Submitted:
09 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
Predators are dependent on the capture of prey to meet their energetic and nutritive requirements, imposing a regime of predation risk. Consequently, prey experience both consumptive and non-consumptive effects. Non-consumptive effects may manifest through altered growth and ontogenetic trajectories of prey species, a dynamic modulated by olfactory or other sensory cues from predators. Bactrocera dorsalis Hendel represents a great invasive threat to global horticulture. While earlier research has primarily centered on the consumptive interactions between B. dorsalis and its natural enemies, the potential consequences of non-consumptive interactions on the development of B. dorsalis have been overlooked. In this study, we investigated the impact of predation risk effects, induced by both visual exposure to the predatory mantis Hierodula patellifera Serville and its associated odor, on the life history traits of B. dorsalis. Female B. dorsalis demonstrated reduced development time in the presence of isolated H. patellifera or H. patellifera odor, yet evidenced significantly increased fecundity. Conversely, males displayed no significant change in development time. Additionally, neither female nor male body weight at death was significantly influenced by predation risk from isolated predators or predator odor. These results indicated adaptive developmental strategies of B. dorsalis when faced with risks from predators and highlight the important role of predator odor, which has the potential to lay the groundwork for improving biological control efforts and innovative approaches within pest control.
Keywords:
Bactrocera dorsalis Hendel
; non-lethal effects
; phenotypic plasticity
; predator-prey interaction
; pest management
Introduction
The interaction between predators and prey is a fundamental form of biological interaction [1,2]. Predators depend on prey to obtain energy and nutrients, while prey are subject to the pressure and control of predators [3]. The interaction between predators and prey can be divided into two main types: consumptive and non-consumptive effects [4,5]. Consumptive effects reduce prey density through lethal predation, while non-consumptive effects affect prey populations by influencing prey fitness [6,7,8,9,10]. Analyses have shown that non-consumptive effects of predators on prey may exceed the impact of consumptive effects on prey populations [3,11]. However, research on non-consumptive effects is limited, and there is even less research on non-consumptive effects in insects.
When organisms experience non-consumptive effects, they often exhibit a trade-off strategy [12,13]. Acquiring resources for development and avoiding predators constitutes an important trade-off, with organisms allocating energy accordingly [14,15]. In the presence of predators, prey often allocate more energy to defense mechanisms against predation [7]. For example, when detecting the presence of predation stress, potato beetle larvae (Leptinotarsa decemlineata) reduce feeding to avoid predation [16]. Female damselflies (Ischnura cervula) in water exhibit reduced feeding desire when they sense predator cues, allocating more resources to finding predator-free habitats [17].
Such defensive strategies often lead to shifts in growth rates and fecundity [18,19]. For example, butterfly larvae (Pararge aegeria) develop rapidly when facing predation risks because they allocate more energy resources to growth and development, despite the increased risk of immediate mortality [20]. Long-term exposure to predation threat, such as that of the six-spotted ladybug (Menochilus sexmaculatus), significantly reduces the lifespan of adult cotton aphids (Aphis gossypii) [21]. In the case of the herbivorous mite (Tyrophagus putrescentiae), individuals in control conditions exhibit a significantly longer lifespan than those subjected to predation risk, regardless of whether the stress occurs throughout their entire life cycle or only at specific stages [22]. The duration of predation risks also affects lifespan; adult Drosophila melanogaster experiences a significantly shorter lifespan under continuous predation risk from ladybugs (Propylea japonica) compared to those facing intermittent risk [23]. Predation risk often results in a decrease in prey fecundity; for instance, Sitobion miscanthi reduces its net reproduction rate in response to isolated predator presence [24]. Female Bactrocera tryoni lay fewer eggs in the presence of predator olfactory cues [25]. However, there are instances where predation risk has no significant effect on prey fecundity, as seen in Philetairus socius [26].
Prey can detect predator cues in the environment through vision, hearing, smell, and touch, using this information to adjust their behavior and avoid predation [27,28]. Visual features such as appearance, body shape, color, and movement patterns of predators help prey identify potential hazards and trigger avoidance behavior [29,30,31]. Additionally, specific vocalizations of predators serve as a means for various prey species to assess threatening situations [32]. However, prey face the disadvantage of requiring closer proximity to predators when using visual and auditory cues to judge predation risk, potentially leading to misjudgments, especially with ambush predators [33]. Prey animals themselves release or store characteristic chemical cues, which other animals detect and use to alter their behavior to reduce predation risk [34,35,36]. Accurate assessment of predation risk enables animals to exhibit effective anti-predator behaviors. For example, Bactrocera tryoni significantly reduces foraging, oviposition, and mating in the presence of predator volatiles [25]. The negative reaction of Leptinotarsa decemlineata to male volatile cues from predators (Podisus maculiventris) was particularly strong [16]. However, research on the effects of predator odor-mediated threats is currently limited
The mantis, Hierodula patellifera Serville, is widely distributed in East Asia and Southeast Asia. It is considered an efficient predator, known for its sit-and-wait hunting strategy, and serves as an important natural enemy of the B. dorsalis and other dipteran and lepidopteran pest in orchards [37,38]. Bactrocera dorsalis, commonly known as the oriental fruit fly, is a widespread fruit pest native to tropical and subtropical regions of Asia. Since its introduction to China, it has inflicted considerable damage on various crops and fruits, with its range steadily expanding [39,40]. The use of chemical control methods currently employed has detrimental effects on the environment and contributes to the development of pesticide resistance in B. dorsalis [41,42,43,44]. Biological control presents a promising alternative for managing B. dorsalis populations. However, a comprehensive understanding of the predation risk posed by potential natural predators is essential for optimizing the effectiveness of this approach.
