
Submitted:
07 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
The emergence of the H3N2 influenza virus in 1968 marked a significant event as it crossed the species barrier. Since then, ongoing mutational dynamics have led to the formation of antigenic clusters, prompting the World Health Organization to advocate for regular updates to H3N2 vaccines. Research in the Western Pacific region underscores the necessity for heightened awareness and effective control strategies. Stemming from avian influenza A, the 1968 H3N2 influenza pandemic resulted in the deaths of one million people globally, with its seasonal variants primarily affecting older individuals and causing severe illness due to antigenic drift. To address the challenge of vaccine efficacy against H3N2 mutations, researchers are exploring innovative strategies such as precise antigenic material administration, controlled release patterns, understanding immune system mechanisms, and glycan engineering. This review comprehensively examines various aspects of the Influenza A (H3N2) virus, encompassing its virological characteristics, evolutionary trends, global epidemiology, vaccination strategies, antiviral interventions, and emerging diagnostic approaches. It underscores the impact of antigenic variation on vaccine design and effectiveness, seasonal outbreak patterns, pandemic potential, and the interplay between viral factors and host immune responses. Moreover, the review evaluates antiviral therapies and the issue of drug resistance, emphasising the necessity for multidisciplinary approaches involving researchers, healthcare professionals, and policymakers to comprehend H3N2 and enhance public health interventions.
Keywords:
H3N2 Influenza
; Antigenic Variation
; Vaccine Efficacy
; Pandemic Potential
; Multidisciplinary Approach
1. Introduction
Zoonotic influenza remains a significant global public health threat [1,2,3]. Specially avian influenza, in different forms, represents a menace for a future pandemic [4]. In the case of H3N2, this is one of the emerging forms currently concerning in some areas of the world [5]. For some experts, the risk of spillover from this and other forms seems imminent for multiple epidemics [6,7]. This review examines the host's humoral immune response, primarily targeting primary epitopes carried by the hemagglutinin (HA) surface glycoprotein in influenza A viruses. The swift mutational evolution of HA leads to "antigenic drift," allowing viruses to evade the host's adaptive immune defence [8,9]. Notably, the H3N2 influenza virus crossed the species barrier, initiating human infection in 1968, and recent antigenic modifications have led to the emergence of antigenic clusters [9]. Influenza A virus causes acute respiratory illness, contributing to a global annual mortality estimate of 250,000 to 500,000 individuals, with notable pandemics including the 1918 H1N1, 1957 H2N2, 1968 H3N2, and 2009 H1N1 outbreaks [10,11,12]. Influenza A viruses are distinguished from types B and C by their nucleoprotein (NP) and matrix (MI) proteins belonging to the Orthomyxoviridae family. The influenza A virus genome comprises 11 protein segments on a single-stranded negative-sense RNA sequence [13]. There are 144 different HA-NA combinations in the influenza A virus family, with 16 HA and 9 NA subtypes identified by neuraminidase (NA or N) and hemagglutinin (HA or H) proteins on the virus surface [14,15,16].
The World Health Organization (WHO) recommends regular updates to the H3N2 component in influenza vaccines due to ongoing mutational dynamics within H3N2 viruses [17]. Amino acid alterations at residues 222 and 225 in the hemagglutinin chain have been identified, affecting receptor-binding characteristics [18,19]. Research in the Western Pacific region aims to assess the impact of H3N2 viruses on public health and mortality rates, emphasising the need for control strategies [20]. Predicting antigenic characteristics of A (H3N2) viruses outside E-SE Asia could improve vaccine strain selection and reduce morbidity and mortality [21]. H1N1 and H3N2 subtypes have circulated in humans since 1977, with age-specific variations likely due to childhood imprinting [22]. Fever is the most common symptom for both influenza A subtypes, with higher body temperatures during the A/H3N2 season, while myalgia, coughing, and sore throats were more common during the A/H1N1 season [23]. Antigenicity characterisation poses challenges for HA inhibitory antibodies, with differences in neutralising antibody titers between children and adults, suggesting the NI assay's potential in H3N2 infection testing and vaccine selection [24]. Figure 1 provides a visual overview of the Influenza A (H3N2) virus, emphasising recent occurrences and research methodologies for analysis.
2. Global Impact and Evolution of H3N2 Influenza: A Historical Perspective
The H3N2 influenza virus originated from avian influenza A and coupled the N2 neuraminidase from the 1957 H2N2 virus with the distinct H3 hemagglutinin to generate the 1968 pandemic. When this virus was first identified in September 1968, it was thought to have killed one million people worldwide, of whom 100,000 died in the United States of America (USA) [25]. The H3N2 seasonal influenza A virus continues its worldwide circulation. These seasonal H3N2 variants frequently undergo antigenic drift, mainly affecting older individuals and leading to severe illness [26]. Since 1977, human-adapted subtypes of influenza A viruses (H1N1, H1N2, H3N2) have posed a simultaneous infection risk. While these viruses infect various animals, their primary hosts are wild birds. Influenza outbreaks affect 10–30% of the world's population and result in 290,000–650,000 deaths yearly, mostly in adults 65 years of age and older. Sub-Saharan Africa and Southeast Asia have the highest death rates from influenza epidemics [20]. In 2011, 12 cases of human infection with the new A(H3N2) virus were identified in the US, with potential person-to-person transmission [27]. Between 2011 and 2018, A(H3N2) was the cause of 39.2% of confirmed influenza infections in 27 Asian countries. Epizootiology research confirmed the H3N2 pandemic strain resulted from avian-human virus reassortment. Various H3N2 variants have amino acid changes at five antigenic sites on the H3 head, and single amino acid substitutions can alter the virus. A 2021-2022 study investigated 161 A(H3N2) virus strains, showing high genetic linkage [28]. Table 1 simplifies the origin and impact of the 1968 H3N2 influenza pandemic by representing the year-wise occurrences of H3N2 human infections, aiding in a clearer understanding of its global impact.
3. Pathophysiology of H3N2 Influenza: Complex Interactions and Inflammatory Response Mechanisms
The H3N2 influenza virus invades the upper respiratory tract, developing within respiratory epithelial cells and triggering an inflammatory response. This response, orchestrated by viral infection, produces inflammatory mediators like chemokines and cytokines. Cytokines recruit leukocytes and activate immune reactions, while chemokines attract leukocytes to infection sites, thus contributing to inflammation. However, this intricate cascade can lead to excessive inflammation, resulting in tissue damage and organ dysfunction in critical illnesses related to the virus [36]. This is comparable to the complex pathophysiology of the H3N2 influenza virus, which starts in the upper respiratory system and progresses to the lower tract. Influenza virus H3N2 damages respiratory epithelium, causing inflammation and airway obstruction [37]. Inside the body, the H3N2 influenza virus attaches to cell receptors, infiltrates host cells, and initiates replication, prompting immune responses that restrict viral replication via antibodies and cytokines.
The virus, however, can also directly damage tissues, leading to complications like pneumonia [38]. Influenza transmission studies using ferrets reveal a significant route through the soft palate, favoured by human influenza viruses' hemagglutinin proteins and similar receptors in humans [39,40]. Influenza's primary pathophysiology involves lung inflammation and dysfunction due to viral infection of the respiratory epithelium, compounded by immune responses. Chronic inflammation can lead to multiorgan failure, with predominant effects on the lungs, causing severe respiratory distress [41]. The influenza virus directly affects the respiratory tract or weakens the immune system. Lung obstruction, alveolar structural loss, and extracellular matrix deterioration are possible outcomes. Acute pneumonia is identified in 30–40% of hospitalised patients with laboratory-confirmed influenza; older patients (more than 65), Caucasians, younger than five, and inhabitants of nursing homes are at higher risk of developing pneumonia. Moreover, influenza can result in secondary bacterial infections and severe pneumonia, which increase the risk of bacterial sepsis and ARDS. Influenza A is the most frequent virus that causes adult acute respiratory distress syndrome (ARDS) [36]. H3N2 Influenza Viruses (IAVs) have evolved rapidly since 1968, incorporating N-linked glycans, increased HA molecule net charge, and altered receptor binding preferences. Researchers have modified antigenic characterisation assays to adapt to these changes. The HAI assay's use of guinea pig red blood cells and 20nM oseltamivir carboxylate enables a more precise evaluation of contemporary IAVs (Figure 2) [42].
4. Evolving Vaccination Strategies
The primary method used to create inactivated influenza vaccines is to develop viruses in chicken eggs, which have been regulated for over 70 years. The FDA distributes, tests, and adapts vaccine viruses following the WHO's strain selection process. The global manufacturing infrastructure can produce 1.5 billion doses annually. Influenza vaccination efficacy is assessed through randomised clinical trials and observational studies. Since the mid-2000s, however, nothing has been known about the effectiveness of vaccines targeted to a particular strain. A meta-analysis of influenza vaccine clinical trials conducted between 1967 and 2011 identified eight placebo-controlled trials, most showing no particular benefit against H3N2. With the development of molecular diagnostic tests, influenza infections may now be detected with high sensitivity and specificity. An inventive 'test-negative' observational study design was employed in Canada in 2005 to quantify vaccine effectiveness for the first time using RT-PCR. This methodology is used in annual VE surveys conducted in the US, Canada, Europe, and Australia [43]. The primary public health approach to curbing influenza, including the H3N2 influenza virus, revolves around vaccination. Annual vaccines are advised for individuals who are deemed to be at risk, such as the elderly and those with high rates of morbidity and death, according to the WHO [44]. Seasonal influenza, primarily H3N2, is a significant cause of respiratory disease and mortality, with increased rates of hospitalisation and excess mortality among the elderly [43].
