatin is made up of large, compact aggregates where the distance between separate aggregates exceeds the effective range of the hydroxyl (.OH) radicals. Further, inside the large aggregates the amount of DNA damaging .OH radicals produced are less due to reduced water content as compared to when DNA is more evenly distributed in a given volume [90]. Similarly, Takata et al. observed that the level and composition of proteins were similar between condensed and decondensed chromatin in Hela cells, and concluded that the protective effect is due to the concentration of chromatin and not level of chromatin-associated proteins as previously proposed [23]. They reasoned that a lower chromatin concentration contains more water molecules, with subsequent increase in reactive radical that are formed. It remains contentious whether the opposing observations between Takata et al. and Warters et al. could be due to the different chromatin manipulation methods used in the respective studies [23].
4.2. DNA Damage Induced Chromatin Modifications
Evidence from earlier biochemical studies pointed out that induction of double strand breaks causes remodelling of chromatin structure around the damaged site [85,91]. Subsequently, Lisby et al. reported that in yeast Saccharomyces cerevisiae, DNA DSB localizes at repair foci which contain Rad52 protein, which suggests that multiple DSBs can be repaired by one Rad52 repair foci [92]. These findings implied that there is mobility of chromatin to allow DSBs to localize at one repair site [92,93]. The authors however acknowledged that the localization and interaction of DSB observed in yeast Saccharomyces cerevisiae might be due to the small nuclear size in yeast as compared to the nuclei of mammalian cells, rather than due to mobility of chromatin. Also, as compared to mammalian cells, homologous recombination is a dominant repair mechanism in yeast, which would explain the co-localization to Rad52 foci in yeast and not in mammalian cells [91,92]. Similarly, in a later study, relocalisation of DSB to nuclear periphery before recruitment of Rad51 was reported in Drospohlilla [94].
A contradictory observation was made by Kruhlak et al. in mammalian cells. The authors noted that remodelling of chromatin architecture at DSB sites does not involve large scale mobility of chromatin to assemble at repair centers, but rather small-scale mobility in micrometer range [91]. The authors reported chromatin expansion at 20 seconds after irradiation which lasted up to 180 second after UV irradiation in HeLa cells. In an attempt to explain the local expansion in chromatin that was observed in the area around the DSB, the authors conceptualised that after DSB formation, the break causes the damaged chromatin region to unfold relieving the tortional stress exerted by packaging of DNA, resulting in expansion and relaxation of chromatin [91]. They concluded that the observed chromatin relaxation and expansion might be due to the presence of DSB sensor proteins which possess chromatin unwinding properties [91]. Indeed, the increased acetylation of DDR proteins histones H2A and H4 at DSB sites was reported to lead to the rapid formation of open chromatin by a number of studies [95,96]. Perhaps, the reasoning offered by Takata et al. that chromatin relaxation after DSB induction was part of evolutionary conservation of the genome to allow access for repair protein, should be given consideration [23].
Another important observation made was an increase in the size of foci as chromatin becomes open [89]. A reasonable explanation was later offered by Kruhlak et al. that the time dependent increase in the size of individual foci which was noted was not due to merging of multiple DSBs as a result of mobility of chromatin, but rather due to the spreading of H2AX phosphorylation over large chromatin area, which subsequently acts as docking site for DNA damage proteins [91]. An akin reasoning was given by Krawczyk et al. that mobility of foci might simply be due to relaxation of chromatin not due to mobility of chromatin [97].
4.3. Chromatin Modification and Type of Radiation
The protective effect of heterochromatin against DSB induction has been linked to the type of radiation, i.e., low or high LET radiation [23]. Earlier reports associated the protective effect with low LET radiation. This was since radiolysis of water molecules with subsequent formation of hydroxyl radicals has long been acknowledged as the major contributor to DNA damage particularly when low LET radiation is used. It was therefore argued that, since heterochromatin has fewer water molecules per chromatin, less hydroxyl radicals are produced as discussed in section 4.2. The opposite is true for decondensed chromatin [23]. However, Takata et al. reported that heterochromatin protects against DNA damage not only from low LET radiation, but from heavy ions as well. The authors made this conclusion after observing a 7-fold increase in radioprotection in heterochromatin when carbon ion was used in HeLa cells [23]. Further, using Monte Carlo simulations, the complexity of DNA damage induction caused by low and high LET types of radiation was observed to be the same in heterochromatin, as well as in euchromatin. However, inefficient repair was noted in heterochromatin [23,85]. This lends support to the arguments presented Takata et al. that the protective effect of heterochromatin hinders efficient repair of DSB [23].
5. Sequencing of HDACi Treatment and Radiation
The use of HDAC inhibitors in combination with radiation therapy remains to be a matter of ongoing research. Evidence from numerous studies points that there is agreement on HDACi treatment before radiation (pre-irradiation HDACi protocol), with only a few studies having investigated the HDACi post-irradiation (post-irradiation HDACi protocol) [7,98,99]. In the studies using pre-irradiation HDACi protocols, HDACi treatment was given at different timepoints (3,6,16,18,24,48 hours) pre-irradiation in different studies [7,10,11,14,17,18,73,100,101,102,103,104,105,106,107,108]. Kim et al. determined that a 18 and 24 hour pre-irradiation incubation of A549 cell line with trichostatin A (TSA) resulted in enhanced radiation sensitisation to 2-8Gy X-rays as compared to HDACi treatment at 3,6, and 12 hours post irradiation [98]. In U251 glioma cell lines, dose enhancement factors of 1.38, 1.4 and 1.46 were reported when cells were exposed to 1.5 mmol/L valproic acid (VPA) at 6 hours, 24 hours and immediately, respectively, post-irradiation [6]. A greater dose enhancement factor of 1.71 was noted with a 16 hour pre- and post-incubation in VPA leading to a conclusion that pre-and post-exposure to HDACi is necessary for maximal radiosensitisation [7]. Noteworthy is also that a higher dose enhancement factor (1.46) was reported when VPA was administered immediately after radiation, as compared to when VPA was administered 6 hours and 24 hours post-irradiation. It is tempting to speculate that the modestly higher dose enhancement factor might in part be due to radiation induced chromatin modifications. As set forth by numerous studies [24,84,85,86,89,94], following induction of DSB by radiation, chromatin relaxation around the DSB occurs. Therefore, one could mechanistically reason that the addition of HDACi immediately post IR, coincides with the rapid chromatin changes that occurs post irradiation. In support, Kruhlak et al. reported chromatin changes as early as 20 seconds after irradiation [91].
