Submitted:
05 April 2024
Posted:
08 April 2024
You are already at the latest version
Abstract
The maintenance of traditional agricultural practices in protected small islands is closely linked to the preservation of the land, the protection of natural resources, and the conservation of local traditions. For this purpose, the Tuscan Archipelago National Park promoted the census of ancient horticultural varieties preserved by some farmers on the island of Elba. The survey revealed a deep-rooted historical, cultural, social, and economic link between this germplasm and the island’s territory. A scientific characterization of the landrace under ethnobotanical, morphological, and genetic aspects was carried out. Among these local varieties, two landraces of the Brassica species and three landraces of the Phaseolus species were identified and analyzed in this study. To ascertain their distinguishability and uniqueness, similar varieties were also identified based on morpho-physiological characteristics or territorial proximity and compared with the Elban landraces. Morpho-phenological data were collected, and molecular analyses were carried out using Simple Sequence Repeats (SSR) markers. Ten different SSR loci were considered for the Phaseolus species and nine for the Brassica species. The Tuscan Archipelago landraces showed a high level of internal variability but were clearly distinct from each other and from the analyzed varieties, as shown by AMOVA (Analysis of Molecular Variance). Their distinctness was further highlighted by the UPGMA dendrogram obtained from the genetic distance matrix. The same results were confirmed by Principal Coordinate Analysis (PCoA). This study confirms the peculiarity of the Elban landraces, their biodiversity is a heritage to be safeguarded, which can also be used in future breeding programs. These ancient Elban landraces were allocated to local farmers (most were young farmers) to protect them from extinction and to promote more resilient varieties cultivation compared to commercial hybrids widely spread in the area. Local farmhouses also play a strategic role in safeguarding the germplasm and dissemination of sustainable agricultural practices. There is high demand for ancient Elban landraces, this has led to the development of a new local economy balanced between agricultural tradition and innovation.
Keywords:
landrace
; local germplasm
; on-farm conservation
; Biosphere Reserve
; custodian farmers
; SSR markers
; population structure
1. Introduction
The central role of agriculture and farmers in biodiversity conservation is recognized by the UN 2030 Agenda, and the European Biodiversity Strategy, implemented in Italy with the National Biodiversity Strategy (2023). This document sets out the lines of action to ensure the integration between the country's development objectives and biodiversity protection. In Italy, the expansion of sustainable agricultural practices is a priority of the 'National Strategy for a Sustainable and Inclusive Agricultural, Food and Forestry System'. This document aims to promote the conservation and enhancement of plant genetic resources important for agriculture and food, with a particular focus on landraces threatened by erosion or extinction [1].
Landraces have traditionally been cultivated on small plots since at least the 1950s, have been naturally selected by the environment and farmers, and are characterised by the fact that they retain more genetic variability over time than commercial varieties [2]. In addition, landraces have generally been developed with reduced agronomic inputs and under more sustainable growing conditions than commercial varieties [3]. The Elba landraces are perfectly adapted to their environment and are clearly identified and usually referred to by a local name. Elba’s local varieties have therefore not been selected through 'formal' genetic improvement but are characterised by a specific adaptation to the environmental conditions of a well-defined area of cultivation, such as the islands and islets of the Tuscan archipelago. Landraces generally have characteristics that make them tolerant to the biotic and abiotic stresses of these regions. They are closely connected with the customs, knowledge, habits, dialects and traditional practices of the local population involved in their cultivation and conservation [4].
It should also be considered that the richness of agricultural biodiversity (i.e. genetic traits of the species) is a relevant component in the ability of landraces to adapt to abiotic stresses such as drought and high temperatures [5]. This can increase crop resilience and improve the response to the challenges faced by agricultural production systems affected by climate change [6]. However, the use of landraces is currently limited, with only a few traditional cultivars of noteworthy reputation being preserved over time, while many others have been gradually abandoned [7].
Genetic erosion, defined as 'the loss over time of genetic diversity between and within populations or landraces of the same species, or the reduction of the genetic base of a species due to human intervention or climate change', is due to many factors: mechanisation and modern farming techniques, socio-economic changes and a lack of interest among younger generations in preserving seeds on farms have also led to an ineffective transmission of knowledge between generations, resulting in a loss of historical memory and, consequently, to a reduction in the number of traditional local varieties cultivated. [8].
Some Italian regions, including Tuscany, the first in Italy, have taken regulatory action against genetic erosion to protect and safeguard agricultural biodiversity through national and European legislation, creating the "Regional Directory of Local Genetic Resources of Tuscany" [9]. The procedures for registering landraces in the regional repertoire have been defined, the figure of the Tuscany Region's custodian farmer has been established and germplasm banks have been activated within the framework of the Conservation and Security Network [10 G. Bellet, et al 2019]. The identification, collection, characterisation and conservation of local genetic resources are therefore of particular importance, especially in the field of landraces, for some of which Italy has historically been very rich in variability. [7]. In this context, the establishment of Protected Areas contributes to the conservation of habitats and biodiversity and to the preservation of agroecosystems and landscapes. In these zones, it is possible to reconcile the need to preserve the natural environment with sustainable forms of economic development, promoting a model of agriculture in balance with the natural environment [11].
The Tuscan Archipelago National Park includes the area of the seven islands off the coast of Tuscany, a territory that contains a great biological diversity, representative of the Mediterranean region. It should also be noted that this area has been acknowledged by UNESCO as a MAB (Man and Biosphere Programme) Biosphere Reserve, due to the long-term preservation of natural resources and the ongoing promotion of sustainable economic development to benefit local communities [12].
Figure 1.
Map of the island of Elba.
In this specific area, traditional agricultural practices have been kept alive, allowing local farmers to select landraces, particularly suited to the specific conditions of the island environment. These landraces have been integrated into the context of traditional local cuisine, especially on Elba island, the largest and the widest agricultural area of the Tuscan islands [13].
Several fruit and vegetable landraces with potential value for the conservation of native germplasm and genetic improvement were identified among elderly farmers on the island of Elba. The risk of genetic erosion of these landraces proved to be quite high from the start of the research. At the time, there was only one Tuscan custodian farmer on the island of Elba, and no landraces from the island were listed in ‘Regional Directory of Endangered Local Varieties’. Faced with this situation, efforts were made to census, characterise, and promote the value of these landraces to safeguard the agrobiodiversity of the islands.
Furthermore, the horticultural varieties under study were characterized from an ethnobotanical, morphological, and genetic point of view to obtain registration in the Regional Directory of Local Genetic Resources of Tuscany and the National Register of Agricultural and Food Biodiversity.
Genetic characterization was carried out with SSR (Simple Sequence Repeats) or microsatellite markers [14]. These molecular markers are widely used in varietal description studies due to their characteristics such as good polymorphism, high repeatability of data obtained, specificity to the amplified locus, and codominance [15].
The joint use of morpho-physiological characterisation and genetic profiling has permitted us to obtain an accurate description of the landraces, to know the level of intra- and inter-varietal variability, and to explore possible genetic relationships between similar varieties. Furthermore, the definition of the typical genetic profiles of each landrace has allowed them to be identified in order to protect and conserve their biodiversity.
2. Materials and Methods
2.1. Preliminary Investigation and Identification of Ancient Varieties
Historical bibliographic research was conducted to uncover details about the agricultural history of the Elba island region and the varieties previously grown there, this process helped determine the specific areas to focus on. Subsequent surveys were carried out to gather information on antique plant varieties, including fruits, grains, and vegetables, and to physically locate and identify them. Multiple sources were consulted for this task, including farmers, farm managers, farmhouse operators, and hobbyist farmers with knowledge of agriculture on Elba island. Samples of the ancient Elban crop varieties were obtained from these sources, then reproduced and multiplied avoiding cross-contamination.
