
Submitted:
05 April 2024
Posted:
08 April 2024
You are already at the latest version
Abstract
The social- and self-directed behaviours of two golden lion tamarins (Leontopithecus rosalia) was investigated while having residence in the tropical house of Aalborg Zoo, Denmark. The behaviour of the female had been distinct from the male since 2018. Despite several diagnostic examinations and medicinal treatments, the cause of the aberrant and apparently stereotypical behavioural patterns remained undetermined, which initiated this study. The pair were recorded throughout a predetermined observation period and these recordings were analysed to illuminate the behaviours of the individuals.
The results showed that there were differences in the type, frequency, and durations of both social- and self-directed behaviours between the sick female and the healthy male. The female expressed both social- and self-directed behaviours more frequently and for longer durations, while the male were more socially reserved and did not engage in social behaviours as frequently. Furthermore, subsequent necropsy of the female revealed uro-genital disease.
Keywords:
Animal Welfare
; Captivity
; Environmental conditions
; Leontopithecus rosalia
; Primates
; Stereotypical behaviour
; Surveillance
; Time budget
; Zoo
Introduction
The golden lion tamarin (GLT), Leontopithecus rosalia, is a small, social and adaptable frugi-faunivore primate of the Callitrichidae family (Kierulff, et al., 2012) (Stolwijk, 2013). They are endemic to the lowland Atlantic Forest region of Rio de Janeiro, Brazil (Kierulff, et al., 2012) (Stolwijk, 2013) (Lapenta, et al., 2007). The golden lion tamarin has a long, silky, and amber pelage as well as a dark bare face surrounded by the mane. The species have elongated arms and fingers, which are used to forage for fruits, small prey, and plant exudes (gums) (Kierulff, et al., 2012) (Snyder, 1974). Wild animals live in matriarchal social groups (called troops) consisting of about four to eight individuals (Stolwijk, 2013) (Lapenta, et al., 2007), typically composed of a breeding pair and their offspring (Lapenta, et al., 2007). The social structure of the species has been shaped by the fact that 78 % of dominant females pregnancies result in twin births, which has led to cooperative breeding in the species (Snyder, 1974) (Stolwijk, 2013) (Lapenta, et al., 2007).
In 1982 GLT was added to IUCN Red List of Threatened Species (Ruiz-Miranda, et al., 2019) due to declining habitat availability and quality (Stolwijk, 2013). They were reclassified as critically endangered from 1996-2000 (Ruiz-Miranda, et al., 2019) (Stolwijk, 2013) (Dietz, 2000). Although the species is now in recovery, there are still complications regarding breeding issues in some captive environments (Snyder, 1974) (Stolwijk, 2013) (Burell & Altman, 2006).
Golden lion tamarins raised in captivity are exposed to other environments and stimuli than their wild-born counterparts (Ruvio & Stevenson, 2017) (Hosey, 2005) (Lutz, et al., 2022). Therefore, they could express different behaviors, new behaviours or possibly lose existing behavioural patterns essential for their social interactions within the troop in addition to their survival in the wild (Hosey, 2005) (Ruvio & Stevenson, 2017). Furthermore, this may result in social instability within a captive-reared troop (Anzenberger & Falk, 2012). These behaviours can include the deficient acquisition of essential survival skills (Sanders & Fernandez, 2020), or detrimental and unnatural stereotypical behaviour, which is often observed in captive animals (Hosey, 2005) (Gottlieb, et al., 2013). Stereotypical behaviours are atypical and repetitive behaviours with no apparent purpose, such as auto-aggression and autoerotism (Gottlieb, et al., 2013) (Lutz, et al., 2022) (Linder, et al., 2020). These behavioural patterns usually indicate that an animals’ psychological welfare is sub-optimal (Lutz, et al., 2022) (Mason & Rushen, 2006) (Linder, et al., 2020). Stereotypical behaviour is often accentuated by insufficient stimuli in the enclosure or may occur as a stress response to the environment or other individuals in the social hierarchy (Lutz, et al., 2022) (Redshaw & Mallinson, 1991) (Carlstead & Shepherdson, 2000). The abnormal stereotypical behaviour may not necessarily be in direct relation to the current welfare state of the animal but could also be a trait from previous triggers (Lutz, et al., 2022) that has become a habitual or automatic response that persists (Lutz, et al., 2022). Auto-aggression and other self-directed behaviours might likewise develop because of restricted physical contact during the first months of life for Callitrichidae (Mallapur & Choudhury, 2003).
Another factor which might induce a stress response in captive GLT in a zoo enclosure, is the presence of larger family troops of other contending Callitrichidae species in proximity (Kierulff, et al., 2012). Studies show that Callitrichidae species should not be housed adjacent to each other, even if they cannot physically interact (Kierulff, et al., 2012) (Snyder, 1974). If there is visual and/or olfactory contact, it could result in severe querulousness (Snyder, 1974). Additionally, GLT will scent mark their territory more frequently when in proximity of contending Callitrichidae (Snyder, 1974). Scent marking activity might increase after sexual encounters, alterations in territory and aggressive intra- and interspecies interactions (Snyder, 1974). The impacts of the zoo environment on the behaviour of captive primates such as GLT have been researched, but concrete conclusions have not been found (Hosey, 2005).
Zoos generally run a higher risk of having an environment that is not optimally suited for the animal (Hosey, 2005) (Sanders & Fernandez, 2020), although modern zoos try to negate this issue to assure the welfare of the animals (Hosey, 2005). Zoos might provide climate control in the enclosures, variation in stimuli and attempt to promote natural behaviours seen in wild GLT, by for instance promoting troop-compositions which could exist in a natural environment (Hosey, 2005) (Snyder, 1974) (Stolwijk, 2013), playing (Snyder, 1974) (Lutz, et al., 2022), self- and allogrooming (Snyder, 1974), resting during the day and bedding down together in dens at lights out (Snyder, 1974), via enrichment programs (Linder, et al., 2020).
While much is known of the social structure and -behaviours of captive Callitrichidae in general, less is known of the influence of an incomplete troop on individual stereotypical behaviour and social interaction. Even less is known of the influence of a sick individual in a captive, small troop.
The aim of this study was to investigate whether the frequency and duration of social- and self-directed behaviours in a captive pair of golden lion tamarins are influenced by a single sick individual. It was anticipated that allogrooming and other positive social interactions would be observed frequently. In lieu of this investigation, potential stereotypical behavioural patterns was examined. This study highlighted the importance of animal welfare in captivity, social- and self-directed behaviours and interactions in social groups.
Methods and Materials
Participants and Setting
This behavioural study included one female (GLT36) and one male (GLT37) golden lion tamarin in Aalborg Zoo (Aalborg, Denmark). The female (born 2011) originated from Olomouc Zoo in the Czech Republic, while the male (born 2012) originated from the Lisbon Zoo in Portugal. The female arrived to Aalborg Zoo in 2014, and she was introduced to the male in July 2019, when he arrived to the zoo as a replacement for another male. Upon reception they were quarantined together for a month before being moved into the enclosure. The female had exhibited signs of discontent and disease after experiencing a potential miscarriage in 2020; she was incontinent and had alopecia on the tail and lower body. After their introduction, it was observed that the male exhibited antagonistic interactions with the female, and since 2020 no mating efforts have been noted between them. Likewise, it was noted that the female exhibited stereotypical behaviour, as she would gnaw on a specific branch several times a day for different durations.