Van Nifterik reported a dose enhancement factor of 1.3 and 1.4 for D384 medulloblastoma cells and 1.7 and 1.5 for T98 glioblastoma cells for 24- and 48-hour pre-incubation periods with 5mM VPA [99]. The authors further reported having not observed any enhancement, with dose enhancement factors of 1.1 for D384 cells and 1.0 for T98 cells, when cells were incubated in VPA for 24 hours post-irradiation only. It is tempting to speculate that the difference in radiosensitivity between the pre-and post-irradiation HDACi protocols may in part, be due to two important factors, i.e., different plating methods, i.e., pre-irradiation plating (pre-IR plating) and post-irradiation (post-IR plating), as well as the period of incubation in HDACi. Typically, radiation sensitivity studies are conducted using colony survival assays [109]. In the pre-IR plating setting, cells are seeded, allowed to attach and treated. In the post-IR setting, cells are treated followed by trypsinization and required numbers of cells are seeded in plates. In addition, the post-IR plating method has two methods that can be used, immediate plating (IP) and delayed plating (DP). In the IP method cells are seeded immediately after radiation and in the DP method cells are seeded hours after irradiation. Of the two post-IR plating methods IP was observed to exhibit a lower survival than DP [110,111]. The difference between the resultant cell survival curves when using the two methods was explained by the cell's capacity to repair potentially lethal damage [110,111]. Moreover, in most studies, it is not specified whether DP or IP was used which also poses a challenge for data integration. Oike et al. reported consistent SF2, SF4, SF6, SF8, SF10, D10 and D50 values between pre-IR plating and post-IR plating methods, using lung cancer (A549) and submandibular gland (HSG) cells [109]. The study, however, did not investigate the possible impact of delayed platting in the post-IR plating setting [109]. It remains undetermined whether consistent results observed by Oike et al. can extrapolated to other cell lines, most importantly, in the context of this review, whether the results can be applied to the combination treatment of HDACi and radiation.
Furthermore, whether a post-IR HDACi protocol or pre-IR HDACi protocol is used, the total period of HDACi incubation used in the various studies is different. As an example, Chinnaiyan et al. evaluated both pre- and post-IR HDACi protocols. In the post-IR HDACi protocol, cells were seeded in 6 well plates, VPA added at immediately 6 hours and 24 hours after irradiation. Cells were incubated in VPA containing media for the remainder of the assay [7]. When using the pre-irradiation HDACi protocol, cells were treated with VPA for 16 hours, irradiated and rinsed with PBS before fresh HDACi-free media was added. In the pre-and post-protocol cells were pre-treated with VPA for 16 hours and returned for incubation post IR. The authors reported improved radiosensitisation (factor of 1.7) when cells were exposed to VPA pre- and post-irradiation, as compared to factors of 1.3 when VPA was administered pre-IR only. This suggests that the removal of VPA containing media at plating could be reason for the observed non-enhancement. Van Nifterik et al. cells were treated with VPA 24 and 48 hours before irradiation and HDACi-containing media was removed during delayed plating at 24 hours post VPA administration [99]. Also, taking into account the review the modes of HDACi-induced cell death in section 2.1, HDACi induced autophagy is reported to depend on the duration and dose of HDACi [2]. It is enticing to speculate that different periods of HDACi incubation as well as different HDACi concentrations noted in the different studies, might have played a role on the mode of cell death, with subsequent differences in the observed results.
6. Conclusions
The combination therapy of HDAC inhibitor and radiation is complex. The matter is further complicated by the pleotropic effects of HDACi on histone and non-histone targets in cells. The biologic rationale for this combination therapy rests on the ability of HDACi to modulate epigenetics to maximise the radiation effect. Mechanistically, pre-IR HDACi treatment induces chromatin relaxation to facilitate increased DSB induction by radiation. However, from existing reports, different temporal sequencing protocols of HDACi and radiation have been used. Some studies employed the pre-IR HDACi protocol and others used post-IR HDACi protocol. The different plating methods, i.e., pre-IR or post-IR plating delayed plating or immediate plating also pose a challenge with the integration of data from different studies. Considering the ability of radiation and HDACi to modify chromatin structure, as well as the paradoxical relationship between apoptosis and autophagy under different conditions the molecular interplay of the two modalities is bound to be complex.
Several reports have emphasized that HDACi treatment depends on cell type, period of incubation with HDACi, as well as dose of HDACi. To date, evidence points out that incubation for 24 hours pre-irradiation to be most optimal sequence. However, in view of the complex mechanism of action of HDACi under different conditions, it seems reasonable that optimal temporal sequencing protocol of HDACi and radiation, as well as the optimal period of HDACi incubation be first determined for each cell. The mechanisms involved remain to be elusive. Inhibition of DNA DSB repair has traditionally been hailed as the main mechanism of HDACi induced radiosensitisation, however, emerging evidence from different cell lines suggests otherwise. For example, numerous studies reported that HDACi impairs DSB repair as evidenced by the prolonged appearance γ-H2AX foci. However, Moertl et al. reported not having observed any prolongation of foci after treatment with SAHA and CUDC-101 in pancreatic cell lines (Su.86.86, MIA Paca-2 and T3M-4) [100]. Clearly, more studies using different cell lines and different HDACi are required to fully unravel the mechanisms of radiosensitisation. For future studies, analysis of DSB repair proteins in addition to the appearance of γ-H2AX foci, as well as investigation of other modes of cell death such as autophagy and ROS production, may aid to fully elucidate the mechanisms involved.
