
Submitted:
03 April 2024
Posted:
04 April 2024
You are already at the latest version
Abstract
Variable and sometimes contrasting effects of using artificial rhizobium inocula to increase grain legume performance have been reported. The objectives of this systematic review and meta-analysis were therefore to (i) estimate the overall effect of rhizobium inocula on grain yield and protein content and (ii) identify potential confounding factors that explain the observed variation. A web of Science database literature search was carried out in 2016. Data from all field experiments, greenhouse and pot experiments that compared grain yields and protein contents in Rhizobium/Bradyrhizobium-inoculated with non-inoculated control crops were extracted and analyzed using previously described unweighted and weighted meta-analysis protocols. GRADE assessments of the strength of evidence of results from the weighted meta-analyses were carried which included an assessment of the risk of publication bias. Results: For unweighted meta-analyses we identified eligible data in 236 and 36 studies for grain yield and protein content respectively. For weighted meta-analyses we identified eligible data in 92 and 8 studies for grain yield and protein content respectively. Rhizobium inoculation was found to (i) significantly increase grain yield in both the unweighted (mean percent difference: 41%, 95%CI 36,46, p<0.0001) and weighted meta-analysis (Standardized mean difference: 1.56, 95%CI 1.26,1.87, p<0.0001) and (ii) significantly increase protein content in the unweighted meta-analysis (mean percent difference: 11%, 95%CI 5,18, p<0.0001), but not the weighed meta-analyses which only detected a trend (0.1>p>0.05) towards higher protein content in inoculated grain legume crops. Multi-level model-based weighted meta-analyses suggest that the (i) type of experiment, (ii) type of fertilizer used in experiments, (iii) grain legume type/species and (iv) the country in which experiments were carried out, were confounding factors. The main limitation of the evidence is that a large number of publications did not report measures of variation and could therefore not be included in the more scientifically sound weighted meta-analyses. However based on the large evidence-base available for grain yield it can be concluded that overall Bradyrhizobium/Rhizobium seed inoculation results can increase grain yield by ~40% compared with non-inoculated grain legume crops.
Keywords:
Rhizobium
; Bradyrhizobium
; grain legumes
; grain yield
; grain protein content
; bean
; soybean
; chickpea
; lentil
1. Introduction
Grain legumes are important food and feed crops and account for ~13% of the grain crops (cereals, pulses and oilseeds) produced globally [1,2]. Legumes play an important role in human nutrition, particularly in developing countries and have been described as “poor man’s meat” due to their high protein content and also being a good source of slow release carbohydrates, minerals and vitamins [3]. Grain legumes also account for a substantial proportion of total protein intake in individuals following a vegetarian or vegan diet and are an important component of traditional Mediterranean diets, which have been linked to a range of health benefits including a lower incidence of chronic and degenerative diseases including type-2 diabetes, certain cancers, cardiovascular diseases, dementia and obesity [3,4,5].
Grain legume crops have the capacity to fix atmospheric nitrogen via a symbiotic relationship with rhizobia (species of the soil bacterial genera Rhizobium and Bradyrhizobium) in the root nodules of legume plants. Root nodules are specialist root organs of legume plants, which are formed in response to infection of roots by rhizobia and provide favourable environmental conditions (e.g., maintenance of low levels of free oxygen via the O2-buffering activity of leghaemoglobin proteins) required for optimum N-fixation by the bacterial symbionts [6,7,8]. N-fixation by the bacterial symbionts is an energy-requiring process, which is fuelled by a substantial proportion of the carbohydrate produced by the legume plants photosynthetic capacity and this partially explains the lower grain yields per unit area in grain legumes compared to cereal crops [7,8,9]. However, the capacity of symbiotic N-fixation also (i) explains the higher protein content in legume compared with cereal grain and (ii) reduces the requirement for N-fertilizer inputs and thereby greenhouse gas emissions/carbon footprint of arable crop rotations [7,8,9]. Additional benefits of including grain legumes into arable rotations can be increases in above- and below-ground biodiversity resulting in a reduction in weed, pest and disease pressure in subsequent cereal crops [7,8,9]. The use of grain legumes as break crops in arable rotations is therefore particularly beneficial in organic farming systems, which prohibit the use of synthetic, chemical crop protection products and mineral N-fertilizers [9,10,11,12].
Different groups of rhizobia are known to form functioning root nodules (=nodulate) with the main grain legume species and a recent review described that that there are now 14, 11, 6, 5, 5, 4, 3 and 2 rhizobia species known to nodulate common bean (Phaseolus vulgaris L.), soybean (Glycine max L.), cowpea (Vigna unguiculata L.), chickpea (Cicer arietinum L.), peanut (Arachis hypogaea), lentil (Vicia lens L.), faba bean (Vicia faba L.) and pea (Pisum sativum), respectively [13]. For example, species that efficiently nodulate common bean and faba bean are Rhizobium leguminosarum sv. phaseoli and Rhizobium leguminosarum sv. viciae respectively, while Bradyrhizobium japonicum and Rhizobium fredii are known to efficiently nodulate soybean [13]. The establishment of N-fixing root nodules therefore relies on high enough populations of legume species-specific rhizobium species being present in soil and the number of active root nodules and N-fixation capacity in a legume crop is correlated to the population density of rhizobia in the soil, which is known to decrease if suitable legume host plants are not grown for many years in the rotation [6,12,14,15].
It is also important to note that, both, (i) the establishment and subsequent N-fixing capacity of root nodules and (ii) legume crop performance, are known to be affected by environmental (including soil type, pH, available N-concentrations and organic matter content, precipitation, irradiation and temperature) and agronomic factors (including N-fertilizer inputs, irrigation) [12,14,15,16,17,18,19,20]. Also, legume plants have efficient mechanisms to regulate the level of nodulation in response to environmental factors (e.g., N-availability in soils) to prevent symbiotic N-fixation from becoming a metabolic burden and having a negative effect on plant growth and grain yield of legume crops [18,19,20].
One strategy to increase the performance (e.g., N-fixation, grain yield and protein content) of legume crops has been to inoculate soils or seeds with artificial Rhizobium/Bradyrhizobium inocula to compensate for (i) the absence of suitable rhizobia in soil (e.g., when non-indigenous legume species are grown for the first time) or (ii) a rapid decline in Rhizobium population density (e.g., where climatic conditions and/or infrequent growing of grain legume crops creates unfavorable conditions for persistence of rhizobia in soils) [21,22,23,24,25]. Commercial seed inoculum products, based on Rhizobium/Bradyrhizobium strains isolated from soils or root nodules, are now available for all major grain legume species [21]. There are also now efficient protocols to (i) remove bacteriophages from Rhizobium/Bradyrhizobium strains, (ii) produce inoculum by liquid fermentation technology and (iii) formulate rhizobia (e.g., with peat and/or clay carrier materials), and commercial seed inoculum products are often described to have superior nodulation and N-fixation capacity compared with strains naturally found in agricultural soils [21,22,23].
However, controlled experiments which compared grain yields and/or protein contents in grain legume crops grown with and without artificial inoculation have reported variable and sometimes contrasting effects. For example, studies by Elsheikh et al. [24], Sanora & Mallik [25] and Provorov et al. [26] reported significantly higher, similar and significantly lower grain yield in grain legume crops produced from inoculated compared with non-inoculated seed respectively. Also, a range of studies reported significant differences in nodulation/N-fixation capacity between strains of the same rhizobia species and a recent review concluded that rhizobia strains isolated in the target environments have superior nodulation/N-fixation capacity under field conditions, compared with strains selected for high N-fixation capacity in bio-assays and/or pot trials carried out under controlled growth chamber/greenhouse conditions [22,27,28].