Animals in the Tropical House
The two individuals were kept in a medium sized callitrichid enclosure (approximately 5 meters broad by 2.5-3 meters deep and 3.5-4 meters high) with access to an outside area, which mimics the natural habitat of the species. The tropical house Goeldi’s marmoset (Callimico goeldii) and Emperor tamarin (Saguinus imperator), both of which have larger family troops, vocalise excessively, and scent mark their enclosures. The Emperor tamarin enclosure was placed right next to the GLT enclosure, while the Goeldi’s marmoset enclosure was a bit further away, but still in olfactory and vocal vicinity. Furthermore, the tropical house also holds fish, reptiles, amphibians, and insects.
Data Collection
Observations were conducted by filming the GLT enclosure inside the Tropical house with six Kitvision Venture 4K action cameras. The cameras filmed continuously on October 20th, 23th, 24th and 27th and again on November 1st, 3rd and 8th – all days from 8:45 AM to 2:15 PM.
An ethogram (Table 1) was developed and used to analyse the behaviours observed (Altmann, 1974). As our study focuses on the social- and self-directed behaviour of both individuals, thus behaviours deemed irrelevant to this have been categorized together as “others”.
Data Analysis
To do statistical analysis of the data, seven days were chosen at random from the observation period. As all data were not normally distributed, non-parametric statistical analysis was applied. Statistical analyses were performed in Excel, Past version 4.03 and RStudio. Histograms was used to illustrate the frequency distribution for each behaviour and frequency distribution for interim between potentially related stereotypical behaviours. Cumulative graphs were used to illustrate how much time each of the two individual cumulatively spent on specific behaviours throughout the days for the entire observation period.
Results
Distribution of Behaviours
Frequency of Duration for the Expressed Behaviours
The frequencies of behaviour durations of observed behaviours are presented on Figure 2-5.
Figure 2A shows that the female was allogrooming more frequently and at longer durations compared to the male. Most instances of allogrooming for the female lasted approximately seven seconds or less, while the male would be allogrooming infrequently with only one occurrence of allogrooming lasting more than 10 seconds. Figure 2B, the frequency for the majority of instances of negative social interactions lasted six seconds or less, with both having one occurrence lasting 20 seconds or longer.
Figure 3A shows that the male exhibited positive social interaction most frequently for approximately 10 seconds or less. The female exhibited positive social interactions 24 times and the duration was usually less than 10 seconds. As illustrated in Figure 3B, the female was scent marking more frequently than the male, with more than 300 instances lasting for less than 10 seconds. The female also had occurrences of scent marking with a duration of 40 seconds or longer.
Figure 4A illustrates that the female had 17 instances of self grooming lasting at least 50 seconds. She also had over 500 occurrences of self grooming lasting 10 seconds or less. The male had more than 200 occurrences of self grooming last 10 seconds or less. Figure 4B shows that he had two instances of interacting with the specific branches used for this behaviour. The female had more than 400 instances, of which most are at 20 seconds or less and 10 instances had a duration of 30 seconds or more.
Figure 5 shows that for more than 200 instances, approximately 10 seconds or less would pass from the female interacting with a branch to initiation of self grooming.
Cumulative Amount of Time Spent on Behaviours
Figure 6a illustrates that the amount of time spent on allogrooming increases from 11:30 to 14:15, from approximately 100 seconds to over 800 seconds for the female, while the male ends at approximately 50 seconds total spent. On Figure 6b it is illustrated that the male had a total cumulative time spent on negative social interaction of approximately 60 seconds, which is nearly double the approximately 35 seconds in total for the female. Both had a period from 10:15 to 12:30 with very little negative social interaction surrounded by singular occurrence of long periods of negative interaction from either of the two.
Figure 7A shows that the female had a total cumulative time of less than 200 seconds and the males’ total cumulative time was more 1000 seconds. At around 11:30, the male had less than 200 seconds cumulative of positive social interactions, after which a steep increase was observed. Figure 7b illustrates the females’ total cumulative time spent on scent marking was approximately 4.000 seconds, while it was only 450 seconds for the male.
Figure 8a illustrates that the females’ total cumulative time spent on self grooming was approximately 16.000 seconds, while only approximately 1.700 seconds for the male. Figure 7b illustrates that the female spent a total cumulative time of approximately 4300 seconds interacting with a branch, while the male spent approximately 26 seconds. The female regularly spends time on this behaviour, while the male was only observed expressing this behaviour during a specific timeframe, and only for a comparably short amount of time.
Discussion
Expression of Individual Behaviours
Different responses by individuals to uniform and familiar stimuli have been conservatively studied in Callitrichidae and primates in general. A significant difference in the time spent on different behaviours between individuals can influence the ability to predict how captivity affect GLT in general, because the response of each individual in the population may vary depending on the individuals’ ability to cope (Morgan & Tromborg, 2007) (Hosey, 2005) (Davey, 2007).
The Time Distribution of Behaviours Exhibited Vary
It is indicated (Figure 1a and 1b) that the proportion of time allocated to socially important behaviours which are time-consuming in wild GLT, such as allogrooming (Morgan & Tromborg, 2007) (Snyder, 1974), are quite minimal for both the female and the male, although the female initiated allogrooming more frequently (Figure 2a). This contradicts other studies, which show that male allogroom female GLT more frequently, compared to female GLT grooming male GLT (Kleiman, 1981). When the female would initiate allogrooming, it would at times lead to the male immediately moving away from her. His hesitance to engage in allogrooming might stem from different factors. On the one hand, according to the EAZA Guidelines for Callitrichidae (Ruvio & Stevenson, 2017), social animals housed together in captivity can tend to interact too much (Ruvio & Stevenson, 2017), which would lead to interactive issues between the individuals of the troop (Ruvio & Stevenson, 2017). On the other hand, a study conducted on cotton-top tamarins (Saguinus oedipus), found that the environmental conditions of captivity would reduce the overall display of positive social interactions, such as allogrooming (Morgan & Tromborg, 2007) and play (which was not at all noted during the observations). However, yet another study found a correlation between primates being reared in isolation or near-isolation and a diminished ability to engage in social interactions and other more "normal" behaviours (Gottlieb, et al., 2013). Considering that the males’ history before being received by Aalborg Zoo is unknown, this could also play a role in their relationship, however it is unlikely. In general, the female spent significantly more time allogrooming than the male (Figure 2A and 6A). She would often try to initiate allogrooming, mostly when the pair is on a specific branch located right outside their nestbox.
For the female and the male, there was differences in the type, frequency, and duration of both social- and self-directed behaviours, which indicate that they express different behavioural patterns to the same environmental stimuli (i.e. captivity and sickness within the pair). This includes self-directed behaviours such as self grooming and scent marking. Considering the disease background of the female it might be an expression of displacement activities (i.e autoerotism, self grooming) caused by stress (Troisi, 2002) (Carlstead & Shepherdson, 2000) (Gottlieb, et al., 2013) (Lutz, et al., 2022). Displacement activities are common when motivational conflicts appear (Maestripieri, 1992), and can often be expressed by displaying more than one activity at once (Troisi, 2002), as the female does during interaction with a branch and self grooming according to observations (Figure 5). The motivational conflict can stem from both the conspecific of the family troop (in this case, either the male, who has expressed antagonistic behaviours towards the female) or the presence of contending Callitrichidae family troops that are nearby (Snyder, 1974).
