
Submitted:
05 April 2024
Posted:
08 April 2024
You are already at the latest version
Abstract
Opuntia ficus-indica (L.) Mill. belongs to the Cactaceae family and the genus Opuntia; it is a succulent plant that adapts to extreme climatic conditions. The aerial part of the plant consists of the cladodes, morphological changes of branches that appear green, are covered with thorns, and are essential to reduce excessive perspiration of water. The composition of cladodes is very varied, and the main constituents are water, fibers, polysaccharides, proteins, fatty acids, vitamins, sterols, minerals, and polyphenols. Polyphenols are responsible for many beneficial activities for human health, such as antioxidant, anti-inflammatory, antitumor, and nutritional properties.
The purpose of this manuscript was to compare the properties of cladodes belonging to the same plant but with different stages of maturity. Relative extracts were tested both in vitro and on a cell line and antioxidant and anti-apoptotic properties were found. The antioxidant activity was tested by the ORAC test, the DPPH test, and the measurement of cellular accumulation of ROS. Anti-apoptotic activity was evaluated by the annexin/PI assay and measurement of caspases 9 and 3 expression.
The results obtained have shown that the extracts considered possess antioxidant and anti-apoptotic properties. However, the different stages of maturity of cladodes are essential for the performance of both functions. In addition, important variations have been made in the dissolution of extracts that have brought greater safety in their use. In conclusion, this manuscript provides further information on cladodes of Opuntia ficus-indica, which can be used as adjuvants in many human pathologies.
Keywords:
Opuntia ficus-indica
; cladodes
; polyphenols
; antioxidant activity
; anti-apoptotic property
1. Introduction
Opuntia ficus-indica (L.) Mill. is a dicotyledonous plant of the category of Angiosperms, belonging to the Cactaceae family and the genus Opuntia. This plant grows preferably in areas with arid and semi-arid climates, like most tropical and subtropical plants [1]. The country of origin of O. ficus-indica is Mexico, where still today the plant is used for gastronomic but also curative purposes [2,3]. However, the geographical areas in which the plant is most present are Africa, the countries of the Mediterranean area, South and Central America, and Australia [4]. O. ficus-indica is a succulent plant that adapts to extreme climatic conditions and grows also in soils with low minerals [5]. Morphological changes of the branches form the aerial part of the plant, called "cladodes", which appear green, covered with thorns, and are indispensable to reducing the excessive transpiration of water [6]. All parts of this plant (including flowers, leaves, and fruits) are used, but in this manuscript, the cladodes will be the main protagonists. The structure of cladodes is a dense molecular network capable of water retention and absorption, which transforms these organs into "tanks" of water. The main constituents of cladodes are water, fibers, polysaccharides, proteins, fatty acids, vitamins, sterols, minerals, and polyphenols [7]. The latter is responsible for many beneficial activities for human health, such as antioxidant, anti-inflammatory, antitumor, and nutritional properties. Polyphenols are secondary metabolites of plants and constitute a vast group of about 10,000 natural products; chemically, they are characterized by at least one aromatic ring linked with other phenolic-, hydroxyl-, carbon- or other chemical groups, and their structure can vary from a simple molecule to an oligomer or polymer complex with high molecular weight [8]. Many foods, such as fruits, vegetables, whole grains, wine, chocolate, and tea are rich sources of polyphenols [9]. The more adopted classification of polyphenols implies the subdivision of these compounds into two main groups: flavonoids (i.e., flavanols, flavanones, flavones, isoflavones, anthocyanidins, and flavan-3-oils) and not flavonoids (i.e., phenolic acids, lignans, condensed and hydrolyzable tannins, stilbenes, hydroxycinnamic and hydroxybenzoic acids) [10]. Polyphenols are produced by plants to protect and defend against pathogens, biotic and abiotic stressors [11]; high daily consumption of these compounds has been linked to reduced risks of many chronic diseases. Polyphenols in cladodes, as already said, are responsible for antioxidant activity both in vitro and in vivo [12] neutralizing free radicals, donating an electron or hydrogen atom, and stopping the oxidative chain reactions [13]. In addition to radical scavenging, polyphenols are also known to affect metal chelation and increase the activity of endogenous antioxidant enzymes such as catalase, superoxide dismutase, and glutathione peroxidase [14]. The excessive production of reactive oxygen or nitrogen species can also determine the occurrence of different inflammatory stimuli with the synthesis and secretion of pro-inflammatory cytokines, such as the nuclear factor kappa B (NF-𝜅B/) or tumor necrosis factor-alpha (TNF-𝛼): polyphenols can modulate these inflammatory processes [15]. Finally, polyphenols have a protective effect against apoptosis [16] and play an important role in some pathologies such as metabolic syndrome, obesity, and hypercholesterolemia [17]. The identification of the polyphenols contained in the cladodes was carried out through HPLC and, among the most known compounds are present: naringin, ferulic acid, rutin, kaempferol-rutinoside, narcissin, tricin, eucomic acid, quercetin, p-coumaric acid 3-O-glucoside, piscidic acid, malic acid, eicosanoic acid [18]. The chemical composition of the cladodes varies according to many factors including climatic conditions, the age of the plant, the cultivar, environmental factors, soil, and salinity, among others [19], demonstrating a purely dynamic attitude. For example, it has been shown that the concentration of calcium ions in cladodes increases with plant maturity [20], while soluble fibers, proteins, and bioactive compounds are more contained in middle-aged cladodes [21]. It has recently been confirmed that young cladodes have a high concentration of carbohydrates [22] and all these considerations show that their composition is dependent on their maturity stage. In previous work, carried out by our research group, it was pointed out how the amount of insoluble fiber contained in the cladodes of O. ficus-indica varies with the harvest season of these organs [23]. With these premises, the present manuscript aims to assess whether the effects of cladodes (collected on the same plant and in the same season) with different stages of maturity, can affect some activities such as antioxidant, anti-inflammatory, and anti-apoptotic. In this direction, smaller and younger cladodes (early-cladodes, e-C), medium-sized cladodes (m-C), and older and larger cladodes (late-cladodes, l-C) were compared. Finally, these extracts were also tested on a cell line (Huvec).
2. Results
2.1. Extracts Obtained from O. ficus-indica
Three extracts have been obtained and tested in this manuscript:
1) e-C obtained from smaller and younger cladodes
2) m-C obtained from medium-sized and age cladodes
3) l-C obtained from the largest and least young cladodes
2.2. Use of Ethanol Instead of Methanol
The corresponding literature indicates that cladodes powder is dissolved in a hydroalcoholic solution of water and methanol (20:80). Since we wanted to remove methanol from O. ficus-indica extracts, we replaced it with ethanol, but to ensure equal solubility efficiency, we had to make further changes in the dissolution protocol. First, we have demonstrated higher cellular toxicity of extracts (e-C, m-C, and l-C) when dissolved in methanol instead of ethanol and this information has been represented in Figure 2. The observed toxicity was not attributable to the extract, as it was also present in untreated cells (vehicle). Samples dissolved in ethanol showed a significant reduction in toxicity compared to those dissolved in methanol. When we dissolved the extracts in a hydroalcoholic solution (water-ethanol 20:80) the solubility was lower than that with methanol and, to make it optimal, it was necessary to raise the temperature to 30 °C.
2.3. Antioxidant Activity
2.3.1. ORAC Test
The ORAC test found a different antioxidant rate in e-C, m-C, and l-C and the results obtained are shown in Figure 3. In panel a, it is highlighted that e-C extract has the lowest antioxidant activity of the three considered, and the corresponding curve is placed just below the curve with a Trolox concentration of 15,25 µg/ml. The m-C extract (panel b) has much higher antioxidant activity, and the corresponding curve is adjacent to the curve with Trolox concentrations of 7.6 µg/ml. In panel c, the l-C curve is slightly lower than the e-C.
2.3.2. DPPH Test
The results of the DPPH test (Figure 4, panels a-c) also showed an excellent radical scavenging effect of the extracts. Different concentrations (0 - 0.25 mg/ml) of extracts were tested and even the lowest were in line with ascorbic acid, used as a positive control. The results were expressed as inhibition % and IC50 value, representing the extract concentration necessary to scavenge the 50% DPPH radicals. In addition, the antioxidant properties of the extracts were confirmed by the IC50 values of 0.054 ± 0.0042 mg/ml; 0.040 ± 0.0027 mg/ml; 0.048 ± 0.0056 mg/ml for e-C, m-C, and l-C respectively. These values showed that m-C was the best extract with the lowest IC50 compared to the others.
2.4. Results on Cell Viability
After conducting several studies on the powder of the different extracts obtained from O. ficus-indica, we decided to evaluate their effects on the Huvec cell line. First, we selected a concentration that could be used without reducing cell viability. As can be seen in Figure 5 panel a, the concentrations between 0,01 and 0,1 mg/ml of e-C, m-C, and l-C did not cause cell mortality and extracts behaved in the same way. On the contrary, only the concentration of 0.2 mg/ml showed a slight reduction in viability. In panel b, the treatment with H2O2 (50 µM, 9 hours) resulted in a significant reduction of cell viability, compared to untreated cells. Pre-treatment with the considered extracts, followed by exposure to H2O2, has shown important variations: the concentration 0.02 mg/ml is the one that protects better than the others. In addition, once again the extract m-C had a greater action than e-C and l-C. The results obtained have suggested to use of the extracts at the concentration of 0.02 mg/ml.