Author Contributions
Conceptualization, E.S.; writing—original draft preparation, E.S.; writing—review and editing, E.S., N.S., C.V., A.J.; final approval of the version published, E.S., N.S., C.V. and A.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This review includes an image from previous publication from our research group. Pharmaceuticals 2023, 16(2), 227; https://doi.org/10.3390/ph16020227. Biorender was used for creating the images.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, J. H.; Choy, M. L.; Ngo, L.; Foster, S. S.; Marks, P. A. Histone deacetylase inhibitor induces DNA damage, which normal but not transformed cells can repair. Proc Natl Acad Sci U S A 2010, 107, 14639–14644. [Google Scholar] [CrossRef] [PubMed]
- Mrakovcic, M.; Bohner, L.; Hanisch, M.; Fröhlich, L. F. Epigenetic Targeting of Autophagy via HDAC Inhibition in Tumor Cells: Role of p53. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Camphausen, K.; Scott, T.; Sproull, M.; Tofilon, P. J. Enhancement of xenograft tumor radiosensitivity by the histone deacetylase inhibitor MS-275 and correlation with histone hyperacetylation. Clin Cancer Res 2004, 10 Pt 1, 6066–6071. [Google Scholar] [CrossRef] [PubMed]
- Camphausen, K.; Tofilon, P. J. Inhibition of histone deacetylation: a strategy for tumor radiosensitization. J Clin Oncol 2007, 25, 4051–4056. [Google Scholar] [CrossRef] [PubMed]
- Groselj, B.; Sharma, N. L.; Hamdy, F. C.; Kerr, M.; Kiltie, A. E. Histone deacetylase inhibitors as radiosensitisers: effects on DNA damage signalling and repair. Br J Cancer 2013, 108, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Damaskos, C.; Garmpis, N.; Valsami, S.; Kontos, M.; Spartalis, E.; Kalampokas, T.; Kalampokas, E.; Athanasiou, A.; Moris, D.; Daskalopoulou, A.; et al. Histone Deacetylase Inhibitors: An Attractive Therapeutic Strategy Against Breast Cancer. Anticancer Res 2017, 37, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Chinnaiyan, P.; Cerna, D.; Burgan, W. E.; Beam, K.; Williams, E. S.; Camphausen, K.; Tofilon, P. J. Postradiation sensitization of the histone deacetylase inhibitor valproic acid. Clin Cancer Res 2008, 14, 5410–5415. [Google Scholar] [CrossRef] [PubMed]
- Schlaff, C. D.; Arscott, W. T.; Gordon, I.; Tandle, A.; Tofilon, P.; Camphausen, K. Radiosensitization Effects of Novel Triple-Inhibitor CUDC-101 in Glioblastoma Multiforme and Breast Cancer Cells In Vitro. International Journal of Radiation Oncology*Biology*Physics 2013, 87. [Google Scholar] [CrossRef]
- Chiu, H. W.; Yeh, Y. L.; Wang, Y. C.; Huang, W. J.; Chen, Y. A.; Chiou, Y. S.; Ho, S. Y.; Lin, P.; Wang, Y. J. Suberoylanilide hydroxamic acid, an inhibitor of histone deacetylase, enhances radiosensitivity and suppresses lung metastasis in breast cancer in vitro and in vivo. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Baschnagel, A.; Russo, A.; Burgan, W. E.; Carter, D.; Beam, K.; Palmieri, D.; Steeg, P. S.; Tofilon, P.; Camphausen, K. Vorinostat enhances the radiosensitivity of a breast cancer brain metastatic cell line grown in vitro and as intracranial xenografts. Mol Cancer Ther 2009, 8, 1589–1595. [Google Scholar] [CrossRef]
- Chen, X.; Wong, P.; Radany, E.; Wong, J. Y. HDAC inhibitor, valproic acid, induces p53-dependent radiosensitization of colon cancer cells. Cancer Biother Radiopharm 2009, 24, 689–699. [Google Scholar] [CrossRef]
- Munshi, A.; Kurland, J. F.; Nishikawa, T.; Tanaka, T.; Hobbs, M. L.; Tucker, S. L.; Ismail, S.; Stevens, C.; Meyn, R. E. Histone deacetylase inhibitors radiosensitize human melanoma cells by suppressing DNA repair activity. Clin Cancer Res 2005, 11, 4912–4922. [Google Scholar] [CrossRef]
- Antrobus, J.; Parsons, J. L. Histone Deacetylases and Their Potential as Targets to Enhance Tumour Radiosensitisation. Radiation 2022, 2, 149–167. [Google Scholar] [CrossRef]
- Gerelchuluun, A.; Maeda, J.; Manabe, E.; Brents, C. A.; Sakae, T.; Fujimori, A.; Chen, D. J.; Tsuboi, K.; Kato, T. A. Histone Deacetylase Inhibitor Induced Radiation Sensitization Effects on Human Cancer Cells after Photon and Hadron Radiation Exposure. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Barazzuol, L.; Jeynes, J. C.; Merchant, M. J.; Wéra, A. C.; Barry, M. A.; Kirkby, K. J.; Suzuki, M. Radiosensitization of glioblastoma cells using a histone deacetylase inhibitor (SAHA) comparing carbon ions with X-rays. Int J Radiat Biol 2015, 91, 90–98. [Google Scholar] [CrossRef]
- Johnson, A. M.; Bennett, P. V.; Sanidad, K. Z.; Hoang, A.; Jardine, J. H.; Keszenman, D. J.; Wilson, P. F. Evaluation of Histone Deacetylase Inhibitors as Radiosensitizers for Proton and Light Ion Radiotherapy. Front Oncol 2021, 11, 735940. [Google Scholar] [CrossRef]
- Yu, J. I.; Choi, C.; Shin, S. W.; Son, A.; Lee, G. H.; Kim, S. Y.; Park, H. C. Valproic Acid Sensitizes Hepatocellular Carcinoma Cells to Proton Therapy by Suppressing NRF2 Activation. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Choi, C.; Lee, G. H.; Son, A.; Yoo, G. S.; Yu, J. I.; Park, H. C. Downregulation of Mcl-1 by Panobinostat Potentiates Proton Beam Therapy in Hepatocellular Carcinoma Cells. Cells 2021, 10. [Google Scholar] [CrossRef]
- Groselj, B.; Kerr, M.; Kiltie, A. E. Radiosensitisation of bladder cancer cells by panobinostat is modulated by Ku80 expression. Radiother Oncol 2013, 108, 429–433. [Google Scholar] [CrossRef]