The primary objective of this study was to investigate the predation risk effects of H. patellifera on the development and reproduction of B. dorsalis. Two distinct rearing conditions were established for B. dorsalis. In the first scenario, B. dorsalis was reared in an environment isolated from the predator H. patellifera, enabling detection of the predator’s presence through visual cues and odors. In the second scenario, B. dorsalis was exposed solely to the odors associated with H. patellifera, allowing detection of the predator exclusively through olfactory cues. These experimental setups enabled us to elucidate the mechanisms through which predator risk influences the developmental and reproductive traits of B. dorsalis.
Methods and Materials
Experimental Insects
Larval B. dorsalis utilized in this study were sourced from the Haidian campus of Hainan University in Haikou, China. Maintained under controlled conditions of 27±1°C temperature, 65% relative humidity, and a 16:8h (L: D) photoperiod, the larvae were nourished on a diet comprising paper, banana, corn flour, sugar, yeast powder, concentrated hydrochloric acid, sodium benzoate, and sterile water [45]. Adults were housed in a screened cage measuring 80×80×80cm, where they were supplied with an artificial diet consisting of sugar, yeast, honey, agar, and water, alongside access to sterile water, with fresh provisions replenished regularly. Female B. dorsalis were collected for fecundity using centrifuge tubes containing fresh orange juice [25] (Figure 1).
The experimental population of H. patellifera utilized in this study originated from a mixed clone, initiated by hatching eggs gathered from the field (Haidian campus of Hainan University, Haikou, China), and purchased from an online store (https://m.tb.cn/h.5IUR2hy?tk=DazqW8llIlO). In the laboratory, the breeding protocol for H. patellifera involved feeding first and second instar larvae with an artificial liquid diet composed of pure milk and honey, with intervals of 72 hours, and subsequently providing third instar to adult H. patellifera with a mixture of B. dorsalis, Bactrocera cucurbitae, and Zelugodocus tau at similar intervals, supplemented with sterile water
The Effect of Isolated-Predator on the Development and Fecundity of B. dorsalis
Adult B. dorsalis that emerged within a 24-hour period were selected for the study. For the isolation treatment, 40 B. dorsalis individuals were housed in a 20×20×20 cm insect rearing cage, maintaining a 1:1 male-to-female ratio, with daily provision of diet. Seven-instar H. patellifera (starved for 24 hours) were confined in a transparent box (15×10×5 cm) with a mesh window (10×5 cm) on the lid, positioned at the bottom of the rearing cage. This setup prevented direct physical contact or feeding interactions between H. patellifera and B. dorsalis, while allowing for the transmission of risk cues through visual and olfactory channels. Test mantises were replaced every 24 hours. The blank control group consisted of an empty box without H. patellifera. Each treatment was replicated five times, and daily mortality and the weight of deceased B. dorsalis individuals were recorded. On the 8th day of the experiment, an egg-collecting tube (Figure 1) containing 5 ml of fresh orange juice was introduced into the cage from 15:00 to 18:00, coinciding with the period of highest daily egg production for B. dorsalis. Eggs laid by the B. dorsalis were collected and counted daily until the demise of all female B. dorsalis.
The Effect of Predator Odor and Development and Fecundity of B. dorsalis
The H. patellifera were subjected to a 24-hour starvation period before odor collection. Each H. patellifera individual was then placed in a plastic box (19×12.5×7.5 cm, cleaned with 75% ethanol to remove any residual chemicals before introduction) with a moist tissue paper (13×9.5 cm, referred to as the ‘risk paper’) positioned at the bottom. After 24 hours of free movement, the H. patellifera were removed, and the ‘risk paper’ served as the predator odor. For control treatments, a clean moist tissue paper was used. The other materials and process were identical to the described procedure, with live mantises replaced by ‘risk paper’ in the risk treatments or a control paper in the control treatments. Test tissue papers were replaced every 24 hours. Each treatment was replicated five times, and daily mortality, weight, and the fecundity of deceased B. dorsalis were recorded as described previously.
Data Analysis
The normality and homogeneity of variance of the data were assessed using Shapiro-Wilk and Levene’s tests, respectively. Survival analysis was conducted to examine the development of B. dorsalis under both risk and control treatments, considering differences between sexes. A Cox proportional hazards model was employed to evaluate variations in survival times, while the log-rank test was utilized to compare survival curves. Fecundity analysis involved a linear mixed model (LMM) to assess the impacts of predation risk and time on B. dorsalis fecundity, with time designated as the random effect. Additionally, a linear mixed model was used to analyze the effects of predation risk, sexes, and time on the body weight of deceased B. dorsalis, with time of death considered as the random effect. Normality and variance homogeneity were confirmed using the qqPlot function from the ‘car’ package and the plot.lme function from the ‘nlme’ package, respectively. All statistical analyses were conducted in R (R Core Team, 2023).
Results
Development Time
For isolated predator treatments, the development time of adult B. dorsalis were significantly affected by predation risk (Table 1a). Female B. dorsalis exposed to isolated predator had a significant shorter development time than that of adult in control treatment (the median survivals in predation risk and control treatments were 21 and 27 respectively; Figure 2a). However, The development time of male were not affected (the median survivals in predation risk and control treatments were 22 and 20 respectively; Figure 2c). For predator odor treatment, the development time of adult B. dorsalis were significantly affected by predation risk (Table 1b). The development time of female is significantly short than that in the control treatment (the median survivals in predation risk and control treatments were 13.5 and 25 respectively; Figure 2b). And the development time of male is short than that in the control treatment, but no significance was found (the median survivals in predation risk and control treatments were 14 and 17.5 respectively; Figure 2d).