2.2. Plant Material
In particular, the present study concerns one landrace of Brassica oleracea L., named Patresi black cabbage, and three landraces of Phaseolus vulgaris L. Miracolo scritto, Miracolo non scritto, and Jolly Campesi.
In addition, a wild species of Brassica oleracea subsp. Robertiana (Gay) Rouy et Fouc synonym Brassica montana Pourret (Palmaiola cabbage) found on the islet of Palmaiola in front of Elba island in the Piombino Channel has been considered. This wild cabbage has been present on the island for a long time (1800) and may have food potential, as well as being an interesting source of variability for genetic improvement.
2.3. Morphological and Agronomic Analyses
Field trials were conducted in various locations on Elba island to analyze the morphology and agronomy of bean and cabbage local varieties. The morphology of Phaseolus varieties was described using the technical protocol CPVO-TP/012/3, while the UPOV TG/90/6 guidelines were used for the Brassica variety. The experimental plots were set up according to the local agricultural traditions for planting distances and types of cultivation.
The trials were expertly managed through a collaborative effort between farmers and researchers. The farmers were entrusted with the agronomic management of the plots, while the researchers conducted meticulous observations, sampling, and surveys on the various replications, adhering to UPOV and CPVO guidelines.
2.3.1. Beans
The bean landraces were sown in open fields at the end of April. For the Jolly Campesi variety, three seeds were placed per posthole, with a spacing of approximately 50cm between them in the row and 60cm between rows. A drip irrigation system was installed.
The Miracolo beans (scritti and non scritti) were sown again with three seeds per posthole, with a spacing of about 50cm between them in the row and 100cm between rows to enable the supports to be positioned. The Elban tradition of farming involves using the capannello technique (as shown in the Figure S1).
This technique requires one cane per posthole and three canes tied to the top. The CPVO-TP/012/3 protocol was used to assess the characteristics of the plants and pods.
Measurements were taken on fresh pods, including length (excluding the beak), width at the widest point, transversal width, length of the beak, weight of 20 fresh pods, and number of seeds per pod. The weight of 100 seeds refers to dry seeds.
Sampling was carried out on 20 plants grown in three different farms located in two different places on Elba island (Loc. La Lecciola in the Municipality of Rio and Loc. Schiopparello in the Municipality of Portoferraio).
2.3.2. Cabbages
Patresi black cabbage was planted in open fields in early September with rows spaced 100cm apart and 50cm apart. Regular irrigation was provided during the first month to promote germination and growth in the initial phase, due to low rainfall.
Subsequently, irrigation was provided as needed based on the seasonal rainfall pattern. Sampling was conducted on 20 plants from two farms and a hobbyist farmer in three different locations on Elba (Loc. La Lecciola in the Municipality of Rio, Loc. Schiopparello in the Municipality of Portoferraio, and Loc. Patresi in the Municipality of Marciana). The plant's characteristics were evaluated following the guidelines specified in UPOV TG/90/6.
Fifteen individuals from a wild population of Palmaiola cabbage were measured on Palmaiola island, located within the “A” integral protection zone of the Tuscan Archipelago National Park (as per the Park Plan approved by the Tuscan Regional Council Deliberation no.87 on 23/12/2009).
All surveys and sampling were conducted during a single visit in late spring, revealing individuals of various ages on the island, including young, mature, and senescent plants simultaneously within the population. The younger individuals were in the vegetative stage, while the more mature ones were at the flowering, fruiting, or ripening stage. This allowed for the collection of mature siliques and seeds for potential future replication.
2.4. Molecular Analysis
In addition to the Elban landraces currently in the process of characterization and conservation in the Tuscan Archipelago, other commercial varieties with similar morpho-physiological characteristics or geographical proximity have been identified and compared with the Elban local varieties to determine their specificity and uniqueness.
Five Brassica oleracea L. accessions were analysed, including two traditional varieties from the Tuscan Archipelago and three other varieties.
For the species Phaseolus vulgaris L., three landraces were analysed, consisting of seven accessions from the island of Elba and nine commercial varieties grown in the area.
The following tables (Table 1.) list the varieties included in this study.
Genomic DNA was extracted from approximately 200 mg of young seedlings obtained from seeds 10 days after germination in a controlled growth chamber. Seven individual samples and a pool of twenty individuals were analysed for all cabbage accessions except Palmaiola. In this case, due to the expected high variability, thirty individual plant leaf samples collected on the Palmaiola island were analysed.
For the bean accessions, three individual samples and a pool of twenty individuals were analysed. DNA extraction was carried out using the C-TAB method [18,19].
Molecular analyses were carried out using simple sequence repeat (SSR) markers selected based on their degree of polymorphism and genomic position. Ten SSR loci for the species Phaseolus [20,21,22] and nine SSR loci for the species Brassica [23] were considered (Tables S1 and S2).
PCR reactions were carried out as previously described in a relevant bibliography [24,25], using primers labelled with an M13 tail.
The amplification products were separated by capillary electrophoresis using an ABI PRISM 3500 Genetic Analyzer (Applied Biosystems, Waltham, MA, USA) and the molecular profiles obtained were recorded using Gene Mapper 5.0 software (Applied Biosystems).
2.5. Data Analysis
2.5.1. Morfological Analysis
Morphological measurements were taken on fresh pods and dry seeds from twenty plants of bean varieties. The mean and standard deviation of pod length (excluding beak), maximum width, transversal width, beak length, weight of 20 fresh pods, number of seeds per pod, and weight of 100 dry seeds were calculated for each variety.
Cabbage median fresh leaves taken from twenty plants were sampled. For both landraces, mean and standard deviation were calculated for the characters, plant diameter, leaf blade length, and width.
2.5.2. Genetic Analysis
The discriminatory power of each SSR marker was determined by calculating the Polymorphic Index Content (PIC) value [26]. The genetic variability at each locus in the accessions analysed was assessed by measuring several parameters [27]. These included the number of observed alleles per locus (Na), the effective number of alleles (Ne), the presence of rare and private alleles (allele frequency < 0.05), and the levels of expected heterozygosity (He) and observed heterozygosity (Ho) and the Inbreeding coefficient (f).
Analysis of Molecular Variance (AMOVA) [28] was performed to assess intra- and inter-varietal genetic variability. The Excel Microsatellites Toolkit version 3.1 [29,30], PowerMarker v3.25 [31] and GenAlEX [32] were used for these analyses.
A UPGMA dendrogram using the PHYLIP package v3.695 [33] was constructed to visualise the genetic relationships among the accessions studied, based on the genetic distances between individuals calculated using the ASM Allele Share Matrix.
Furthermore, a multivariate Principal Coordinate Analysis (PCoA) [34] was implemented to visualise the genetic relationships between individuals and varieties. Genetic variability is presented in a two-dimensional graph.
3. Results
3.1. Morpho-Physiological Analyses
3.1.1. Family: Fabaceae, Genus: Phaseolus, Species: vulgaris L.
Landrace: Jolly Campesi
The typical production area for this variety is Elba island, and it's believed to have originated in the municipality of Campo nell‘Elba. This variety has successfully adapted to the island's climatic conditions, which are characterized by a semi-arid climate [35]. It exhibits robust growth, high disease resistance, enhanced tolerance to drought, and a longer growing season compared to commercial varieties (Figure S2.). This variety's general agronomic needs align with traditional practices for cultivating green beans.
The Jolly Campesi bean has epigeous seedlings, adult plants with a dwarf habit, tufted and of medium height (50cm). The leaf is compound, trilobed, medium green, medium rugosity, terminal leaflet shape from circular to quadrangular and a medium acuminate apex. Flowering time is between June and July, the white flower is in foliage. The pod, green, of medium length (14cm). It is characterized by the absence of stringiness on the ventral suture and absent or very weak constrictions (Figure S3.). The weight of 20 fresh pods is 87g with 7/8 seeds per pod. The seeds with an elliptical median longitudinal section and a circular median cross-section are white (Figure S4.) with a weight of 100 dry seeds around 22/25g.