- Jenke, R.; Ressing, N.; Hansen, F. K.; Aigner, A.; Buch, T. Anticancer Therapy with HDAC Inhibitors: Mechanism-Based Combination Strategies and Future Perspectives. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Maier, P.; Hartmann, L.; Wenz, F.; Herskind, C. Cellular Pathways in Response to Ionizing Radiation and Their Targetability for Tumor Radiosensitization. Int J Mol Sci 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Shabason, J. E.; Tofilon, P. J.; Camphausen, K. Grand rounds at the National Institutes of Health: HDAC inhibitors as radiation modifiers, from bench to clinic. J Cell Mol Med 2011, 15, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Hanafusa, T.; Mori, T.; Shimura, M.; Iida, Y.; Ishikawa, K.; Yoshikawa, K.; Yoshikawa, Y.; Maeshima, K. Chromatin Compaction Protects Genomic DNA from Radiation Damage. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, P.; Panyutin, I. V.; Remeeva, E.; Neumann, R. D.; Panyutin, I. G. Effect of Chromatin Structure on the Extent and Distribution of DNA Double Strand Breaks Produced by Ionizing Radiation; Comparative Study of hESC and Differentiated Cells Lines. International Journal of Molecular Sciences 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Arlinghaus, L. R.; Cardin, D. B.; Goff, L.; Berlin, J. D.; Parikh, A.; Abramson, R. G.; Yankeelov, T. E.; Hiebert, S.; Merchant, N.; et al. Phase I trial of vorinostat added to chemoradiation with capecitabine in pancreatic cancer. Radiotherapy and Oncology 2016, 119, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Anderson, S. K.; Miller, C. R.; Sarkaria, J. N.; Jaeckle, K.; Buckner, J. C.; Ligon, K. L.; Ballman, K. V.; Moore, D. F., Jr; Nebozhyn, M.; et al. Phase I/II trial of vorinostat combined with temozolomide and radiation therapy for newly diagnosed glioblastoma: results of Alliance N0874/ABTC 02. Neuro-Oncology 2017, 20, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Gurbani, S. S.; Yoon, Y.; Weinberg, B. D.; Salgado, E.; Press, R. H.; Cordova, J. S.; Ramesh, K. K.; Liang, Z.; Vega, J. V.; Voloschin, A.; et al. Assessing Treatment Response of Glioblastoma to an HDAC Inhibitor Using Whole-Brain Spectroscopic MRI. Tomography 2019, 5, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Teknos, T. N.; Grecula, J.; Agrawal, A.; Old, M. O.; Ozer, E.; Carrau, R.; Kang, S.; Rocco, J.; Blakaj, D.; Diavolitsis, V.; et al. A phase 1 trial of Vorinostat in combination with concurrent chemoradiation therapy in the treatment of advanced staged head and neck squamous cell carcinoma. Investigational New Drugs 2019, 37, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Richon, V.; Ni, X.; Talpur, R.; Duvic, M. Selective induction of apoptosis by histone deacetylase inhibitor SAHA in cutaneous T-cell lymphoma cells: relevance to mechanism of therapeutic action. J Invest Dermatol 2005, 125, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb Perspect Med 2016, 6. [Google Scholar] [CrossRef]
- Mrakovcic, M.; Kleinheinz, J.; Frohlich, L. F. Histone Deacetylase Inhibitor-Induced Autophagy in Tumor Cells: Implications for p53. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Bolden, J. E.; Peart, M. J.; Johnstone, R. W. Anticancer activities of histone deacetylase inhibitors. Nat Rev Drug Discov 2006, 5, 769–784. [Google Scholar] [CrossRef]
- Fotheringham, S.; Epping, M. T.; Stimson, L.; Khan, O.; Wood, V.; Pezzella, F.; Bernards, R.; La Thangue, N. B. Genome-wide loss-of-function screen reveals an important role for the proteasome in HDAC inhibitor-induced apoptosis. Cancer Cell 2009, 15, 57–66. [Google Scholar] [CrossRef]
- Everix, L.; Seane, E. N.; Ebenhan, T.; Goethals, I.; Bolcaen, J. Introducing HDAC-Targeting Radiopharmaceuticals for Glioblastoma Imaging and Therapy. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Dokmanovic, M.; Clarke, C.; Marks, P. A. Histone Deacetylase Inhibitors: Overview and Perspectives. Molecular Cancer Research 2007, 5, 981–989. [Google Scholar] [CrossRef]
- Smalley, J. P.; Cowley, S. M.; Hodgkinson, J. T. Bifunctional HDAC Therapeutics: One Drug to Rule Them All? Molecules 2020, 25. [Google Scholar] [CrossRef]
- Rajak, H.; Singh, A.; Raghuwanshi, K.; Kumar, R.; Dewangan, P. K.; Veerasamy, R.; Sharma, P. C.; Dixit, A.; Mishra, P. A structural insight into hydroxamic acid based histone deacetylase inhibitors for the presence of anticancer activity. Curr Med Chem 2014, 21, 2642–2664. [Google Scholar] [CrossRef]
- Passaro, E.; Papulino, C.; Chianese, U.; Toraldo, A.; Congi, R.; Del Gaudio, N.; Nicoletti, M. M.; Benedetti, R.; Altucci, L. HDAC6 Inhibition Extinguishes Autophagy in Cancer: Recent Insights. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Zhang, J.; Zhong, Q. Histone deacetylase inhibitors and cell death. Cell Mol Life Sci 2014, 71, 3885–3901. [Google Scholar] [CrossRef]
- Frew, A. J.; Johnstone, R. W.; Bolden, J. E. Enhancing the apoptotic and therapeutic effects of HDAC inhibitors. Cancer Lett 2009, 280, 125–133. [Google Scholar] [CrossRef]
- Mrakovcic, M.; Kleinheinz, J.; Fröhlich, L. F. p53 at the Crossroads between Different Types of HDAC Inhibitor-Mediated Cancer Cell Death. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Insinga, A.; Minucci, S.; Pelicci, P. G. Mechanisms of selective anticancer action of histone deacetylase inhibitors. Cell Cycle 2005, 4, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Peart, M. J. Novel mechanisms of apoptosis induced by histone deacetylase inhibitors.full. CANCER RESEARCH 2003, 63, 4460–4471. [Google Scholar]
- Gong, P.; Wang, Y.; Jing, Y. Apoptosis Induction byHistone Deacetylase Inhibitors in Cancer Cells: Role of Ku70. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Diao, H.; Dong, N.; Su, X.; Wang, B.; Mo, Q.; Yu, H.; Wang, X.; Chen, C. Histone deacetylase inhibitor induces cell apoptosis and cycle arrest in lung cancer cells via mitochondrial injury and p53 up-acetylation. Cell Biol Toxicol 2016, 32, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. J.; Chee, C. E.; Huang, S.; Sinicrope, F. A. The role of autophagy in cancer: therapeutic implications. Mol Cancer Ther 2011, 10, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Gao, Z.; Marks, P. A.; Jiang, X. Apoptotic and autophagic cell death induced by histone deacetylase inhibitors. Proc Natl Acad Sci U S A 2004, 101, 18030–18035. [Google Scholar] [CrossRef] [PubMed]
