
Submitted:
03 April 2024
Posted:
03 April 2024
You are already at the latest version
Abstract
Dolichandrone spathacea is a rare and endangered semi-mangrove plant species with an extremely small population, naturally distributed only in Zhanjiang City and the east coast of Hainan Island, China. Regrettably, the present situation of the D. spathacea population in China has not yet been carefully investigated. In this study, we aim to evaluate the current resource status and population dynamic characteristics of D. spathacea on Hainan Island, China, in order to propose sustainable conservation and management strategies for future ecological restoration of its natural population. Our results show that D. spathacea on Hainan Island can be divided into four populations. All the D. spathacea populations present a state of overall dispersion, local concentration, and occasionally sporadic existence, and they exist in geographical isolation. The young, middle, and old D. spathacea plants account for 20.42%, 66.20%, and 13.38%, respectively, indicating that the overall age class of the D. spathacea population on Hainan Island belongs to a declining type. Furthermore, instead of temporal structure, we chose diameter at breast height (DBH) to establish a static life table, draw a population survival curve, and quantify the future development trend through population dynamic analysis and time-series prediction. These results directly point out that the D. spathacea population on Hainan Island is sensitive to external disturbances and possesses two main increases in mortality rate—namely, in its juvenile and mature stages—due to weak vitality, competition, and anthropogenic interferences, which are the most important reasons for its endangered status. Depending on the current conditions of the D. spathacea population, we should conserve and expand mature trees in situ, preserve their germplasm resources, rehabilitate their habitats to promote provenance restoration, and conduct artificial cultivation and spreading planting in order to realize the sustainable conservation and management of D. spathacea.
Keywords:
Mangrove
; Dolichandrone spathacea
; population dynamic characteristics
; natural population
; diameter at breast height
; static life table
1. Introduction
Mangrove forests are special vegetated and coastal ecosystems, globally dominating around 1.7 × 105 km2 along the intertidal estuaries and coasts of tropical and subtropical areas and functioning as one of the vital regions for major biological activities [1,2,3]. Mangrove wetland ecosystems are considered as a transitional zone between terrestrial and marine environments, and they comprise approximately 75% of the acreage of coastlines worldwide [4]. Mangrove forests possess multiple properties, including high productivity, high decomposition rate, high restitution rate, and high stress resistance [5]. Meanwhile, mangrove ecosystems can supply large-scale ecological value to local communities, thereby continually stabilizing the intertidal regions [1,4,5,6]. In particular, they can purify the ocean through phytoremediation, offer detritus and superior refuges for aquatic animals, produce biomass energy resources for local residents, maintain climatic stability through their huge sequestration and storage abilities, and preserve coastal buffer zones against natural disasters such as sea waves, tides, and storm events due to their large coverage and strong root systems [1,4,5,6].
Unfortunately, because of their special ecological niche between the land and ocean, mangrove forests are extremely susceptible to both anthropogenic and natural destruction in a global context. First, mangrove forests are frequently exposed to the effects of climate changes, such as extreme temperature and precipitation, sea level rise, biotic and abiotic stresses, and sediment property changes in terms of natural factors [7,8,9]. Furthermore, with the rapid development of industrialization and urbanization, aquaculture activity, seawall construction, metal smelting, and industrial wastewater discharge have consequently followed in recent decades [7,8,9]. In addition, due to the highly valuable wood of mangrove trees, coastal dwellers persistently carry out unsustainable land clearing, illegal logging, and over-exploitation [10]. As a result, mangrove forests are among one of the fastest disappearing ecosystems, with a decline rate of 1–2% per year globally, which is equal to (or even greater than) decreases in tropical rainforests and their adjacent coral reefs [11]. Consequently, around 16% of mangrove species are on the verge of extinction on Earth [12]. In terms of Chinese mangroves, the mangrove resource investigation conducted by the National Investigation of Forest Resource in 1956 reported that their initial acreage approximately covered 2.5 × 105 ha; however, they dramatically dropped to 4.2 × 104 ha [13]. The worse news is that China’s mangrove area sharply plunged to 2.1 × 104 ha and 1.5 × 104 ha from 1970 to 1980, respectively, due to land reclamation by seaside residents [13]. In a word, the reserve of Chinese mangrove forests has declined by around a half from 1950 to 2001 due to a series of anthropogenic and natural factors [13]. Hence, preventing the continued loss of natural mangrove resources and protecting endangered mangrove species requires more attention by humans.
Dolichandrone spathacea (L.f.) K. Schum. (also named mangrove trumpet tree) is a semi-mangrove species belonging to the genus Dolichandrone of the Bignoniaceae, which can wildly grow on land without tide effects as well as in waterlogged beaches and estuaries [14]. It is naturally found in regions of southern India, Sri Lanka, and New Caledonia (island) [15,16] and is also sparsely naturally distributed in areas of China including Zhanjiang City, Guangdong Province, and the east coast of Hainan Island [17]. In the natural world, its common companion species are Nypa fruticans Wurmb, Bruguiera gymnorhiza (L.) Savigny, Bruguiera sexangula (Lour.) Poir., Pongamia pinnata (L.) Pierre, Cerbera manghas L., and so on [17]. As an excellent coastal shelterbelt mangrove species, D. spathacea possesses various medicinal, economic, and ecological values [17]. For example, the leaves of this traditional medicinal plant have been widely used as a tonic, as well as an emmenagogue for post-partum females [18]; their juice can also cure oral thrush, flatulence, and bronchitis [19,20]. Its bark can be used as laxative and for treatment of allergies [21]. In addition, its fruits and timber are edible and can be used for wood products and construction materials, respectively [18]. Furthermore, this mangrove tree also plays a vital part in muddy sand (sediment) stabilization [18]. However, due to severe destruction of its habitat in China, D. spathacea was added to the List of Key Protected Wild Plants from Hainan Island in 2006. Unfortunately, to the best of our knowledge, there have been scarce contributions in the existing literature regarding the conservation and management of D. spathacea in Hainan Province.
Therefore, in this work, based on our latest experiences gained through field investigation of Laguncularia racemosa (L.) C. F. Gaertn. over the whole of Hainan Island [22], we aimed to evaluate the current population resource status and dynamic characteristics of the endangered mangrove species D. spathacea across Hainan Island, China in detail through a genecological analysis. We also summarize a sequence of reasonable conservation and management policies for D. spathacea resources, based on our obtained results. We cordially hope that this study, as a typical case, can provide theoretical references for the conservation, management, restoration, and rehabilitation of endangered mangrove species in the future.
2. Materials and Methods
2.1. Study Area
This research was conducted along the entire circular coastline of Hainan Island, China, spanning from 108°36′ E to 111°30′ E and from 18°80′ N to 20°10′ N (Figure 1), during July and August of 2022. Hainan Island (officially called Hainan Province in China) is located in the southernmost part of the country and neighbors Guangdong Province to the north across the Qiongzhou Strait. As the second-largest island in China, Hainan Island covers an area of approximately 3.39 × 106 ha, and is characterized by a typical tropical monsoon climate. The vast majority of the coastline terrain of Hainan Island is low and flat, but the central insular area contains large-scale mountains. Due to this special landform, the western part of the island experiences an annual average rainfall of 1600 mm, while the eastern part has an average precipitation of 2.0–2.4 × 103 mm yearly [23]. Combining the suitable annual average precipitation and temperature (around 25 °C), Hainan Island possesses extremely rich tropical and subtropical forest resources, with a stable forest cover of over 60%, and boasts one of the most impressive ecotopes in China. The 1823 km long coastline of Hainan Island is dotted with plentiful rivers and creeks that run into the ocean, creating a complicated network of estuary deltas which offers an ideal living circumstance for the existence and procreation of mangrove forests. As a consequence, Hainan Island has one of the widest distributions of mangroves in China, with the greatest species variety and the richest biodiversity [24].
In the present study, all the four cities on Hainan Island were discovered to harbor our target endangered species, D. spathacea, after our overall investigation, and the specific information of the geographical research area is exhibited in Figure 1.
2.2. Experimental Design
We chose sample quadrats for evaluation of the endangered mangrove species following the basic criteria of large space, absence of animal interference, and dominance of D. spathacea [25]. A large enough size of 20 × 20 m2 was chosen for each sample plot, due to the sparse distribution of D. spathacea trees, and 8 quadrat surveys in 12 regions of 4 cities on Hainan Island were established, in order to ensure the accuracy and universality of this investigation. The survey was carried out mainly based on the Diameter at Breast Height (DBH) of trees in all quadrats. Apart from DBH, the following specific ecological information was also noted during the investigation for every quadrat:
- (1)
- Plant species, dominant species, height, number, and canopy density;
- (2)
- City, specific distribution point, longitude, latitude, sediment type, intertidal zone condition, and interference condition.
