
Submitted:
08 April 2024
Posted:
11 April 2024
You are already at the latest version
Abstract
The domestic ducks (Anas platyrhynchos domesticus) are resistant to most of the highly pathogenic avian influenza virus (HPAIV) infections. In this study, we characterized the lung proteome and phosphoproteome of ducks infected with HPAI H5N1 virus (A/duck/India/02CA10/2011/Agartala) at 12 hr, 48 hr and 5 days post infection condition. Total 2082 proteins were differentially expressed and 320 phosphorylation sites mapping to 199 phosphopeptides corresponding to 129 proteins were identified. Functional annotation of proteome data analysis reveals activation of RIG-I-like receptor and Jak-STAT signaling pathways, that lead to induction of interferon stimulated genes (ISGs) expression. The pathway analysis of phosphoproteome datasets also confirms the activation of RIG I, Jak-STAT signaling, NF-kappa B signaling and MAPK signaling pathways in the lung tissues. The induction of ISGs proteins (STAT1, STAT3, STAT5B, STAT6, IFIT5 and PKR) establishes protective antiviral immune response in duck lung tissue. Further, the protein-protein interaction network analysis identified proteins like AKT1, STAT3, JAK2, RAC1, STAT1, PTPN11, RPS27A, NFKB1 and MAPK1 as main hub proteins that might play important roles in disease progression in ducks. Together, functional annotation of proteome and phosphoproteome datasets reveals the molecular basis of the disease progression and disease resistance mechanism in ducks infected with HPAI H5N1 virus.
Keywords:
Duck
; avian influenza virus
; proteomics
; phosphoproteome
; disease resistance
; hub proteins
Introduction
Ducks (Anas platyrhynchos domesticus) act as natural reservoir species of avian influenza viruses (AIVs). Most of the highly pathogenic avian influenza virus (HPAIV) infections in domestic ducks are asymptomatic or cause only mild transient clinical signs. This host protection mechanism against influenza infection in ducks may be conferred by increasing innate resistance to infection. System level understanding of host innate resistance mechanism in ducks, can help in designing efficient prophylactic and therapeutic strategies against influenza infection [1,2]. The high-throughput genomic methods are better suitable tools for system level understanding of the innate immune responses, as, they provide global views of the complex innate immune system and reveal the intertwined molecular events that are responsible for disease resistance mechanism [2]
Transcriptome analysis is presently one of the high-throughput omics technologies that can be easily performed, and these are often used as a proxy for protein abundance. Global host immune gene responses using microarrays/RNA-Seq have been investigated in ducks infected with different AIVs [3,4,5,6,7,8].These transcriptomics studies revealed that ducks produce an early RIG-I immune response and expression of cytokines, IFNs, iNOS, STAT3 and IFITM genes [5,6,8,9,10,11,12,13,14,15]. These differential expression patterns of innate immune genes play a crucial role in the disease progression of ducks to AIVs infection. Further, RIG I immune response is well known general concept in ducks for disease resistant, however an actual explanation of which molecular pathways constitutes this innate resistance immune response is lacking.
On the other hand, viral–host interactions are multidimensional nature and transcriptome represents only a single facet of the host response. Other types of high-throughput omic technologies might bring different insights on disease pathogenesis [2,8,16]. The influenza viruses also modulate post-transcriptional regulation and translation, which cannot be addressed by transcriptomics data alone, but requires integration of proteomic data and other omics profiling data [16,17]. The quantities of mRNAs and the corresponding proteins do not always correlate because proteins are modulated by complex post-transcriptional modification (PTM) processes [18]. Furthermore, PTMs such as phosphorylation, ubiquitination, acetylation, glycosylation and many others are required for regulating signal transduction and protein interactions of cellular, biological and immunological processes [18]. Hence, proteomics analysis along with phosphoproteomic data can elucidate the molecular mechanisms of influenza disease progression to remarkably greater extent. Still now only few proteomics studies available for avian species against avian influenza infection [8,19,20,21]. Further, most current research into innate immune processes against influenza virus infection is biased towards human model systems. Future research work needed for the wild and domestic aquatic birds, because these birds were recognized as the reservoirs of most influenza A viruses [22]. Hence, this study was planned to analysis both proteome and phosphoproteome datasets, in order to further understand the molecular pathways and proteomic determinants responsible for the innate resistant immune mechanisms in ducks infected with HPAI H5N1 virus.
Materials and Methods
Experimental Infection of Ducks
Six weeks old healthy domestic ducks, sero-negative for AIV were used for this study. The animal experiments were approved by the Institutional Animal Ethics Committee of ICAR-NIHSAD (Approval no. 68/IAEC/HSADL/12 dated 11.05.2012) and all the experiments were conducted in biosafety level 3 containment facility of ICAR-National Institute of High Security Animal Diseases, Bhopal, India. The ducks were separated into four groups (n=5 birds/group). Among the four groups, three groups were intranasally inoculated with 106 EID50 of H5N1 virus (A/duck/India/02CA10/2011/Agartala) and one group (control) was inoculated with PBS. The birds were observed daily for clinical signs. Lung tissues were collected from five birds from each infected group at 12hr, 48 hr, and 5 days post-infection. Lung tissues were also collected from the control group at 12hr post-inoculation. The tissues were snap chilled in Liquid Nitrogen and stored at -800C until protein extraction. Avian influenza virus infection of lung tissues was confirmed by virus isolation upon inoculation in embryonated chicken eggs (ECEs) and RT-PCR.
Protein Extraction
150mg lung tissue from each sample was washed in 50mM NH4HCO3 washing buffer. The lung tissue was cut into small pieces and 650 μl of SDS protein extraction lysis buffer [0.1% SDS (Invitrogen); 50mM NH4HCO3 (Sigma); 1X Complete™ Protease Inhibitor Cocktail (Roche-11836145001)] was added. Tissue samples were homogenized in LZ-Lyser homogenizer at 30 HZ for 2 min. After complete homogenization, the tissue lysate was incubated on ice for 90 mins for complete protein lysis. The lysate was centrifuged at 20,000g for 60 mins at 40C and the supernatant was collected. The supernatants were immediately snap heat treated at 560C for 30 min in a dry bath for inactivation of HPAIV H5N1 in the protein extracts. All the heat-treated samples were stored at -800C for mass spectrometry analysis.
Sample Preparation for LC-MS Analysis
The protein amount in each sample was estimated using BCA assay. The quality of the lung proteins was checked in SDS-PAGE. A pool for each time point was prepared by pooling 150μg protein lysate each from 3 best samples at that time point. Protein samples were reduced for 20 minutes at 600C in 10mM dithiothreitol solution followed by alkylation for 10 min in 20 mM iodacetamide in the dark at room temperature. To remove SDS from the samples, the samples were subjected to acetone precipitation (6X). The protein pellet obtained from each condition was then centrifuged at 13,000 x g for 10 min at 40C to remove the acetone and impurities. The pellets were then resuspended in 100 mM TEABC buffer and subjected to trypsin digestion (enzyme: substrate 1:20) at 370C overnight. After the trypsin digestion, the samples were subjected to Tandem Mass Tag™ labeling (TMT) labeling. The samples were labeled with the following TMT channels such as Control- 127C; D12H—128N; D48H—129C; D5D—130N as per manufacture protocol. After the TMT labeling, 5µl labeled sample was taken from each condition, pooled and the pooled sample were subjected to high pH reverse phase liquid chromatography separation. Total 96 fractions were collected and which was subsequently pooled to 12 fractions. 1/10th of the volume from these were transferred to eppendorf tubes and used for LC-MS/MS analysis in Orbitrap Fusion Tribrid mass spectrometer to determine the expression of total quantitative proteome. The remaining samples were pooled to obtain 6 fractions which were subjected for phosphoproteomics analysis. Each fraction was subjected to phosphopeptide enrichment using TiO2 beads. Briefly, the peptide samples were incubated with TiO2 beads in a ratio of 1:10 for 15 minutes followed by washing the bound beads and eluting the bound phosphopeptides. LC-MS/MS analysis was carried out in the Orbitrap Fusion Tribrid mass spectrometer to determine the extent of phosphorylation across various time points in the enriched fractions. Peptide fragmented by HCD fragmentation with collision energy of 34. The data was acquired in data dependent mode (DDA) with top 10 MS2.