- Rebecca, V. W.; Amaravadi, R. K. Emerging strategies to effectively target autophagy in cancer. Oncogene 2016, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kimmelman, A. C. The dynamic nature of autophagy in cancer. Genes Dev 2011, 25, 1999–2010. [Google Scholar] [CrossRef]
- Pagotto, A.; Pilotto, G.; Mazzoldi, E. L.; Nicoletto, M. O.; Frezzini, S.; Pastò, A.; Amadori, A. Autophagy inhibition reduces chemoresistance and tumorigenic potential of human ovarian cancer stem cells. Cell Death & Disease 2017, 8, e2943–e2943. [Google Scholar] [CrossRef]
- El-Khoury, V.; Pierson, S.; Szwarcbart, E.; Brons, N. H.; Roland, O.; Cherrier-De Wilde, S.; Plawny, L.; Van Dyck, E.; Berchem, G. Disruption of autophagy by the histone deacetylase inhibitor MGCD0103 and its therapeutic implication in B-cell chronic lymphocytic leukemia. Leukemia 2014, 28, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Hrzenjak, A.; Kremser, M. L.; Strohmeier, B.; Moinfar, F.; Zatloukal, K.; Denk, H. SAHA induces caspase-independent, autophagic cell death of endometrial stromal sarcoma cells by influencing the mTOR pathway. J Pathol 2008, 216, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, L. F.; Mrakovcic, M.; Smole, C.; Zatloukal, K. Molecular mechanism leading to SAHA-induced autophagy in tumor cells: evidence for a p53-dependent pathway. Cancer Cell Int 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. L.; Yang, P. M.; Shun, C. T.; Wu, M. S.; Weng, J. R.; Chen, C. C. Autophagy potentiates the anti-cancer effects of the histone deacetylase inhibitors in hepatocellular carcinoma. Autophagy 2010, 6, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Gilardini Montani, M. S.; Granato, M.; Santoni, C.; Del Porto, P.; Merendino, N.; D'Orazi, G.; Faggioni, A.; Cirone, M. Histone deacetylase inhibitors VPA and TSA induce apoptosis and autophagy in pancreatic cancer cells. Cell Oncol (Dordr) 2017, 40, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Richon, V. M.; Sandhoff, T. W.; Rifkind, R. A.; Marks, P. A. Histone deacetylase inhibitor selectively induces p21WAF1 expression and gene-associated histone acetylation. Proc Natl Acad Sci U S A 2000, 97, 10014–10019. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yang, Y.; Liu, S.; Lu, J.; Huang, B.; Zhang, Y. HDAC inhibitor PAC-320 induces G2/M cell cycle arrest and apoptosis in human prostate cancer. Oncotarget 2018, 9, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, H. A.; Chu, K. B.; Moon, E. K.; Kim, S. S.; Quan, F. S. Sensitization to oxidative stress and G2/M cell cycle arrest by histone deacetylase inhibition in hepatocellular carcinoma cells. Free Radic Biol Med 2020, 147, 129–138. [Google Scholar] [CrossRef]
- Hrgovic, I.; Doll, M.; Kleemann, J.; Wang, X.-F.; Zoeller, N.; Pinter, A.; Kippenberger, S.; Kaufmann, R.; Meissner, M. The histone deacetylase inhibitor trichostatin a decreases lymphangiogenesis by inducing apoptosis and cell cycle arrest via p21-dependent pathways. BMC Cancer 2016, 16. [Google Scholar] [CrossRef]
- Schoepflin, Z. R.; Shapiro, I. M.; Risbud, M. V. Class I and IIa HDACs Mediate HIF-1α Stability Through PHD2-Dependent Mechanism, While HDAC6, a Class IIb Member, Promotes HIF-1α Transcriptional Activity in Nucleus Pulposus Cells of the Intervertebral Disc. J Bone Miner Res 2016, 31, 1287–1299. [Google Scholar] [CrossRef]
- Jeong, J. W.; Bae, M. K.; Ahn, M. Y.; Kim, S. H.; Sohn, T. K.; Bae, M. H.; Yoo, M. A.; Song, E. J.; Lee, K. J.; Kim, K. W. Regulation and destabilization of HIF-1alpha by ARD1-mediated acetylation. Cell 2002, 111, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Deroanne, C. F.; Bonjean, K.; Servotte, S.; Devy, L.; Colige, A.; Clausse, N.; Blacher, S.; Verdin, E.; Foidart, J. M.; Nusgens, B. V.; et al. Histone deacetylases inhibitors as anti-angiogenic agents altering vascular endothelial growth factor signaling. Oncogene 2002, 21, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Kuljaca, S.; Tee, A.; Marshall, G. M. Histone deacetylase inhibitors: multifunctional anticancer agents. Cancer Treat Rev 2006, 32, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Iordache, F.; Buzila, C.; Constantinescu, A.; Andrei, E.; Maniu, H. Histone deacetylase (HDAC) inhibitors down-regulate endothelial lineage commitment of umbilical cord blood derived endothelial progenitor cells. Int J Mol Sci 2012, 13, 15074–15085. [Google Scholar] [CrossRef] [PubMed]
- Ellis, L.; Pili, R. Histone Deacetylase Inhibitors: Advancing Therapeutic Strategies in Hematological and Solid Malignancies. Pharmaceuticals (Basel) 2010, 3, 2411–2469. [Google Scholar] [CrossRef]
- Munshi, A.; Tanaka, T.; Hobbs, M. L.; Tucker, S. L.; Richon, V. M.; Meyn, R. E. Vorinostat, a histone deacetylase inhibitor, enhances the response of human tumor cells to ionizing radiation through prolongation of γ-H2AX foci. Molecular Cancer Therapeutics 2006, 5, 1967–1974. [Google Scholar] [CrossRef]
- Ediriweera, M. K.; Tennekoon, K. H.; Samarakoon, S. R. Emerging role of histone deacetylase inhibitors as anti-breast-cancer agents. Drug Discov Today 2019, 24, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Luu, T. H.; Morgan, R. J.; Leong, L.; Lim, D.; McNamara, M.; Portnow, J.; Frankel, P.; Smith, D. D.; Doroshow, J. H.; Wong, C.; et al. A phase II trial of vorinostat (suberoylanilide hydroxamic acid) in metastatic breast cancer: a California Cancer Consortium study. Clin Cancer Res 2008, 14, 7138–7142. [Google Scholar] [CrossRef]
- Vitti, E. T.; Parsons, J. L. The Radiobiological Effects of Proton Beam Therapy: Impact on DNA Damage and Repair. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Bian, L.; Meng, Y.; Zhang, M.; Li, D. MRE11-RAD50-NBS1 complex alterations and DNA damage response: implications for cancer treatment. Molecular Cancer 2019, 18. [Google Scholar] [CrossRef]
- Hall Eric J, A. J. G. Radiobiology for the Radiologist; LIPPINCOT WILLIAMS AND WILKINS, 2012.