2.3. Representations of D. spathacea Population Dynamic Characteristics
2.3.1 Population Structure
Due to the low number of the endangered D. spathacea trees on Hainan Island, it is difficult to obtain their ages accurately. Therefore, their population structure was analyzed by means of DBH, instead of temporal structure [26]. In the present work, we divided the age class of D. spathacea into 7 levels based on its DBH (Table 1). Additionally, according to Frost and Rydin (2000), the 7 levels of age classes were divided into 3 stages for the convenience of this study, among which the level I plants were considered as young individuals, levels II to IV indicated middle individuals, and levels V and above were identified as old individuals [27].
2.3.2 Analysis of Quantitative Population Dynamics
The analysis of quantitative D. spathacea population dynamics was conducted according to Shen et al. (2008) [28]. The series of formulas were as follows:
where Vx is the dynamic index of the number of individuals in the D. spathacea population from the xth to the x+1th age class; Sx and Sx+1 are the numbers of D. spathacea individuals within the xth and the x+1th age classes, respectively; Vpi is the dynamic index of the entire D. spathacea population structure; k is the number of age classes in the D. spathacea population; V’pi is the dynamic index of the entire D. spathacea population structure considering external disturbances; and Pmax is the risk probability of D. spathacea population with respect to external random disturbances. In addition, Vx∈[-1, 1], where positive, negative, and 0 values of Vx reflect the dynamic relationships of growth, decline, and stability in the number of individuals between two adjacent age classes (or the age structure of the whole population), respectively. Either Vpi or V’pi reflects a consistent dynamic relationship with Vx. Vpi is considered to have the greatest effects only when P value reaches its maximum.
2.3.3 Compilation of Static Life Table
A static life table can describe the static state of a plant population, and reflects the causes of death and change factors of different age classes of population [29]. As the target mangrove species in this research belonged to a natural population, it was inevitable that the mortality rate will be negative due to systematic sampling error, which is inconsistent with the mathematical hypothesis using the number of individuals with different age class structures to study the population dynamics within a specific time period. Therefore, many scholars use the smoothing slip technique to process the statistical data. In this study, the method of equation fitting was used to smooth the result, and the age class and the number of surviving D. spathacea were used as the independent and dependent variables to fit the equation, respectively [29]. The series of formulas were as follows:
where a0 is the actual number of survivors in the xth age class, ax is the current number of surviving individuals in the xth age class after smoothing, lx is the standardized number of surviving individuals in the xth age class, dx is the standardized number of dead individuals between two adjacent age classes, qx is the mortality rate for the xth age class, Lx is the average number of surviving individuals from the xth to the x+1th age class, Tx is the total number of surviving individuals from the xth to the higher xth age class, ex is the life expectancy of an individual in the xth age class, Kx is the vanishing rate for the xth age class, and Px is the survival rate for the xth age class.
2.3.4 Drawing of Survival Curve
The survival curve, which is drawn with the age class as the horizontal coordinate and the pair value (lnan) of surviving individuals as the vertical coordinate, can describe the death rate at different age stages according to the number of deaths of individual organisms, and can directly indicate the survival state and the changing trend of population dynamics [30]. In this study, the equations Nx = N0 e-bx (where Nx is the value of lnlx, x is the diameter class, and N0 and b are constants) and Nx = N0 x-b (where Nx is the value of lnlx, x is diameter class, and N0 and b are constants) were used to verify the type of survival curve of D. spathacea, according to Deevey’s theory [31].
2.3.5 Compilation of Population Survival Analysis Functions
In order to visually display the dynamic characteristics of the D. spathacea population, the survival rate function (S(i)), the cumulative mortality rate function (F(i)), the mortality density function (f(t)), and the hazard rate function (λ(t)) were used for analysis of its survival status [32]. The associated formulas are as follows:
where Px is the survival rate of the xth age class, Sx is the number of D. spathacea individuals within the xth age class, and ℎx is the length of the interval.
2.3.6 Analysis of Time-Series
In order to understand the future change trend of each age class of the D. spathacea population, the survival numbers after 2, 4, and 6 age classes were predicted using the one-time moving average method [28]. The formula is as follows:
where Mn is the moving average of the nth cycle, which is the number of individuals in the xth age class group after n age classes; n is the age classes that need to be predicted; x is the age class; and Dx is the population size of the xth age class.
2.4. Description of Statistical Analysis
All the obtained data were treated using Excel (version 2016), and all the figures in this work were created using Origin (version 2020). All the DBH data were calculated for at least three replicates.
3. Results
3.1. Basic Information and Current Distribution Status of D. spathacea on Hainan Island, China
After the detailed investigation of wild D. spathacea across Hainan Island, we found that it was only discovered in the cities of Wenchang, Wanning, and Sanya (Table 2). D. spathacea presented a state of overall dispersion, local concentration, and occasionally sporadic existence on Hainan Island. In addition, we also discovered that it was mainly distributed at the edge of mangrove forests, in land areas that could be flooded or completely unaffected by tides during high tides (Table 2). The altitude for D. spathacea survival on Hainan Island was 2.5–10.2 m (Table 2). The number and canopy density of D. spathacea on Hainan Island were 284 (in total) and 0.5–0.9 (Table 2), and its largest DBH and height were 48 cm and 13 m, respectively. The dominant tree species in the D. spathacea community on Hainan Island included D. spathacea, Heritiera littoralis Dryand., Cocos nucifera L., Rhizophora stylosa Griff., B. sexangula, Sonneratia caseolaris (L.) Engler, Rhizophora apiculata Bl., Ceriops tagal (Perr.) C. B. Rob., Sonneratia paracaseolaris Ko, E. Y. Chen et W. Y. Chen, Xylocarpus granatum J. Koenig, Talipariti tiliaceum (L.) Fryxell, Areca catechu L., N. fruticans, Lumnitzera racemosa Willd., and Sonneratia alba Sm. in Ress (Table 2). The sediment type characterizing the habitat of D. spathacea was simply composed of sandy and silty, while the disturbance types were complicated, including aquaculture, dam, roadblock, and deforestation (Table 2).
Moreover, D. spathacea on Hainan Island could be divided into four populations. Specifically, the population in Wenchang City was located along the west coast of Bamen Gulf, between its mangroves and seven surrounding villages (Table 2); the population in Wanning City was located in Qiexin Estuary; and the two populations in Sanya City were located in the Qingmei Harbor Mangrove Reserve and Tielu Harbor Mangrove Reserve, respectively. Notably, these two latter D. spathacea populations were geographically isolated.
3.2. Age Class Structure Analysis of D. spathacea Population
Among the investigated 284 D. spathacea trees on Hainan Island, their age classes were as follows: There were 58, 188, and 38 young, middle, and old individuals, accounting for 20.42%, 66.20%, and 13.38% of the population, respectively (Figure 2). Specifically, the numbers of young individuals in the D. spathacea populations of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) only accounted for 29.61% (45), 0% (0), 8.16% (4), and 24.32% (9), while the numbers of middle individuals accounted for up to 56.58% (86), 76.90% (35), 83.67% (41), and 70.27% (26) in the four above-mentioned populations (Figure 2). The numbers of old individuals in these D. spathacea populations were the fewest, occupying only a proportion of 13.82% (21), 23.91% (11), 8.16% (4), and 5.41% (2), respectively (Figure 2).
3.3. Quantitative Population Dynamic Analysis of D. spathacea Population
The dynamic index of the D. spathacea population was used to evaluate the variation characteristics of its different age classes on Hainan Island more accurately. As shown in Table 3, the data ranges of V1 to V7 for the four D. spathacea populations in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) on Hainan Island were irregular, suggesting that there were large fluctuations between age classes in the process of population development (Table 3). Furthermore, the V1 values for the four D. spathacea populations were all negative, suggesting that all four of the D. spathacea populations exhibited a decline in age class I; namely, there was a large loss of D. spathacea seedlings at the initial stage (Table 3). In addition, negative values also appeared for V5 of the Bamen Gulf (Wenchang) population and V2 of the Qiexin Estuary (Wanning) population, indicating that the D. spathacea populations in these stages also exhibited similar decreasing trends (Table 3).
