Submitted:
04 April 2024
Posted:
05 April 2024
You are already at the latest version
Abstract
Relative to several model bacteria, the ethanologenic bacterium Zymomonas mobilis is shown here to have an elevated resistance to exogenous antimicrobial peptides (AMPs) – both, with regard to peptide bulk concentration in the medium, and the numbers of peptide molecules per cell. By monitoring integration of the AMPs in the bacterial cell membrane and observing the resulting effect on membrane energy-coupling, it is concluded that the membranotropic effects of the tested AMPs in Z. mobilis and in Escherichia coli are comparable. The advantage of Z. mobilis over E. coli apparently results from its uncoupled mode of energy metabolism that, in contrast to E. coli, does not rely on oxidative phosphorylation, and hence, is less vulnerable to disruption of its energy-coupling membrane by AMPs. It is concluded that the high resistance to antimicrobial peptides (AMPs) observed in Z. mobilis not only proves crucial for its survival in its natural environment but also offers a promising platform for AMP production and sheds light on potential strategies for novel resistance development in clinical settings.
Keywords:
Antimicrobial peptides
; Minimal inhibitory concentrations
; Zymomonas mobilis
; uncoupled growth
; Langmuir-Blodgett compression isotherm
; Chemiosmotic coupling
1. Introduction
The ethanologen Zymomonas mobilis is a facultatively anaerobic alpha-proteobacterium, with an efficient ethanol fermentation pathway, based on the Entner-Doudoroff glycolysis in combination with pyruvate decarboxylase and alcohol dehydrogenase activities [1]. It has an exceptionally high ethanol productivity, several times exceeding that of the yeast Saccharomyces cerevisiae [2]. That is primarily because the catabolism of Z. mobilis is loosely coupled to the energy demands of its anabolism [3], and proceeds with an excessive rate, channeling up to 98% of substrate carbon to ethanol. Although Z. mobilis is an obligately fermentative bacterium, able to grow only on glucose, fructose and sucrose, it also possesses an active, constitutive aerobic respiratory chain. In line with Z. mobilis uncoupled mode of metabolism, its respiratory chain also operates with a low energy-coupling efficiency. It does not contribute to oxidative ATP synthesis and biomass yield in aerobically growing culture [4,5,6,7], yet it performs rapid regeneration of intracellular NAD(P)+ pools [8]. These characteristics of Z. mobilis physiology are relevant to its applications in biotechnology, and have attracted the interest of researchers over several decades. Successful attempts have been made to engineer its catabolic pathways and to design novel bioprocesses [9,10], involving also co-cultivations with other producer microorganisms [11,12]
However, co-cultivations with Z. mobilis often prove to be problematic. Early work on conjugation with E. coli [13], co-cultivations with yeast [14], successful use of Z. mobilis biomass for treating infections [15,16,17], and other related evidence (for a review, see [18]) strongly point to antagonistic interactions between Z. mobilis and a wide variety of other microorganisms. It was suggested that this bacterium might be producing some unidentified antimicrobial substances (‘zymocins’). Lima et al. hypothesized that it might be an antimicrobial peptide [19]. We could not find evidence in support of the peptide nature of Z. mobilis antimicrobial activity. Yet, we identified acetate and propionate as the principal antimicrobial compounds of this bacterium. We showed that the raise of the antimicrobial activity by the respiratory metabolism of aerobic culture was paralleled by accumulation of both acids, while the non-respiring mutant lacking the respiratory NADH dehydrogenase (ndh), or the wild-type under strictly anaerobic growth conditions, did not accumulate these acids, and at the same time, had a negligible antimicrobial activity [20].
In a broader sense, the ability to produce antimicrobial compounds represents just one side of the interspecies competitiveness. Another side of the coin is the resistance to inhibitors, produced by other species of the same ecological niche, including bacteria, fungi and plants. Although Z. mobilis might not be synthesizing antimicrobial peptide(s) (AMPs) of its own, resistance to those coming from other organisms might well add to its competitiveness and ability to suppress its neighbors. A priori, there seems to be some rational basis to expect resistance to AMPs in bacteria with uncoupled energy metabolism. Since bacterial cell membrane is the key target of most AMPs [21], which function as pore-forming agents that increase transmembrane permeability, one might speculate that interference with membrane chemiosmotic energy-coupling plays a central role in their antimicrobial effects. Accordingly, the survival of a bacterium like Z. mobilis, relying completely upon substrate-level ATP generation during ethanol fermentation, could be less vulnerable to disruption of their membrane energy-coupling. In the present work we actually demonstrate a substantially higher resistance of Z. mobilis to selected AMPs, than that of E. coli, S. aureus, and several other bacteria, and discuss the relation of this finding to the uncoupled energy metabolism, as well as its potential biotechnological relevance.