2.5. O. ficus-indica Extracts Reduce the Accumulation of ROS in the Huvec Cell Line
To evaluate the antioxidant potential of the extracts of O. ficus-indica, we have also measured the accumulation of ROS on cell cultures pre-treated with e-C, m-C, and l-C and successively exposed to hydrogen peroxide (150 µM, 20 min). This study was carried out using cytofluorimetric readings, which allowed us to appreciate the displacement of fluorescence about the content of ROS. Panel a of Figure 6, highlighted the results obtained from the treatment with the extracts: the pre-treatment with e-C and l-C has reduced ROS generated from the exposition to H2O2, which were however significantly more abundant than the untreated cells. m-C has strongly reduced ROS compared to e-C and l-C. In panel b, the respective quantification was shown.
2.6. O. ficus-indica Extracts and Apoptosis
To evaluate the effect of extracts in a model of induced inflammation, the cells were treated with lipopolysaccharide (LPS, 1 µg/mL for 24 h), one of the components of the outer layer of the cell wall of gram-negative bacteria, capable of causing strong immune responses. As can be seen in Figure 7 panel a, the annexin/PI test was used to assess viability or type of cell death resulting from appropriate treatments. Prolonged cellular treatment with LPS induces irreparable damage that can be assessed as apoptotic death. Untreated cells or those exposed only to the extracts of cladodes (e-C, m-C, and l-C) showed no death and the cells appeared fully viable. Apoptotic cell death induced by LPS has been evidenced by the shift of cells from Q1 to Q3. Surprisingly pre-treatment with l-C, before exposure to LPS, resulted in substantial cellular protection visible through reduced apoptosis and enrichment of viable cells. Pre-treatment with e-C did not reduce cell death at all, while m-C generated an intermediate effect between e-C and l-C, reducing apoptosis and increasing cell viability: however, both variations were only slight. In panel b, the respective quantification is shown.
2.7. O. ficus-indica Extracts and Caspases 9 and 3
The expression of two of the main apoptotic markers (cleaved fraction of caspase 9 and 3) was evaluated by cytofluorescence, as can be seen in Figure 8, panels a and b respectively. In the first box of panel a, the difference in expression of caspase 9, between untreated cells and those exposed to LPS, was highlighted. Inflammatory treatment has caused a shift of fluorescence to the right, which can be interpreted as an increased expression of the apoptotic marker. While e-C and m-C cannot revert the increased expression of caspase 9, l-C extract is the only one that significantly reduces LPS-induced damage. In panel b, the expression of cleaved caspase 3 has been reported. In this case, already pre-treatment with e-C and m-C reduces the expression of caspase, but l-C can report its expression levels similar to those of untreated cells. Summing up, it is possible to say that in the case of induced apoptosis (Figure 7 and Figure 8), l-C extract is the one that works most significantly reducing the induced damage.
2.8. Determination of Total Phenolic and Flavonoid Content
Figure 9, panels a and b, shows the total content of polyphenols and flavonoids respectively, present in the tested extracts. As can be seen in the figure, the extracts e-C, m-C, and l-C have the same amount of both polyphenols and flavonoids.
3. Discussion
The first important result of this work was to make the extracts (e-C, m-C, and l-C) less toxic: the literature indicated as preferential protocol their dissolution in a hydroalcoholic solution consisting of water and methanol (ratio 20:80). However, the presence of methanol increased the level of toxicity and the risk of not being able to administer e-C, m-C, and l-C at high concentrations. Methanol is the simplest of alcohols and its chemical formula is CH3OH; it is very volatile, extremely flammable, and toxic inducing depression of the central nervous system and damage to the optic nerve and retina [24]. Ethanol (CH3CH2OH) is very similar to methanol but contains one carbon and two other hydrogen atoms in its molecule. Despite the minimal structural difference, ethanol is less toxic and safer than methanol [25]. The lower toxicity of ethanol suggested replacing methanol with this alcohol. However, the simple substitution was not sufficient to guarantee the same rate of solubility and it was necessary to help the dissolution process by increasing the temperature up to 30 °C.
The results obtained in this manuscript have indicated that specific stages of maturity of cladodes of O. ficus-indica influence the aptitude to specific properties. For example, m-C exercises antioxidant properties and this potential is absent in both e-C and l-C demonstrating a condition of uniqueness only for m-C. Similarly, l-C has an anti-apoptotic activity that is essentially typical of this extract. The antioxidant property can presumably be carried out by polyphenols, as widely demonstrated and particularly abundant in cladodes [26,27,28]. It is important to remember that plants are known to be sources of antioxidant compounds and that their amounts depend on a variety of conditions that can increase or reduce their content, such as climate, soil, and stress factors, among others [29,30]. The observed apoptotic activity is a consequence of an inflammatory stimulus-induced: the cells were treated with LPS (1 µg/mL for 24 h) and the pro-inflammatory prolonged process caused irreversible damage resulting in apoptotic death. Polyphenols seem also to be widely involved in inflammatory protection [31,32] and therefore they could be responsible for the property carried out by l-C. Given these considerations, it would be expected that the polyphenolic content would be different in all the fractions considered and probably higher in m-C and l-C. On the contrary, surprisingly the content of polyphenols and flavonoids was unchanged in e-C, m-C, and l-C. The only explanation capable of justifying this phenomenon is that a qualitative variation of the polyphenols occurs instead of a quantitative one. In this direction, it is likely to imagine that m-C is made up of polyphenols that more and better perform the antioxidant functions while l-C would have a polyphenolic content more capable of performing an anti-inflammatory and anti-apoptotic activity; this substantial qualitative difference would not involve the polyphenolic content. It is important not to forget that polyphenols are secondary metabolites of plants that play a key role in both their defense and signaling [33], by organizing defense mechanisms against herbivores, pathogenic bacteria, fungi, viruses, and parasites, developing tolerance to various forms of stress and fostering relations with pollinating insects [34]. Cladodes are therefore likely to produce specific polyphenols depending on the stage of maturity. Another hypothesis capable of explaining the qualitative polyphenolic variations of plants is that some physical characteristics (the age of the plant or seasonality) can modulate gene expression, increasing the formation of specific secondary metabolites [35,36]. In conclusion, it is possible to say that further trials must be carried out on this subject, but if the results obtained were confirmed, the use of O. ficus-indica could increase and this plant could guarantee specific remedies for human health. In addition, it would be desirable shortly to identify which polyphenols are directly responsible.
4. Materials and Methods
4.1. Plant Materials and Sample Preparation
The cladodes of Opuntia ficus-indica were collected at Roccelletta di Borgia, Calabria, Italy (February 2023, Temperature: 10° C): latitude 31, 57' 22'' N, longitude 12,23, 21' 38’’E, from the same plant, but with different degrees of maturity. It has been useful to distinguish between early, medium, and late cladodes. To respect this condition, some parameters have been chosen such as the number of days of presence on the plant, the size, the different weights, and its length. These parameters are reported and listed in Table 1. The cladodes chosen were collected, and washed with distilled water; the plugs were removed and cut into pieces of about 1 cm. Subsequently, the cladodes were dried in a laboratory oven (ENCO, Venice, Italy), at a temperature of 40 °C for 4 days; the dried pieces were ground (2000 W, velocity 25,000 r/min, running time 5 min with an interval time of 10 min) and sieved through 200 µm mesh sieves to obtain a green powder with an intermediate granulometry. The powder obtained was stored in an airtight glass bottle at 4° C, away from light and moisture, until further use [23]. At the time of use, the powder was dissolved in a solution of water and ethanol (20:80), at 30 ° C, and the mixture obtained was stirred for 2 hours in the dark. Using this approach, three extracts were obtained and tested: e-C, m-C, and l-C.
4.2. Antioxidant Activity
4.2.1. The ORAC Assay
The antioxidant activity of e-C, m-C, and l-C was determined by the Oxygen Radicals Absorbance Capacity (ORAC) assay. The loss of fluorescence of fluorescein (used as a probe) is measured over time and this fluorescence is due to the formation of peroxylic radicals, following the spontaneous degradation of 2,2'-azobis-2-methyl-propanimidamide, dihydrchloride (AAPH). The peroxylic radical oxidizes the fluorescein, causing the gradual loss of the fluorescence signal. Antioxidants suppress this reaction and inhibit signal loss: 6-hydroxy-2,5,7,8-tetramTethylchroman-2-carboxylic acid (Trolox) is a water-soluble analog of vitamin E that inhibits fluorescence decay in a dose-dependent manner. Finally, e-C, m-C, and l-C were used at a concentration of 0.02 mg/ml. The evaluation of fluorescent decay for fluorescein was conducted using 485 and 520 nm as excitation and emission wavelengths respectively. The temperature was maintained at 37 ° C. Measurements were made in triplicate, and data obtained from fluorescence vs. time curves are reported as the average antioxidant efficacy of the antioxidant compound. A regression equation was constructed by comparing the net area below the fluorescein decay curve and the Trolox concentration. The area under the curve has been calculated with the following equation:
i = 90
AUC = 1 + Σ f1/f1
i = 1
4.2.2. DPPH Test
The antioxidant activity of the extracts obtained from the cladodes of O. ficus-indica was measured using the stable radical 2,20 -difenyl-1 picrylhydrazyl (DPPH). Experimentally, 850 µl of DPPH solution was added to 50 µl of various extract concentrations (0.02-0.25 mg/ml), keeping the mixture in the dark for 20 min. Subsequently, the absorbance was read by a UV-Vis spectrophotometer (Multiskan GO, Thermo Scientific, Denver, CO, USA) at 517 nm, room temperature, and its reduction in absorbance was visible as a change in color from purple to yellow. The results obtained were expressed in % inhibition and IC50. The latter represents the concentration of matter in powder required to remove 50% of DPPH radicals.