3.1.2. Family: Fabaceae, Genus: Phaseolus, Species: vulgaris L.
Landrace: Miracolo scritto, Miracolo non scritto
The landraces are widespread throughout the island. These varieties have high vigour and good disease tolerance. In the environmental conditions of Elba, two production cycles can be carried out with sowing in spring and summer.
The bean Miracolo Scritto e non Scritto share common morphological characteristics in their vegetative development. They are climbing beans, and they have epigeous seedlings. The leaves are medium green with medium rugosity. The terminal leaflet shape is from triangular to circular and it has a medium acuminate apex. The flowering time differs between the two varieties, being medium in the case of the bean Miracolo Scritto and medium/early for the bean Miracolo non Scritto.
The flower in both cases presents a double colouring with a white standard and violet wing colour. As far as the pod is concerned, the two varieties present very similar characteristics, while they differ in shape and characteristics of the seed. In the case of the bean Miracolo Scritto, the seed is medium-sized (100 seed weight 55g) with an elliptical median longitudinal section, beige with brown veins, and a different coloured hilum to the seed (Figures S5 and S6).
The bean Miracolo non Scritto on the other hand, has a small seed (100 seed weight 35-40g) that is white without veins (Figure S7), with a white hilum and an elliptical/rectangular median longitudinal section. In the case of the bean Miracolo non Scritto, there is an additional accession with beige seeds.
3.1.3. Family: Brassicaceae, Genus: Brassica, Species: oleracea L.
Landrace: Patresi black cabbage
The traditional cultivation area of the Patresi black cabbage is in the municipality of Marciana on Elba island but is expanding to other Elban municipalities. It is not found in other parts of Italy. The growing cycle spans from planting in September to flowering in late spring. The leaves are harvested during the winter. In Elba's ethnobotanical tradition, the leaves are usually used for preparing soups; it is currently employed in the preparation of 'pesto' manufactured by a canning company in the area.
The plant has a flattened shape and medium height, with green leaves of medium intensity and absent anthocyanin coloration. The leaf blade of average length and width is elliptical in shape with weak midrib curvature (Figure S8). In the middle part of the plant, the leaves have very weak or absent density of “curling” with semi-erect or horizontal petioles of average length (Figure S9). The yellow flower is gathered in racemose inflorescences.
3.1.4. Family: Brassicaceae, Genus: Brassica, Species: oleracea L. subsp. Robertiana (Gay) Rouy et Fouc sinonimo Brassica montana Pourret
Landrace: Palmaiola cabbage
The variety is typical of the island of Palmaiola, where, however, cultivation was abandoned fifty years ago, and it therefore occurs in the wild. Its area of distribution is limited to the island of Palmaiola. There is no knowledge of the presence of the same variety in other parts of Italy. Its location on the island of Palmaiola, which has been uninhabited for over fifty years and is denied to visitors, suggests the uniqueness of this variety.
The plant is flattened and of medium height, with a diameter of approx. 17mm. The leaf, which is partially anthocyanin-colored (Figure S10), is blue-green in the juvenile stage, turning to medium-green when fully developed. The elliptical leaf blade has an average length of about 15.5cm and an average width of about 11.4cm. It has a weakly curved midrib and, a short horizontal petiole of medium width. The flowers are yellow, gathered in a racemose inflorescence.
The wild population of Palmaiola has individuals at different vegetative stages simultaneously, and flowering is staggered from April to June, depending on the weather.
3.2. Genetic Analysis Results
3.2.1. Bean
The amplification of ten microsatellite loci for the genetic description of the Elban varieties and their relative comparison with the reference varieties yielded 38 different alleles, ranging from a minimum of two alleles found for the markers DROUGH1, AY1 and PVM118 to a maximum of 7 alleles for the markers PVM115 and PVM127, with an average of 3.4 alleles per locus. The effective number of alleles (Ne) ranges from 4.87 (PVM115) to 1.14 (PVM118). The average value of PIC is 0.5, five SSRs have values above 0.5. The major allele frequencies were lowest in PVM115 (MAF=0.35) and highest in PVM118 (MAF=0.93). The mean observed heterozygosity (Ho) is 0.92. Six out of ten alleles showed no heterozygosity and the inbreeding coefficient varied between 0.23 and 1. The parameters described are reported in the Table S3.
Two of the thirteen studied varieties were genetically homogeneous (He=0) and did not show any heterozygous individuals (Ho=0). The remaining varieties displayed similar values of observed heterozygosity. Private alleles were not identified for the Elban landraces, but rare alleles were present in all analyzed varieties (Table S5). The Analysis of Molecular Variance (AMOVA) calculation, based on the genetic distance matrix and 999 permutations, indicates that there is greater variability among accessions (95% of total variability) than within accessions (5%). These data are reported in Table S7. Four out of the ten studied SSRs showed variability within accessions.
The dendrogram obtained from the matrix of genetic distances, based on the frequency of shared alleles, using the UPGMA method shows that all three Elban varieties are distinct from each other and from the reference varieties (Figure 2). Both Elban varieties and three reference varieties displayed internal heterogeneity. However, all individuals belonging to the same variety were grouped into a single cluster, confirming the lower significance of within-variety variability compared to that among varieties, as indicated by the AMOVA analysis.
The two accessions of the “Miracolo non scritto” with white seeds can be distinguished from each other by only one out of the ten analyzed SSR loci. The selection of the “Miracolo non scritto” with beige seeds (2016 selection) differs from the white-seed accession, despite their close genetic relationship. The two accessions, beige and white, are also morphologically similar, with the only difference being the colour of the seed. Both accessions cluster together, indicating inherent variability within the variety.
The two accessions of the Jolly Campesi variety analysed share a single genetic profile that represents the variety.
The two 'Miracolo scritto' accessions have identical genetic profiles. All the Elban varieties are clearly distinguishable from each other and from the varieties studied for similarity and geographical proximity.
To further illustrate the relationship between the analyzed varieties, a Principal Coordinates Analysis (PCoA) was conducted (Figure 3). The first two principal components, with an Eigen value greater than 1, account for 59% of the total variability of the 10 SSR markers.
This two-dimensional representation confirms the distinguishability of the landraces identified in the cluster analysis. It also highlights the similarity of individuals belonging to the same landrace, even in cases of heterogeneity.
The graph shows two main groups based on growth type. The first group (blue circle) consists of dwarf bean varieties with determinate growth. The second group (red circle) includes all climbing bean varieties with indeterminate growth. Although determinate in growth, the Cocco Bianco variety is more like indeterminate growth the climbing varieties.
3.2.2. Cabbage
The genetic description of the Patresi black cabbage, Palmaiola cabbage, and reference varieties was made possible by amplifying nine microsatellite loci.
The analysis revealed a high level of genetic variability among the five examined varieties, with a total of 35 different alleles detected. The SSR Bo1652 had the lowest number of observed alleles (2), while Bo0836 had the highest (8), with an average of 3.9 alleles per locus. The effective number of alleles ranged from 1.4 (Bo1652) to 5.6 (Bo0836). Seven out of nine alleles had a PIC above 0.5, resulting in an average PIC value of 0.54. Bo1740 had the lowest major allele frequency (MAF=0.38), while Bo1652 had the highest (MAF=0.81). The observed heterozygosity (Ho) has an average value of 0.092, while the inbreeding coefficient ranges from 0.27 to 0.93. The parameters described are reported in the Table S4.
All studied varieties exhibited positive values of observed heterozygosity (Ho), with the highest values observed in the Patresi and Palmaiola landrace cabbages. A total of eleven private alleles were identified among the five varieties, with one for the Patresi Cabbage variety and six for the Palmaiola Cabbage variety. In contrast, no rare alleles were present in all analyzed varieties (Table S6).