Fecundity
In isolated predator treatments, the fecundity of B. dorsalis was significantly influenced by predation risk, time, and their interaction (Table 2a). Females subjected to isolated predator treatment exhibited higher fecundity compared to those in the control treatment (Figure 3a). The predicted curve indicates a decrease in fecundity with increasing time. Similarly, in the presence of predator odor treatment, the fecundity of adult B. dorsalis was significantly affected by predation risk and time, although there was no significant interaction effect between predation risk and time on fecundity (Table 2b). Female B. dorsalis exposed to predator odor treatment displayed higher fecundity compared to adults in the control treatment, with the predicted curve also illustrating a decrease in fecundity over time (Figure 3b).
Body Weight at Death
For isolated predator treatments, although the sex effects on body weight at death were highly significant, there were no effects observed for risk treatments, sexes, or their interactions on body weight at death (Table 3a; Figure 4a,b). For predator odor treatments, significant effects were found in sexes and time, but no significant effects were found in risk treatments. Additionally, there were no significant effects of the interaction between treatments, time, and sex (Table 3b; Figure 4c,d).
Discussion
This study demonstrated that predator risk treatment significantly affected the development and fecundity of B. dorsalis, although it did not have a significant effect on body weight at death. B. dorsalis subjected to isolated predator and predator odor treatments exhibited shorter development times and higher fecundity. These findings suggested a potential correlation between predation risk imposed by H. patellifera and developmental changes in B. dorsalis. Moreover, they indicated that only predator odor could induce a predation risk effect on the development of B. dorsalis similar to that induced by a real predator.
Studies have shown that under predation risk, prey face a trade-off between responding to predation threats and lifespan [46,47]. For instance, as predation pressure increases, the development time of the larvae of Tyrophagus putrescentiae extends, while the adult lifespan shortens [48]. In our study, we observed that the development time of B. dorsalis decreased following exposure to isolated predator risk. Similarly, exposure to predator odors also reduced the development time of B. dorsalis compared to the control, suggesting that when faced with predation risks, B. dorsalis might allocate more resources and energy to defense rather than to development, consequently altering their development time. This finding is consistent with previous research where, under the risk of predation by bats, Spodoptera litura exhibited a reduced lifespan [49].
Furthermore, we discovered that the impact of predation risk on prey B. dorsalis development time was gender-specific; female B. dorsalis treated with isolated predator had a shorter development time than those in the control treatment, while this was not observed in males. This disparity was also found in the phytophagous mite (Tyrophagus putrescentiae), where regardless of being subjected to predator stress throughout the life cycle or at different life stages, the lifespan of males was significantly longer than that of females [22]. The variation in development time responses between females and males could be attributed to the fact that females bear the responsibility for fecundity and need to allocate most of their resources to reproduction, leaving less for development under predation risks [48,50,51].
A trade-off between predation risks and reproduction was also indicated by a bulk of studies [25,47,48]. For example, predation-stressed phytophagous mites (Tyrophagus putrescentiae) reduce fecundity and allocate resources to anti-predation [22], which is contrary to our observations. The fact that fecundity increased under exposure to predation risks (both isolated predator and predator odor treatments) was observed. The difference may be due to differences in prey species, predator type, hunting model, and the strength of the predation risks, and prey may adjust their reproduction accordingly [52,53].
However, the development time of female B. dorsalis treated with predation risk decreased, but the fecundity was higher than that of the control treatment. A similar situation has also been observed in some studies, where, when threatened by long-term predation, the lifespan of aphid Rhopalosiphum padi will decrease, but the fecundity rate will increase [54]. The increased number of offspring may allow the individual that is eaten by a predator to be compensated [55]. This suggests that in our studies, the resources or energy diverted from development by female B. dorsalis may be allocated to fecundity or anti-predation, or to both of them. As the increased number of offspring may result in a satiety effect of the predator [56], allowing enough individuals surviving to establish a new population.
Our study showed that there was no significant difference in body weight at death between the control and isolated predator or predator odor treatments. This differs from the results of previous studies. Under predation stress, the development of Helicoverpa armigera was accelerated, and the adult size became larger [57]. Studies also indicated that prey tend to become larger when facing predation risk [58]. It is still unknown why body weight is not affected in our present results, however, we did observe hesitation of B. dorsalis to approach and feed on the food in the experimental cage under predation risk. As food is an important factor influencing body weight, this behavior should reduce its body weight. A probable reason is that the stressed B. dorsalis may increase the quantity and/or quality of its food (for example, eat more or increase food conversion rate), thus suffering fewer effects on body weight.
To conclude, this study showed that predation risk from the predatory mantis H. patellifera could shorten the developmental time of B. dorsalis, but it could increase fecundity without changing biomass. We highlight the role of predation risk in shaping the developmental traits when B. dorsalis is faced with a decision between anti-predation and development, which represents an important trade-off within organisms in nature. Furthermore, we observed that only predator odor could induce a risk response comparable to that of an actual predator, suggesting a potential for novel and more accessible pest control methods utilizing predator odors instead of real predators in the field. However, it is crucial to identify the chemical nature of the predator odor before implementing such strategies.
References
- Barbosa, P.; Lgnacio, C. Ecology of predator-prey interactions. Oxford University Press. 2005.