4.3. Cell Cultures
The Human Endothelial Cell line (Huvec) has been isolated from the vein of the umbilical cord and acquired by the American Type Culture Collection (ATCC, 20099 Sesto San Giovanni, Milan, Italy). Huvec cells were grown in culture in the modified Dulbecco Eagle medium (DMEM), (enriched with 100 U/ml penicillin, 100 µg/ml streptomycin, and 10% fetal bovine serum) and kept in a humidified 5% CO2 atmosphere at 37% C. The medium was changed every 2-3 days and when the cell lines reached 50% of the confluence, they were treated with e-C, m-C, and l-C (0.02 mg/ml) for 24 hours or pre-treated with these extracts (0.02 mg/ml) for 24 hours and then exposed to H2O2 50µM for 9 h or to LPS 1 µg/ml for 24 h.
4.4. Proliferation Assay
Cell proliferation was measured through the 3-(4,5-dimethyl-2-yl)-2,5-diphenyltetrazole bromide (MTT) test; this dye, when contained in living cells with functioning mitochondria, can be reduced resulting in colorimetric variation. MTT reduction measurement provides information on cell viability and metabolic activity. 8 x 103 cells were plated in 96-well plates and, after 24 hours, the culture medium was replaced with fresh medium containing e-C, m-C, or l-C for 24 hours. At the end of treatment time, the medium was replaced with a phenol-free medium containing a solution of MTT (0,5 mg/ml) and after 4h incubation, 100 µl of 10% SDS were added to each well to solubilize the formazan crystals. Finally, the optical density was measured at wavelengths of 540 and 690 nm using a spectrophotometer reader (X MARK Microplate Bio-Rad Spectrophotometer).
4.5. ROS Measurement in the Huvec Cell Line
The H2DCF-DA probe can be used to measure reactive oxygen species accumulated in cells, taking advantage of its peculiarities: 1) it easily enters cells: 2) H2DCF-DA is cleaved, by intracellular esterase, to form H2DCF; 3) H2DCF is no longer able to leave the cells; 4) If H2DCF is oxidized, it binds to the ROS forming the compound highly fluorescent DCF. The quantification of the DCF probe provides the content of ROS in the cell. Experimentally, 6 x 104 cells were plated in 96-well microplates, and the following day, Huvec cells were pre-treated with e-C, m-C, or l-C for 24 hours at the concentration of 0.02 mg/ml for 24h. At the end of the treatment, the growth medium was replaced by a phenol-free fresh medium containing H2DCF-DA (25µM) and, after 30 minutes of exposure to 37 °C, the cells were washed with PBS, centrifuged, resuspended in PBS, and exposed or not at H2O2 (100 µM, 20 min). Finally, the fluorescence was evaluated by cytometric analysis (FACS Accury, Becton Dickinson, Franklin Lakes, NJ, USA).
4.6. Annexin V/PI Staining
After treating the cells as previously indicated, trypsinized and washed with PBS, the Huvec cells were resuspended in 1x Binding Buffer (Metabolic Activity/AnnexinV/Dead Cell Apoptosis Kit). One hundred microliters of the suspension were transferred to a 5 ml culture tube and 5 µl of FITC Annexin V (BD Biosciences, San Jose, CA, USA) was added. The samples were vortexated and incubated for 15 minutes in the dark, at 25° C. Finally, 400 µl of 1 binding buffer and 5 µl of propyl iodide (PI) were added to each tube and samples were analyzed with flow cytometry (Accury FACS, Becton Dickinson laser cytometer).
4.7. Measurement of Protein Expression through Immuno-Cytofluorometry
To measure the expression of caspases 9 and 3, monoclonal antibodies, linked to fluorochromes, have been used and the samples have been analyzed through cytofluorometry. The cells were washed with PBS, trypsinized, and collected in a growth medium in cytofluorometer tubes. Subsequently, they were treated with BSA in 10% animal serum and kept for 60 min at room temperature to block non-specific sites. At the end of the indicated time, the cells were incubated for 2 hours at 37° C with the desired antibodies. For caspase 9, a rabbit monoclonal antibody (2D5) was purchased from Invitrogen, at 1:200 dilution. For caspase 3, a rabbit monoclonal antibody (E87) was purchased from Abcam (197202) at 1:200 dilution. After appropriate washing with PBS to remove excess primary antibodies, secondary antibodies, conjugated with Fitc, diluted in 5% animal serum for 1 hour at room temperature, and 1:500 dilution, were added. After further washing in PBS, the cytofluorometer reading was carried out.
4.8. Determination of Total Phenolic and Flavonoid Content
The total content of polyphenols in extracts was calculated using the Folin-Ciocalteu colorimetric assay. First, several solutions of gallic acid with different concentrations (0, 25, 50, 100, 200, and 300 µg/ml) were used. Subsequently, 1g of powdered cladodes of O. ficus-indica was mixed with 20 ml ethanol/water 80:20 (w/w) and this mixture was shaken for 24 hours. 400 μl of the mixture were put in one cuvette and 0.8 ml of Folin-Ciocalteu reagent diluted 10 times were added. After 3 minutes of stirring, 0.8 ml of sodium carbonate 7% (w/v) was added and the resulting mixture was left to stand for a further 2 hours and stirred constantly until color developed. The relative absorbance was measured at 760 nm with a Prism V-1200 spectrophotometer and the phenolic content was determined by the linear equation of the standard curve. The total phenolic content of the extract was expressed in mg Gallic acid equivalent (GAE)/g dry weight. For determination of total flavonoid content, the aluminium-chloride colorimetric test was used. 1 ml of extract was mixed with 1 ml of 2% aluminum chloride in methanol. After 30', the absorbance at 430 nm has been measured and quercetin equivalents per gram of extract (mg QE/g extract) are used to represent the estimated flavonoid content.
Author Contributions
: J.M. and V.M. have conceptualized the manuscript. S.N., M.S., and R.C. have validated and curated the manuscript. F.O., R.M., and F.S. have participated in the original draft preparation. J.M., C.M., E.P., S.R., and V.M. have written and supervised the manuscript.
Funding
The work was supported by public resources from the Italian Ministry of Research.
Acknowledgments
This work was supported by PON-MIUR 03PE000_78_1, PON-MIUR 03PE000_78_2 and PONa3 00359.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pimienta-Barrios, E. Prickly pear (Opuntia spp.): a valuable fruit crop for the semiarid land of Mexico. J Arid Environ 1994, 28, 1–11. [Google Scholar] [CrossRef]
- de Albuquerque, J.G.; de Souza Aquino, J.; de Albuquerque, J.G.; de Farias, T.G.S.; Escalona-Buendía, H.B.; Bosquez-Molina, E.; Azoubel, P.M. Consumer perception and use of nopal (Opuntia ficus-indica): A cross-cultural study between Mexico and Brazil. Food Res Int. 2019, 124, 101–108. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, H.I.; Elemlegy, A.A.; Eldesoky, S.E.; Safwat, G. Phytochemical screening, antimicrobial, antioxidant, anticancer activities and nutritional values of cactus (Opuntia ficus indica) pulp and peel. Fresenius Environ. Bull. 2019, 28, 1534–1551. [Google Scholar]
- Piga, A. Cactus Pear: A fruit of nutraceutical and functional importance. J. Prof. Assoc. Cactus Dev. 2004, 6, 9–22. [Google Scholar]
- Aragona, M.; Lauriano, E.R.; Pergolizzi, S.; Faggio, C. Opuntia ficus-indica (L.) Miller as a source of bioactivity compounds for health and nutrition. Natural product research 2018, 32, 2037–2049. [Google Scholar] [CrossRef]
- Sinicropi, M.S.; Baldino, N.; Ceramella, J.; Iacopetta, D.; Scali, E.; Basile, G.; Saturnino, C.; Catalano, A. Opuntia ficus indica (L.) Mill. An Ancient Plant Source of Nutraceuticals. Curr Top Med Chem. 2022, 22, 1736–1749. [Google Scholar] [CrossRef] [PubMed]