Likewise, the pathogenicity of the female could affect both her social- and self-directed behaviours, as well as the social behaviour of the male towards her (Freeland, 1976). Sick individuals will often adapt their behaviour to avoid infecting the troop with disease, and troops will be expected to have disease-controlling behaviours, in which they avoid contact with the sick individual (Freeland, 1976). However, the female did not suffer any infectious disease, neither ectoparasites or fungal infections. She did however have an ovarian cyst, which could have affected the endocrine functions, which is essential for communication in some Callitrichidae (Candland, et al., 1980). To our knowledge, no research has so far examined the impact of a sick individual in a pair, or anything related to such situations.
Most of the behaviours expressed by the male fall into the ‘others’ category, and he did not express the social- or self-directed behaviours observed in another male Callitrichidae as frequently as expected (Kleiman, 1981).
Gauging an animal’s internal state through the observation of its external behaviours can however be tricky (Maestripieri, 1992). In a social species, the absence and/or presence of conspecifics in the enclosure can confirm or disrupt expectations, interrupt, or initiate a behavioural sequence or create motivational conflicts (Maestripieri, 1992). Overall, the type, frequency, and duration of self-directed- and social behaviours between both the female and the male varies.
The Repetitive Self-Directed- and Social Behaviours of the Individuals
According to an earlier study, some marmosets are known to display repetitive or purposeless (Linder, et al., 2020) behaviour, which can be expressed in response to sub-optimal physical and/or mental state. Stereotypical behaviours might for instance be carried out to reduce the psychological stress experienced by the marmoset (Kaplan & Rogers, 2012). It was observed that the female was prone to perform scent marking more frequently and for overall longer than the male (Figure 3B and 7B). This might be influenced by the instincts of the female; in wild social hierarchies, female GLT act as the dominant matriarch, and they can often be seen displaying dominance by scent marking their territory (Snyder, 1974). Likewise, male Callitrichidae are known to influence the scent marking activities of female conspecifics, as the males will urine-wash throughout the enclosure during breeding season (Candland, et al., 1980).
Furthermore, GLT have been observed to engage in aggressive interactions with Goeldi’s marmosets when separated in adjacent enclosures (Snyder, 1974). This observation, collected by Snyder (Snyder, 1974), could insinuate that the Goeldi’s marmosets adjacent to the GLT may lead the female to perform scent marking more frequently, in order to display dominance and mark her territory (Roberts, 2012). Furthermore, the excessive scent marking performed by the female could possibly be the cause of (or at least worsen the condition of) the dermatitis, which was noted on her lower abdomen, vaginal area, and thighs. As the female scent marks, she drags her abdomen, perineum and thighs across wooden surfaces, which could arguably cause pain due to the dermatitis; despite this, she continues to do so. This might be an attempt to reduce psychological stress in order to cope with an infringed territory or due to frustration, as described by Gottlieb and coworkers (Gottlieb, et al., 2013), induced by the presence of competing species in the Zoo (Snyder, 1974) (Hosey, 2005) (Gottlieb, et al., 2013). The study also mentions how abnormal behaviours, such as auto-aggression, could be detected, which correlates with the female continuing to scent mark despite bleeding from the lower abdomen and vaginal area. Auto-aggression does not occur in captivity very frequently (Mason & Rushen, 2006), however. Furthermore, it has been observed that GLT performs self grooming frequently, possibly to obtain social advantages similar to those of allogrooming (Snyder, 1974). Additionally, the female can be seen self grooming more frequently than the male (Figure 2A). The female has also been observed to ascertain a certain stereotypical behavioural pattern, where the individual will observe and interact with some specific different branches in the enclosure, some more than others, especially one located near the nestbox and then start to self groom shortly after (Figure 5). This pattern could be caused by the males’ lack of interest in allogrooming, causing her to perhaps find comfort in performing allogrooming-like interactions with branches and then almost directly after starting to self groom (Figure 2A and 6A).
Captive family troops of marmosets, whose composition would not be seen naturally occurring, might be more susceptible to display abnormal behaviours (Linder, et al., 2020) (Morgan & Tromborg, 2007), and this might be factor in the unusual (Snyder, 1974) proportion of time allocated to doing abnormal and stereotyped behaviours (Lutz, et al., 2022) in both individuals.
Limitations
This study examines only one pair of GLT for a limited time period, which limits the reliability of the obtained data. Furthermore, several factors could have influenced the results of this study, as there are many variables which influence the behaviours of primates. Furthermore, the design does not allow distinguishing between gender differences and differences between sick and healthy individuals.
Conclusion
The female and the male golden lion tamarins showed differences in type, frequency, and duration of both social- and self-directed behaviours. The female expressed both social- and self-directed behaviours more frequently and for longer durations, while the male were more socially reserved and did not engage in social behaviours as frequently. Stereotypical self-directed behaviours of the female could be caused by frustration or motivational conflicts, and the reservedness of the male could be influenced by the female´s health problems. Due to a small sample and a limited amount of knowledge about sick individuals reactions in social groupings, these conclusions may be precautious.
Acknowledgements
Thanks to Aalborg Zoo and especially the animal keepers, Nicolai Bach Thomas and Dennis G. Lynggren for taking care of the animals.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Altmann, J., 1974. Observational Study of Behaviour: Sampling Methods. Behaviour.
- Anzenberger, G. & Falk, B., 2012. Monogamy and family life in callitrichid monkeys: deviations, dynamics and captive management. International Zoo Yearbook. [CrossRef]
- Bryan, K., Harrison, S. B., Price, E. & Wormell, D., 2017. The Impact of Exhibit Type on Behaviour of Caged and Free-ranging Tamarins. Applied Animal Behaviour Science. [CrossRef]
- Bufalo, F. S., Galetti, M. & Culoy, L., 2016. Seed Dispersal by Primates and Implications for the Conservation of a Biodiversity Hotspot, the Atlantic Forest of South America. International Journal of Primatology. [CrossRef]
- Burell, A. M. & Altman, J. D., 2006. The Effect of the Captive Environment on Activity of Captive Cotton-top Tamarins (Saquinus oedipus). Applied Animal Welfare Science.
- Candland, D. K., Blumer, E. S. & Mumford, M. D., 1980. Urine as a communicator in a New World primate, Saimiri sciureus. Animal Learning and Behaviour. [CrossRef]
- Carlstead, K. & Shepherdson, D. J., 2000. The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare - Alleviating stress in zoo animals with environmental enrichment. s.l.:CABI Publishing.
- Davey, G., 2007. Visitor's Effects on the Welfare of Animals in the Zoo. Applied Animal Welfare Science.
- Dietz, J. M., 2000. Endangered Animals. s.l.:Green Wood Press.
- Franklin, S. P., 2004. Predator Influence on Golden Lion Tamarin Nesy Choice and Presleep Behaviour. s.l.:Departmnet of Biology.