The Analysis of Molecular Variance (AMOVA), calculated from the matrix of genetic distances with 999 permutations, indicates that 55% of the total variability is due to differences among varieties, while 45% is due to differences among individuals within the same variety (Table S8). It was observed that all varieties showed internal heterogeneity. In addition, the data are strongly influenced by the larger sample size used for the Palmaiola cabbage population. This is due to the high level of genetic variation observed among individuals in this group.
The genetic analysis allowed us to determine whether the Patresi and Palmaiola landrace are distinguishable from the reference varieties. The UPGMA dendrogram (Figure 4) shows that both varieties have a distinguishable molecular profile and group into distinct clusters, clearly differentiated from those of the Nero Pesciatino, Mercato di Copenhagen 2, and Scartoccio varieties. This confirms the relatively lower significance of intra-variety variability compared to inter-variety variability, as highlighted by the AMOVA analysis.
One individual from the Scartoccio variety was considered an off type because it is genetically distant from the profiles obtained from the other representative individuals of the variety. Each variety studied is easily distinguishable from the others, forming distinct clusters (highlighted by different colored boxes).
To further illustrate the relationship between the analyzed varieties, a Principal Coordinates Analysis (PCoA) was conducted Figure 5). The first two principal components (with an Eigen value greater than 1) describe 40.2% of the total variability.
In the two-dimensional PCoA plot, the distribution of the individual points highlights the high level of genetic variability. This further confirms the distinguishability of the Patresi and Palmaiola cabbages from the other three varieties analysed.
3.3. Registering in the Regional Directory
The bean varieties and the landrace Patresi have been entered in the Regional Directory of Local Genetic Resources of Tuscany.
To proceed with the enrolment, it was necessary to submit a specific application, which was evaluated by the technical-scientific Commission in charge of expressing the opinion for the enrolment in the Directory and in the National Register (LR 64/2004 Protection and valorization of the heritage of local livestock and varieties of agricultural, zootechnical and forestry interest).
The Palmaiola cabbage variety has not yet been entered into the Tuscany Region's germplasm register and is at risk of genetic erosion.
4. Discussion
The characterisation of Elba's traditional varieties of cabbages and beans is based on the analysis of different factors: historical, to understand their relationship with the territory, morphological, physiological, and agronomic, to improve their use and commercialisation, and genetic, to assess their specific and unique characteristics. The presence of these vegetable landraces, which are typical of the island of Elba, is attested by the old farmers from time immemorial, even if their cultivation has been largely abandoned in the past. The origin of these varieties is linked to the history of the island, which, from the Medici to the Napoleonic period, has seen the introduction of many species and varieties of fruit and vegetables [36]. In addition, in the last century the island was affected by migratory flows of workers from southern Italy, Tuscany and Emilia (the so-called Lombards), employed in mining, agriculture and prison management. These workers brought with them new varieties of fruit and vegetables from their places of origin, enriching the island's varietal landscape, which then underwent further selection and adaptation until the present day [37,38]. Isolation from the mainland has also contributed to selective forces linked to the island's environmental conditions and human pressure.
The three varieties of beans, "Miracolo scritto Elba," "Miracolo non scritto bianco Elba," and "Jolly Campesi" (Phaseolus vulgaris L.), are mainly grown in the eastern and central regions of Elba.
The enhancement of the agronomic and qualitative characteristics of these landraces achieved through this characterization study has contributed to expanding the cultivation area. Currently, production is spread across various locations on the island, with cultivation areas in the process of expansion. These varieties are not cultivated in other parts of Italy.
The key features of these bean landraces are their strong vigour and low susceptibility to diseases. Throughout the field trials conducted during the study, no phytosanitary treatments were applied, and no diseases were observed on the specimens, demonstrating their vigour and disease resistance/tolerance. All landraces are well adapted to the Elba climatic conditions, allowing for longer production compared to commercial varieties. The three varieties have stringless pods. Green pods are used, and for the "miracolo" beans, both fresh and dry seeds are utilized.
Genetic analysis confirmed the distinctiveness of all three bean landraces compared to very similar or locally present varieties. The bean landraces showed different levels of intra-varietal variability, as shown in the UPGMA dendrogram. Both the Jolly Campesi and Miracolo scritto varieties were genetically homogeneous within themselves, suggesting a rigorous breeding effort by farmers that led to the selection of these two varieties.
Conversely, the Miracolo non scritto bean showed the presence of three different accessions, similar but clearly distinguishable. There are two genetically closely related white-seeded accessions and one genetically distinct beige-seeded selection. All three accessions fall under the same cluster in the dendrogram, highlighting an intra-varietal variability characteristic of local conservation varieties [39,40,41,42].
The research carried out in the entire territory of Elba identified the Patresi black cabbage (Brassica oleracea L.), widely used in the culinary tradition of the Elba people. It is cultivated on land terraces in the western part of the island in the municipality of Marciana [43]. This delicately flavoured variety is regularly consumed in the area and is currently also used for pesto production by a canning company. This production is expected to promote its reintroduction throughout the island of Elba, significantly reducing the risk of genetic erosion.
Genetically, the variety proved to be distinct, albeit heterogeneous. In fact, a unique profile was identified for each analysed individual. This intra-varietal richness is typical of the species, as reported in other studies [44,45]. In the case of both Patresi and Palmaiola, the inter-varietal variability is even greater, which makes them unique compared to the varieties considered. This information allows us to classify them as local landraces with a rich biodiversity heritage.
The cultivation of Palmaiola cabbage is documented in the first half of the 19th century [46,47,48]. Already at that time there were botanical recognition and identification studies [49,50], showing a great interest in the species. After the automation of the lighthouse in 1974, the island remained uninhabited and the last report of a cultivated garden dates to the testimony of the daughter of one of the last lighthouse keepers [48]. Recently, the presence of Palmaiola cabbage has been confirmed by park guides and residents of the island of Elba, who have seen populations of this variety close to the sea, even without landing on the island.
The difference between naturalized populations of B. oleracea L. and B. montana is mainly determined by leaf colour. B. oleracea L. has greyish-green leaves, while B. montana has slightly greyish leaves that can range from pure green to bluish. This distinction is primarily based on leaf colour, according to Snogerup et al. [51]. The morphological data comparison between Palmaiola cabbage (bluish green) and Patresi black cabbage (green) cultivated on the island of Elba, revealed differences in the colour of young leaves and notable drought resistance of Palmaiola cabbage. These differences led to their classification as B. montana for Palmaiola cabbage and B. oleracea L. for Patresi cabbage (Figures S8–S11).
The population on Palmaiola island does not receive irrigation. Despite low rainfall and high population density in a small area, the vegetation is lush, there is abundant flowering, and significant seed production.
To cultivate the variety, it is sown in a seedbed at the beginning of September and then transplanted into rows spaced 100cm apart with a 50cm row spacing. The leaves are harvested during winter, and it flowers at the end of spring. B. Montana has potential agronomic value due to its drought and insect tolerance, as well as its high glucosinolate content [52]. Further study and preservation of the variety is warranted based on these characteristics [53,54].
The Palmaiola cabbage displays significant genetic diversity among its populations. We analyzed 30 individuals and obtained 22 molecular profiles. The molecular markers have identified high variability, indicating a broad genetic background within this landrace. Therefore, it is crucial to conserve this population to maintain its original diversity. Safeguarding this valuable germplasm should be an urgent priority. Currently, Palmaiola cabbage is not included in the Tuscany region's germplasm catalogue and is at risk of genetic erosion.
5. Conclusions
The aim of this study was to characterise ethnobotanically, morphologically, and genetically three bean and two cabbage varieties of ancient, endangered vegetable plant varieties of the Elba island. These landraces have been preserved over time by local farmers who have safeguarded them from extinction. The research has collected testimonies and information on endangered agricultural biodiversity, promoting the conservation of sustainable agricultural practices and developing cultural value for the territory. This has given voice to the cultural tradition of the islands.