As a result, there is still considerable controversy on whether and to what extent the use of artificial rhizobium inocula will significantly improve the yield and protein content and/or the economic performance of grain legume crops. Also, recent studies in Northern Europe have contributed to this uncertainty, because they showed that (i) even fields with no recent legume cultivation have sufficiently high rhizobia populations in soil for optimum nodulation/N-fixation of grain legume crops like faba bean and (ii) grain yield of non-inoculated faba beans, peas and lupins in long term experiments were as stable as those recorded for other spring crops (e.g., cereals) [28,29,30].
The objectives of this systematic review and meta-analysis were therefore to (i) estimate the overall effect of seed inoculation on grain yields and protein contents and (ii) identify potential confounding factors (fertilization, grain legume type/species, study type, country is which studies were carried out) that may explain the observed variation and/or contrasting effects reported in the scientific literature.
2. Materials and Methods
2.1. Criteria for Selecting Studies for the Review
All studies comparing grain yield and protein content of legumes inoculated with Rhizobia versus non-inoculated plants were included. The study included both field and glasshouse/controlled environment experiments which was included as a factor in the analysis. In addition, all species of legumes and Rhizobium or Bradyrhizobium bacteria were initially included in the review. The main outcomes were seed yield and protein content to reflect the effectiveness of Rhizobium inoculation. Yield was expressed as unit weight per given area, i.e., kg/ha, t/ha and g/plant, while protein was expressed as a percentage.
2.2. Literature Search Strategy
The literature search strategy was based on previously published protocols by Brandt et al. [31] and Baranski et al. [32]. Relevant publications were identified through an initial search of the literature with Web of Science using two search terms: “*rhizobium” and “inocula*”. The search terms were combined with Boolean logic (AND) and with truncation (*) in order to find all contrasting interventions and participants for this study. There were no language restrictions, all papers in different languages were included, and translation of papers was done by colleagues and/or using Google translate.
2.3. Screening Procedure
An EndNote library was constructed, and all duplicates were removed before the results were screened according to the eligibility criteria. Papers found in the initial search were filtered in a two-stage process:
- (i)
- Title and abstract screening. In order to minimize the risk of error two independent researchers read the title and abstract as recommended by Higgins & Thompson [32]. All studies relevant to the topic were included and if there was any doubt about suitability then the papers were included for stage two. In case the abstract was not available at this stage then the paper was also included for stage two.
- (ii)
- Full text screening. The full text of all relevant studies identified in Stage 1 was read and assessed for relevance.
2.4. Data Management and Extraction
All data relevant to primary objectives of the review were extracted from the included studies i.e., number of participants, mean value, standard deviation or standard error of the outcome measurement in each intervention group. Results reported as numerical values in the text or tables were copied directly into a spreadsheet, while data published in graphical form was enlarged, printed, measured (using a ruler) and then entered as described previously [32,33].
Authors of publications for which only the abstract was available were contacted by email and requested for full text together with any unpublished data from experiments which were within the scope of this study. Additionally, authors of all collected papers were asked for any other publications or unpublished results that could be added to this review. In order to obtain possible missing data (including measures of variation i.e., SD, SEM) the authors of the study were contacted, but very few responded and provided the additional information requested.
2.5. Data Synthesis and the Strength of Evidence Tests
The standardized mean difference (SMD) was used as an effect size to summarize treatment effects in each trial as previously recommended [32,34,35,36]. Results of the meta-analysis were visualized on forest plots which show SMD values together with 95% confidence intervals (CI) as a measure of variability (Supplementary Figures S3 and S4). The importance of each study was also expressed as a weight equal to the inverse variance which is used to calculate study weights. The SMD was calculated in relation to non-inoculated (control) plants, therefore a negative value indicates a higher performance (yield or protein content) of non-inoculated plants. Weighted meta-analysis was employed according to Baranski et al. [32] using the ‘metafor’ package of the R statistical environment [37]. The SMD from a single study was calculated using standard formulas within ‘metafor’ as follows
where is the mean value for inoculated samples, and is the mean value for non-inoculated samples. Swithin is the pooled standard deviation of the two groups, and J is a factor used to correct for small sample size.
J was calculated as:
where and are the sample sizes in the two groups
and is calculated as:
where and are the standard deviation in the two groups (organic and conventional).
The pooled SMD (SMDtot) across all studies was computed as:
where is a sampling variance estimated as:
The pooled or summary effect () was calculated for all parameters reported in a minimum of three studies, using the methods described by Lipsey and Wilson [38].
A positive SMD value indicates that the mean trait i.e., protein and yield are higher in inoculated plants, while a negative SMD indicates that the mean effect is higher in non-inoculated plants. Confidence intervals for each SMD were estimated based on standard methods using the ‘metafor’ package [37]
Unweighted meta-analyses were carried out to allow inclusion of studies that have only mean values and when measures of variability and/or sample size were unavailable. The effect size was computed as an ln-transformed ratio of inoculated means: non-inoculated means and was expressed as a percentage as shown below. The resampling method was used to evaluate comparison of the arithmetic average of samples with . P values were derived from Fisher’s one-sample randomization test and p<0.05 was considered statistically significant [32].
The mean percentage difference (MPD) was calculated to facilitate value judgements regarding the nutritional importance of the relative effect magnitudes, and to compare these between weighted and unweighted meta-analysis. The MPD was expressed as ‘% higher ‘in inoculated or non-inoculated plants, and provide estimates for the magnitude of differences. For each data-pair used for SMD calculation the mean percentage difference was computed as:
2.5. Assessment of Heterogeneity
Heterogeneity between studies resulting from contrasting methodology used in each trial was assessed using Q statistics and I2 statistics [39]. The results of these assessments were reported and discussed. A significant heterogeneity was assumed if the I2 was less than 25% and the P value for the Q statistics was greater than 0.01. In order to explore causes of between-study variability (heterogeneity) the effects of moderators were analyzed using a group meta-analytical approach.
2.6. Assessment of the Risk of Bias
The article quality and risk of bias was assessed using a critical appraisal of each paper that provided suitable data for the weighted meta-analyses. It helped to monitor a number of quality criteria that have the potential to impact on the results of the study. Study quality was evaluated using the method described by Yin [40]. The assessment was done using a form of statements for which the following responses were possible: ‘Yes’ when the statement reflected the content of the paper, ‘No’ when there was no information in the paper described by the statement or ‘Unclear’ when information provided was inconsistent with the statement (see Supplementary Table S1. tor details of the results of the quality assessments of studies included in the weighted meta-analyses).
The final rating of the overall methodological quality was carried out using the method described by Guyatt et al. [41]. Only data/means from studies which also reported the number of replicates (n) used and measures of variation were included in the weighted meta-analyses, while data/means from all studies were included in the unweighted meta-analyses.
An overall assessment of the strength of evidence was performed using an adaption of GRADE (Grading of Recommendations, Assessment, Development and Evaluation) system, which included information about risk of bias for all studies that provided date suitable for the weighted meta-analyses, as well as inconsistency, indirectness and imprecision of the results, and publication bias [40]. Publication bias was assessed by funnel plot inspection as recommended by Mlinaric et al. [42].
3. Results
3.1. Literature Search
A total of 5767 publications were identified in the initial literature search, of which 4430 were excluded, because the information given in the title and abstract suggested that they did not include suitable data (see Figure 1 for the PRISMA 2020 flow diagram for the systematic review). The full text of the remaining 1337 publications were then read and publications that did not report suitable data or contained the same data as other publications were rejected (Figure 1). This resulted in suitable data for unweighted meta-analyses of differences in grain yield and protein content between Rhizobium inoculated and non-inoculated crops to be identified in 236 and 92 publications respectively (see reference lists in the supplementary materials). A list of publications included in meta-analyses that met the inclusion criteria is provided in the supplementary materials. A large proportion of these publications did not report measures of variation (e.g., standard error, standard deviation, 95% confidence intervals) and only yield and protein content data extracted from 36 and 9 publications could therefore be included in the weighted meta-analysis (Figure 1).