- Perucini-Avendaño, M.; Nicolás-García, M.; Jiménez-Martínez, C.; de Jesús Perea-Flores, M.; Gómez-Patiño, M.B.; Arrieta-Báez, D.; Dávila-Ortiz, G. Cladodes: Chemical and structural properties, biological activity, and polyphenols profile. Food Sci Nutr 2021, 9, 4007–4017. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: chemistry, bioavailability and effects on health. Nat Prod Rep 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- El Gharras, H. Polyphenols: Food sources, properties and applications. A review. International Journal of Food Science and Technology, 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid Redox Signal 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- López-Romero, P.; Pichardo-Ontiveros, E.; Avila-Nava, A.; Vázquez-Manjarrez, N.; Tovar, A. R.; Pedraza-Chaverri, J.; Torres, N. The effect of nopal (Opuntia ficus indica) on postprandial blood glucose, incretins, and antioxidant activity in Mexican patients with type 2 diabetes after consumption of two different composition breakfasts. Journal of the Academy of Nutrition and Dietetics 2014, 114, 1811–1818. [Google Scholar] [CrossRef]
- Avila-Nava, A.; Calderón-Oliver, M.; Medina-Campos, O.N.; Zou, T.; Gu, L.; Torres, N.; Tovar, A.R.; Pedraza-Chaverri, J. Extract of cactus (Opuntia ficus indica) cladodes scavenges reactive oxygen species in vitro and enhances plasma antioxidant capacity in humans. Journal of Functional Foods 2014, 10, 13–24. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Guo, H.; Lou, H. Grape seed polyphenols protect cardiac cells from apoptosis via induction of endogenous antioxidant enzymes. J. Agric. Food Chem. 2007, 55, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Salzano, S.; Checconi, P.; Hanschmann, E-M. ; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S. et al. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc Natl Acad Sci U S A 2014, 111, 12157–12162. [Google Scholar] [CrossRef]
- Alimi, H.; Hfaeidh, N.; Bouoni, Z.; Sakly, M.; Rhouma, K.B. Ameliorative effect of Opuntia ficus indica juice on ethanol-induced oxidative stress in rat erythrocytes. Experimental and Toxicologic Pathology 2013, 65, 391–396. [Google Scholar] [CrossRef]
- Petruk, G.; Di Lorenzo, F.; Imbimbo, P.; Silipo, A.; Bonina, A.; Rizza, L.; Piccoli, R.; Monti, D.M.; Lanzetta, R. Protective effect of Opuntia ficus-indica L. cladodes against UVA-induced oxidative stress in normal human keratinocytes. Bioorg Med Chem Lett. 2017, 27, 5485–5489. [Google Scholar] [CrossRef] [PubMed]
- Abbas, E.Y.; Ezzat, M.I.; El Hefnawy, H.M.; Abdel-Sattar, E. An overview and update on the chemical composition and potential health benefits of Opuntia ficus-indica (L.) Miller. J Food Biochem. 2022, 46, e14310. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Sobeh, M.; Badr, W.K.; Abdelfattah, M.A.O.; Ali, Z.Y.; El-Tantawy, M.E.; Rabeh, M.A.; Wink, M. HPLC-PDA-MS/MS profiling of secondary metabolites from Opuntia ficus-indica cladode, peel and fruit pulp extracts and their antioxidant, neuroprotective effect in rats with aluminum chloride induced neurotoxicity. Saudi J Biol Sci 2020, 27, 2829–2838. [Google Scholar] [CrossRef]
- Rykaczewski, K.; Jordan, J.S.; Linder, R.; Woods, E.T.; Sun, X.; Kemme, N.; Manning, K.C.; Cherry, B.R.; Yarger, J.L.; Majure, L.C. Microscale Mechanism of Age Dependent Wetting Properties of Prickly Pear Cacti (Opuntia). Langmuir 2016, 32, 9335–9341. [Google Scholar] [CrossRef]
- Hernández-Becerra, E.; Gutiérrez-Cortez, E.; Del Real, A.; Rojas-Molina, A.; Rodríguez-García, M.; Rubio, E.; Quintero-García, M.; Rojas-Molina, I. Bone Mineral Density, Mechanical, Microstructural Properties and Mineral Content of the Femur in Growing Rats Fed with Cactus Opuntia ficus indica (L.) Mill. (Cactaceae) Cladodes as Calcium Source in Diet. Nutrients 2017, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Mounir, B.; Younes, E.G.; Asmaa, M.; Abdeljalil, Z.; Abdellah, A. Physico-chemical changes in cladodes of Opuntia ficus-indica as a function of the growth stage and harvesting areas. J Plant Physiol. 2020, 251, 153196. [Google Scholar] [CrossRef] [PubMed]
- Caminiti, R.; Serra, M.; Nucera, S.; Ruga, S.; Oppedisano, F.; Scarano, F.; Macrì, R.; Muscoli, C.; Palma, E.; Musolino, V.; Statti, G.; Mollace, V.-; Maiuolo, J. Antioxidant Activity and Seasonal Variations in the Composition of Insoluble Fiber from the Cladodes of Opuntia ficus-indica (L.) Miller: Development of New Extraction Procedures to Improve Fiber Yield. Plants 2024, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Kotikova, K.; Klepis, P.; Ridzon, P.; Hlusicka, J.; Navratil, T.; Rulisek, J.; Zak, I.; Zakharov, S. Peripheral polyneuropathy after acute methanol poisoning: Six-year prospective cohort study. Neurotoxicology. 2020, 79, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Toxicology and the biological role of methanol and ethanol: Current view. Biomed Pap. Med Fac Univ Palacky Olomouc Czech Repub 2016, 160, 54–63. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Izuegbuna, O.; Otunola, G.; Bradley, G. Chemical composition, antioxidant, anti-inflammatory, and cytotoxic activities of Opuntia stricta cladodes. PLoS One. 2019, 14, e0209682. [Google Scholar] [CrossRef] [PubMed]
- Perucini-Avendaño, M.; Perea-Flores, M.J.; Gómez-Patiño, M.B.; Arrieta-Báez, D.; Dávila-Ortiz, G. Cladodes: Chemical and structural properties, biological activity, and polyphenols profile. Food Sci Nutr. 2021, 9, 4007–4017. [Google Scholar] [CrossRef] [PubMed]
- Witkowska-Banaszczak, E.; Radzikowska, D.; Ratajczak, K. Chemical profile and antioxidant activity of Trollius europaeus under the influence of feeding aphids. Open Life Sci. 2018, 13, 312–318. [Google Scholar] [CrossRef]
- Vanacker, H.; Guichard, M.; Bohrer, A.-S.; Issakidis-Bourguet, E. Redox Regulation of Monodehydroascorbate Reductase by Thioredoxin y in Plastids Revealed in the Context of Water Stress. Antioxidants 2018, 7, 183. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients. 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid Med Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Kowalczewski, P.; Radzikowska, D.; Ivanišová, E.; Szwengiel, A.; Kačániová, M.; Sawinska, Z. Influence of Abiotic Stress Factors on the Antioxidant Properties and Polyphenols Profile Composition of Green Barley (Hordeum vulgare L.). Int J Mol Sci 2020, 21, 397. [Google Scholar] [CrossRef] [PubMed]
- Fejér, J.; Gruľová, D.; Eliašová, A.; Kron, I. Seasonal Variability of Juniperus communis L. Berry Ethanol Extracts: 2. In Vitro Ferric Reducing Ability of Plasma (FRAP) Assay. Molecules. 2022, 27, 9027. [Google Scholar] [CrossRef]
- Scioneaux, A.N.; Schmidt, M.A.; Moore, M.A.; Lindroth, R.L.; Wooley, S.C.; Hagerman, A.E. Qualitative variation in proanthocyanidin composition of Populus species and hybrids: genetics is the key. J Chem Ecol 2011 37, 57–70. [CrossRef]
- Murai, Y.; Setoguchi, H.; Ono, E.; Iwashina, T. Flavonoids and their qualitative variation in Calystegia soldanella and related species (Convolvulaceae). Nat Prod Commun 2015, 10, 429–432. [Google Scholar] [CrossRef]
Figure 1.
Conceptual map of the manuscript. Panel a describes the collection of cladodes from the same plant of O. ficus-indica: 1) a small and young cladode was chosen and collected (e-C); 2) a medium-sized and age cladode was chosen and collected (m-C); 3) a large and least young cladode was chosen and collected (l-C). Panel b highlights the continuation of work: e-C, m-C, and l-C have been obtained as described and tested for antioxidant and anti-apoptotic properties.
Figure 1.
Conceptual map of the manuscript. Panel a describes the collection of cladodes from the same plant of O. ficus-indica: 1) a small and young cladode was chosen and collected (e-C); 2) a medium-sized and age cladode was chosen and collected (m-C); 3) a large and least young cladode was chosen and collected (l-C). Panel b highlights the continuation of work: e-C, m-C, and l-C have been obtained as described and tested for antioxidant and anti-apoptotic properties.