- Abrams, P. The evolution of predator-prey interactions: Theory and evidence. Annual Review of Ecology and Systematics. 2000, 31, 79–105. [Google Scholar] [CrossRef]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to death? The effects of intimidation and consumption in predator-prey interactions. Ecology. 2005, 86, 501–509. [Google Scholar] [CrossRef]
- Orrock, J.L.; Grabowski, J.H.; Pantel, J.H.; Peacor, S.D.; Peckarsky, B.L.; Sih, A.; Werner, E.E. Consumptive and nonconsumptive effects of predators on metacommunities of competing prey. Ecology. 2008, 89, 2426–35. [Google Scholar] [CrossRef]
- Ritchie, E.G.; Johnson, C.N. Predator interactions, mesopredator release and biodiversity conservation. Ecology Letters. 2009, 12, 982–998. [Google Scholar] [CrossRef] [PubMed]
- Hawlena, D.; Schmitz, O.J. Herbivore physiological response to predation risk and implications for ecosystem nutrient dynamics. Proc Natl Acad Sci U S A. 2010, 107, 15503–15507. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, C.; Lohroff, T.; Biefel, F.; Cocherell, D.E.; Carson, E.W.; Hung, T.C.; Connon, R.E.; Fangue, N.A.; Todgham, A.E. Effects of turbidity, temperature and predation cue on the stress response of juvenile delta smelt. Conserv Physiol. 2023, 11, coad036. [Google Scholar] [CrossRef]
- Nicieza, A.G. Interacting effects of predation risk and food availability on larval anuran behaviour and development. Oecologia. 2000, 123, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Culshaw-Maurer, M.; Sih, A.; Rosenheim, J.A. Bugs scaring bugs: Enemy-risk effects in biological control systems. Ecology Letters. 2020, 23, 1693–1714. [Google Scholar] [CrossRef]
- Weed, A.S.; Frank, J.H. Oviposition behavior of pheropsophus aequinoctialis L. (coleoptera: Carabidae): A natural enemy of scapteriscus mole crickets (orthoptera: Gryllotalpidae). Journal of Insect Behavior. 2005, 18, 707–723. [Google Scholar] [CrossRef]
- Cresswell, W. Non-lethal effects of predation in birds. Ibis. 2008, 150, 3–17. [Google Scholar] [CrossRef]
- Johnson, E.C.; Braco, J.T.; Whitmill, M.A. Connecting nutrient sensing and the endocrine control of metabolic allocation in insects. Insect Science. 2014, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Worm, B.; Karez, R. Competition, coexistence and diversity on rocky shores. In Competition and coexistence, Ecological Studies. Springer, 2002; 133–163. [Google Scholar]
- Werner, E.E. Individual behavior and higher-order species interactions. The American Naturalist. 1992, 140, S5–S32. [Google Scholar] [CrossRef]
- Werner, E.E.; Anholt, B.R. Ecological consequences of the trade-off between growth and mortality rates mediated by foraging activity. The American Naturalist. 1993, 142, 242–272. [Google Scholar] [CrossRef] [PubMed]
- Hermann, S.L.; Thaler, J.S. Prey perception of predation risk: Volatile chemical cues mediate non-consumptive effects of a predator on a herbivorous insect. Oecologia. 2014, 176, 669–676. [Google Scholar] [CrossRef]
- Siepielski, A.M.; Fallon, E.; Boersma, K. Predator olfactory cues generate a foraging-predation trade-off through prey apprehension. Royal Society. 2016. [Google Scholar] [CrossRef] [PubMed]
- Koch, N.; Lynch, B.; Rochette, R. Trade-off between mating and predation risk in the marine snail, littorina plena. Invertebrate Biology. 2007, 126, 257–267. [Google Scholar] [CrossRef]
- Urban, M.C. The growth-predation risk trade-off under a growing gape-limited predation threat. Ecology. 2007, 88, 2587–97. [Google Scholar] [CrossRef] [PubMed]
- Gotthard, K. Increased risk of predation as a cost of high growth rate: An experimental test in a butterfly. Journal of Animal Ecology. 2000, 69, 896–902. [Google Scholar] [CrossRef]
- Lin, X.M.; Cui, X.X.; Tang, J.H.; Zhu, J.W.; Li, J.H. Predation risk effects of lady beetle menochilus sexmaculatus (fabricius) on the melon aphid, aphis gossypii glover. insects. 2024, 15, 13. [Google Scholar] [CrossRef]
- Wei, X.; Liu, J.; Zhang, Z.-Q. Predation stress experienced as immature mites extends their lifespan. Biogerontology. 2023, 24, 67–79. [Google Scholar] [CrossRef]
- Li, Y.P.; Ge. F. Effect of prey stress from Propylea japonica on development and fecundity of Drosophila melanogaster in successive three generations. Entomological Knowledge. 2010, 47, 139–145. [Google Scholar]
- Wang, L.; Atlihan, R.; Chai, R.; Dong, Y.; Luo, C.; Hu, Z. Assessment of non-consumptive predation risk of coccinella septempunctata (coleoptera: Coccinellidae) on the population growth of sitobion miscanthi (hemiptera: Aphididae). insects. 2022, 13, 524. [Google Scholar] [CrossRef]
- Kempraj, V.; Park, S.J.; Taylor, P.W. Forewarned is forearmed: Queensland fruit flies detect olfactory cues from predators and respond with predator-specific behaviour. Scientific Reports. 2020, 10, 7297. [Google Scholar] [CrossRef]
- Fortuna, R.; Covas, R.; D'Amelio, P.B.; Silva, L.R.; Parenteau, C.; Bliard, L.; Rybak, F.; Doutrelant, C.; Paquet, M. Interplay of cooperative breeding and predation risk on egg allocation and reproductive output. Behavioral Ecology. 2024, 35. [Google Scholar] [CrossRef] [PubMed]
- Kral, K. Visually guided search behavior during walking in insects with different habitat utilization strategies. Journal of Insect Behavior. 2019, 32, 290–305. [Google Scholar] [CrossRef]
- Zanuzzo, F.S.; de, C.B.A.L.; Pereira, R.T.; Valença-Silva, G.; Barcellos, L.J.G.; Barreto, R.E. Innate response based on visual cues of sympatric and allopatric predators in nile tilapia. Behavioural Processes. 2019, 109–114. [Google Scholar] [CrossRef]
- Kats, L.B.; Dill, L.M. The scent of death: Chemosensory assessment of predation risk by prey animals. Écoscience. 1998, 5, 361–394. [Google Scholar] [CrossRef]