Bioinformatics Analysis
The data obtained from both the total proteome and phosphoproteome datasets were analyzed in Proteome Discoverer 2.1 software suite. The data were searched using SequestHT algorithm. The reference duck proteome dataset was downloaded from NCBI database. The important parameters were used for database search as follows, Peptide mass error tolerance level-10ppm, Fragment mass error tolerance level—0.05Da, number of missed cleavage -2, and carbamidomethylation of cysteine, TMT label at N-term of peptides and lysine residue was set as fixed modification and oxidation of methionine was set as variable modification. In addition to above parameters, for identification of phosphoproteins phosphorylation at serine (S), threonine (T) and tyrosine (Y) was set as variable modification. The relative ratio for each phosphopeptide was calculated by dividing the intensity of each phosphorylated peptide over the intensity of the corresponding peptide. The high confident PTM sites were filtered by PhosphoRS score above 75%. The data were searched against target decoy database and the false discovery rate was set to1% at the protein and peptide level. Functional classification of the proteins and phosphoproteins were performed for gene ontology (GO) in Database for Annotation, Visualization and Integrated Discovery (DAVID) [23] and pathway analysis in NetworkAnalyst web server using Kyoto Encyclopedia of Genes and Genomes (KEGG) database [24,25]. We used online web server NetworkAnalyst for construction of protein-protein interaction (PPI) networks [24]. The main driving or hub proteins were identified on the basis of two topological measures, degree centrality and betweenness centrality. Heatmap generated using ‘Clustvis’ web tool [26].
Results and Discussion
Differential Protein Expression Analysis
The duck lung raw proteome dataset contained total 1, 20,000 MS spectra and 30,000 MS/MS spectra. The 30,000 MS/MS spectra were searched against NCBI duck proteome database. Total 2082 proteins were differentially expressed in ducks lung tissues infected with HPAI H5N1 virus at 1% FDR level. The analysis of differential expression of proteins at 12 hrs interval showed 876 proteins were upregulated and 1179 proteins were downregulated. A total of 898 proteins were upregulated and 1159 proteins were downregulated at 48 hr post-infection. At 5 days post infection, 1095 proteins upregulated and 962 proteins were downregulated in duck lung tissues (Table 1). The protein profile showed that 113, 72 and 205 proteins were exclusively up regulated in H5N1 infected duck lung tissue at 12hr, 48hr and 5days post-infection, respectively (Figure 1). Downregulation of 154, 112 and 49 proteins were observed exclusively at 12 hr, 48hr and 5days post-infection, respectively (Figure 1). The fold change value of the upregulated proteins ranged from 11.8 to 1. Interestingly, at 12 and 48 hr time point, higher number of proteins were downregulated as compare to 5 days time point post-infection condition (Table 1). Heatmap of differentially expressed proteins of HPAIV infected duck lung tissues was represented in Figure 2. This result indicates that many of the host proteins were downregulated at the initial stage of infection and then later host adjust the cellular homeostasis to recover from virus induced differential expression of proteins.
Gene ontology (GO) analysis of the exclusively upregulated and downregulated lung tissue proteins of H5N1 infected duck were done using DAVID tools. This analysis result provides information how the disease progress from commencement of disease to end of disease progression in duck infected with HPAI H5N1 virus. At 12 hr interval, upregulated protein list enriched Fc-epsilon receptor signaling pathway, MAPK cascade, Wnt signaling pathway, antigen processing and presentation of exogenous peptide antigen via MHC class I, NIK/NF-kappaB signaling, tumor necrosis factor-mediated signaling pathway, endocytosis, small GTPase mediated signal transduction GO terms. The downregulated protein list enriched ATP binding, glycolytic process, protein refolding, extracellular matrix, intracellular ribonucleoprotein complex, gluconeogenesis, NAD binding, translational initiation, mRNA transport and protein transport GO terms related with cellular homeostasis (Figure 3). These patterns indicate that at 12 hr time interval virus host interaction resulted activation host signalling pathways and inhibit the cellular homeostasis process. At 48 hr post infection stage, upregulated proteins enriched movement of cell component, mRNA splicing, focal adhesion, actin cytoskeleton, spliceosomal complex and the down regulated proteins enriched Golgi membrane, endoplasmic reticulum lumen, lysosome, calcium ion transport GO terms (Figure 4). At 5 days post infection stage, most of cellular homeostasis process like regulation of mRNA stability, protein polyubiquitination, cytoskeleton organization, regulation of translational initiation, tricarboxylic acid cycle, regulation of organelle assembly, positive regulation of cellular protein catabolic process, transport vesicle, GTPase activity, kinase activity GO terms were enriched (Figure 5). In summary, these results indicate that the inhibition of the cellular homeostasis and activation of signalling pathways at the initial stage of infection and host resume normal cellular homeostasis at later stage of infection.
Differential Phosphoproteomic Expression Analysis
Post-translational modifications (PTMs) are crucial regulatory cellular process in which proteins are enzymatically modified to reversibly modulate the activity, subcellular localization, conformation, and/or protein–protein interactions of the target protein. Thus, PTMs are essential for the dynamic regulation of intracellular pathogen sensing signal transduction pathways [27]. In RIG I and interferon-mediated signaling pathway, ubiquitination and phosphorylation events are important for induction of antiviral innate immune response. Phosphorylation is the most important PTMs, which involves the transfer of a phosphate group by a protein kinase to serine (S), threonine (T), and tyrosine (Y) residue of a target protein substrate.
In this study we investigated the phosphorylation-regulated host proteins and signaling pathways of HPAI H5N1 infected duck lung tissues at different time points post infection. We identified 320 phosphorylation sites mapping to 199 phosphopeptides corresponding to 129 proteins. In all, 93, 93 and 104 phosphosites were differentially phosphorylated (1.5-fold up and down) in 12 hr, 48 hr and 5days post infection respectively in duck lung tissues. The phosphoproteome profile showed that 36, 27 and 40 sites were hyperphosphorylated in duck lung tissue proteins at 12hr, 48hr and 5days post-infection, respectively. Further, 57, 66 and 64 sites were observed hypophosphorylated in the lung tissues proteins at 12 hr, 48hr and 5days post-infection, respectively. Our phosphoproteome profile indicates that HPAI H5N1 virus has a substantial impact on host protein phosphorylation. Most proteins were phosphorylated at Ser residues in comparison with Thr and Tyr residues, which is agreed with a previous study [28,29]. To elucidate the kinase family involved in influenza infection condition, we used group-based prediction system (GPS) [30]. The GPS ranks the likelihood that a particular kinase or kinase family phosphorylates a given phosphorylation site by considering the amino acids surrounding the phosphorylation site. The AGC, CK1, CMGC and CAMK protein kinase families were predicted to be activated during HPAI H5N1 infection condition in ducks. Activation of these protein kinase family members in influenza infection was reported in previous study [28,29,31].