- Fontana, A. O.; Augsburger, M. A.; Grosse, N.; Guckenberger, M.; Lomax, A. J.; Sartori, A. A.; Pruschy, M. N. Differential DNA repair pathway choice in cancer cells after proton- and photon-irradiation. Radiother Oncol 2015, 116, 374–380. [Google Scholar] [CrossRef]
- Kachhap, S. K.; Rosmus, N.; Collis, S. J.; Kortenhorst, M. S.; Wissing, M. D.; Hedayati, M.; Shabbeer, S.; Mendonca, J.; Deangelis, J.; Marchionni, L.; et al. Downregulation of homologous recombination DNA repair genes by HDAC inhibition in prostate cancer is mediated through the E2F1 transcription factor. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Miller, K. M.; Tjeertes, J. V.; Coates, J.; Legube, G.; Polo, S. E.; Britton, S.; Jackson, S. P. Human HDAC1 and HDAC2 function in the DNA-damage response to promote DNA nonhomologous end-joining. Nat Struct Mol Biol 2010, 17, 1144–1151. [Google Scholar] [CrossRef]
- Thurn, K. T.; Thomas, S.; Raha, P.; Qureshi, I.; Munster, P. N. Histone deacetylase regulation of ATM-mediated DNA damage signaling. Mol Cancer Ther 2013, 12, 2078–2087. [Google Scholar] [CrossRef]
- Kao, G. D.; McKenna, W. G.; Guenther, M. G.; Muschel, R. J.; Lazar, M. A.; Yen, T. J. Histone deacetylase 4 interacts with 53BP1 to mediate the DNA damage response. J Cell Biol 2003, 160, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Cubizolles, F.; Zhang, Y.; Reichert, N.; Kohler, H.; Seiser, C.; Matthias, P. Histone deacetylases 1 and 2 act in concert to promote the G1-to-S progression. Genes Dev 2010, 24, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Hai, R.; He, L.; Shu, G.; Yin, G. Characterization of Histone Deacetylase Mechanisms in Cancer Development. Front Oncol 2021, 11, 700947. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Fernández, A.; Cabrero, J. R.; Serrador, J. M.; Sánchez-Madrid, F. HDAC6: a key regulator of cytoskeleton, cell migration and cell-cell interactions. Trends Cell Biol 2008, 18, 291–297. [Google Scholar] [CrossRef]
- Bhaskara, S.; Knutson, S. K.; Jiang, G.; Chandrasekharan, M. B.; Wilson, A. J.; Zheng, S.; Yenamandra, A.; Locke, K.; Yuan, J. L.; Bonine-Summers, A. R.; et al. Hdac3 is essential for the maintenance of chromatin structure and genome stability. Cancer Cell 2010, 18, 436–447. [Google Scholar] [CrossRef]
- Nishimoto, K.; Niida, H.; Uchida, C.; Ohhata, T.; Kitagawa, K.; Motegi, A.; Suda, T.; Kitagawa, M. HDAC3 Is Required for XPC Recruitment and Nucleotide Excision Repair of DNA Damage Induced by UV Irradiation. Mol Cancer Res 2020, 18, 1367–1378. [Google Scholar] [CrossRef]
- M, Z. HDAC6 Deacetylates and Ubiquitinates MSH2 to Maintain Proper Levels of MutSα. Molecular Cell 2014, 55, 31–46. [Google Scholar]
- Wang, R.-H. Impaired DNA Damage Response, Genome Instability, and Tumorigenesis in SIRT1 Mutant Mice.
- Polo, S. E.; Almouzni, G. Chromatin dynamics after DNA damage: The legacy of the access-repair-restore model. DNA Repair (Amst) 2015, 36, 114–121. [Google Scholar] [CrossRef]
- Fortuny, A.; Polo, S. E. The response to DNA damage in heterochromatin domains. Chromosoma 2018, 127, 291–300. [Google Scholar] [CrossRef]
- Etier, A.; Dumetz, F.; Chéreau, S.; Ponts, N. Post-Translational Modifications of Histones Are Versatile Regulators of Fungal Development and Secondary Metabolism. Toxins (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Warters, R. L.; Lyons, B. W. Variation in radiation-induced formation of DNA double-strand breaks as a function of chromatin structure. Radiat Res 1992, 130, 309–318. [Google Scholar] [CrossRef]
- Cowell, I. G.; Sunter, N. J.; Singh, P. B.; Austin, C. A.; Durkacz, B. W.; Tilby, M. J. gammaH2AX foci form preferentially in euchromatin after ionising-radiation. PLoS One 2007, 2. [Google Scholar] [CrossRef]
- Kim, J. A.; Kruhlak, M.; Dotiwala, F.; Nussenzweig, A.; Haber, J. E. Heterochromatin is refractory to gamma-H2AX modification in yeast and mammals. J Cell Biol 2007, 178, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Nygren, J.; Ljungman, M.; Ahnström, M. Chromatin Structure and Radiation-induced DNA Strand Breaks in Human Cells: Soluble Scavengers and DNA-bound Proteins Offer a Better Protection Against Single- than Double-strand Breaks. International Journal of Radiation Biology 1995, 68, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Kruhlak, M. J.; Celeste, A.; Dellaire, G.; Fernandez-Capetillo, O.; Müller, W. G.; McNally, J. G.; Bazett-Jones, D. P.; Nussenzweig, A. Changes in chromatin structure and mobility in living cells at sites of DNA double-strand breaks. J Cell Biol 2006, 172, 823–834. [Google Scholar] [CrossRef]
- Lisby, M.; Mortensen, U. H.; Rothstein, R. Colocalization of multiple DNA double-strand breaks at a single Rad52 repair centre. Nat Cell Biol 2003, 5, 572–577. [Google Scholar] [CrossRef]