Nevertheless, compared to other subtypes, the efficiency of the existing influenza vaccinations against H3N2 viruses is lacking [20,45]. This discrepancy is partially attributed to the H3N2 virus's rapid and unpredictable evolutionary pace in contrast to other seasonal flu viruses [46]. Further contributing to the reduced vaccine effectiveness is the composition of trivalent inactivated vaccine (TIV) influenza vaccines, which incorporate antigens from two A subtypes (A H3N2 and A H1N1) and only one B lineage. This setup frequently leads to mismatches between circulating and vaccine B strains. Consequently, the Quadrivalent influenza vaccine (QIV) has demonstrated enhanced immunogenicity compared to TIV across various age groups [44]. Furthermore, a prime-boost vaccination strategy involving the Ad5-HA + Ad5-NP vaccine followed by an inactivated H3N2 vaccine has proven effective in inducing cross-reactive immunity against H3N2 viruses in swine[47]. To improve vaccinations, yearly assessments of vaccine efficacy and genetic analysis of circulating influenza viruses are essential. The evidence base for choosing influenza vaccine viruses may be strengthened by combining clinical protection with virologic data [48]. The efficacy of influenza vaccines against influenza A(H3N2) viruses was lower than that of influenza B viruses, necessitating improved effectiveness. However, the 2016–2017 influenza vaccination trials demonstrated modest protection against outpatient influenza [49]. The antigenic distance hypothesis states that harmful interference from the previous season's immunisation may have an adverse effect on this season's protection from influenza. During three outbreaks (2010-2011, 2012-2013, 2014-2015), a study conducted in Canada assessed the effectiveness of vaccines against influenza A(H3N2) disease that required medical attention and laboratory confirmation. Consistent with the ADH, the results revealed considerable variations in the preceding vaccination effects by season. In 2014–2015, adverse effects were evident and statistically significant, indicating that low vaccine effectiveness in subsequent epidemics since 2010 could be attributed to influenza vaccinations administered more than once [50]. The discussion on H3N2 vaccine effectiveness involves summarising and presenting data from studies or clinical trials, providing a general outline to structure the discussion in Table 2. Vaccination remains the chief strategy to contain the H3N2 influenza virus. However, formulating effective vaccines encounters challenges due to the virus's rapid evolutionary rate.
4.1. Current Influenza Vaccines
The influenza vaccination now in use, which targets the H3N2 strain, has been updated for the flu season of 2022–2023 [56,57]. A component of the virus similar to A/Darwin/9/2021 (H3N2) is included in this vaccination [56,57]. However, it's crucial to understand that the flu vaccine's ability to fend off H3N2 viruses changes according to the season. Vaccinations against influenza A(H3N2) are often less effective against influenza B viruses and more effective against influenza A(H1N1) viruses [58]. The H3N2 strain in the flu season of 2021–2022 showed antigenic dissimilarity from the vaccine virus, which led to a decrease in the efficacy of the vaccine against H3N2 viruses [59]. The World Health Organization (WHO) has recommended the A/Darwin/6/2023 (H3N2) component of influenza vaccinations for the 2023–2024 influenza season in the northern hemisphere [60]. Maintaining a yearly vaccination regimen is pivotal in guarding against the flu, given the ongoing variations in the viruses [56].
4.2. Effectiveness and Limitations of Vaccines
H3N2 vaccines' effectiveness varies annually due to the virus's continuous evolution and adaptation. The potential deterioration in the effectiveness of the H3N2 vaccination can be attributed to several factors, including antigenic mismatch, vaccine component egg-adaptive alterations, and age-related effects, which allow older individuals to be less protected against A(H3N2) viruses due to previous exposure to non-A(H3N2) influenza viruses. According to recent studies, immunisation lowers the risk of influenza hospitalisation in younger, immunocompetent adults during seasons when the vaccine virus and influenza A(H3N2) are antigenically different. In 2020–2021, the COVID-19 pandemic saw a low level of influenza circulation; in 2021–2022, the circulation level increased. Immunisations decreased the likelihood of hospitalisation for younger persons but not for those over 65. Antivirals, vaccinations, and preventative measures all require advancements [59]. Among participants <50 years old, influenza vaccines displayed a 36% effectiveness against A(H3N2)-related illnesses [61].
Studies show that repeated annual influenza vaccine shots are less effective against influenza. In a trial, ferrets were given a prime-boost vaccination regimen twice and once, and they were then challenged with A/Hong Kong/4801/2014 (H3N2). The RV group lost weight more slowly and shed more virus, indicating that variations in the quality of the immune response could influence protection following recurrent immunisation [62]. Despite the continued widespread use of egg-based influenza vaccinations, emerging vaccine platforms promise to resolve drawbacks [20]. The investigation compared the H3N2-specific antibody responses of mice immunised with mRNA-LNP vaccines encoding wild-type and egg-adapted H3 antigens. The results showed that mRNA-LNP encoding wild-type H3 was superior to egg-adapted H3 or the egg-based Fluzone vaccination in neutralising the wild-type 3c.2A H3N2 virus. Both mRNA-LNP vaccinations produced significant levels of group 2 HA stalk-reactive antibodies, suggesting that mRNA-LNP-based vaccines modified with nucleosides can avoid problems associated with egg adaptation in the most recent 3c.2A H3N2 viruses. Because of their distinct glycosylation site, 3c.2A H3N2 viruses, a distinct offshoot of the 1968 H3N2 strain, came to light during the 2014–2015 influenza season and are still circulating worldwide [63,64]. Those with moderate diseases, such as mild upper respiratory tract infections, fever, or diarrhoea, can benefit from the nasal spray flu vaccine. Even if the vaccine does not precisely match the strain that is now circulating, immunisation remains the most effective means of protection against the flu overall [65].
4.3. Novel Approaches to Vaccine Design
Influenza H3N2 undergoes continuous mutations, presenting a challenge for vaccine creation. Nonetheless, researchers have explored innovative avenues to enhance vaccine efficacy. These strategies include the accurate administration of antigenic material, control over release patterns, and well-informed design based on a more profound comprehension of immune system mechanisms and pathogen-host interactions [66]. One study employed an H3N2 microneedle vaccine, which produced a cross-protective immune response against many H3N2 antigenic variants [67]. Alternate approaches encompass employing conserved antigens like HA, NA, matrix, and internal proteins, coupled with diverse vaccine platforms such as recombinant antigen/protein-based, virus-vectored, nanoparticle-based, DNA/RNA-based, virus-like particle (VLP), and multiplex vaccines [68,69,70]. How to expose highly cross-protective epitopes to the immune system has been investigated using glycan engineering of HA and NA proteins [68]. Researchers have also devised chimeric HAs by transferring unique HA globular head domains from exotic novel strains to the HA stalk domains of presently circulating human influenza viruses [71]. Moreover, adenoviruses have been transformed into vaccine vectors by disabling genes responsible for their replication, exhibiting the potential for developing a "universal" flu vaccine. Overall, pandemic-focused vaccine development aims to mitigate public health repercussions and societal disruption [72].
5. Antiviral Therapies of H3N2 Influenza Virus
Several antiviral medications are accessible for treating influenza, including H3N2. The four antiviral medications recommended for influenza treatment are oseltamivir, peramivir, zanamivir, and baloxavir [73,74]. These medications function by stopping the surfaces of infected host cells from releasing influenza virions [75]. Antiviral therapy should be started as soon as possible for hospitalised patients with suspected or confirmed influenza who have severe, complicated, or worsening symptoms or are at higher risk of developing influenza-related complications. It is not necessary to wait for laboratory proof of influenza virus infection before starting antiviral therapy in suspected influenza cases [73]. Vaccination is the most reliable method of preventing H3N2 influenza; it is recommended for all individuals six months of age and older and is typically available in the fall. Keeping up with proper sanitary practices is also crucial. These include avoiding direct contact with sick people, frequently washing hands with soap and water, and covering the mouth and nose when coughing or sneezing [75].
5.1. Neuraminidase Inhibitors (Oseltamivir, Zanamivir)
Neuraminidase inhibitors, such as oseltamivir and zanamivir, belong to a drug class that obstructs the neuraminidase enzyme, a vital component for influenza replication. The FDA has approved using oseltamivir, with a twice-daily dosing schedule, for treating acute, uncomplicated influenza within two days of the onset of illness. These antiviral drugs are commonly used to battle influenza viruses A and B [73]. Even in severe influenza, where rhabdomyolysis is present, zanamivir has proven beneficial when treatment is initiated more than 48 hours after the onset of symptoms. When used early in influenza treatment, Neuraminidase inhibitors effectively lower the incidence of severe cases and fatality [76]. However, individuals with severe immunosuppression face the highest risk of developing oseltamivir- and peramivir-resistant influenza virus infections during or after treatment with these drugs [73]. Ongoing observation of oseltamivir resistance is necessary to further protect public health due to the evolution of antiviral medication resistance among influenza viruses [77]. Public health organisations advise using neuroliminase inhibitors to treat and prevent seasonal and pandemic influenza infections (Figure 3) [78]. Sialic acid receptors are crucial for virus attachment and entry into host cells. Blocking these receptors and inhibiting virus-host cell interactions is the best way to control and prevent infection. Neuraminidase inhibitors, including oseltamivir and zanamivir, are the most efficient drugs for treating influenza A and B virus infections [79].
5.2. Polymerase Inhibitors (Baloxavir Marboxil)
Baloxavir marboxil, a polymerase inhibitor, serves as a treatment for uncomplicated influenza [80,81]. It prevents the viral polymerase complex's cap-dependent endonuclease activity, which is essential for viral replication [82]. About its use against H3N2 influenza, here are key insights: Clinical trials have established the efficacy of baloxavir marboxil against H3N2 influenza [81]. However, because to changes in the polymerase acidic protein, certain H3N2 viruses have shown reduced susceptibility to baloxavir marboxil [80,83]. Unlike neuraminidase inhibitors (NAIs) like zanamivir and oseltamivir, which are usually advised twice daily for five days, baloxavir marboxil is only given orally once [81]. In vitro, investigations have revealed that baloxavir acid, the active metabolite of baloxavir marboxil, can be combined with other inhibitors, such as NAIs and favipiravir, to augment antiviral efficacy against seasonal influenza A viruses [84]. Baloxavir marboxil exhibits potential as a treatment choice for uncomplicated influenza, including H3N2 strains. Nevertheless, the emergence of resistant strains underscores the ongoing necessity for research and the development of novel antiviral therapies.