2. Results
2.1. Comparative Screening of Z.mobilis MIC Values for the Selected AMPs
Comparative evaluation of the resistance against 11 AMPs revealed that Z. mobilis possessed significantly higher resistance than other bacteria that we had examined earlier [22], including conventional pathogens such as P. aeruginosa and S. aureus (Table 1).

Table 1.
Minimal inhibitory concentrations (MIC) of antimicrobial peptides against selected bacteria, μg mL‐1.
Table 1.
Minimal inhibitory concentrations (MIC) of antimicrobial peptides against selected bacteria, μg mL‐1.
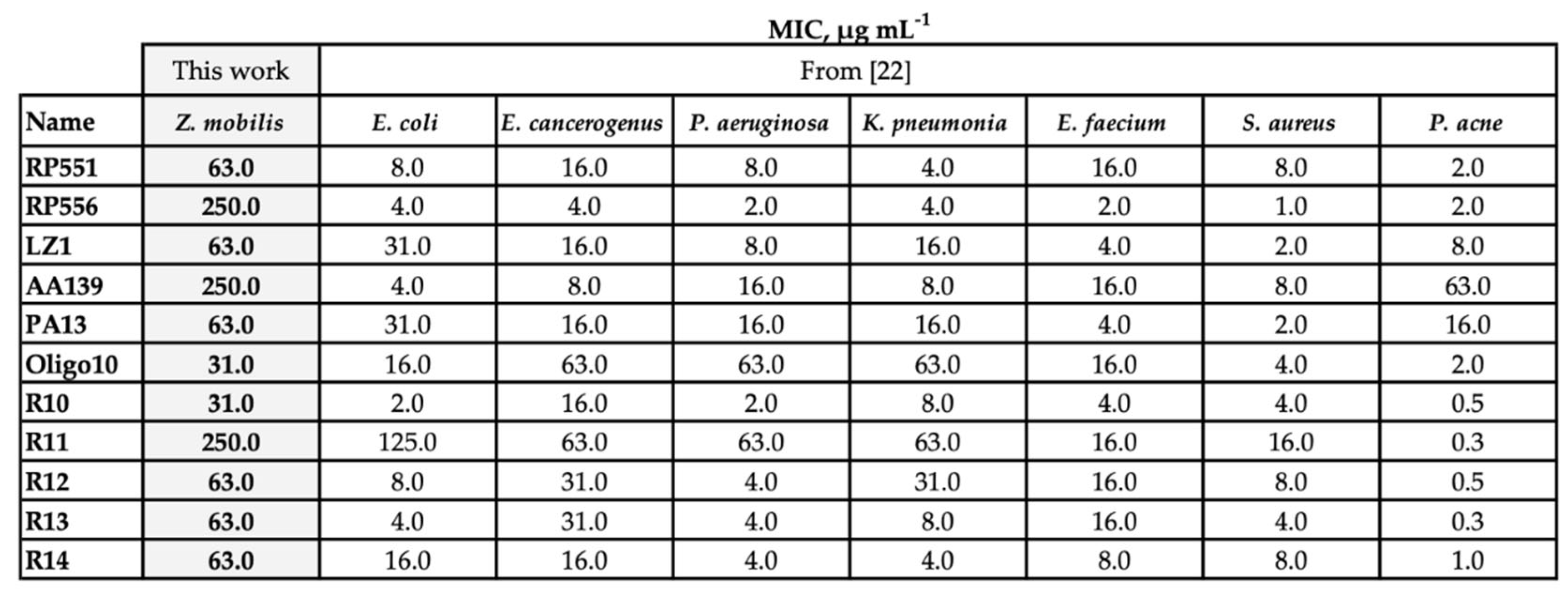
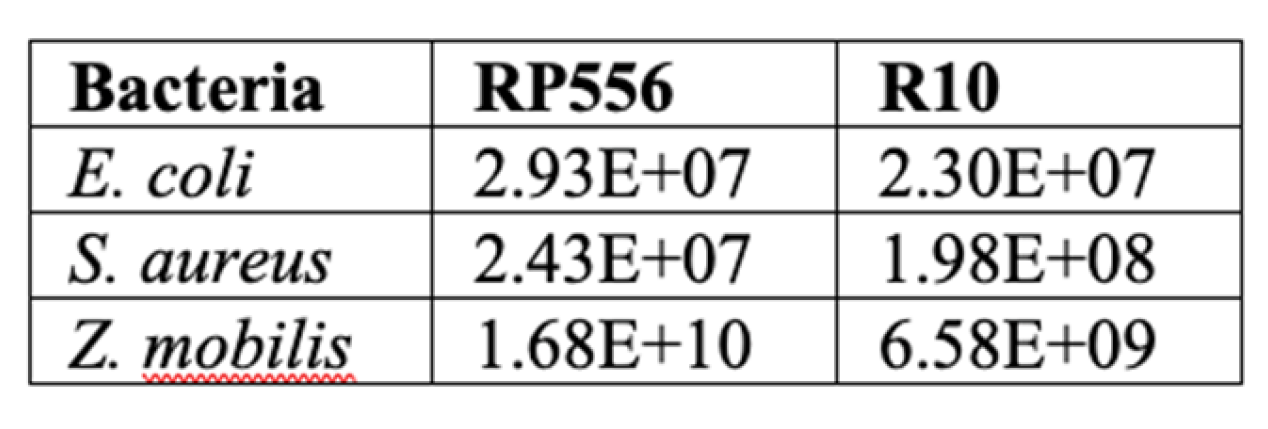
We noted, however, that there was a substantial difference between the specific cultivation conditions of Z. mobilis and the rest of tested bacteria: Z. mobilis required higher concentration of inoculum. Higher amount of biomass introduced at the start of cultivation might mitigate the antimicrobial effect of the AMPs, since that decreases the number of peptide molecules available to penetrate the membrane of each individual cell. Indeed, it was shown that at least 107 bound peptide molecules per bacterial cell are needed to kill it [23]. To examine, if it was the large inoculum of Z. mobilis (and hence, too small amount of peptide molecules per cell) that caused the observed elevated resistance to AMPs, we compared the Z. mobilis resistance to that of other microorganisms by normalizing the added amounts of AMPs to the respective inocula size. For these experiments, we chose one Gram-positive and one Gram-negative model strain (S.aureus and E.coli, respectively) and two AMPs, RP556 and R10. As the cells of all three bacterial species have a roughly comparable size, calculation of specific inhibitory concentrations for all strains was done by using the E. coli cell size parameters [24].
Table 2.
Normalized inhibitory concentrations of antimicrobial RP556 and R10 peptides against
selected bacteria. Values represent peptide molecules per cell needed to inhibit bacterial growth.
Table 2.
Normalized inhibitory concentrations of antimicrobial RP556 and R10 peptides against
selected bacteria. Values represent peptide molecules per cell needed to inhibit bacterial growth.
2.2. Confocal Microscopy Examination of Intracellular Localization of the FITC-Labelled AMPs
Confocal microscopy images for E. coli and Z. mobilis showed similar results for both bacteria (Figure 1). Taken together, our examination of FITC-labeled AMPs interacting with the E. coli and Z. mobilis membranes unveiled the ability of labeled AMP to spontaneously insert into phospholipid membranes. At the same time, it is worth noting that the utilization of FITC fluorescent tags in the confocal microscope experiments is based on assumption that the tagging process has not affected the physiochemical characteristics of the respective peptides. That might as well not be true. To address this potential concern, we delved deeper into the interactions between these AMPs and membranes by employing the Langmuir-Blodgett monolayer technique.
2.3. The AMP Effect on Langmuir-Blodgett Compression Isotherms of Model Membranes
By providing a controlled and well-defined experimental platform, the Langmuir-Blodgett (LB) monolayer technique can yield valuable information about the mechanisms of interaction of AMPs with biological membranes. It is well established that monitoring of the surface pressure via the monolayer area, or the so called π–A isotherms of such mixed lipid/AMPs Langmuir monolayers, may reveal the lipid-peptide interactions [25]. To see if weak AMP—membrane interaction might be the underlying reason of Z. mobilis high AMP resistance, we applied the LB monolayer technique, and compared the obtained results with our previously published data on the interaction of E. coli phospholipid model membranes with the same AMPs [22]. In E. coli, as a model Gram-negative bacteria, the membrane lipid composition is well characterized [26]. Its inner membrane is composed mainly of phosphatidylethanolamine (PE), phosphatidylglycerol and cardiolipin (CL). Z. mobilis membrane phospholipid composition is less well studied. According to [27], it contains less phosphatidylethanolamine, but is enriched with phosphatidylserine and phosphatidylcholine (Table 1). Of these phospholipids, phosphatidylcholine is the less negatively charged one and, at the same time, it is more commonly found in eukaryotic cell membranes (accordingly, the eukaryotic cell membranes carry less negative charge, and thus are less susceptible to the positively charged AMPs).
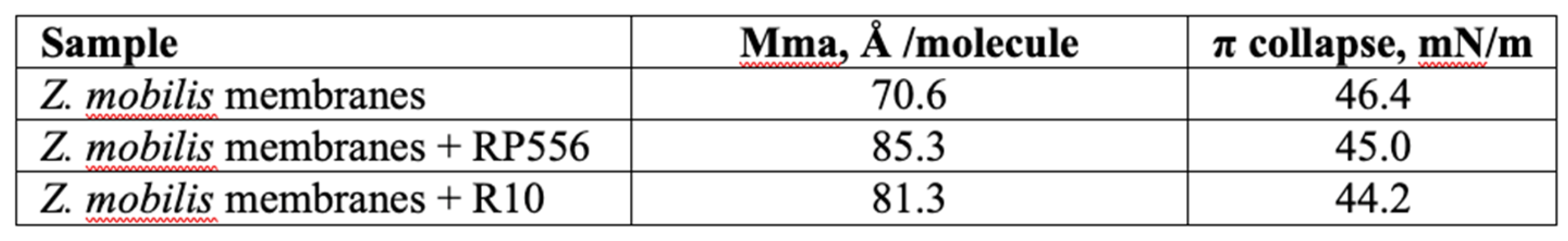
Analysis of the π –A isotherms confirmed that both peptides, RP556 and R10, intercalated in the Z. mobilis phospholipid model membrane, and slightly reduced its π–collapse values (Figure 2). The same effect was observed previously with E. coli phospholipid model membrane [22].