Furthermore, the values of Vpi for the four D. spathacea populations in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) on Hainan Island were 27.46%, 38.22%, 38.55%, and 38.55%, respectively. Considering the potential external random disturbances, the values of V’pi accounted for 0.65%, 5.46%, 2.75%, and 2.75%, indicating that the growth of the D. spathacea populations was severely limited at these stages (Table 3). Fortunately, both Vpi and V’pi presented positive values, suggesting that these four populations possessed an overall potential to grow (Table 3). The values of V’pi were much lower than those of Vpi, and all the Pmax values were positive, indicating that all four D. spathacea populations on Hainan Island showed sensitivity to external random disturbances, which could thus have serious impacts on the D. spathacea population (Table 3).
3.4. Analysis of Static Life Table of D. spathacea Population
As can be seen from the static life table of D. spathacea, all four of the D. spathacea population structures exhibited significant fluctuation (Table 4). In particular, we discovered that, after standardized smoothing treatment, the number of surviving individuals (lx) gradually slowed down and levelled off with an increase in the age class. In addition, we found the trend in the data for ax was consistent with that of lx. The peak values of the mortality rate (qx) and vanishing rate (Kx) were all observed in the highest age classes in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya). The two highest mortality rates (100.00%) of D. spathacea populations both occurred in Sanya City (Table 4). In addition, all individual life expectancy (ex) values were relatively small in the four D. spathacea populations, suggesting that the population might be continually exposed to great fluctuations (Table 4).
3.5. Survival Curve Analysis of D. spathacea Population
The survival curve of D. spathacea population on Hainan Island was drawn with the age class as horizontal coordinate and lglx as the vertical coordinate, and the results of fitting equations and verifications (R²) are shown in Table 5. All of the survival curves for the D. spathacea populations in the Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) areas tended to the Deevey II pattern, as the fitting effects of exponential function equations for them were better than those of power function equations (Table 5), indicating that the mortality rates of D. spathacea populations in these regions were generally stable across the age classes.
Moreover, the mortality and vanishing rates for the D. spathacea populations in the Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) areas were basically consistent, showing a similar trend of first decreasing and then increasing, suggesting that D. spathacea populations were more susceptible to death at the young and old age stages (Figure 3), In addition, the vanishing rates were obviously higher than the mortality rates for each age class of the four D. spathacea populations (Figure 3).
3.6. Population Survival Analysis of D. spathacea Population
In terms of the survival rate function and the cumulative mortality rate function of the D. spathacea populations on Hainan Island, they exhibited two exactly contradictory trends (Figure 4). In particular, the survival rate functions of the D. spathacea populations in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) were decreasing, while the cumulative mortality rate functions increased continuously with the growth of their age classes (Figure 4). The intersection point of these two curves appeared around age class II and, interestingly, the values at the intersections for all four of the D. spathacea populations were 50% (Figure 4).
The mortality density function and hazard rate function of the D. spathacea populations on Hainan Island were also explored. Similar to the survival rate function and the cumulative mortality rate function, these two curves in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) also presented an opposite trend, where the former decreased while the latter increased dramatically (Figure 5). It is worth mentioning that the early slopes of the two functions were higher than the late ones (Figure 5), indicating that the life state of the D. spathacea population was unstable in the young age, with a significant number of individuals dying; however, as they grew older, the mortality risk gradually declined.
3.7. Analysis of Age Class Time-Series of D. spathacea Population
For the age class time-series of the four D. spathacea populations, we observed that the initial numbers in the Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) areas were visibly irregular (Table 5). After going through the next predicted 2, 4, and 6 age classes, the elderly individuals in the D. spathacea populations were supplemented, to a certain extent (Table 5); however, due to the lack of young individuals in the areas of Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya), the number of middle-aged individuals decreased and the D. spathacea population renewal potential was insufficient. Eventually, this would lead to an obvious decline.
4. Discussion
4.1. Analysis of D. spathacea Population Structure
It is generally accepted that mangroves play key roles in coastal wetland ecosystems in tropical and subtropical areas, such as coastal protection and maintenance of coastal ecosystem function [1]. However, due to human interference (e.g., urban development and aquaculture), mangrove deforestation has occurred in an endless stream, causing the acreage of mangrove forests in the world to decrease by 30–50% over the last century [11,12]. As a result, a large number of mangrove species are globally in the process of becoming endangered species, and they may even face extinction. In the present study, we tried to evaluate the current resource status and the population dynamic characteristics of an endangered semi-mangrove species, D. spathacea, on Hainan Island, China, in order to propose sustainable conservation and management strategies for future ecological restoration of its natural population. This research may allow us to better understand how to effectively protect, manage, and rehabilitate endangered mangrove species in the future.
The population structure can reflect the current situation and the development direction, which plays an important role in analyzing the history and predicting the future dynamics of a population [33]. In the present study, we first found the number of seedlings was significantly higher than that in the larger-age classes; furthermore, the D. spathacea population structure was mainly composed of young individuals in age classes I, II, and III (0 cm ≤ DBH ≤ 12.5 cm), which could survive in the understory of the forest community, suggesting that the seed germination and seedling growth of D. spathacea could be well-adaptively completed in a shaded situation. This result was particularly similar to that of another endangered terrestrial tree species—Taxus cuspidata Siebold et Zucc—in China reported by Liu et al. (2023) [34]. Additionally, all four of the D. spathacea populations possessed a large number of young individuals and 3–4 species of other dominant trees in the community, implying that competition for light, water, and nutrients might exist within and among the various species. Indeed, our previous study on the ecological patterns of three kinds of terrestrial medicinal plants in Altay Prefecture, China, also found that competition was one of the most important factors that crucially determined the ecological niche of a species [35]. In this work, the higher D. spathacea population mortality rate in the early age class may be a result of the enhanced intraspecific and interspecific competition. Based on these deductions, it was no wonder that the young seedlings could not grow into medium- and large-sized trees, leading to their ultimate death. Except for the effects of drastic competition, it was certainly the case that the higher mortality rate of D. spathacea population in its initial age class made it insufficient to provide enough young individuals to supplement the older age classes, thus affecting the renewal and development of its population. Moreover, the small plant population was found to be extremely sensitive to changes in the external environment, making it unable to maintain long-term stability and, so, the D. spathacea population might stay low [36]. As for the older age class, the populations of D. spathacea in the Bamen Gulf (Wenchang) and Qingmei Harbor (Sanya) areas presented fluctuations, indicating that there might be strong anthropogenic disturbances in these two regions which caused them to deviate from a stable population structure. During the period from the 1970s to 1980s, unlawful logging for construction and manufacturing were common, particularly targeting higher age class mangrove plants [13]. These behaviors had severely detrimental influences on the stand type and structure of mangrove forests.
4.2. Analysis of D. spathacea Population Dynamics
After analysis of theD. spathacea population dynamic index, we found that there were large fluctuations between age classes in the process of population development. Furthermore, the V1 values for the four D. spathacea populations on Hainan Island were all negative, suggesting a large loss of D. spathacea seedlings at the initial stage, which was in keeping with our analysis of the mortality and vanishing rate curves. The values of V’pi were much lower than those of Vpi, and all the Pmax values were positive, indicating that all four D. spathacea populations on Hainan Island showed sensitivity to external random disturbances, which could have serious impacts on the D. spathacea population. These results were broadly similar to those of a previous study on T. cuspidate [34].
From the results of the static life table and other dynamic indices for the D. spathacea population, we discovered that the D. spathacea population experienced two fluctuations in its growth cycle: one in the juvenile stage and the other during the mature stage. The first fluctuation in the immature stage of the D. spathacea population may be mainly caused by their weak vitality and the intraspecific and interspecific competition for light, water, and nutrients. As for the second fluctuation, emerging in the mature stage, we propose that it may be attributed to anthropogenic interferences with high probability. This speculation can be summarized according to two main evidential aspects. On one hand, all the survival curves for the age classes of the four D. spathacea populations displayed a standard Deevey II type, characterized by an exponential curve, implying that their mortality rates were significantly stable throughout the process of population development, even in the last age class. This indicated that external factors could obviously affect the population development of D. spathacea [34]. On the other hand, the reflection of the irregular initial number of D. spathacea trees could correspond to our hypothesis of unsustainable deforestation. In general, a healthy forest population would present a more balanced distribution of age classes, similar to the predicted time-series, with a gradual decrease in number with enhancement of the age class [34].
