
Submitted:
10 April 2024
Posted:
29 April 2024
You are already at the latest version
Abstract
Depression marsh intra- and interspecific facilitation in Southeast Florida, dominated by Hypericum fasciculatum, Stillingia aquatica, and Rhynchospora tracyi were studied as a third installment to the author’s ongoing interest in “Hypericum-Stillingia (HS) marsh” vegetation. The project includes a contextual overview in relation to the HS marsh environmental filter, stress-gradient-hypothesis, and harsh-habitat trait convergence vs. divergence. Using field measurements, transects, and quadrats, the effort focused mainly on two research questions: 1. Does the Hypericum fasciculatum microdistribution relate to marsh microtopography? 2. Does H. fasciculatum have positive or negative associations with other species? Tempered by “cause and effect” concerns, the results supported positive feedback autogenic site alteration by H. fasciculatum along with interspecific facilitation.
Keywords:
freshwater marsh
; microtopography
; limiting similarity
; environmental filter
; facilitation
; nurse plants
; autogenic site alteration
; environmental switch
1. Introduction
Portions of Florida are dotted with thousands of shallow depression wetlands ranging from meters to kilometers in diameter. Their geologic origins vary, those to the north reflecting karst solution pockets, those in south Florida more likely drainage-impeded dips in the varied sandy-clay substrates [1]. Inputs and outputs are mostly rain and evaporation, with unclear connections to underlying hydrology. The water is usually acid, nutrient poor if unpolluted, and nutritionally phosphorus-limited [1]. Depths and hydroperiods vary substantially in time and space. Some depressions are year-round lakes or ponds. The shallowest and best-drained are wet meadows or prairies. The nomenclature applied to Florida shallow wetlands is confusingly inconsistent. The depression marshes in the present project are of intermediate depths, often covering vast areas with water often more or less uniformly deep. With annual variation, the low-rain season extends from late winter to late spring, with peak rain June to September. The marshes of present interest spend time with their surfaces exposed, although with a lag between drying weather and exposure. Submersion often exceeds exposed time, and the subsurface soil typically remains moist during the drier season. Fire may occur some years during dry periods [1].
The Southeast Florida study area marshes (Figure 1a) are peppered abundantly by two woody shrubs 0.5-1.5 m tall, generally found sympatrically in our area: Hypericum fasciculatum (Clusiaceae, Figure 1b) and Stillingia aquatica (Euphorbiaceae, Figure 1c). Freshwater marshlands dominated by this pair define the study habitat, here called “Hypericum-Stillingia [HS] marshes.” (In wetlands outside the study area additional shrubby Hypericum species are abundant to dominant.) Appearing in varied proportions, the two dominant herbaceous species are Rhynchospora harperi (1d, e) and R. tracyi (Figure 1f). As Kral [2] observed,” Rhynchospora harperi is most abundant in a very special habitat referred to here as the “Hypericum pond.” Also sometimes encountered, mostly on exposed wet marsh bottom, is R. divergens, a usually tiny, grassy species having capillary leaves. Xyris smalliana is abundant in the study area, and young individuals are often physically adjacent to the H. fasciculatum trunk. Weakley et al. [3] noted X. smalliana as “adjacent” to Hypericum (and to Pontederia) in the intermediate zones of basin marshes in peninsular Florida. And long before that, Kral [4] mentioned X. smalliana in “Hypericum ponds.”
The HS marsh is a harsh habitat. Establishment entails all life phases surviving the alternating flooding-drying environmental filter exacerbated by oligotrophy, periphyton blanket, full exposure to wind and subtropical sun, and perhaps occasional fire. For these reasons, the system prompts attention to the question of character trait convergence vs. divergence in the face of a strong filter, discussed below. It also intersects with the stress-gradient-hypothesis (SGH), which suggests a shift in interactive balance from competition to facilitation along increasingly stressful environmental gradients [5,6]. Accordingly, facilitation is of special concern in exploring the challenging HS marsh ecology.
The present project is a continuation of the author’s extended interest in the local HS marshes, with prior individual attention to H. fasciculatum and S. aquatica, largely their reproductive biology [7,8]. The third installment, here based mostly on field measurements, takes a broader look with a special eye to autogenic site alteration and to intra- and interspecific facilitation. Aided by improving technology, there has been recent interest in autogenic environmental heterogeneity [9,10]. That such recent work embraces positive feedback (aka “switches”) [11,12] in marshes prompted the present research question: beyond stochastic competition and generalized growth and decay, is there detectable plant-mediated habitat alteration? And if so, does that provide intraspecific positive feedback and/or interspecific facilitation?
As the main goal of the present project, this compound question was addressed by field measurements centered chiefly on Hypericum fasciculatum and its context. Data-acquisition included two components. First, two rounds of microtopographic depth measurements. Second, a series of near-neighbor, nearest-neighbor, photographic, and transect and quadrat studies summarized in Table 1. The project began with a contextual marsh overview of the local HS marsh community before narrowing to the stated research questions.
2. Materials and Methods
Species are listed in Appendix A. Authorships and families, given there, are not repeated in the text. Taxonomic details generally follow Wunderlin et al. [13]. The study sites were two depression marsh systems in Palm Beach County, Florida. The “Mack Dairy Rd.” marsh (N 26.949417, W -80.222182) in the Cypress Creek Natural Area is a small (150 m diam) HS marsh surrounded by wet pinewoods. The “Casuarina” HS marsh (Figure 1a, named informally for a landmark stand of trees) was the primary study zone. It is multiple kilometers in diameter within the Florida Fish and Wildlife Conservation Commission John C. and Mariana Jones/Hungryland Wildlife and Environmental Area and adjoining Pal-Mar Natural Area, separated by an E-W road. Most of the work was within Casuarina. Data were collected at four subareas referred to here as: Casuarina-South (N 26.929162, W -80.288498), Casuarina-North (26.939811, -80.289019), Casuarina-West (26.941800, -80.308328), and Casuarina-East (26.943650, -80.291711, Figure 1a). The study period was November through March 2023/2024. Water depths were maximum early in the project to off-peak yet still submerged for almost all of the study. Most of the work was in standing water (8)20(30) cm deep. Weather ranged from heavily rainy in November through much of January to partly drying unevenly through February, and mostly dry in March, with marsh bottom partly exposed in the final days of the study. State and county research permits prohibited destructive sampling. Transect placements were restricted to minimally invasive access by existing vehicular and animal trails from marsh access points. (Transects paralleled trails, and were not within them.) During the study period, the road providing access to Casuarina-North and -South was closed unexpectedly, forcing relocation mostly to Casuarina-East.
Graphics and calculations were prepared using R [14] employing the Tidyverse package. The dendrogram was generated using the R dist (binary) and hclust (as.dendrogram) methods applied to Supplementary Materials “SM” dataset J available in [15]. All field data tables are in that repository. Species abbreviations used in the graphics are given in the caption to Figure 2.