6. Advancements in Molecular Detection Methods
6.1. Rapid Diagnostic Methods
Serological testing, antigen detection tests, and molecular assays are quick diagnostic techniques for H3N2 influenza. In ten to fifteen minutes, antigen detection tests known as Rapid Influenza Diagnostic Tests (RIDTs) can identify influenza viral antigens in respiratory specimens with low sensitivity. The antigen-detection assays known as RIDTs have moderate sensitivity and may detect influenza virus antigens in respiratory specimens in 10 to 15 minutes. These immunoassays detect influenza A and B viral nucleoprotein antigens in respiratory samples and yield a qualitative result (positive vs. negative). RIDTs have excellent specificity; however, they only offer modest sensitivity. On the other hand, outstanding sensitivity and specificity are offered by Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and other molecular techniques for identifying influenza virus RNA or nucleic acids in respiratory materials [85,86]. Among the techniques used in serological research are the Hemagglutination Inhibition Assay (HAI), Single Radial Hemolysis (SRH), Virus Neutralization Assay (VN) or Microneutralization, and Enzyme-Linked Immunosorbent Assay (ELISA). These tests detect influenza virus antibodies in serum or other bodily fluids. However, their applicability for early influenza diagnosis is limited due to the need for paired serum samples collected at least 2 weeks apart. For H3N2 influenza, molecular assays and antigen detection tests stand out as the most common quick diagnostic techniques [87].
6.2. Molecular Detection Techniques
There are numerous molecular detection methods available for identifying the H3N2 influenza virus. Reverse transcription-polymerase chain reaction (RT-PCR) and other nucleic acid amplification tests are the most widely used methods for detecting influenza virus RNA or nucleic acids in respiratory samples because of their exceptional sensitivity and specificity. Several molecular techniques can distinguish between influenza A and B infections as well as identify seasonal influenza A virus subtypes, such as A(H1N1) pdm09 or A (H3N2) [87,88,89,90]. Loop-mediated isothermal amplification (LAMP), an alternative molecular-based influenza diagnostic, provides a rapid, accurate, and dependable molecular detection technique. This technique provides a stable foundation for influenza and COVID-19 testing [91,92,93]. The Multiplex One-Step Real-Time RT-PCR technology increases efficiency and simultaneously identifies the human H3N2 virus, the pandemic (H1N1) 2009 virus, and the reassortant avian H7N9 virus [94]. Recommended by the WHO, these molecular assays offer high accuracy and efficacy for diagnosing the H3N2 influenza virus [87,88,89,90].
6.3. Point-of-Care Testing Advances
Point-of-care testing (POCT) for influenza is experiencing rapid growth, and multiple studies have demonstrated its benefits in diagnosing influenza among patients with acute respiratory tract infections [95]. POCTs represent swift diagnostic assessments performed at the point of care, whether in a doctor's office or an emergency department, delivering results within minutes [95,96]. Initially criticised for limited sensitivity and result variability, recent research has highlighted the high specificity and sensitivity of POCTs, establishing them as reliable and practical tools for early influenza identification [96,97,98]. POCTs have the potential to enhance patient flow, reduce hospitalisations, and facilitate targeted treatments [99,100]. Predominant POCT options for influenza encompass antigen detection tests and molecular assays like RT-PCR [95,96,98]. Additionally, the molecular-based Loop-mediated isothermal amplification (LAMP) assay introduces an innovative avenue for molecular diagnosis [99,101]. The introduction of POCTs offers fresh prospects for managing healthcare patients, as illustrated in Table 3.
7. Future Directions
In moving forward, this review has illuminated crucial paths for future research and development in effectively combatting the challenges of Influenza A (H3N2). These essential directions encompass strengthening global surveillance through advanced sequencing technologies and real-time data sharing, enabling rapid detection of new strains and swift public health responses. Predictive modelling holds promise in guiding the formulation of more effective vaccines by anticipating antigenic shifts. Universal vaccines targeting conserved viral regions could offer enduring protection against diverse strains. Investigating host immune responses can provide insights into disease severity and susceptibility, informing targeted interventions. Exploring combination antiviral therapies and integrating advanced diagnostic tools into public health strategies can enhance treatment and outbreak management. For comprehensive solutions, promoting multidisciplinary collaboration among professionals is essential. Investment in worldwide education and pandemic preparedness will improve the ability to control possible epidemics. Influenza A (H3N2) 's dynamic complexity must be anticipated, understood, and efficiently managed. Proactive collaboration, cutting-edge technology, and a multidisciplinary approach are essential.
8. Conclusion
This review delves into the humoral immune response to influenza A viruses, mainly focusing on recognising hemagglutinin surface glycoprotein epitopes. It emphasises the significant impact of the H3N2 virus, which breached the species barrier in 1968, leading to acute respiratory illness and global mortality. The WHO advocates for regular updates to H3N2 vaccines due to mutational dynamics, with research in the Western Pacific region underscoring the importance of awareness and control strategies. The devastating 1968 H3N2 influenza pandemic claimed one million lives globally, with seasonal variants continuing to cause severe illness. Since 1977, human-adapted subtypes have posed a significant infection risk, affecting a substantial portion of the world's population annually. Influenza A (H3N2) spreads through various means and exhibits mutability, causing sporadic outbreaks and pandemics. Notably, H3N2 has caused fatal outcomes in India, and its molecular epidemiology indicates the possibility of reinfection by the same subtype in a short time frame. The review highlights the challenge posed by H1N1 and H3N2 mutations, affecting vaccination effectiveness. The WHO strongly recommends yearly influenza vaccination, particularly for the 2023-2024 flu season. However, H3N2 mutations present obstacles to vaccine development, prompting exploration into innovative strategies such as alternative antigen formulations and pandemic-focused vaccine development. Despite longstanding regulation of inactivated influenza vaccines, their effectiveness against H3N2 viruses has been limited due to the virus's rapid evolutionary pace. The review comprehensively explores the evolution, epidemiology, clinical manifestations, vaccination strategies, and Influenza A (H3N2) antiviral interventions, underscoring the importance of diagnostic advancements and multidisciplinary collaboration for pandemic preparedness. It is a valuable resource for healthcare professionals and policymakers combating H3N2.
Funding
Universidad Continental covered the APC for this publication.
Abbreviations
| H3N2 | Hemagglutinin 3 and Neuraminidase 2 |
| CDC | Centres for Disease Control and Prevention |
| WHO | World Health Organization |
| HA | Hemagglutinin |
| RNA | Ribonucleic Acid |
| IBVs | Influenza B Viruses |
| A(H3N2) | Influenza A Subtype H3N2 |
| rRT-PCR | Real-Time Reverse Transcription Polymerase Chain Reaction |
| M | Matrix |
| ILI | influenza-like illness |
| PA | Polymerase acid |
| PB1 | polymerase basic 1 |
| PB2 | polymerase basic 2 |
| TIV | trivalent inactivated vaccine |
| QIV | Quadrivalent influenza vaccine |
| RIDTs | Rapid Influenza Diagnostic Tests |
| HAI | Hemagglutination Inhibition Assay |
| VN | Virus Neutralization Assay |
| SRH | Single Radial Homolysis |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| RT-PCR | Reverse Transcription-Polymerase Chain Reaction |
| LAMP | Loop-Mediated Isothermal Amplification |
| POCT | Point-of-care testing |
| VE | Vaccine Effectiveness |
| H3N2 | Hemagglutinin 3 and Neuraminidase 2 |
| H3N2v | H3N2 variant |
References
- Bonilla-Aldana, D.K.; Aguirre-Florez, M.; Villamizar-Peña, R.; Gutiérrez-Ocampo, E.; Henao-Martínez, J.F.; Cvetkovic-Vega, A.; Dhama, K.; Rabaan, A.; Sah, R.; Rodriguez-Morales, A.J.; et al. After SARS-CoV-2, will H5N6 and other influenza viruses follow the pandemic path? Infez Med 2020, 28, 475–485. [Google Scholar]
- Choudhary, P.; Shafaati, M.; Abu Salah, M.A.H.; Chopra, H.; Choudhary, O.P.; Silva-Cajaleon, K.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. Zoonotic diseases in a changing climate scenario: Revisiting the interplay between environmental variables and infectious disease dynamics. Travel Med Infect Dis 2024, 58, 102694. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Cardona-Ospina, J.A.; Collins, M.H. Editorial: Emerging and Re-emerging Vector-borne and Zoonotic Diseases. Front Med (Lausanne) 2021, 8, 714630. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Bonilla-Aldana, D.K.; Paniz-Mondolfi, A.E. Concerns about influenza H5N8 outbreaks in humans and birds: Facing the next airborne pandemic? Travel Med Infect Dis 2021, 41, 102054. [Google Scholar] [CrossRef]
- Priyanka; Khandia, R.; Chopra, H.; Choudhary, O.P.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. The re-emergence of H3N2 influenza: An update on the risk and containment. New Microbes New Infect 2023, 53, 101147. [Google Scholar] [CrossRef]
- Ahmad, F.; Haque, S.; Tawil, S.; Husni, R.; Bonilla-Aldana, D.K.; Montenegro-Idrogo, J.J.; Rodriguez-Morales, A.J. Avian influenza spillover to humans: Are we prepared to deal with another potential pandemic? Travel Med Infect Dis 2023, 55, 102634. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Paniz-Mondolfi, A.E.; Faccini-Martínez Á, A.; Henao-Martínez, A.F.; Ruiz-Saenz, J.; Martinez-Gutierrez, M.; Alvarado-Arnez, L.E.; Gomez-Marin, J.E.; Bueno-Marí, R.; Carrero, Y.; et al. The Constant Threat of Zoonotic and Vector-Borne Emerging Tropical Diseases: Living on the Edge. Front Trop Dis 2021, 2, 676905. [Google Scholar] [CrossRef]
- Yu, J.; Li, F.; Wang, D. The first decade of research advances in influenza D virus. J Gen Virol 2021, 102. [Google Scholar] [CrossRef]
- Forghani, M.; Khachay, M. Feature Extraction Technique for Prediction the Antigenic Variants of the Influenza Virus. International Journal of Medical and Health Sciences 2018, 12, 525–530. [Google Scholar]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. The Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Sahu, M.; Singh, N.; Shukla, M.K.; Potdar, V.A.; Sharma, R.K.; Sahare, L.K.; Ukey, M.J.; Barde, P.V. Molecular and epidemiological analysis of pandemic and post-pandemic influenza A (H1N1) pdm09 virus from central India. Journal of Medical Virology 2018, 90, 447–455. [Google Scholar] [CrossRef]
- Somerville, L.K.; Basile, K.; Dwyer, D.E.; Kok, J. The impact of influenza virus infection in pregnancy. Future microbiology 2018, 13, 263–274. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiological reviews 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Vemula, S.V.; Zhao, J.; Liu, J.; Wang, X.; Biswas, S.; Hewlett, I. Current approaches for diagnosis of influenza virus infections in humans. Viruses 2016, 8, 96. [Google Scholar] [CrossRef]
- Riedel, S. Crossing the species barrier: the threat of an avian influenza pandemic. In Proceedings of Baylor University Medical Center Proceedings; pp. 16-20.