- Grubb, T.C., Jr. Antipredator defenses in birds and mammals. The Auk 2006, 123, 601–605. [Google Scholar] [CrossRef]
- Munoz, N.E.; Blumstein, D.T. Multisensory perception in uncertain environments. Behavioral Ecology. 2012, 23, 457–462. [Google Scholar] [CrossRef]
- Hettena, A.M.; Munoz, N.; Blumstein, D.T. Prey responses to predator's sounds: A review and empirical study. Ethology. 2014, 120, 427–452. [Google Scholar] [CrossRef]
- Brown, G.E.; Magnavacca, G. Predator inspection behaviour in a characin fish: An interaction between chemical and visual information? Ethology. 2003, 109, 739–750. [Google Scholar] [CrossRef]
- Binz, H.; Bucher, R.; Entling, M.H.; Menzel, F. Knowing the risk: Crickets distinguish between spider predators of different size and commonness. Ethology. 2014, 120, 99–110. [Google Scholar] [CrossRef]
- Schmitz, O.A.O. Predator and prey functional traits: Understanding the adaptive machinery driving predator-prey interactions. F1000Researc, 2017; 6, 1767. [Google Scholar]
- Poulin, R.X.; Lavoie, S.; Siegel, K.; Gaul, D.A.; Weissburg, M.J.; Kubanek, J. Chemical encoding of risk perception and predator detection among estuarine invertebrates. Proc Natl Acad Sci U S A. 2018, 115, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Wu, J.; Zhao, D.X. Research progress on Hierodula patellifera serville. Journal of Southern Agriculture. 2014; 45, 53–57. [Google Scholar]
- Wang, S.J.; Wu, J.; Zhao, Y.A.; Li, R.X.; Zhao, D.X. Functional response of adult Hierodula patellifera (Serville, 1839) (Mantodea: Mantidae) to Tessaratoma papillosa (Drury) (Hemiptera:Tessaratomidae). International Journal of Tropical Insect Science. 2020, 4, 1053–1058. [Google Scholar] [CrossRef]
- Lin, J.T.; Zeng, L.; Lu, Y.Y.; Liang, G.W.; Xu, Y.J. Research Advances in biology and control of Bactrocera (Bactrocera) dorsalis (Hendel). Joumal of ZhongKai Agrotechnical College. 2004, 60–67. [Google Scholar]
- Huang, J.F.; Zhang, Y.J. Research progress of oriental fruit fly Bactrocera dorsalis (Hendel) (Diptera: Tetriphitidae). Deciduous Fruits, 2023; 55, 68-71+3. [Google Scholar]
- Jin, Y.X.; Zhang, D.M.; Xie, C.F.; Li, M.M.; Meng, L.L.; Shang, M.Q.; Zhou, H.-X. Research advance on green prevention and control technology of Bactrocera dorsalis H. Plant Quarantine, 2022; 36, 1–6. [Google Scholar]
- Zhu, Y.F.; Shang, M.Q.; Teng. Z.W.; Tan, X.M; et al. Analysis of Invasion, Distribution and Spreding Trend of Bactrocera dorsalis. Shandong Agricultiral Sciences. 2020, 52, 141–149. [Google Scholar]
- Zhu, X.S.; Liu, Y.; Dai, S.Z.; Luo, R.; Jia, H.S.; He, P.; Zhao, L. The control experiment of three kinds of potion on bactrocera dorsalis in apple orchard. Yunnan Agricultural Science and Technology. 2021, 03, 9–10. [Google Scholar]
- Quan, J.C.; Chen, G.F.; Jiang, Y.H. Damage Investigation and Field Control Test of Bactrocera dorsalis in Guangxi. South China Fruits. 2019, 48, 86–91. [Google Scholar]
- Yuan, R.L.; Zheng, C.W.; Feng, F.D. Study on new feeding method of bactrocera dorsalis. Agricultural Technolofg Service. 2020, 37, 27–30. [Google Scholar]
- Duong, T.M.; McCauley, S.J. Predation risk increases immune response in a larval dragonfly (leucorrhinia intacta). Ecology. 2016, 97, 1605–1610. [Google Scholar] [CrossRef]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction-immunity trade-offs in insects. Annu Rev Entomol. 2016, 61, 239–56. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Z.Q. Level-dependent effects of predation stress on prey development, lifespan and reproduction in mites. Biogerontology. 2022, 23, 515–527. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Wang, Z.; Lin, T.; Feng, J.; Jiang, T. Effects of predation risks of bats on the growth, development, reproduction, and hormone levels of spodoptera litura. Front. Ecol. Evol. 2023, 11. [Google Scholar] [CrossRef]
- Li, G.Y.; Zhang, Z.Q. Development, lifespan and reproduction of spider mites exposed to predator-0induced stress across generations. Biogerontology. 2019, 20, 871–882. [Google Scholar] [CrossRef]
- Chandrasegaran, K.; Kandregula, S.R.; Quader, S.; Juliano, S.A. Context-dependent interactive effects of non-lethal predation on larvae impact adult longevity and body composition. PLoS ONE. 2018, 13, e0192104. [Google Scholar] [CrossRef] [PubMed]
- Segev, O.; Verster, R.; Weldon, C. Testing the link between perceived and actual risk of predation: Mosquito oviposition site selection and egg predation by native and introduced fish. J Appl Ecol. 2017, 54, 854–861. [Google Scholar] [CrossRef]
- Dumont, F.; Lucas, É.; Alomar, O. Oviposition behavior of the mirid Macrolophus pygmaeus under risk of intraguild predation and cannibalism. Insect Science. 2021, 28, 224–230. [Google Scholar] [CrossRef]
- Ninkovic, V.; Feng, Y.; Olsson, U.L.; Pettersson, J. Ladybird footprints induce aphid avoidance behavior. Biological Control. 2013, 65, 63–71. [Google Scholar] [CrossRef]
- Chamberlain, J.D.; Clifton, I.T.; Gifford, M.E. Influence of prey size on reproduction among populations of Diamond-backed Watersnakes (Nerodia rhombifer). Canadian Journal of Zoology. 2017, 95, 929–935. [Google Scholar] [CrossRef]
- Mills, N.J. Satiation and the functional response: A test of a new model. Ecological Entomology. 1982, 7, 305–315. [Google Scholar] [CrossRef]
- Xiong, X.; Michaud, J.P.; Li, Z.; Wu, P.; Chu, Y.; Zhang, Q.; Liu, X. Chronic, predator-induced stress alters development and reproductive performance of the cotton bollworm, helicoverpa armigera. BioControl. 2015, 60, 827–837. [Google Scholar] [CrossRef]
- Mikolajewski, D.J; Brodin, T.; Johansson, F.; Joop, G. ; Phenotypic plasticity in gender specifc life-history: Efects of food availability and predation. Oikos. 2005, 110, 91–100. [Google Scholar] [CrossRef]
Figure 1.