The 236 studies which provided data for unweighted meta-analyses were published between 1976 and 2015 (Supplementary Figure S1) and most (199) reported results from field experiments, although 25 studies based on pot experiments and 12 studies based on greenhouse experiments also contributed to the evidence base used in meta-analyses. Field experiments were carried out across 41 different countries in Africa, Asia, North, Central and South America and Southern Europe, with a notable absence of data from Northern Europe (Supplementary Figure S2). Most studies were on soybean (Glycine max L.), other bean species (including Vicia faba L., Phaseolus vulgaris L., Vigna radiata L., Vigna mungo L., Phaseolus acutifolius A.Gray and/or Phaseolus aureus L.), chickpea (Cicer arietinum L.) and lentil (Vicia lens L.) and for these species data suitable for both weighted and unweighted meta-analyses were available (Supplementary Figures 3 and 4). Only a small number of publications reported data for pea (Lathyrus oleraceus Lam.), pigeon pea (Cajanus cajan (L.) Huth) and peanut (Arachis hypogaea L.) (8, 6 and 8 publications respectively), and none of these publications provided data suitable for weighted meta-analyses. Data for these grain legume species were therefore excluded from both the weighted and unweighted meta-analyses.
A much smaller number of studies provided data suitable for weighted meta-analyes of grain/seed yield (36 studies) and protein content (8 studies), with 34 and 3 studies reporting results from field experiments and and pot trials respectively (Supplementary Figures S3 and S4). All studies that provided data for weighted meta-analyses were of acceptable (21) or high (16) quality (Supplementary Table 1).
3.2. Standard Unweighted and Weighted Meta-Analyses
There was considerable variation in the results reported in (i) the 236 studies that compared grain yield and (ii) 92 studies that compared protein content in crops produced from Rhizobium inoculated and non-inoculated seed. Specifically, for yield 82% of studies reported significantly higher yields with inoculation, 3% reported significantly higher yields with non-inoculated seed and 15% reported no significant effect of inoculation. Similarly, for protein content 71% of studies reported significantly higher yields with inoculated seed, 11% reported significantly higher yields with non-inoculated seed and 17% reported no significant effect of inoculation.
Standard unweighted and weighted meta-analysis methods were carried out to investigate whether overall (across all studies available) Rhizobium-inoculation had a significant effect on grain/seed yield and protein content.
For grain yield, results from both the unweighted and standard weighted meta-analysis found that Rhizobium inoculation results in significantly (p<0.0001) higher yields (Table 1 and Table 2). Based on the results from the unweighted meta-analyses (which was based on 236 studies and 597 observations data-points) Rhizobium inoculation can be estimated to increased grain yield by around 40% (Table 1).
For protein content only the unweighted meta-analyses found that Rhizobium inoculation results in significantly (p<0.0001) higher protein content (Table 1), although the weighted meta-analysis also identified a trend (0.1>p>0.01) towards higher protein content in crops produced from Rhizobium-inoculated crops (Table 2). Based on the results from the unweighted meta-analysis (which was based on 92 studies and 240 observations/data-points), Rhizobium inoculation can be estimated to increase the protein content of grain legumes by around 10% (Table 1).
Heterogeneity was moderate (I2 <75%) for yield and low (I2 ≤2%) for protein content in the standard weighted meta-analyses (Table 2). Supplementary Figure S3 and S4 show the forest plots for grain yield and protein content respectively which provide detailed information on the SMD and variation observed in individual experiments/data-points included in the standard weighted meta-analyses.
GRADE assessment of the strength of evidence from the weighted meta-analyses identified that the overall reliability was moderate for both grain yield and protein content (see Table 3 for a details).
3.3. Multilevel Model Weighted Meta-Analyses
Multilevel-model weighted (MLMw) meta-analysis to investigate potential confounding effects of (i) study type (field versus pot trials) (ii) grain legume type/species (beans, chickpea, lentil, soybean) and (iii) country in which experiments were carried out (Table 4 and Table 5).
Results provide evidence that differences in type of fertilizer and experiment, and grain legume type/species augmented the effect of Rhizobium inoculation on both grain yield and protein content (Table 4 and Table 5).
Specifically, SMDs for grain yield obtained for different fertilizer types varied between 0.91 (NS) when both organic and mineral fertilizers were applied to 5.78 (p<0.0001). Similarly the SMD obtained for (i) field and pot experiments were 1.44 and 2.91, (ii) different grain legume types/species varied between 1.34 for beans and 2.61 for lentils.
The greatest variation in the effect of Rhizobium-inoculation on grain yield was detected between countries (Table 4). Notably, the MLMw meta-analysis results show that Rhizobium inoculation resulted in (i) a significantly lower grain yield (SMD: -5.54, p=0.003) in an experiment carried out in Uzbekistan, (ii) no significant effect on yield in experiments in Bangladesh, Canada, China, Nepal, New Zealand, Spain, Taiwan and Tunesia, and (iii) significantly higher grain yields in experiments carried out in
Columbia, India, Iran, Pakistan, South Africa, Sudan, Tanzania, Tukey and the USA, with the highest SMD (7.66, p<0.0001) found for experiments in South Africa (Table 4). (Table 4).
For protein content, the MLMw meta-analysis found that Rhizobium-inoculation resulted in a significant increase in protein content in (i) chick pea (but not beans, lentils and soybeans) and (ii) experiments carried out in Pakistan (but not in India, Iran and Turkey) (Table 5).
3. Discussion
3.1. Impact of Including Grain Legumes in Crop Rotations
Grain legumes are recognized to provide important ecosystem services in arable crop rotations [9,43]. Most importantly their capacity for symbiotic N-fixation significantly reduces the need for N-fertilizer inputs in arable crop rotations and results in lower greenhouse gas emissions and carbon footprints [2,6,7,17]. Diversifying arable rotations with grain legumes was also shown to reduces weed, pest and disease pressure in cereal crops grown after grain legume crops [8,9,43,44,45].
However, grain yields of legumes are lower than cereals crops, primarily because legume plants invest a significant proportion of the energy/carbohydrate from photosynthesis to establish and “fuel” N-fixation by rhizobia in root nodules [46]. Also, residual soil-N levels after grain legume crops are significantly lower than those recorded after forage legume crops (e.g., clover, lucerne, vetch), because a substantial proportion of the N assimilated via symbiotic N-fixation is utilized for seed protein synthesis and removed with the harvested grain [44,45,46]. Grain legumes, are therefore usually considered as N-neutral (= not reducing soil N, but also not producing a net-N input) in the N-budgets of intensive arable crop rotations, and may even result in a decrease in soil N and C (e.g., when grown with conventional tillage in rotations without cover crops) [44,45,46,47]. However, replacing a cereal crop (e.g., maize, wheat or rice) with an N-neutral grain legume crop in the rotation, allows N-fertilizer inputs to be reduced by between 200 and 300 kg/ha in intensive, stockless, conventional cereal crop rotations that are widely used in Europe, Japan and China [44,45,46].
3.2. Effects of Rhizobium Inoculation on the Performance of Grain Legume Crops
It is well documented that rhizobia may not be present or not be present in high enough numbers in agricultural soils for optimum nodulation and symbiotic N-fixation by legume crops, especially if (i) a suitable host legume species was not previously grown or not grown for many years in the field/region or (ii) pedoclimatic conditions result in a rapid decline in rhizobia populations in soils [48,49,5051,52,53,54].
Since rhizobia can be readily isolated from soils and multiplied by fermentation technology, the use of artificial rhizobium inocula has been one strategy to optimize symbiotic N-fixation, grain yield and protein content of grain legume crops [22,23,27]. However, as described in the introduction, this has generated variable and sometimes contrasting effects and resulted in uncertainty about (i) the overall impact of using artificial rhizobia inocula and (ii) the factors (e.g., legume species, agronomic and climatic background conditions) that may explain the variability observed.