- Potter, C.; Jennings, R.; Clark, A.; Ali, M. Interference following dual inoculation with influenza A (H3N2) and (H1N1) viruses in ferrets and volunteers. Journal of Medical Virology 1983, 11, 77–86. [Google Scholar] [CrossRef]
- Lin, Y.; Wharton, S.A.; Whittaker, L.; Dai, M.; Ermetal, B.; Lo, J.; Pontoriero, A.; Baumeister, E.; Daniels, R.S.; McCauley, J.W. The characteristics and antigenic properties of recently emerged subclade 3C. 3a and 3C. 2a human influenza A (H3N2) viruses passaged in MDCK cells. Influenza and other respiratory viruses 2017, 11, 263–274. [Google Scholar] [CrossRef]
- Xu, R.; Krause, J.C.; McBride, R.; Paulson, J.C.; Crowe Jr, J.E.; Wilson, I.A. A recurring motif for antibody recognition of the receptor-binding site of influenza hemagglutinin. Nature structural & molecular biology 2013, 20, 363–370. [Google Scholar]
- Lin, Y.P.; Xiong, X.; Wharton, S.A.; Martin, S.R.; Coombs, P.J.; Vachieri, S.G.; Christodoulou, E.; Walker, P.A.; Liu, J.; Skehel, J.J. Evolution of the receptor binding properties of the influenza A (H3N2) hemagglutinin. Proceedings of the National Academy of Sciences 2012, 109, 21474–21479. [Google Scholar] [CrossRef]
- Kang, M.; Zanin, M.; Wong, S.-S. Subtype H3N2 influenza A viruses: an unmet challenge in the Western Pacific. Vaccines 2022, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.A.; Jones, T.C.; Barr, I.G.; Cox, N.J.; Garten, R.J.; Gregory, V.; Gust, I.D.; Hampson, A.W.; Hay, A.J.; Hurt, A.C. The global circulation of seasonal influenza A (H3N2) viruses. Science 2008, 320, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Gostic, K.M.; Bridge, R.; Brady, S.; Viboud, C.; Worobey, M.; Lloyd-Smith, J.O. Childhood immune imprinting to influenza A shapes birth year-specific risk during seasonal H1N1 and H3N2 epidemics. PLoS pathogens 2019, 15, e1008109. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Ryu, G.; Lee, K.-I. Symptomatic Differences between Influenza A/H3N2 and A/H1N1 in Korea. Journal of Clinical Medicine 2023, 12, 5651. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Lin, X.; Guo, J.; Liu, L.; Li, Z.; Lan, Y.; Liu, L.; Guo, J.; Lu, J.; Huang, W. The antibody response against neuraminidase in human influenza A (H3N2) virus infections during 2018/2019 flu season: focusing on the epitopes of 329-N-glycosylation and E344 in N2. Frontiers in Microbiology 2022, 13, 845088. [Google Scholar] [CrossRef]
- Jester, B.J.; Uyeki, T.M.; Jernigan, D.B. Fifty years of influenza A (H3N2) following the pandemic of 1968. American journal of public health 2020, 110, 669–676. [Google Scholar] [CrossRef]
- CDC. 1968 Pandemic (H3N2 virus). Availabe online: https://www.cdc.gov/flu/pandemic-resources/1968-pandemic.html (accessed on.
- Lindstrom, S.; Garten, R.; Balish, A.; Shu, B.; Emery, S.; Berman, L.; Barnes, N.; Sleeman, K.; Gubareva, L.; Villanueva, J. Human infections with novel reassortant influenza A (H3N2) v viruses, United States, 2011. Emerging infectious diseases 2012, 18, 834. [Google Scholar] [CrossRef]
- Galli, C.; Pellegrinelli, L.; Giardina, F.; Ferrari, G.; Renteria, S.C.U.; Novazzi, F.; Masi, E.; Pagani, E.; Piccirilli, G.; Mauro, M.V. On the lookout for influenza viruses in Italy during the 2021-2022 season: Along came A (H3N2) viruses with a new phylogenetic makeup of their hemagglutinin. Virus Research 2023, 324, 199033. [Google Scholar] [CrossRef]
- Viboud, C.; Grais, R.F.; Lafont, B.A.; Miller, M.A.; Simonsen, L. Multinational impact of the 1968 Hong Kong influenza pandemic: evidence for a smoldering pandemic. The Journal of infectious diseases 2005, 192, 233–248. [Google Scholar] [CrossRef]
- Yu, H.; Hua, R.-H.; Zhang, Q.; Liu, T.-Q.; Liu, H.-L.; Li, G.-X.; Tong, G.-Z. Genetic evolution of swine influenza A (H3N2) viruses in China from 1970 to 2006. Journal of Clinical Microbiology 2008, 46, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Scholtissek, C.; Rohde, W.v.; Von Hoyningen, V.; Rott, R. On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology 1978, 87, 13–20. [Google Scholar] [CrossRef]
- Wen, F.; Bedford, T.; Cobey, S. Explaining the geographical origins of seasonal influenza A (H3N2). Proceedings of the royal society b: biological sciences 2016, 283, 20161312. [Google Scholar] [CrossRef]
- Pearce, M.B.; Jayaraman, A.; Pappas, C.; Belser, J.A.; Zeng, H.; Gustin, K.M.; Maines, T.R.; Sun, X.; Raman, R.; Cox, N.J. Pathogenesis and transmission of swine origin A (H3N2) v influenza viruses in ferrets. Proceedings of the National Academy of Sciences 2012, 109, 3944–3949. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Vincent, A.L.; Kitikoon, P.; Holmes, E.C.; Gramer, M.R. Evolution of novel reassortant A/H3N2 influenza viruses in North American swine and humans, 2009–2011. Journal of virology 2012, 86, 8872–8878. [Google Scholar] [CrossRef]
- Skowronski, D.M.; Janjua, N.Z.; De Serres, G.; Purych, D.; Gilca, V.; Scheifele, D.W.; Dionne, M.; Sabaiduc, S.; Gardy, J.L.; Li, G. Cross-reactive and vaccine-induced antibody to an emerging swine-origin variant of influenza A virus subtype H3N2 (H3N2v). The Journal of infectious diseases 2012, 206, 1852–1861. [Google Scholar] [CrossRef]
- Kalil, A.C.; Thomas, P.G. Influenza virus-related critical illness: pathophysiology and epidemiology. Critical care 2019, 23, 258. [Google Scholar] [CrossRef]
- Sun, H.; Blackmon, S.; Yang, G.; Waters, K.; Li, T.; Tangwangvivat, R.; Xu, Y.; Shyu, D.; Wen, F.; Cooley, J. Zoonotic risk, pathogenesis, and transmission of avian-origin H3N2 canine influenza virus. Journal of virology 2017, 91, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, J.E.; Kuchel, G.A.; Zhou, X.; Swain, S.L.; Haynes, L. T-cell immunity to influenza in older adults: a pathophysiological framework for development of more effective vaccines. Frontiers in immunology 2016, 7, 180251. [Google Scholar] [CrossRef]
- Lakdawala, S.S.; Jayaraman, A.; Halpin, R.A.; Lamirande, E.W.; Shih, A.R.; Stockwell, T.B.; Lin, X.; Simenauer, A.; Hanson, C.T.; Vogel, L. The soft palate is an important site of adaptation for transmissible influenza viruses. Nature 2015, 526, 122–125. [Google Scholar] [CrossRef]
- Nicholls, J.M.; Bourne, A.J.; Chen, H.; Guan, Y.; Peiris, J.M. Sialic acid receptor detection in the human respiratory tract: evidence for widespread distribution of potential binding sites for human and avian influenza viruses. Respiratory research 2007, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zangrillo, A.; Biondi-Zoccai, G.; Landoni, G.; Frati, G.; Patroniti, N.; Pesenti, A.; Pappalardo, F. Extracorporeal membrane oxygenation (ECMO) in patients with H1N1 influenza infection: a systematic review and meta-analysis including 8 studies and 266 patients receiving ECMO. Critical Care 2013, 17, 1–8. [Google Scholar] [CrossRef]
- Allen, J.D.; Ross, T.M. H3N2 influenza viruses in humans: Viral mechanisms, evolution, and evaluation. Human vaccines & immunotherapeutics 2018, 14, 1840–1847. [Google Scholar]
- Belongia, E.A.; McLean, H.Q. Influenza vaccine effectiveness: defining the H3N2 problem. Clinical Infectious Diseases 2019, 69, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Barberis, I.; Martini, M.; Iavarone, F.; Orsi, A. Available influenza vaccines: immunization strategies, history and new tools for fighting the disease. Journal of preventive medicine and hygiene 2016, 57, E41. [Google Scholar] [PubMed]
- Flannery, B.; Fry, A.M. Comparing influenza vaccine types: the path toward improved influenza vaccine strategies. Oxford University Press US: 2019; Vol. 220, pp 1237-1239.