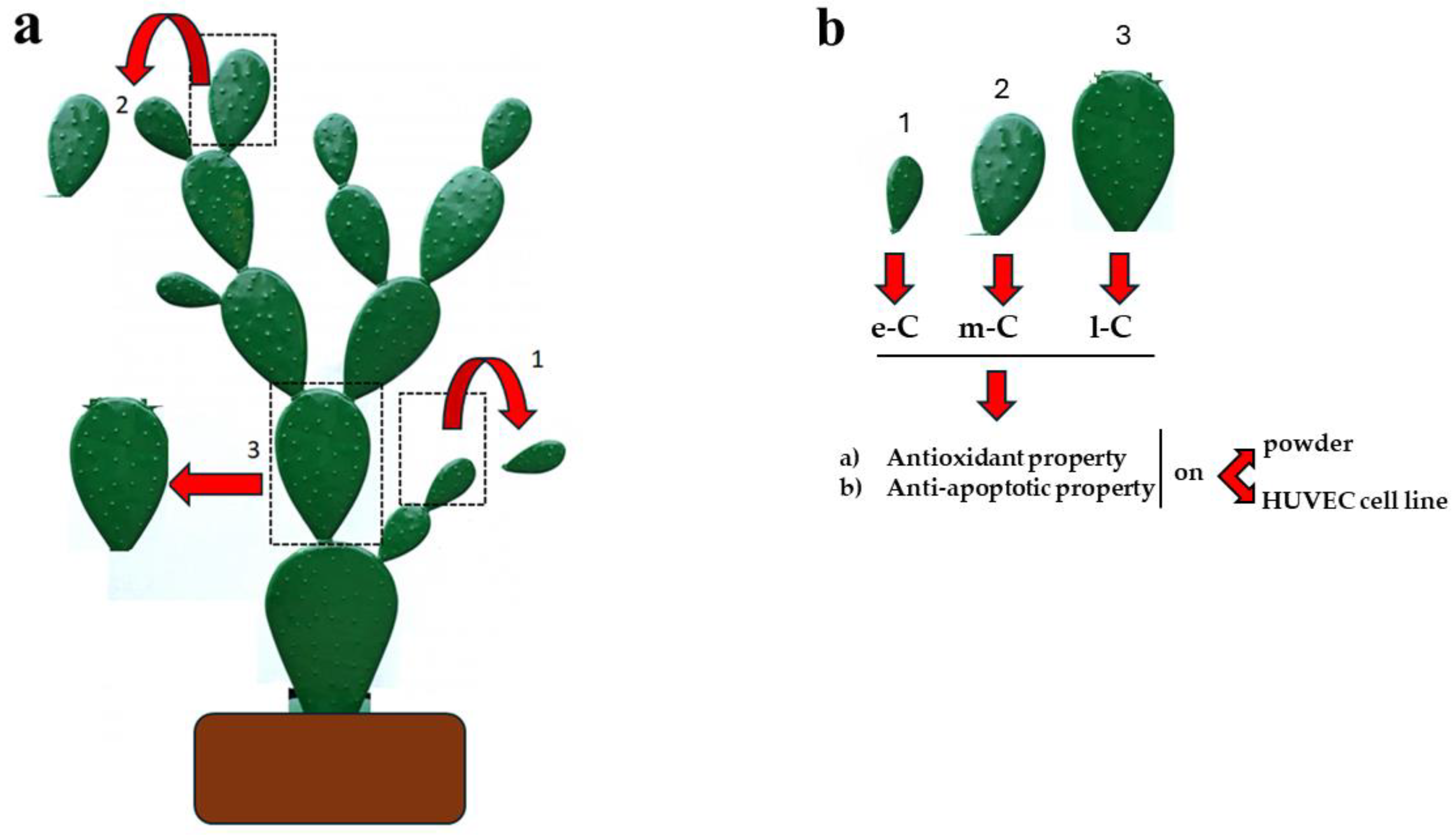
Figure 2.
Comparison of extracts dissolved in methanol or ethanol. The extracts obtained from the cladodes of O. ficus-indica were dissolved in two hydroalcoholic solutions (1) water-methanol 20:80 or 2) water-ethanol 20:80, and their viability has been measured. As can be seen, all the extracts dissolved in the ethanol solution are significantly less toxic than the counterparts dissolved in the methanol solution. This difference does not depend on the composition of the extracts, as demonstrated by the same reaction in untreated cells in which the same amount of alcohol used for extracts is present. Three independent experiments were performed, and the values were expressed as the mean ± sd. § denotes p < 0.05 vs. the counterparty dissolved in methanol. Variance analysis (ANOVA) was followed by a Tukey–Kramer comparison test.
Figure 2.
Comparison of extracts dissolved in methanol or ethanol. The extracts obtained from the cladodes of O. ficus-indica were dissolved in two hydroalcoholic solutions (1) water-methanol 20:80 or 2) water-ethanol 20:80, and their viability has been measured. As can be seen, all the extracts dissolved in the ethanol solution are significantly less toxic than the counterparts dissolved in the methanol solution. This difference does not depend on the composition of the extracts, as demonstrated by the same reaction in untreated cells in which the same amount of alcohol used for extracts is present. Three independent experiments were performed, and the values were expressed as the mean ± sd. § denotes p < 0.05 vs. the counterparty dissolved in methanol. Variance analysis (ANOVA) was followed by a Tukey–Kramer comparison test.
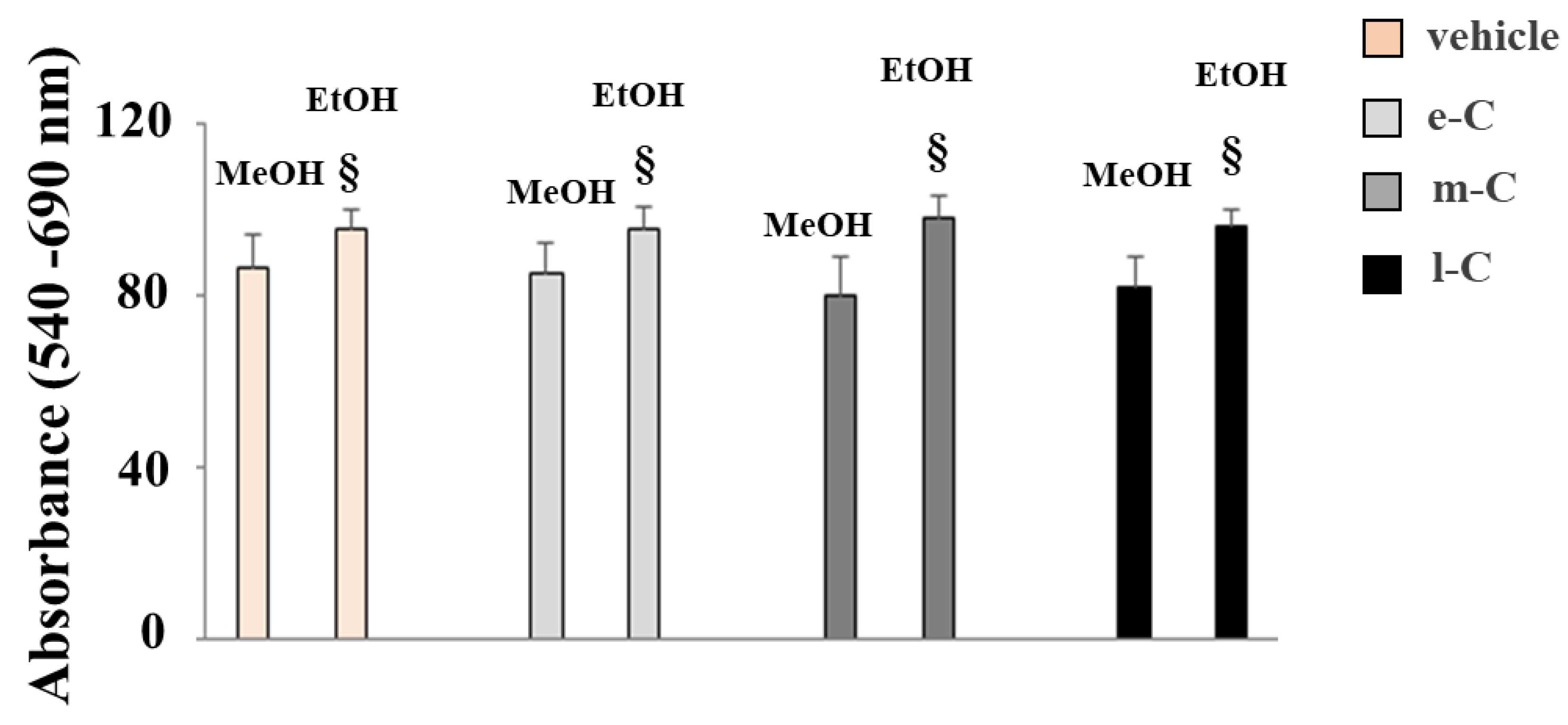
Figure 3.
ORAC assay. The fluorescence decay curves for e-C, m-C, and l-C, (from a representative experiment) were shown in the a, b, and c panels respectively.
Figure 3.
ORAC assay. The fluorescence decay curves for e-C, m-C, and l-C, (from a representative experiment) were shown in the a, b, and c panels respectively.

Figure 4.
DPPH assay. Antiradical activity (% inhibition) and IC50 of e-C, m-C, and l-C are highlighted and compared to positive control (ascorbic acid). Nonlinear regression was used to calculate IC50 values. A representative experiment is shown.
Figure 4.
DPPH assay. Antiradical activity (% inhibition) and IC50 of e-C, m-C, and l-C are highlighted and compared to positive control (ascorbic acid). Nonlinear regression was used to calculate IC50 values. A representative experiment is shown.