To quantitatively assess the interactions within membrane monolayers, we utilize a parameter commonly employed in LB techniques: the limiting area per molecule (Aπ→0). This value is derived by extrapolating the linear segment of a densely packed solid monolayer’s π–A isotherm to zero surface pressure. In our context, Aπ→0 signifies the limiting area occupied by each membrane-forming unit, and this linear segment typically falls within the surface pressure range of 25–40 mN/m. As the surface pressure (π) of 30 mN/m generally represents the lipid packing density of a cell membrane’s outer leaflet [28], the results obtained from our model system hold biological relevance. Consistent with our findings in the E. coli membrane model system [22], the present results with Z. mobilis membranes show that within the lipid layer, R556 occupies a larger area than R10, resulting in a higher Aπ→0 value (see Table 3).
Table 3.
Aπ→0 and π‐collapse values of Z. mobilis phospholipid model membrane with peptides,
derived from π‐A isotherms.
Table 3.
Aπ→0 and π‐collapse values of Z. mobilis phospholipid model membrane with peptides,
derived from π‐A isotherms.
Altogether, confocal microscopy and the LB experiments confirm membranothropic behavior of both peptides in Z. mobilis. Our study thus strongly indicates that the Z. mobilis membrane composition does not prevent its interaction with AMPs, and apparently cannot serve as an explanation for the elevated AMP resistance of this bacterium.
2.4. AMP Uptake by Cells
While it has been established that the tested AMPs interact with bacterial membranes, the inherent amphiphilic nature of AMPs still raises questions about the proportion of added AMPs that remain in the aqueous solution, whether in the buffer system or cultivation medium. This information holds significance as it can aid in more accurately estimating the precise number of molecules required to reach the critical threshold for pore formation in biological membranes. To assess this, we introduced FITC-labeled R10 peptide to thermally inactivated E. coli and Z. mobilis cells at room temperature. Subsequently, we monitored the time-course of fluorescence changes between the buffer, where the initially labeled peptides were introduced, and the inactivated cells (Figure 3). Thus, after integration in bacterial membranes, peptides transfer the fluorescence signal from the medium to the cells.
As seen from the steep increase of the fluorescence ratio, AMP integration in cell membranes occurs within the first seconds of the experiment, capturing vast majority of added AMPs. Of course, rapid integration in membranes does not necessary mean that the final pore conformation is arranged, yet it highlights the integration speed and proportions of peptide residing in the cell membranes versus the fraction remaining in the aqueous media. In all experiments the increase of fluorescence ratio cells/ buffer was close to 5. This observation indicates that the previously obtained normalized peptide inhibitory concentrations per cell might largely correspond to the membrane-integrated AMP fraction (even bearing in mind that to some extent integration in the membrane itself might alter the fluorescence).
2.5. The Interference of AMPs with ATP Synthesis by Artificially Induced Transmembrane pH Gradient
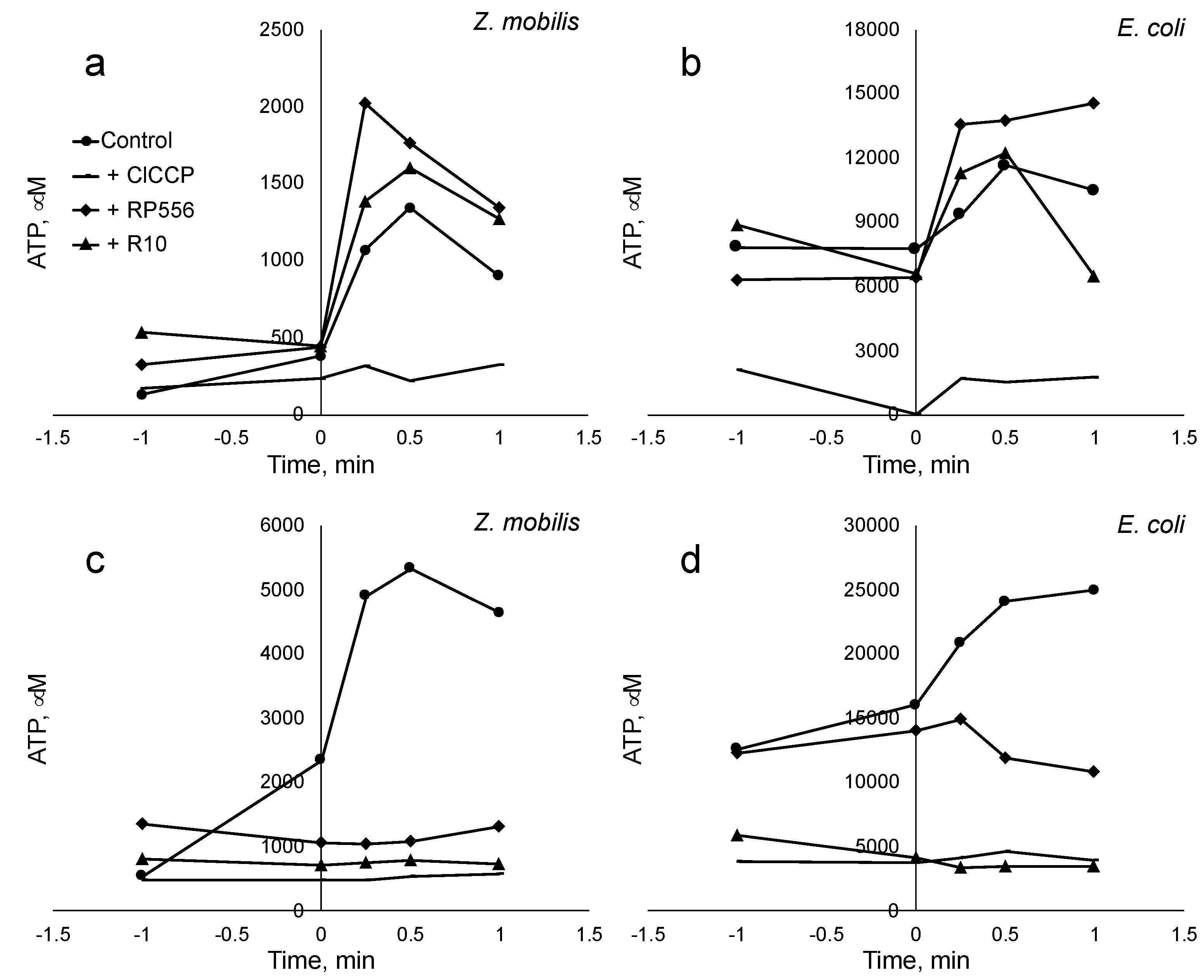
All the evidence accumulated by the above experiments demonstrates interaction of the peptides with membranes of both bacteria. The key question however is, to what extent the AMPs interact specifically with the cytoplasmic membrane in each bacterium, and whether that affects energy-coupling. In particular, that is relevant for Z. mobilis, since this Gram-negative bacterium is known for the low permeability of its outer membrane. In principle, an AMP-impermeable outer membrane could be a plausible alternative explanation of Z. mobilis high resistance to the tested AMPs. To gain a direct indication of the AMPs’ interference with energy-coupling in the cytoplasmic membranes we chose to monitor ATP synthesis in response to artificially induced transmembrane pH gradient, and to examine the putative effect of AMP addition upon the membrane energy-coupling, added in concentrations above and below the inhibitory threshold.