A pilot study tabulated the vegetation pattern along the depth gradient at 16 different local HS marsh sites. Diversity was highest near shores, intermediate at medium depths (used for most of the present resulting project), and diminished in permanent or nearly permanent standing water. In the pilot study, non-dominant species varied considerably in presence and abundance, even in sites with generally similar appearances and water levels. The pilot results were not incorporated directly into the present analysis; they are, nonetheless, available as SM dataset G [15]. Multiple additional datasets for the project were compiled using different combinations of transects and quadrats, depending on specific study questions, and on changing circumstances [15]. There is no intermixing of datasets, except juxtaposition of the Mack Rd. and Casuarina rank abundance graphs based on the same methodologies. Methods used for compiling each dataset were as follows:
Substudy 1. Species abundances. Species abundances were determined at the Mack Dairy Rd. marsh and at the Casuarina-North site using 30 quadrat placements at both marshes during December and January 2023/2024. The 40 cm x 60 cm quadrat dimensions match the aspect ratio and mounting height of a camera used to photograph quadrats. Quadrat placements in this substudy were blind over-the-shoulder quadrat tosses following a depth contour of ca. 20 cm along a total distance of 60 m. “Individuals” (including ramets) rooted within the frame were counted for each placement, including those with root crowns overlapping the inner frame edge (SM E, F).
Substudy 2. Qualitative characteristics. In order to discuss functional traits, qualitative information was tabulated for the common species (SM I) by direct field examination, by on-line examination of herbarium specimens at the University of South Florida (USF), and by literature review. To constrain subjectivity, the list of traits designated as key to surviving the marsh environmental filter came from outside sources: xeromorphy in seasonal marsh plants [16] and from Keddy’s [17] general characteristics of wetland and marsh species: aerenchyma, rhizomes, pointed shoots, and low protected growing tips. Stillingia aquatica resprouts from latent buds near the highwater line after top damage, and thus was included in the protected buds category. (Such sprouting happens in H. fasciculatum but only rarely, and was not included.) Xeromorphy was here taken to be one or more of the following traits: reduced photosynthetic surfaces, terete or nearly terete (including narrowly channeled) main photosynthetic structures, upright primary photosynthetic surfaces, subsucculence, and thick or waxy covering (e.g., Dichanthelium erectifolium). The last-mentioned grass makes broad leaves at the marsh bottom when submerged, and on its rising culm forms erect narrow leaves. Although Eriocaulon compressum has a soft, lax spreading rosette when submerged, its leaves are upright and relatively stiff on exposed soil. Also with foliar dimorphy, Hypericum fasciculatum has broad leaves on young plants as opposed to the needle leaves on mature individuals. Rhynchospora harperi makes small, grassy, thin leaves when submerged or partly submerged (and can be fertile then), and makes much larger leaves with age and emersion (Figure 1d).
Substudy 3. Herbaceous species nearest-neighbors. As a check on interspecific associations among herbaceous plants, at Casuarina-North a 100-m string transect was extended along the ca. 20-cm depth line. The transect was sampled for the six most abundant nondominant herbaceous species at that locality (Aristida palustris, Dichanthelium erectifolium, Eriocaulon compressum, Panicum tenerum, Rhynchospora harperi, Xyris smalliana). Sampling points were determined by the presence within two m of the transect of Panicum tenerum due to it being by far the least abundant species. Each of the other species nearest each P. tenerum transect-encounter had its nearest nonconspecific neighbor recorded (SM A).
Substudy 4. Woody species neighbors. Seeking interspecific associations for H. fasciculatum and for S. aquatica, nearest neighbors to Hypericum fasciculatum, to Stillingia aquatica, and to a randomly repeatedly placed fiberglass “null” vertical pole were recorded at Casuarina-West. For this a 60-m study strip two m wide extended parallel to shore in standing water. Nearest neighbors were tabulated for each non-seedling Hypericum fasciculatum, Stillingia aquatica, and pole location, 30 samples each. The pole was placed using random X-Y coordinates within the full width and length of the study strip (SM H). In a related separate tabulation to permit reappraisal, at Casuarina-South all neighbors within 10 cm of H. fasciculatum, S. aquatica, and the randomly placed pole, 30 each, were tabulated (SM B1-B3).
Substudy 5. Hypericum fasciculatum and marsh microtopography. To determine if the H. fasciculatum microdistribution is related to soil elevation beyond the root cone, 30 sequential individuals (trunks > 9 cm diam 20 cm above marsh bottom) were measured along a trail parallel to shore at Casuarina-East. Water depths were recorded at 0, 10, 20, 30, 40, 50, and 60 cm from each H. fasciculatum trunk base along the N,S,E,W radii with deviations when a radius was physically blocked. Individuals were skipped if situated among crowded conspecifics (SM C1).
For a separate appraisal of the same question, and for comparison with Stillingia aquatica, along a new linear transect parallel to shore in standing water at Casuarina-East water depths were recorded at 5 cm intervals, as were the locations of all H. fasciculatum and S. aquatica individuals touching the string. (In spots where a depth reading would occur at a H. fasciculatum root cone, in order to avoid the distortion of measuring depth to the cone top, the two readings on either side of the cone were averaged (SM C2)).
Substudy 6. Hypericum fasciculatum and interspecific facilitation. Association of H. fasciculatum with microtopographical elevations in substudy 5 raised the question of possible facilitation of herbaceous vegetation on the same microelevations. As a first step, the Casuarina-East site was searched for visual patterns of sedge (mostly R. tracyi, R. harperi, Scleria baldwinii) height and/or density association with H. fasciculatum, with examples photographed. To quantify that relationship, a transect was stretched parallel to shore in standing water for approx. 23 m at Casuarina-East. The full length of the transect was tallied for all sedge clump bases (or isolated ramets) in 463 5 X 5 cm grid squares arranged serially along the string with no gaps except the wire grid margins (SM D).
As a separate reappraisal of the relationship between sedge density and distance from H. fasciculatum, a second study was conducted at Casuarina-East. Rather than follow a transect, the reappraisal study tabulated 30 individuals of H. fasciculatum (over 9 cm diam at 20 cm above the base). The H. fasciculatum shrubs were selected using a spinner. A single radius leading outward from each selected H. fasciculatum trunk was then chosen, again by spinner (redone if the radius was physically blocked or within ca. 1 m of a different H. fasciculatum). For each H. fasciculatum the radial samples consisted of 15 serially adjacent 5 X 5 cm wire grid squares proceeding outward from the trunk base, the numbers of sedge bases or isolated ramets in each square recorded. Because the result echoed closely the transect study, the second data table and resulting plot are not included here, and are in the SM as “D reappraisal.”

Table 1.
Summary of the substudies. Cas = Casuarina site Mack = Mack Dairy Rd. site. SM = Supplementary Materials. NN = nearest neighbor.
Table 1.
Summary of the substudies. Cas = Casuarina site Mack = Mack Dairy Rd. site. SM = Supplementary Materials. NN = nearest neighbor.