4.3. Recommendations of Conservation and Management for D. spathacea Population
Based on the above discussion, we propose some recommendations for the sustainable conservation and management of this endangered semi-mangrove species in China: (1) In situ conservation and expanding D. spathacea trees: We must make sure that there are enough viable seeds or seedlings being naturally produced. Considering the geographical isolation among the four D. spathacea populations on Hainan Island, so we should encourage endogenous gene exchange through sexual reproduction as much as possible, increase genetic diversity within the population and, thus, enhance population stability. Kramer et al. (2008) reported that the detrimental influences of serious habitat fragmentation can be partially offset by frequent gene exchange between internal populations [37]. (2) Preserving germplasm resources: A diverse range of genetic materials such as pollens, seeds, and tissues should be stored to form a source of genetic diversity that can serve the purpose of gene exchange in the future. (3) Rehabilitating the habitats to promote provenance restoration: We observed various external disturbances, such as aquaculture, dams, and roadblocks, affecting the D. spathacea populations on Hainan Island. There is no doubt that these disturbances impose serious restrictions on their development and, hence, we should return ponds to forests and reinforce habitat restoration to provide favorable conditions for their recovery and growth. (4) Artificial cultivation and spreading planting: Last, but not least, we should plant more of this species in its local land or other suitable planting regions in order to enlarge its population quantity, as there are no obstacles to the anthropogenic propagation and breeding of D. spathacea in China [17].
5. Conclusions
D. spathacea is a typical endangered semi-mangrove species in China that needs to be protected and managed urgently. In this research, our findings indicated only four main D. spathacea populations, distributed in the areas of Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya), on Hainan Island, China. These populations presented a state of overall dispersion, local concentration, and occasionally sporadic existence, and existed in geographical isolation. The number of young, middle, and old D. spathacea plants accounted for 20.42%, 66.20%, and 13.38% of the population, respectively, suggesting that the age class distribution of the D. spathacea population on Hainan Island belongs to a decline type. In addition, the analysis of D. spathacea population dynamic characteristics directly indicated that the D. spathacea population on Hainan Island was sensitive to external disturbances and possessed two main increases in mortality rate—namely, in the juvenile and mature stages—due to weak vitality, competition, and anthropogenic interferences, which are the most important reasons for its endangered status. Hence, in order to protect D. spathacea resources, we should conserve and expand the mature D. spathacea trees in situ, preserve its germplasm resources, rehabilitate its habitats to promote provenance restoration, and conduct artificial cultivation and spreading planting, in order to realize the sustainable conservation and management of D. spathacea.
Author Contributions
M.T., T.L. and H.Z. conceived of the original research project and selected methods. M.T., X.K., M.L., K.D., Y.Y., Z.F., C.Z., S.L. and Z.Z. performed most of the experiments. N.F.-y.T., T.L. and H.Z. supervised the experiments and provided technical assistance to M.T., X.K., M.L., K.D., Y.Y., Z.F., C.Z., S.L. and Z.Z. M.T. and T.L. wrote the article. H.Z. and N.F.-y.T. refined the project and revised the writing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was jointly supported by the Hainan Province Science and technology Special Fund (ZDYF2021SHFZ259), the Foundation for the Scientific Research Base and Distinguished Talents in Guangxi (Guike AD22035015; Guike AC22080006), the National Natural Science Foundation of China (32101367), the Guangdong Basic and Applied Basic Research Foundation (2022A1515010698), the Science, Technology, and Innovation Commission of Shenzhen Municipality (JCYJ20220818095601003; KCXFZ20230731092801002), and the Guangxi First-class Disciplines (Agricultural Resources and Environment, BSUFCD-KF202402).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alongi, D.M. Impact of Global Change on Nutrient Dynamics in Mangrove Forests. Forests 2018, 9, 596. [Google Scholar] [CrossRef]
- Bayen, S. Occurrence, bioavailability and toxic effects of trace metals and organic contaminants in mangrove ecosystems: A review. Environ. Int. 2012, 48, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Feller, I.; Lovelock, C.; Berger, U.; McKee, K.; Joye, S.; Ball, M. Biocomplexity in Mangrove Ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef]
- Lacerda, L.D. Mangrove ecosystems: function and management. Springer Verlag, Berlin. 2002.
- Wang, Y.S. Molecular ecology of mangroves. Beijing: Science Publishing Press. in Chinese. 2019.
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological role and services of tropical mangrove ecosystems: a reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Ward, R.D.; Friess, D.A.; Day, R.H.; MacKenzie, R.A. Impacts of climate change on mangrove ecosystems: a region by region overview. Ecosyst. Health Sustain. 2016, 2, e01211. [Google Scholar] [CrossRef]
- Carugati, L.; Gatto, B.; Rastelli, E.; Martire, M.L.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef]
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proc. Natl. Acad. Sci. USA 2016, 113, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Duke, N.C.; Meynecke, J.-O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; Koedam, N.; Lee, S.Y.; Marchand, C.; Nordhaus, I.; Dahdouh-Guebas, F. A world without mangroves? Science 2007, 317, 41–42. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Wang,W. ; Wang, M. The mangroves of China; Science Press: Beijing, China, 2007. [Google Scholar]
- Lin, P. A review on the mangrove research in China. J. Xiamen Univ. (Natural Sci.) 2001, 40, 592–603. [Google Scholar]
- Mahesh, G.; Patil, K.H. Dynamics of seedling distribution of Dolichandrone spathacea(L.f.)K.Schum. A proceeding of national conference on recent advances in physics and natural sciences. Maharashtra, India: Karmaveer Bhaurao Patil College, Urun, Islampur, 2018.
- Suman, D.O. Mangrove management: Challenges and guidelines. In coastal wetlands: An integrated ecosystem approach, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Tian, G.H.; Li, M.; Yang, X.B.; Liao, B.W.; Liu, W.H.; Lei, Z.S. Study on Introduction of Dolichandrone spathacea on Qi-ao Island. J. Anhui Agr. Sci. 2011, 39, 9618–9619. [Google Scholar]
- Nguyen, V.T.; Do, L.Q.; Nguyen, T.A.; Nguyen, T.T.; Ho, N.A.; Tran, V.C.; Tran, T.P.T. New cycloartanes and new iridoids from Dolichandrone spathacea collected in the mangrove forest of Soc Trang province, Vietnam. J. Asian Nat. Prod. Res. 2017, 20, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Chandra Prasad, P.R.; Reddy, C.; Redy, S.H.; Dutt, C.B.S. Folkore medicinal plants of North Andaman Island, India. Fitoterapia 2008, 79, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Wiart, C. Medicinal plants of the Asia-Pacific: Drugs for the future? WSP Singapore, Singapore, 2006.
- Ho, P.H. Medicinal trees in Vietnam; Young Publishers: Ho Chi Minh, Vietnam, 2006. [Google Scholar]
- Cheng, C.; Ke, X.; Lang, T.; Zhong, C.; Lv, X.; Zhang, M.; Chen, Y.; Fang, Z.; Zhou, H.; Chen, Y. Current Status and Potential Invasiveness Evaluation of an Exotic Mangrove Species, Laguncularia racemosa (L.) C.F. Gaertn, on Hainan Island, China. Forests 2023, 14, 2036. [Google Scholar] [CrossRef]
- Hsieh, C.M.; Zhong, G.F. Hainan–The island of South Sea. A new province in China. Geosci. J. 1990, 20, 385–391. [Google Scholar]
- Li, M.; Lee, S. Mangroves of China: a brief review. For. Ecol. Manag. 1997, 96, 241–259. [Google Scholar] [CrossRef]
- Wang, B.; Yu, S. Handbook of Plant Community Experiments; Guangdong Education Press: Guangzhou, China, 1996. [Google Scholar]
- Hong, W.; Liu, J.; Wu, C.Z. A study on structure and spatial distribution pattern of Castanopsis hystrix population Sci. Silvae Sin. 2001, 31, 6–10, (In Chinese with English abstract). [Google Scholar]
- Frost, I.; Rydin, H. Spatial patem and size distribution of the anima-dispersed tree Quercus robur in two spruce-dominated forests. Ecoscience, 2000, 7, 38–44. [Google Scholar] [CrossRef]
- Shen, S.; Ma, H.; Wang, Y.; Wang, B.; Shen, G. Structure and dynamics of natural populations of the endangered plant Euryodendron excelsum HT Chang. Front. Forest. China, 2009, 4, 14–20. [Google Scholar] [CrossRef]
- Farahat, E.A. Age structure and static life tables of the endangered Juniperus phoenicea L. in North Sinai Mountains, Egypt: implication for conservation. J. Mt. Sci. 2020, 17, 2170–2178. [Google Scholar] [CrossRef]
- Hett, J.M.; Loucks, O.L. Age Structure Models of Balsam Fir and Eastern Hemlock. J. Ecol. 1976, 64, 1029. [Google Scholar] [CrossRef]
- Deevey, E.S. Life Tables for Natural Populations of Animals. Q. Rev. Biol. 1947, 22, 283–314. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z. (. A unified survival-analysis approach to insect population development and survival times. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Harcombe, P.A. 1987; 37.
