
Submitted:
08 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
Probiotics play an important role as immunostimulants in improving growth and health status in fish. However, little is known on the effects of probiotics when fish offered a low fish meal diet. This study evaluated the effects of three probiotic bacteria isolated from the fish intestine to replace antibiotics in a reduced fish meal diet fed to the juvenile olive flounder Paralichthys olivaceus. A basal diet without feed additives was used as a control (CON) and five other diets were formulated by supplementation with Bacillus subtilis WB60 (BSWB60) at 1×108 CFU/g, B. subtilis SJ10 (BSSJ10) at 1×108 CFU/g, Enterococcus faecium SH30 (EFSH30) at 1×107 CFU/g, amoxicillin (AMO) at 4 g/kg and oxytetracycline (OTC) at 4 g/kg of the basal diet. Triplicate groups of 25 fish averaging 12.1±0.09 g (mean±SD) were fed one of the six experimental diet groups for 8 weeks. Then, 45 fish from each group were intraperitoneally injected with the pathogenic bacteria, Edwardsiella tarda, and mortality was recorded for 14 days. At the end of the feeding trial, weight gain, specific growth rate and feed efficiency of fish fed the BSWB60 diet were significantly greater than those of fish fed the CON, OTC and AMO diets (P < 0.05). Furthermore, feeding efficiency and protein efficiency ratio of fish fed the BSWB60 diet were significantly greater than those of fish fed the CON, OTC and AMO diets. However, there were no significant differences in survival, hepatosomatic indices, viscerosomatic indices and condition factors among the fish fed the experimental diets. The whole-body proximate composition of the fish was unaffected by the dietary treatments. The serum biochemistry parameters, such as aspartate aminotransferase levels were significantly greater in fish fed the BSSJ10 diet than in those fed the BSWB60 diet. However, serum alanine aminotransferase, glucose and total protein levels were not significantly affected by the experimental diets. The lysozyme activity of fish fed the BSSJ10, BSWB60 and EFSH30 diets was significantly greater than that of the CON, OTC and AMO diets. The myeloperoxidase activity of fish fed the BSWB60 diet was significantly greater than that of the fish fed the CON and AMO diets. The flounder growth hormone levels of fish fed the BSSJ10, BSWB60, EFSH30 and AMO diets were significantly greater than that of the fish fed the CON diet. The interleukin 1β (IL-1β) gene expression levels in fish fed the BSSJ10 and EFSH30 diets were significantly greater than those in fish fed the CON, OTC and AMO diets. The interleukin 10 (IL-10) gene expression levels in fish fed the BSSJ10, BSWB60, EFSH30 and OTC diets were significantly greater than those of fish fed the CON and AMO diets. Intestinal histology revealed that the average villus length of fish fed the BSSJ10, BSWB60, and EFSH30 diets was significantly greater than that of fish fed the CON, OTC and AMO diets. The cumulative survival rates of fish fed the EFSH30 and BSWB60 diets were significantly greater than those of fish fed the CON diet after the 15th day of the challenge test. Overall, the results of the present study demonstrated that the supplementation of autochthonous bacteria, B. subtilis (108 CFU/g diet) or E. faecium (107 CFU/g diet) in the diet could be ideal probiotics to replace antibiotics based on growth performance, the serum biochemical composition, immune responses and disease resistance in olive flounder fed a low fish meal diet..
Keywords:
probiotics
; growth
; immune genes
; histology
; disease resistance
; olive flounder
1. Introduction
Olive flounder (Paralichthys olivaceus) is a flat fish native to the temperate coastal waters of East Asia that represents both an important capture as well as aquaculture industry. Olive flounder is most popular in Northeast Asian nations such as Korea, China, and Japan [1,2]. In South Korea, it is the most cultured species at more than 46,000 MT and accounts for more than 50% of the country’s overall production, which was 91,000 MT in 2022 [3]. As olive flounder culture exceeds the capture fishery, there is an ever increasing pressure to reduce its reliance on fish meal (FM). FM accounts for up to 50% of the cost of olive flounder aqua feed. Although FM is an ideal source of essential amino acids (EAAs) and fatty acids (FAs), it comes at the expense of increased production for producers and great pressure on wild fisheries. Since the world’s production of FM is in decline and will unlikely increase in the future [4], the sustainability of the aquaculture industry is dependent on reducing reliance on this commodity moving forward.
Fish meal replacement has been a major objective for aquaculture nutritionists for several decades. There are many candidates for FM replacement, such as meat and bone meal [5], poultry by-product meal [6], blood meal [7] and plant-based protein sources such as soybean meal [8]. Proteins obtained from animal sources have great potential with some of the most popular sources being poultry byproduct meals, blood meals, and meat and bone meal [9]. However, diets with low FM often suffer from issues related to growth, digestibility, palatability, and disease resistance. To overcome these problems, fish farms have started to rely on various feed additives and antibiotics. The excessive application of the antibiotics is becoming a major health and environmental hazard since antibiotic-resistant strains are becoming increasingly prevelant [10,11]. Thus, the development of safe and sustainable alternatives to these pharmaceuticals has become an ever expanding field of research.
One of the most promising tools for reducing antibiotic reliance in the aquaculture industry is probiotics. Probiotics are microbes that help to correct imbalances in the microflora of the intestines and confer benefits to the host organism’s health status when consumed in sufficient amounts [12]. Some of the specific benefits of probiotics include their ability to retard and outcompete the growth of pathogens [13], aid in digestion by contributing to enzymatic activities [14], antiviral properties, and enhancing immune responses [15]. Recently, there has been tremendous interest in this field of research within the aquaculture industries. Therefore, the present study has been designed to evaluate the effects of dietary supplementation of the three intestinally isolated (autochthonous) probiotics, Bacillus subtilis WB60, Bacillus subtilis SJ10 and Enterococcus faecium on the growth and health status of juvenile olive flounder fed a low FM-diet.
2. Materials and Methods
2.1. Ethics Statement
This experiment was conducted under the guidelines of Institutional Animal Care and Use Committee Regulations, No. 554, issued by Pukyong National University, Busan, Republic of Korea. Every effort was taken to minimize the number of fish sacrificed.
2.2. Bacterial isolation and culture condition
The strain B. subtilis WB60 was isolated from the intestines of healthy Japanese eel and was identified by cluster analysis via 16S rDNA sequencing. The B. subtilis WB60 was isolated according to Lee et al. [16] and incubated at 30°C for 72 h in Luria-Bertani broth (LB broth; Sigma-Aldrich, St. Louis, USA), after which the optical density (OD600) was measured at 600 nm using spectrophotometry. The B. subtilis SJ10 was isolated from jeotgal, a traditional Korean fermented dish made from salt-preserved seafood such as squid, pollock roe, and shrimp, according to Hasan et al. [17] and it was incubated from a single colony on lysogeny broth (LB, USB Corporation, USA) agar, and was subsequently cultured in 10 ml of LB broth for 16 h at 37°C in a shaking incubator. Furthermore, E. faecium SH30 was isolated from the intestine of healthy Nile tilapia, and the bacteria were grown in MRS (deMan, Rogosa, and Sharpe) broth at 36°C for 48 h according to Xia et al. [18]. All probiotics were washed in sterile saline and the concentration of the final suspension was calculated to be 1 × 108 CFU/g for WB60 and SJ10, and 1 × 107 CFU/g for SH30 in the diets.