The systematic literature and standard model weighted and unweighted meta-analyses reported here were designed to address this uncertainty and found that overall (across all studies carried out with different grain legume species) the use of artificial rhizobia inoculum increased grain yields and protein contents, although it should be noted that the increase in grain protein from inoculation was only significant when the unweighted meta-analysis protocol was used. Based on the results of the unweighted meta-analyses the average increase in grain yield and protein content was estimated at ~40% and ~10% respectively. This suggests that overall the use of inoculum is economically viable for farmers, since the cost of commercial seed inoculum products are now relatively low (usually less than 10 US $/ha) and recent economic analyses showed that relatively small increases in grain legume yields will significantly improve net-income for farmers, without economic benefits of increased residual-N (higher yields and/or lower fertilizer costs) in crops grown after legumes being taken into consideration [55,56,57,58,59]. However, since (i) crude protein content is estimated by measuring grain N-content and (ii) because grain N-content is correlated to total symbiotic N-fixation, the residual soil-N from inoculated crops with a 40% higher grain yield and 10% higher protein content, can also be estimated to be at least 40% higher compared with a non-inoculated crop.
3.3. Factors Confounding the Effects of Rhizobium Inoculation on Grain Legume Performance
The multi-level model based weighted meta-analyses were designed to identify factors that potentially confound the impact of rhizobium inoculation on grain legume yield and protein content. Results suggest that the relative effect of using inoculum differs considerably between (i) grain legume species, (ii) fertilization practices, and (iii) countries (and associated pedoclimatic conditions) in which experiments were carried out.
When different legume species were compared rhizobium inoculation increased grain yields in all species for which data were available, but the relative effect differed substantially between species. For example, standardized mean difference (SMD) was two-times higher in lentils compared with beans. Also, although a positive SMD was found for all grain legume species, a significant effect of inoculation on protein content was only detected for chick pea, but not beans, lentils and soybeans.
When the effect of inoculation was compared in crops grown with different fertilization regimes the increase in yield from inoculation (=SMD) decreased with increasing fertilization intensity (non-fertilized crops > organic fertilizers only > mineral fertilizers only > organic and mineral fertilizers) and no significant effect of inoculation was detected in crops grown with both mineral and organic fertilizers. These results are consistent with previous studies which reported that fertilization (especially mineral N-fertilizer inputs) results in (i) reduced nodulation and N-fixation by legume plants and (ii) fertilizer inputs becoming a more important yield determining factor in legume crops than symbiotic N-fixation [18,19,27].
When the effect of inoculation was compared in crops grown in different countries (and associated pedo-climatic conditions) the variation in SMDs for grain yield was even larger than the variation found between species and fertilization regime; specifically it ranged from significantly lower grain yield (SMD -5.5) in Usbekistan (which has an arid continental climate) to significantly higher grain yields (SMD 7.7) in South Africa (which has regions with sub-tropical and Mediterranean climatic conditions).
Results from the multilevel meta-analyses reported here therefore confirm previous studies which concluded that the contrasting responses to Rhizobium inoculation, may be due to complex interactions between (i) the genetics, physiology and environmental adaptation of both different legume species and Rhizobium species/strains used as inoculation and (ii) contrasting agronomic protocols (e.g., rotation, fertilization, irrigation) used for grain legumes and/or (iii) pedoclimatic background conditions [6,7,8,12,14,15,16,17,18,19,20,22,23,24,25,27,30,48,49,50,51,52,53,54]. However, a range of limitations in the evidence base made it impossible to (i) estimate the relative contribution of specific explanatory variables (e.g., soil, climatic or agronomic factors) and/or (ii) investigate interactions between these factors with respect to the impact of rhizobium inoculation on grain legume performance (see also section 3.2 Study limitations). Future research should therefore focus on studying these interactions to (i) clearly define the environmental window of opportunity (=range of climatic and agronomic background conditions) required to achieve N-fixation, crop yield and economic grains from rhizobium inocula in each grain legume species, (ii) thereby allowing better decision making by farmers (e.g., on whether and how frequently to use Rhizobium inocula).
In this context, it is important to note that water deficiency, mineral N-fertilizer application, a soil pH lower than 5.5 and low soil organic matter content were reported to reduce Rhizobium persistence, symbiotic N-fixation, and/or the yield gain from rhizobium inoculation [14,17,27,44,49,52,54]. In contrast, high soil organic matter content, the use of manure as fertilizer and temperate climates were described to be conducive to rhizobium persistence in soils [14,22,24,29]. These factors may partially explain large variation observed between (i) studies carried out in different countries and (ii) different studies carried out in the same country in the multi-level model meta-analyses reported here. They may also may explain (i) the results of a recent study which reported that “Fields with no recent legume cultivation have sufficient nitrogen-fixing rhizobia for crops of faba bean” in the UK, a country with a maritime, temperate climate and soils with a relatively high organic matter content [29] and (ii) why rhizobium inoculum products are not widely used or recommended for the main grain legume crops traditionally grown in Northern Europe (faba beans, peas and lentils), while they are recommended and used in crops that were more recently introduced in this region (e.g., soybean and chick pea) [60,61,62,63].
Over the last 10 years a range of studies has focused on improving the isolation, selection, production and formulation of rhizobium strains used in inoculum products and it may be possible that recently developed products will deliver N-fixation, yield and economic grains also in the traditional grain legume crops grown in Northern Europe. This view is supported by a recent study from the UK which reported evidence that Rhizobium inoculation of clover seed results in a substantial increase in nodulation and N-fixation by red clover leys, although rhizobium inoculation is not recommended for clover in the UK [64].
For grain yield there were also enough data to compare results obtained in pot and field experiments. The finding that pot experiments (which are often carried out under controlled conditions in greenhouses or growth chambers) reported higher yield grains from rhizobium inoculation suggests that pot trials do not provide an accurate estimate of the yield improvements that can be achieved with rhizobium inocula under field conditions.
3.4. Study Limitations
The main limitation of the evidence is that a large number of publications did not report measures of variation (e.g., standard errors, standard deviation, 95% confidence intervals) and could therefore not be included in the more scientifically sound weighted meta-analyses. This reduced the statistical power of the weighted meta-analyses, especially for protein content.
Another limitation was that the limited background information provided in most articles that contributed to the evidence base and as a result, we could not investigate interactions between soil, climate and agronomic explanatory variables, that may have explained the variation between legume species, fertilization regime and country/pedoclimatic background conditions identified in the multi-level model based weighted meta-analysis. Specifically, limitations included (i) insufficient information on soil, climatic and agronomic background conditions, (ii) different rhizobium strains and/or grain legume species being used in countries where experiments being carried out, (iii) the relatively small number of studies available from most countries, and (iv) the variation in study design and agronomic protocols used.
5. Conclusions
The study provided strong evidence that overall, the use of rhizobium inocula will result in a substantial increase in grain yields in bean, chickpea, lentil and soybean crops.
However, results also confirm previous studies which reported that (i) the yield gap between inoculated and non-inoculated crops decreases with increasing fertilization intensity (especially mineral N-fertilizer inputs) and (ii) under certain pedoclimatic and/or agronomic background conditions, inoculation may have no significant effect or result in lower yields compared with non-inoculated crops.
The finding that rhizobium inoculation resulted in similar or higher protein content suggests that N-fixation and residual-N available to crops grown after grain legumes will also increase substantially if inocula are used. This confirms that the use of rhizobium inoculum has a significant positive impact not only on (i) grain yields and associated economic gains for farmers, but also (ii) substantially increases environmental benefits (reduced need for mineral N-fertilizers and associated reduction in greenhouse gas emissions) of including grain legumes in arable rotations.