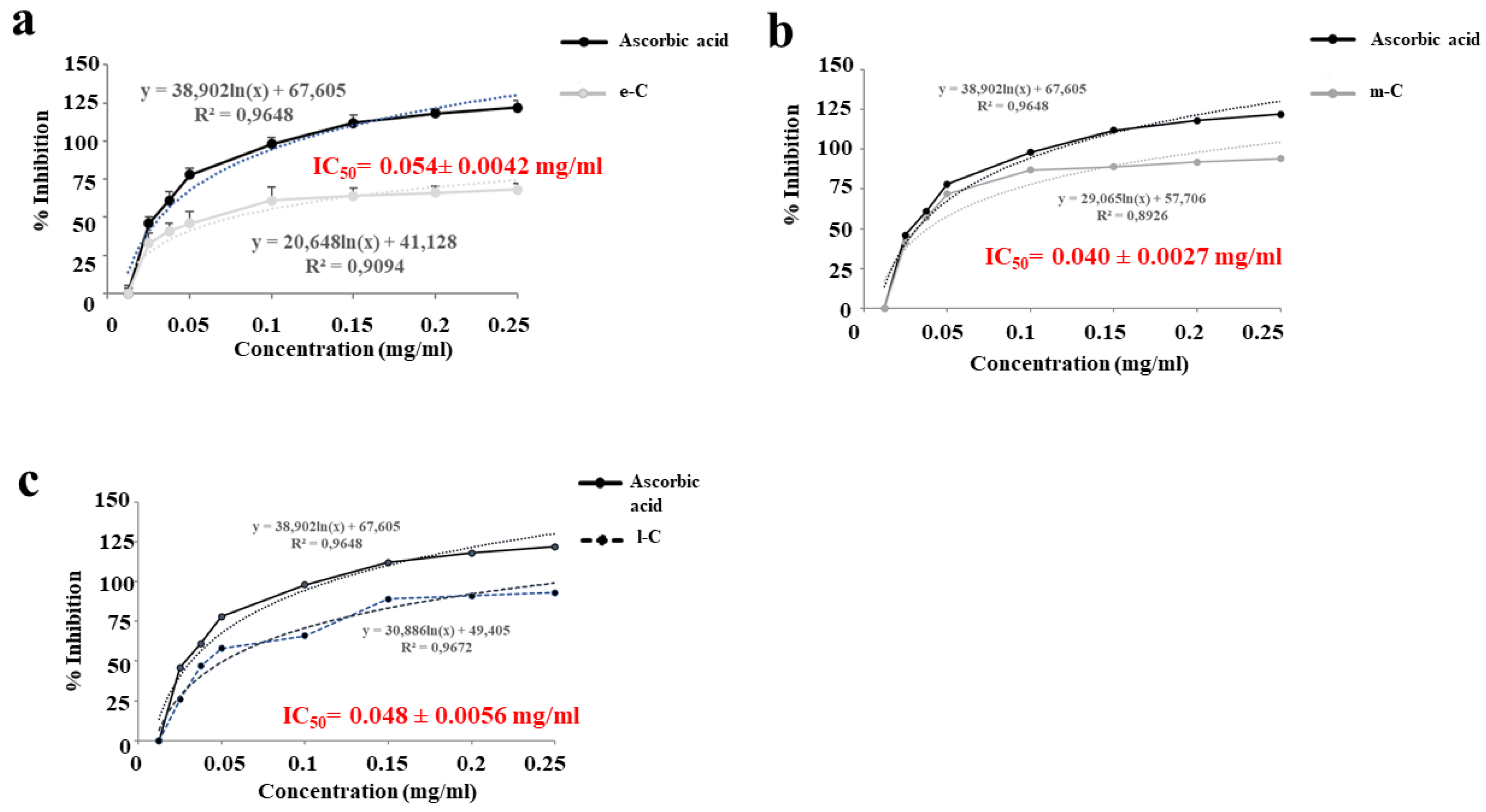
Figure 5.
Effects of e-C, m-C, and l-C on cell viability. Panel a shows the curves of viability generated on Huvec cells by increasing concentrations of extracts. Panel b highlights the protection exerted by extracts on the damage induced by cell exposure to hydrogen peroxide (50 µM, 9 hours). Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. untreated cells; °° denotes p < 0.01 vs. H2O2; § denotes p < 0.05 vs. respective m-C. Variance analysis (ANOVA) was followed by a Tukey–Kramer comparison test.
Figure 5.
Effects of e-C, m-C, and l-C on cell viability. Panel a shows the curves of viability generated on Huvec cells by increasing concentrations of extracts. Panel b highlights the protection exerted by extracts on the damage induced by cell exposure to hydrogen peroxide (50 µM, 9 hours). Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. untreated cells; °° denotes p < 0.01 vs. H2O2; § denotes p < 0.05 vs. respective m-C. Variance analysis (ANOVA) was followed by a Tukey–Kramer comparison test.
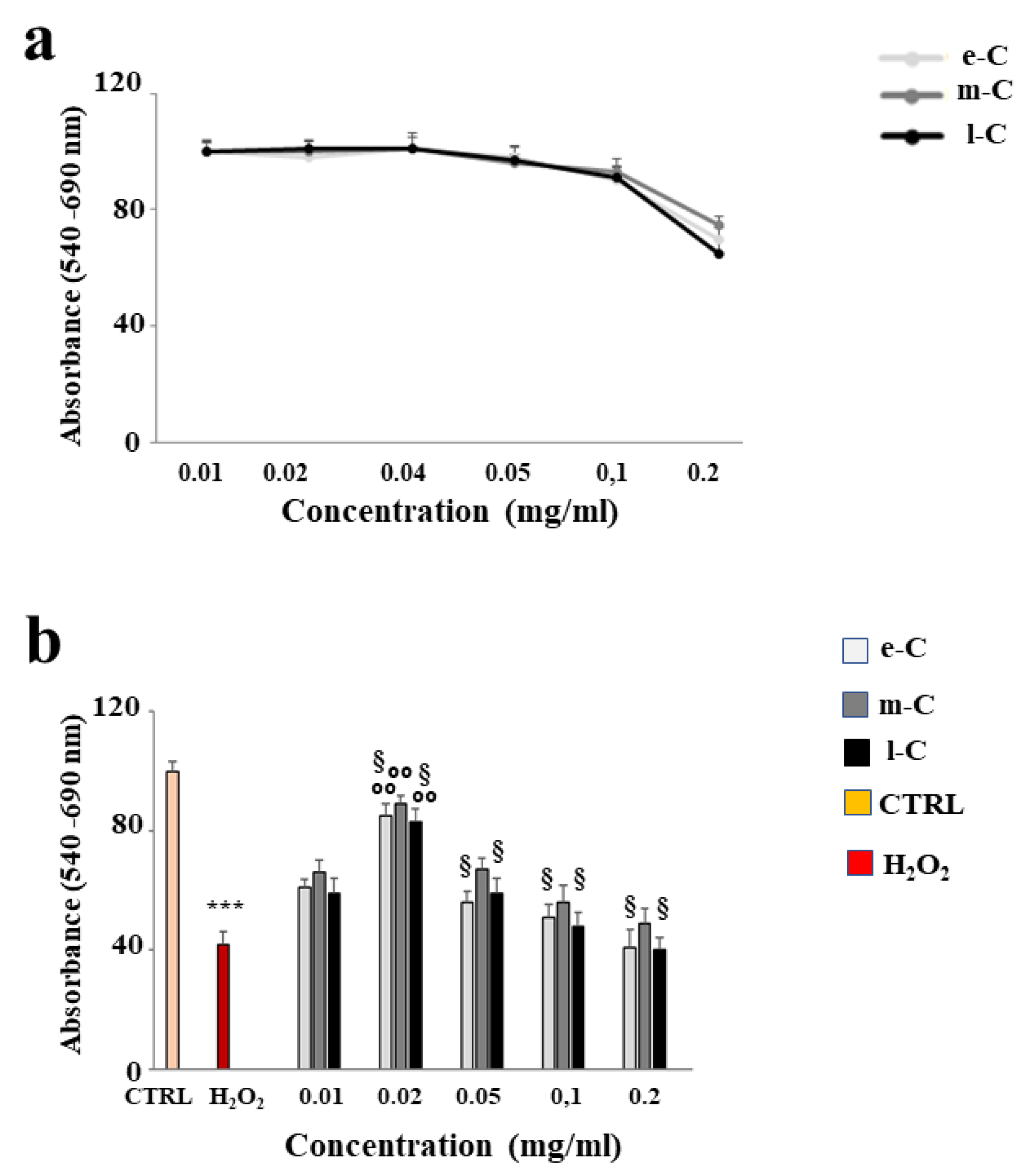
Figure 6.
Measurement of the accumulation of ROS. In panel a, the x-axis indicates the fluorescence of the fluorochrome FITC (connected to the probe that binds the ROS). The shift to the right indicates increased fluorescence and ROS accumulation. The hydrogen peroxide was used as a positive control (150 µM, 20 min). The y-axis is relative to the number of cells that we decided to acquire (20,000). Panel b highlights the quantification of the experiment reported in panel a. Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. the control; ** denotes p < 0.01 vs. the control; * denotes p < 0.05 vs. the control; § denotes p < 0.05 vs. m-C + H2O2. The Analysis of Variance (ANOVA) was followed by the Tukey–Kramer comparison test.
Figure 6.
Measurement of the accumulation of ROS. In panel a, the x-axis indicates the fluorescence of the fluorochrome FITC (connected to the probe that binds the ROS). The shift to the right indicates increased fluorescence and ROS accumulation. The hydrogen peroxide was used as a positive control (150 µM, 20 min). The y-axis is relative to the number of cells that we decided to acquire (20,000). Panel b highlights the quantification of the experiment reported in panel a. Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. the control; ** denotes p < 0.01 vs. the control; * denotes p < 0.05 vs. the control; § denotes p < 0.05 vs. m-C + H2O2. The Analysis of Variance (ANOVA) was followed by the Tukey–Kramer comparison test.