Pathway Analysis of Proteomics Datasets
In domestic ducks, most HPAI virus infections cause no or mild clinical signs and lesions [32,33]. Understanding the disease resistance immune responses in ducks in HPAIV infection may provide key insights into immune pathways required for prophylactic or therapeutic protection. Previous literature specifies that, HPAI virus-infected ducks produce an early protective type I interferon response and thereby recovers from influenza induced inflammatory pathology [5,6,14,34,35]. However, the relationship between type I interferon and inflammation is complex and multifactorial cellular process [36]. Further, the several signaling pathways activated during type I interferon response are yet to be confirmed at protein and phosphoproteome level in ducks. Hence, the proteome and phosphoproteome datasets were used for pathway analysis in order to identify the signaling pathways that are activated in ducks during HPAIVs infection. Molecular pathway analysis of differential expressed proteins shows activation RIG-I-like receptor signaling pathway, Jak-STAT signaling pathway, PI3K-Akt signaling pathway, MAPK signaling pathway, NOD-like receptor signaling pathway, Toll-like receptor signaling pathways in lung tissue of ducks (Table 2).
RIG-I-Like Receptors Signaling Pathway
Retinoic acid-inducible gene I is thought to be an important pattern recognizing receptor (PRR) during influenza infection [37]. The RLR family includes three members: RIG-I, melanoma differentiation-associated gene 5 (MDA5) and laboratory of genetics and physiology 2 (LGP2) [38]. We observed upregulation of expression of MDA5 and LGP2 proteins in the duck H5N1 infected lung tissues. Significant upregulation of MDA5 and LGP2 have been observed in the lung tissues of Muscovy ducks infected with HPAI H5N1 infection [39,40]. In ducks, both MDA5 and LGP2 activate IRF-7-dependent signaling pathway and induce interferons (IFNs) as well as interferon-stimulated genes (ISGs) production, both of which mediate the antiviral and pro-inflammatory responses during HPAI H5N1 virus infection [8,39,40,41,42,43,44]. The tripartite motif (TRIM) family of proteins, tripartite motif 25 (TRIM25) and ubiquitin carboxyl-terminal hydrolase 15 (USP15) were expressed in the duck lung tissues (Figure 6). The IFN-inducible E3 ubiquitin ligase TRIM25 is a key regulator of the RIG-I-mediated IFN response and modifies RIG-I with K63-linked polyubiquitination [43,45]. The CARD domains of RIG-I and MDA5 were ubiquitinated by TRIM25 [46,47,48]. TRIM25 stabilizes the RIG-I–2CARD: MAVS–CARD helical structure by furnishing short chains of K63-linked ubiquitin molecules and amplifies MAVS signaling [45]. Further, duck TRIM25 provides both anchored and unanchored K63-linked polyubiquitin chains to CARD domains of RIG-I [48]. USP15 is a deubiquitinases, which removes K48-linked ubiquitination from the TRIM25 SPRY domain thus stabilizing TRIM25 and inducing a sustained cytokine response. Taken together, RIG-I like receptors ubiquitination by TRIM25 protein induces RIG-I oligomerization and its interaction with MAVS activates RIG-I signaling pathway and induce antiviral gene expression in lung tissues of HPAI H5N1 virus infected ducks.
Interferon Signaling: The Jak-Stat Pathway
The Janus Kinases 2 (JAK2) are ubiquitously expressed and bind to heterodimers of IFN-γ receptors 1 (IFNGR1) and 2 (IFNGR2) chains on the inner side of the membrane. JAKs provide receptors with stability, facilitate their cell surface localization, and serve as key components of IFN signaling complexes [49,50,51]. Up regulation of JAK2 protein in the all three-post infection condition was observed in the HPAIV infected duck lung tissues (Figure 6). Upon binding of type II IFN to the IFNGR1/2, the receptor complex leads to phosphorylation of preassociated JAK1 and JAK2 tyrosine kinases, and transphosphorylation of the receptor chains leads to recruitment and phosphorylation of signal transducers and activators of transcription 1 (STAT1) [52]. Homodimers of phosphorylated STAT1 proteins form the IFN-γ activation factor (GAF). The GAF translocate into the nucleus and bind in the gamma-activated sequence promoter elements and resulting in expression of ISGs [53].
STAT proteins have the dual function of signal transduction and activation of transcription [54]. In duck proteome dataset, expression of STAT proteins like STAT1, STAT3, STAT5B and STAT6 were observed (Figure 6). It is also previously reported that Influenza A virus induces expression STATs protein in the lung tissues of duck [6,13,55]. STAT1 is critical in signal transduction from the type I IFNs and the type II IFN [52,56,57]. Previous studies showed SOCS-3 expression negatively affects STATs phosphorylation [58,59]. The STAT3 protein act as a transcription factor, plays a critical role in the IFN signalling pathways and is required for a robust IFN-induced antiviral response [60]. STAT3 protein has antagonistic effect on the inflammatory cytokine response and promotes a strong anti-inflammatory response [6,13,55]. STAT5B is activated in response to a variety of cytokines and binds to the gamma interferon activation site element and activates prolactin induced transcription [61,62].
Interferon-induced protein with tetratricopeptide 5 (IFIT5) exhibited antiviral activity against influenza virus by sequestering single strand viral RNA with 5′triphosphate [63]. IFIT5 protein at 12 hrs interval was down regulated and in later post infection condition IFIT5 showed upregulation to counter viral infection. Similar kind of upregulation were observed in duck lungs and spleens in response to HPAI H5N1 viruses [6,35,64,65]. PKR is an interferon-induced protein with antiviral, anti-proliferative, and pro-apoptotic functions. PKR is upregulated by type I and type III IFNs signaling pathways [66]. PKR has a known anti-influenza A virus effect by phosphorylating the alpha subunit of eukaryotic initiation factor 2 (EIF2α), which shuts down cellular and influenza A viral protein synthesis, thereby effectively reducing viral replication [67,68]. PKR is highly upregulated in the lungs of HPAI-infected ducks and the same pattern has been reported (Figure 6) [6,65]. In summary, our datasets showed evidence that protective interferon responses were activated through stimulation of RIG-I-like receptor signaling and Jak-STAT signaling pathways that resulted in induction of expression of ISGs (STAT1, STAT3, STAT5B, STAT6, IFIT5 and PKR).
Pathway Analysis of Phosphoproteomics Datasets
To further gain insight into the activation of specific signal transduction pathways by HPAI H5N1 infection condition in ducks, we analyzed the phosphoproteome data using the KEGG pathways database. The most notable cellular pathways activated for differentially regulated phosphoproteins in ducks lung tissues included Jak-STAT signaling pathway, NF-kappa B signaling pathway, MAPK signaling pathway, ErbB signaling pathway, PI3K-Akt signaling pathway, Rap1 signaling pathway, regulation of actin cytoskeleton, prolactin signaling pathway, chemokine signaling pathway, T cell receptor signaling pathway, endocytosis, tight junction etc (Table 3). Among these pathways, Jak-STAT signaling pathway, NF-kappa B signaling pathway and MAPK signaling pathway were interrelated with RIG I and interferon-mediated signaling pathways. Hence, our phosphoproteome datasets pathway analysis also confirm the activation of RIG I and interferon-mediated signaling pathway in HPAIV infection condition in ducks. Activation of MAPK signaling pathway, endocytosis and tight junction in influenza virus infection condition was reported [29]. Furthermore, activation of focal adhesions and actin cytoskeleton pathway were involved in the early stages of IAV infection was also reported [29,31]. Lakadamyali et al., 2003 suggested the endocytic pathway toward late endosomes: endosome maturation and initial acidification, occurs in the perinuclear region [69]. Sun and Whittaker (2007) reported indispensable roles of a dynamic actin cytoskeleton for influenza virus entry into the epithelial cells [70]. Thus, activation of these cellular functions is essential for influenza virus entry into host cells.