There is considerably scope to further improve the efficacy of rhizobium inocula through (i) research that improves the understanding of the complex interactions between legumes, rhizobium, other plant growth promoting microbes including VA mycorrhizal fungi, soil physical, chemical and biological and climatic factors and (ii) the development of improved strain isolation, efficacy screening and formulation methods for Rhizobium inocula.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Figure S1. Number of articles included in the meta-analysis by year of publication; Supplementary Figure S2. Number of articles from which data were extracted for meta-analysis by country (field experiments only); Supplementary Figure S3. Forest Plot showing using standardized mean differences (SMD) and 95% confidence intervals of comparisons of grain/seed yields in rhizobium-inoculated and non-inoculated legume crops in studies/experiments included in the standard random effect model-based weighted meta-analysis. Supplementary Figure S4. Forest Plot showing using standardized mean differences (SMD) and 95% confidence intervals of comparisons of grain protein content in rhizobium inoculated and non-inoculated legume crops in studies/experiments included in the standard random effect model-based weighted meta-analysis. Supplementary Table S1. Results of the article quality assessments.
Author Contributions
Conceptualization, E.K.S., MB and C.L.; methodology, E.K.S., M.B., and C.L.; software, E.K.S., M.B. and L.R.; validation, P.B., M.B. and C.L.; formal analysis, E.K.S., M.B., and L.R.; investigation, E.K.S., G.H., J.W., L.R. and M.B.; resources, P.B. and C.L.; data curation, E.K.S. and M.B.; writing—original draft preparation, E.K.S., P.B. and C.L.; writing—review and editing, E.K.S., G.H., J.W., P.B., L.R., N.V., M.B. and C.L.; visualization, E.K.S. and M.B.; supervision, P.B., L.R., M.B. and C.L.; project administration, P.B. and C.L.; funding acquisition, P.B. and C.L. All authors have read and agreed to the published version of the manuscript
Funding
This research was funded by the European Union Integrated project Quality Low Input Food (Grant number 506358) (2004−2009) and a Professorial Fellowship awarded to Prof Carlo Leifert from the Sheepdrove Trust.
Data Availability Statement
Data will be made available upon reasonable request by the 1st author Enas Khalid Sufar or the 5th author Leonidas Rempelos.
Acknowledgments
The authors gratefully acknowledge the administrative support from Lois Bell, Dave Whittock and English language proof reading of the manuscript by Catherine A. Leifert.
Conflicts of Interest
All authors declare no conflict of interest.
References
- FAOstat. Statistics Database of the Food and Agriculture Organization of the United Nations. 2016. Available online: http://www.fao.org/statistics/databases/en/ (accessed on 21 October 2023).
- Watson, C.A.; Reckling, M.; Preissel, S.; Bachinger, J.; Bergkvist, G.; Kuhlman, T.; Lindström, K.; Nemecek, T.; Topp, C.F.E.; Vanhatalo, A.; et al. Grain Legume Production and Use in European Agricultural Systems. Adv. Agron. 2017, 144, 235–303. [CrossRef]
- Tharanathan, R.N.; Mahadevamma, S. 'Grain legumes—A boon to human nutrition'. Trends Food Sci. Technol. 2003, 14, 507-518. [CrossRef]
- Ortega, R.M. Importance of functional foods in the Mediterranean diet. Public Health Nutr. 2006, 9, 1136–1140. [CrossRef]
- Guasch-Ferré, M.; Willett, W.C. The Mediterranean diet and health: A comprehensive overview. J. Intern. Med. 2021, 290, 549–566. [CrossRef]
- Goyal, R.K.; Mattoo, A.K.; Schmidt, M.A. Rhizobial-Host Interactions and Symbiotic Nitrogen Fixation in Legume Crops Toward Agriculture Sustainability. Front. Microbiol. 2021, 12, 669404. [CrossRef]
- Dutta, A.; Trivedi, A.; Nath, C.P.; Gupta, D.S.; Hazra, K.K. A comprehensive review on grain legumes as climate-smart crops: Challenges and prospects. Environ. Chall. 2022, 7, 100479. [CrossRef]
- Cusworth, G.; Garnett, T.; Lorimer, J. Agroecological break out: Legumes, crop diversification and the regenerative futures of UK agriculture. J. Rural. Stud. 2021, 88, 126–137. [CrossRef]
- Robson, M.C.; Fowler, S.M.; Lampkin, N.H.; Leifert, C.; Leitch, M.; Robinson, D.; Watson, C.A.; Litterick, A.M. The agronomic and economic potential of break crops for ley/arable rotations in temperate organic agriculture. Adv. Agron. 2002, 77, 370–427. [CrossRef]
- EU. Regulation (EU) 2018/848 of the European Parliament and of the Council of 30 May 2018. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:02018R0848-20220101 (accessed on 21 October 2023).
- Rempelos, L.; Baranski, M.; Wang, J.; Adams, T.N.; Adebusuyi, K.; Beckman, J.J.; Brockbank, C.J.; Douglas, B.S.; Feng, T.; Greenway, J.D.; et al. Integrated Soil and Crop Management in Organic Agriculture: A Logical Framework to Ensure Food Quality and Human Health? Agronomy 2021, 11, 2494. [CrossRef]
- Barbieri, P.; Starck, T.; Voisin, A.-S.; Nesme, T. Biological nitrogen fixation of legumes crops under organic farming as driven by cropping management: A review. Agric. Syst. 2023, 205, 103579. [CrossRef]
- Shamseldin, A.; Abdelkhalek, A.; Sadowsky, M.J. Recent changes to the classification of symbiotic, nitrogen-fixing, legume-associating bacteria: A review. Symbiosis 2017, 71, 91-109. [CrossRef]
- Peoples, M.B.; Brockwell, J.; Hunt, J.R.; Swan, A.D.; Watson, L.; Hayes, R.C.; Li, G.D.; Hackney, B.; Nuttall, J.G.; Davies, S.L. et al. Factors affecting the potential contributions of N2 fixation by legumes in Australian pasture systems. Crop Pasture Sci. 2021, 63, 759-786. [CrossRef]
- Liu, L.; Knight, J.D.; Lemke, R.L.; Farrell, R.E. A side-by-side comparison of biological nitrogen fixation and yield of four legume crops. Plant Soil 2019, 442, 169–182. [CrossRef]
- Stoddard and Schauman. There Is a Grain Legume for Every Field. University of Helsinki. Legumes Translated Practice Note 66. 2022. Available online: https://www.legumehub.eu/wp-content/uploads/2022/04/PN66-There-is-a-grain-legume-forevery-field.pdf (accessed on 21 October 2023).
- Smith, C.J.; Chalk, P.M. Grain legumes in crop rotations under low and variable rainfall: Are observed short-term N benefits sustainable? Plant Soil 2020, 453, 271–279. [CrossRef]
- Downie, J.A. Legume nodulation. Current Biology, 2014, 24, 184-190. [CrossRef]
- Ferguson, B.J.; Mens, C.; Hastwell, A.H.; Zhang, M.; Su, H.; Jones, C.H.; Chu, X.; Gresshoff, P.M. Legume nodulation: The host controls the party. Plant Cell Environ. 2019, 42, 41–51. [CrossRef]
- Ferguson, B.J.; Lin, M.H.; Gresshoff, P.M. Regulation of legume nodulation by acidic growth conditions. Plant Signal Behav. 2013 8, e23426. [CrossRef]
- Newedge microbials. Legume Inoculant Group Chart. Available online: https://microbials.com.au/innoculant-selector/ (accessed on 11 March 2024).