Egg-collecting tube (a) made of 50ml centrifuge tube. B. dorsalis deposits their eggs directly into the egg-collecting tube (b).
Figure 1.
Egg-collecting tube (a) made of 50ml centrifuge tube. B. dorsalis deposits their eggs directly into the egg-collecting tube (b).

Figure 2.
The lifespan of female (a and b) and male (c and d) B. dorsalis when exposed to an isolated predator (a and c) and predator odor (b and d), and curves were fitted using log-rank test.
Figure 2.
The lifespan of female (a and b) and male (c and d) B. dorsalis when exposed to an isolated predator (a and c) and predator odor (b and d), and curves were fitted using log-rank test.
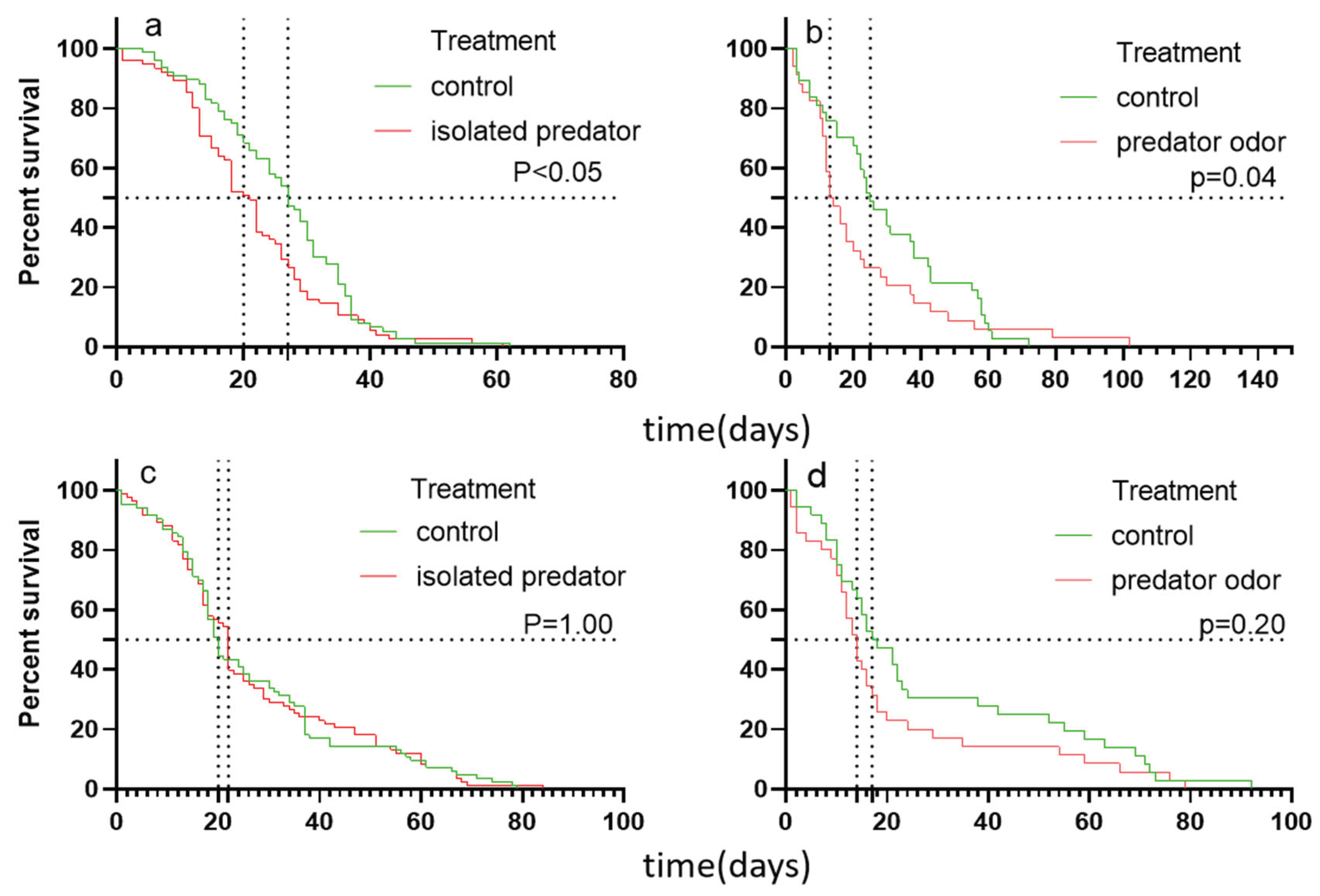
Figure 3.