Influenza virus has ability to hijack the various intracellular signaling pathways to support their efficient replication. Among the signalling pathways, activation of PI3K/Akt signalling, MAPK (Raf/MEK/ERK) signalling pathway, and NF-κB signalling pathways were reported as essential pathways responsible for virus-supportive replication. The influenza NS1 protein binds to the SH2 domain of the p85 sub8unit and activates PI3K/Akt signalling pathways [71,72]. Activation of this pathway prevents premature apoptosis thereby promoting efficient virus replication at a late step of the infection [22,73]. MAPK (Raf/MEK/ERK) signalling pathways encompass cascades of kinases, which convert extracellular signals into cellular responses [74]. Activation of the Raf/MEK/ERK signalling pathway facilitates influenza virus replication supportive roles by inducing nuclear export of viral ribonucleoprotein complexes (RNPs) at late stages of the viral life cycle [75]. Further, influenza virus infection impairs Ras-Raf-MEK-ERK pathway by downregulating MEK1 SUMOylation to facilitate viral RNP export and virus propagation [76]. Activation of the NF κB signalling pathway is one of the essential cellular responses regulating antiviral cytokine and interferon-β expression [77]. A number of previous studies suggest that influenza viruses-elicited NF-κB activity, which helps in virus replication and spreading and the NF-κB inhibition also impairs IAV propagation [78,79,80]. In nutshell, phosphoproteome datasets analysis provide insights into multiple kinase-mediated signaling pathways were activated in HPAI H5N1 virus infected duck lung tissues.
Identification of Proteomic Determinants
Biological network analysis is a powerful approach to gain systems-level understanding of disease pathogenesis. To know main driving or hub proteins responsible for disease resistance to HPAIV infection condition in ducks, we constructed protein–protein interaction (PPI) networks for differentially expressed lung proteins of ducks. Among different molecular networks, PPI networks have emerged as an important resource because protein interactions play fundamental roles in structuring and mediating essentially all biological processes. PPI networks are often presented as undirected graphs with nodes as proteins and edges indicating interactions between two connecting proteins.
The proteins such as AKT1, STAT3, JAK2, RAC1, STAT1, PTPN11, RPS27A, NFKB1, MAPK1 etc were identified with high degree centrality and betweenness centrality measures as main hub proteins responsible for disease pathogenesis in ducks (Table 4) (Figure 7). The functional proteomics and phosphoproteomics analysis of our datasets shows that activation of RIG I signaling and Jak-STAT signaling pathways were responsible for disease resistance against HPAI H5N1 virus infection in ducks. The JAK2, STAT1, STAT3 and NFKB1 proteins were involved in these pathways were identified as main hub proteins for disease pathogenesis in ducks. These proteins act as main signaling molecules for induction of expression of ISGs and establishment of antiviral state in the host cells.
Akt (Protein kinase B, PKB) is a serine/threonine kinase and Akt is activated by the lipid products of phosphoinositide 3′-kinase (PI3K). Upon activation, Akt regulates the activities related to proliferation, cell cycle, glycogen synthesis, angiogenesis, and telomerase [81]In particular, PI3K–Akt pathway plays a critical role in uptake of virus during viral entry, prevention of premature apoptosis, and viral RNA expression and RNP localization [72,73,82]. RAC1 belongs to the family of Rho GTPases that regulate a wide variety of cellular processes. RAC1 is reported to have virus-supportive as well as virus-suppressive functions for different viruses [83,84,85,86]. In case of influenza virus infections condition, RAC1 has antiviral role by inducing interferon-β production and crucial virus-supportive role in the activity of the viral polymerase complex [87,88]. Influenza A virus (IAV) blocks RAC1-mediated host cell signal transduction through the NS1 protein to facilitate its own replication [89]. Further, RAC1 is proposed as cellular target for therapeutic treatment of influenza virus infections as inhibition of RAC1 by small chemical compound NSC23766 resulted in impaired replication of a wide variety of influenza viruses [88].
Protein tyrosine phosphatase non-receptor type 11 (PTPN11) is a member of the protein tyrosine phosphatase family. Expression of PTPN11 genes has been reported in influenza infection condition [90,91]. SHP-2, the protein encoded by PTPN11, interacts with the transcriptional activator STAT3 [92]. PTPN11 interacts with long intergenic noncoding RNA LINC00673, which promotes cell growth and proliferation by activating SRC-ERK signaling and inhibiting STAT1 signaling [93]. PTPN11 positively regulates MAPK signal transduction pathway [94]. NFKB1 is a subunit of the NF-kappaB (NF-κB) protein complex. Activation of NF-κB plays an important role in the inhibition of virus replication by stimulating the synthesis of IFN-α/β, which leads to the expression of antiviral genes [95,96]. However, the NS1 protein of influenza A virus has the ability to prevent NF-κB activation thereby subvert the IFN innate immune system [97].
MAPK1 is a member of the MAP kinase family. MAPKs play critical roles in regulating the expression of various proinflammatory cytokines, including IL-6 and TNF-alpha in response to microbial infection [98]. Activation of p38 mitogen-activated protein kinase (p38 MAPK) induces dysregulation of cytokine expression in HPAIV infected primary human monocyte-derived macrophages and bronchial epithelial cells [99,100,101]. Further, the inhibition of p38 MAPK significantly reduces hyper induction of cytokines and prevents cytokine-induced pathogenicity in HPAIV infected mice and human macrophages [99,102]. Therefore, interference with the p38 MAPK pathway is also proposed for therapeutic treatment for HPAIV infection [102]. RPS27A and UBA52 genes code for ubiquitin and this ubiquitin is a highly conserved protein that has a major role in targeting cellular proteins for degradation in the 26S proteosome [103,104]. Further, UBA52 host protein interacts with viral RNA polymerase acidic protein (PA), PA-N155, and PA-N182 and promotes the replication of highly pathogenic H5N1 avian influenza virus [105].
The Ras homolog gene family member A (RhoA) is a small GTPase that controls gene transcription, actin polymerization, cell cycle progression, and cell transformation. Influenza virus NS1 protein induces G0/G1 cell cycle arrest mainly through interfering with the RhoA/pRb signaling cascade [106]. In summary, we identified many proteomic determinants involved in the RIG I, Jak-STAT and other signaling pathways responsible for disease progression and disease resistance in ducks HPAIV infection by PPI network analysis. Most of the proteomic determinants identified in this study were previously reported in human model systems and showed evidence as play a very essential role in disease pathogenesis in influenza infection conditions. Here we identified the expression of these proteins in duck lung tissues and functionally annotated their role in disease progression. Further, some of these proteomic determinants were already tested as novel cellular targets for the therapeutic treatment of influenza virus infections. However, further in vivo studies are required to validate the role of proteomic determinants in disease progression and find novel host proteins as a therapeutic or prophylactic treatment in HPAIV infection in ducks.
Conclusions
In conclusion, bioinformatics analysis of proteome and phosphoproteome datasets revealed activation of RIG-I-like receptors signaling and Jak-STAT signaling pathways resulted into expression of ISGs proteins (STAT1, STAT3, STAT5B, STAT6, IFIT5 and PKR) thus establish protective antiviral immune response in HPAI H5N1 virus infected lung tissues. Activation of these pathways and expression of interferon stimulated proteins may restrict viral replication and recovery from influenza virus induced inflammatory changes thereby ducks may develop disease resistance against HPAI H5N1 virus infection condition. Through PPI network analysis, we identified many proteomic determinants (AKT1, STAT3, JAK2, RAC1, STAT1, PTPN11, RPS27A, NFKB1 and MAPK1) that might play important roles in disease progression and disease resistance in ducks.
Author Contributions
Conceived and designed the experiments: A.M., T.S.K.P., A.A.R. and P.V. Performed the experiments: A.M., R.S., Y.S., A.A.R. and P.V. Analyzed the data: S.M.P. and P.V. Contributed reagents/materials/analysis tools: S.M.P. and T.S.K.P. Wrote the paper: P.V. and A.M. All authors have read and approved the manuscript.
Funding
This work was funded by the Department of Biotechnology (grant number:BT/IN/Indo-UK/FADH/48/AM/2013). The funders had no role in study design, data collection, analysis of data and preparation of the manuscript.