2.3. Experimental Fish and Feeding Trial
Juvenile olive flounder were obtained from a private farm (JUNGANG Fisheries, Chungcheongnam-do, Taean-gun, Republic of Korea). Prior to the start of the feeding trial, the apparent health status of the fish was checked visually, and the fish were starved for 24 h. All the fish were then fed a commercial diet for two weeks prior to the start of the feeding trial to acclimatize to the laboratory conditions. On average, 12.1±0.04 g (mean±SD) of fish were weighed, divided into triplicate groups of 25 fish corresponding to the dietary treatment, and randomly distributed into twelve 40 L indoor fiberglass tanks receiving a constant flow (1.2 L/min) of filtered seawater. During the experiment, supplemental aeration was provided in each tank to maintain adequate dissolved oxygen. The temperature was maintained at 19.0±1.0 ºC throughout the experiment by electric heaters in a concrete reservoir. Fish were fed twice a day (09:00 and 19:00) for 8 weeks at a rate of 2.5~5% body weight per day. Dead fish were immediately removed and weighed, after which the amount of feed provided to the remaining fish was adjusted. The uneaten feed was siphoned 1 hour after feeding. The inside of the tanks was scrubbed once per week to minimize algal and fungal growth.
2.4. Experimental DIETS
The basal diet formulation is shown in Table 1. Anchovy fish meal (68.75% CP) and soybean meal (47.04% CP) were used as the main protein sources, while fish oil was used as the main lipid source. The feed additives (probiotics) used in this experiment were B. subtills WB60 (1 × 108 CFU/g), B. subtills SJ10 (1 × 108 CFU/g), E. faecium SH30 (1 × 107 CFU/g), OTC (oxytetracycline 4 g/kg) and AMO (amoxicillin 4 g/kg). The procedures for feed manufacturing and preparation were performed as previously described by Lee et al. [16]. According to the feed formulation table, all fine powdered ingredients were mixed thoroughly with an electric mixer (HYVM-1214, Hanyoung Food Machinery, Republic of Korea). Then, a stiff dough was formed by adding fish oil and the desired amount of water (~10%). The dough was passed through a pellet machine (SFD-GT, Shinsung, Republic of Korea) with a 0.2 cm die. The prepared diets were air-dried in a drying room for 48 hours, broken into smaller pieces and stored at -20oC. According to the proximate composition analysis, shown in Table 2, all the diets were iso-nitrogenous and iso-lipidic.
2.5. Sample Collection and Analysis
At the end of the feeding trial, fish were starved for 24 h prior to sample and data collection. The fish were subsequently counted and weighed to calculate the final weight (FW), weight gain (WG), specific growth rate (SGR), feed efficiency (FE) protein efficiency ratio (PER) and survival rate (SUR). Four fish from each tank were selected at random, weighed individually, and dissected to obtain liver and visceral metrics for calculation of hepatosomatic index (HSI) and visceral somatic index (VSI); thereafter, the same intestinal samples were used for histological observation and enzyme activity. Three additional fish per tank were captured at random and anesthetized with ethylene glycol phenyl ether (200 mg/L for 5–10 min). After this, blood was drawn from the caudal vein, which was subsequently centrifuged at 5000 ×g for 10 min to obtain the serum. Serum samples were then stored at -70°C for the analysis of non-specific immune responses, such as superoxide dismutase (SOD), lysozyme, and myeloperoxidase (MPO) activities, in addition to biochemical parameters, including aspartate aminotransferase (AST), alanine aminotransferase (ALT), glucose and total protein (TP) levels. The serum levels of AST, ALT, glucose, and total protein were determined by a chemical analyzer (Fuji DRI-CHEM 3500i, Fuji Photo Film Ltd., Tokyo, Japan) following the manufacturer’s instructions.
Three additional fish from each tank were collected for whole-body proximate composition analysis. Proximate composition analyses of both whole fish and experimental diets were performed by the standard methods of AOAC [19]. Whole fish and diet samples were dried at 105°C to a constant weight to determine their moisture content. The ash content was determined by incinerating the samples at 550°C. The protein concentration was determined by using the Kjeldahl method (N×6.25) after acid digestion. Crude lipids were measured by Soxhlet extraction using Soxhlet system 1046 (Tacator AB, Hoganas, Sweden) after the samples were freeze-dried for 20 h.
2.6. Antioxidant Capacity and Non-Specific Immune Response Analyses
Superoxide dismutase (SOD) activity was measured by the superoxide radical dependent reaction inhibition rate of enzyme with water soluble tetrazolium dye (WST-1) substrate and xanthine oxidase using a SOD Assay Kit (Sigma-Aldrich, 19160) according to the manufacturer's instructions. The absorbance at 450 nm (the absorbance wavelength for the colored product of the WST-1 reaction with superoxide) was monitored after 20 minutes of reaction at 37°C. The percentage of inhibition was normalized to mg of protein and is expressed as SOD units/mg. Olive flounder serum lysozyme activity was analyzed as follows: 0.1 ml of test serum was added to 2 ml of a suspension of Micrococcus lysodeikticus (0.2 mg/ml) in 0.05 M sodium phosphate buffer (pH 6.2). The reactions were carried out at 20℃, and the absorbance was measured at 530 nm. Measurements were taken between 0.5 min and 4.5 min on a spectrophotometer. One lysozyme activity unit was defined as the amount of enzyme that produced a decrease in absorbance corresponding to 0.001/min. Myeloperoxidase activity was measured according to the method described by Quade and Roth [19]. Briefly, 20 µL of serum was diluted with Hank’s balanced salt solution (HBSS) without Ca2+ or Mg2+ (Sigma- Aldrich) in 96-well plates. Then, 35 µL of 3, 3’, 5, 5’ tetramethylbenzidine hydrochloride (TMB, 20 mM) (Sigma-Aldrich) and H2O2 (5 mM) were added. The color change reaction was stopped after 2 min by adding 35 µL of 4 M sulfuric acid. Finally, the optical density was read at 450 nm in a microplate reader.
2.7. Real-Time PCR
Tissue fragments from HK were obtained and immediately stored at -80 °C in TRIzol reagent (Thermo Fisher Scientific) for RNA extraction. Total RNA was extracted from 0.5 g of olive flounder tissue using TRIzol Reagent (Thermo Fisher Scientific, San Jose, CA, USA). Afterwards, the RNA was quantified and the purity was assessed spectrophotometrically. The RNA was then treated with DNase I (Cosmogenetech, Seoul, Republic of Korea) to remove genomic DNA contamination. Complementary DNA (cDNA) was synthesized using M-MuLV reverse transcriptase (Cosmogenetech). The expressions of four selected immune-related genes were analyzed by real-time quantitative polymerase chain reaction (RT-qPCR), which was performed with a Bio-Rad CFX96 (Bio-Rad, Hercules, CA, USA) using SYBR Green PCR Core Reagents (Cosmogenetech). The relative expression levels of the target gene transcripts (FGH, IL-1B, IL-10) were measured with GAPDH as an internal control were using CFX Manager software version 2.0 (Bio-Rad) (Table 3). In all the cases, each PCR was performed with triplicate samples.
2.8. Challenge Test
After sampling, seven fish from each tank were redistributed into 27 tanks in a non-recirculating system without water renewal to perform the 9-challenge test. The pathogenic bacterium, Edwardsiella tarda (E. tarda) FSW910410 was obtained from the Department of Biotechnology, Pukyong National University, Busan, Rep. Korea. The bacteria were originally sourced from diseased olive flounder and cultured on tryptic soy agar (TSA, Sigma) plates (24 h at 27°C). All the fish were subjected to intraperitoneal injection of 50 μl of E. tarda (3×108 CFU/ml) solution. The water temperature was maintained at 19±1.0 °C (mean±SD) during 15 days of challenge test and fish mortalities were recorded daily from each tank. Dead fish were necropsied and kidney samples were taken and streaked on Salmonella-Shigella agar (SS agar, Difco). The presence of black pigments confirmed E. tarda infection.