- Perofsky, A.C.; Nelson, M.I. The challenges of vaccine strain selection. Elife 2020, 9, e62955. [Google Scholar] [CrossRef] [PubMed]
- Mancera Gracia, J.C.; Pearce, D.S.; Masic, A.; Balasch, M. Influenza A virus in swine: epidemiology, challenges and vaccination strategies. Frontiers in Veterinary Science 2020, 7, 647. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Kondor, R.J.G.; Chung, J.R.; Gaglani, M.; Reis, M.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S. Spread of antigenically drifted influenza A (H3N2) viruses and vaccine effectiveness in the United States during the 2018–2019 season. The Journal of infectious diseases 2020, 221, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Chung, J.R.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; McLean, H.Q.; Gaglani, M.; Murthy, K.; Zimmerman, R.K.; Nowalk, M.P. Influenza vaccine effectiveness in the United States during the 2016–2017 season. Clinical Infectious Diseases 2019, 68, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Chambers, C.; De Serres, G.; Sabaiduc, S.; Winter, A.-L.; Dickinson, J.A.; Gubbay, J.B.; Fonseca, K.; Drews, S.J.; Charest, H. Serial vaccination and the antigenic distance hypothesis: effects on influenza vaccine effectiveness during A (H3N2) epidemics in Canada, 2010–2011 to 2014–2015. The Journal of infectious diseases 2017, 215, 1059–1099. [Google Scholar] [CrossRef]
- Kissling, E.; Pozo, F.; Buda, S.; Vilcu, A.-M.; Rizzo, C.; Gherasim, A.; Horváth, J.K.; Brytting, M.; Domegan, L.; Meijer, A. Effectiveness of influenza vaccine against influenza A in Europe in seasons of different A (H1N1) pdm09 and the same A (H3N2) vaccine components (2016–17 and 2017–18). Vaccine: X 2019, 3, 100042. [Google Scholar] [CrossRef]
- Skowronski, D.M.; Leir, S.; Sabaiduc, S.; Chambers, C.; Zou, M.; Rose, C.; Olsha, R.; Dickinson, J.A.; Winter, A.-L.; Jassem, A. Influenza vaccine effectiveness by A (H3N2) phylogenetic subcluster and prior vaccination history: 2016–2017 and 2017–2018 epidemics in Canada. The Journal of Infectious Diseases 2022, 225, 1387–1398. [Google Scholar] [CrossRef]
- Rose, A.M.; Kissling, E.; Gherasim, A.; Casado, I.; Bella, A.; Launay, O.; Lazăr, M.; Marbus, S.; Kuliese, M.; Syrjänen, R. Vaccine effectiveness against influenza A (H3N2) and B among laboratory-confirmed, hospitalised older adults, Europe, 2017-18: A season of B lineage mismatched to the trivalent vaccine. Influenza and other respiratory viruses 2020, 14, 302–310. [Google Scholar] [CrossRef]
- Trebbien, R.; Fischer, T.K.; Krause, T.G.; Nielsen, L.; Nielsen, X.C.; Weinreich, L.S.; Lis-Tønder, J.; Skov, M.N.; Christiansen, C.B.; Emborg, H.-D. Changes in genetically drifted H3N2 influenza A viruses and vaccine effectiveness in adults 65 years and older during the 2016/17 season in Denmark. Journal of Clinical Virology 2017, 94, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Chambers, C.; Sabaiduc, S.; Dickinson, J.A.; Winter, A.-L.; De Serres, G.; Drews, S.J.; Jassem, A.; Gubbay, J.B.; Charest, H. Interim estimates of 2016/17 vaccine effectiveness against influenza A (H3N2), Canada, January 2017. Eurosurveillance 2017, 22, 30460. [Google Scholar] [CrossRef] [PubMed]
- CDC. Frequently Asked Influenza (Flu) Questions: 2022-2023 Season. Availabe online: https://www.cdc.gov/flu/season/faq-flu-season-2022-2023.htm (accessed on.
- FDA, U. Influenza Vaccine for the 2022-2023 Season. Availabe online: https://www.fda.gov/vaccines-bloodbiologics/lot-release/influenza-vaccine-2022-2023-season (accessed on.
- CDC. Vaccine Effectiveness: How Well Do Flu Vaccines Work? Availabe online: https://www.cdc.gov/flu/vaccines-work/vaccineeffect.htm (accessed on.
- Tenforde, M.W.; Patel, M.M.; Lewis, N.M.; Adams, K.; Gaglani, M.; Steingrub, J.S.; Shapiro, N.I.; Duggal, A.; Prekker, M.E.; Peltan, I.D. Vaccine effectiveness against influenza A (H3N2)–associated hospitalized illness: United States, 2022. Clinical Infectious Diseases 2023, 76, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- WHO. Recommendations announced for influenza vaccine composition for the 2023-2024 northern hemisphere influenza season. Availabe online: https://www.who.int/news/item/24-02-2023-recommendations-announced-for-influenza-vaccine-composition-for-the-2023-2024-northern-hemisphere-influenza-season (accessed on.
- Price, A.M.; Flannery, B.; Talbot, H.K.; Grijalva, C.G.; Wernli, K.J.; Phillips, C.H.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; McLean, H.Q. Influenza vaccine effectiveness against influenza A (H3N2)-related illness in the United States during the 2021–2022 influenza season. Clinical Infectious Diseases 2023, 76, 1358–1363. [Google Scholar] [CrossRef] [PubMed]
- Music, N.; Tzeng, W.-P.; Liaini Gross, F.; Levine, M.Z.; Xu, X.; Shieh, W.-J.; Tumpey, T.M.; Katz, J.M.; York, I.A. Repeated vaccination against matched H3N2 influenza virus gives less protection than single vaccination in ferrets. npj Vaccines 2019, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Gouma, S.; Furey, C.; Santos, J.J.; Parkhouse, K.; Weirick, M.; Muramatsu, H.; Pardi, N.; Fan, S.H.; Weissman, D.; Hensley, S.E. Nucleoside-modified mRNA-based influenza vaccines circumvent problems associated with H3N2 vaccine strain egg adaptation. Journal of Virology 2023, 97, e01723–01722. [Google Scholar] [CrossRef] [PubMed]
- Gouma, S.; Kim, K.; Weirick, M.E.; Gumina, M.E.; Branche, A.; Topham, D.J.; Martin, E.T.; Monto, A.S.; Cobey, S.; Hensley, S.E. Middle-aged individuals may be in a perpetual state of H3N2 influenza virus susceptibility. Nature Communications 2020, 11, 4566. [Google Scholar] [CrossRef]
- CDC. Live Attenuated Influenza Vaccine [LAIV] (The Nasal Spray Flu Vaccine). Availabe online: https://www.cdc.gov/flu/prevent/nasalspray.htm (accessed on.
- Wallis, J.; Shenton, D.; Carlisle, R. Novel approaches for the design, delivery and administration of vaccine technologies. Clinical & Experimental Immunology 2019, 196, 189–204. [Google Scholar]
- Shin, Y.; Kim, J.; Seok, J.H.; Park, H.; Cha, H.-R.; Ko, S.H.; Lee, J.M.; Park, M.-S.; Park, J.-H. Development of the H3N2 influenza microneedle vaccine for cross-protection against antigenic variants. Scientific reports 2022, 12, 12189. [Google Scholar] [CrossRef]
- Dormitzer, P.; Tsai, T.; Del Giudice, G. New technologies for influenza vaccines. Human vaccines & immunotherapeutics 2012, 8, 45–58. [Google Scholar]
- Nguyen, Q.-T.; Choi, Y.-K. Targeting antigens for universal influenza vaccine development. Viruses 2021, 13, 973. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-J.; Crank, M.C.; Shiver, J.; Graham, B.S.; Mascola, J.R.; Nabel, G.J. Next-generation influenza vaccines: opportunities and challenges. Nature reviews Drug discovery 2020, 19, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Chen, C.; Han, X.; Lin, S.; Ao, X.; Han, X.; Wang, J.; Ye, H. Structural insights for anti-influenza vaccine design. Computational and structural biotechnology journal 2019, 17, 475–483. [Google Scholar] [CrossRef] [PubMed]
- CDC. Interim Updated Planning Guidance on Allocating and Targeting Pandemic Influenza Vaccine during an Influenza Pandemic. Availabe online: https://www.cdc.gov/flu/pandemic-resources/nationalstrategy/planning-guidance/index.html (accessed on.
- CDC. Influenza Antiviral Medications: Summary for Clinicians. Availabe online: https://www.cdc.gov/flu/professionals/antivirals/summary-clinicians.htm (accessed on.
- CDC. Variant Influenza Virus Treatment. Availabe online: https://www.cdc.gov/flu/swineflu/variant-treatment.htm (accessed on.
- De Clercq, E. Antiviral agents active against influenza A viruses. Nature reviews Drug discovery 2006, 5, 1015–1025. [Google Scholar] [CrossRef]
- Alchikh, M.; Obermeier, P.E.; Schweiger, B.; Rath, B.A. Successful Treatment of Complicated Influenza A (H3N2) Virus Infection and Rhabdomyolysis with Compassionate Use of IV Zanamivir. Pharmaceuticals 2023, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- CDC. Antiviral Drug Resistance among Influenza Viruses. Availabe online: https://www.cdc.gov/flu/professionals/antivirals/antiviral-drug-resistance.htm (accessed on.
- Jefferson, T.; Jones, M.A.; Doshi, P.; Del Mar, C.B.; Hama, R.; Thompson, M.J.; Spencer, E.A.; Onakpoya, I.J.; Mahtani, K.R.; Nunan, D. Neuraminidase inhibitors for preventing and treating influenza in adults and children. Cochrane database of systematic reviews 2014. [Google Scholar] [CrossRef]
- Mahal, A.; Duan, M.; Zinad, D.S.; Mohapatra, R.K.; Obaidullah, A.J.; Wei, X.; Pradhan, M.K.; Das, D.; Kandi, V.; Zinad, H.S. , et al. Recent progress in chemical approaches for the development of novel neuraminidase inhibitors. RSC Adv 2021, 11, 1804–1840. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Zagribelnyy, B.; Pascua, P.N.Q.; Bezrukov, D.S.; Barman, S.; Okda, F.; Webby, R.J.; Ivanenkov, Y.A.; Govorkova, E.A. Influenza A virus polymerase acidic protein E23G/K substitutions weaken key baloxavir drug-binding contacts with minimal impact on replication and transmission. PLoS Pathogens 2022, 18, e1010698. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Morita, H.; Ogawa, R.; Nakamura, K.; Fujisaki, S.; Shirakura, M.; Kuwahara, T.; Kishida, N.; Watanabe, S.; Odagiri, T. Susceptibility of influenza viruses to the novel cap-dependent endonuclease inhibitor baloxavir marboxil. Frontiers in microbiology 2018, 9, 3026. [Google Scholar] [CrossRef]
- Mifsud, E.J.; Hayden, F.G.; Hurt, A.C. Antivirals targeting the polymerase complex of influenza viruses. Antiviral research 2019, 169, 104545. [Google Scholar] [CrossRef]
- Takashita, E.; Kawakami, C.; Ogawa, R.; Morita, H.; Fujisaki, S.; Shirakura, M.; Miura, H.; Nakamura, K.; Kishida, N.; Kuwahara, T. Influenza A (H3N2) virus exhibiting reduced susceptibility to baloxavir due to a polymerase acidic subunit I38T substitution detected from a hospitalised child without prior baloxavir treatment, Japan, January 2019. Eurosurveillance 2019, 24, 1900170. [Google Scholar] [CrossRef] [PubMed]
- Checkmahomed, L.; Padey, B.; Pizzorno, A.; Terrier, O.; Rosa-Calatrava, M.; Abed, Y.; Baz, M.; Boivin, G. In vitro combinations of baloxavir acid and other inhibitors against seasonal influenza A viruses. Viruses 2020, 12, 1139. [Google Scholar] [CrossRef]
- CDC. Rapid Influenza Diagnostic Tests. Availabe online: https://www.cdc.gov/flu/professionals/diagnosis/clinician_guidance_ridt.htm (accessed on.