- Liu, D.; Guo, Z.; Cui, X.; Fan, C. Estimation of the Population Dynamics of Taxus cuspidata by Using a Static Life Table for Its Conservation. Forests 2023, 14, 2194. [Google Scholar] [CrossRef]
- Lang, T.; Pan, L.; Liu, B.; Guo, T.; Hou, X. Vegetation Characteristics and Response to the Soil Properties of Three Medicinal Plant Communities in Altay Prefecture, China. Sustainability 2020, 12, 10306. [Google Scholar] [CrossRef]
- Pala, N.A.; Negi, A.K.; Gokhale, Y.; Bhat, J.A.; Todaria, N.P. Diversity and regeneration status of Sarkot Van Panchyat in Garhwal Himalaya, India. J. For. Res. 2012, 23, 399–404. [Google Scholar] [CrossRef]
- Kramer, A.T.; Ison, J.L.; Ashley, M.V.; Howe, H.F. The Paradox of Forest Fragmentation Genetics. Conserv. Biol. 2008, 22, 878–885. [Google Scholar] [CrossRef]
Figure 1.
A combination figure showing the geographical research area of the forests of D. spathacea Hainan Island, China. ① Distribution of D. spathacea in Touyuan Village; ②distribution of D. spathacea in Xiachang Village; ③ distribution of D. spathacea in Danchang Village; ④ distribution of D. spathacea in Xibian Village; ⑤ distribution of D. spathacea in Liangfeng Village; ⑥ distribution of D. spathacea in Haitou Village; ⑦ distribution of D. spathacea in Sanduo Village; ⑧ distribution of D. spathacea in Jiaxin Estuary; ⑨ distribution of D. spathacea in Qingmei Harbor; ⑩ distribution of D. spathacea in Tielu Harbor.
Figure 1.
A combination figure showing the geographical research area of the forests of D. spathacea Hainan Island, China. ① Distribution of D. spathacea in Touyuan Village; ②distribution of D. spathacea in Xiachang Village; ③ distribution of D. spathacea in Danchang Village; ④ distribution of D. spathacea in Xibian Village; ⑤ distribution of D. spathacea in Liangfeng Village; ⑥ distribution of D. spathacea in Haitou Village; ⑦ distribution of D. spathacea in Sanduo Village; ⑧ distribution of D. spathacea in Jiaxin Estuary; ⑨ distribution of D. spathacea in Qingmei Harbor; ⑩ distribution of D. spathacea in Tielu Harbor.
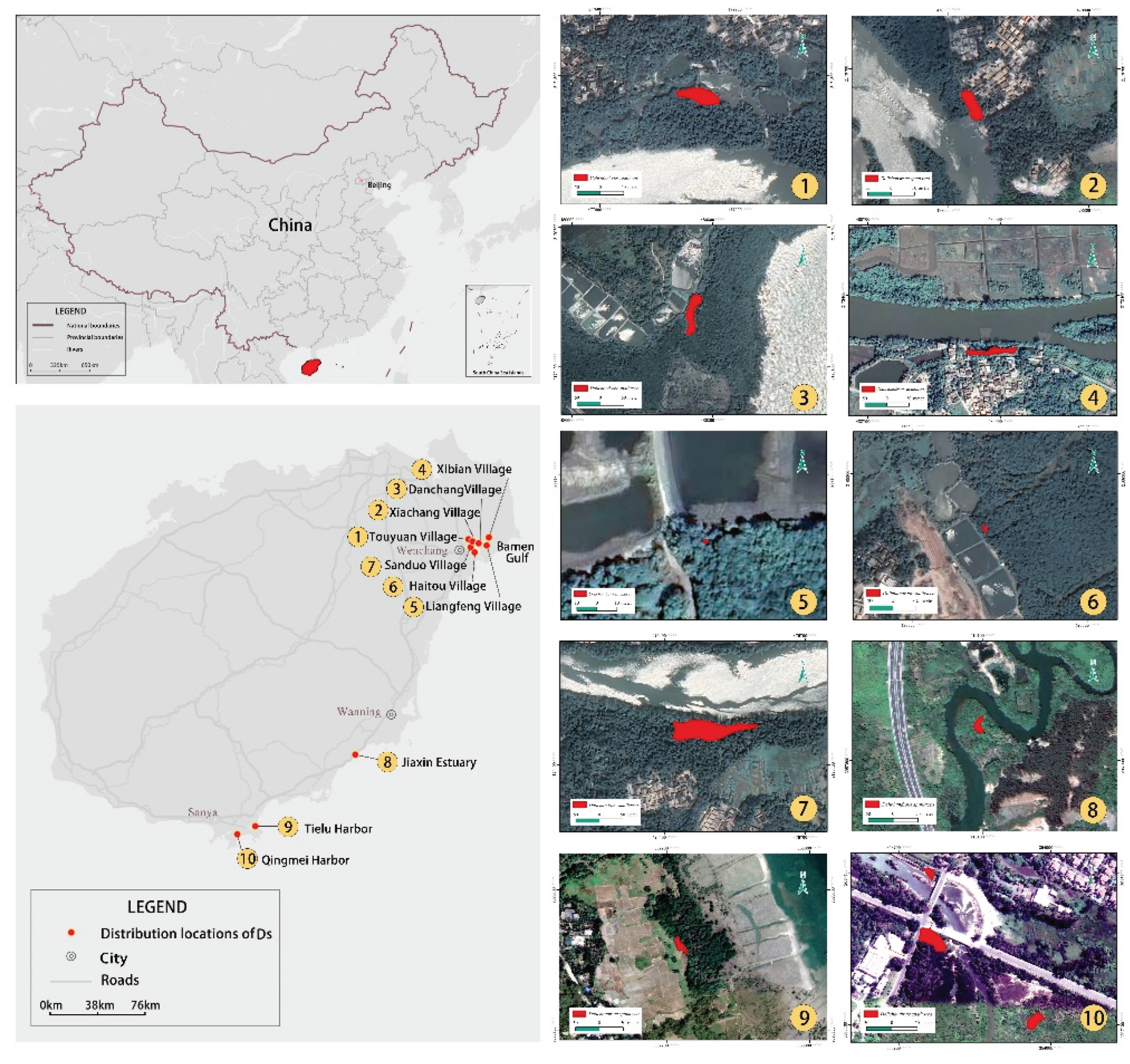
Figure 2.
Age class structures of D. spathacea populations in Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) on Hainan Island, China.
Figure 2.
Age class structures of D. spathacea populations in Bamen Gulf (Wenchang), Qiexin Estuary (Wanning), Qingmei Harbor (Sanya), and Tielu Harbor (Sanya) on Hainan Island, China.