The fecundity of B. dorsalis when exposed to an isolated predator (a) and predator odor (b), and predicted curves were fitted using mixed models.
Figure 3.
The fecundity of B. dorsalis when exposed to an isolated predator (a) and predator odor (b), and predicted curves were fitted using mixed models.
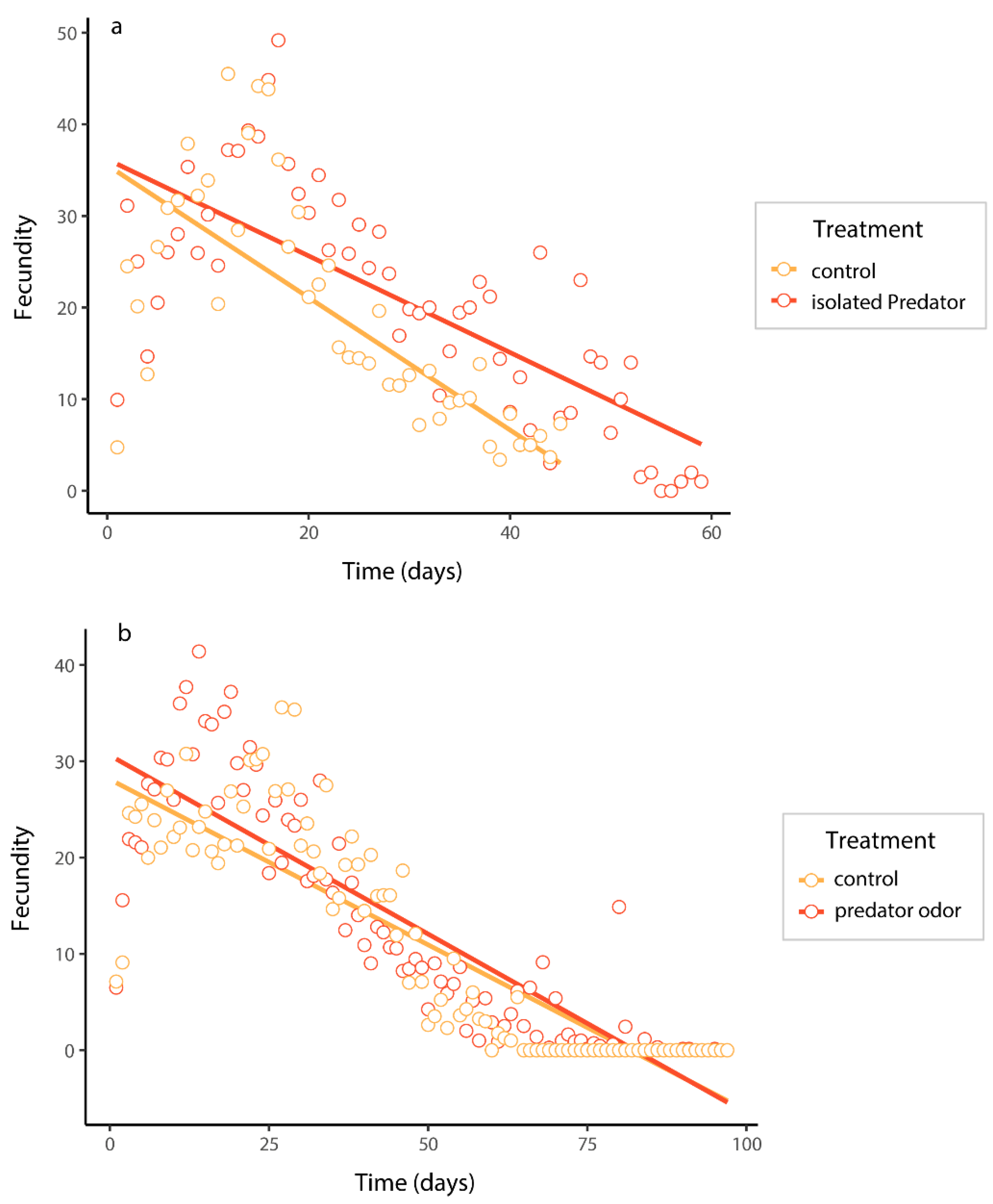
Figure 4.
The body weight at death of female (a and c) and male (b and d) B. dorsalis when exposed to an isolated predator (a and b) and predator odor (c and d), and predicted curves were fitted using linear mixed models.
Figure 4.
The body weight at death of female (a and c) and male (b and d) B. dorsalis when exposed to an isolated predator (a and b) and predator odor (c and d), and predicted curves were fitted using linear mixed models.