The use of SSR molecular markers has proved useful both in confirming the distinctiveness of these landraces in relation to the closest varieties and in assessing the level of intra-varietal variability. This information is currently being used to define appropriate seed conservation and multiplication strategies and, in the future, could be a tool for verifying these procedures to maintain the authentic landraces over time. In addition, the molecular profiles can be used, to confirm the traceability and authenticity of even processed products. The results obtained from the morphological characterisation and genetic analysis have shown the presence of high genetic diversity within among the populations, confirming the uniqueness of these landraces. This has led to the inclusion of the three beans and one cabbage (Patresi) studied, in the Tuscan Regional Catalogue and the Italian National Register database, officially recognising their importance as local genetic resources. The Palmaiola cabbage also has very interesting agronomic and genetic characteristics and therefore needs to be studied in more detail with a view to its conservation and valorisation.
Researchers and farmers collaborated productively in managing experimental plots during the project. This collaboration has increased the number of custodian farmers from one to six. The people involved, farmer and researcher, have established a network committed to promoting and spreading awareness about Elban varieties to safeguard agrobiodiversity and preserve these unique varieties. This will preserve them from disappearance through in situ/on-farm conservation [43,44,45,46,47,48,49,50,51,52,53,54,55]. In addition to the older farmers in the area, who represent a historical memory, some young people have shown an interest in experimenting in their home area and in the potential for economic diversification of their activities and production.
The custodian farmers are actively collaborating with the Park Authority to provide a future perspective on the valorisation and protection of the agricultural biodiversity of the territory. The annually collected seeds of these landraces are stored ex situ at the germplasm bank of the Tuscan Regional Earth Authority. This process secures the local genetic resources of agricultural and food interest on the island of Elba.
The region's significant allure to tourists, combined with the cooperation between the National Park and agricultural businesses, has facilitated the promotion of landraces and best agricultural practices through guided tours and educational workshops. The positive impact of these efforts has extended beyond the island's borders, helping to promote an agricultural model based on short supply chains, traditional products and environmental sustainability.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Ten microsatellites used for molecular characterisation in P. vulgaris landrace. Table S2. Nine microsatellites used for molecular characterisation in Brassica landrace. Table S3. Genetic diversity parameters from the 10 SSR loci used for the analysis of the P. vulgaris collection from the island of Elba. Table S4. Genetic diversity parameters from the 9 SSR loci used for the analysis of the Brassica collection from the island of Elba. Table S5. Genetic diversity parameters for the P. vulgaris landrace. Table S6. Genetic diversity parameters for the Brassica landrace. Table S7. Analysis of molecular variance among and within landraces of the P. vulgaris landrace. Table S8. Analysis of molecular variance among and within landraces of the B. oleracea landrace. Figure S1. Capannello: Elba's traditional farming technique. Figure S2. The image displays the Jolly Campesi landrace on the left and a commercial variety on the right. The Jolly Campesi landrace is more vigorous, even late in the season. Its production cycle is significantly longer than that of the commercial variety. Figure S3. Jolly Campesi pod. It is green, of medium length (14cm) and characterized by the absence of stringiness on the ventral suture and absent or very weak constrictions. Figure S4. Campesi white seeds with an elliptical median longitudinal section and a circular median cross-section. Figure S5. The Miracolo Scritto bean. The seed is medium-sized (100 seed weight 55g) with an elliptical median longitudinal section, beige with brown veins, and a different colored hilum to the seed. Figure S6. The Miracolo Scritto bean, dry seeds. Figure S7. The bean Miracolo non Scritto bianco has a small seed (100 seed weight 35-40g) that is white without veins. Figure S8. Patresi cabbage leaf is of average length and width is elliptical in shape with weak midrib curvature. Figure S9. Patresi plant. In the middle part of the plant, the leaves have very weak or absent density of “curling” with semi-erect or horizontal petioles of average length. Figure S10. Palmaiola landrace: the leaf, which is partially anthocyanin-colored is blue-green in the juvenile stage, turning to medium-green when fully developed. Figure S11. Palmaiola leaf.
Author Contributions
Conceptualization, A.S. and C.D.; methodology, L.A.; G.S, C.M, F.C; investigation, F.C., L.A., A.V, G.S.; resources A.S., G.S.; data curation, L.A., G.S.; writing—original draft preparation, C.M., L.A., G.S., G.G.; G.A; writing—review and editing, L.A, C.M, G.S, A.G, and C.D.; supervision, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been carried out in the framework of the project “Censimento, caratterizzazione e valorizzazione del patrimonio frutticolo, orticolo e cerealicolo legato al territorio dell’Arcipelago Toscano” funded by PSR FEASR Regione Toscana 2014- 2020 “Sottomisura 10.2 Sostegno alla conservazione e all’uso sostenibile delle risorse genetiche in agricoltura (10.2 Support for the conservation and sustainable use of genetic resources in agriculture)”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Programma Rete Rurale Nazionale 2014-20, Piano di azione biennale 2021-23.
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an Evolved Concept of Landrace. Front. Plant Sci. 2017, 08, 145. [Google Scholar] [CrossRef]
- Santamaria, P. , Ronchi L. Varietà da conservazione in Italia: lo stato dell’arte per le specie orticole. Italus Hortus 23 (2), pp. 29-44, Italia, 2016.
- MIPAAF. Linee Guida per la conservazione e la caratterizzazione della biodiversità vegetale di interesse per l’agricoltura. Piano Nazionale sulla Biodiversità di Interesse Agricolo. INEA, Roma., Italia, 2013.
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace Germplasm for Improving Yield and Abiotic Stress Adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- FAO. Coping with climate change – the roles of genetic resources for food and agriculture. Rome, Italia, 2015.
- Piotto, B. , Giacanelli V., Ercole S. La conservazione ex situ della biodiversità delle specie vegetali spontanee e coltivate in Italia. Stato dell’arte, criticità e azioni da compiere. Manuali e linee guida ISPRA 54/2010, Italia, 2010.
- Marino, M. , Trisorio A., Bianchi P.G., Bocci R., Bravi R., Dalla Ragione I., Di Matteo A., Fideghelli C., Fontana M., Macchia M., Maggioni L., Negri V., Pignone D., Porfiri O., Schneider A., Sottile F., Vazzana C. Linee guida per la conservazione e la caratterizzazione della biodiversità vegetale di interesse per l’agricoltura. Inea, Roma. Italia, 2013.
- https://www.regione.toscana.it/documents/10180/25064512/LR+64+2004+testo+integrato_ok.pdf/240c83d9-c12e-85c0-64c9-d72880b377b5?t=1590480895986.
- Bellet, G. , Conte G., Marescot A., Mele M., Scaramuzzi S., Serra A. Tutela e valorizzazione dell’agrobiodiversità vegetale e animale in Toscana: analisi e indicazioni di policy, Italia, 2019.
- Servadei, L. Quale agricoltura nelle aree protette? ECOSCIENZA n.4/2022, pp. 20-21, Italia, 2022.
- https://en.unesco.org/biosphere/eu-na/tuscan-islands.
- Action Plan of the Tuscan Islands Biosphere Reserve 2021-2025 (2021).
- Gupta, P. K., H. S. Balyan, P. C. Sharma, and B. Ramesh. “Microsatellites in Plants: A New Class of Molecular Markers.” Current Science 70, no. 1 (1996): 45–54. Available from: http://www.jstor.org/stable/24097472.
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: an overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Andreani, L.; Camerini, G.; Delogu, C.; Fibiani, M.; Scalzo, R.L.; Manelli, E. How to save a landrace from extinction: the example of a winter squash landrace (Cucurbita maxima Duchesne) in Northern Italy (Lungavilla-Pavia). Genet. Resour. Crop. Evol. 2021, 69, 1163–1178. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar] [CrossRef]
- Doyle, J.J. and Doyle, J.L. (1987) A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochemical Bulletin, 19, 11-15.