2.9. Histology
The anterior intestinal tissues from the fish were dissected and fixed in 10% neutral buffered formalin, dehydrated in a graded ethanol series and embedded in paraffin. The tissue blocks were sectioned (4 μm thick) and stained with hematoxylin and eosin (H&E). At least 6 tissue sections from each sample were examined under an AX70 Olympus (Japan) microscope.
2.10. Statistical Analysis
All the data were analyzed by one-way ANOVA (Statistix 3.1; Analytical Software, St. Paul, MN, USA) to test the effects of the dietary treatments. When a significant treatment effect was observed, an LSD test was used to compare the means. Treatment effects were considered significant at the P<0.05 level.
3. Results
3.1. Growth Performances
Table 4 shows the growth performances of juvenile olive flounder fed different experimental diets for 8 weeks. At the end of the feeding trial, WG and SGR of fish fed the BSWB60 diet were significantly higher greater those of fish fed the CON diet (P>0.05). The FE of fish fed the BSWB60 diet was significantly greater than that of fish fed the CON, OTC, or AMO diets (P>0.05). The PER of fish fed the BSWB60 diet was significantly greater than that of fish fed the CON and AMO diets (P>0.05). Moreover, probiotics supplemented diets were not significantly different from fish fed the BSWB60 diet (P>0.05) in terms of growth performance. Furthermore, there were no significant differences (P>0.05) in terms of SUR, CF, HSI or VSI among fish fed the experimental diets.
3.2. Whole-Body Proximate Composition
There were no significant differences (P>0.05) in terms of crude protein, lipid, moisture or ash content among any of the group of fish fed the experimental diets for 8 weeks (Table 5).
3.3. Hematological Parameters
The blood parameters of the juvenile olive flounder fed the experimental diets are shown in Table 6. The aspartate aminotransferase (AST) level in the fish fed the BSSJ10 diet was significantly greater than that of fish fed the BSWB60 diet (P>0.05). There were no significant differences in alanine aminotransferase (ALT), glucose, or total protein content among any of the group of fish fed the experimental diets for 8 weeks.
3.4. Non-Specific Immune Responses
Table 7 shows the non-specific immune responses of juvenile olive flounder fed different experimental diets for 8 weeks. The lysozyme activities of fish fed the BSSJ10, BSWB60, EFSH30 and AMO diets were significantly greater than that of fish fed CON and OTC diets (P>0.05). Myeloperoxidase activity of fish fed BSWB60 was significantly higher than those from the CON and AMO diets (P>0.05).
3.5. Growth and Immune Related Gene Expressions
The gene expression profiles of the immunological parameters in the intestine of olive flounder fed diets supplemented with probiotics are presented in Figure 1. The mRNA expression levels of the flounder growth hormone gene (FGH) in fish fed the BSSJ10, BSWB60, EFSH30 and AMO diets were significantly greater than those in fish fed the CON diet (P˂0.05). Moreover, interleukin 1-1β (IL-1β) expressions in fish fed the BSSJ10 and EFSH30 diets were significantly greater than those in fish fed the CON and AMO diets (P˂0.05). Interleukin 10 (IL-10) expressions in fish fed the BSSJ10, BSWB60, EFSH30 and OTC diets were significantly greater than those in fish fed the CON and AMO diets (P˂0.05). Moreover, the fish fed the BSSJ10 and EFSH30 diets presented higher FGH, IL-1β, and IL-10 expressions than did the fish fed all the other experimental diets (P˂0.05).
3.6. Histology
Histological analysis of the anterior intestine of olive flounder fed different experimental diets for 8 weeks is shown in Figure 2. The fish fed the BSSJ10, BSWB60, EFSH30, OTC and AMO diets clearly exhibited better intestinal histomorphology with more massive villus in comparison to the CON diet (Fig 2A). In addition, image (2a), which corresponds to the CON group (Fig. 2b), shows certain untidiness and improper arrangement of villi compared to those of the other groups.
3.7. Challenge Test
The percent cumulative survival of juvenile olive flounder challenged with E. tarda for 15 days is shown in Figure 3. During the challenge test, the first mortalities occurred on the second day. At the end of the 15 day challenge, the percent cumulative survivals of fish fed the BSWB60 and EFSH30 diets were significantly greater than that of fish fed the CON diet (P<0.05).
4. Discussion
Research into the use of probiotics in aquaculture nutrition has attracted much interest due to their health benefits and because they are considered environmentally friendly [20]. In recent years, probiotic effects have been studied in different fish species [16,21]. Additionally, many trials have investigated the extraction of probiotic strains from the intestines of various fish, which are subsequently added to aquafeeds. This strategy of sourcing and using probiotics from these species has improved growth performance, feed efficiency and immune response [22,23]. Therefore, to build on this growing body of knowledge, the present study utilized three different probiotics according to the results of previous experiments.
The two probiotics used were Bacillus subtilis extracted from the intestines of Japanese eel and jeotgal [24,25], because most other probiotic studies have focused on the use of Bacillus spp. The other probiotics used was E. faecium isolated from the intestine of healthy Nile tilapia. Additionally, a previous experiment in which Arapaima gigas was fed diets containing E. faecium at a concentration of 1 × 108 CFU/g showed increased weight gain compared to that in the control treatment group [26]. The results showed that all the experimental diets containing probiotics resulted in increased weight gain and feed efficiency compared with the control diet. These results are likely due to the increased secretion of proteolytic enzymes, which increase feed efficiency, similar to the findings of probiotic experiments in olive flounder [21]. Likewise, beneficial effects of probiotics have also been reported on growth performance in terms of improving weight gain, specific growth rate, and feed efficiency in Oreochromis niloticus [27,28,29,30].
Modulation of the immune system is one of the most common benefits of probiotics [29]. Lysozyme activity is frequently used as an indicator of non-specific immune functions and is the principle means of combating infections in fish. This enzyme not only has bacteriolytic activity against gram-positive and gram-negative bacteria [30], but also has anti-inflammatory and antiviral properties. Myeloperoxidase (MPO) is another important enzyme that utilizes oxidative radicals to produce hypochlorous acid, which kills pathogens. In the present study, the immune parameters, including MPO activity were measured and the beneficial effects of both probiotic bacteria on nonspecific immune related enzyme responses, were clearly shown to be greatest for the olive flounder fed B. subtilis, at the 108 CFU/g (BSWB60, BSSJ10) and E. faecium, at the 108 CFU/g (EFSH30).
Growth hormone is a hormone that stimulates the secretion of IGF-1 in the liver, increases the concentration of glucose and vitreous acid [31], produces IGF-1 induced protein synthesis [32], and is reported to be an indicator of growth factors in fish such as promoting cell division [33]. In the present study, olive flounder fed probiotics supplemented diets exhibited significantly greater FGH expression than did those fed the control diet (Fig 1-1). Similarly, previous studies in which probiotics were added showed high FGH values [34,35].