Synthesis of ATP induced by an artificial pH gradient is related to the activity of the proton-dependent ATP synthase of the bacterial cytoplasmic membrane. pH gradient-induced ATP synthesis has been previously demonstrated both in Z. mobilis and E. coli [6,29,30]. We here assumed that any effect the AMPs exert on the pH-induced ATP synthesis, would indicate a direct interaction between the peptide and the cytoplasmic membrane. After induction of a pH gradient of 3.5-4.0 units in cell suspensions of both Z. mobilis and E. coli, we observed an abrupt increase of the intracellular ATP concentration, reaching its maximum value within a time span between 30 sec and 1 min. The magnitude of the observed ATP concentration rise approximately corresponded to previously published values. Addition of the protonophoric uncoupler ClCCP at 10 μM concentration caused an almost complete deenergization and a dramatic drop of the ATP yield. Notably, the AMPs, when added in amount exceeding the threshold value of about E10+7 molecules per cell (Figure 4, panel c and d), also largely eliminated the ATP jump after the pH pulse, although in both bacteria peptides caused some steady elevation of the intracellular ATP level during incubation already before the pH shift. When added in amounts below the threshold value (panels a and b), peptides had no inhibitory effect on the ATP response. These results indicate that in both bacteria these peptides deenergize the cytoplasmic membrane to a comparable degree. Yet, such an effect elicits a lot stronger physiological response in E. coli, than in Z. mobilis.
3. Discussion
In the present study we demonstrate that the ethanologenic bacterium Z. mobilis, in addition to its broad-spectrum antimicrobial activity, manifests elevated resistance to exogenous AMPs. The MIC values of 11 tested AMPs for Z. mobilis appeared to be substantially higher than almost all MIC values of the same peptides for several other Gram-positive and Gram-negative bacteria [22], including pathogens. For two AMPs, selected from the list of eleven, we calculated the amount of peptide molecules per bacterial cell needed to induce the inhibitory effect. For Z. mobilis, these normalized values also were high, exceeding those of S. aureus approximately 10 times and those of E. coli even by 3 orders of magnitude.
Several experiments confirmed that the antibacterial effects of the two selected AMPs resulted from their interaction with the cell membranes in both, E. coli and Z. mobilis. In model membranes composed from phospholipid mixtures, simulating, respectively, the composition of E. coli [22] and Z. mobilis (present work) cytoplasmic membranes, integration of AMPs in the phospholipid layer was demonstrated by Langmuir-Blodgett compression isotherms. Confocal microscopy of whole cells of both bacteria using FITC-labelled AMP clearly showed localization of the fluorescing peptide molecules near the cell membranes. When the labelled peptides were added to suspensions of thermally inactivated bacteria, the fluorescent signal rapidly accumulated in the biomass, while simultaneously, the fluorescence decreased in the extracellular medium, showing that the AMPs penetrated cell membranes via passive diffusion and accumulated there at high proportion.
In order to see, if the AMPs penetrated also the cytoplasmic membrane (not just the outer membrane and / or the periplasmic space) and thus could directly interfere with the membrane energy-coupling in both bacteria, we applied induction of ATP synthesis by means of artificial transmembrane pH gradient as a test assay. We assumed that forming peptide pores in the lipid bilayer [21] would elevate its proton conductance and reduce the fraction of protons crossing the cytoplasmic membrane via the ATP synthase channel in its Fo part, in result lowering the ATP yield. ATP synthesis after pH shift was indeed strongly reduced by the peptides, indicating that the AMPs tested here interacted with cytoplasmic membranes of both, E. coli and Z. mobilis. Apparently, these peptides interferred with the hemiosmotic energy coupling via the cytoplasmic membrane F1Fo-type H+-ATP synthase in both bacteria. Although the absolute value of the ATP response to an artificial pH gradient in each bacterium was different, and close to previously reported values [6,30], in both bacteria we observed a pronounced deenergizing effect of AMPs (when added in amounts exceeding the inhibitory threshold), resembling that of the protonophoric uncoupler ClCCP. The above data suggest that the difference between E. coli and Z. mobilis with respect to AMP resistance does not result from a higher permeability barrier of Zymomonas outer membrane, or from poor integration of the peptides in the lipid bilayer of its cytoplasmic membrane, relative to E. coli. Rather, it reflects the specifics of the uncoupled Z. mobilis energy metabolism per se. In contrast to the energy metabolism of E. coli, energy supply in (micro)aerobically growing Z. mobilis culture is based exclusively on substrate-level phosphorylation in the E-D pathway [1,6,31] Therefore, disruption of the energy-coupling via the H+-ATP synthase is less damaging for its overall viability. In other words, the inherent decoupling of energy metabolism in this bacterium, coupled with its dependence on substrate-level ATP production, enhances its resilience to inhibitory compounds that target hemiosmotic energy coupling, suggesting the existence of a novel resistance mechanism against AMPs.
The high AMP resistance of Z. mobilis should have a strong ecological relevance for this bacterium. Apart from competitive advantage against other microorganisms, this trait is essential for its survival in its natural media, namely, plants. Z. mobilis is a member of plant microbiota, inhabiting sugary plant materials [16,18]. Plants synthesize a vast variety of AMPs, which serve for plant defense against pathogens and also affect plants own growth and development [32]. Hence, ability to resist media containing various AMPs of plant origin might represent a significant selective advantage in this ecological niche.
Given the significant challenge of masking the potentially lethal effects of AMPs on the bacterial host in recombinant production, it becomes all the more tempting to capitalize on the novel AMP-resistance exhibited by Z. mobilis for the overproduction of heterologous AMPs. That, however raises several issues, the main of which might be the relatively low cell yield of this bacterium that directly results from its inefficient energetics – which, at the same time, might be the very basis of its high AMP-resistance, as we propose here. Z. mobilis can be regarded as a potential producer organism of AMPs, but design of a technologically feasible production may still require a lot of research.
As the development of resistance to antimicrobial compounds remains one of the most pressing challenges for healthcare systems, the identification of this novel super-resistance mechanism to antimicrobial peptides (AMPs) observed in this study holds critical importance beyond the scope of Z. mobilis physiology.
4. Materials and Methods
4.1. Strains and Cultivation
Z. mobilis strains, Zm6 (ATCC 29191) E. coli (ATCC 25404) and S. aureus strain (DSM 20231) were subjected to a screen for AMP resistance. Z. mobilis strains were routinely grown on the standard culture medium containing (per liter) 5 g of yeast extract, 1 g of KH2PO4, 1 g of (NH4) 2SO4, and 0.5 g of MgSO4·7H2O, supplemented with 40 mL of 50% glucose solution. E. coli was grown on LB medium, containing (per liter) 10 g of tryptone, 5 g of yeast extract, and 10 g of NaCl. S. aureus was grown on Mueller Hinton Broth 21 g per 1 litre, at 30 oC.
4.2. Design and Synthesis of Antimicrobial Peptides
Six AMP amino acid sequences used in this study were published earlier, while five were novel peptides designed by the present project team [21]. All AMPs, including the fluorescein isothiocyanate (FITC) labeled RP556 and R10 peptides, were synthesized at a 1 mg scale by ProteoGenix in France, utilizing the solid-phase synthesis method. The purity of these peptides, assessed through mass spectroscopy and high-performance liquid chromatography, consistently exceeded 98%. While, according to APPTEST calculations, RP556, AA139 and R11 peptides where antiparallel beta sheet-structured, all the others formed alpha helices.
4.3. Quantification of Antibacterial Activity
The quantification of antimicrobial resistance against various AMPs was conducted by monitoring the bacterial growth in the presence of peptides at serial dilutions. This assay was performed in 96-well plates using the Infinite® M200 PRO Multimode Microplate Reader from Tecan in Maennedorf, Switzerland, as described in our recent report [19]. All bacterial strains were cultivated at 32°C for 13–18 hours at 200 rpm. Optical density measurements (λ = 550 nm) were automatically recorded at 10-minute intervals. The antimicrobial resistance activity was quantified as the minimum inhibitory concentration (MIC), which represents the lowest peptide concentration capable of preventing the growth of bacterial cells.