- Hanai, L.R.; Santini, L.; Camargo, L.E.A.; Fungaro, M.H.P.; Gepts, P.; Tsai, S.M.; Vieira, M.L.C. Extension of the core map of common bean with EST-SSR, RGA, AFLP, and putative functional markers. Mol. Breed. 2009, 25, 25–45. [Google Scholar] [CrossRef]
- Lioi, L.; Piergiovanni, A.R.; Pignone, D.; Puglisi, S.; Santantonio, M.; Sonnante, G. Genetic diversity of some surviving on-farm Italian common bean (Phaseolus vulgaris L.) landraces. Plant Breed. 2005, 124, 576–581. [Google Scholar] [CrossRef]
- Guerra-Sanz, J.M. New SSR markers of Phaseolus vulgaris from sequence databases. Plant Breed. 2004, 123, 87–89. [Google Scholar] [CrossRef]
- Li, H.; Younas, M.; Wang, X.; Li, X.; Chen, L.; Zhao, B.; Chen, X.; Xu, J.; Hou, F.; Hong, B.; et al. Development of a core set of single-locus SSR markers for allotetraploid rapeseed (Brassica napus L.). Theor. Appl. Genet. 2012, 126, 937–947. [Google Scholar] [CrossRef]
- Durand, J.; Bodénès, C.; Chancerel, E.; Frigerio, J.-M.; Vendramin, G.; Sebastiani, F.; Buonamici, A.; Gailing, O.; Koelewijn, H.-P.; Villani, F.; et al. A fast and cost-effective approach to develop and map EST-SSR markers: oak as a case study. BMC Genom. 2010, 11, 570–570. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Weber, J.L. Informativeness of human (dC-dA)n · (dG-dT)n polymorphisms. Genomics 1990, 7, 524–530. [Google Scholar] [CrossRef]
- Masatoshi, N. Molecular Evolutionary Genetics., Columbia University Press, 1987. ISBN, 0231063210, 9780231063210.
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Park SDE. 2001. The Excel microsatellite toolkit (version 3.1). Animal Genomics Laboratory, University College Dublin. https://animalgenomics.ucd.ie/sdepark/ms-toolkit/.
- Park SDE (2001) Trypanotolerance in West African cattle and the population genetic effects of selection. PhD thesis. University of Dublin. Trypanotolerace in West African cattle and the population genetic effects of selection (tcd.ie).
- Liu, K.; Muse, S.V. PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. genalex 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- PHYLIP Felsenstein, J. (1993). 5 c. Joseph Felsenstein. 1993.
- Zuur, A. F. , Ieno, E. N., & Smith, G. M. (2007). Principal coordinate analysis and non-metric multidimensional scaling. Analysing ecological data, 259-264. Available from:. [CrossRef]
- https://www.lamma.toscana.it/sites/all/files/doc/energia/2_Territorio. 2024.
- Pullè, G. Monografia agraria del circondario della Isola dell'Elba, Tipografia Elbana, Portoferraio, Italia, 1879.
- AA.VV. Il germoplasma della Toscana. ARSIA - Regione Toscana, Firenze, Italia, 2000.
- AA.VV. I segni d’Elisa. Scienza e governo del territorio del principato napoleonico di Piombino, Felici Editore, Pisa, Italia, 2006.
- Catarcione, G.; Paolacci, A.R.; Alicandri, E.; Gramiccia, E.; Taviani, P.; Rea, R.; Costanza, M.T.; De Lorenzis, G.; Puccio, G.; Mercati, F.; et al. Genetic Diversity and Population Structure of Common Bean (Phaseolus vulgaris L.) Landraces in the Lazio Region of Italy. Plants 2023, 12, 744. [Google Scholar] [CrossRef]
- Falcione, M.; Simiele, M.; Renella, A.; Scippa, G.S.; Di Martino, P.; Trupiano, D. A Multi-Level Approach as a Powerful Tool to Identify and Characterize Some Italian Autochthonous Common Bean (Phaseolus vulgaris L.) Landraces under a Changing Environment. Plants 2022, 11, 2790. [Google Scholar] [CrossRef]
- Raggi, L.; Tiranti, B.; Negri, V. Italian common bean landraces: diversity and population structure. Genet. Resour. Crop. Evol. 2012, 60, 1515–1530. [Google Scholar] [CrossRef]
- Piergiovanni, A.R.; Lioi, L. Italian Common Bean Landraces: History, Genetic Diversity and Seed Quality. Diversity 2010, 2, 837–862. [Google Scholar] [CrossRef]
- Camangi, F. , Stefani A. I Quaderni del Parco n.8. Piante, essenze e tradizioni nell’Arcipelago Toscano, Italia, 2017.
- El-Esawi, M.A.; Germaine, K.; Bourke, P.; Malone, R. Genetic diversity and population structure of Brassica oleracea germplasm in Ireland using SSR markers. Comptes Rendus Biol. 2016, 339, 133–140. [Google Scholar] [CrossRef]
- Louarn, S.; Torp, A.M.; Holme, I.B.; Andersen, S.B.; Jensen, B.D. Database derived microsatellite markers (SSRs) for cultivar differentiation in Brassica oleracea. Genet. Resour. Crop. Evol. 2007, 54, 1717–1725. [Google Scholar] [CrossRef]
- ASFI, Fortezze e Fabbriche, Fabbriche Lorenesi, 2124 (https://archiviodistatofirenze.cultura.gov.it/inventari/s/fortezze_fabbriche_dis/dati/parte2h.html).
- 2806.
- Monti I, Zecchini M, L’isola di palmaiola nell’arcipelago toscano. l’eremo, la torre e il faro. 2023.
- Caruel, T. Prodromo della flora toscana, ossia, Catalogo metodico delle piante che nascono salvatiche in Toscana e nelle sue isole, o che vi sono estesamente coltivate, con la indicazione dei luoghi nei quali si di Teodoro Caruel.; Smithsonian Institution: Washington, DC, United States, 1860. [Google Scholar]
- Supplemento generale al “Prodromo della Flora toscana di T. Caruel”: 71, Società Botanica Italiana,Italia,1897. https://www.google.it/books/edition/Supplemento_generale_al_Prodromo_della_f/t78ZAAAAYAAJ?hl=it&gbpv=1&dq=inauthor:%22Eugenio+Baroni%22&printsec=frontcover.
- von Bothmer, R.; Gustafsson, M.; Snogerup, S. Brassica sect.Brassica (Brassicaceae). Genet. Resour. Crop. Evol. 1995, 42, 165–178. [Google Scholar] [CrossRef]
- Maggioni, L.; Alessandrini, A. The occurrence of Brassica montana Pourr. (Brassicaceae) in the Italian regions of Emilia-Romagna and Marche, and in the Republic of San Marino. Ital. Bot. 2019, 7, 1–16. [Google Scholar] [CrossRef]
- Warwick, S.I. , Francis A., Gugel R.K. Guide to wild germplasm of Brassica and allied crops (tribe Brassiceae, Brassicaceae) (3rd edn) – Part IV – Wild crucifer species as sources of agronomic traits, 2009.
- Pelgrom, K.T.B.; Broekgaarden, C.; Voorrips, R.E.; Bas, N.; Visser, R.G.F.; Vosman, B. Host plant resistance towards the cabbage whitefly in Brassica oleracea and its wild relatives. Euphytica 2014, 202, 297–306. [Google Scholar] [CrossRef]
- Camangi, F. , Claudi A., Peria G., Sardi M., Berger H.G., M. Nardi M., Stefani A. L’Orto dei Semplici nell’Eremo di Santa Caterina a Rio nell’Elba. Frutta antica e piante spontanee nella tradizione alimentare elbana, Edizione ETS Pisa, Italia, 2014.