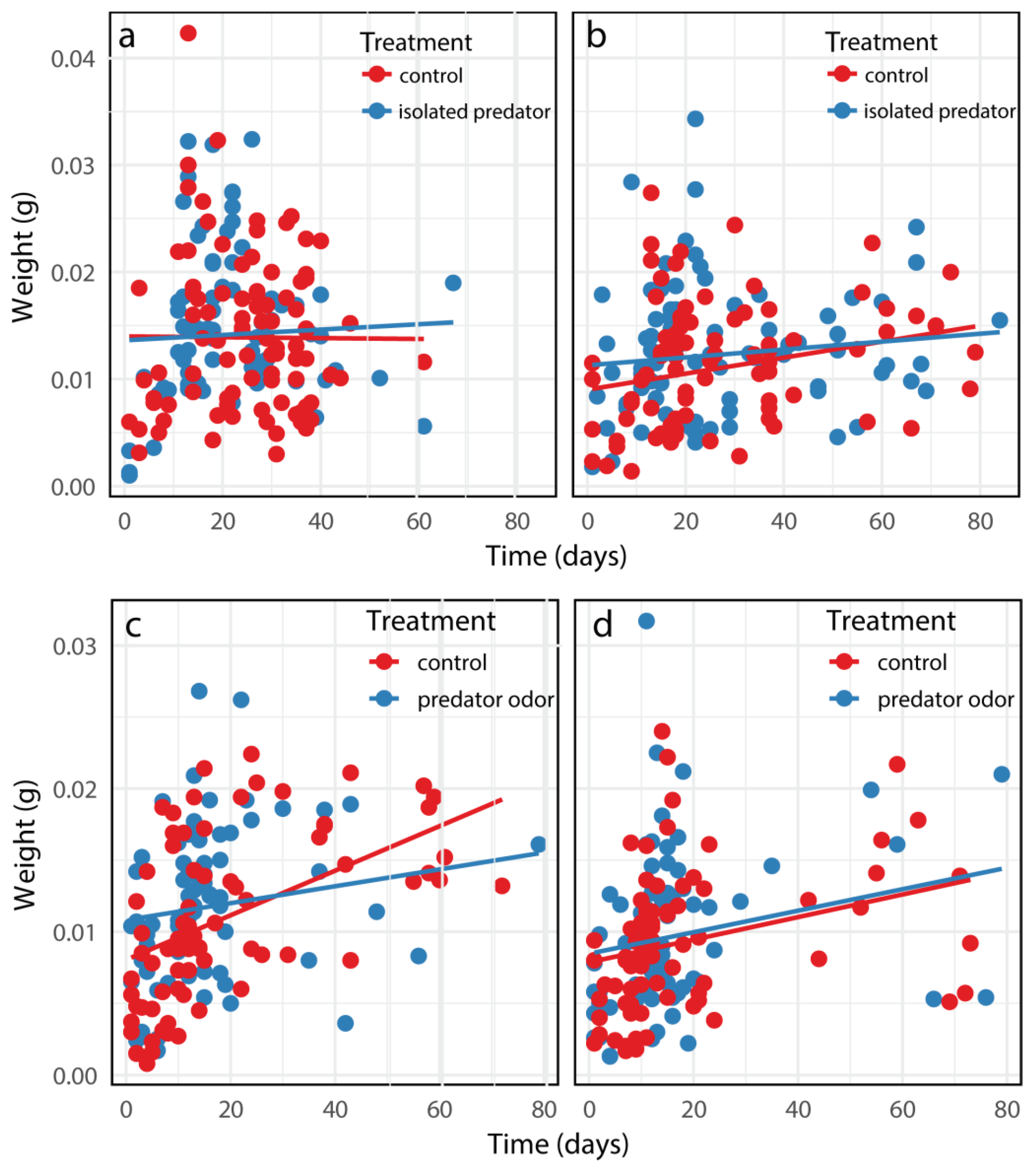

Table 1.
The COX proportional hazards model examined the effects of predation risk treatments, sexes, and their interaction on the survival of B. dorsalis under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
Table 1.
The COX proportional hazards model examined the effects of predation risk treatments, sexes, and their interaction on the survival of B. dorsalis under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
| Response variables | Predictor variables | Coef | Exp (coef ) | Se (coef) | Z | Pr (>|z|) |
| Development time (isolated predator experiments) | treatments | 0.349 | 1.417 | 0.164 | 2.132 | 0.033 |
| sexes | -0.107 | 0.898 | 0.163 | -0.656 | 0.512 | |
| Treatments: sexes | -0.346 | 0.708 | 0.226 | -1.529 | 0.126 | |
| Development time break//(predator odor experiments) | treatments | 0.395 | 1.485 | 0.172 | 2.304 | 0.021 |
| sexes | -0.025 | 0.975 | 0.172 | -0.145 | 0.885 |
Table 2.
The ANOVA analysis examined the effects of predation risk treatments, fecundity time, and their interaction on the fecundity of B. dorsalis under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
Table 2.
The ANOVA analysis examined the effects of predation risk treatments, fecundity time, and their interaction on the fecundity of B. dorsalis under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
| Response variables | Predictor variables | df | chisq | Pr (>F) |
| Fecundity (isolated predator experiments) | treatment | 1 | 33.10 | <0.001 |
| time | 1 | 83.48 | <0.001 | |
| treatment: time | 1 | 8.33 | 0.004 | |
| Fecundity (predator odor experiments) | treatment | 1 | 3.95 | <0.001 |
| time | 1 | 359.95 | 0.047 | |
| treatment: time | 1 | 1.76 | 0.183 |
Table 3.
The linear mixed model was used to analyze the impact of predation risk, sexes and time on the body weight of B. dorsalis at death under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
Table 3.
The linear mixed model was used to analyze the impact of predation risk, sexes and time on the body weight of B. dorsalis at death under isolated predator (a) and predator odor experiments (b). The bold value is the significant value.
| Response variables | Predictor variables | df | chisq | Pr (>F) |
| (a) Body weight at death (isolated predator experiments) | time | 1 | 2.599 | 0.107 |
| treatment | 1 | 1.214 | 0.271 | |
| sex | 1 | 13.437 | <0.001 | |
| time:treatment | 1 | 0.171 | 0.679 | |
| time:sex | 1 | 0.818 | 0.366 | |
| treatment:sex | 1 | 0.487 | 0.485 | |
| time:treatment:sex | 1 | 0.471 | 0.493 | |
| (b) Body weight at death (predator odor experiments) | time | 1 | 19.753 | <0.001 |
| treatment | 1 | 2.012 | 0.156 | |
| sex | 1 | 9.781 | 0.002 | |
| time:treatment | 1 | 1.440 | 0.230 | |
| time:sex | 1 | 1.313 | 0.252 | |
| treatment:sex | 1 | 0.280 | 0.597 | |
| time:treatment:sex | 1 | 1.574 | 0.210 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