4.4. Confocal Microscopy
For confocal microscopy analysis, E. coli and Z. mobilis cells were cultured to the mid-logarithmic phase, harvested by centrifugation at 4000 rpm for 5 minutes, and washed twice with PBS buffer. The cells, at a concentration of approximately 0.3 mg dry weight mL-1, were incubated with labeled AMPs, following the methodology described previously [32]. In brief, FITC-labeled R10 peptides (2 µg/mL) were incubated with the cells for 30 minutes at 37°C. Subsequently, the cells were washed twice with PBS to remove any unbound labeled peptides. The cells were then fixed with 4% paraformaldehyde for 20 minutes and washed again with PBS. The localization of the peptides within the bacteria was observed using a confocal laser scanning microscope Leica DM RA-2 (Germany) equipped with a TCS-SL confocal scanning head.
4.5. Langmuir−Blodgett Compression Isotherms
Langmuir-Blodgett compression isotherm experiments were conducted at a controlled temperature of 30 ± 1 ◦C on an antivibration table using the KSV LB instrument (KSV MiniMicro, Helsinki, Finland) equipped with two movable barriers, collectively offering a total area of 273 cm². Surface pressure measurements were performed using a platinum Wilhelmy plate with a perimeter of 39.24 mm, in accordance with the methodology outlined in [25].. The process of obtaining surface pressure versus area per molecule isotherms involved a computer-controlled incremental compression technique. The monolayer of the membrane was compressed by a defined area increment (225 mm² in this case), and then allowed to relax. The relaxation process was considered complete when the changes in surface pressure became smaller than 0.02 mN/m per second. At this point, the equilibrium surface pressure of the monolayer (π) was determined. This incremental compression-relaxation process was repeated until all isotherms were recorded and the monolayer eventually collapsed. Herein, for the study of interactions between antimicrobial peptides (AMPs) and phospholipids bacterial model membranes were used. In E. coli as a model of Gram negative strains, according [25], the inner membrane is mainly composed of phosphatidylethanolamine (PE), while Z. mobilis membrane composition is more diverse with less PE content [26]. Detailed model membrane compositions used in this study are presented in the Table 4.
Table 4.
Phospholipid composition of the bacterial model membranes used in this study.
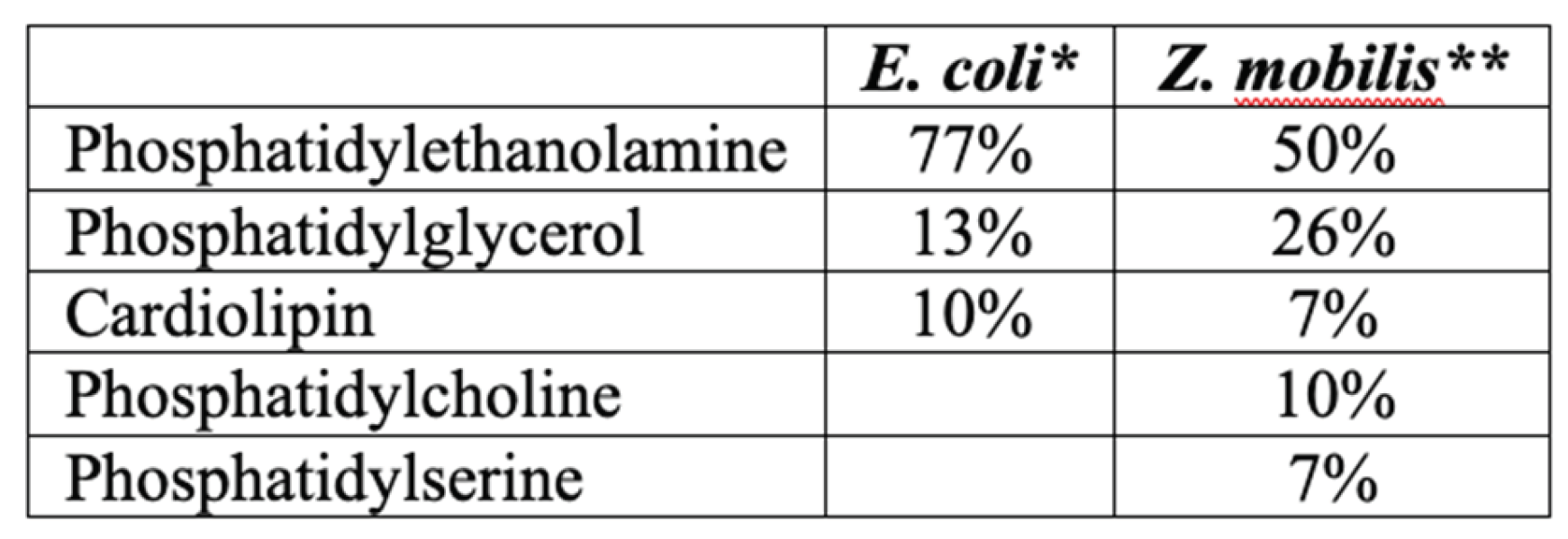
*Hoyo et al. 2020; ** Barrow et al. 1983
To investigate AMP-membrane interactions, 50 μL of model membranes at the concentration of 0.2 mg protein mL-1 were gently deposited onto the surface of deionized water (27.2 MΩ·cm) using a Hamilton micro syringe. In experiments involving AMPs, a solution containing 5 μL of the peptide dissolved in ethanol (at the concentration of 1 mg mL-1) was mixed with the model membranes before deposition on the water surface. After allowing complete solvent evaporation, measurements of surface pressure (π) versus mean molecular area (Mma) isotherms were taken in 10 minute intervals. The interactions between AMPs and the membranes were characterized using the collapse pressure (π collapse) that indicated the surface pressure at which the monolayer became densely packed, and further pressure increase was restricted. This collapse leads to the formation of a condensed monolayer state, limiting the membrane area per unit (Aπ→0) as π approaches 0 mN/m. To derive the Aπ→0 values, the linear portion of the π–A isotherms (within the π range of 25–40 mN m-1) was approximated by the linear function π = Mma * x + b, where ‘Mma’ and ‘b’ are fitting constants, and then the limiting area, corresponding to π = 0, was calculated using the equation Aπ→0 = −b/a.
4.6. ATP Synthase Activity Induced by Artificial pH Gradient
ATP synthase activity induced by artificial pH gradient was measured in non-growing starved cell suspensions. For preparation of non-growing cell suspension, cells were harvested at late exponential phase, sedimented, washed, and resuspended in 100 mM potassium phosphate buffer (pH 7.0), to a biomass concentration of 1.5–2.0 g dry wt L-1. Artificial transmembrane pH gradients of 3.5–4.0 pH units were induced 4 h after cell starvation on orbital shaker (120 rpm) by the addition of 16 μL of 1 M HCl to 200 μL aliquot of cell suspension (with 3.8–4.0 mg dry wt mL-1 biomass concentration). For ATP determination, 20 μL samples were quenched in ice-cold 10% TCA and assayed by the standard luciferin-luciferase method [7] using a TECAN Infinite 200Pro microplate reader. To evaluate the impact of AMPs and the protonophoric uncoupler ClCCP upon ATP synthesis, 5 μL of peptide (10 mg mL-1), or ClCCP (at 10 μM final concentration) was added to 200 μL of starved cell suspension, 2 h (for peptides), or 5 min (for ClCCP) prior to the pH shift by HCl.