| Substudies | Sites and dates | Methods | Data access | Questions addressed |
Summarized results | Related graphics |
| 1 | Cas-N and Mack 12/9/23-1/15/24 |
Quadrats along 20-cm depth contours | SM: E, F | Inventory | Species compositions differ between similar sites | Figure 2 |
| 2 | All sites, plus literature & online herbarium |
Qualitative trait assessments | SM: I, J | Convergence and divergence of plant traits in relation to alpha and beta niches |
Convergence in beta niche, divergence in alpha niche | Figure 3b (Figure 3c based on SM J) |
| 3 | Cas-N 1/3-1/11/24 |
Transect along ca. 20-cm depth contour | SM: A | Exploratory, seeking herbaceous species NN patterns | Inconsequential. | |
| 4 | Data B1-B3: Cas -S 2/1-2/3 2024 Data H: Cas-W 2/6-2/8 2024 |
60 m X 2 m study strip 100 m X 2 m study strip |
SM: B1-B3 SM: H |
B1-B3: all neighbors within 10 cm of H. fasciculatum, of S. aquatica, and of random pole H: NN to H. fasciculatum, to S. aquatica, and to random pole |
Hypericum fasciculatum had more close neighbor species than S. aquatica or random pole, with R. harperi disproportionately as NN, at high water |
Figures 4b, 6 |
| 5 | Cas-E 2/19-2/25 2024 |
Transect parallel to shore for depths around bases (C1), same transect measured at 5 cm intervals for depth profile and shrub placements (C2) |
SM: C1, C2 |
Does H. fasciculatum placement reflect microtopography? | On average H. fasciculatum had soil raised around its base to ca. 50 cm radius (C1), and H. fasciculatum tended to be on raised marsh bottom (C2) | Figure 7a , b |
| 6 | Cas-E 2/26-3/4 2024 |
For Cyperaceae, transect with 5 cm square micro- quadrats |
SM: D | Does H. fasciculatum influence vegetation beyond its adjacent small neighbors? | H. fasciculatum appears to influence neighbors out to over 30 cm | Figure 7c, also “D reappraisal” data and plot in SM |
3. Results
3.1. Marsh Overview
A substantial mat of periphyton and/or suspended organic debris was almost constantly present. Periphyton can tie up and redistribute nutrients [18]. Tests for orthophosphate (EPA650 Capillary Ion Electrophoresis) in the Mack Dairy Rd. and Casuarina sites near high water in January 2024 yielded no detectable results for open water. The same occurred at Casuarina for nitrite and nitrate using Industrial Test Systems 480009 test strips in open water, mud, and H. fasciculatum root crowns. At different times and places the periphyton mat can have widely varying composition and appearance: almost pure filamentous green algae (e.g., Mougeotia sp.), or a matrix of floating Utricularia with clinging algae, or a slurry of organic debris, fungi, protists, nematodes, diatoms, green algae, and Cyanobacteria. During submersion the mat plus undecayed dead foliage is thick enough to impede light transmission to the marsh floor. During dry times the mat coats surfaces as a sodden or crisp crust. The marsh bottom is sand with a variable usually thin organic layer sometimes loose and thin enough to wash away during water movement to expose the sand marsh floor, with sediments to several cm deep collecting in other places.
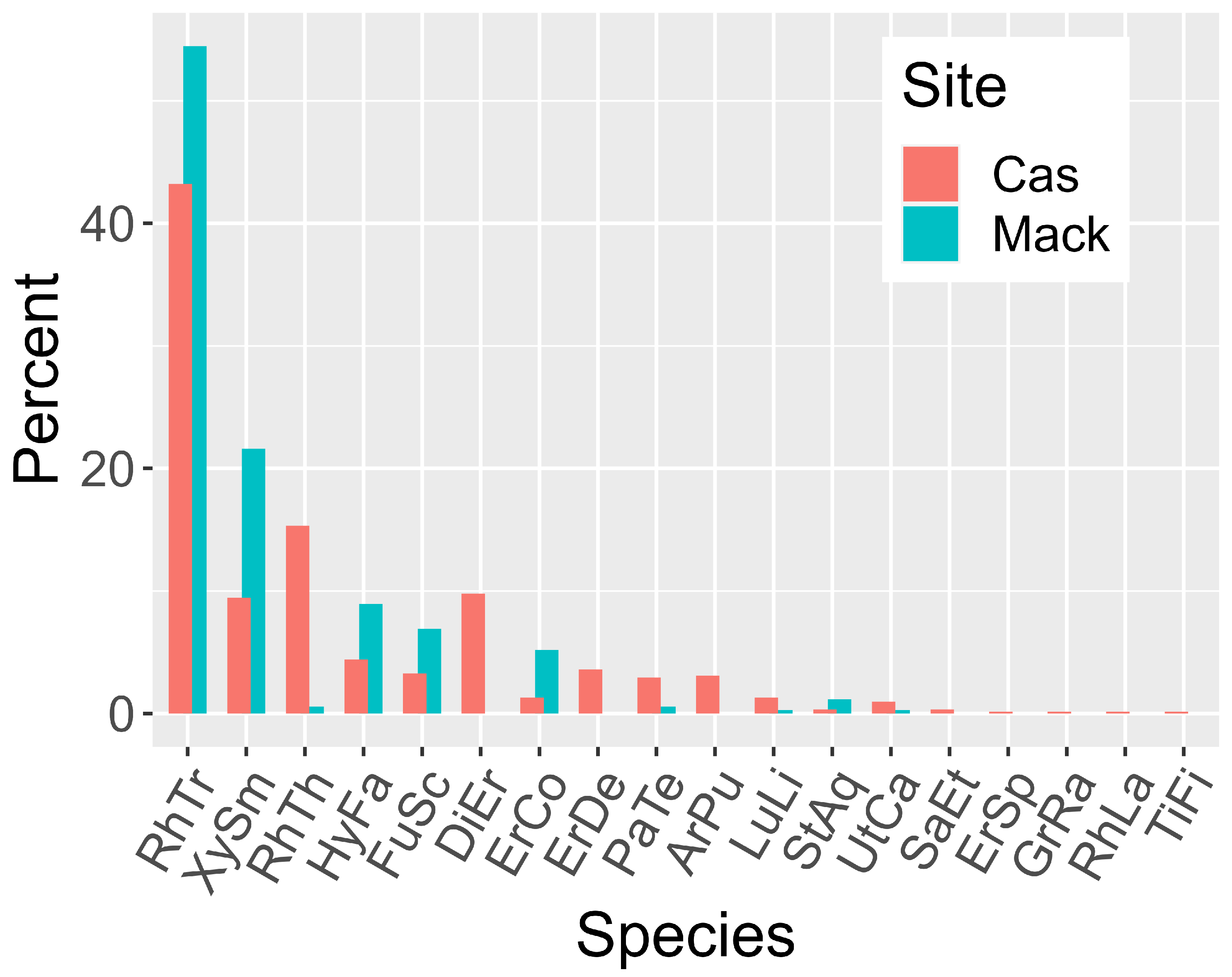
3.2. Substudy 1.
To assess the local HS marsh as an environmental filter using the Wunderlin et al. database [13], the number of native vascular plants in Palm Beach County is 868; the obligate wetland species count is 138; my pilot study of 16 marshes (10 cm, 30 cm, and more or less permanent water) turned up 77 species. The master list for the present project (Appendix A) amounts to 34 species, and the quadrat studies at Casuarina-North and Mack Dairy Rd. (Figure 2) gave 18 species. Beyond the two dominant shrubs and Rhynchospora species, often-abundant species were Aristida palustris, Dichanthelium erectifolium, Eriocaulon compressum, E. decangulare, Fuirena scirpoidea, Utricularia (mostly U. purpurea), and Xyris smalliana. At these and additional study sites, also often present, if spotty, were Bacopa caroliniana, Centella asiatica, Proserpinaca pectinata, Rhynchospora divergens, Scleria baldwinii, Tiedemannia filiformis, and additional, unusual, species, especially near shores.
Figure 2.
A. Rank abundances at Casuarina-North and Mack Dairy Rd. sites. Left bars=Casuarina-N site. Right bars = Mack Dairy Rd. marsh. Species abbreviations for this and other graphics: ArPu Aristida palustris, BaCa Bacopa caroliniana, DiEr Dichanthelium erectifolium, ErCo Eriocaulon compressum, ErDe E. decangulare, ErSp Eragrostis spectabilis, FuSc Fuirena scirpoidea, GrRa Gratiola ramosa, HyFa Hypericum fasciculatum, LuLi Ludwigia linifolia, PaTe Panicum tenerum (Coleataenia tenera), PrPe Proserpinaca pectinata, RhDi Rhynchospora divergens, RhLa R. latifolia, RhTh R. harperi, RhTr R. tracyi, SaEt Samolus ebracteatus, ScBa Scleria baldwinii, StAq Stillingia aquatica, TiFi Tiedemannia filiformis, UtCa Utricularia purpurea, XySm Xyris smalliana.