- Freeland, W. J., 1976. Pathogens and the Evolution of Primate Sociality. Biotropica. [CrossRef]
- Gottlieb, D. H., Capitanio, J. P. & McCowan, B., 2013. Risk factors for stereotypic behavior and self-biting in rhesus macaques (Macaca mulatta): Animal's history, current environment, and personality. American Journal of Primatology.
- Hosey, G. R., 2005. How does the zoo environment affect the behaviour of captive primates?. Applied Animal Behaviour Science. [CrossRef]
- Kaplan, G. & Rogers, L. M., 2012. Stress and stress reduction in common marmosets. Applied Animal Behaviour Science. [CrossRef]
- Kierulff, M. C. M. et al., 2012. The Golden lion tamarin Leontopithecus rosalia: a conservation success story. Zoological Society of London, 28 February, pp. 36-45.
- Kleiman, D. G., 1981. Mammalian species - Leontopithecus rosalia. American Society of Mammalogists.
- Lapenta, M. J., Procópio de Pliveira, P. & Nogueira-Neto, P., 2007. Daily activity period, home range and sleeping sites of golden lion tamarins (Leontopithecus rosalia) translocated to the União Biological Reserve, RJ-Brazil. De Gruyter, 4 October.
- Linder, A. C. et al., 2020. Using Behavioral Instability to Investigate Behavioral Reaction Norms in Captive Animals: Theoretical Implications and Future Perspectives. Symmetry. [CrossRef]
- Lutz, C. K. et al., 2022. Nonhuman primate abnormal behavior: Etiology, assessment, and treatment. American Journal of Primatology. [CrossRef]
- Maestripieri, D., 1992. A modest proposal: displacement activities as an indicator of emotions in primates. Animal Behaviour. [CrossRef]
- Mallapur, A. & Choudhury, B. C., 2003. Behavioural Abnormalities in Nonhuman Primates. Journal of Applied Animal Welfare Science.
- Mallinson, J. J. C., 1995. Conservation breeding programmes: an important ingredient for species survival. Biodiversity and Conservation. [CrossRef]
- Mason, G. & Rushen, J., 2006. Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare. s.l.:s.n.
- Morgan, K. N. & Tromborg, C. T., 2007. Sources of Stress in Captivity. Applied Animal Behaviour Science.
- Pertoldi, C., Bahrndorff, S., Novicic, Z. K. & Rohde, P. D., 2016. The Novel Concept of Behavioural Instability and its Potential Applications. Symmetry. [CrossRef]