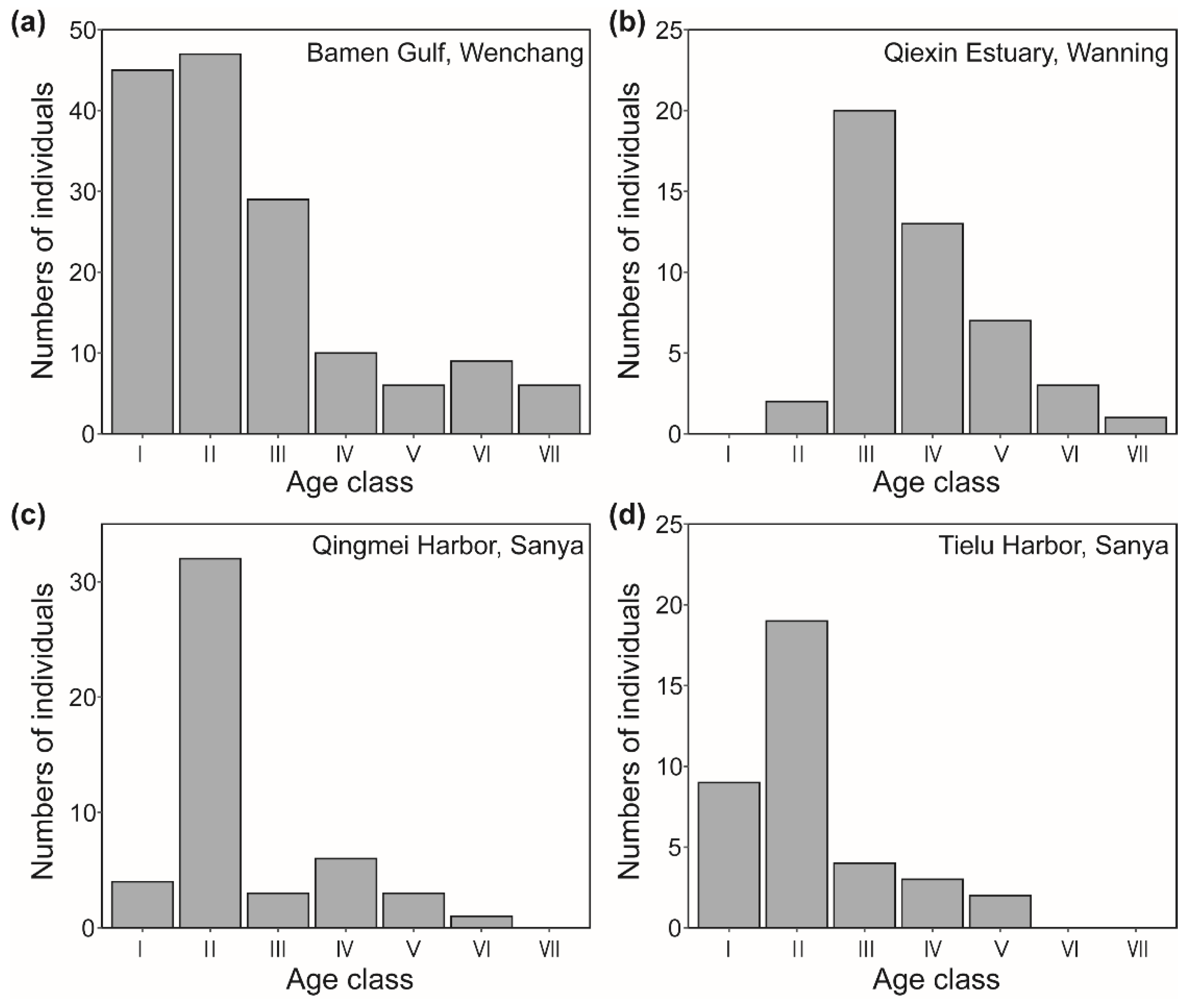
Figure 3.
Curves of the mortality and vanishing rates of the D. spathacea populations on Hainan Island, China.
Figure 3.
Curves of the mortality and vanishing rates of the D. spathacea populations on Hainan Island, China.
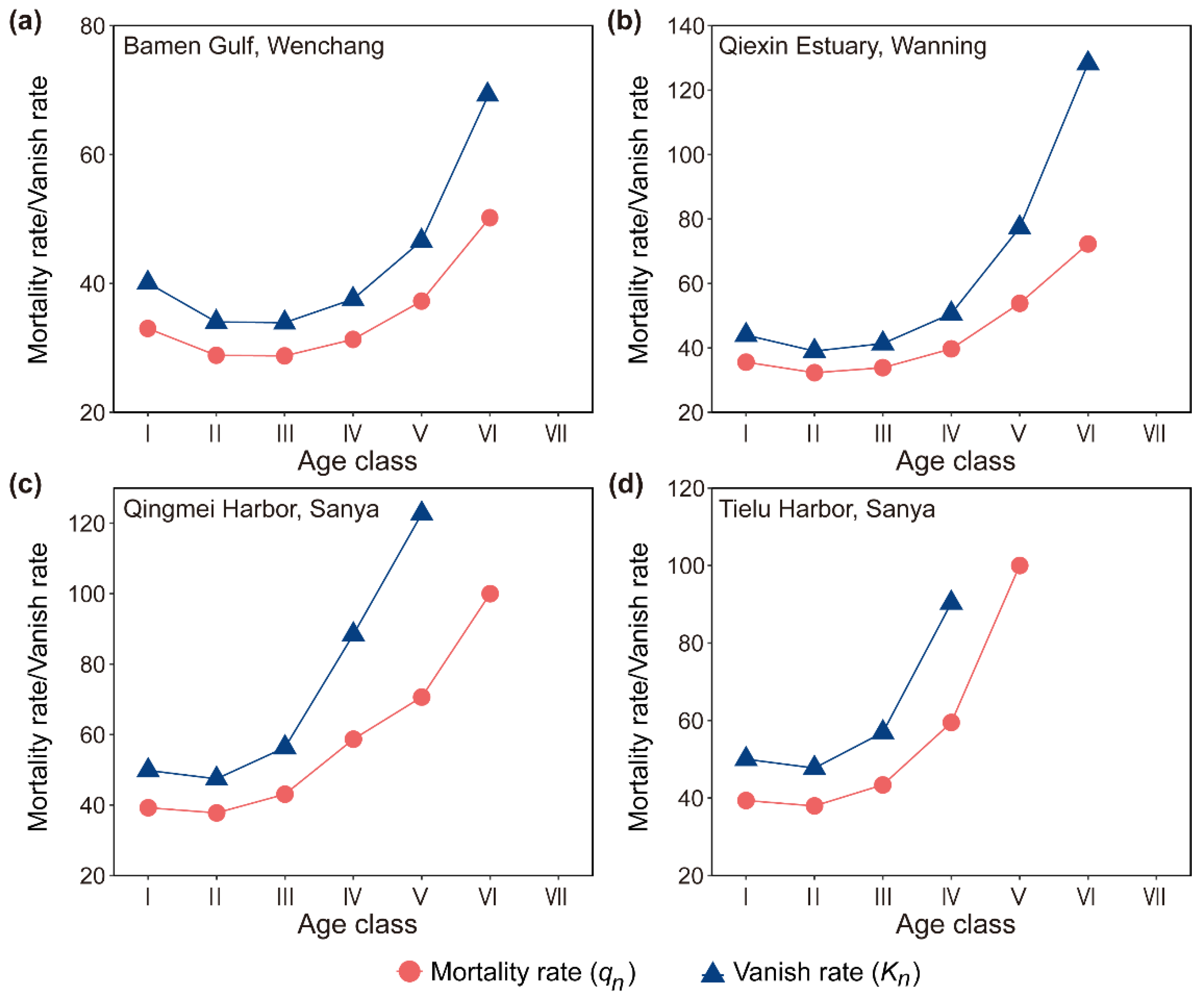
Figure 4.
Curves of the survival rate function and the cumulative mortality rate function of D. spathacea population on Hainan Island, China.
Figure 4.
Curves of the survival rate function and the cumulative mortality rate function of D. spathacea population on Hainan Island, China.
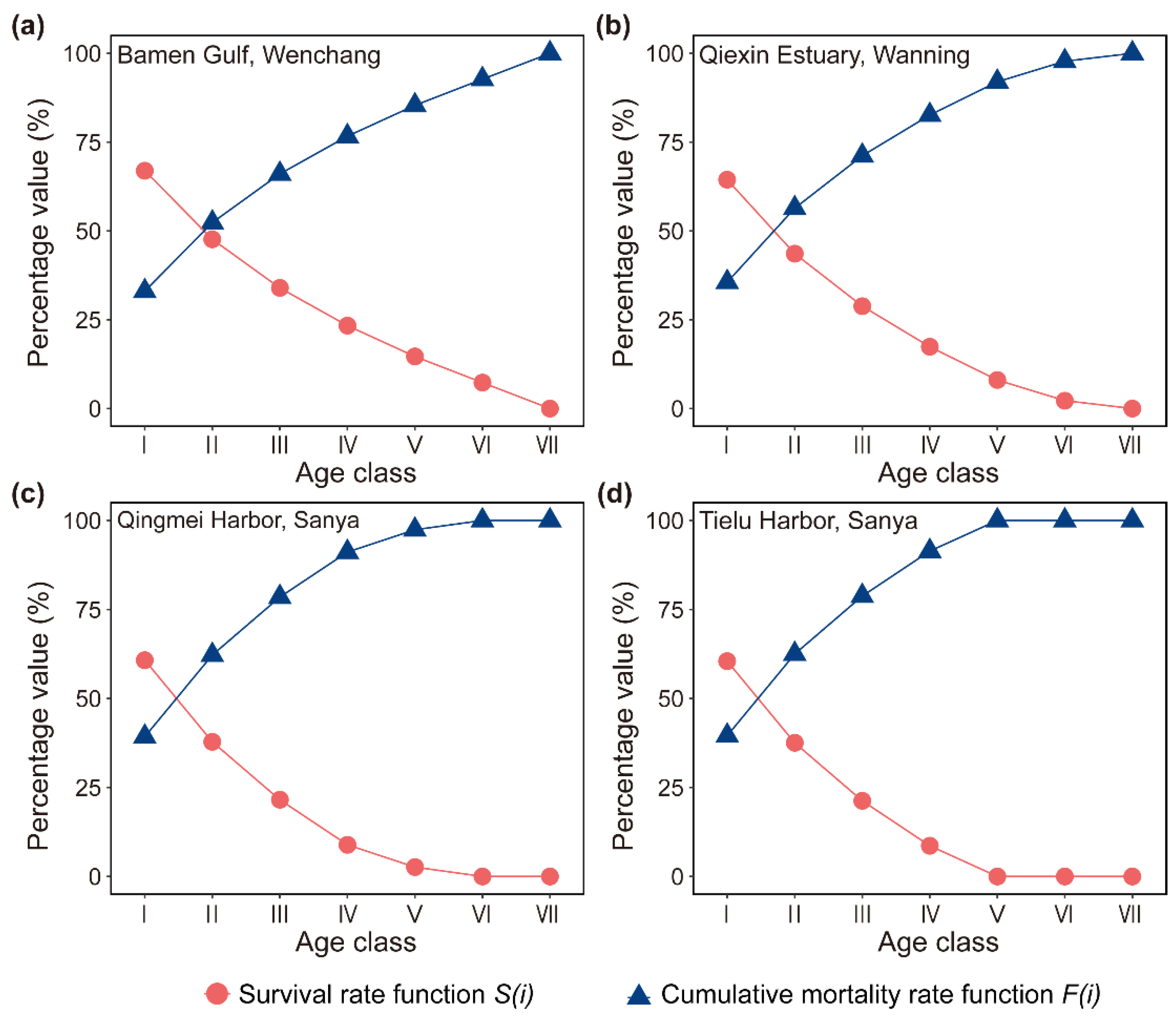
Figure 5.