Figure 2.
A. Rank abundances at Casuarina-North and Mack Dairy Rd. sites. Left bars=Casuarina-N site. Right bars = Mack Dairy Rd. marsh. Species abbreviations for this and other graphics: ArPu Aristida palustris, BaCa Bacopa caroliniana, DiEr Dichanthelium erectifolium, ErCo Eriocaulon compressum, ErDe E. decangulare, ErSp Eragrostis spectabilis, FuSc Fuirena scirpoidea, GrRa Gratiola ramosa, HyFa Hypericum fasciculatum, LuLi Ludwigia linifolia, PaTe Panicum tenerum (Coleataenia tenera), PrPe Proserpinaca pectinata, RhDi Rhynchospora divergens, RhLa R. latifolia, RhTh R. harperi, RhTr R. tracyi, SaEt Samolus ebracteatus, ScBa Scleria baldwinii, StAq Stillingia aquatica, TiFi Tiedemannia filiformis, UtCa Utricularia purpurea, XySm Xyris smalliana.
3.3. Substudy 2.
There is an apparent qualitative distinction in the study area between convergent “beta niche” traits connected mostly with the physical environment as opposed to divergent biological “alpha niche” traits connected mostly with close-distance competitive resource acquisition. A summary of the tabulation is in Figure 3a, with traits regarded as beta niche the five left-most columns (aerenchyma, xeromorphy, safe (low protected) buds, pointed growing tips, rhizomes). Figure 3b is a dendrogram of qualitative traits sorting the main species into five roughly defined potential functional guilds, showing a considerable phylogenetic component.
3.4. Substudy 3
The nearest-neighbor frequencies of each herbaceous species with respect to the usually dominant R. tracyi almost all reflected the 47% overall representation of R. tracyi. However, only 28% of Aristida palustris had R. tracyi as nearest neighbor. A second minor anomaly was that Panicum tenerum tended to be especially close to its nearest neighbors, 1.54 cm on average, as opposed to 2.62-3.62 cm for the other species.
3.5. Substudy 4
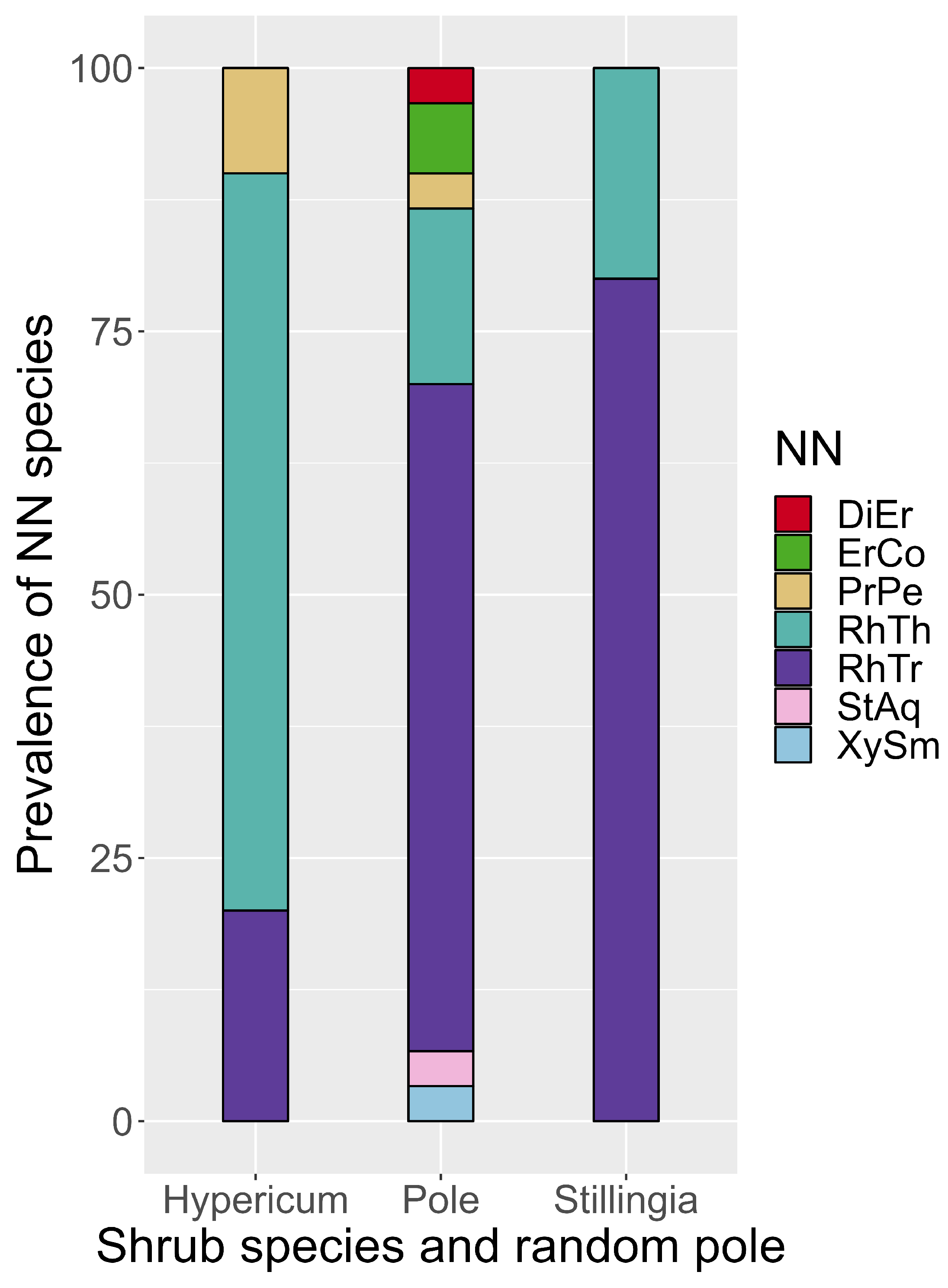
Hypericum fasciculatum had more species as nearest-neighbors at Casuarina-West than did S. aquatica, likewise for the randomly placed “null” pole (SM H). In the similar but separate tally at Casuarina-South for all neighbors within 10 cm, Hypericum. fasciculatum had 19 different neighbor species, S. aquatica and the pole both only 13. The neighbors unique to H. fasciculatum were Burmannia capitata, Gratiola ramosa, Fuirena scirpoidea, Pinus elliottii seedling, Scleria reticularis, and Tiedemannia seedlings. Hypericum fasciculatum had Rhynchospora harperi disproportionately as nearest-neighbor compared with nearest-neighbors to S. aquatica and to the pole (Figure 6, SM B1-B3).
Figure 4.
Hypericum fasciculatum root cones. a. Root cone with detritus. b. Root-trunk junction with young Rhynchospora harperi (left) and Xyris smalliana (right). c and d. Root cones cut open.
Figure 4.
Hypericum fasciculatum root cones. a. Root cone with detritus. b. Root-trunk junction with young Rhynchospora harperi (left) and Xyris smalliana (right). c and d. Root cones cut open.