- Redshaw, M. E. & Mallinson, J. J. C., 1991. Stimulation of natural patterns of behaviour: studies with golden lion tamarins and gorillas. s.l.:s.n.
- Roberts, S. C., 2012. On the Relationship between Scent-Marking and Territoriality in Callitrichid Primates. Int J Primatol. [CrossRef]
- Ruiz-Miranda, C. R., Kierulff, C. & Jerusalinsky, L., 2019. Leontopithecus rosalia, Golden Lion Tamarin, s.l.: International Union for Conservation of Nature and Natural Resources.
- Ruvio, E. B. & Stevenson, M. F., 2017. EAZA Best Practice Guidelines for Callitrichidae, s.l.: EAZA.
- Sanders, K. & Fernandez, E. J., 2020. Behavioral Implications of Enrichment for Golden Lion Tamarins: A Tool for Ex Situ Conservation. Journal of Applied Animal Welfare Science. [CrossRef]
- Snyder, P. A., 1974. Behavior of Leontopithecus rosalia (Golden-lion marmoset) and related species: A review. Journal of Human Evolution, March, 3(2). [CrossRef]
- Stolwijk, R., 2013. The golden lion tamarin (Leontopithecus rosalia): a flagship species for the Atlantic Forest of Brazil. Universiteit Utrecht.
- Troisi, A., 2002. Displacement Activities as a Behavioral Measure of Stress in Nonhuman Primates and Human Subjects. [CrossRef]
Figure 1.
Time budgets for the two golden lion tamarins the female (A) and the male (B) spent on each behaviour in percentage of the time. Behaviours designated “other” comprises of all behaviours not relevant to this study.
Figure 1.
Time budgets for the two golden lion tamarins the female (A) and the male (B) spent on each behaviour in percentage of the time. Behaviours designated “other” comprises of all behaviours not relevant to this study.
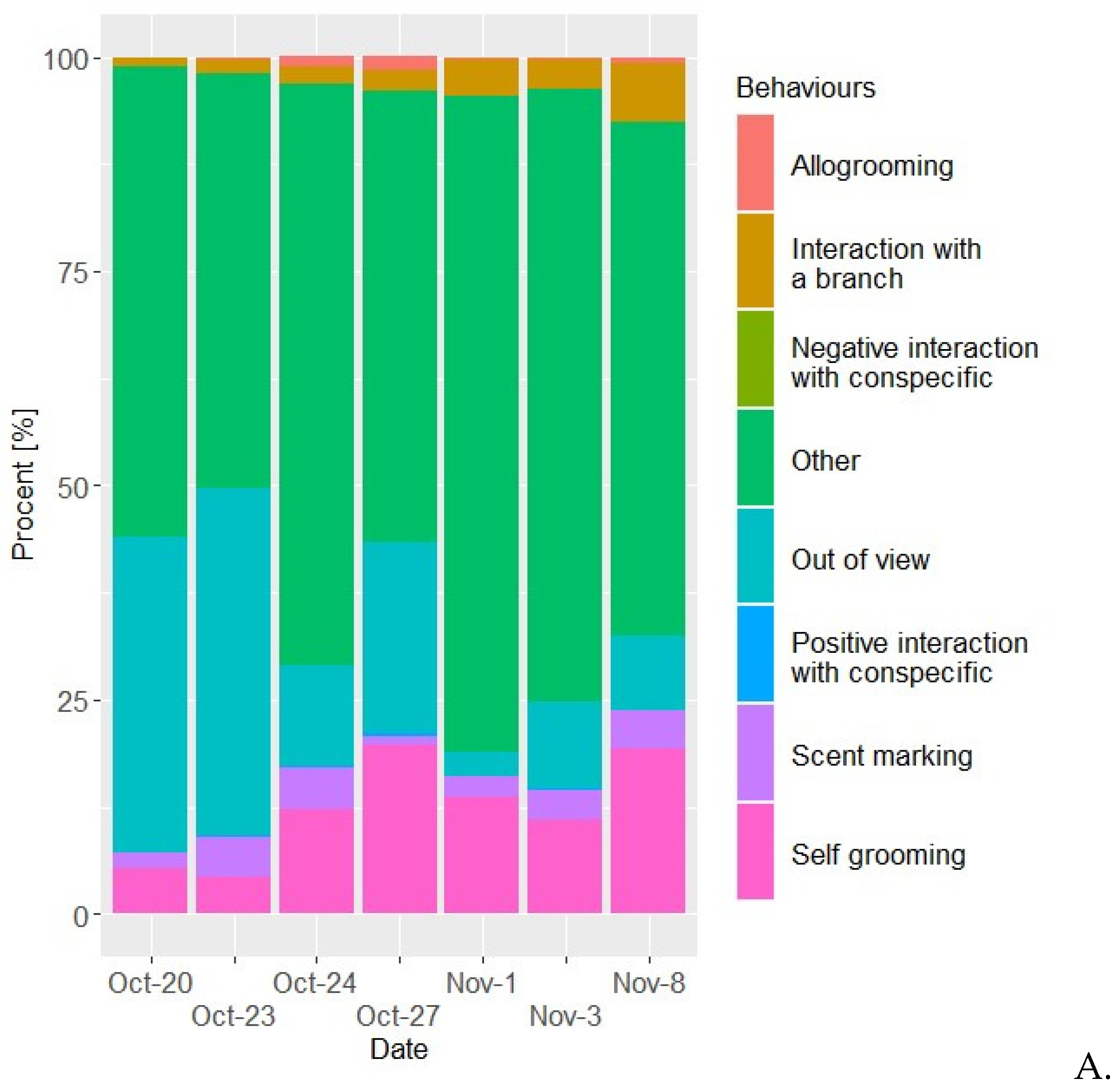
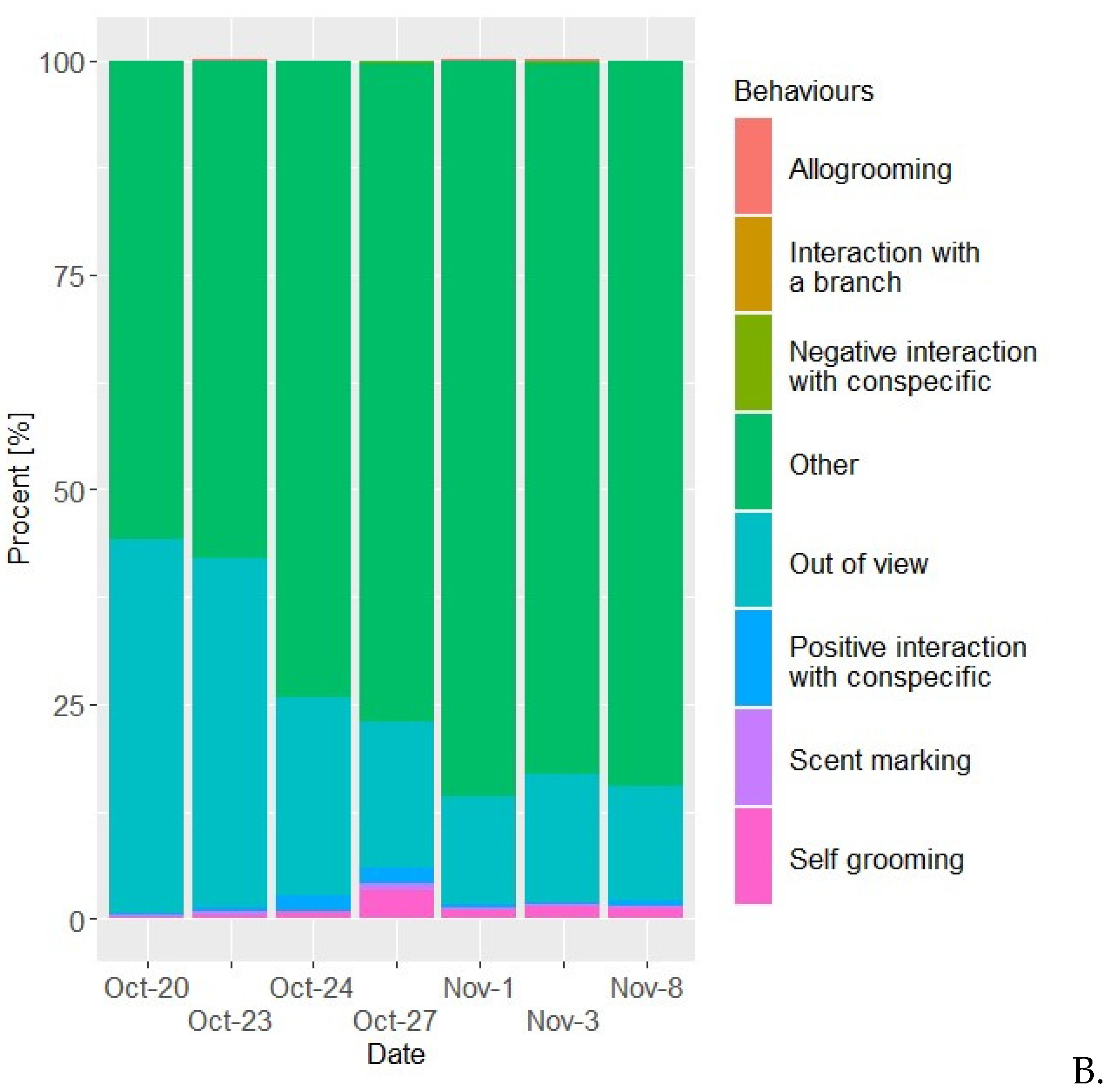
Figure 2.
Frequency histogram illustrating the frequency for duration of the behaviours A. Allogrooming and B. negative interaction with conspecific for the golden lion tamarin individuals, the female and the male.
Figure 2.
Frequency histogram illustrating the frequency for duration of the behaviours A. Allogrooming and B. negative interaction with conspecific for the golden lion tamarin individuals, the female and the male.
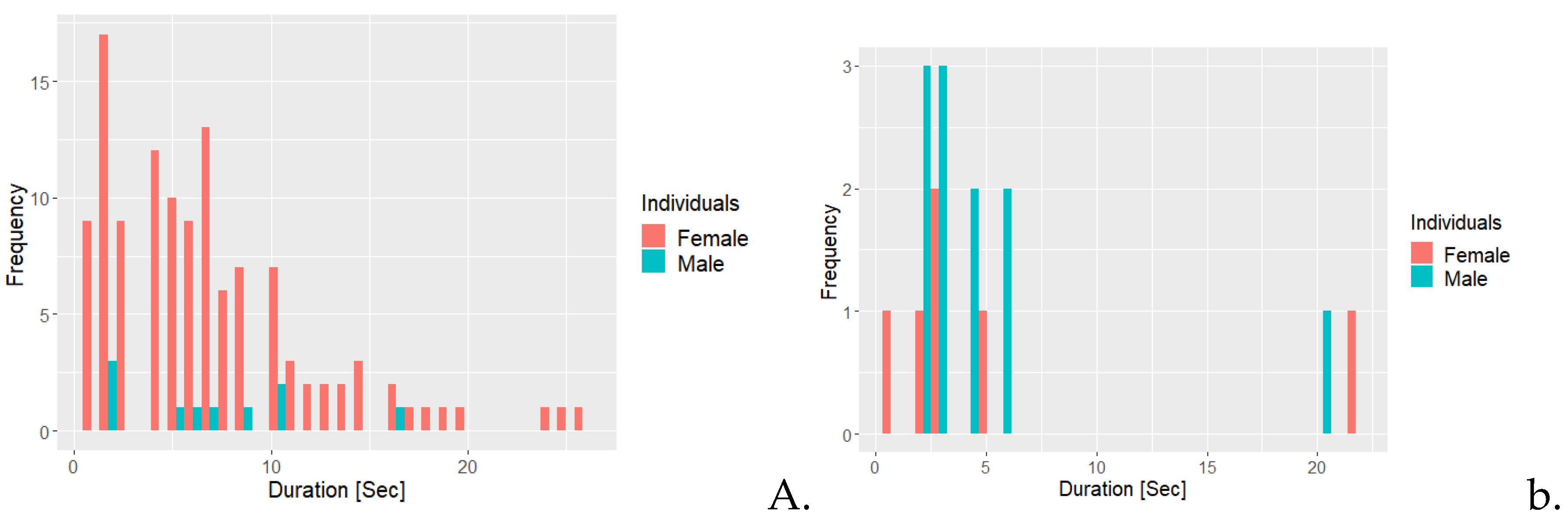
Figure 3.