- Mendoza-Suárez, M.; Andersen, S.U.; Poole, P.S.; Sánchez-Cañizares, C. Competition, Nodule Occupancy, and Persistence of Inoculant Strains: Key Factors in the Rhizobium-Legume Symbioses. Front. Plant Sci. 2021, 12, 690567. [CrossRef]
- Fudge, R. Rhizobia Inoculation for Organic farming Systems. University of Minnesota Extension 2024. Available online: https://www.printfriendly.com/p/g/CqS8L9 (accessed on 11 March 2024).
- Elsheikh, E.A.E.; Salih, S.S.M.; Elhussein, A.A.; Babiker, E.E. Effects of intercropping, Bradyrhizobium inoculation and chicken manure fertilisation on the chemical composition and physical characteristics of soybean seed. Food Chem. 2009, 112, 690-694. [CrossRef]
- Sanoria, C.L; Mallik, M.K. (1981) 'The effect of seed inoculation with Rhizobium and Azotobacter on yield and quality of lentils (Lens esculenta). J. Agric. Sci. 1981, 97, 237-239. [CrossRef]
- Provorov, N.A.; Saimnazarov, U.B.; Bahromov, I.U.; Pulatova, D.Z.; Kozhemyakov, A.P.; Kurbanov, G.A. Effect of rhizobia inoculation on the seed (herbage) production of mungbean (Phaseolus aureus Roxb.) grown at Uzbekistan. J. Arid Environ. 1998, 39, 569-575. [CrossRef]
- Thilakarathna, M.S.; Raizada, M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol. Biochem. 2017, 105, 177-196. [CrossRef]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V.; Hunegnaw, D.K. Legume-rhizobium specificity effect on nodulation, biomass production and partitioning of faba bean (Vicia faba L.). Sci Rep. 2021, 11, 3678. [CrossRef]
- Reckling, M.; Döring, T.F.; Bergkvist, G.; Stoddard, F.L.; Watson, C.A.; Seddig, S.; Chmielewski, F.-M.; Bachinger, J. Grain legume yields are as stable as other spring crops in long-term experiments across northern Europe. Agron. Sustain. Dev. 2018, 38, 63. [CrossRef]
- Maluk, M.; Ferrando-Molina, F.; Lopez del Egido, L.; Langarica-Fuentes, A.; Yohannes, G.G.; Young, M.W.; Martin, P.; Gantlett, R.; Kenicer, G.; Hawes, C.; et al. Fields with no recent legume cultivation have sufficient nitrogen-fixing rhizobia for crops of faba bean (Vicia faba L.). Plant Soil 2022, 472, 345–368. [CrossRef]
- Brandt, K.; Srednicka-Tober, D.; Baranski, M.; Sanderson, R.; Leifert, C.; Seal, C. (2013) 'Methods for Comparing Data across Differently Designed Agronomic Studies: Examples of Different Meta-analysis Methods Used to Compare Relative Composition of Plant Foods Grown Using Organic or Conventional Production Methods and a Protocol for a Systematic Review', J. Agric. Food Chem. 2013, 61, 7173-7180. [CrossRef]
- Baranski, M.; Srednicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, C. et al. (2014) 'Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analyses', Br. J. Nutr. 2014, 112, 794-811. [CrossRef]
- Lajeunesse, M.J. Recovering missing or partial data from studies: A survey of conversions and imputations for meta-analysis. In: Handbook of meta-analysis in ecology and evolution; ed. Koricheva, J., Gurevitch, J., Mengersen, K.; Princeton University Press, Princeton, USA, 2013; pp. 196-485.
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [CrossRef]
- Stewart, G. Meta-analysis in applied ecology. Biol. Lett. 2010, 6, 78-81. [CrossRef]
- Koricheva, J.; Gurevitch, J. (2013) Place of meta-analysis among other methods of research synthesis. In Handbook of Meta-analysis in Ecology and Evolution; ed. Koricheva, J., Gurevitch, J., Mengersen, K.; Princeton University Press, Princeton, USA, 2013; pp. 3-14. [CrossRef]
- Viechtbauer, W. Conducting Meta-Analyses in R with the metafor Package. J. Stat. Softw. 2010, 36, 1-48. [CrossRef]
- Lipsey, M.W. and Wilson, D.B. (2001) Practical meta-analysis; Sage Publications, Inc., Thousand Oaks, USA, 2001; pp.154-196.
- Higgins, J.P. and Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Statist. Med. 2002, 21, 1539–1558. [CrossRef]
- Yin, R.K. Case Study Research. Design and Methods. Z. Personalforsch. 2009, 26, 93-101.
- Guyatt, G.H.; Oxman, A.D.; Montori, V.; Vist, G.; Kunz, R.; Brozek, J.; Alonso-Coello, P.; Djulbegovic, B.; Atkins, D.; Falck-Ytter, Y. et al. GRADE guidelines: 5. Rating the quality of evidence-publication bias. J Clin Epidemiol, 2011, 64, 1277-82. [CrossRef]
- Mlinaric, A.; Horvat, M; Smolcic, V.S. (2017) Dealing with the positive publication bias: Why you should really publish your negative results. Biochem. Med. 2017, 27, 447-452. [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005.
- Jangir, C.K.; Thakur, A.; Bijani, H.; Thakur, P.; Kumar, S.; Meena, R.S.; Bedwal, S.; Rani, K.; Shukla, U.N.; Meena, A.K.; et al. Chapter 7 - Residual nitrogen for succeeding crops in legume-based cropping system. In Advances in Legumes for Sustainable Intensification. Ram Swaroop Meena, R.S., Kumar S. Eds.; Academic Press, Cambridge, USA, 2022; pp. 113-132.
- Gogoi, N., Baruah, K.K., Meena, R.S. (2018). Grain Legumes: Impact on Soil Health and Agroecosystem. In Legumes for Soil Health and Sustainable Management. Meena, R.; Das, A.; Yadav, G.; Lal, R. Eds.; Springer Nature Singapore Pte Ltd., Singapore, 2018, pp 1-16.
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [CrossRef]
- Plaza-Bonilla, D.; Nolot, J.-M.; Passot, S.; Raffaillac, D.; Justes, E. Grain legume-based rotations managed under conventional tillage need cover crops to mitigate soil organic matter losses. Soil Tillage Res. 2016, 156, 33-43. [CrossRef]
- Rupela, O.P.; Toosan, B.; Mittal, S.; Dart, P.J.; Thompson J.A. Chickpea Rhizobium populations: Survey of influence of season, soil depth and cropping pattern. Soil Biol. Biochem. 1987, 19, 247-252. [CrossRef]
- Issa, S.; Wood, M. Multiplication and survival of chickpea and bean rhizobia in dry soils: The influence of strains, matric potential and soil texture. Soil Biol. Biochem. 1995, 27, 785-792. [CrossRef]
- Hirsch, P. R. Population Dynamics of Indigenous and Genetically Modified Rhizobia in the Field. New Phytol. 1996, 133, 159–171. [CrossRef]
- Nzeyimana, F. Onwonga, R.N.; Ayuke, F.O.; Chemining'wa, G.N.; Nabahungu, N.L.; Bigirimana, J.; Josiane U.K.N. Determination of abundance and symbiotic effectiveness of native rhizobia nodulating soybean and other legumes in Rwanda. Plant Environment Interactions 2024, 5, e10138. [CrossRef]
- Slattery, J.F.; Coventry D.R.; Slattery W.J. Rhizobial ecology as affected by the soil environment. Aust. J. Exp. Agric. 2001, 41, 289 – 298. [CrossRef]
- Zengeni, R.; Mpepereki, S.; Giller, K.E. Manure and soil properties affect survival and persistence of soyabean nodulating rhizobia in smallholder soils of Zimbabwe. Appl. Soil Ecol., 2006, 32, 232-242. [CrossRef]
- Romdhane, S.B.; Trabelsi, M.; Aouani, M.E.; de Lajudie, P.; Mhamdi, R. The diversity of rhizobia nodulating chickpea (Cicer arietinum) under water deficiency as a source of more efficient inoculants. Soil Biology Biochem. 2009, 41, 2568-2572. [CrossRef]
- Mmbaga, G.W.; Mtei, K.M.; Ndakidemi, P.A. Yield and Fiscal Benefits of Rhizobium Inoculation Supplemented with Phosphorus (P) and Potassium (K) in Climbing Beans (Phaseolus vulgaris L.) Grown in Northern Tanzania. Agricultural Sciences 2015, 6, 783-797. [CrossRef]
- Ulzen, J.; Abaidoo, R.C.; Mensah, N.E.; Masso, C.; AbdelGadir, A.H. Bradyrhizobium Inoculants Enhance Grain Yields of Soybean and Cowpea in Northern Ghana. Front. Plant Sci. 2016, 7, 1770. [CrossRef]
- Paudel, D.; Wang, L.; Poudel, R.; Acharya, J.P.; Victores, S.; Lopes de Sousa, C.H., Rios, E.; Wang, J. Elucidating the effects of organic vs. conventional cropping practice and rhizobia inoculation on rhizosphere microbial diversity and yield of peanut. Environ. microbiome 2023, 18, 60. [CrossRef]
- Messias, M.; Asobia, P.C., de Brito Ferreira, E.P. Economic analysis of rhizobia and azospirilla co-inoculation in common beans. Bull. Natl. Res. Cent. 2023, 47, 173. [CrossRef]
- Chandarana, K.A.; Amaresan, N. Rhizobium Biofertilizers: Mass Production Process and Cost-Benefit Ratio Analysis. In: Agricultural Microbiology Based Entrepreneurship. Microorganisms for Sustainability, vol 39. Springer Nature Singapore Pte Ltd., Singapore, 2023.