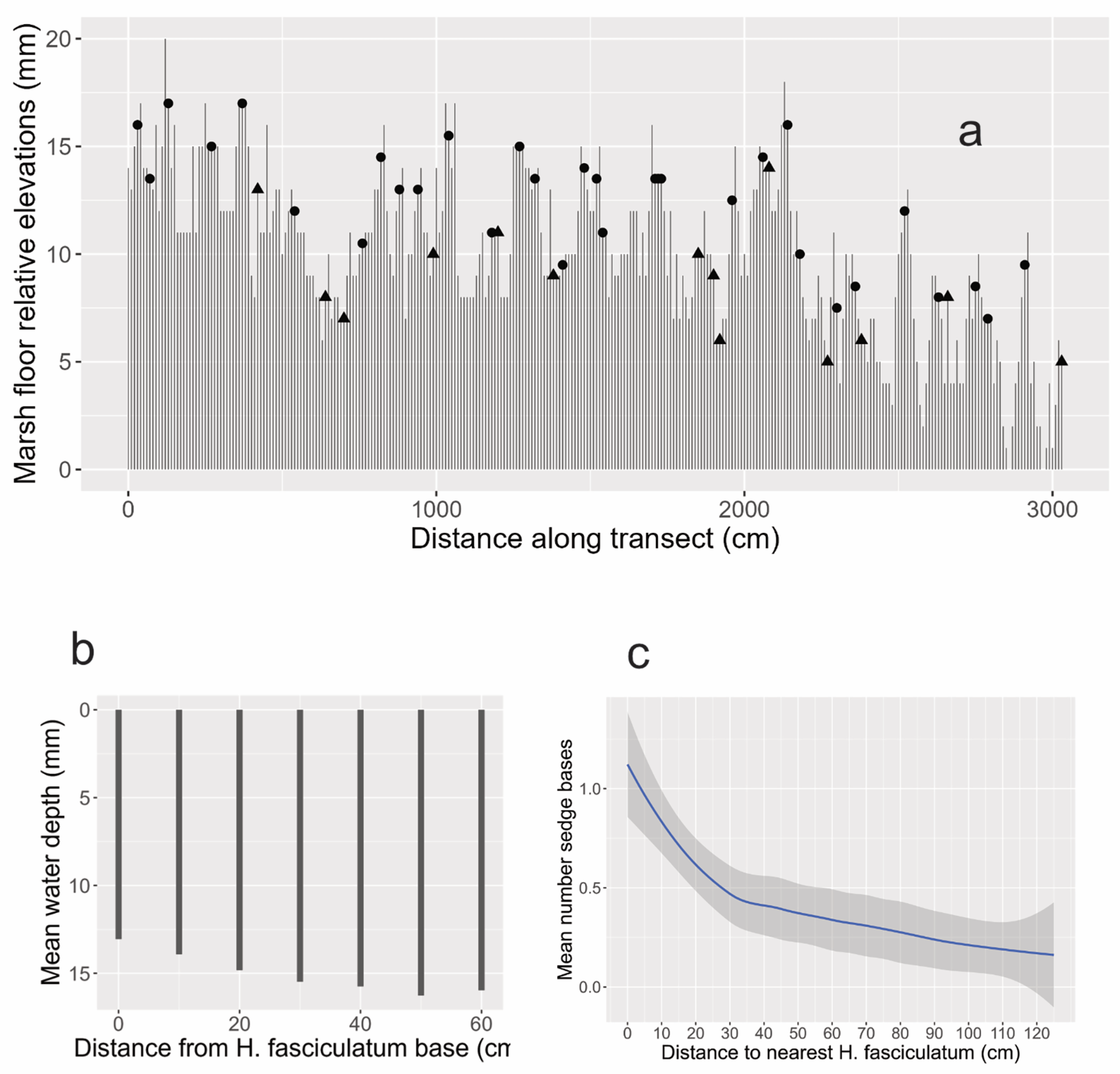
3.6. Substudy 5
A common attribute of wetland plants is adventitious roots, and H. fasciculatum develops these abundantly forming the root cone. The top of its root cone is often several cm above the marsh bottom and near the highwater line, the base where the cone joins the mud is about as wide as the cone is tall. Alternatively, in places the cone can be hidden under soft sediments, and the cone sizes vary. The cone when not sediment-covered, tends to comb organic matter (and propagules) from the water (Figure 4). Beneath large mature individuals having well developed cones accumulates a layer of decaying organic matter mixed with roots. The “soil” in this mass is peaty and different from the basic sandy marsh bottom (Figure 5). The adventitious root system and the new soil around and beneath it rise by having new roots emerge higher on the cone replacing older roots below. The rising root-soil mass creates a raised, porous, detritus-enriched “garden bed” for small plants (Figs. 4b, 6). As an informal test, the dark soil from the cone and light soil from the separate marsh bottom was left in the containers after Figure 5b was taken and kept moist in a screen enclosure. As of this writing approx.. 3 week later, the dark cone soil sprouted 17 seedlings, and the light marsh floor soil sprouted five. The most salient difference is that the dark-soil seedlings include eight young threadlike rhynchosporas, the light soil just one. In addition to the roots making up the root cone, shallow H. fasciculatum cordlike roots can extend a meter or more radially from the trunk base. These sometimes have small erect branch roots [8]. Substudy 5 showed the ground around mature H. fasciculatum to most often to be elevated (Figure 7a), with the marsh bottom on average sloping away from the trunk out to a radius of about 40-50 cm (Figure 7b). (Hypericum fasciculatum seedlings can be on deep marsh bottom or on elevations.)
3.7. Substudy 6
Dense and/or tall sedge growth is sometimes observable around H. fasciculatum (Figure 8). To quantify the relationship, Figure 7c shows elevated sedge density out to about 30 cm, with slight enhancement beyond. (For a second and separate similar study showing essentially the same results see SM “D reappraisal.”)
Figure 7.
a. Transect profile showing water depths and locations of H. fasciculatum and S. aquatica. Dots = H. fasciculatum. Triangles = S. aquatica. Sample points at 5 cm intervals. b. Soil elevation measurements around H. fasciculatum, means in cm. c. Mean numbers of sedge bases per 5 X 5 cm grid squares along transect, vs. distances to nearest H. fasciculatum. C.I. = 95%. For species abbreviations see Figure 2 caption.
Figure 7.
a. Transect profile showing water depths and locations of H. fasciculatum and S. aquatica. Dots = H. fasciculatum. Triangles = S. aquatica. Sample points at 5 cm intervals. b. Soil elevation measurements around H. fasciculatum, means in cm. c. Mean numbers of sedge bases per 5 X 5 cm grid squares along transect, vs. distances to nearest H. fasciculatum. C.I. = 95%. For species abbreviations see Figure 2 caption.
4. Discussion
Keddy and Laughlin [19] observed that competition is often between individual species and a single dominant species, as opposed to chiefly with each other. In the present study rhizomatous R. tracyi is usually the dominant herbaceous species, although sometimes there is abundant (nonrhizomatous) R. harperi. The additional herbaceous species are intermixed in gaps among shrubs and among the large rhynchosporas, as Boutin and Keddy [20] described as typical for other marshes.
The harsh HS marsh environmental filter suggests a balanced expression of two opposing trends in plant traits: As several authors have discussed [12,19,21,22,23], on one hand the abiotic filter hypothetically selects largely for convergence in traits pertaining mostly to surviving harsh physical conditions. On the other hand, the filtered survivors must then compete, promoting limiting similarity and functional trait divergence especially in traits relating chiefly to competitive architecture and to spatial extension [21]. As suggested by Figure 3a, the present HS marsh species met expectations of convergence with respect to the physical environmental filter (aerenchyma, xeromorphy, protected buds, rhizomes) and divergence with respect to short-distance competitive foliar and root diversity.