Interleukin-1 beta (IL-1β) is one of the earliest expressed pro-inflammatory cytokines and enables organisms to respond promptly to infection by inducing a cascade of reactions leading to inflammation [36]. Many of the effector roles of IL-1β are mediated through the up- or down-regulation of the expressions of other cytokines and chemokines [37]. Mammalian IL-1β is produced by a wide variety of cells, but mainly by blood monocytes and tissue macrophages. IL-1β was the first interleukin to be characterized in fish and has since been identified in a number of fish species, such as rainbow trout [38], carp [39], seabass [40], gilt head seabream [41], haddock [42], tilapia [43]. Interleukin-10 (IL-10) on the other hand, is an anti-inflammatory cytokine that down-regulates the expression of pro-inflammatory cytokines [44]. Additionally, interleukin-10 (IL-10) was initially discovered to be an inhibitory factor for the production of Th1 cytokines. Subsequently, pleiotropic inhibitory and stimulatory effects of IL-10 on various types of blood cells were described, including its role as a survival and differentiation factor for B cells. IL-10, which is produced by activated monocytes, T cells and other cell types, such as keratinocytes, appears to be a crucial factor for at least some forms of peripheral tolerance and a major suppressor of the immune response and inflammation. The inhibitory function of IL-10 is mediated by the induction of regulatory T cells [45]. In the present study, the activities of IL-1β and IL-10 in the blood of the fish that were administered probiotics were significantly greater than those in the control with low FM diet. Therefore, dietary probiotics appears to increase the immune function of fish.
Intestinal morphological parameters (villus length and muscular layer thickness) are indicative of a healthy gut in fish. The intestine is very important for the digestion and absorption of nutrients. The length of the intestinal villi determines the absorption of nutrients in the GI tract (gastrointestinal tract) [46,47]. Thus, digestive function is associated with intestinal development [29,47]. In this study, the beneficial effects of probiotics on intestinal morphology were clearly observed. The length of the villi increased in a dose dependent manner and villi length was significantly highest for the olive flounder fed the BSSJ10, BSWB60, and EFSH30 diets (Fig. 2a). In the same manner, Lee et al. [16] reported that probiotics are capable of increasing the villus length in the proximal intestine of Japanese eel. With regard to disease resistance, olive flounder fed with probiotics at 107 and 108 CFU/g in the diet exhibited the highest disease resistance compared to other treatment groups.
5. Conclusions
In conclusion, the present study revealed the potential benefits of supplementation with the bacteria species, B. subtilis and E. faecium as probiotics in the diet of olive flounder. Therefore, B. subtilis at 108 CFU/g and E. faecium at 107 CFU/g could be ideal probiotics for improving growth performance, immune responses, enzyme activity and disease resistance, while replacing the dietary supplementation of antibiotics in juvenile olive flounder fed a reduced FM diet.
Author Contributions
W.C.: planned the experiment; formulated the feed; determined the growth, somatic indices, non-specific immune enzyme activities, and gene expressions; and drafted the manuscript. M.M.: performed the statistical analyses, drafted and finalized the manuscript. J.B. and S.L. (Seunghan Lee): performed the investigation and statistical analyses and drafted the manuscript. T.M.: reviewed the manuscript. S.L. (Seunghyung Lee): reviewed and finalized the manuscript. S.C.B.: planned the experiment, supervised the project and gave final approval of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Feeds & Foods Nutrition Research Center (FFNRC), Pukyong National University (PKNU), Busan, Republic of Korea.
Institutional Review Board Statement
This experiment was conducted under the guidelines of Institutional Animal Care and Use Committee Regulations, No. 554, issued by Pukyong National University, Busan, Republic of Korea. Every effort was taken to minimize the number of fish sacrificed.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Acknowledgments
Mohammad Moniruzzaman acknowledge the Brain Pool program funded by the Ministry of Science and ICT through the National Research Foundation of Korea (grant number 2019H1D3A1A01101555).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fuji, K; Kobayashi, K. ; Hasegawa, O.; Coimbra, M.R.M.; Sakamoto, T.; Okamoto, N. Identification of a single major genetic locus controlling the resistance to lymphocystis disease in Japanese flounder (Paralichthys olivaceus). Aquaculture, 2006, 254, 203–210. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Won, S.; Lee, S.; Lee, S.; Farris, N.W.; Bai, S.C. Nutrition and feeding of olive flounder Paralichthys olivaceus: A Review. Rev. Fish. Sci. Aquac. 2020, 28, 340–357. [Google Scholar] [CrossRef]
- Statistics Korea. Results of the year 2022 fish farming trend survey. Text by Department of Agriculture and Fisheries Trends Department, In: Fish farming trend survey. Daejeon., 2023.https://kostat.go.kr/board.es?mid=a10301080400&bid=225&act=view&list_no=424510 Accessed 20 February 2024.
- Boyd, C.E. Overview of aquaculture feeds: Global impacts of ingredient use, in: Feed Feed. Pract. Aquac., Elsevier, 2015, pp. 3–25.
- Bharadwaj, A.S.; Brignon, W.R.; Gould, N.L.; Brown, P.B.; Wu, Y.V. Evaluation of meat and bone A.S. meal in practical diets fed to juvenile hybrid striped bass Morone chrysops × M. saxatilis. J. World. Aquac. Soc. 2002, 33, 448–457. [Google Scholar] [CrossRef]
- Markey, J.C.; Amaya, E.A.; Davis, D.A. Replacement of poultry by-product meal in production diets for the Pacific white shrimp, Litopenaeus vannamei. J. World. Aquac. Soc. 2010, 41, 893–902. [Google Scholar] [CrossRef]
- Kang, Y.J.; Lee, S.M.; Yang, S.G.; Bai, S.C. Effects of meat meal, blood meal or soybean meal as a dietary protein source replacing fish meal in parrot fish, Oplegnathus fasciatus. J. Aquacult. 1999, 12, 205–212. [Google Scholar]
- McGoogan, B.B.; Gatlin, III D. M. Effects of replacing fish meal with soybean meal in diets for red drum Sciaenops ocellatus and potential for palatability enhancement. J World Aquac Soc. 1997, 28, 374–385. [Google Scholar] [CrossRef]
- Tacon, A.G.J. Feed ingredients for warmwater fish, fish meal and other processed feedstuffs., FAO Fish. Circ. (FAO). No. 856.
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial use in aquaculture re-examined: its relevance to antimicrobial resistance and to animal and human health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Done, H.Y.; Venkatesan, A.K.; Halden, R.U. Does the recent growth of aquaculture create antibiotic resistance threats different from those associated with land animal production in agriculture? AAPS J 2015, 17, 513–524. [Google Scholar] [CrossRef]
- Balcázar, J.L.; De Blas, I.; Ruiz-Zarzuela, I.; Cunningham, D.; Vendrell, D.; Múzquiz, J.L. The role of probiotics in aquaculture. Vet. Microbiol. 2006, 114, 173–186. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Vendrell, D.; De Blas, I.; Ruiz-Zarzuela, I.; Muzquiz, J.L. Probiotics: a tool for the future of fish and shellfish health management. J. Aquac. Trop. 2004, 19, 239–242. [Google Scholar]
- Garriques, D. An evaluation of the production and use of a live bacterial isolate to manipulate the microbial flora in the commercial production of Penaeus vennamei postlarvae in Ecuador., Swim. through Troubl. Water. Proc. Spec. Sess. Shrimp Farming, Aquac. World Aquac Soc 1995, pp 53–59.