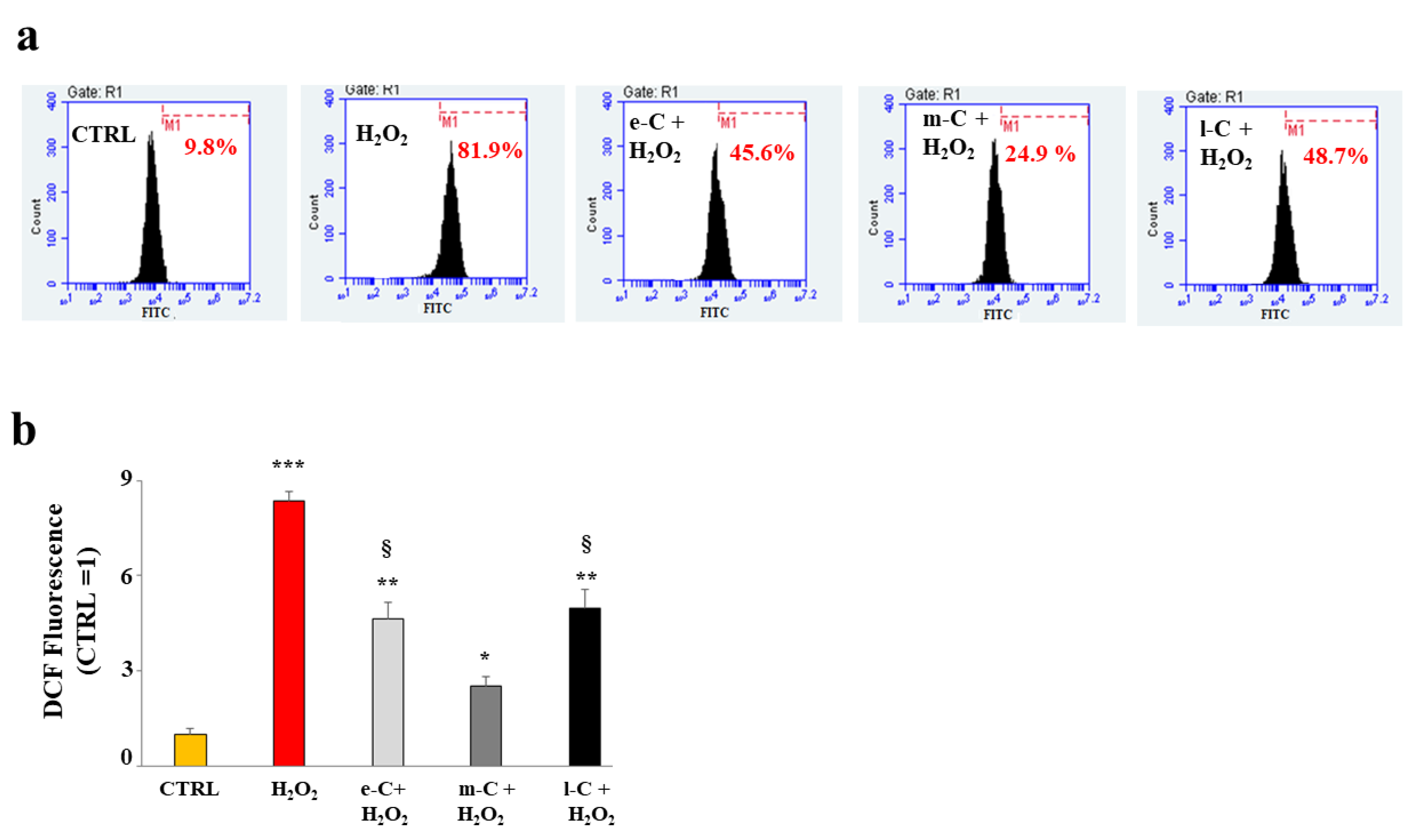
Figure 7.
Effects of extracts of O. ficus-indica, on cellular apoptosis. To evaluate the effect of extracts of O. ficus-indica on the apoptotic process, citofluorimetric experiments with annexin V/PI were conducted. In panel a, a representative experiment of three independent experiments was reported. Each box is divided into four quadrants (Q1, Q2, Q3, and Q4). Q1 refers to Annexin V negative/PI negative cells (viable cells). Q2 refers to Annexin V positive/PI negative cells (early apoptosis). Q3 refers to Annexin V positive/PI-positive cells (late apoptosis); Q4 refers to Annexin V negative/PI-positive cells (necrosis). In panel b, the respective quantification is shown. Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. Q1 of the control; ** denotes p < 0.01 vs. Q1 of the control; * denotes p < 0.05 vs. Q1 of the control; °°° denotes p < 0.001 vs. Q3 of the control; °° denotes p < 0.01 vs. Q3 of the control; ° denotes p < 0.05 vs. Q3 of the control. The Analysis of Variance (ANOVA) was followed by the Tukey–Kramer comparison test.
Figure 7.
Effects of extracts of O. ficus-indica, on cellular apoptosis. To evaluate the effect of extracts of O. ficus-indica on the apoptotic process, citofluorimetric experiments with annexin V/PI were conducted. In panel a, a representative experiment of three independent experiments was reported. Each box is divided into four quadrants (Q1, Q2, Q3, and Q4). Q1 refers to Annexin V negative/PI negative cells (viable cells). Q2 refers to Annexin V positive/PI negative cells (early apoptosis). Q3 refers to Annexin V positive/PI-positive cells (late apoptosis); Q4 refers to Annexin V negative/PI-positive cells (necrosis). In panel b, the respective quantification is shown. Three independent experiments were performed, and the values were expressed as the mean ± sd. *** denotes p < 0.001 vs. Q1 of the control; ** denotes p < 0.01 vs. Q1 of the control; * denotes p < 0.05 vs. Q1 of the control; °°° denotes p < 0.001 vs. Q3 of the control; °° denotes p < 0.01 vs. Q3 of the control; ° denotes p < 0.05 vs. Q3 of the control. The Analysis of Variance (ANOVA) was followed by the Tukey–Kramer comparison test.
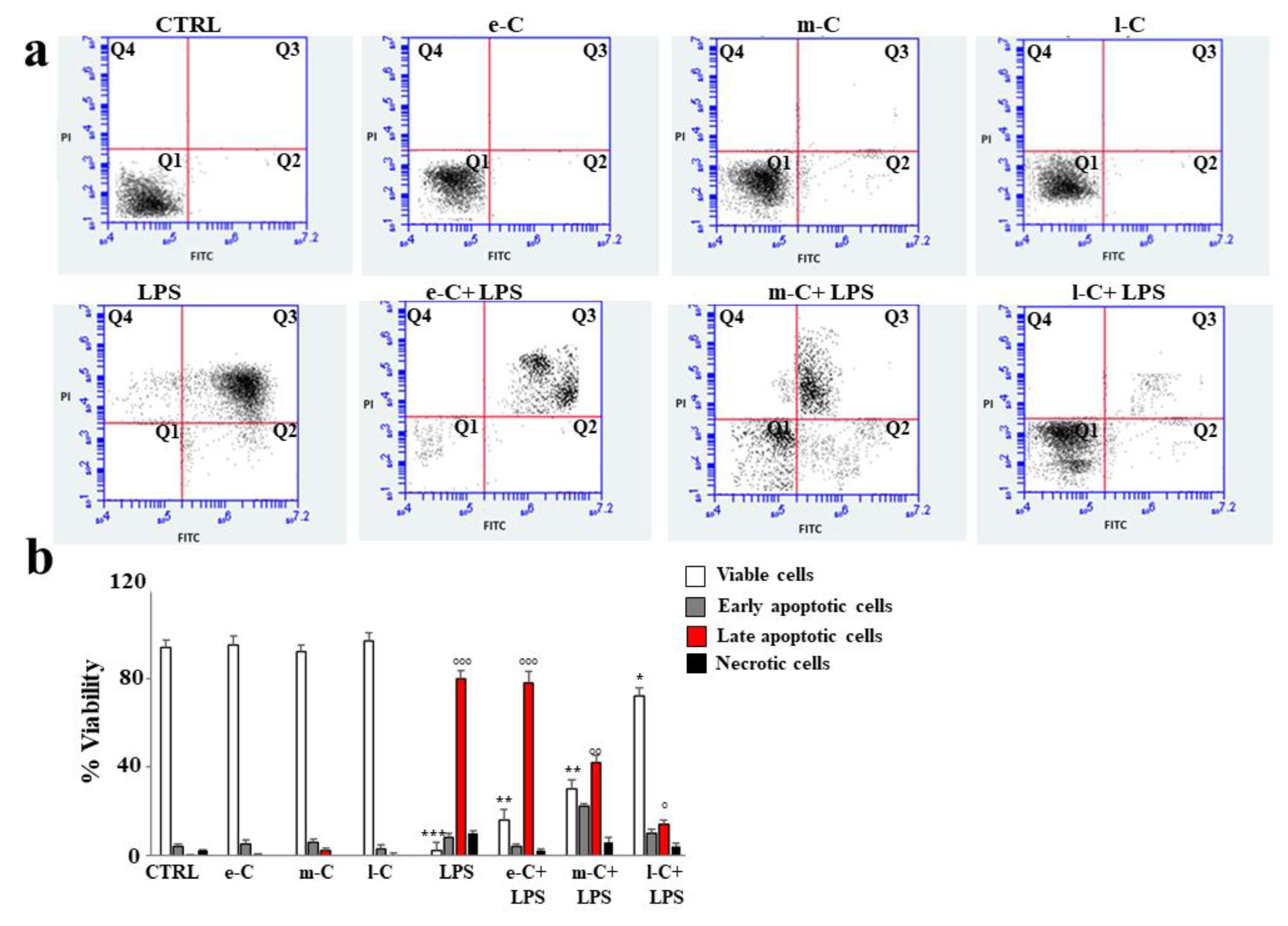
Figure 8.