Frequency histogram illustrating the frequency for duration of the behaviours A. positive interaction with conspecific and B. scent marking for the golden lion tamarin individuals, the female and the male.
Figure 3.
Frequency histogram illustrating the frequency for duration of the behaviours A. positive interaction with conspecific and B. scent marking for the golden lion tamarin individuals, the female and the male.
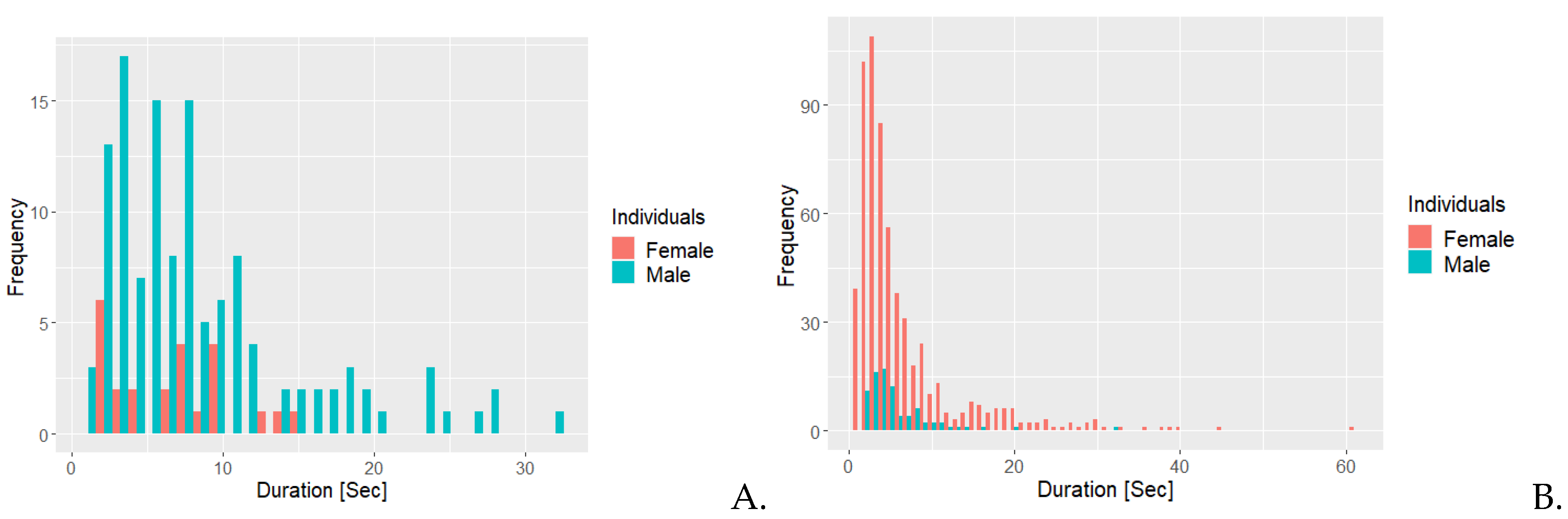
Figure 4.
Frequency histogram illustrating the frequency for duration of the behaviours A. self grooming and B. interaction with a branch for the golden lion tamarin individuals, female and male.
Figure 4.
Frequency histogram illustrating the frequency for duration of the behaviours A. self grooming and B. interaction with a branch for the golden lion tamarin individuals, female and male.
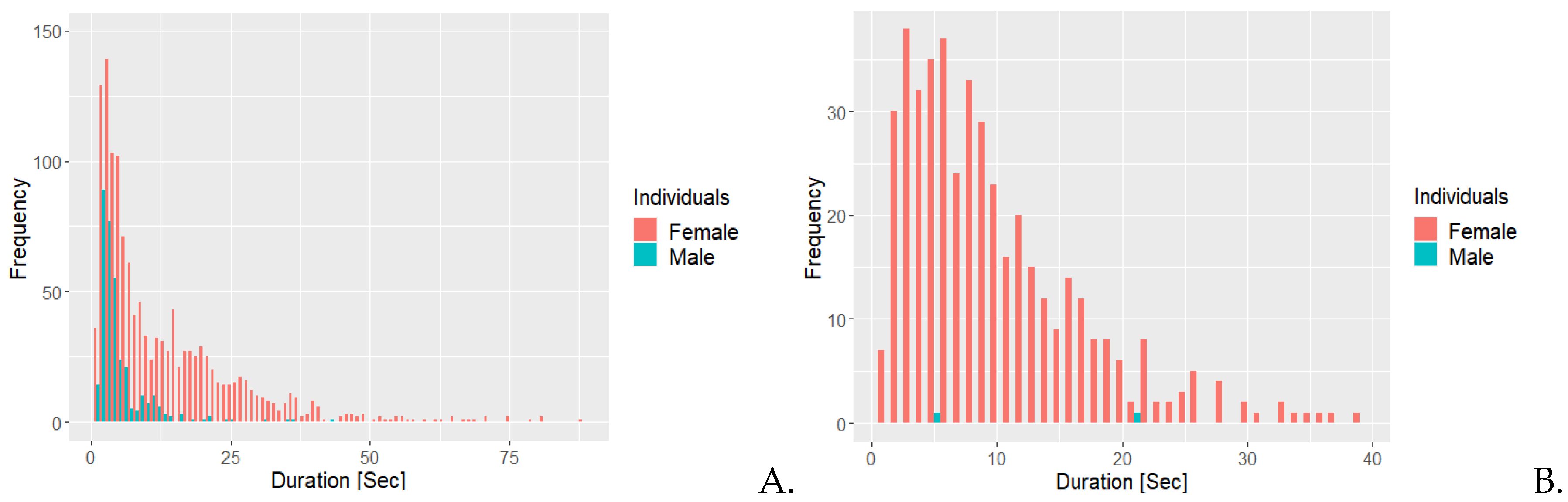
Figure 5.
Frequency histogram illustrating the sequence of time between golden lion tamarin, the female, interactions with a branch and attendance of tail during self grooming. The behaviour pattern is regarded as stereotypical behaviour.
Figure 5.
Frequency histogram illustrating the sequence of time between golden lion tamarin, the female, interactions with a branch and attendance of tail during self grooming. The behaviour pattern is regarded as stereotypical behaviour.