Curves of the mortality density function and the hazard rate function of D. spathacea population on Hainan Island, China.
Figure 5.
Curves of the mortality density function and the hazard rate function of D. spathacea population on Hainan Island, China.
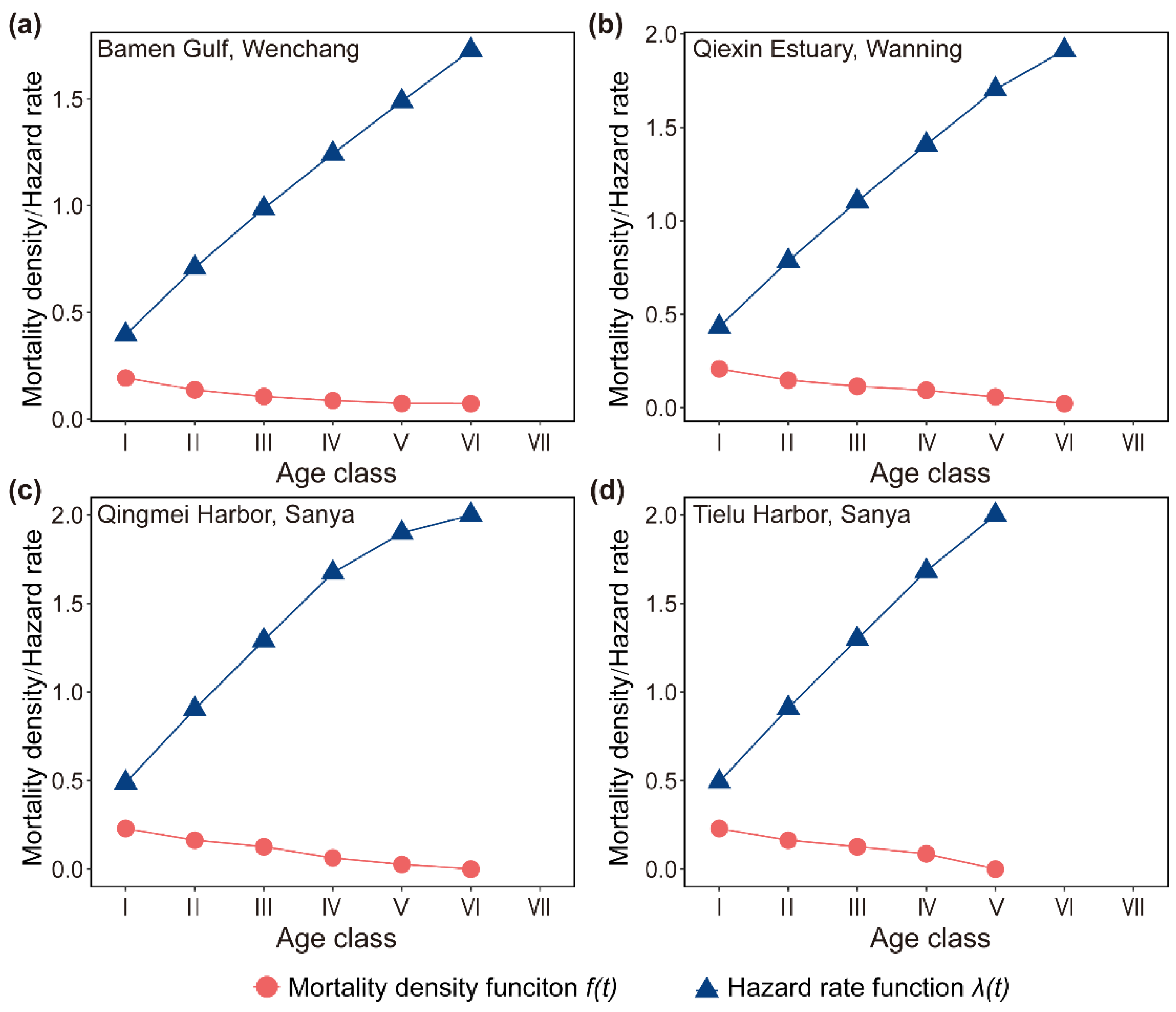

Table 1.
Definition of age class levels of D. spathacea based on its DBH on Hainan Island, China.
| DBH range | Level |
| DBH ≤ 2.5 cm | I |
| 2.5 cm < DBH ≤ 7.5 cm | II |
| 7.5 cm < DBH ≤ 12.5 cm | III |
| 12.5 cm < DBH ≤ 17.5 cm | IV |
| 17.5 cm < DBH ≤ 22.5 cm | V |
| 22.5 cm < DBH ≤ 27.5 cm | VI |
| DBH > 27.5 cm | VII |
Table 2.
Basic information and current distribution status of D. spathacea on Hainan Island, China.
| City | Distribution Point | Longitude | Latitude | Altitude (m) |
Quantity | Canopy Density | Dominant Tree Species | Tide Situation | Sediment Type | Disturbance Type | |
| Wenchang | Bamen Gulf | Touyuan Village | 110°47′19″E | 19°37′42″N | 2.5 | 13 | 0.9 | S. caseolaris, B. sexangula | Spring Tides | Silty | Dam |
| Xiachang Village | 110°47′43″E | 19°37′22″N | 6.5 | 24 | 0.7 |
S. caseolaris, B. sexangula, S. paracaseolaris |
Spring Tides | Sandy and Silty | Roadblock | ||
| Danchang Village | 110°48′42″E | 19°37′18″N | 3.2 | 36 | 0.8 |
R. stylosa, B. sexangula |
Spring Tides | Silty | Dam | ||
| Xibian Village |
110°53′40″E | 19°38′50″N | 10.2 | 17 | 0.7 |
C. nucifera, S. caseolaris, H. tiliaceus |
No Tides | Silty | Aquaculture | ||
| Liangfeng Village | 110°51′34″E | 19°37′43″N | 3.5 | 1 | 0.5 | R. apculata, C. tagal | No Tides | Sandy and Silty | Dam | ||
| Haitou Village | 110°47′46″E | 19°36′03″N | 2.6 | 2 | 0.6 | R. stylosa, X. granatum | No Tides | Sandy and Silty | Dam | ||
| Sanduo Village | 110°46′32″E | 19°37′17″N | 3.4 | 59 | 0.9 | H. littoralis, D. spathacea, C. nucifera | Spring Tides | Silty | Aquaculture | ||
| Wanning | Jiaxin Estuary | 110°10′36″E | 18°36′02″N | 3.5 | 46 | 0.9 |
D. spathacea, A. catechu, N. fruticans |
No Tides | sandy and Silty | Desertification | |
| Sanya | Tielu Harbor | 109°42′07″E | 18°15′47″N | 4.2 | 37 | 0.7 |
R. apculata, S. alba, L. racemosa |
Spring Tides | Sandy | – | |
| Qingmei Harbor | 109°37′03″E | 18°13′57″N | 5.2 | 49 | 0.6 |
L. racemosa, C. tagal, R. apculata |
Spring Tides | Sandy | Roadblock | ||
Table 3.