- CDC. Overview of Influenza Testing Methods. Availabe online: https://www.cdc.gov/flu/professionals/diagnosis/overview-testing-methods.htm (accessed on.
- Ravina; Manjeet; Mohan, H.; Narang, J.; Pundir, S.; Pundir, C.S. A changing trend in diagnostic methods of Influenza A (H3N2) virus in human: a review. 3 Biotech 2021, 11, 1–13. [Google Scholar] [CrossRef]
- CDC. Interim Guidance for Enhanced Influenza Surveillance: Additional Specimen Collection for Detection of Influenza A (H3N2) Variant Virus Infections. Availabe online: https://www.cdc.gov/flu/swineflu/variant/h3n2v-surveillance.htm (accessed on.
- CDC. Overview of Influenza Testing Methods. Availabe online: https://www.cdc.gov/flu/professionals/diagnosis/overview-testing-methods.htm (accessed on.
- CDC. Information on Rapid Molecular Assays, RT-PCR, and other Molecular Assays for Diagnosis of Influenza Virus Infection. Availabe online: https://www.cdc.gov/flu/professionals/diagnosis/molecularassays.htm (accessed on.
- Poon, L.L.; Leung, C.S.; Chan, K.H.; Lee, J.H.; Yuen, K.Y.; Guan, Y.; Peiris, J.S. Detection of human influenza A viruses by loop-mediated isothermal amplification. Journal of clinical microbiology 2005, 43, 427–430. [Google Scholar] [CrossRef]
- Kang, J.-S.; Seo, M.-R.; Chung, Y.-J. Development of reverse-transcription loop-mediated isothermal amplification assays for point-of-care testing of human influenza virus subtypes H1N1 and H3N2. Genomics & Informatics 2022, 20. [Google Scholar]
- Heithoff, D.M.; Barnes, L.; Mahan, S.P.; Fox, G.N.; Arn, K.E.; Ettinger, S.J.; Bishop, A.M.; Fitzgibbons, L.N.; Fried, J.C.; Low, D.A. Assessment of a smartphone-based loop-mediated isothermal amplification assay for detection of SARS-CoV-2 and influenza viruses. JAMA network open 2022, 5, e2145669–e2145669. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Zhao, D.; Xie, G.; Yang, X.; Huo, Z.; Zheng, S.; Yu, F.; Chen, Y. Simultaneous detection of influenza A subtypes of H3N2 virus, pandemic (H1N1) 2009 virus and reassortant avian H7N9 virus in humans by multiplex one-step real-time RT-PCR assay. Springerplus 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Egilmezer, E.; Walker, G.J.; Bakthavathsalam, P.; Peterson, J.R.; Gooding, J.J.; Rawlinson, W.; Stelzer-Braid, S. Systematic review of the impact of point-of-care testing for influenza on the outcomes of patients with acute respiratory tract infection. Reviews in medical virology 2018, 28, e1995. [Google Scholar] [CrossRef]
- Dickson, E.M.; Zambon, M.; Pebody, R.; de Lusignan, S.; Elliot, A.J.; Ellis, J.; Lackenby, A.; Smith, G.; McMenamin, J. Do point-of-care tests (POCTs) offer a new paradigm for the management of patients with influenza? Eurosurveillance 2020, 25, 1900420. [Google Scholar] [CrossRef]
- Nelson, P.P.; Rath, B.A.; Fragkou, P.C.; Antalis, E.; Tsiodras, S.; Skevaki, C. Current and future point-of-care tests for emerging and new respiratory viruses and future perspectives. Frontiers in cellular and infection microbiology 2020, 10, 181. [Google Scholar] [CrossRef]
- Maignan, M.; Viglino, D.; Hablot, M.; Termoz Masson, N.; Lebeugle, A.; Collomb Muret, R.; Mabiala Makele, P.; Guglielmetti, V.; Morand, P.; Lupo, J. Diagnostic accuracy of a rapid RT-PCR assay for point-of-care detection of influenza A/B virus at emergency department admission: A prospective evaluation during the 2017/2018 influenza season. PloS one 2019, 14, e0216308. [Google Scholar] [CrossRef] [PubMed]
- Melhuish, A.; Vargas-Palacios, A.; Yaziji, N.; Selfridge, J.; Pisavadia, M.; Sagoo, G.S.; Minton, J. Cost evaluation of point-of-care testing for community-acquired influenza in adults presenting to the emergency department. Journal of Clinical Virology 2020, 129, 104533. [Google Scholar] [CrossRef] [PubMed]
- Fjelltveit, E.B.; Cox, R.J.; Østensjø, J.; Blomberg, B.; Ebbesen, M.H.; Langeland, N.; Mohn, K.G. Point-of-Care Influenza Testing Impacts Clinical Decision, Patient Flow, and Length of Stay in Hospitalized Adults. The Journal of Infectious Diseases 2022, 226, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Weitzel, T.; Schnabel, E.; Dieckmann, S.; Börner, U.; Schweiger, B. Evaluation of a new point-of-care test for influenza A and B virus in travellers with influenza-like symptoms. Clinical microbiology and infection 2007, 13, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Ravina; Dalal, A.; Mohan, H.; Prasad, M.; Pundir, C. Detection methods for influenza A H1N1 virus with special reference to biosensors: a review. Bioscience reports 2020, 40, BSR20193852. [Google Scholar] [CrossRef] [PubMed]
- Storms, S.M.; Shisler, J.; Nguyen, T.H.; Zuckermann, F.A.; Lowe, J.F. RT-LAMP as Diagnostic Tool for Influenza—A Virus Detection in Swine. Veterinary Sciences 2023, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.N.; McCarthy, C.; Lantigua, D.; Camci-Unal, G. Development of diagnostic tests for detection of SARS-CoV-2. Diagnostics 2020, 10, 905. [Google Scholar] [CrossRef]
- Stockton, J.; Ellis, J.; Saville, M.; Clewley, J.; Zambon, M. Multiplex PCR for typing and subtyping influenza and respiratory syncytial viruses. Journal of clinical microbiology 1998, 36, 2990–2995. [Google Scholar] [CrossRef]
- Zhou, B.; Deng, Y.-M.; Barnes, J.R.; Sessions, O.M.; Chou, T.-W.; Wilson, M.; Stark, T.J.; Volk, M.; Spirason, N.; Halpin, R.A. Multiplex reverse transcription-PCR for simultaneous surveillance of influenza A and B viruses. Journal of Clinical Microbiology 2017, 55, 3492–3501. [Google Scholar] [CrossRef]
- Dziąbowska, K.; Czaczyk, E.; Nidzworski, D. Detection methods of human and animal influenza virus—current trends. Biosensors 2018, 8, 94. [Google Scholar] [CrossRef]
- Lin, X.; Liu, X.-Y.; Zhang, B.; Qin, A.-Q.; Hui, K.-M.; Shi, K.; Liu, Y.; Gabriel, D.; Li, X.J. A rapid influenza diagnostic test based on detection of viral neuraminidase activity. Scientific Reports 2022, 12, 505. [Google Scholar] [CrossRef] [PubMed]
- Hľasová, Z.; Košík, I.; Ondrejovič, M.; Miertuš, S.; Katrlík, J. Methods and current trends in determination of neuraminidase activity and evaluation of neuraminidase inhibitors. Critical Reviews in Analytical Chemistry 2019, 49, 350–367. [Google Scholar] [CrossRef] [PubMed]
- Faye, O.; Faye, O.; Dupressoir, A.; Weidmann, M.; Ndiaye, M.; Sall, A.A. One-step RT-PCR for detection of Zika virus. Journal of Clinical Virology 2008, 43, 96–101. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The figure depicts an enveloped influenza A virus, with each viral protein encoded by eight separate segments of single-stranded RNA. The viral RNA segments and polymerase produce viral ribonucleoprotein by binding to nucleoproteins (vRNP). The viral RNA-dependent RNA polymerases, including polymerase acid (PA), polymerase basic 1 (PB1), and polymerase basic 2 (PB2), are responsible for replication. Two glycoproteins, hemagglutinin (HA) and neuraminidase (NA) bind to the viral surface. Neuraminidase facilitates the virus's escape from infected cells, while hemagglutinin mediates viral entry into host cells. The matrix protein (M1) forms a coat inside the virus envelope, providing structural stability and organisation to the viral components. Additionally, the membrane protein (M2) serves as a proton ion channel, contributing to the acidification of the virus inside the host cell endosome.
Figure 1.
The figure depicts an enveloped influenza A virus, with each viral protein encoded by eight separate segments of single-stranded RNA. The viral RNA segments and polymerase produce viral ribonucleoprotein by binding to nucleoproteins (vRNP). The viral RNA-dependent RNA polymerases, including polymerase acid (PA), polymerase basic 1 (PB1), and polymerase basic 2 (PB2), are responsible for replication. Two glycoproteins, hemagglutinin (HA) and neuraminidase (NA) bind to the viral surface. Neuraminidase facilitates the virus's escape from infected cells, while hemagglutinin mediates viral entry into host cells. The matrix protein (M1) forms a coat inside the virus envelope, providing structural stability and organisation to the viral components. Additionally, the membrane protein (M2) serves as a proton ion channel, contributing to the acidification of the virus inside the host cell endosome.