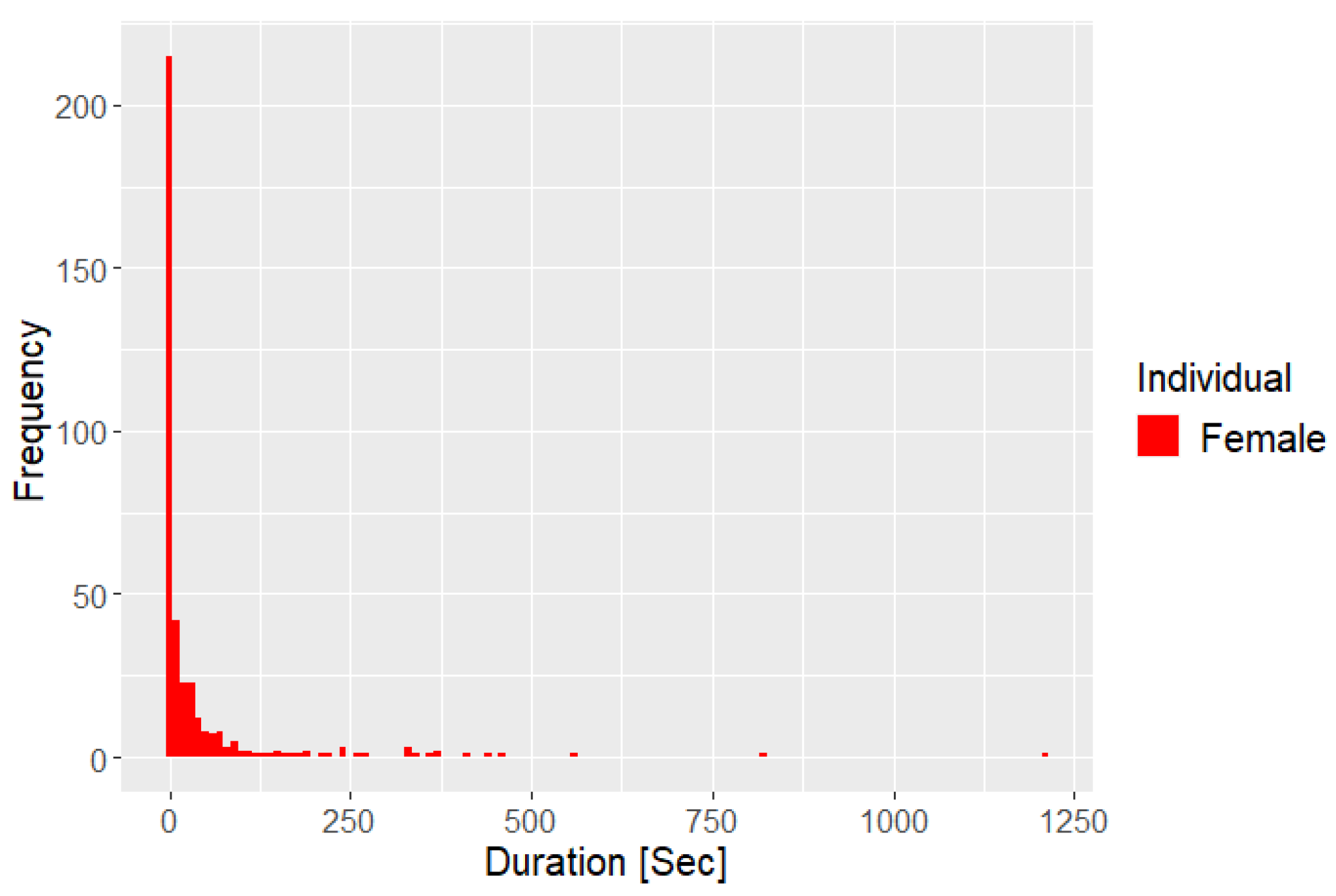
Figure 6.
Cumulative graph illustrating the cumulative time spent on A. allogrooming and B. negative interaction with conspecific by the two golden lion tamarin individuals, the female and the male, during observations.
Figure 6.
Cumulative graph illustrating the cumulative time spent on A. allogrooming and B. negative interaction with conspecific by the two golden lion tamarin individuals, the female and the male, during observations.
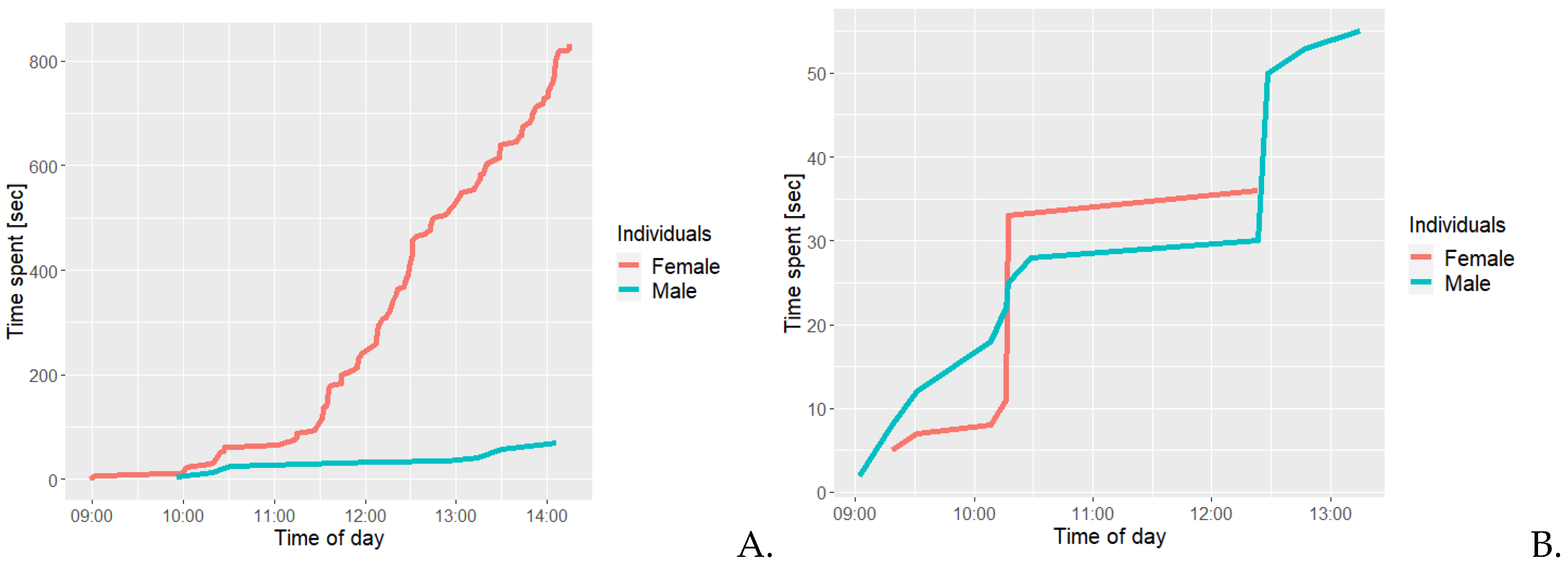
Figure 7.
Cumulative graph illustrating the cumulative time spent on being A. positive interactions with conspecific and B. scent marking by the two golden lion tamarin individuals, the female and the male, during observations.
Figure 7.
Cumulative graph illustrating the cumulative time spent on being A. positive interactions with conspecific and B. scent marking by the two golden lion tamarin individuals, the female and the male, during observations.