Figure 2.
Dendrogram calculated from the matrix of shared alleles. The landraces on the island of Elba are shown in bold.
Figure 2.
Dendrogram calculated from the matrix of shared alleles. The landraces on the island of Elba are shown in bold.
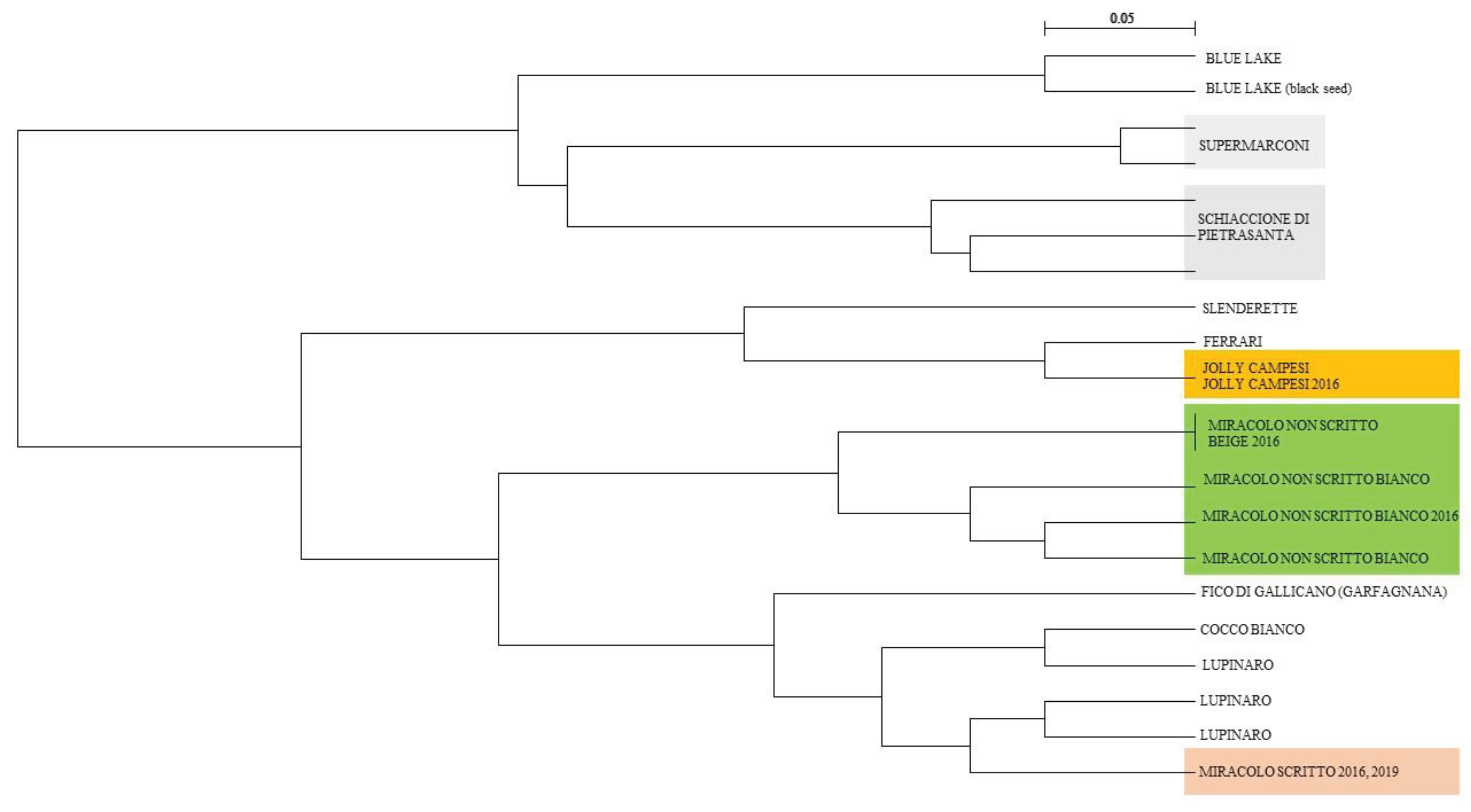
Figure 3.
PCoA score plot. The landraces on the island of Elba are highlighted in bold.
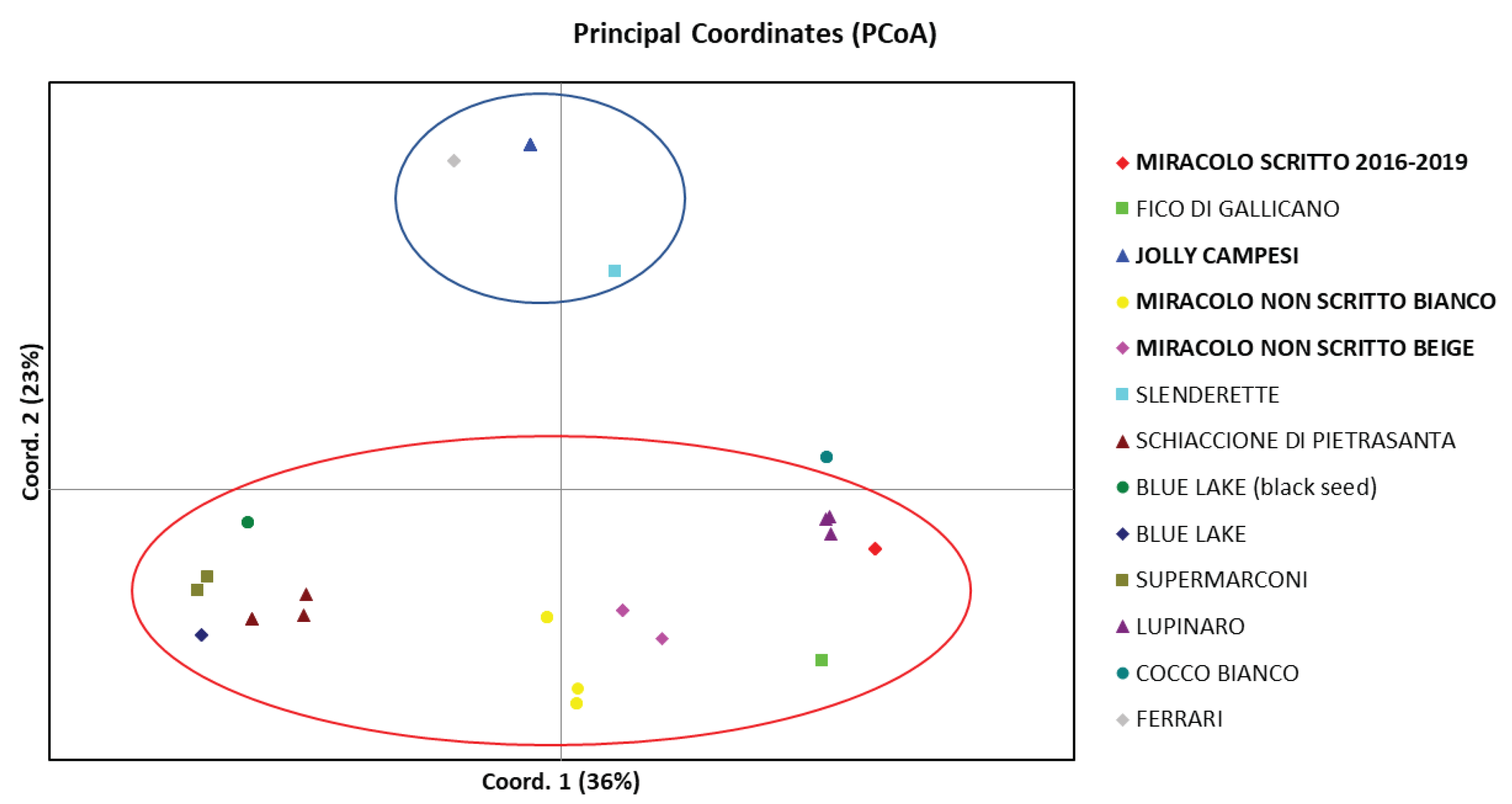
Figure 4.