- Direkbusarakom, S.; Yoshimizu, M.; Ezura, Y.; Ruangpan, L.; Danayadol, Y. Vibrio spp., the dominant flora in shrimp hatchery against some fish pathogenic viruses. J. Mar. Biotechnol. 1998, 6, 266–267. [Google Scholar] [PubMed]
- Lee, S.; Katya, K.; Hamidoghli, A.; Hong, J.; Kim, D.J.; Bai, S.C. Synergistic effects of dietary supplementation of Bacillus subtilis WB60 and mannanoligosaccharide (MOS) on growth performance, immunity and disease resistance in Japanese eel, Anguilla japonica. Fish Shellfish Immunol. 2018, 83, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.T.; Jang, W.J.; Kim, H.; Lee, B.J.; Kim, K.W.; Hur, S.W.; Lim, S.G.; Bai, S.C.; Kong, I.S. Synergistic effects of dietary Bacillus sp. SJ-10 plus β-glucooligosaccharides as a synbiotic on growth performance, innate immunity and streptococcosis resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2018, 82, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutri. 2020, 6, 69–79. [Google Scholar] [CrossRef] [PubMed]
- AOAC, Official Methods of Analysis, Aquac. Res. sixteenth (1995).
- Quade, M.J. , Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Cha, J.H.; Yang, S.Y.; Woo, S.H.; Song, J.W.; Oh, D.H.; Lee, K.J. Effects of dietary supplementation with Bacillus sp. on growth performance, feed utilization, innate immunity and disease resistance against Streptococcus iniae in olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2012, 45, 35–42. [Google Scholar]
- Park, Y.; Kim, H.; Won, S.; Hamidoghli, A.; Hasan, M.T.; Kong, I.S.; Bai, S.C. Effects of two dietary probiotics (Bacillus subtilis or licheniformis) with two prebiotics (mannan or fructo oligosaccharide) in Japanese eel, Anguilla japonica. Aquac. Nutr. 2020, 26, 316–327. [Google Scholar] [CrossRef]
- Vazirzadeh, A.; Roosta, H.; Masoumi, H.; Farhadi, A.; Jeffs, A. Long-term effects of three probiotics, singular or combined, on serum innate immune parameters and expressions of cytokine genes in rainbow trout during grow-out. Fish Shellfish Immunol. 2020, 98, 748–757. [Google Scholar] [CrossRef]
- Kim, Y.R.; Kim, E.Y.; Lee, J.M.; Kim, J.K.; Kong, I.S. Characterisation of a novel Bacillus sp. SJ-10 β-1, 3--1, 4-glucanase isolated from jeotgal, a traditional Korean fermented fish. Bioprocess Biosyst. Eng. 2013, 36, 721–727. [Google Scholar] [CrossRef] [PubMed]
- da Costa, Sousa, N. et al. Effects of an Enterococcus faecium-based probiotic on growth performance and health of Pirarucu, Arapaima gigas. Aquac. Res. 2019, 50, 3720–3728. [Google Scholar] [CrossRef]
- Abumourad, I.M.K.; et al. Enterococcus faecium probiotic as a growth promoter and its impact on the expression of the host innate immune in cultured Oreochromis niloticus. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 1747–1761. [Google Scholar]
- Lara-Flores, M.M.; Olvera-Novoa, A. The use of lactic acid bacteria isolated from intestinal tract of Nile tilapia (Oreochromis niloticus), as growth promoters in fish fed low protein diets. Lat. Am. J. Aquat. Res. 2013, 41, 490–497. [Google Scholar] [CrossRef]
- Wang, Y.B.; Tian, Z.Q.; Yao, J.T.; Li, W. Effect of probiotics, Enteroccus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture 2008, 277, 203–207. [Google Scholar] [CrossRef]
- Aly, S.M.; Mohamed, M.F.; John, G. Effect of probiotics on the survival, growth and challenge infection in Tilapia nilotica (Oreochromis niloticus). Aquac. Res. 2008, 39, 647–656. [Google Scholar] [CrossRef]
- Helal, R.; Melzig, M.F. In vitro effects of selected saponins on the production and release of lysozyme activity of human monocytic and epithelial cell lines. Sci. Pharm. 2011, 79, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, F.C.; Landon, J. Growth hormone secretion in response to stress in man. Nature 1966, 210, 540–541. [Google Scholar] [CrossRef] [PubMed]
- Bornfeldt, K.E.; Arnqvist, H.J.; Dahlkvist, H.H.; Skottner, A.; Wikberg, J.E.S. Receptors for insulin-like growth factor-I in plasma membranes isolated from bovine mesenteric arteries. Eur. J. Endocrinol. 1988, 117, 428–434. [Google Scholar] [CrossRef]
- Clemmons, D.R.; Busby, W.; Clarks, J.B.; Parker, A.; Duan, C.; Nam, T.J. Modifications of insulin-like growth factor binding proteins and their role in controlling IGF actions. Endocr. J. 1998, 45, S1–S8. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Tang, J.; Cai, J.; Yu, H.; Wang, Z.; Abarike, E.D.; Lu, Y.; Li, Y.; Afriyie, G. In vivo assessment of the probiotic potentials of three host-associated Bacillus species on growth performance, health status and disease resistance of Oreochromis niloticus against Streptococcus agalactiae. Aquaculture 2020, 527, 735440. [Google Scholar] [CrossRef]
- Back, S.J.; et al. The effects of dietary heat-killed probiotics bacteria additives in low-fishmeal feed on growth performance, immune responses, and intestinal morphology in juvenile olive flounder Paralichthys olivaceus. Aquac. Rep. 2020, 18, 100415. [Google Scholar] [CrossRef]
- Reyes-Cerpa, S.; Maisey, K.; Reyes-López, F.; Toro-Ascuy, D.; Sandino, A.M.; Imarai, M. Fish cytokines: current research and application. Fish. Sci. 2021, 87, 1–9. [Google Scholar]
- Dinarello, C.A. Interleukin-1 and its biologically related cytokines. Adv. Immunol. 1989, 44, 153–205. [Google Scholar] [PubMed]
- Pleguezuelos, O.; Zou, J.; Cunningham, C.; Secombes, C.J. Cloning, sequencing, and analysis of expression of a second IL-1 β gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics 2000, 51, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, K.; Shin, D.H.; Nakao, M.; Yano, T. Molecular cloning and expression analysis of carp (Cyprinus carpio) interleukin-1β, high affinity immunoglobulin E Fc receptor γ subunit and serum amyloid A. Fish Shell. Immunol. 2000, 10, 229–242. [Google Scholar] [CrossRef]
- Scapigliati, G.; Buonocore, F.; Bird, S.; Zou, J.; Pelegrin, P.; Falasca, C.; Prugnoli, D.; Secombes, C.J. Phylogeny of cytokines: molecular cloning and expression analysis of sea bass Dicentrarchus labrax interleukin-1β. Fish Shell. Immunol. 2001, 11, 711–726. [Google Scholar] [CrossRef]
- Pelegrín, P.; García-Castillo, J.; Mulero, V.; Meseguer, J. Interleukin-1β isolated from a marine fish reveals up-regulated expression in macrophages following activation with lipopolysaccharide and lymphokines. Cytokine 2001, 16, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Corripio-Miyar, Y.; Bird, S.; Tsamopoulos, K.; Secombes, C.J. Cloning and expression analysis of two pro-inflammatory cytokines, IL-1β and IL-8, in haddock (Melanogrammus aeglefinus). Mol. Immunol. 2007, 44, 1361–1373. [Google Scholar] [CrossRef]
- Lee, D.S.; Hong, S.H.; Lee, H.J.; Jun, L.J.; Chung, J.K.; Kim, K.H.; Jeong, H.D. Molecular cDNA cloning and analysis of the organization and expression of the IL-1β gene in the Nile tilapia, Oreochromis niloticus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 143, 307–314. [Google Scholar] [CrossRef]
- Huo, H.J.; Chen, S.N.; Li, L.; Nie, P. Functional characterization of IL-10 and its receptor subunits in a perciform fish, the mandarin fish, Siniperca chuatsi. Dev. Comp. Immunol. 2019, 97, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Yang, H.J.; Liu, Y.; Chen, S.J.; Guo, D.Q.; Yu, Y.; Tian, L.X. Effects of graded levels of threonine on growth performance, biochemical parameters and intestine morphology of juvenile grass carp Ctenopharyngodon idella. Aquaculture 2014, 424, 113–119. [Google Scholar] [CrossRef]
- Klurfeld, D.M. Nutritional regulation of gastrointestinal growth. Front. Biosci. 1999, 4, D299–302. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative expression levels of a) fish growth hormone (FGH), b) interleukin-1beta (IL-1β) and c) interleukin-10 (IL-10) mRNA expressions of intestine from olive flounder fed the experimental diets for 8 weeks.