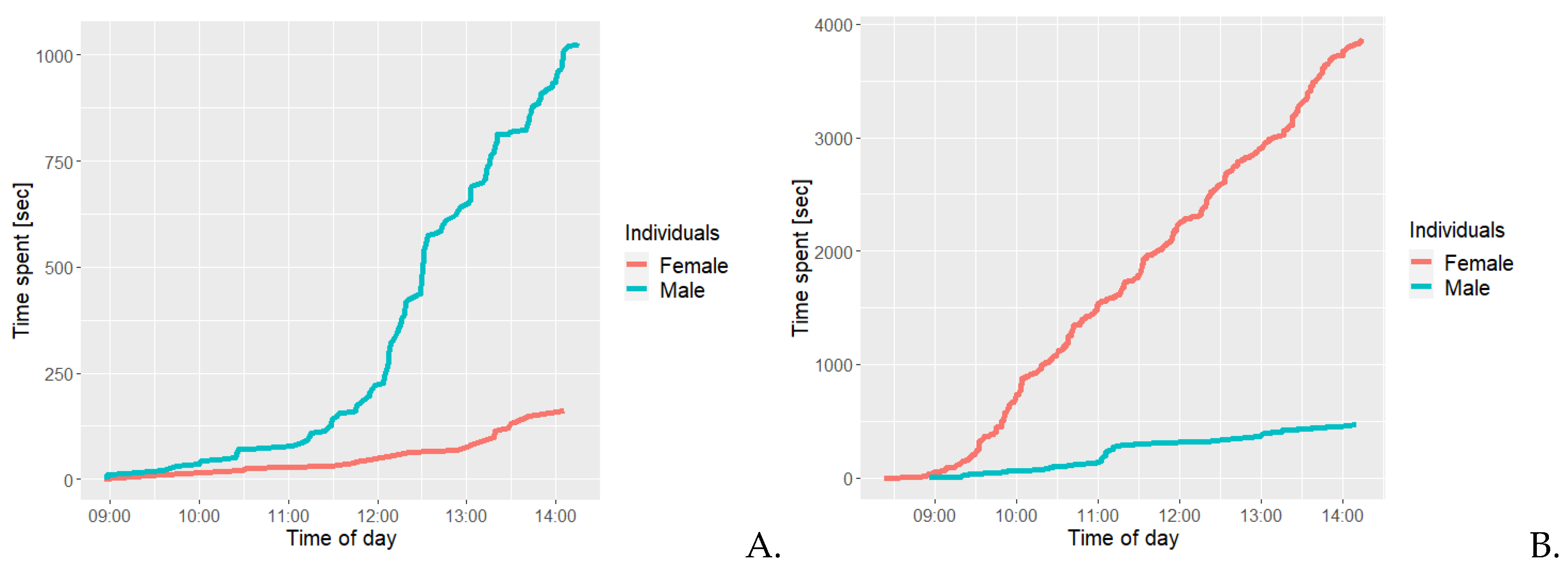
Figure 8.
Cumulative graph illustrating the cumulative time spent on being A self grooming and B interactions with a branch by the two golden lion tamarin individuals, the female and the male , during observations.
Figure 8.
Cumulative graph illustrating the cumulative time spent on being A self grooming and B interactions with a branch by the two golden lion tamarin individuals, the female and the male , during observations.
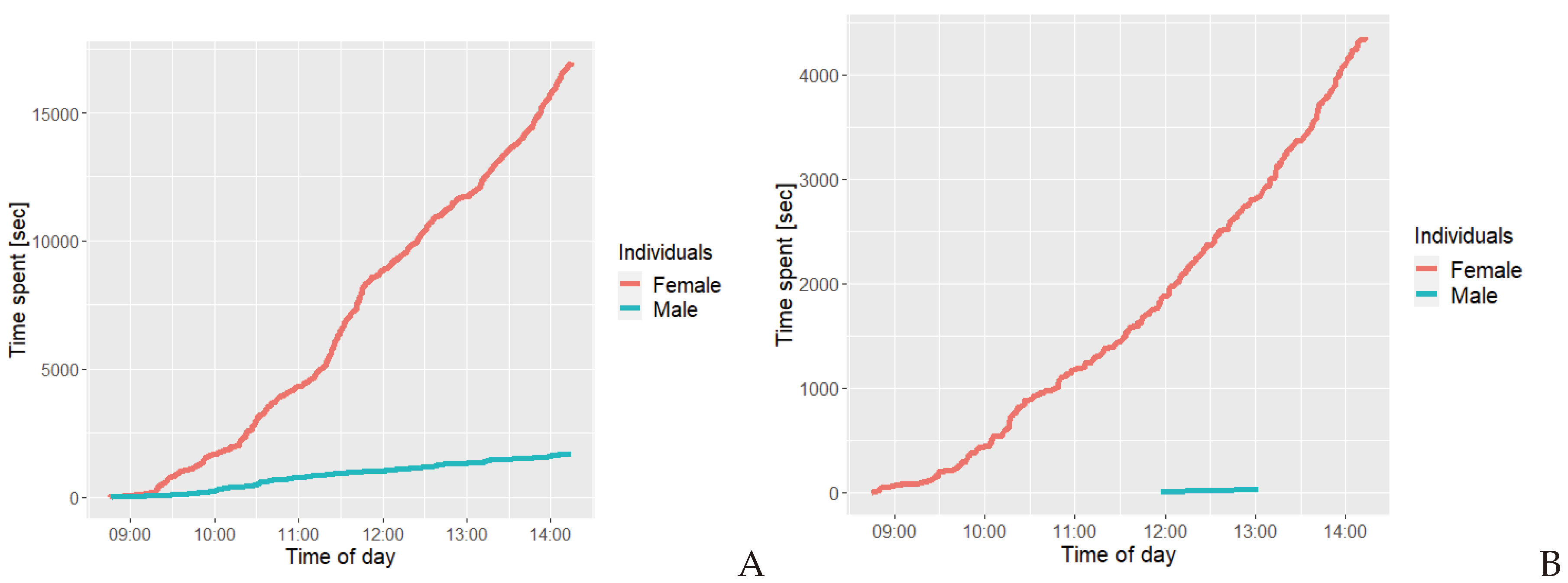

Table 1.
Ethogram describing the behaviours noted during observations of the two golden lion tamarin individuals.
Table 1.
Ethogram describing the behaviours noted during observations of the two golden lion tamarin individuals.
| Behaviour | Description |
|---|---|
| Allogrooming | Licking, scratching, and pruning the other individual gently |
| Negative interaction with conspecific | Recipient of antagonistic interactions from conspecific (threat display, teeth baring, stealing food) |
| Positive interaction with conspecific | Recipient of gentle or playful interactions from conspecific without evidence of antagonistic behaviour |
| Scent marking | Individual rubbing abdomen or genitalia on branches in enclosure |
| Self grooming | Licking, pruning, or scratching self |
| Interactions with a branch | Touching, rubbing against, or looking at specific branches in the enclosure |
| Others | Locomotion, scanning, foraging, going in nestbox, excretion, yawning, vocalisation, inactive, decreasing and increasing distance from conspecific, out of view |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