- PGRO. Online Pulse Agronomy Guide. Available on line: www.pgro.org/pulse-agronomy-guide/ (accessed 25 March 2024).
- FiBL. Organic soya in Switzerland – Optimizing the Swiss production chain from seeds to the final product. Available online: www.fibl.org/en/themes/projectdatabase/projectitem/project/1053/ (accessed 25 March 2024).
- BIOAktuell.ch. Biosoja. Available online: https://www.bioaktuell.ch/pflanzenbau/ackerbau/koernerleguminosen/soja (accessed 25 March 2024).
- Ökolandbau. De. Ökologischer Kichererbsenanbau. https://www.oekolandbau.de/landwirtschaft/oekologischer-pflanzenbau/spezieller-pflanzenbau/ackerbau/koernerleguminosen/kichererbsen/ (accessed 25 March 2024).
- Wilkinson, A.;Wilkinson, J.N.; Shotton, P.; Eyre, M.; Hasanaliyeva, G.; Bilsborrow, P.; Leifert, C.; Rempelos, L. Effect of Clover Sward Management on Nitrogen Fixation and Performance of Following Spring- andWinter Wheat Crops; Results of a 3-YearPilot Study. Agronomy 2022, 12, 2085. [CrossRef]
Figure 1.
PRISMA 2020 flow diagram providing a summary of the on-line search results and selection of studies included in the weighted and unweighted meta-analyses. uwMA, unweighted meta-analysis; wMA, weighted meta-analysis; 1, number of comparisons/data points extracted from studies.
Figure 1.
PRISMA 2020 flow diagram providing a summary of the on-line search results and selection of studies included in the weighted and unweighted meta-analyses. uwMA, unweighted meta-analysis; wMA, weighted meta-analysis; 1, number of comparisons/data points extracted from studies.
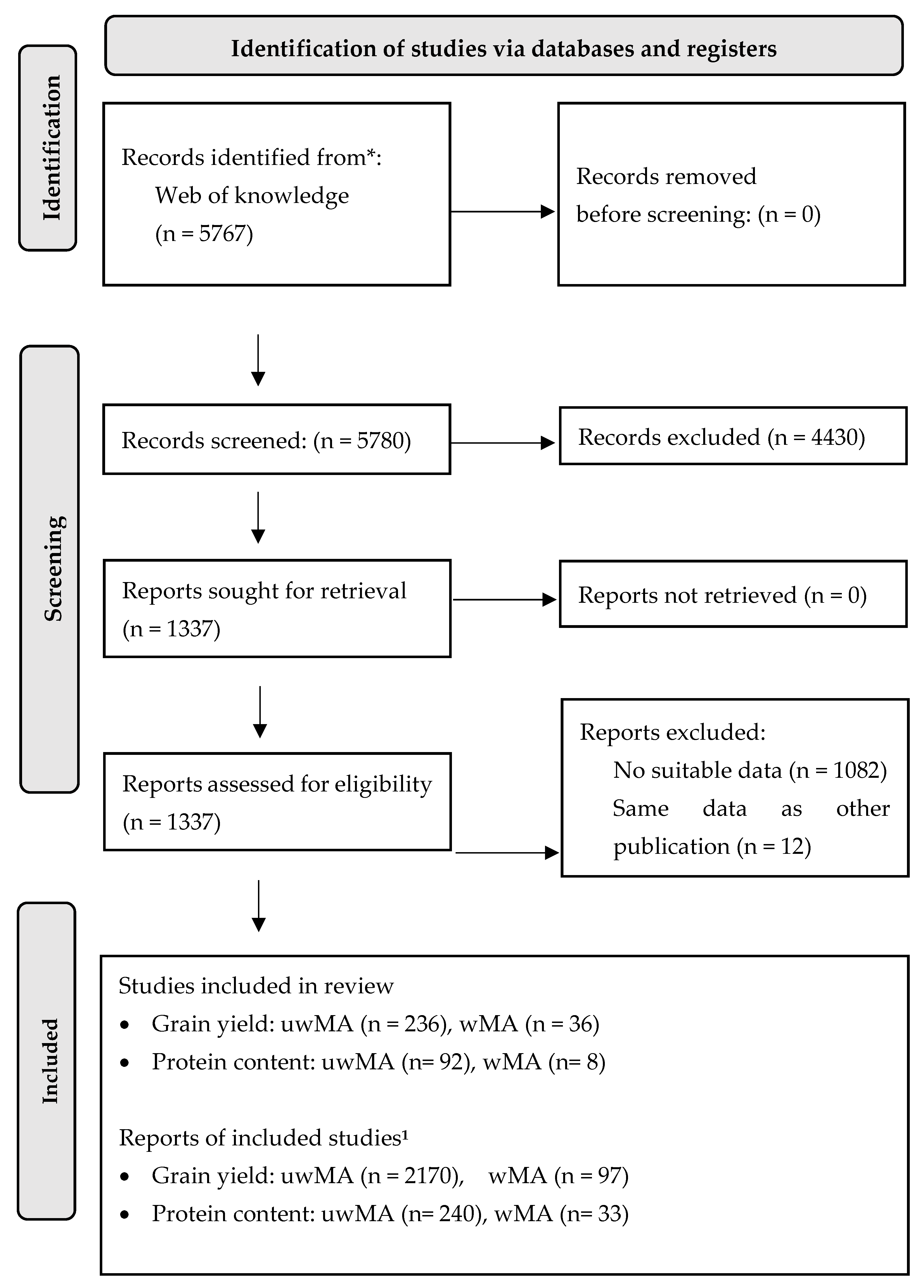

Table 1.
Effects of Rhizobium inoculation of grain legume seed on yield and protein content; results of the unweighted meta-analysis.
Table 1.