Figure 1.
Relative expression levels of a) fish growth hormone (FGH), b) interleukin-1beta (IL-1β) and c) interleukin-10 (IL-10) mRNA expressions of intestine from olive flounder fed the experimental diets for 8 weeks.
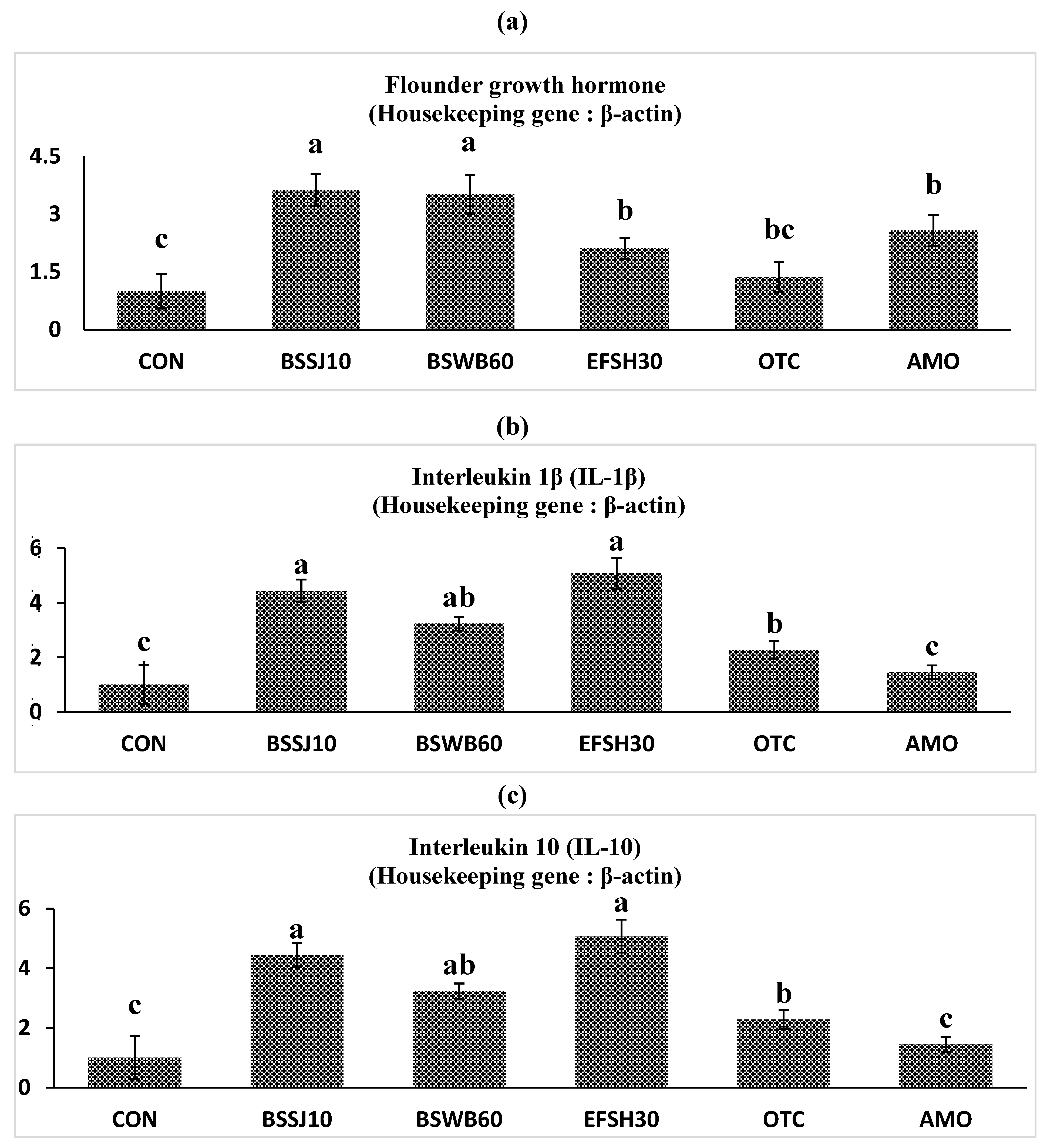
Figure 2.
Intestinal histology of juvenile olive flounder fed the experimental diets for 8 weeks; a) villi lengths in fish fed the different diets; b) histological photomicrographs of (A) CON (B) BSSJ10 (C) BSWB60 (D) EFSH30 (E) OTC (F) AMO diet groups; (scale bar=100 μm; original magnification×40).
Figure 2.
Intestinal histology of juvenile olive flounder fed the experimental diets for 8 weeks; a) villi lengths in fish fed the different diets; b) histological photomicrographs of (A) CON (B) BSSJ10 (C) BSWB60 (D) EFSH30 (E) OTC (F) AMO diet groups; (scale bar=100 μm; original magnification×40).