Ethics statement
The experiments were approved by the Institutional Animal Ethics Committee of ICAR-NIHSAD (Approval no. 68/IAEC/HSADL/12 dated 11.05.2012), and performed under the guidance of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Govt. of India.
Acknowledgements
We thank Director, ICAR-National Institute of High Security Animal Diseases, Director, ICAR-Indian Veterinary Research Institute, and Indian Council of Agricultural Research, India for providing necessary facilities to carry out this work.
Competing Interests
The authors declare no competing interests.
References
- Herold, S.; Becker, C.; Ridge, K.M.; Budinger, G.R.S. Influenza Virus-Induced Lung Injury: Pathogenesis and Implications for Treatment. Eur Respir J 2015, 45, 1463–1478. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Vijayakumar, P.; Raut, A.A. Emerging Avian Influenza Infections: Current Understanding of Innate Immune Response and Molecular Pathogenesis. Int Rev Immunol 2017, 36. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Y.; Burt, D.W.; Chen, H.; Zhang, Y.; Qian, W.; Kim, H.; Gan, S.; Zhao, Y.; Li, J.; et al. The Duck Genome and Transcriptome Provide Insight into an Avian Influenza Virus Reservoir Species. Nat Genet 2013, 45, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Maughan, M.N.; Dougherty, L.S.; Preskenis, L.A.; Ladman, B.S.; Gelb, J.; Spackman, E. V.; Keeler, C.L. Transcriptional Analysis of the Innate Immune Response of Ducks to Different Species-of-Origin Low Pathogenic H7 Avian Influenza Viruses. Virol J 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Smith, N.; Yu, L.; Paton, I.R.; Gutowska, M.W.; Forrest, H.L.; Danner, A.F.; Seiler, J.P.; Digard, P.; Webster, R.G.; et al. A Comparative Analysis of Host Responses to Avian Influenza Infection in Ducks and Chickens Highlights a Role for the Interferon-Induced Transmembrane Proteins in Viral Resistance. BMC Genomics 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Vijayakumar, P.; Gandhale, P.N.; Ranaware, P.B.; Kumar, H.; Kulkarni, D.D.; Raut, A.A.; Mishra, A. Genome-Wide Gene Expression Pattern Underlying Differential Host Response to High or Low Pathogenic H5N1 Avian Influenza Virus in Ducks. Acta Virol 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Klees, S.; Schlüter, J.S.; Schellhorn, J.; Bertram, H.; Kurzweg, A.C.; Ramzan, F.; Schmitt, A.O.; Gültas, M. Comparative Investigation of Gene Regulatory Processes Underlying Avian Influenza Viruses in Chicken and Duck. Biology (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, P.; Raut, A.A.; Chingtham, S.; Murugkar, H.V.; Kulkarni, D.D.; Sood, R.; Singh, V.P.; Mishra, A. Proteomic Analysis of Differential Expression of Lung Proteins in Response to Highly Pathogenic Avian Influenza Virus Infection in Chickens. Arch Virol 2022, 167. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.W.; Aldridge, J.R.; Webster, R.G.; Magor, K.E. Association of RIG-I with Innate Immunity of Ducks to Influenza. Proc Natl Acad Sci U S A 2010, 107, 5913–5918. [Google Scholar] [CrossRef]
- Sarmento, L.; Afonso, C.L.; Estevez, C.; Wasilenko, J.; Pantin-Jackwood, M. Differential Host Gene Expression in Cells Infected with Highly Pathogenic H5N1 Avian Influenza Viruses. Vet Immunol Immunopathol 2008, 125, 291–302. [Google Scholar] [CrossRef]
- Liang, Q. long; Luo, J.; Zhou, K.; Dong, J. xin; He, H. xuan Immune-Related Gene Expression in Response to H5N1 Avian Influenza Virus Infection in Chicken and Duck Embryonic Fibroblasts. Mol Immunol 2011, 48, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Burggraaf, S.; Bingham, J.; Payne, J.; Kimpton, W.G.; Lowenthal, J.W.; Bean, A.G.D. Increased Inducible Nitric Oxide Synthase Expression in Organs Is Associated with a Higher Severity of H5N1 Influenza Virus Infection. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Kuchipudi, S. V.; Tellabati, M.; Sebastian, S.; Londt, B.Z.; Jansen, C.; Vervelde, L.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; Chang, K.C. Highly Pathogenic Avian Influenza Virus Infection in Chickens but Not Ducks Is Associated with Elevated Host Immune and Pro-Inflammatory Responses. Vet Res 2014, 45. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.K.; Fleming-Canepa, X.; Webster, R.G.; Magor, K.E. Tissue Specific Transcriptome Changes Upon Influenza A Virus Replication in the Duck. Front Immunol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Su, Q.; Luo, J.; Li, M.; Wu, Q.; Chang, H.; Du, J.; Huang, C.; Ma, J.; Han, S.; et al. Differences in Highly Pathogenic H5N6 Avian Influenza Viral Pathogenicity and Inflammatory Response in Chickens and Ducks. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Josset, L.; Tisoncik-Go, J.; Katze, M.G. Moving H5N1 Studies into the Era of Systems Biology. Virus Res 2013, 178, 151–167. [Google Scholar] [CrossRef]
- Zak, D.E.; Tam, V.C.; Aderem, A. Systems-Level Analysis of Innate Immunity. Annu Rev Immunol 2014, 32, 547–577. [Google Scholar] [CrossRef]
- Meissner, F.; Mann, M. Quantitative Shotgun Proteomics: Considerations for a High-Quality Workflow in Immunology. Nat Immunol 2014, 15, 112–117. [Google Scholar] [CrossRef]
- Zou, W.; Ke, J.; Zhang, A.; Zhou, M.; Liao, Y.; Zhu, J.; Zhou, H.; Tu, J.; Chen, H.; Jin, M. Proteomics Analysis of Differential Expression of Chicken Brain Tissue Proteins in Response to the Neurovirulent H5N1 Avian Influenza Virus Infection. J Proteome Res 2010, 9, 3789–3798. [Google Scholar] [CrossRef]
- Li, Y.; Ming, F.; Huang, H.; Guo, K.; Chen, H.; Jin, M.; Zhou, H. Proteome Response of Chicken Embryo Fibroblast Cells to Recombinant H5N1 Avian Influenza Viruses with Different Neuraminidase Stalk Lengths. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Guo, K.; Lin, X.; Li, Y.; Qian, W.; Zou, Z.; Chen, H.; Zhou, H.; Jin, M. Proteomic Analysis of Chicken Embryo Fibroblast Cells Infected with Recombinant H5N1 Avian Influenza Viruses with and without NS1 EIF4GI Binding Domain. Oncotarget 2017, 9, 8350–8367. [Google Scholar] [CrossRef]
- Hale, B.G.; Jackson, D.; Chen, Y.H.; Lamb, R.A.; Randall, R.E. Influenza A Virus NS1 Protein Binds P85beta and Activates Phosphatidylinositol-3-Kinase Signaling. Proc Natl Acad Sci U S A 2006, 103, 14194–14199. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat Protoc 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Xia, J.; Benner, M.J.; Hancock, R.E.W. NetworkAnalyst--Integrative Approaches for Protein-Protein Interaction Network Analysis and Visual Exploration. Nucleic Acids Res 2014, 42. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Gack, M.U. Post-Translational Control of Intracellular Pathogen Sensing Pathways. Trends Immunol 2017, 38, 39–52. [Google Scholar] [CrossRef]