4.7. Uptake of FITC-Labeled AMPs in Inactivated Cells
To quantify the uptake rate of AMPs in the inactivated cells, FITC-labeled peptides were used. For this, E. coli DH5α and Z. mobilis ZM4 cells were grown to midlogarithmic phase, harvested by centrifugation 5000 rpm for 5 min and washed with sterile dH2O. The bacterial cells were resuspended in 800 µL sterile dH2O and inactivated by heating for 15 min at 85 °C. Then 7.2 mL of dH2O was added to reach optical density OD600 1, and 1 mL of the sample was taken as control for the fluorescence measurements. Afterwards, FITC-labeled R10 peptide (5 µg/mL) was added. 1 mL samples were taken at 0.5, 1, 2.5, 5, 7.5, 10 min and centrifuged immediately at 13 000 rpm for 1 min to separate cells from the supernatant. The cells were resuspended in 1 mL dH2O for fluorescence measurements. The FITC fluorescence measurements for the cells and supernatant were done with FluoroMax®-3 (HORIBA Jobin Yvon Inc.) spectrofluorometer at λ ex =490 nm and λ em =530 nm.
5. Conclusions
Compared to several model bacteria, Z. mobilis demonstrated significantly elevated resistance to various AMPs, both in terms of peptide concentration in the medium and the number of peptide molecules per cell. This resistance extends beyond conventional pathogens like E. coli and S. aureus, which suggests a unique defense mechanism possessed by Z. mobilis. By examining the integration of AMPs into the bacterial cell membrane and their impact on membrane energy coupling, it was revealed that Z. mobilis, despite similarities with E. coli in membrane interaction, possesses a distinct advantage due to its uncoupled mode of energy metabolism. Unlike E. coli, Z. mobilis does not rely on oxidative phosphorylation, rendering it less susceptible to disruptions in energy-coupling caused by AMPs. The high resistance of Z. mobilis to AMPs not only contributes to its survival in its natural environment, particularly in sugary plant materials, but also presents opportunities for biotechnological applications. Z. mobilis could serve as a platform for AMP production due to its resistance properties, offering insights into potential strategies for combating antimicrobial resistance in clinical settings. While the AMP resistance of Z. mobilis holds promise for biotechnological applications, such as heterologous AMP production, challenges remain, particularly concerning cell yield and energy efficiency. Further research is needed to address these challenges and harness the full potential of Z. mobilis in AMP biotechnological production.
Overall, the study sheds light on the intricate interplay between microbial physiology, membrane interactions, and antimicrobial resistance mechanisms, underscoring the importance of interdisciplinary approaches in addressing challenges related to antimicrobial resistance and microbial biotechnology.
Author Contributions
RR and UK conceived and designed research. MR, IS, ZL and RR conducted experiments. RR contributed new reagents and analytical tools. RR and UK analyzed data. UK, RR and MR wrote the manuscript. All authors read and approved the manuscript.
Funding
This work was supported by the European Regional Development Fund project No.1.1.1.1/ 19/A/097 (“Novel antimicrobial peptides from Zymomonas mobilis for treatment of skin and soft tissue infections”).
Conflicts of Interest
“The authors declare no conflicts of interest.”.
References
- Sprenger G (1996) Carbohydrate metabolism in Zymomonas mobilis: a catabolic highway with some scenic routes. FEMS Microbiol Lett 145:301-307. [CrossRef]
- Rogers PL, Lee KJ, Skotnicki ML, Tribe DE (1982) Ethanol production by Zymomonas mobilis. In: Microbial Reactions. Advances in Biochemical Engineering, vol. 23. Springer, Berlin, pp 37–84.
- Belaich JP, Senez JC (1965) Influence of aeration and pantothenate on growth yields of Zymomonas mobilis. J Bacteriol 89: 1195–1200.
- Strohdeicher M, Neuß B, Bringer-Meyer S, Sahm H (1990) Electron transport chain of Zymomonas mobilis. Interaction with the membrane-bound glucose dehydrogenase and identification of ubiquinone 10. Arch Microbiol 154:536–543. [CrossRef]
- Kalnenieks U, Galinina N, Bringer-Meyer S, Poole RK (1998) Membrane d-lactate oxidase in Zymomonas mobilis: Evidence for a branched respiratory chain. FEMS Microbiol Lett 168:91–97.
- Rutkis R, Galinina N, Strazdina I, Kalnenieks U (2014) The inefficient aerobic energetics of Zymomonas mobilis: Identifying the bottleneck. J Basic Microbiol 54:1090–1097. [CrossRef]
- Rutkis R, Strazdina I, Balodite E, Lasa Z, Galinina N, Kalnenieks U (2016) The low energy-coupling respiration in Zymomonas mobilis accelerates flux in the Entner–Doudoroff pathway. PLoS ONE, 11:4. [CrossRef]
- Pentjuss A, Odzina I, Kostromins A, Fell DA, Stalidzans E, Kalnenieks U (2013) Biotechnological potential of respiring Zymomonas mobilis: A stoichiometric analysis of its central metabolism. J Biotechnol 165:1–10. [CrossRef]
- Wang X, He Q, Yang Y, Wang J, Haning K, Hu Y, Wu B, He M, Zhang Y, Bao J, Contreras LM, Yang S (2018) Advances and prospects in metabolic engineering of Zymomonas mobilis. Metab Eng 50:57-73. [CrossRef]
- Kalnenieks U, Pappas KM, Bettenbrock K (2020) Zymomonas mobilis metabolism: novel tools and targets for its rational engineering. Adv Micr Physiol 77:37-88. [CrossRef]
- Fuchino, K. , Bruheim, P. (2020) An assessment of serial co-cultivation approach for generating novel Zymomonas mobilis strains. BMC Res Notes 13, 422. [CrossRef]
- Strazdina I, Klavins L, Galinina N, Shvirksts K, Grube M, Stalidzans E, Kalnenieks U (2021) Syntrophy of Crypthecodinium cohnii and immobilized Zymomonas mobilis for docosahexaenoic acid production from sucrose-containing substrates. J. Biotechnol 338:63–70. [CrossRef]
- Haffie TL, Louie PW, Khachatourians GG (1985) Isolation of noninhibitory strains of Zymomonas mobilis. Appl Environ Microbiol 49:1007-1009.
- Fu N, Peiris P, Markham J, Bavor J (2009) A novel co-culture process with Zymomonas mobilis and Pichia stipitis for efficient ethanol production on glucose/ xylose mixtures. Enz Microb Technol 45: 210-217. [CrossRef]
- Souza C, Souza LAG (1973) Colpites and vulvovaginitis treatment using Zymomonas mobilis var. recifensis. 8: Rev Inst Antib 13.