- Aten, J. A.; Stap, J.; Krawczyk, P. M.; van Oven, C. H.; Hoebe, R. A.; Essers, J.; Kanaar, R. Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science 2004, 303, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Amaral, N.; Ryu, T.; Li, X.; Chiolo, I. Nuclear Dynamics of Heterochromatin Repair. Trends in Genetics 2017, 33, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Murr, R.; Loizou, J. I.; Yang, Y. G.; Cuenin, C.; Li, H.; Wang, Z. Q.; Herceg, Z. Histone acetylation by Trrap-Tip60 modulates loading of repair proteins and repair of DNA double-strand breaks. Nat Cell Biol 2006, 8, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, Y.; Jiang, X.; Ayrapetov, M. K.; Moskwa, P.; Yang, S.; Weinstock, D. M.; Price, B. D. The p400 ATPase regulates nucleosome stability and chromatin ubiquitination during DNA repair. J Cell Biol 2010, 191, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, P. M.; Borovski, T.; Stap, J.; Cijsouw, T.; ten Cate, R.; Medema, J. P.; Kanaar, R.; Franken, N. A.; Aten, J. A. Chromatin mobility is increased at sites of DNA double-strand breaks. J Cell Sci 2012, 125 Pt 9, 2127–2133. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. H.; Kim, I. H.; Shin, J. H.; Kim, H. J.; Kim, I. A. Sequence-Dependent Radiosensitization of Histone Deacetylase Inhibitors Trichostatin A and SK-7041. Cancer Res Treat 2013, 45, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Van Nifterik, K. A.; Van den Berg, J.; Slotman, B. J.; Lafleur, M. V.; Sminia, P.; Stalpers, L. J. Valproic acid sensitizes human glioma cells for temozolomide and gamma-radiation. J Neurooncol 2012, 107, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Moertl, S.; Payer, S.; Kell, R.; Winkler, K.; Anastasov, N.; Atkinson, M. J. Comparison of Radiosensitization by HDAC Inhibitors CUDC-101 and SAHA in Pancreatic Cancer Cells. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Munshi, A.; Tanaka, T.; Hobbs, M. L.; Tucker, S. L.; Richon, V. M.; Meyn, R. E. Vorinostat, a histone deacetylase inhibitor, enhances the response of human tumor cells to ionizing radiation through prolongation of gamma-H2AX foci. Mol Cancer Ther 2006, 5, 1967–1974. [Google Scholar] [CrossRef]
- Shanthi Adimoolam*, M. S. , Jun Chen*, Patti Thiemann*, James M. Ford†, and Joseph J. Buggy. HDAC-inhibitor-PCI24781-decreases-RAD51-expression-and-inhibits-homologous-recombination. PNAS 104, 9482. [Google Scholar] [CrossRef]
- Mueller, S.; Yang, X.; Sottero, T. L.; Gragg, A.; Prasad, G.; Polley, M. Y.; Weiss, W. A.; Matthay, K. K.; Davidoff, A. M.; DuBois, S. G.; et al. Cooperation of the HDAC inhibitor vorinostat and radiation in metastatic neuroblastoma: efficacy and underlying mechanisms. Cancer Lett 2011, 306, 223–229. [Google Scholar] [CrossRef]
- Chen X, W. P. , Radany E, Wong JY. HDAC inhibitor, valproic acid, induces p53-dependent radiosensitization of colon cancer cells. Cancer Biother Radiopharm. [CrossRef]
- Flatmark, K.; Nome, R. V.; Folkvord, S.; Bratland, A.; Rasmussen, H.; Ellefsen, M. S.; Fodstad, O.; Ree, A. H. Radiosensitization of colorectal carcinoma cell lines by histone deacetylase inhibition. Radiat Oncol 2006, 1, 25. [Google Scholar] [CrossRef] [PubMed]
- Saelen, M. G.; Ree, A. H.; Kristian, A.; Fleten, K. G.; Furre, T.; Hektoen, H. H.; Flatmark, K. Radiosensitization by the histone deacetylase inhibitor vorinostat under hypoxia and with capecitabine in experimental colorectal carcinoma. Radiation Oncology 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Shoji, M.; Ninomiya, I.; Makino, I.; Kinoshita, J.; Nakamura, K.; Oyama, K.; Nakagawara, H.; Fujita, H.; Tajima, H.; Takamura, H.; et al. Valproic acid, a histone deacetylase inhibitor, enhances radiosensitivity in esophageal squamous cell carcinoma. Int J Oncol 2012, 40, 2140–2146. [Google Scholar] [CrossRef] [PubMed]
- Perona, M.; Thomasz, L.; Rossich, L.; Rodriguez, C.; Pisarev, M. A.; Rosemblit, C.; Cremaschi, G. A.; Dagrosa, M. A.; Juvenal, G. J. Radiosensitivity enhancement of human thyroid carcinoma cells by the inhibitors of histone deacetylase sodium butyrate and valproic acid. Mol Cell Endocrinol 2018, 478, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Oike, T.; Hirota, Y.; Dewi Maulany Darwis, N.; Shibata, A.; Ohno, T. Comparison of Clonogenic Survival Data Obtained by Pre- and Post-Irradiation Methods. J Pers Med 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg-Schwager, M.; Frankenberg, D.; Harbich, R. Potentially lethal damage repair is due to the difference of DNA double-strand break repair under immediate and delayed plating conditions. Radiat Res 1987, 111, 192–200. [Google Scholar] [CrossRef]
- Reddy, N. M.; Kapiszewska, M.; Lange, C. S. Detection of X-ray damage repair by the immediate versus delayed plating technique is dependent on cell shape and cell concentration. Scanning Microsc 1992, 6, 543–555; [Google Scholar]
Figure 1.
Overview of the broad effects of HDAC inhibitors [34].
Figure 1.
Overview of the broad effects of HDAC inhibitors [34].
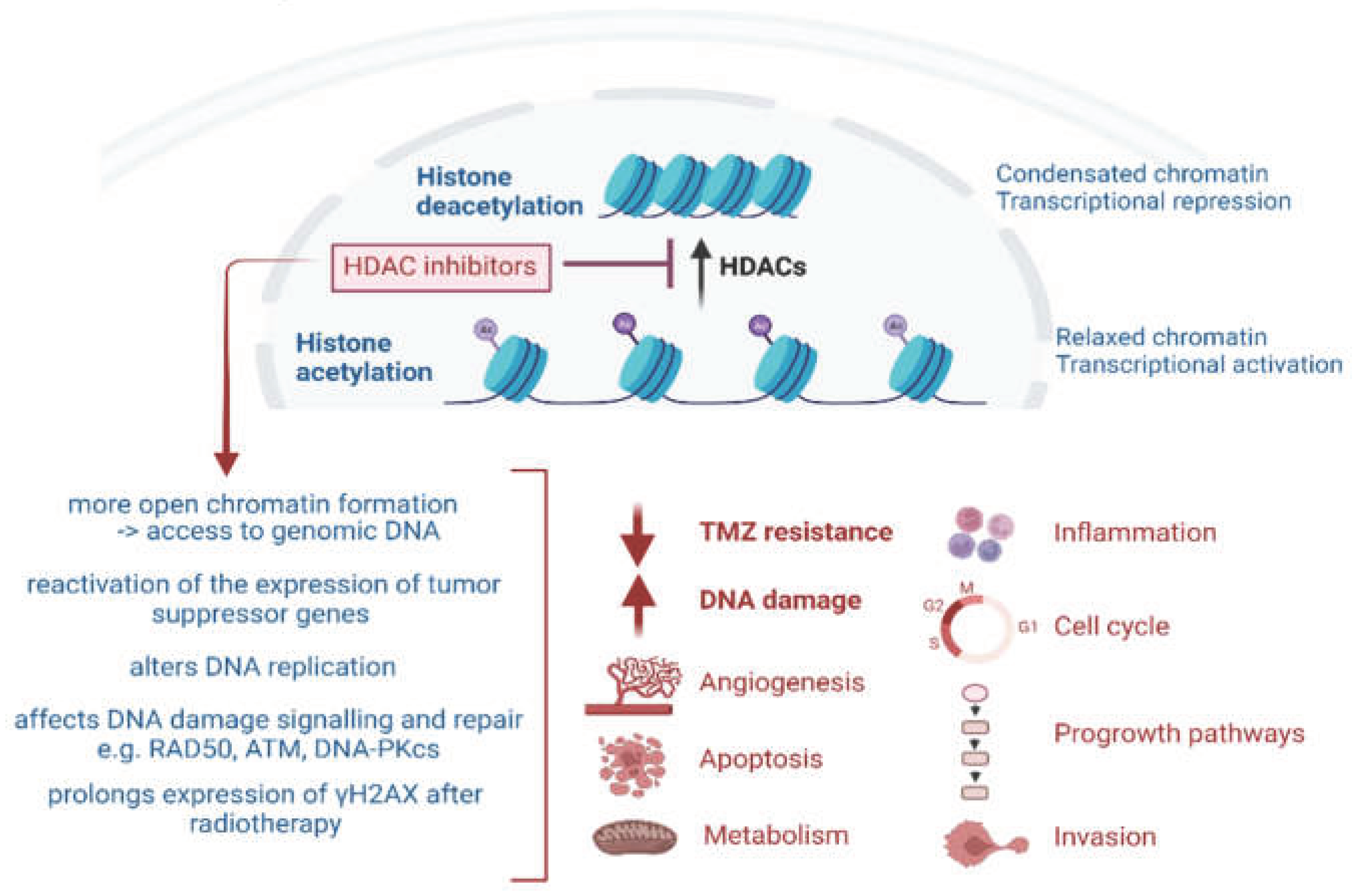
Figure 2.