- Soderholm, S.; Kainov, D.E.; Ohman, T.; Denisova, O. V.; Schepens, B.; Kulesskiy, E.; Imanishi, S.Y.; Corthals, G.; Hintsanen, P.; Aittokallio, T.; et al. Phosphoproteomics to Characterize Host Response During Influenza A Virus Infection of Human Macrophages. Mol Cell Proteomics 2016, 15, 3203–3219. [Google Scholar] [CrossRef] [PubMed]
- Yángüez, E.; Hunziker, A.; Dobay, M.P.; Yildiz, S.; Schading, S.; Elshina, E.; Karakus, U.; Gehrig, P.; Grossmann, J.; Dijkman, R.; et al. Phosphoproteomic-Based Kinase Profiling Early in Influenza Virus Infection Identifies GRK2 as Antiviral Drug Target. Nat Commun 2018, 9. [Google Scholar] [CrossRef]
- Xue, Y.; Liu, Z.; Cao, J.; Ma, Q.; Gao, X.; Wang, Q.; Jin, C.; Zhou, Y.; Wen, L.; Ren, J. GPS 2.1: Enhanced Prediction of Kinase-Specific Phosphorylation Sites with an Algorithm of Motif Length Selection. Protein Eng Des Sel 2011, 24, 255–260. [Google Scholar] [CrossRef]
- Weber, A.; Dam, S.; Saul, V. V.; Kuznetsova, I.; Müller, C.; Fritz-Wolf, K.; Becker, K.; Linne, U.; Gu, H.; Stokes, M.P.; et al. Phosphoproteome Analysis of Cells Infected with Adapted and Nonadapted Influenza A Virus Reveals Novel Pro- and Antiviral Signaling Networks. J Virol 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Parsons, G.; Manvell, R.J. Experimental Assessment of the Pathogenicity of Eight Avian Influenza A Viruses of H5 Subtype for Chickens, Turkeys, Ducks and Quail. Avian Pathol 1986, 15, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Perkins LE; Swayne DE Pathogenicity of a Hong Kong-Origin H5N1 Highly Pathogenic Avian Influenza Virus for Emus, Geese, Ducks, and Pigeons. Avian Dis. 2002, 46, 53–63. [CrossRef]
- Cagle, C.; Wasilenko, J.; Adams, S.C.; Cardona, C.J.; To, T.L.; Nguyen, T.; Spackman, E.; Suarez, D.L.; Smith, D.; Shepherd, E.; et al. Differences in Pathogenicity, Response to Vaccination, and Innate Immune Responses in Different Types of Ducks Infected with a Virulent H5N1 Highly Pathogenic Avian Influenza Virus from Vietnam. Avian Dis 2012, 56, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Saito, L.B.; Diaz-Satizabal, L.; Evseev, D.; Fleming-Canepa, X.; Mao, S.; Webster, R.G.; Magor, K.E. IFN and Cytokine Responses in Ducks to Genetically Similar H5N1 Influenza A Viruses of Varying Pathogenicity. J Gen Virol 2018, 99, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Type I Interferon: Friend or Foe? J Exp Med 2010, 207, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential Roles of MDA5 and RIG-I Helicases in the Recognition of RNA Viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Matsumiya, T.; Stafforini, D.M. Function and Regulation of Retinoic Acid-Inducible Gene-I. Crit Rev Immunol 2010, 30, 489–513. [Google Scholar] [CrossRef]
- Wei, L.; Cui, J.; Song, Y.; Zhang, S.; Han, F.; Yuan, R.; Gong, L.; Jiao, P.; Liao, M. Duck MDA5 Functions in Innate Immunity against H5N1 Highly Pathogenic Avian Influenza Virus Infections. Vet Res 2014, 45. [Google Scholar] [CrossRef]
- Jiao, P.R.; Wei, L.M.; Song, Y.F.; Cui, J.; Zhang, S.; Han, F.; Yuan, R.Y.; Liao, M. Molecular Cloning and Immune Responsive Expression of LGP2 Gene, a Pivotal Member of the RLR Gene Family from Muscovy Duck Cairina Moschata. Poult Sci 2015, 94, 1170–1176. [Google Scholar] [CrossRef]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 Is a Positive Regulator of RIG-I- and MDA5-Mediated Antiviral Responses. Proc Natl Acad Sci U S A 2010, 107, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, J.; Huang, Q.; Zhang, J.; Liu, J.; Xue, Q.; Li, W.; Liao, M.; Jiao, P. Host Innate Immune Response of Geese Infected with Clade 2.3.4.4 H5N6 Highly Pathogenic Avian Influenza Viruses. Microorganisms 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, J.; Xue, Q.; Liu, J.; Huang, B.; He, Z.; Huang, J.; Zu, S.; Chen, Z.; Zhao, B.; et al. Duck TRIM32 Functions in IFN-β Signaling Against the Infection of H5N6 Highly Pathogenic Avian Influenza Virus. Front Immunol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Pal, A.; Baviskar, P. RIGI, TLR7, and TLR3 Genes Were Predicted to Have Immune Response Against Avian Influenza in Indigenous Ducks. Front Mol Biosci 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Gack, M.U.; Shin, Y.C.; Joo, C.H.; Urano, T.; Liang, C.; Sun, L.; Takeuchi, O.; Akira, S.; Chen, Z.; Inoue, S.; et al. TRIM25 RING-Finger E3 Ubiquitin Ligase Is Essential for RIG-I-Mediated Antiviral Activity. Nature 2007, 446, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Gack, M.U.; Albrecht, R.A.; Urano, T.; Inn, K.S.; Huang, I.C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; García-Sastre, A. Influenza A Virus NS1 Targets the Ubiquitin Ligase TRIM25 to Evade Recognition by the Host Viral RNA Sensor RIG-I. Cell Host Microbe 2009, 5, 439–449. [Google Scholar] [CrossRef]
- Jiang, X.; Kinch, L.N.; Brautigam, C.A.; Chen, X.; Du, F.; Grishin, N. V.; Chen, Z.J. Ubiquitin-Induced Oligomerization of the RNA Sensors RIG-I and MDA5 Activates Antiviral Innate Immune Response. Immunity 2012, 36, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Miranzo-Navarro, D.; Magor, K.E. Activation of Duck RIG-I by TRIM25 Is Independent of Anchored Ubiquitin. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Gauzzi, M.C.; Barbieri, G.; Richter, M.F.; Uzé, G.; Ling, L.; Fellous, M.; Pellegrini, S. The Amino-Terminal Region of Tyk2 Sustains the Level of Interferon Alpha Receptor 1, a Component of the Interferon Alpha/Beta Receptor. Proc Natl Acad Sci U S A 1997, 94, 11839–11844. [Google Scholar] [CrossRef]
- Ragimbeau, J.; Dondi, E.; Alcover, A.; Eid, P.; Uzé, G.; Pellegrini, S. The Tyrosine Kinase Tyk2 Controls IFNAR1 Cell Surface Expression. EMBO J 2003, 22, 537–547. [Google Scholar] [CrossRef]
- Haan, C.; Kreis, S.; Margue, C.; Behrmann, I. Jaks and Cytokine Receptors--an Intimate Relationship. Biochem Pharmacol 2006, 72, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Shuai, K.; Schindler, C.; Prezioso, V.R.; Darnell, J.E. Activation of Transcription by IFN-Gamma: Tyrosine Phosphorylation of a 91-KD DNA Binding Protein. Science 1992, 258, 1808–1812. [Google Scholar] [CrossRef] [PubMed]