Variation indices of D. spathacea population dynamics on Hainan Island, China.
| Distribution Point | Variation Index of D. spathacea Population Dynamics (%) | |||||||||
| V1 | V2 | V3 | V4 | V5 | V6 | V7 | Vpi | V'pi | Pmax | |
| Bamen Gulf (Wenchang) | -4.26 | 38.30 | 65.52 | 40.00 | -33.33 | 33.33 | 100.00 | 27.46 | 0.65 | 2.38 |
| Qiexin Estuary ( Wanning ) | -100.00 | -90.00 | 35.00 | 46.15 | 57.14 | 66.67 | 100.00 | 38.22 | 5.46 | 14.29 |
| Qingmei Harbor (Sanya) | -52.63 | 78.95 | 25.00 | 33.33 | 100.00 | 100.00 | 100.00 | 38.55 | 2.75 | 7.14 |
| Tielu Harbor (Sanya) | -52.63 | 78.95 | 25.00 | 33.33 | 100.00 | 100.00 | 100.00 | 38.55 | 2.75 | 7.14 |
Note: Vx is the dynamic index of the number of individuals in the population from the xth to the x+1th age class, Vpi is the dynamic index of the entire population structure, V'pi is the dynamic index of the entire population structure when external disturbances are taken into account, and Pmax is the risk probability of the D. spathacea population with respect to external random disturbances.
Table 4.
Static life table of D. spathacea population on Hainan Island, China.
| Distribution Point | Age class | ax | lx | lgax | dx | qx (%) | Lx | Tx | ex | Kx (%) | Px (%) |
| Bamen Gulf (Wenchang) | I | 52 | 1000 | 3.946 | 330 | 33.0 | 835 | 2402 | 2.402 | 40.1 | 67.0 |
| II | 35 | 670 | 3.545 | 193 | 28.8 | 573 | 1567 | 2.339 | 34.0 | 71.2 | |
| III | 25 | 477 | 3.205 | 137 | 28.8 | 408 | 994 | 2.085 | 33.9 | 71.2 | |
| IV | 18 | 339 | 2.866 | 106 | 31.3 | 286 | 586 | 1.725 | 37.6 | 68.7 | |
| V | 12 | 233 | 2.490 | 87 | 37.3 | 190 | 300 | 1.287 | 46.6 | 62.7 | |
| VI | 8 | 146 | 2.024 | 73 | 50.2 | 110 | 110 | 0.749 | 69.7 | 49.8 | |
| VII | 4 | 73 | 1.326 | – | – | – | – | – | – | – | |
| Qiexin Estuary ( Wanning ) | I | 45 | 1000 | 3.806 | 356 | 35.6 | 718 | 1943 | 1.943 | 44.0 | 64.4 |
| II | 29 | 644 | 3.366 | 208 | 32.3 | 540 | 1225 | 1.902 | 39.0 | 67.7 | |
| III | 20 | 436 | 2.976 | 148 | 33.9 | 305 | 685 | 1.572 | 41.4 | 66.1 | |
| IV | 13 | 288 | 2.562 | 115 | 39.7 | 231 | 380 | 1.319 | 50.6 | 60.3 | |
| V | 8 | 174 | 2.056 | 94 | 53.9 | 98 | 149 | 0.859 | 77.3 | 46.1 | |
| VI | 4 | 80 | 1.283 | 58 | 72.3 | 51 | 51 | 0.639 | 128.3 | 27.7 | |
| VII | 1 | 22 | 1.000 | – | – | – | – | – | – | – | |
| Qingmei Harbor (Sanya) | I | 38 | 1000 | 3.647 | 392 | 39.2 | 804 | 1803 | 1.803 | 49.8 | 60.8 |
| II | 23 | 608 | 3.149 | 230 | 37.8 | 493 | 999 | 1.644 | 47.7 | 62.2 | |
| III | 15 | 378 | 2.674 | 163 | 43.1 | 297 | 506 | 1.339 | 56.3 | 56.9 | |
| IV | 8 | 215 | 2.111 | 126 | 58.7 | 152 | 210 | 0.974 | 88.4 | 41.3 | |
| V | 3 | 89 | 1.227 | 63 | 70.7 | 57 | 57 | 0.647 | 122.7 | 29.3 | |
| VI | 1 | 26 | 0.000 | 26 | 100.0 | 13 | 13 | 0.500 | – | 0.0 | |
| VII | 0 | 0 | – | – | – | – | – | – | – | – | |
| Tielu Harbor (Sanya) | I | 24 | 1000 | 3.184 | 394 | 39.4 | 803 | 1782 | 1.782 | 50.0 | 60.6 |
| II | 15 | 606 | 2.683 | 230 | 38.0 | 491 | 979 | 1.614 | 47.7 | 62.0 | |
| III | 9 | 376 | 2.206 | 163 | 43.4 | 295 | 487 | 1.295 | 56.9 | 56.6 | |
| IV | 5 | 213 | 1.637 | 127 | 59.5 | 150 | 193 | 0.905 | 90.3 | 40.5 | |
| V | 2 | 86 | 0.734 | 86 | 100.0 | 43 | 43 | 0.500 | – | 29.3 | |
| VI | 0 | 0 | – | 0 | – | 0 | 0 | – | – | – | |
| VII | 0 | 0 | – | – | – | – | – | – | – | – |
Note:ax is the current number of surviving individuals in the xth age class after smoothing treatment, lx is the standardized number of surviving individuals in the xth age class, dx is the standardized number of dead individuals between two adjacent age classes, qx is the mortality rate for the xth age class, Lx is the average number of surviving individuals from the xth to the x+1th age class, Tx is the total number of surviving individuals from the xth to the higher age classes, ex is the life expectancy of an individual in the xth age class, Kx is the vanishing rate for the xth age class, and Px is the survival rate for the xth age class.
Table 5.
Survival curves and their verification models of D. spathacea population on Hainan Island, China.
Table 5.
Survival curves and their verification models of D. spathacea population on Hainan Island, China.
| Distribution Point | Nx = N0 e-bx | Nx = N0 x-b |
| Bamen Gulf (Wenchang) | Nx = 5.0355e-0.16x, R² = 0.9235 | Nx = 4.6354x-0.457, R² = 0.7586 |
| Qiexin Estuary ( Wanning ) | Nx =4.986e-0.19x, R² = 0.9314 | Nx = 4.3919x-0.491, R² = 0.7819 |
| Qingmei Harbor (Sanya) | Nx = 5.435e-0.281x, R² = 0.887 | Nx = 4.295x-0.633, R² = 0.7297 |
| Tielu Harbor (Sanya) | Nx = 5.3633e-0.357x, R² = 0.8766 | Nx = 3.9627x-0.301, R² = 0.7149 |
Table 5.
Time-series prediction of D. spathacea population on Hainan Island, China.
| Age class | Bamen Gulf (Wenchang) | Qiexin Estuary ( Wanning ) | Qingmei Harbor ( Sanya ) | Tielu Harbor (Sanya) | ||||||||||||
| Initial Number | M2 | M4 | M6 | Initial Number |
M2 | M4 | M6 | Initial Number | M2 | M4 | M6 | Initial Number | M2 | M4 | M6 | |
| I | 45 | – | – | – | 0 | – | – | – | 4 | – | – | – | 9 | – | – | – |
| II | 47 | 46 | – | – | 2 | 1 | – | – | 32 | 18 | – | – | 19 | 14 | – | – |
| III | 29 | 38 | – | – | 20 | 11 | – | – | 3 | 18 | – | – | 4 | 12 | – | – |
| IV | 10 | 20 | 33 | – | 13 | 17 | 9 | – | 6 | 5 | 11 | – | 3 | 4 | 9 | – |
| V | 6 | 8 | 23 | – | 7 | 10 | 11 | – | 3 | 5 | 11 | – | 2 | 3 | 7 | – |
| VI | 9 | 8 | 14 | 24 | 3 | 5 | 11 | 8 | 1 | 2 | 3 | 8 | 0 | 1 | 2 | 6 |
| VII | 6 | 8 | 8 | 18 | 1 | 2 | 6 | 8 | 0 | 1 | 3 | 8 | 0 | 0 | 1 | 5 |
Note: M2, M4, and M6 are number distributions in the predicted 2, 4, and 6-age classes of D. spathacea population.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