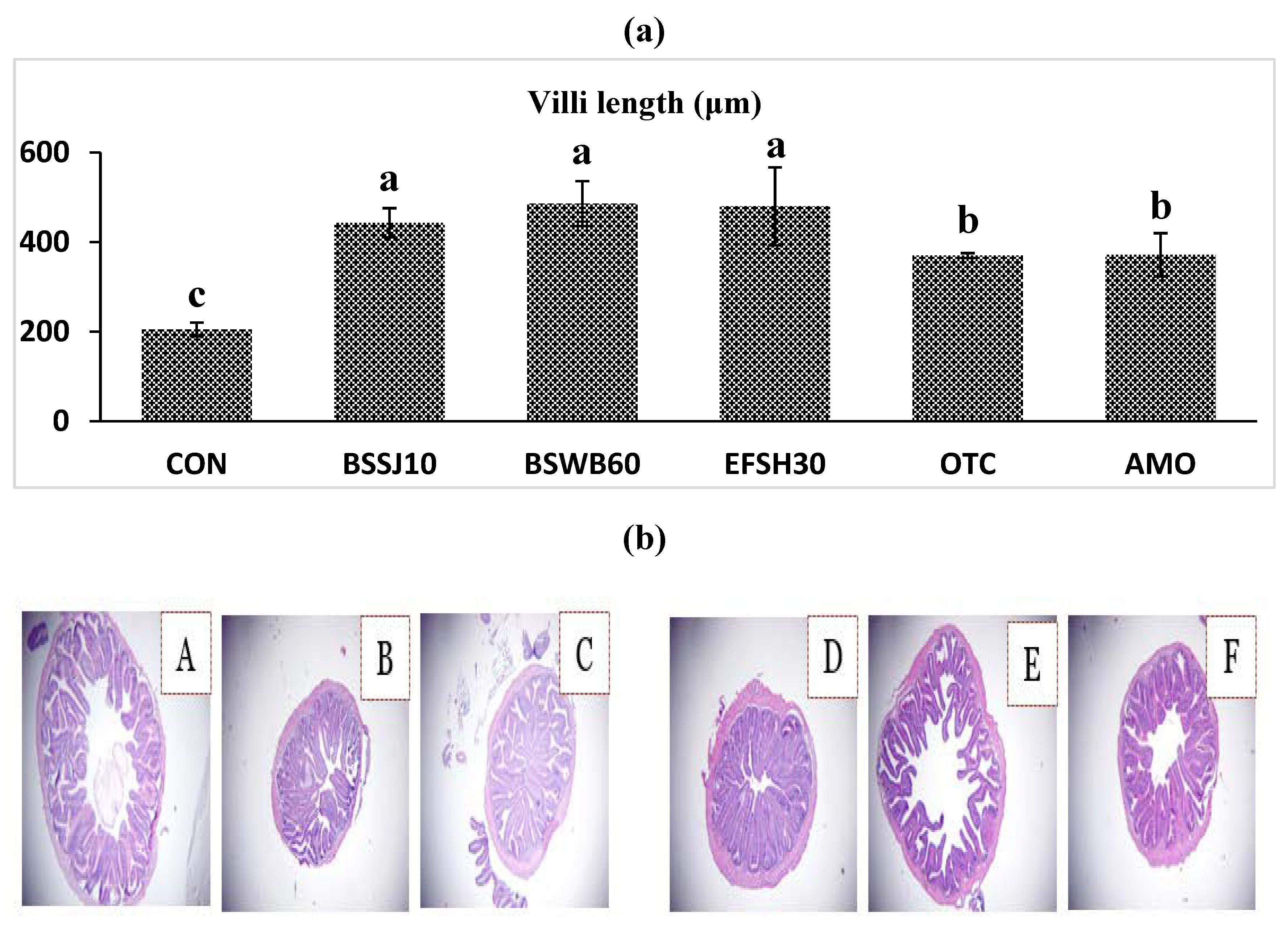
Figure 3.
Cumulative Survival (%) of olive flounder fed the experimental diets for 8 weeks. 1Values are means of triplicate samples from fish where the values in each line with different superscripts are significantly different (P<0.05).
Figure 3.
Cumulative Survival (%) of olive flounder fed the experimental diets for 8 weeks. 1Values are means of triplicate samples from fish where the values in each line with different superscripts are significantly different (P<0.05).
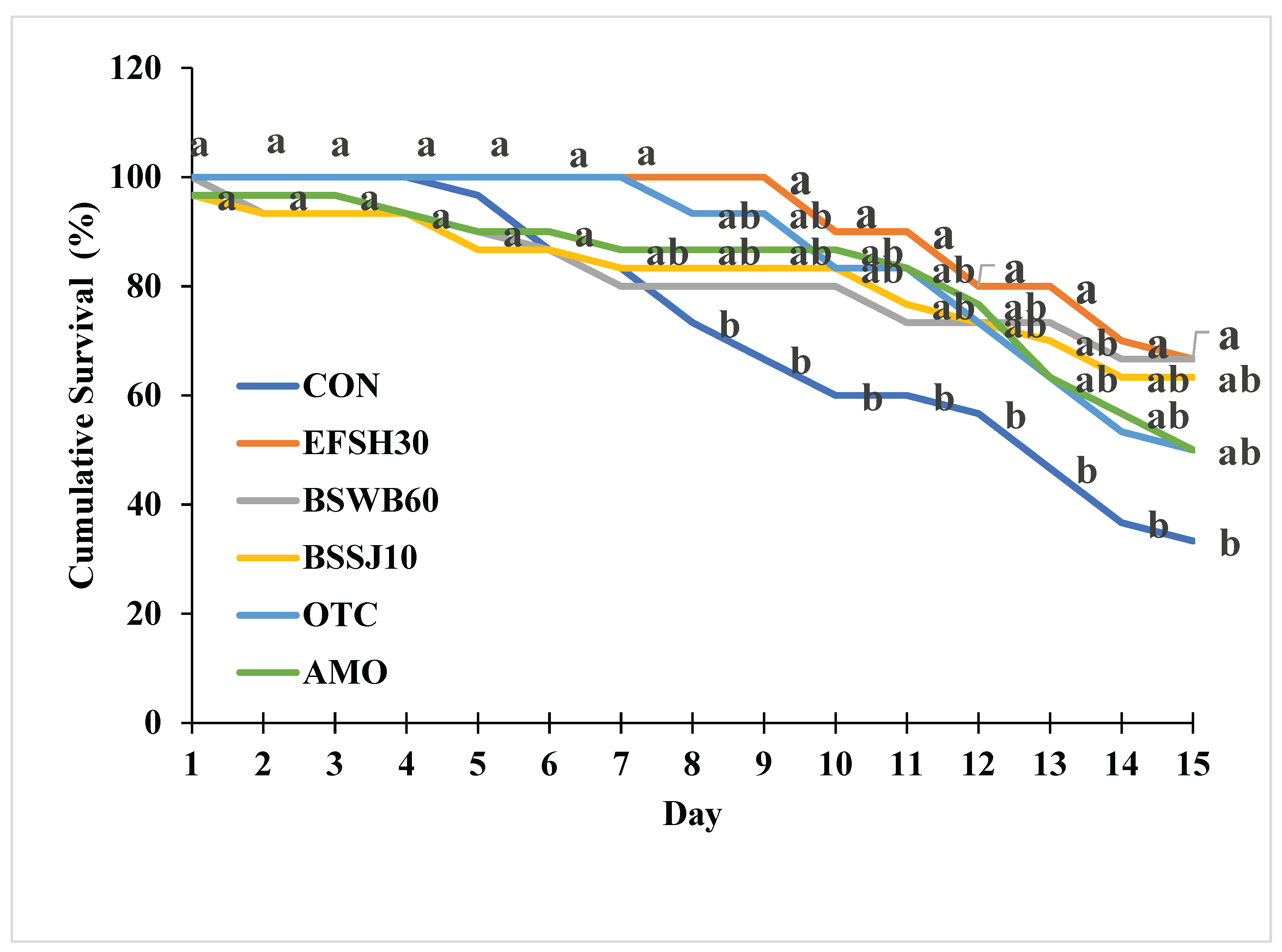

Table 1.
Composition of the basal diet (% of dry matter basis).
| Ingredients | % |
|---|---|
| Anchovy fish meal1 | 45 |
| Soybean meal | 12 |
| Starch2 | 3.8 |
| Wheat flour | 7.0 |
| Blood meal | 4.5 |
| Squid liver powder | 5.6 |
| Meat and bone meal | 8.0 |
| Poultry by product meal | 4.5 |
| Fish oil3 | 4.3 |
| Vitamin premix4 | 1.2 |
| Mineral premix5 | 1.2 |
| etc.6 | 3.0 |
| Proximate analysis (% of DM basis) | |
| Moisture | 8.56 |
| Crude protein | 56.2 |
| Crude lipid | 8.35 |
| Crude ash | 11.4 |
1Suhyup feed Co. Uiryeong, Korea. 2The feed Co. Goyang, Korea. 3Jeil feed Co. Hamman, Korea. 4Contains (as mg/kg in diets) : Ascorbic acid, 300; dl-Calcium pantothenate, 150; Choline bitate, 3000; Inositol, 150; Menadion, 6; Niacin, 150; Pyridoxine · HCl, 15; Rivoflavin, 30; Thiamine mononitrate, 15; dl-α-Tocopherol acetate, 201; Retinyl acetate, 6; Biotin, 1.5; Folic acid, 5.4; Cobalamin, 0.06. 5Contains (as mg/kg in diets) : NaCl, 437.4; MgSO4·7H2O, 1379.8; ZnSO4·7H2O, 226.4; Fe-Citrate, 299; MnSO4, 0.016; FeSO4, 0.0378; CuSO4, 0.00033; Ca(IO)3, 0.0006; MgO, 0.00135; NaSeO3, 0.00025. 6etc.: Calcium phosphate; Lecithin, Batain; Taurine; Choline; Vitamin C; Lysine; Methionine.