- Decker, T.; Kovarik, P.; Meinke, A. GAS Elements: A Few Nucleotides with a Major Impact on Cytokine-Induced Gene Expression. J Interferon Cytokine Res 1997, 17, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E.; Kerr, lan M. ; Stark, G.R. Jak-STAT Pathways and Transcriptional Activation in Response to IFNs and Other Extracellular Signaling Proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- El Kasmi, K.C.; Holst, J.; Coffre, M.; Mielke, L.; de Pauw, A.; Lhocine, N.; Smith, A.M.; Rutschman, R.; Kaushal, D.; Shen, Y.; et al. General Nature of the STAT3-Activated Anti-Inflammatory Response. J Immunol 2006, 177, 7880–7888. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.Y.; Kessler, D.S.; Veals, S.A.; Levy, D.E.; Darnell, J.E. ISGF3, the Transcriptional Activator Induced by Interferon Alpha, Consists of Multiple Interacting Polypeptide Chains. Proc Natl Acad Sci U S A 1990, 87, 8555–8559. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.; Fu, X.Y.; Improta, T.; Aebersold, R.; Darnell, J.E. Proteins of Transcription Factor ISGF-3: One Gene Encodes the 91-and 84-KDa ISGF-3 Proteins That Are Activated by Interferon Alpha. Proc Natl Acad Sci U S A 1992, 89, 7836–7839. [Google Scholar] [CrossRef]
- Pauli, E.K.; Schmolke, M.; Wolff, T.; Viemann, D.; Roth, J.; Bode, J.G.; Ludwig, S. Influenza A Virus Inhibits Type I IFN Signaling via NF-KappaB-Dependent Induction of SOCS-3 Expression. PLoS Pathog 2008, 4. [Google Scholar] [CrossRef]
- Jia, D.; Rahbar, R.; Chan, R.W.Y.; Lee, S.M.Y.; Chan, M.C.W.; Wang, B.X.; Baker, D.P.; Sun, B.; Malik Peiris, J.S.; Nicholls, J.M.; et al. Influenza Virus Non-Structural Protein 1 (NS1) Disrupts Interferon Signaling. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Yang, C.H.; Murti, A.; Pfeffer, L.M. STAT3 Complements Defects in an Interferon-Resistant Cell Line: Evidence for an Essential Role for STAT3 in Interferon Signaling and Biological Activities. Proc Natl Acad Sci U S A 1998, 95, 5568–5572. [Google Scholar] [CrossRef]
- Wang, D.; Stravopodis, D.; Teglund, S.; Kitazawa, J.; Ihle, J.N. Naturally Occurring Dominant Negative Variants of Stat5. Mol Cell Biol 1996, 16, 6141–6148. [Google Scholar] [CrossRef] [PubMed]
- Lin JX; Mietz J; Modi WS; John S; Leonard WJ Cloning of Human Stat5B. Reconstitution of Interleukin-2-Induced Stat5A and Stat5B DNA Binding Activity in COS-7 Cells. J Biol Chem. 1996, 3, 10738–10744. [Google Scholar]
- Abbas, Y.M.; Pichlmair, A.; Górna, M.W.; Superti-Furga, G.; Nagar, B. Structural Basis for Viral 59-PPP-RNA Recognition by Human IFIT Proteins. Nature 2013. [Google Scholar] [CrossRef] [PubMed]
- Vanderven, H.A.; Petkau, K.; Ryan-Jean, K.E.E.; Aldridge, J.R.; Webster, R.G.; Magor, K.E. Avian Influenza Rapidly Induces Antiviral Genes in Duck Lung and Intestine. Mol Immunol 2012, 51, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Fleming-Canepa, X.; Aldridge, J.R.; Canniff, L.; Kobewka, M.; Jax, E.; Webster, R.G.; Magor, K.E. Duck Innate Immune Responses to High and Low Pathogenicity H5 Avian Influenza Viruses. Vet Microbiol 2019, 228, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda Interferon (IFN-Lambda), a Type III IFN, Is Induced by Viruses and IFNs and Displays Potent Antiviral Activity against Select Virus Infections in Vivo. J Virol 2006, 80, 4501–4509. [Google Scholar] [CrossRef]
- Clemens, M.J.; Elia, A. The Double-Stranded RNA-Dependent Protein Kinase PKR: Structure and Function. J Interferon Cytokine Res 1997, 17, 503–524. [Google Scholar] [CrossRef] [PubMed]
- García, M.A.; Gil, J.; Ventoso, I.; Guerra, S.; Domingo, E.; Rivas, C.; Esteban, M. Impact of Protein Kinase PKR in Cell Biology: From Antiviral to Antiproliferative Action. Microbiol Mol Biol Rev 2006, 70, 1032–1060. [Google Scholar] [CrossRef] [PubMed]
- Lakadamyali, M.; Rust, M.J.; Babcock, H.P.; Zhuang, X. Visualizing Infection of Individual Influenza Viruses. Proc Natl Acad Sci U S A 2003, 100, 9280–9285. [Google Scholar] [CrossRef]
- Sun, X.; Whittaker, G.R. Role of the Actin Cytoskeleton during Influenza Virus Internalization into Polarized Epithelial Cells. Cell Microbiol 2007, 9, 1672–1682. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Wolff, T.; Pleschka, S.; Planz, O.; Beermann, W.; Bode, J.G.; Schmolke, M.; Ludwig, S. Influenza A Virus NS1 Protein Activates the PI3K/Akt Pathway to Mediate Antiapoptotic Signaling Responses. J Virol 2007, 81, 3058–3067. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.K.; Liu, Q.; Tikoo, S.K.; Babiuk, L.A.; Zhou, Y. Effect of the Phosphatidylinositol 3-Kinase/Akt Pathway on Influenza A Virus Propagation. J Gen Virol 2007, 88, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Zhirnov, O.P.; Klenk, H.D. Control of Apoptosis in Influenza Virus-Infected Cells by up-Regulation of Akt and P53 Signaling. Apoptosis 2007, 12, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, S. Disruption of Virus-Host Cell Interactions and Cell Signaling Pathways as an Anti-Viral Approach against Influenza Virus Infections. Biol Chem 2011, 392, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza Virus Propagation Is Impaired by Inhibition of the Raf/MEK/ERK Signalling Cascade. Nat Cell Biol 2001, 3, 301–305. [Google Scholar] [CrossRef]
- Wang, C.; Liu, H.; Luo, J.; Chen, L.; Li, M.; Su, W.; Zhao, N.; Liu, S.; Xie, L.; Jia, Y.; et al. HA Triggers the Switch from MEK1 SUMOylation to Phosphorylation of the ERK Pathway in Influenza A Virus-Infected Cells and Facilitates Its Infection. Front Cell Infect Microbiol 2017, 7. [Google Scholar] [CrossRef]
- Schmitz, M.L.; Kracht, M.; Saul, V. V. The Intricate Interplay between RNA Viruses and NF-ΚB. Biochim Biophys Acta 2014, 1843, 2754–2764. [Google Scholar] [CrossRef]
- Mazur, I.; Wurzer, W.J.; Ehrhardt, C.; Pleschka, S.; Puthavathana, P.; Silberzahn, T.; Wolff, T.; Planz, O.; Ludwig, S. Acetylsalicylic Acid (ASA) Blocks Influenza Virus Propagation via Its NF-KappaB-Inhibiting Activity. Cell Microbiol 2007, 9, 1683–1694. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Rückle, A.; Hrincius, E.R.; Haasbach, E.; Anhlan, D.; Ahmann, K.; Banning, C.; Reiling, S.J.; Kühn, J.; Strobl, S.; et al. The NF-ΚB Inhibitor SC75741 Efficiently Blocks Influenza Virus Propagation and Confers a High Barrier for Development of Viral Resistance. Cell Microbiol 2013, 15, 1198–1211. [Google Scholar] [CrossRef]