Effects of Rhizobium inoculation of grain legume seed on yield and protein content; results of the unweighted meta-analysis.
| Parameter | N | N | MPD | 95% CI | p | Fisher’s ratio (p) | |
| Grain yield | 236 | 597 | 41 | 36 | <0.0001 | 134 (<0.0001) | |
| Protein content | 92 | 240 | 11 | 5 | <0.0001 | 109 (<0.0001) |
N, number of studies that provided data suitable for unweighted meta-analysis; n, number of comparisons/data points; MPD, mean percent difference; 95%CI, 95% confidence interval.
Table 2.
Effects of Rhizobium inoculation of grain legume seed on grain yield and protein content; results of the standard weighted meta-analysis.
Table 2.
Effects of Rhizobium inoculation of grain legume seed on grain yield and protein content; results of the standard weighted meta-analysis.
| Parameter | N | SMD | P | 95% CI | Heterogeneity (I2) |
| Grain yield | 97 | 1.56 | <0.0001 | 1.26,1.87 | 66% |
| Grain Protein | 33 | 0.25 | T | -0.02,0.52 | 2% |
n, number of comparative data sets, SMD, standardized mean difference; 95%CI, 95% confidence interval; T, trend (0.1>p>0.05).
Table 3.
GRADE (Grading of Recommendations, Assessments, Development and Evaluation) assessment of the strength of evidence for standard weighted meta-analysis with SMD as effect size for all parameters included.
Table 3.
GRADE (Grading of Recommendations, Assessments, Development and Evaluation) assessment of the strength of evidence for standard weighted meta-analysis with SMD as effect size for all parameters included.
| Parameter | Effect Magnitude1 |
Precision2 | Inconsistency3 | Publication bias4 | Overall Reliability5 |
|---|---|---|---|---|---|
| Yield | Moderate | High | High | No | Moderate |
| Protein | Small | High | Moderate | High | Moderate |
1, Effect magnitude: large effects were defined as >20%, moderate effects as 10-20% and small effects as <10%.2, Precision was based on the width of the pooled effect CI and the extent of overlap in the substantive interpretation of effect magnitude GRADE assessment. 3, Inconsistency was based on the measure of heterogeneity and the consistency of effect direction.4, Publication bias was assessed using visual inspection of funnel plots, Egger tests, two fail-safe number tests and trim and fill. Overall publication bias was considered strong when indicated by two or more methods, moderate when indicated by one method, and low when indicated by none of the methods. 5, The overall quality of evidence was then assessed across domains as recommended for standard GRADE appraisals.
Table 4.
Multilevel model- based weighted meta-analysis of the effect of Rhizobium seed inoculation on the yield of grain legumes; factors included in the model were fertilization, experiment type, legume species and country.
Table 4.
Multilevel model- based weighted meta-analysis of the effect of Rhizobium seed inoculation on the yield of grain legumes; factors included in the model were fertilization, experiment type, legume species and country.
| Factors | n | SMD | 95% CI | p | Heterogeneity (I2) |
|---|---|---|---|---|---|
| All | 97 | 1.56 | 1.26,187 | <0.0001 | 67% |
| Fertilizer type | |||||
| Mineral | 77 | 1.52 | 1.22, 1.83 | <0.0001 | 56% |
| Mineral + organic | 2 | 0.91 | -0.80,2.62 | NS | 56% |
| No-fertiliser | 1 | 5.78 | 3.42,8.14 | <0.0001 | 56% |
| Not-specified | 15 | 0.99 | 0.32,1.65 | 0.0033 | 56% |
| Organic | 2 | 3.66 | 1.57,5.76 | 0.0005 | 56% |
| Experiment type | |||||
| Field | 91 | 1.44 | 1.14,1.74 | <0.0001 | 62% |
| Pot | 6 | 2.91 | 1.80,4.01 | <0.0001 | 62% |
| Species | |||||
| Bean | 53 | 1.34 | 0.92,1.76 | <0.0001 | 67% |
| Chickpea | 10 | 1.86 | 0.87,2.84 | 0.0002 | 67% |
| Lentil | 5 | 2.61 | 1.09,4.13 | 0.0007 | 67% |
| Soybean | 29 | 1.72 | 1.18,2.27 | <0.0001 | 67% |
| Country (climate) | |||||
| Bangladesh (tropical monsoon) | 3 | 1.45 | -0.04,2.95 | T | 60% |
| Canada (various) | 3 | 0.47 | -1.01,1.96 | NS | 60% |
| China (various) | 2 | 0.96 | -0.90,2.84 | NS | 60% |
| Colombia (tropical; sub-tropical) | 7 | 1.45 | 0.41,2.48 | 0.0050 | 60% |
| India (various) | 20 | 1.83 | 1.19,2.47 | <0.0001 | 60% |
| Iran (various) | 12 | 1.70 | 0.84,2.57 | 0.0001 | 60% |
| Nepal (various) | 1 | 0.17 | -2.44,2.78 | NS | 60% |
| New Zealand (maritime, temperate) | 8 | 0.73 | -0.16,1.64 | NS | 60% |
| Pakistan (various) | 5 | 3.80 | 2.22,5.39 | <0.0001 | 60% |
| South Africa (Mediterranean, subtropical) | 2 | 7.66 | 4.47,10.84 | <0.0001 | 60% |
| Spain (mainly Mediterranean) | 2 | 0.92 | -0.86,2.71 | NS | 60% |
| Sudan (transitional; semi desert; semi arid) | 3 | 4.29 | 2.15,6.44 | <0.0001 | 60% |
| Taiwan (humid sub-tropical) | 6 | 1.00 | -0.04,2.04 | T | 60% |
| Tanzania (tropical wet; savanna) | 4 | 1.86 | 0.71,3.01 | 0.001 | 60% |
| Tunisia (Mediterranean) | 5 | 1.07 | -0.05,2.20 | T | 60% |
| Turkey (mainly Mediterranean) | 10 | 1.52 | 0.62,2.41 | 0.0008 | 60% |
| USA (various) | 3 | 2.01 | 0.44,3.57 | 0.0100 | 60% |
| Uzbekistan (arid continental) | 1 | -5.45 | -9.10,-1.80 | 0.0030 | 60% |
n, number of comparative data sets, SMD, standardized mean difference; 95%CI, 95% confidence interval; NS, not significant (p≥0.1); T, trend (0.1>p>0.05).
Table 5.
Multilevel model-based weighted meta-analysis of the effect of Rhizobium seed inoculation on the protein content of grain legumes; factors included in the model were fertilizer type, legume species and country where the experiments were carried out.
Table 5.
Multilevel model-based weighted meta-analysis of the effect of Rhizobium seed inoculation on the protein content of grain legumes; factors included in the model were fertilizer type, legume species and country where the experiments were carried out.
| Factors | n | SMD | 95% CI | p | Heterogeneity (I2) |
|---|---|---|---|---|---|
| All | 33 | 0.25 | -0.02,0.53 | T | 2% |
| Fertility Type | |||||
| Mineral | 28 | 0.19 | -0.10,0.49 | NS | <1% |
| Not-specified | 5 | 0.59 | -0.14,1.33 | NS | <1% |
| Species | |||||
| Bean | 14 | 0.10 | -0.32,0.53 | NS | <1% |
| Chickpea | 7 | 0.70 | 0.10,1.29 | 0.020 | <1% |
| Lentil | 2 | 0.64 | -0.71,2.01 | NS | <1% |
| Soybean | 10 | 0.09 | -0.41,0.59 | NS | <1% |
| Country | |||||
| India | 5 | 0.41 | -0.24,1.06 | NS | <1% |
| Iran | 12 | 0.00 | -0.46,0.46 | NS | <1% |
| Pakistan | 6 | 1.08 | 0.33,1.84 | 0.004 | <1% |
| Turkey | 10 | 0.08 | -0.41,0.59 | NS | <1% |
n, number of comparative data sets, SMD, standardized mean difference; 95%CI, 95% confidence interval; NS, not significant (p≥0.1); T, trend (0.1>p>0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