Nearest-neighbor data among the herbaceous species revealed very little non-stochastic community structure. The particularly low-frequency nearest-neighbor relationship of Aristida palustris with R. tracyi occurred probably because A. palustris has a broad spreading root pattern (or conceivably a preference for shallower microsite). The particularly close proximity of Panicum tenerum (Coleataenia tenera) to its varied nearest neighbors may result from being a thin wispy grass having a tightly knotted rhizome suited to gaps. That species resembles its congener “Panicum longifolium”(Coleataenia longifolia (Torr.) Soreng), which conforms to the “interstitial reed” guild in Boutin and Keddy [20]. Both species have a similar knotty rhizome.
Probably promoting coexistence in the HS marsh are trade-offs between competitive dominance vs. colonization, dispersal, and reproductive pattern [17,24]. Three rhynchosporas may exemplify such tradeoffs. Rhynchospora tracyi is relatively tall and thick, tough, rhizomatous, and nearly dormant aboveground when flooded, becoming green and increasingly fertile as waters recede. By contrast, R. harperi regenerates with small grassy leaves (Figure 4b) and produces flowers even during elevated water levels, both submerged and on relatively elevated spots. Rhynchospora harperi graduates from thin grassy leaves to more substantial foliage (note the mixed leaf sizes in Figure 1d taken as water levels were dropping). While R. tracyi elongates unidimensionally by elongating rhizome growth, R. harperi wanders two-dimensionally by basal budding. Typically much smaller than the other two, R. divergens, aka “spreading beaksedge,” is perennial but inconspicuous during high water, and rises quickly from newly exposed wet substrate.
That large Hypericum fasciculatum accumulates rich, peaty, aerated soil penetrated by its own roots is an example of an autogenic intraspecific positive feedback. Turning to interspecific facilitation, although the stress-gradient-hypothesis has generated controversy and inconsistent data [25], a metastudy by Adams et al. [6] generally supported it for plants, also see [26]. That Hypericum fasciculatum in multimonth standing water selectively facilitates small plants at its elevated base (Figure 4b, 6) is consistent with SGH.
Expanding the perspective now to increasing distance from the base, Figures 7a and 7b indicate soil elevation to extend outward from the root cone. As Figure 7c shows, sedges concentrate to some extent in more or less the same radius. The reappraisal study in the Supplementary Materials echoed nearly identical sedge concentration pattern. (It should be noted that Hypericum fasciculatum occurs in varied wetlands. (It is not contended here that an elevated root cone surrounded by raised soil and concentrated sedge associates is universal for the species, especially in view of large variation in hydroperiods and substrates at non-studied sites.)
The situational associations of H. fasciculatum with elevated microtopography radiating to 30-60 cm and with associated enhanced sedge growth in the same perimeter poses a “cause and effect” concern. Does the shrub raise the marsh floor, or do the H. fasciculatum and the sedges around it merely fare best on higher ground? (Less likely, do the plants all benefit mutually from some additional underlying feature? Does H. fasciculatum cast protective shade, or suppresses non-sedge competitors allelopathically?)
That H. fasciculatum and the sedges benefit from raised marsh bottom regardless of the elevational cause is probably true. As Keddy [17] pointed out, “a few centimeters” wetland elevation can substantially affect species composition. Accordingly, relatively high spots will favor certain species, including it seems H. fasciculatum and associates. That H. fasciculatum is at least a contributor to the elevation is a visible fact, given the peaty substrate collecting at large root cones. Root penetration into that enriched soil, and influence on near neighbor relative frequencies are likewise visible (Figs. 4b, 6) at the cones. The question thus becomes the radial extent and broad ecological significance of the H. fasciculatum makes to soil building and to other species. The shrub is abundant throughout the HM marshes with obvious high turnover, and its collected soil has to go somewhere. In addition to the root cone, the shrub’s long radiating shallow roots would fortify soft loose sediments and, upon decay would add to the substrate beyond the canopy.
5. Conclusions
The studied Hypericum-Stillingia marshes filter 34 of 138 local obligate wetland species into an oligotrophic exposed habitat of alternating extended flooding and seasonal soil exposure. The plants there all are aerenchymatous, most show xeromorphy, and about half are rhizomatous. They sort into about five functional guilds having a phylogenetic component. One of the two woody species, Hypericum fasciculatum, to varying degrees with respect to size, to site conditions, and to individuals, collects periphyton and decaying organic debris on its raised roots, creating dark peaty soil in contrast with the normal marsh bottom sand. At the study sites, characterized by flooding for over half the year, Hypericum fasciculatum and associated sedges ostensibly grow best on elevated microtopography. Hypericum fasciculatum is self-elevating by adventitious root growth associated with soil-building and interspecific facilitation at its base. Elevated marsh bottom and enhanced sedge growth extend outward from the base to 30(60) cm, although with increasing distance, the role of H. fasciculatum as site engineer diminishes in intensity and in certainty.
6. Future Directions
Given that H. fasciculatum has root aerenchyma, and that the radiating roots can produce small upright roots, the roots may disperse exudates or oxygen to the surrounding soil. Of particular interest is the potential role of the root biota in oxygenation and in nutrient cycling, with an eye to nitrogen-fixing Cyanobacteria. The marsh plants have their subterranean systems in close physical proximity, impacting community assembly either randomly or deterministically, especially given broad diversity in root and rhizome structures. The root systems range from small compact “paintbrush” systems to spidery loose spreading roots in Aristida palustris to the “bottlebrush plus taproot” of Stillingia aquatica. The local marshes have a network of busy animal trails with their own mixed fauna, trailside flora, and nocturnal mysteries. A quick literature survey reveals the South Florida marsh mammal-plant ecology to be nearly unstudied.
Figure 8.
Graminoids concentrated around H. fasciculatum.

Supplementary Materials and Data Availability Statement
Supplementary materials are placed in figshare.com, 10.6084/m9.figshare.25513423. These are original field-based datasets for each substudy as listed in Table 1, and a plot and dataset (SM D) for the reappraisal study in substudy 6.
Funding
This research received no external funding.
Acknowledgments
Phosphorus testing was by ETR Laboratories, Leominster, MA. The Palm Beach County Dept. of Environmental Resource Management issued a research permit for the county natural areas. The Florida Fish and Wildlife Conservation Commission granted research permit SUO-82154 for the John C. and Mariana Jones/Hungryland Wildlife and Environmental Area.
Conflicts of Interest
The author declares no conflicts of interest.