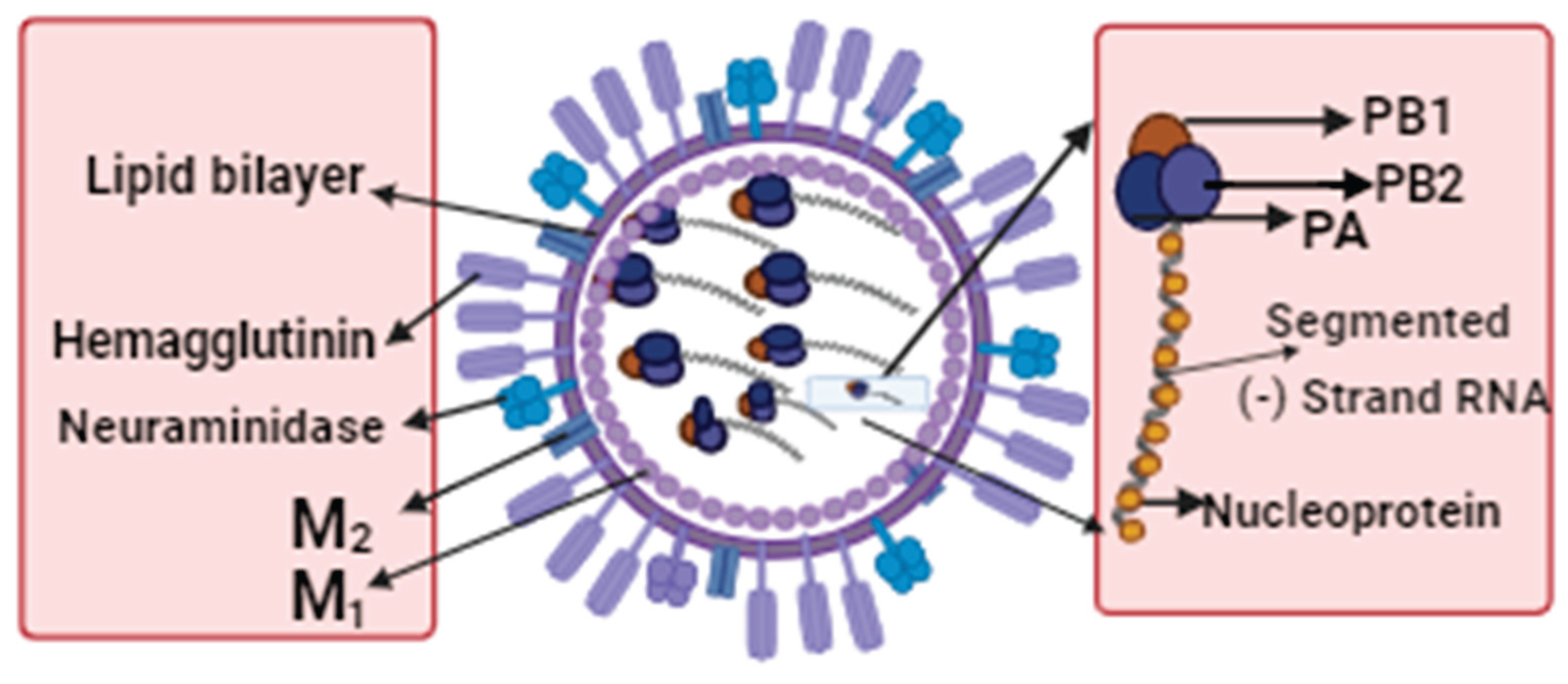
Figure 2.
Pathophysiological progression of H3N2 Influenza virus infection. (a) Initiation of H3N2 Influenza virus infection, (b) Penetration of H3N2 virus into human lung, (c) Intracellular replication of the virus within respiratory epithelial cells, (d) Dissemination of H3N2 variant throughout the tissue, (e) Vascular impairment due to virus-induced damage, (f) Induction of TNF-alpha and pro-inflammatory cytokines, such as Interleukin-6 and Interleukin-8, and (g) Lung infection with H3N2 virus.
Figure 2.
Pathophysiological progression of H3N2 Influenza virus infection. (a) Initiation of H3N2 Influenza virus infection, (b) Penetration of H3N2 virus into human lung, (c) Intracellular replication of the virus within respiratory epithelial cells, (d) Dissemination of H3N2 variant throughout the tissue, (e) Vascular impairment due to virus-induced damage, (f) Induction of TNF-alpha and pro-inflammatory cytokines, such as Interleukin-6 and Interleukin-8, and (g) Lung infection with H3N2 virus.
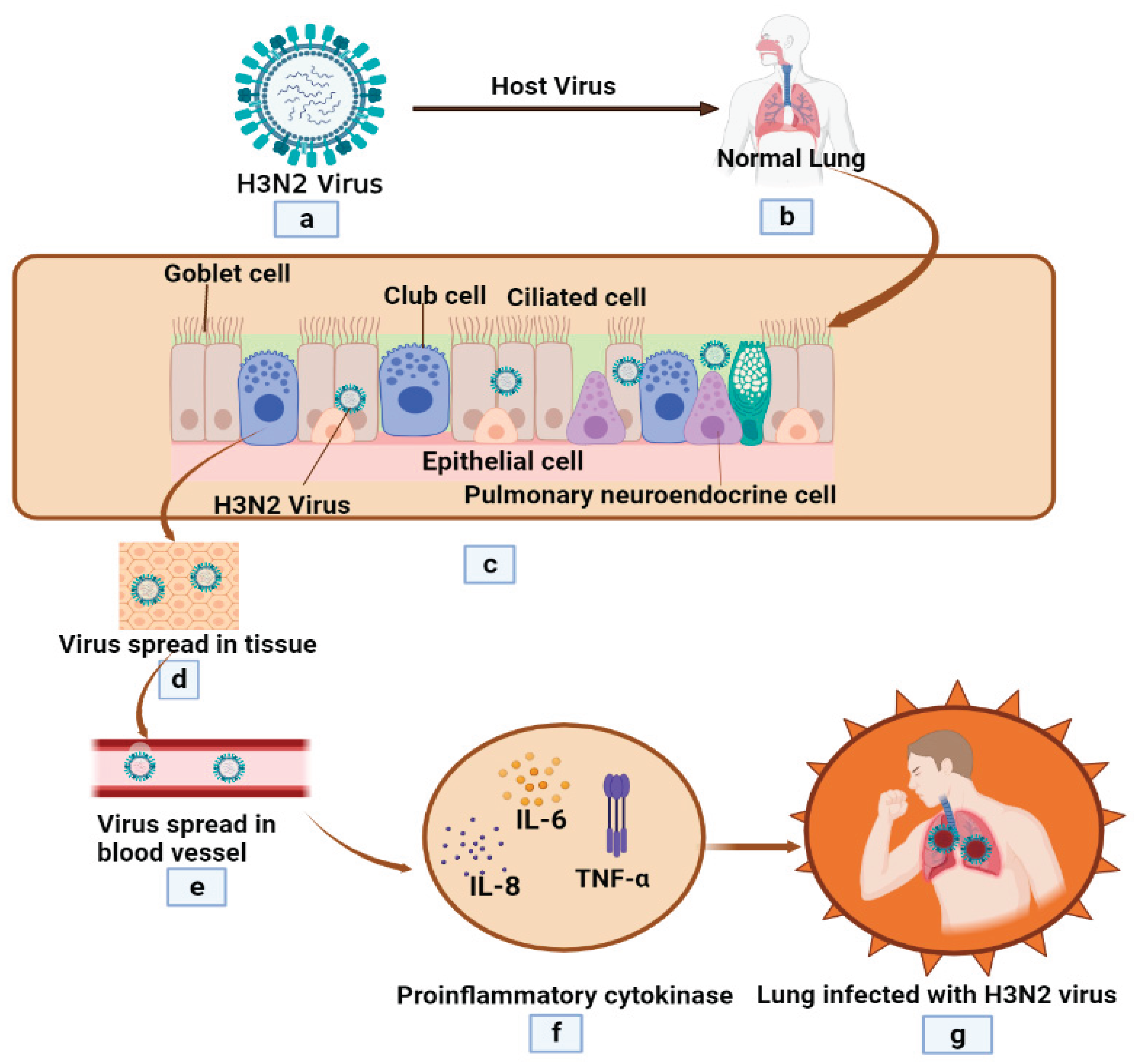
Figure 3.
Mechanism of virus entry (A) Neuraminidase Evolution and (B) Neuraminidase inhibitors.