UPGMA Dendrogram calculated from the matrix of shared alleles. The Patresi variety is highlighted by the orange box, while Palmaiola variety is highlighted by the blue box.
Figure 4.
UPGMA Dendrogram calculated from the matrix of shared alleles. The Patresi variety is highlighted by the orange box, while Palmaiola variety is highlighted by the blue box.

Figure 5.
PCoA score plot. The landraces on the island of Elba, namely Patresi (red dots) and Palmaiola (purple dots), are highlighted and shown in bold.
Figure 5.
PCoA score plot. The landraces on the island of Elba, namely Patresi (red dots) and Palmaiola (purple dots), are highlighted and shown in bold.
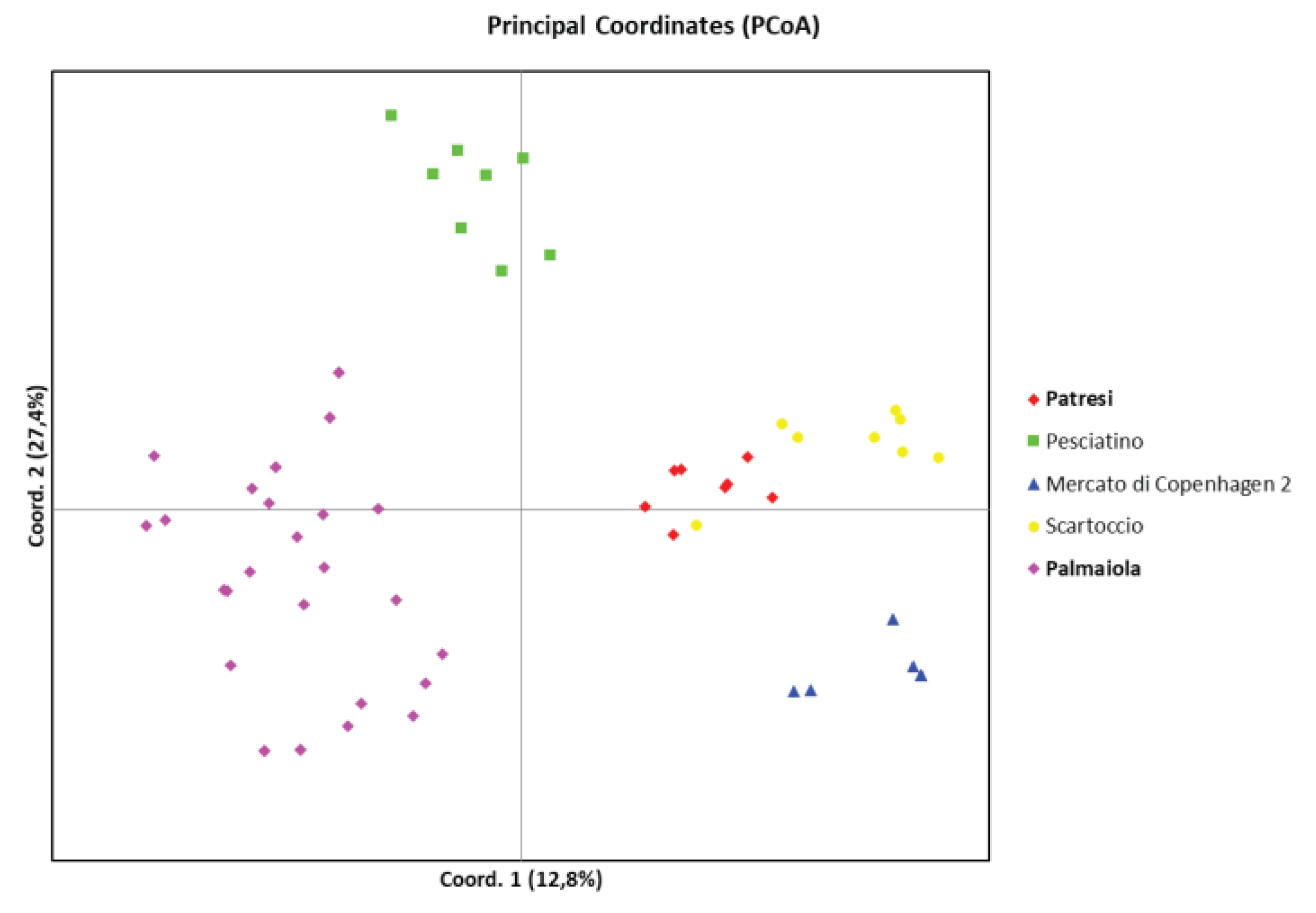

Table 1.
List of Brassica oleracea and Phaseolus vulgaris varieties.
| SPECIE | VARIETIES DESCRIBED |
|---|---|
| Brassica oleracea | Patresi (Elba landrace) |
| Brassica oleracea | Palmaiola (Elba landrace) |
| Brassica oleracea | Pesciatino |
| Brassica oleracea | Copenhagen 2 |
| Brassica oleracea | Scartoccio |
| Phaseolus vulgaris | Miracolo scritto 2016 (Elba landrace) |
| Phaseolus vulgaris | Miracolo scritto 2019 (Elba landrace) |
| Phaseolus vulgaris | Jolly Campesi (Elba landrace) |
| Phaseolus vulgaris | Jolly Campesi 2016 (Elba landrace) |
| Phaseolus vulgaris | Miracolo non scritto bianco (Elba landrace) |
| Phaseolus vulgaris | Miracolo non scritto bianco 2016 (Elba landrace) |
| Phaseolus vulgaris | Miracolo non scritto 2016 Beige (Elba landrace) |
| Phaseolus vulgaris | Slenderette |
| Phaseolus vulgaris | Fico di Gallicano (Garfagnana) |
| Phaseolus vulgaris | Piattella Schiaccione di Pietrasanta |
| Phaseolus vulgaris | Blue Lake (black seed) |
| Phaseolus vulgaris | Blue Lake |
| Phaseolus vulgaris | Supermarconi (white seed) |
| Phaseolus vulgaris | Lupinaro or Aquila |
| Phaseolus vulgaris | Cocco Bianco |
| Phaseolus vulgaris | Ferrari |
Table 2.
Main morphological characteristics used to describe the Elba bean landraces. The populations within each variety were found to be relatively homogeneous based on the results of the morphometric surveys.
Table 2.
Main morphological characteristics used to describe the Elba bean landraces. The populations within each variety were found to be relatively homogeneous based on the results of the morphometric surveys.
| Morphological characteristics | Miracolo non scritto | Miracolo scritto | Jolly Campesi | |
|---|---|---|---|---|
| Pod: length (excluding beak) cm | M SD |
10,81 0,61 |
11,40 0,95 |
14,60 0,74 |
| Pod: width at maximum point cm | M SD |
0,78 0,06 |
1,39 0,10 |
0,60 0,07 |
| Pod: transversal width cm | M SD |
0,61 0,08 |
0,53 0,06 |
0,57 0,05 |
| Pod: length of beak cm | M SD |
0,47 0,16 |
1,51 0,23 |
1,05 0,22 |
| Nr.seeds/pod | M SD |
7,13 1,36 |
5,45 0,98 |
7,20 0,79 |
| 100-Seed weight g | M SD |
37,25 2,68 |
54,40 2,17 |
23,60 1,63 |
| 20-green pod weight g | M SD |
nd nd |
122,80 4,64 |
87,90 5,88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