- Swings J, De Ley J (1977) The biology of Zymomonas. Bacteriol Rev 41:1-46. [CrossRef]
- Santos JFM, Vasconcelos J, Souza JR, Coutinho EM, Montenegro SML, Azevedo-Ximenes E (2004) The effect of Zymomonas mobilis culture on experimental Shistosoma mansoni infection. Rev Soc Brasil Med Tropical 37:502-504. [CrossRef]
- Weir PM (2016) The ecology of Zymomonas: a review. Folia Microbiol 61:385-392. [CrossRef]
- Rutkis R, Ļaša Z, Rubina M, Ščerbaka R, Kalniņš G, Bogans J, Kalnenieks U (2022) Antimicrobial Activity of Zymomonas mobilis Is Related to Its Aerobic Catabolism and Acid Resistance. Fermentation 8:77. [CrossRef]
- Lima GMS, Souza LIO, Rosato YB, Araújo JM (2011) Detection of bacteriocins in Zymomonas mobilis and RAPD fingerprinting of the producer strains. Afr J Pharmacy Pharmacol 5:2132-2139. [CrossRef]
- Corrêa JAF, Evangelista AG, Nazareth TM, Luciano FB (2019) Fundamentals on the molecular mechanism of action of antimicrobial peptides. Materialia 8:100494. [CrossRef]
- Ramata-Stunda A, Boroduskis M, Kaktina E, Patetko L, Kalnenieks U, Lasa Z, Rubina M, Strazdina I, Kalnins G, Rutkis R (2023) Comparative Evaluation of Existing and Rationally Designed Novel Antimicrobial Peptides for Treatment of Skin and Soft Tissue Infections. Antibiotics 12:551. [CrossRef]
- Savini F, Loffredo MR, Troiano C, Bobone S, Malanovic N, Eichmann TO, Caprio L, Canale VC, Park Y, Mangoni ML, Stella L (2020) Binding of an antimicrobial peptide to bacterial cells: Interaction with different species, strains and cellular components. Biochim Biophys Acta Biomembr 1862:183291. [CrossRef]
- Sajed T, Marcu A, Ramirez M, Pon A, Guo A, Knox C, Wilson M, Grant J, Djoumbou Y, Wishart D (2015) ECMDB 2.0: A richer resource for understanding the biochemistry of E. coli. Nucleic Acids Res 44:D495-D501. [CrossRef]
- Rojewska M, Smułek W, Kaczorek E, Prochaska K (2021) Langmuir Monolayer Techniques for the Investigation of Model Bacterial Membranes and Antibiotic Biodegradation Mechanisms. Membranes 11:707. [CrossRef]
- Hoyo J, Torrent-Burgués J, Tzanov T (2020) Lipid-lipid interactions of Escherichia coli mimetic inner membrane at human physiological temperature. Gen Physiol Biophys., 39(2):195-202. [CrossRef]
- Barrow KD, Collins JG, Rogers PL, Smith GM (1983) Lipid composition of an ethanol-tolerant strain of Zymomonas mobilis. Biochim Biophys Acta 753:324-330. [CrossRef]
- Seelig A (1987) Local Anesthetics and Pressure: A Comparison of Dibucaine Binding to Lipid Monolayers and Bilayers. Biochim Biophys Acta 899:196–204. [CrossRef]
- Kalnenieks U, de Graaf AA, Bringer-Meyer S, Sahm H (1993) Oxidative phosphorylation in Zymomonas mobilis. Arch Microbiol 160:74–79. [CrossRef]
- Wilson DM, Alderete JF, Maloney PC, Wilson TH (1976) Protonmotive force as the source of energy for adenosine 5′- triphosphate synthesis in Escherichia coli. J Bacteriol 126:327-337. [CrossRef]
- Kalnenieks U (2006) Physiology of Zymomonas mobilis: some unanswered questions. Adv Microb Physiol 51:73–117. [CrossRef]
- Li J, Hu S, Jian W, Xie C, Yang X (2021) Plant antimicrobial peptides: structures, functions, and applications. Bot Stud 62: 5. [CrossRef]
- Klubthawee N, Adisakwattana P, Hanpithakpong W, Somsri S, Aunpad R (2020) A Novel, Rationally Designed, Hybrid Antimicrobial Peptide, Inspired by Cathelicidin and Aurein, Exhibits Membrane-Active Mechanisms against Pseudomonas Aeruginosa. Sci Rep 10:9117. [CrossRef]
Figure 1.
Localization of the FITC-labeled RP556 peptide in E. coli (a) and Z. mobilis (b) cells, imaged by confocal microscopy.
Figure 1.
Localization of the FITC-labeled RP556 peptide in E. coli (a) and Z. mobilis (b) cells, imaged by confocal microscopy.
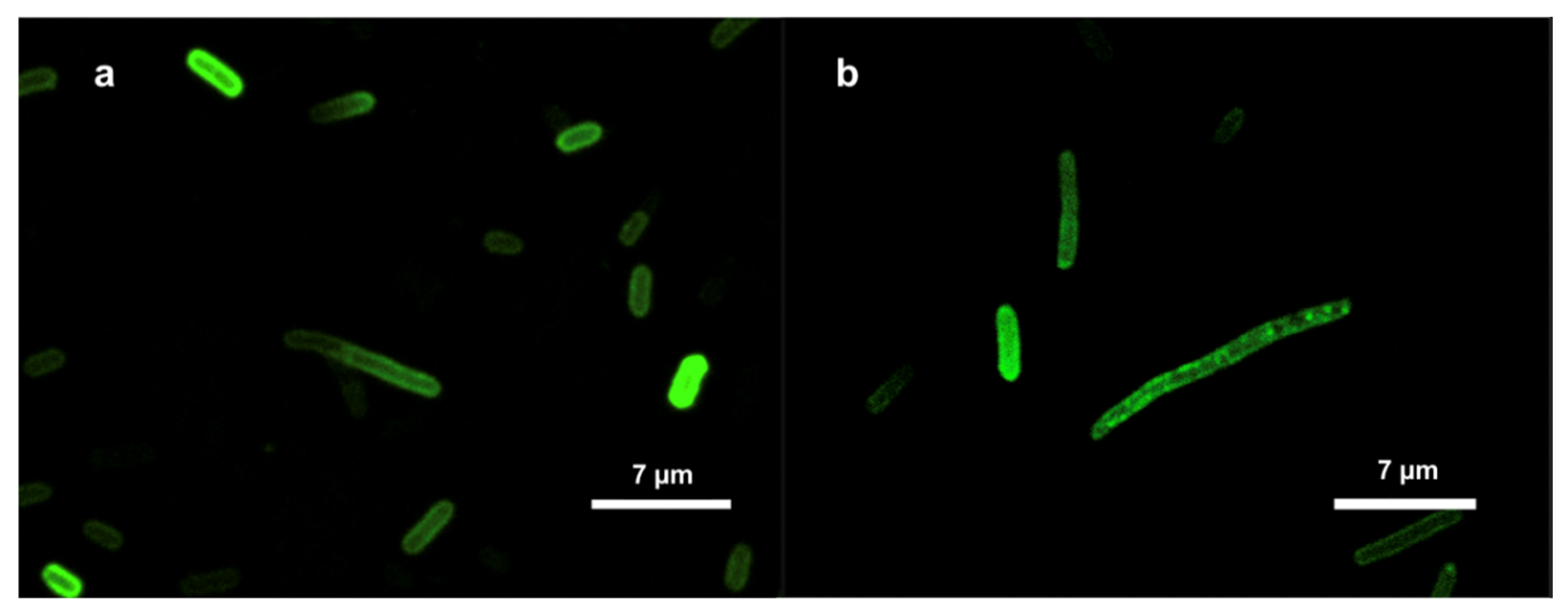
Figure 2.
The π-A isotherms for Z. mobilis model mixtures with peptides R2 (a) and R10 (b). Dashed lines denote the linear part of the π-A isotherms, taken for calculation of the limiting area per molecule (Aπ→0).
Figure 2.
The π-A isotherms for Z. mobilis model mixtures with peptides R2 (a) and R10 (b). Dashed lines denote the linear part of the π-A isotherms, taken for calculation of the limiting area per molecule (Aπ→0).