Table 2.
Proximate composition of the eight experimental diets (% of DM basis)1.
| Parameters | Diets | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | BSWB60 | BSSJ10 | EFSH30 | OTC | AMO | ||||||
| Moisture | 8.56 | 9.01 | 8.91 | 8.56 | 8.91 | 8.71 | |||||
| Crude Protein | 56.2 | 55.9 | 54.8 | 57.1 | 55.9 | 56.1 | |||||
| Crude Lipid | 8.35 | 8.29 | 8.34 | 8.23 | 811 | 8.32 | |||||
| Crude Ash | 11.4 | 10.9 | 11.2 | 10.9 | 11.2 | 11.8 | |||||
1 Values are means of triplicate samples. Values in each row without superscripts are non-significantly different (P>0.05).
Table 3.
Gene specific primers, amplicon lengths and gene bank accession numbers of immune and growth-related genes used in this study.
Table 3.
Gene specific primers, amplicon lengths and gene bank accession numbers of immune and growth-related genes used in this study.
| Name of gene | Sense | Oligonucleotide Sequence (5’ to 3’) | Base pair (bp) |
Gene bank accession number |
|---|---|---|---|---|
| β-actin | F | CAGCATCATGAAGTGTGACGTG | 107 | HQ386788.1 |
| R | CTTCTGCATACGGTCAGCAATG | |||
| FGH | F | CGCCGTATGGAAACTCTGAACT | 160 | M23439.1 |
| R | GGGTGCAGTTAGCTTCTGGAAA | |||
| IL-1β | F | ATGGAATCCAAGATGGAATGC | 250 | KF025662.1 |
| R | GAGACGAGCTTCTCTCACAC | |||
| IL-10 | F | AGCGAACGATGACCTAGACACG | 114 | KF025662.1 |
| R | ACCGTGCTCAGGTAGAAGTCCA |
1 FGH: flounder growth hormone. 2 IL-1β: interleukin-1beta. 3 IL-10: interleukin-10.
Table 4.
Growth performance and feed utilization of olive flounder fed the six experimental diets for 8 weeks1.
Table 4.
Growth performance and feed utilization of olive flounder fed the six experimental diets for 8 weeks1.
| Parameters | Diets | Pooled SEM |
|||||
|---|---|---|---|---|---|---|---|
| CON | BSWB60 | BSSJ10 | EFSH30 | OTC | AMO | ||
| IBW | 12.5ns | 12.4 | 12.2 | 12.4 | 12.4 | 12.4 | 0.04 |
| FBW | 36.7 b | 42.0a | 40.8ab | 40.7ab | 36.9b | 37.0b | 0.98 |
| WG (%) | 194b | 230a | 224ab | 229ab | 202b | 199b | 8.38 |
| SGR (%/day) | 1.95b | 2.24a | 2.16ab | 2.16ab | 2.01b | 1.99b | 0.05 |
| FE (%) | 106b | 118ab | 125a | 115ab | 111b | 110b | 2.79 |
| PER | 0.65b | 0.84a | 0.76ab | 0.76ab | 0.70ab | 0.66b | 0.03 |
| Survival (%) | 93.3ns | 90.7 | 89.3 | 89.3 | 96.0 | 98.7 | 5.57 |
| HSI (%) | 1.24ns | 1.66 | 1.29 | 1.31 | 1.13 | 1.12 | 0.08 |
| VSI (%) 1 | 2.05ns | 1.94 | 1.74 | 1.94 | 1.88 | 1.74 | 0.05 |
| CF | 0.92ns | 0.97 | 0.93 | 0.93 | 0.92 | 0.91 | 0.01 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different (P<0.05); ‘ns’ denotes non-significant differences (P>0.05). IBW: Initial body weight. FBW: Final body weight. WG: Weight gain (%) = [(final wt. - initial wt.) × 100] / initial wt. SGR: Specific growth rates (%) = [(loge final wt. - loge initial wt.) × 100] / days. FE: Feed efficiency (%) = (wet weight gain / dry feed intake) × 100. PER: Protein efficiency ratio = (wet weight gain / protein intake). Survival (%) = [(total fish – dead fish) × 100] / total fish. HSI: Hepatosomatic index = (liver wt. × 100) / body wt. VSI: Viscerosomatic index (%) = (viscera wt. × 100) / body wt. CF: Condition factors = (wet weight / total length3) × 100.
Table 5.
Whole-body proximate composition of olive flounder fed the experimental diets1.
| Parameters | Diets | Pooled SEM |
|||||
|---|---|---|---|---|---|---|---|
| CON | BSWB60 | BSSJ10 | EFSH30 | OTC | AMO | ||
| Moisture | 76.3ns | 75.5 | 75.1 | 75.8 | 76.8 | 76.2 | 0.25 |
| Protein | 20.3ns | 21.0 | 20.8 | 21.5 | 22.3 | 20.7 | 0.29 |
| Lipid | 2.42ns | 2.39 | 2.30 | 2.34 | 2.41 | 2.41 | 0.02 |
| Ash | 4.09ns | 4.25 | 4.21 | 4.16 | 4.14 | 4.19 | 0.02 |
1 Values are means from triplicate groups of fish where the values in each row without superscripts are non-significantly different (P>0.05); ‘ns’ denotes non-significant differences.
Table 6.
Hematological parameters of olive flounder fed the six experimental diets1.
| Parameters | Diets | Pooled SEM |
|||||
|---|---|---|---|---|---|---|---|
| CON | BSWB60 | BSSJ10 | EFSH30 | OTC | AMO | ||
| ALT | 4.67ns | 4.00 | 4.67 | 4.33 | 4.00 | 4.00 | 0.13 |
| AST | 16.3bc | 26.7a | 15.3c | 22.0ab | 20.0bc | 21.7ab | 1.69 |
| GLU | 3.1ns | 3.4 | 3.3 | 3.2 | 3.3 | 3.5 | 0.06 |
| TP | 13.3ns | 11.8 | 16.0 | 15.1 | 14.8 | 13.2 | 1.04 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different (P<0.05), where ‘ns’ denotes non-significant differences. ALT: Alanine aminotransferase (U/l), AST: Aspartate aminotransferase (U/l), GLU: Glucose (mg/dl), Total protein (g/dl).
Table 7.
Non-specific immune responses of olive flounder fed the six experimental diets1.
| Parameters | Diets | Pooled SEM |
|||||
|---|---|---|---|---|---|---|---|
| CON | BSWB60 | BSSJ10 | EFSH30 | OTC | AMO | ||
| Lysozyme2 | 0.22c | 0.69a | 0.65a | 0.67a | 0.30bc | 0.39b | 0.08 |
| MPO3 | 1.36b | 1.81a | 1.55ab | 1.56ab | 1.49ab | 1.45b | 0.06 |
1 Values are means from triplicate groups of fish where the values in each row with different superscripts are significantly different (P<0.05). 2 Lysozyme (U/ml): lysozyme activity. 3 MPO (absorbance): myeloperoxidase (OD at 450 nm).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