Illustration of the role of HDACs in the DDR (Created with BioRender.com).
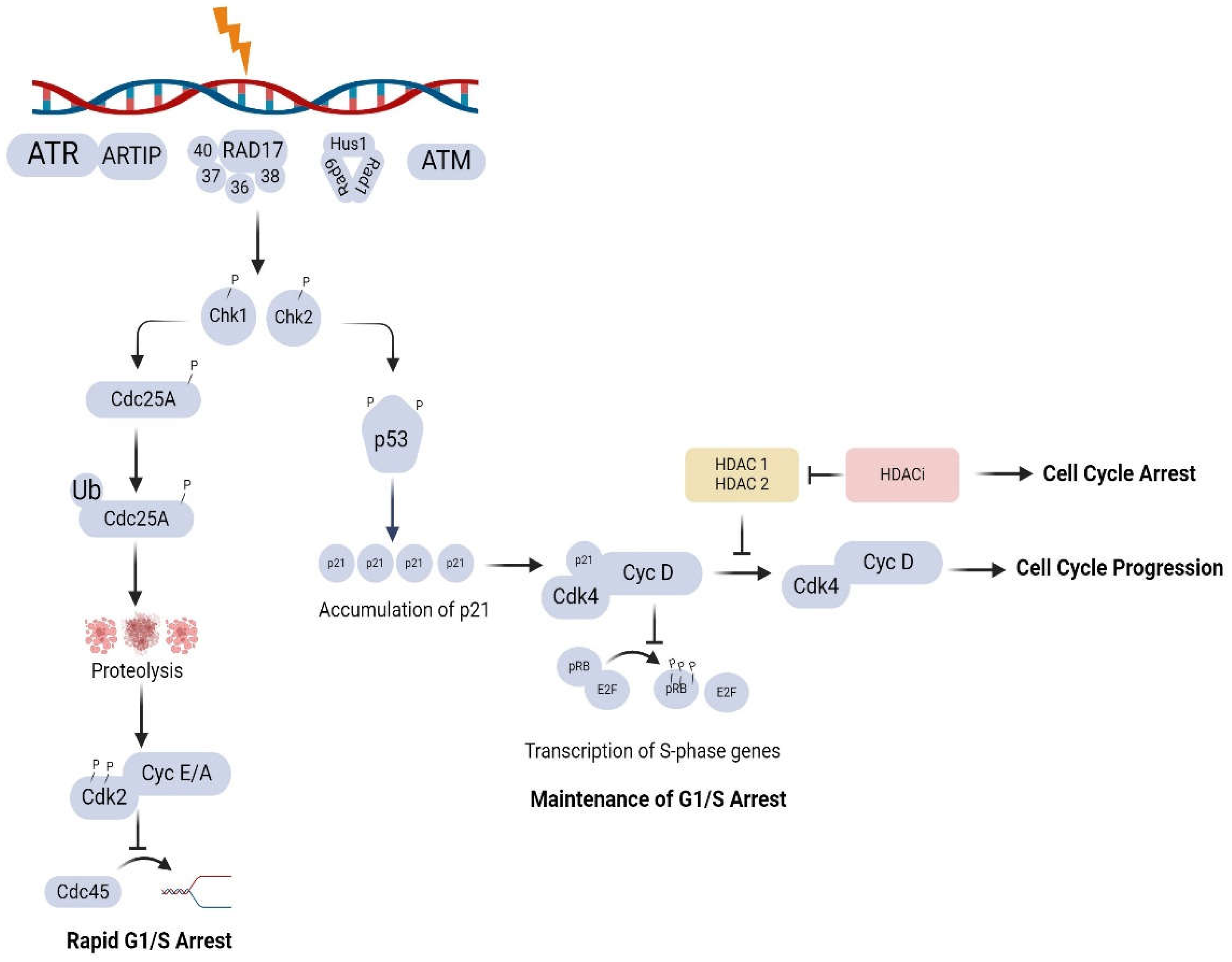

Table 1.
Roles of HDACs in the DDR.
| HDAC | Role | Reference | |
|---|---|---|---|
| HDAC 1 and 2 | DNA-damage signalling | data | |
| Stabilise broken ends during NHEJ | [74] | ||
| Influence persistence of Ku70 and Artemis at DNA damage sites- promoting NHEJ | [13] | ||
| Deactivates the function of p21 and p27 | [77,78] | ||
| Hypo-phosphorylation of RB gene | [79] | ||
| HDAC 3 | Maintenance of chromatin structure and genomic stability | [80] | |
| Essential for DNA DSB repair | [13] | ||
| Recruits Xeroderma Pigmentosum C (XPC) during Nucleotide excision repair (NER) | [81] | ||
| HDAC 4 | Silencing of chromatin near broken ends. | [76] | |
| Co-localises with 53BP foci, and contributes to stability of 53BP | [13] | ||
| HDAC 6 | Reduces cellular sensitivity to damaging agents | [82] | |
| Repair of DNA mismatch | [13] | ||
| HDAC 9 and 10 | DSB repair via the HR pathway | [13] | |
| G2/M transition -regulates transcription of cyclin A2 | [78] | ||
| SIRT1 | Reduces activity of p53Modulation of γ-H2AX foci, BBRCA1, Rad51, NBS foci formation | [83] | |
| SIRT6 | Facilitates DSB repair by activating PARP1 | [78] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