Appendix A. List of species
| SPECIES | FAMILY | NOTES |
| Agalinis sp. | Orobanchaceae | Spotty, occasional in study zones |
| Aristida palustris (Chapm.) Vasey | Poaceae | Common but spotty |
| Bacopa caroliniana (Walter) B. L. Robinson | Plantaginaceae | Common in places, esp. in openings |
| Burmannia capitata (J.F.Gmel.)Mart. | Burmanniaceae | Unusual, limited in context to H. fasciculatum root tops |
| Centella asiatica (L.)Urb. | Apiaceae | Common, esp. in openings |
| Chamaecyse sp. | Euphorbiaceae | Shallow water, wet soil |
| Dichanthelium erectifolium (Nash)Gould & C.A.Clark | Poaceae | Almost ubiquitous |
| Eragrostis spectabilis (Pursh) Steud. | Poaceae | Unusual and spotty |
| Eriocaulon compressum Lam. | Eriocaulaceae | Ubiquitous but small |
| Eriocaulon decangulare L. | Eriocaulaceae | Common but spotty |
| Fuirena scirpoidea Michx. | Cyperaceae | Common but spotty |
| Gratiola ramosa Walter | Plantaginaceae | Spotty, can be abundant but not usually |
| Hypericum fasciculatum Lam. | Clusiaceae | Codominant |
| Hyptis alata (Raf.)Shinners | Lamiaceae | Scattered, occasional, spotty |
| Ludwigia linifolia Poir | Onagraceae | Common in places, spotty |
| Panicum tenerum Beyrich ex Trin. (Coleataenia tenera (Beyrich ex Trin.) Soreng) | Poaceae | Common, spotty, shy |
| Pinus elliottii Engelm. | Pinaceae | Very unusual in context, as seedling on H. fasciculatum |
| Pluchea baccharis (Miller) Pruski | Asteraceae | Becomes frequent during drying, and at margins |
| Proserpinaca pectinata Lam. | Haloragaceae | Common, spotty, mostly in openings |
| Rhynchospora divergens Chapm. ex M.A.Curtis | Cyperaceae | Rhizomeless on wet exposed marsh bottom, spotty, abundant in places |
| Rhynchospora harperi Small | Cyperaceae | Abundance variable in space & time, sometimes codominant w/R. tracyi |
| Rhynchospora inundata (Oakes) Fernald | Cyperaceae | Common locally but very little in immediate study zones |
| Rhynchospora latifolia (Baldwin) W.W.Thomas | Cyperaceae | Spotty |
| Rhynchospora tracyi Britton | Cyperaceae | Variably codominant with R. harperi |
| Sabatia decandra (Walter) R.M.Harper | Gentianaceae | Spotty, occasional in study zones |
| Sabatia grandiflora (A.Gray) Small | Gentianaceae | Spotty, occasional in study zones |
| Samolus ebracteatus Kunth | Primulaceae | Spotty |
| Schoenus nigricans L. | Cyperaceae | Spotty, sometimes plentiful |
| Scleria baldwinii (Torr.) Steud. | Cyperaceae | Spotty, can be highly abundant |
| Stillingia aquatica Chapm. | Euphorbiaceae | Codominant |
| Tiedemannia filiformis (Walter) Feist & S.R.Downie | Apiaceae | Common but spotty |
| Utricularia purpurea Walter | Lentibulariaceae | Abundant, U. purpurea, also U. foliosa and others present |
| Viola lanceolata L. | Violaceae | Uncommon within the study context, although plentiful in wetlands |
| Xyris smalliana Nash | Xyridaceae | Common and widespread |
References
- Kirkman, L.; Smith, L.; Golladay, S. Southeastern Depressional Wetlands. Wetl. Habitats N. Am. Ecol. Conserv. Concerns 2012, 203–215. [Google Scholar]
- Kral, R. Rhynchospora in Flora of North America @ Efloras.Org. Available online: http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=128495 (accessed on 30 March 2024).
- Weakley, A. ; collaborators Flora of the Southeastern US. Available online: https://fsus.ncbg.unc.edu/ (accessed on 30 March 2024).
- Kral, R. The Genus Xyris in Florida. Rhodora 1960, 62, 295–319. [Google Scholar]
- Bertness, M.D.; Callaway, R. Positive Interactions in Communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.E.; Besozzi, E.M.; Shahrokhi, G.; Patten, M.A. A Case for Associational Resistance: Apparent Support for the Stress Gradient Hypothesis Varies with Study System. Ecol. Lett. 2022, 25, 202–217. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G. Notes on the Natural History of Stillingia aquatica (Euphorbiaceae): With Special Attention to Reproductive Biology. J. Bot. Res. Inst. Tex. 2021, 15, 183–199. [Google Scholar] [CrossRef]
- Rogers, G.K. Notes on Hypericum fasciculatum (Clusiaceae) in South Florida Depression Marshes: With Attention to Structural Adaptations, to the Biological Context, and to Floral Biology1. J. Torrey Bot. Soc. 2022, 149. [Google Scholar] [CrossRef]
- Diamond, J.; Mclaughlin, D.; Slesak, R.; Stovall, A. Pattern and Structure of Microtopography Implies Autogenic Origins in Forested Wetlands. Hydrol. Earth Syst. Sci. 2019, 23, 5069–5088. [Google Scholar] [CrossRef]
- Diamond, J; Epstein J.; Cohen, M., and collaborators. A Little Relief: Ecological Functions and Autogenesis of Wetland Microtopography - Diamond - 2021 - WIREs Water - Wiley Online Library. Available online: https://wires.onlinelibrary.wiley.com/doi/abs/10.1002/wat2.1493 (accessed on 30 March 2024).
- Marani, M.; Da Lio, C.; D’Alpaos, A. Vegetation Engineers Marsh Morphology through Multiple Competing Stable States. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, 3259–3263. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Agnew, A.; Roxburgh, S. The Nature of Plant Communities | Plant Science. Available online: https://www.cambridge.org/us/academic/subjects/life-sciences/plant-science/nature-plant-communities. https://www.cambridge.org/us/academic/subjects/life-sciences/plant-science (accessed on 30 March 2024).
- Wunderlin, R. ; collaborators ISB: Atlas of Florida Plants. Available online: https://florida.plantatlas.usf.edu/ (accessed on 30 March 2024).
- R Core Team R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 30 March 2024).
- Rogers, G. Ecological Notes on Seasonal Depression Marshes in Southeast Florida, with Special Attention to Soil Modification and Interspecific Facilitation by Hypericum fasciculatum (Clusiaceae), a Woody Shrub 2024. [CrossRef]
- Kozlowski, T.T. Plant Responses to Flooding of Soil. BioScience 1984, 34, 162–167. [Google Scholar] [CrossRef]
- Keddy, P.A. Wetland Ecology: Principles and Conservation, 2nd ed.; Cambridge University Press: Cambridge, 2010. [Google Scholar]
- Dodds, W.K. The Role of Periphyton in Phosphorus Retention in Shallow Freshwater Aquatic Systems. J. Phycol. 2003, 39, 840–849. [Google Scholar] [CrossRef]
- Keddy, P.A.; Laughlin, D.C. A Framework for Community Ecology: Species Pools, Filters and Traits; Cambridge University Press: Cambridge, 2021; ISBN 978-1-316-51260-9. [Google Scholar]
- Boutin, C.; Keddy, P.A. A Functional Classification of Wetland Plants. J. Veg. Sci. 1993, 4, 591–600. [Google Scholar] [CrossRef]
- Garnier, E.; Navas, M.-L.; Grigulis, and K. Plant Functional Diversity: Organism Traits, Community Structure, and Ecosystem Properties; Oxford University Press: Oxford, New York, 2016. ISBN 978-0-19-875737-5.
- de Bello, F.; Carmona, C.P.; Dias, A.T.C.; Götzenberger, L.; Moretti, M.; Berg, M.P. Handbook of Trait-Based Ecology: From Theory to R Tools; Cambridge University Press: Cambridge, 2021; ISBN 978-1-108-47291-3. [Google Scholar]
- Moreira-Saporiti, A.; Teichberg, M.; Garnier, E.; Cornelissen, J.H.C.; Alcoverro, T.; Björk, M.; Boström, C.; Dattolo, E.; Eklöf, J.S.; Hasler-Sheetal, H.; et al. A Trait-Based Framework for Seagrass Ecology: Trends and Prospects. Front. Plant Sci. 2023, 14, 1088643. [Google Scholar] [CrossRef] [PubMed]
- Miller, A. et al. Coexistence of Many Species under a Random Competition-Colonization Trade-off - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/38261621/ (accessed on 30 March 2024).