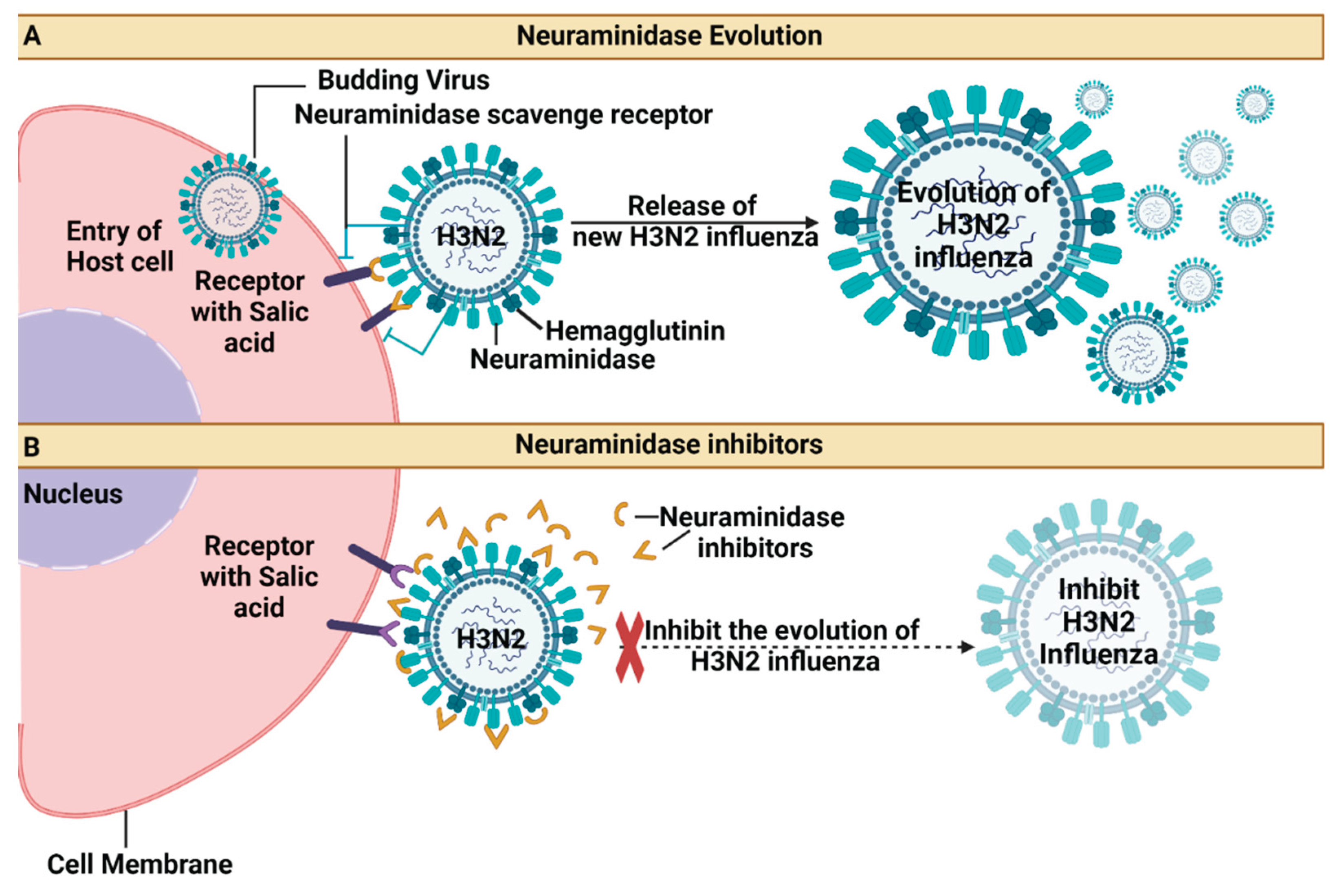

Table 1.
Tracking the Impact: Yearly Incidences of H3N2 Human Infections Worldwide.
| # | Year | Event Description | Impact | Ref. |
|---|---|---|---|---|
| 1 | 1968-1970 | The first influenza A/H3N2 pandemic season (1968/1969) it led to significant US mortality, while the second (1969/1970) caused most deaths in England. This reveals a global mortality pattern. | Mortality patterns in Europe and Asia were delayed until the second pandemic season due to higher neuraminidase immunity and a drift in the antigen during 1969/1970. | [29] |
| 2 | 1970-2006 | It has been discovered that pandemic influenza viruses persist in Chinese pigs as intermediate hosts, with triple-, double-, and entirely human-like H3N2 viruses coexisting. |
This study analyses eight H3N2 virus genes from 1970 to 2006, revealing that pigs serve as mixing vessels for the virus generation. The coexistence of these viruses underscores the importance of reinforcing swine influenza virus surveillance in China. | [30] |
| 3 | 1976 | Eight segments of the Singapore (H2N2) strain's RNA have been tagged, divided, and associated with proteins and gene functions. The base sequence homology between the H2N2 virus and several influenza A strains was discovered using molecular hybridisation. | The Singapore strain exhibits a base sequence homology of about 160% compared to the FM1 strain (HlNl). Still, the H3N2 strain is likely descended from an H2N2 subtype, as evidenced by its retention of four segments and the HA gene from a different strain. | [31] |
| 4 | 2000-2010 |
This work recreated the ecological and evolutionary dynamics of influenza using a host metapopulation representative of the tropical, temperate, and southern regions. |
Results showed that a region's primary reproductive number significantly impacts the antigenic evolution of its viral population and the probability of its strains spreading globally. Seasonality increases the probability of tropical populations exporting evolutionarily successful strains but doesn't predict their antigenically advanced status alone. | [32] |
| 5 | 2009-2011 | Concerns have been raised about the possibility of a pandemic brought on by four A(H3N2) v influenza viruses that were isolated from US people and examined in a study. It was discovered that the viruses may effectively propagate among ferrets living together and infect newly acquired ferrets via respiratory droplets. |
The study found that A(H3N2) v viruses replicated in Calu-3 cells at considerably higher levels than the usual seasonal H3N2 influenza viruses, highlighting the significance of continuous public health surveillance. |
[33] |
| 6 | 2009-2011 | During a phylogenetic analysis of influenza viruses from swine and humans in North America, thirty-four rH3N2p viruses with identical H3, N2, and pM segments to the human-identified H3N2v viruses were found. |
Combination events between H3N2 viruses and the pM segment have produced these viruses about four to ten times since 2009. All H3N2v viruses recovered from humans have an N2 segment originating from a genetically unique N2 lineage, which may affect the development of influenza vaccines and the possibility of pandemics. | [34] |
| 7 | 2010-2012 | Recent US cases of H3N2v influenza infection, primarily among children, are being studied to determine cross-reactive antibody levels and whether seasonal TIV may increase seroprotection. | While teenagers and young adults have cross-reactive H3N2v antibodies, children and older individuals are susceptible. The lack of seroprotection in recent TIV formulations makes a particular vaccine necessary to spread the epidemic. |
[35] |
Table 2.
The effectiveness of the H3N2 influenza vaccine, incorporating relevant data for analysis and comparison.
Table 2.
The effectiveness of the H3N2 influenza vaccine, incorporating relevant data for analysis and comparison.
| # | Population | Study Design | Vaccine formulation | Vaccine effectiveness % | Key finding | Ref. |
|---|---|---|---|---|---|---|
| 1 | Paediatric (2-17 Years) | Observational studies | Trivalent inactivated vaccine | Efficacy of vaccination, 5%; 95% confidence interval, -47 to 39 | Between 2015 and 2016, influenza vaccinations dramatically decreased the likelihood of contracting the illness. | [49] |
| 2 | Adults in the 20–364 age range |
Meta-analysis of TND studies | Trivalent influenza | VE of 65% | According to the ADH, the impact of recurrent influenza vaccination may have contributed to the low VE in recent A(H3N2) epidemics in Canada since 2010. | [50] |
| 3 | General Population | test-negative design and observational studies | trivalent vaccine | VE was 59% | According to the study, the recently developed A(H1N1)pdm09 vaccine offered reasonable defence against circulating strains. However, VE against A(H3N2) was less than 35% in 2016–17 and 2017–18, presumably due to the antigenic mismatch obtained from egg multiplication. | [51] |
| 4 | Aged one year and above. | Test-negative design | vaccine strain uses egg-adaptation mutations | In 2016–17 and even lower in 2017–18, VE against A(H3N2) was below 40%. |
The study suggests that VE, influenced by phylogenetic sub-clusters and vaccination history, exhibits informative heterogeneity. However, it requires larger sample sizes and may be linked to pivotal mutations. | [52] |
| 5 | Age group is greater than 65-79 years. | test-negative design | trivalent vaccine | Influenza A(H3N2) IVE was 24%, while B IVE was 30%, 37%, and 19%. | IVE against influenza B in hospitalised older adults is similar to A(H3N2), highlighting the importance of influenza vaccination. | [53] |
| 6 | Aged 65 and above. | test-negative case-control design | trivalent influenza vaccine | The adjusted VE for inpatients was 7.4%, while outpatients had 19.3%. | Denmark experienced multiple genetically drifted H3N2 viruses during the 2016-17 influenza season, with low estimated VE and varying VEs across four main virus clusters. | [54] |
| 7 | Aged one year and older. | test-negative design | - | The vector error (VE) for Canada's influenza A(H3N2) outbreak in 2016–17 is over forty per cent higher than in 2014–15. | The intermediate vector error (VE) is approximately 40% higher in Canada's 2016–17 influenza A(H3N2) epidemic than in the 2014–15 pandemic. To reduce morbidity and death, particularly in high-risk individuals, further steps are required. |
[55] |
Table 3.
The diagnostic methods of the H3N2 influenza virus, highlighting their descriptions.
| # | Method | Description | Ref. |
|---|---|---|---|
| 1. | Biosensors | Advanced methods to detect H3N2 influenza viruses based on different parameters, aiming to improve specificity and sensitivity. The human influenza virus binds to α, 2–6 glycosidic bonds, while the avian influenza virus binds to α, 2-3 glycosidic bonds. Viruses detect distinct receptors on host cells. Pigs show both genetic re-assortment and antigenic shift since they have both types. | [87,102] |
| 2. | RT-LAMP stands for Reverse Transcription Loop-Mediated Isothermal Amplification. | Molecular diagnostic tool for influenza A viruses, including H3N2. It is quick, easy, cost-effective, sensitive, and specific, suitable for point-of-care testing during outbreaks. The process involves amplifying nucleic acid using reverse transcriptase, DNA polymerase, and oligos, resulting in double-stranded looped DNA structures that can be detected using pH sensitivity, fluorescent response, and turbidity. | [103,104] |
| 3. | Multiplex PCR (Polymerase Chain Reaction). |
The diagnostic tool uses several primer pairs in the same reaction, amplifying different specific amplicons for various targets. Increasingly used for the diagnosis of infectious diseases, including RNA-containing viruses like H3N2 influenza. Using this technique, different influenza viruses' HA, NA, and M gene segments can be amplified simultaneously from clinical specimens or isolates to be sequenced. |
[105,106] |
| 4. | Rapid Influenza Diagnostic Tests, or RIDTs | Rapid influenza tests can detect viral nucleoprotein antigens in respiratory specimens in less than 15 minutes; commercial and laboratory-developed RT-PCR assays are suitable reference tests. | [85,107] |
| 5. | Neuraminidase Activity-Based Assay | A chemiluminescent assay for detecting influenza viral neuraminidase (NA) activity utilises a unique substrate related to NA, a target for newer-generation influenza therapeutic drugs. The main goal of mechanism-based drug design is to locate and create target enzyme competitive inhibitors. | [108,109] |
| 6. | one- step RT-PCR | The one-step RT-PCR assay proved quicker and easier than virus isolation and serological methods. | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