Effects of extracts of O. ficus-indica, on caspases. Monoclonal antibodies linked to fluorochromes were used to measure the expression of caspase 9 (panel a*) and caspase 3 (panel b). A representative experiment of three independent experiments was reported.
Figure 8.
Effects of extracts of O. ficus-indica, on caspases. Monoclonal antibodies linked to fluorochromes were used to measure the expression of caspase 9 (panel a*) and caspase 3 (panel b). A representative experiment of three independent experiments was reported.
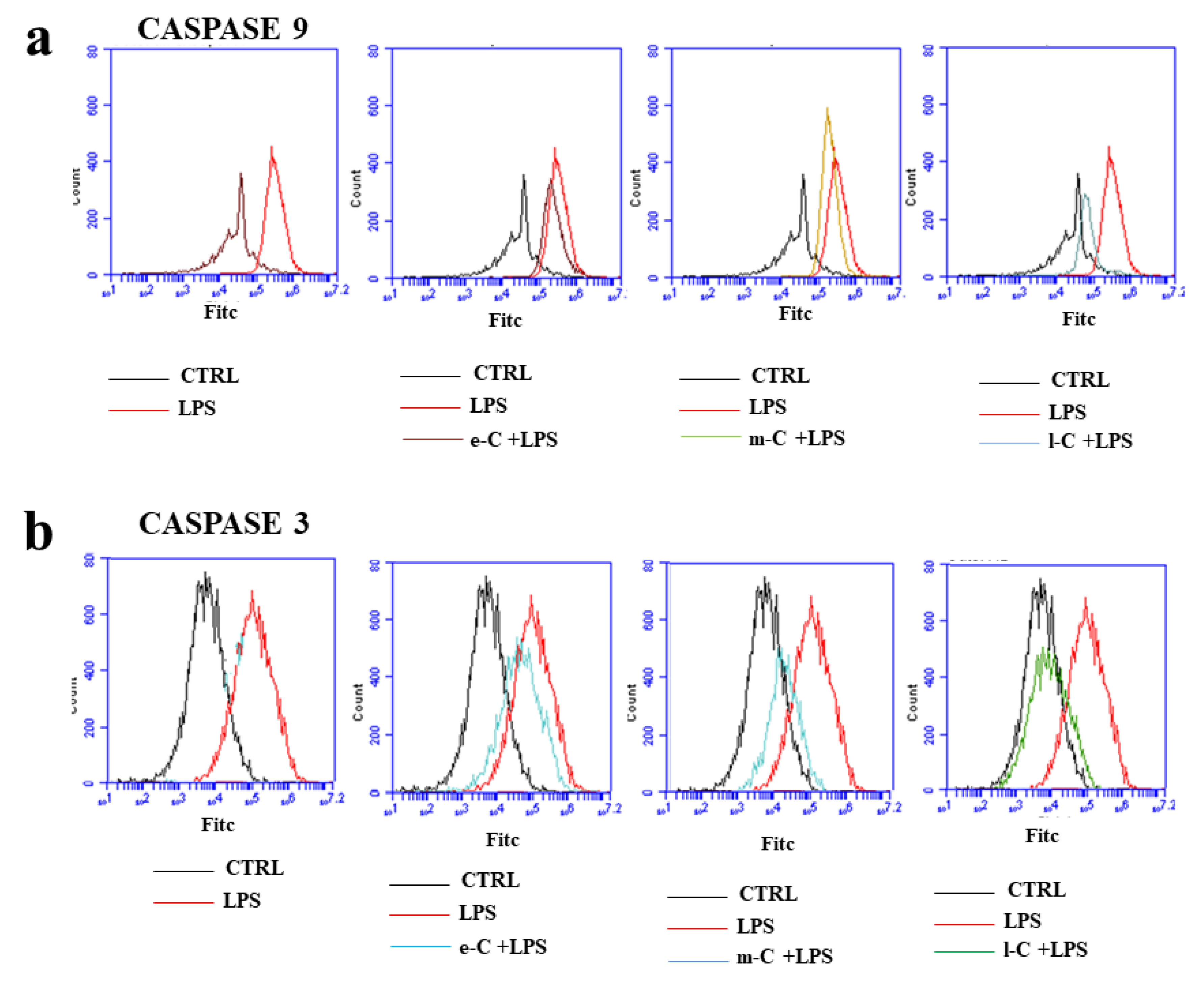
Figure 9.
Total polyphenols and flavonoid content. The polyphenol content of the extracts e-C, m-C, and l-C is shown in panel a. Values are expressed as mg gallic acid equivalents/g extract. In panel b the levels of flavonoids are represented, and the values are expressed as mg quercetin equivalents/g extract. Three independent experiments were performed, and the values were expressed as the mean ± sd.
Figure 9.
Total polyphenols and flavonoid content. The polyphenol content of the extracts e-C, m-C, and l-C is shown in panel a. Values are expressed as mg gallic acid equivalents/g extract. In panel b the levels of flavonoids are represented, and the values are expressed as mg quercetin equivalents/g extract. Three independent experiments were performed, and the values were expressed as the mean ± sd.
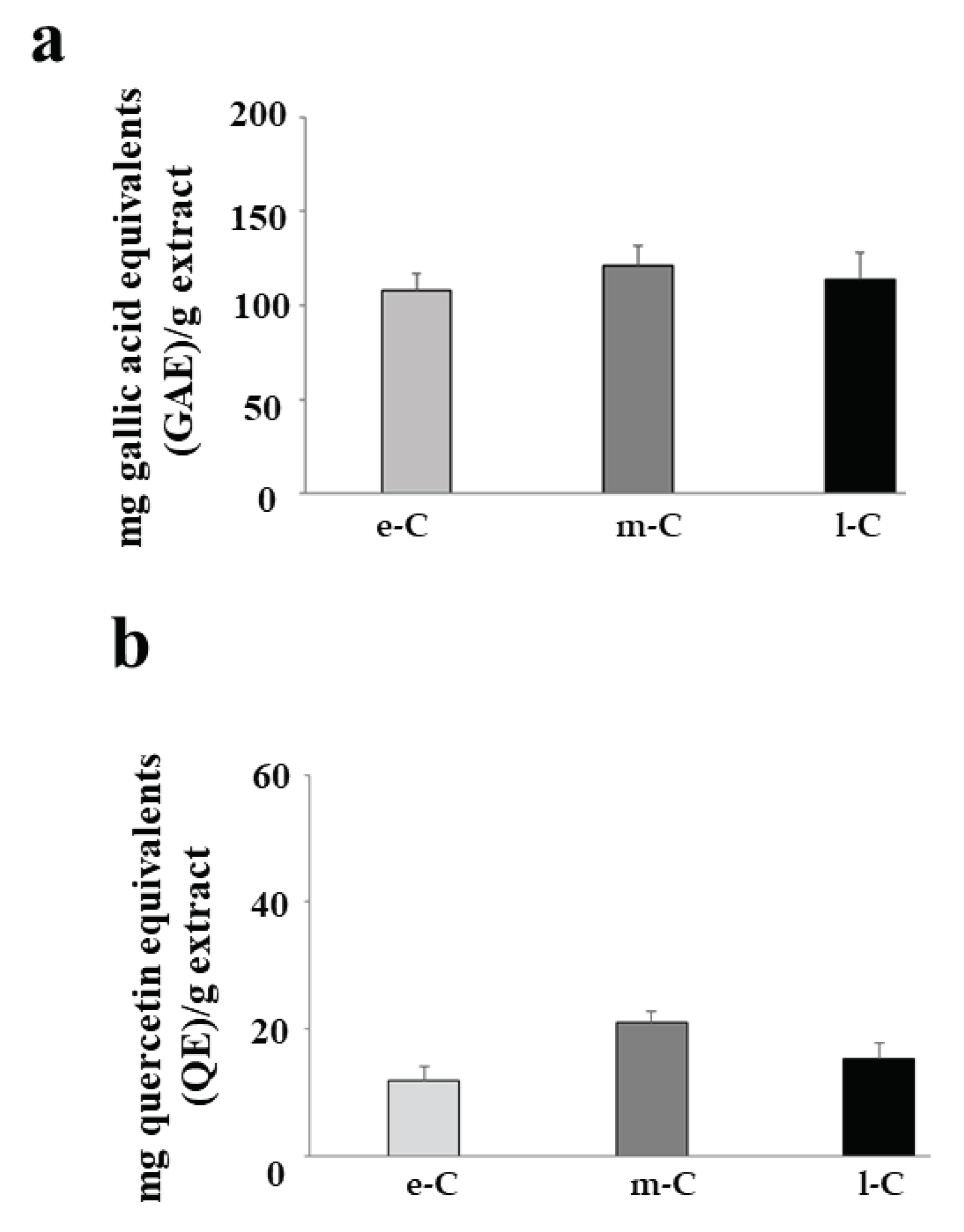

Table 1.
Characteristics of e-C, m-C, and l-C.
| Early cladodes | Medium cladodes | Late cladodes | |
|---|---|---|---|
| Days | 40 | 100 | 150 |
| Grams | 150-220 | 280 | 415 |
| cm | 3-10 cm | 11-25cm | 26-50 cm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