- Campbell, D.; Keddy, P. The Roles of Competition and Facilitation in Producing Zonation Along an Experimental Flooding Gradient: A Tale of Two Tails with Ten Freshwater Marsh Plants. Wetlands 2022, 42, 5. [Google Scholar] [CrossRef]
- Bimler, M.D.; Stouffer, D.B.; Lai, H.R.; Mayfield, M.M. Accurate Predictions of Coexistence in Natural Systems Require the Inclusion of Facilitative Interactions and Environmental Dependency. J. Ecol. 2018, 106, 1839–1852. [Google Scholar] [CrossRef]
Figure 1.
Study area and main plants. a. HS marsh (at Casuarina-East). The marsh is crisscrossed by well-worn animal trails such as the one visible. b. Hypericum fasciculatum. c Stillingia aquatica. d. Rhynchospora harperi. Note small grassy leaves especially on right side of photo. e. R. harperi, achene. Rule marks 0.5 mm. f. R. tracyi, flowering heads.
Figure 1.
Study area and main plants. a. HS marsh (at Casuarina-East). The marsh is crisscrossed by well-worn animal trails such as the one visible. b. Hypericum fasciculatum. c Stillingia aquatica. d. Rhynchospora harperi. Note small grassy leaves especially on right side of photo. e. R. harperi, achene. Rule marks 0.5 mm. f. R. tracyi, flowering heads.

Figure 3.
Species groupings. a. Trait distributions by species. Dark color = possession of trait. “Needle leaves” = resemble short conifer needles roughly 1-1.5 cm long. “Knitting needles” in Figure 3b resemble knitting needles in shape and size multiple cm long. b. Dendrogram suggesting functional guilds, based on SM J. The y-axis shows the proportional distances for binary characters. Species abbreviations are in Figure 2 captions.
Figure 3.
Species groupings. a. Trait distributions by species. Dark color = possession of trait. “Needle leaves” = resemble short conifer needles roughly 1-1.5 cm long. “Knitting needles” in Figure 3b resemble knitting needles in shape and size multiple cm long. b. Dendrogram suggesting functional guilds, based on SM J. The y-axis shows the proportional distances for binary characters. Species abbreviations are in Figure 2 captions.
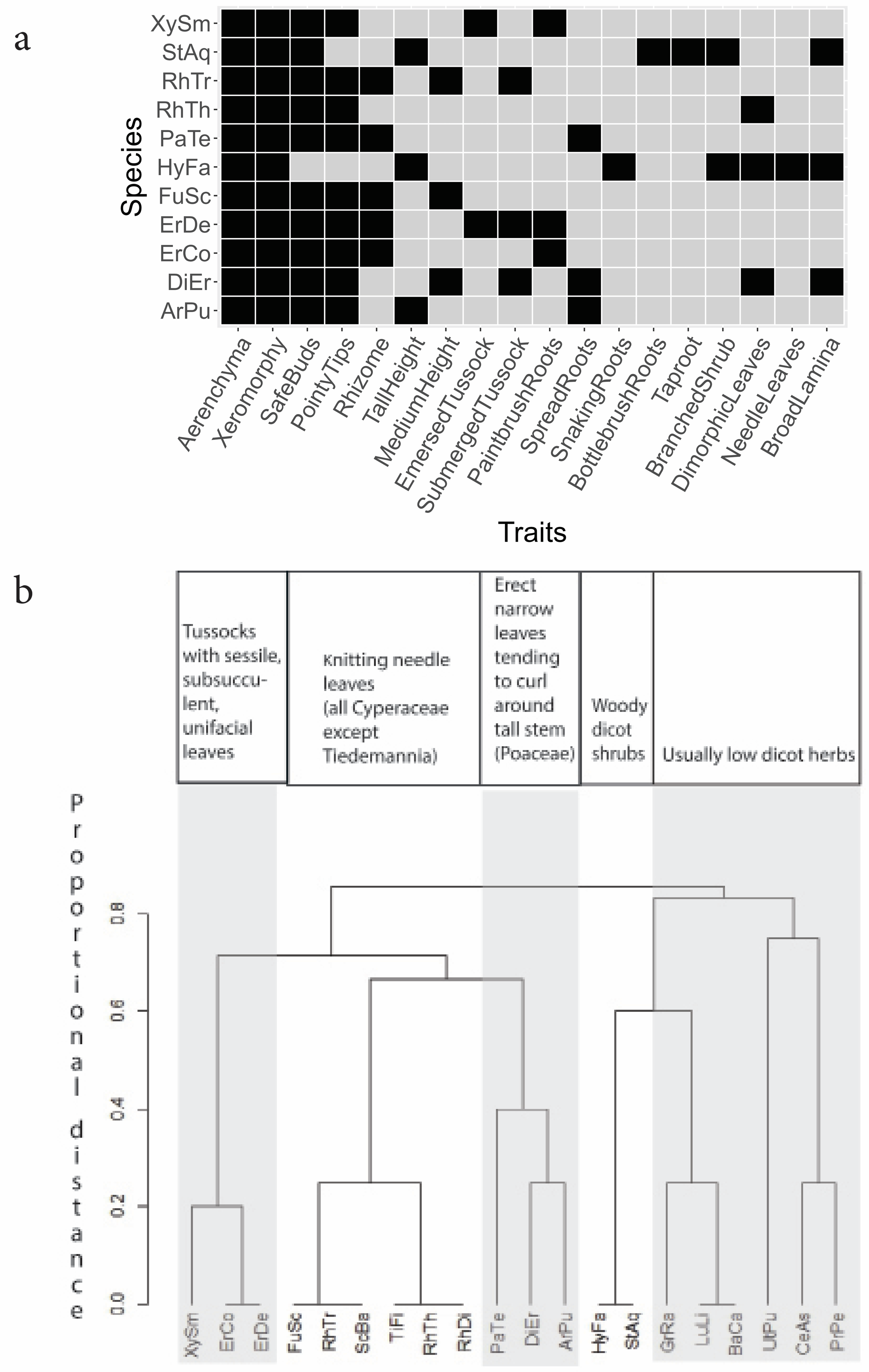
Figure 5.
Hypericum fasciculatum soils. a and b. Soils from marsh bottom. Samples apart from H. fasciculatum (left) and sample from under H. fasciculatum (right). c. Soil core from marsh bottom physically apart from H. fasciculatum. Rule marks = cm. Positions of ruler is meaningless. d. Soil core through H. fasciculatum root cone.
Figure 5.
Hypericum fasciculatum soils. a and b. Soils from marsh bottom. Samples apart from H. fasciculatum (left) and sample from under H. fasciculatum (right). c. Soil core from marsh bottom physically apart from H. fasciculatum. Rule marks = cm. Positions of ruler is meaningless. d. Soil core through H. fasciculatum root cone.

Figure 6.
Prevalence of (young) R. harperi as nearest neighbor to H. fasciculatum, at high water. Species abbreviations in Figure 2 caption.
Figure 6.
Prevalence of (young) R. harperi as nearest neighbor to H. fasciculatum, at high water. Species abbreviations in Figure 2 caption.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