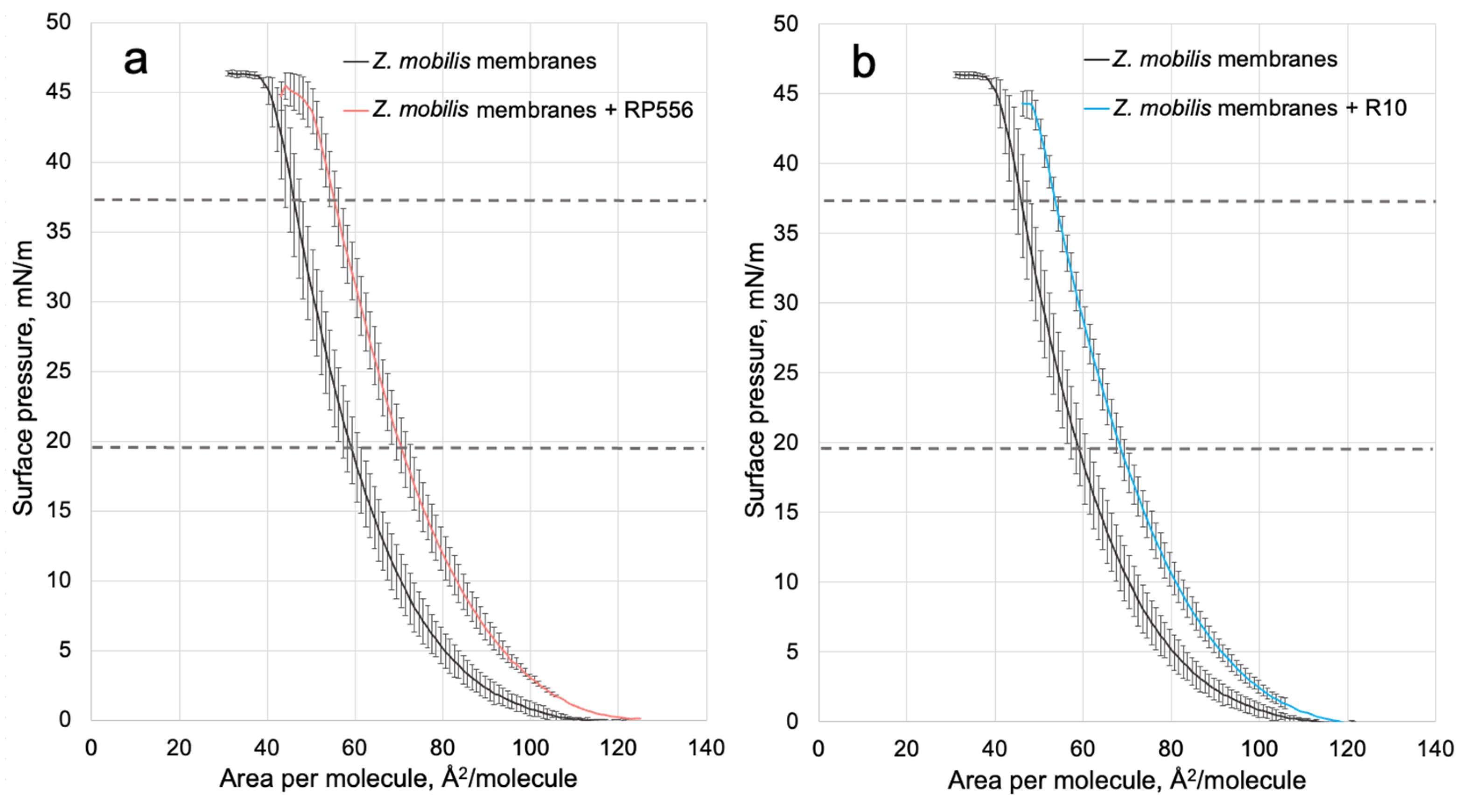
Figure 3.
Time course of the E. coli and Z. mobilis fluorescence ratio between the thermally inactivated cells and extracellular medium.
Figure 3.
Time course of the E. coli and Z. mobilis fluorescence ratio between the thermally inactivated cells and extracellular medium.
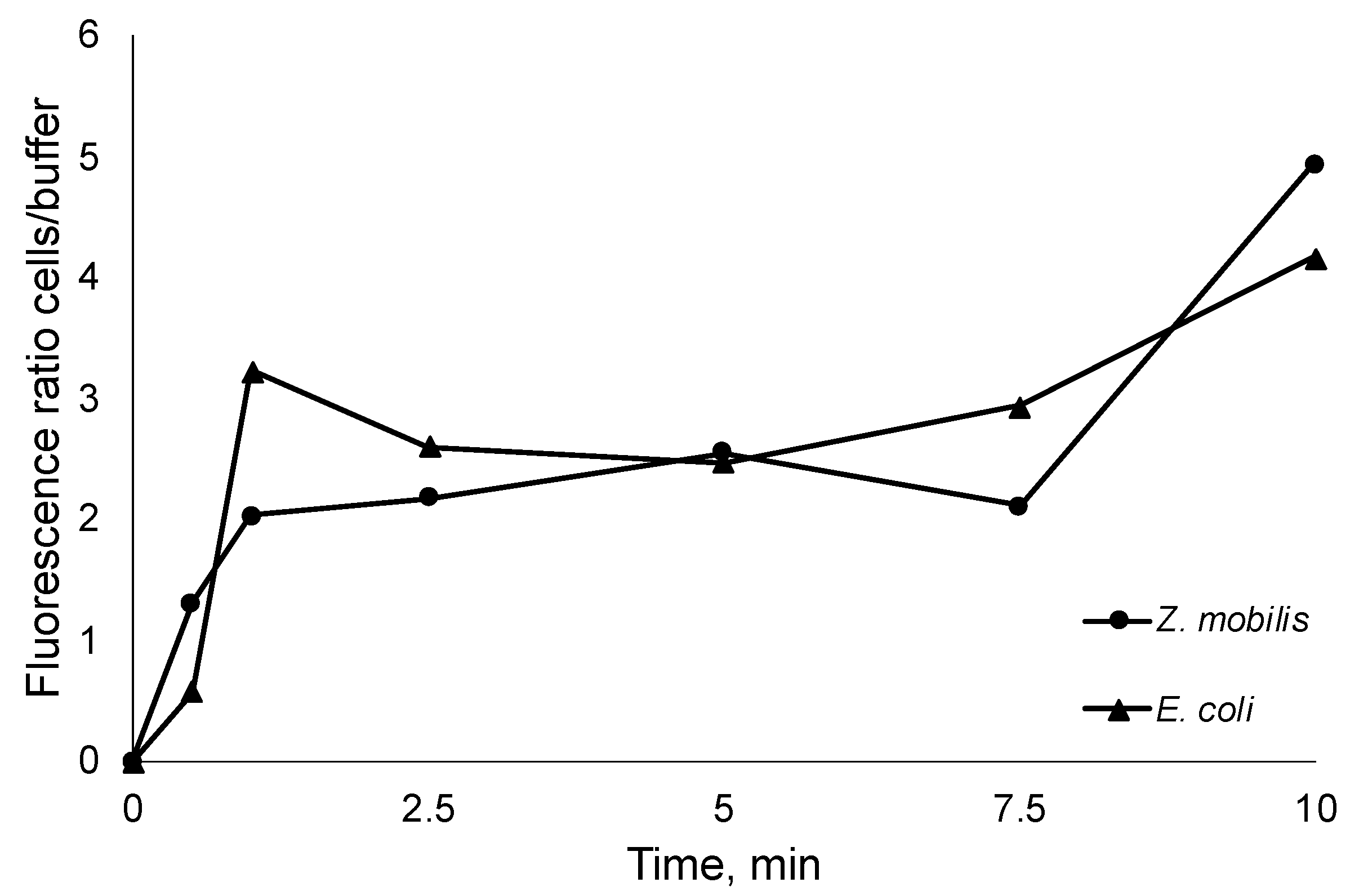
Figure 4.
Synthesis of ATP induced by an artificial pH gradient. In a and b charts peptides were added at the concentration 6.5 E+06 molecules per cell. In c and d charts peptides were added at the concentration 1.27 E+08 molecules per cell. .
Figure 4.
Synthesis of ATP induced by an artificial pH gradient. In a and b charts peptides were added at the concentration 6.5 E+06 molecules per cell. In c and d charts peptides were added at the concentration 1.27 E+08 molecules per cell. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people o Kawada r property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