- Jin, J.; Hu, H.; Li, H.S.; Yu, J.; Xiao, Y.; Brittain, G.C.; Zou, Q.; Cheng, X.; Mallette, F.A.; Watowich, S.S.; et al. Noncanonical NF-ΚB Pathway Controls the Production of Type I Interferons in Antiviral Innate Immunity. Immunity 2014, 40, 342–354. [Google Scholar] [CrossRef]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The Evolution of Phosphatidylinositol 3-Kinases as Regulators of Growth and Metabolism. Nat Rev Genet 2006, 7, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Hirata, N.; Suizu, F.; Matsuda-Lennikov, M.; Edamura, T.; Bala, J.; Noguchi, M. Inhibition of Akt Kinase Activity Suppresses Entry and Replication of Influenza Virus. Biochem Biophys Res Commun 2014, 450, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.; Helenius, A. Vaccinia Virus Uses Macropinocytosis and Apoptotic Mimicry to Enter Host Cells. Science 2008, 320, 531–535. [Google Scholar] [CrossRef]
- Saeed, M.F.; Kolokoltsov, A.A.; Freiberg, A.N.; Holbrook, M.R.; Davey, R.A. Phosphoinositide-3 Kinase-Akt Pathway Controls Cellular Entry of Ebola Virus. PLoS Pathog 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Zhang, J.L.; Chen, W.; Xu, X.F.; Gao, N.; Fan, D.Y.; An, J. Roles of Small GTPase Rac1 in the Regulation of Actin Cytoskeleton during Dengue Virus Infection. PLoS Negl Trop Dis 2010, 4. [Google Scholar] [CrossRef]
- Carter, G.C.; Bernstone, L.; Baskaran, D.; James, W. HIV-1 Infects Macrophages by Exploiting an Endocytic Route Dependent on Dynamin, Rac1 and Pak1. Virology 2011, 409, 234–250. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Kardinal, C.; Wurzer, W.J.; Wolff, T.; Von Eichel-Streiber, C.; Pleschka, S.; Planz, O.; Ludwig, S. Rac1 and PAK1 Are Upstream of IKK-ε and TBK-1 in the Viral Activation of Interferon Regulatory Factor-3. FEBS Lett 2004, 567, 230–238. [Google Scholar] [CrossRef]
- Dierkes, R.; Warnking, K.; Liedmann, S.; Seyer, R.; Ludwig, S.; Ehrhardt, C. The Rac1 Inhibitor NSC23766 Exerts Anti-Influenza Virus Properties by Affecting the Viral Polymerase Complex Activity. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Jiang, W.; Sheng, C.; Gu, X.; Liu, D.; Yao, C.; Gao, S.; Chen, S.; Huang, Y.; Huang, W.; Fang, M. Suppression of Rac1 Signaling by Influenza A Virus NS1 Facilitates Viral Replication. Sci Rep 2016, 6. [Google Scholar] [CrossRef]
- Capitanio, J.S.; Wozniak, R.W. Host Cell Factors Necessary for Influenza A Infection: Meta-Analysis of Genome Wide Studies. 2012. [Google Scholar] [CrossRef]
- Smith, S.B.; Dampier, W.; Tozeren, A.; Brown, J.R.; Magid-Slav, M. Identification of Common Biological Pathways and Drug Targets across Multiple Respiratory Viruses Based on Human Host Gene Expression Analysis. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Bard-Chapeau, E.A.; Li, S.; Ding, J.; Zhang, S.S.; Zhu, H.H.; Princen, F.; Fang, D.D.; Han, T.; Bailly-Maitre, B.; Poli, V.; et al. Ptpn11/Shp2 Acts as a Tumor Suppressor in Hepatocellular Carcinogenesis. Cancer Cell 2011, 19, 629–639. [Google Scholar] [CrossRef]
- Zheng, J.; Huang, X.; Tan, W.; Yu, D.; Du, Z.; Chang, J.; Wei, L.; Han, Y.; Wang, C.; Che, X.; et al. Pancreatic Cancer Risk Variant in LINC00673 Creates a MiR-1231 Binding Site and Interferes with PTPN11 Degradation. Nat Genet 2016, 48, 747–757. [Google Scholar] [CrossRef]
- Pannone, L.; Bocchinfuso, G.; Flex, E.; Rossi, C.; Baldassarre, G.; Lissewski, C.; Pantaleoni, F.; Consoli, F.; Lepri, F.; Magliozzi, M.; et al. Structural, Functional, and Clinical Characterization of a Novel PTPN11 Mutation Cluster Underlying Noonan Syndrome. Hum Mutat 2017, 38, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J.; Alper, D.; Cohen, L.; Leblanc, J.F.; Sportza, L.; Wong, A.; Xanthoudakis, S. Induction of Human Interferon Gene Expression Is Associated with a Nuclear Factor That Interacts with the NF-Kappa B Site of the Human Immunodeficiency Virus Enhancer. J Virol 1989, 63, 2557–2566. [Google Scholar] [CrossRef] [PubMed]
- Lenardo, M.J.; Fan, C.M.; Maniatis, T.; Baltimore, D. The Involvement of NF-Kappa B in Beta-Interferon Gene Regulation Reveals Its Role as Widely Inducible Mediator of Signal Transduction. Cell 1989, 57, 287–294. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Zheng, H.; Muster, T.; Palese, P.; Beg, A.A.; Garcı́a-Sastre, A. Influenza A Virus NS1 Protein Prevents Activation of NF-KappaB and Induction of Alpha/Beta Interferon. J Virol 2000, 74, 11566–11573. [Google Scholar] [CrossRef]
- Lee, J.C.; Laydon, J.T.; McDonnell, P.C.; Gallagher, T.F.; Kumar, S.; Green, D.; McNulty, D.; Blumenthal, M.J.; Keys, J.R.; Land Vatter, S.W.; et al. A Protein Kinase Involved in the Regulation of Inflammatory Cytokine Biosynthesis. Nature 1994, 372, 739–746. [Google Scholar] [CrossRef]
- Lee, D.C.W.; Cheung, C.-Y.; Law, A.H.Y.; Mok, C.K.P.; Peiris, M.; Lau, A.S.Y. P38 Mitogen-Activated Protein Kinase-Dependent Hyperinduction of Tumor Necrosis Factor Alpha Expression in Response to Avian Influenza Virus H5N1. J Virol 2005, 79, 10147–10154. [Google Scholar] [CrossRef]
- Lee, N.; Wong, C.K.; Chan, P.K.S.; Lun, S.W.M.; Lui, G.; Wong, B.; Hui, D.S.C.; Lam, C.W.K.; Cockram, C.S.; Choi, K.W.; et al. Hypercytokinemia and Hyperactivation of Phospho-P38 Mitogen-Activated Protein Kinase in Severe Human Influenza A Virus Infection. Clin Infect Dis 2007, 45, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Jibiki, I.; Asai, Y.; Gon, Y.; Kobayashi, T.; Ichiwata, T.; Shimizu, K.; Hashimoto, S. Analysis of Gene Expression in Human Bronchial Epithelial Cells upon Influenza Virus Infection and Regulation by P38 Mitogen-Activated Protein Kinase and c-Jun-N-Terminal Kinase. Respirology 2008, 13, 203–214. [Google Scholar] [CrossRef]
- Börgeling, Y.; Schmolke, M.; Viemann, D.; Nordhoff, C.; Roth, J.; Ludwig, S. Inhibition of P38 Mitogen-Activated Protein Kinase Impairs Influenza Virus-Induced Primary and Secondary Host Gene Responses and Protects Mice from Lethal H5N1 Infection. J Biol Chem 2014, 289, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, L.S.; Stratakis, C.A. Structure of the Human Ubiquitin Fusion Gene Uba80 (RPS27a) and One of Its Pseudogenes. Biochem Biophys Res Commun 2000, 270, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Tanaka, K. Regulatory Mechanisms Involved in the Control of Ubiquitin Homeostasis. J Biochem 2010, 147, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Q.; Liu, T.; Chang, G.; Sun, Z.; Gao, Z.; Wang, F.; Zhou, H.; Liu, R.; Zheng, M.; et al. Host Interaction Analysis of PA-N155 and PA-N182 in Chicken Cells Reveals an Essential Role of UBA52 for Replication of H5N1 Avian Influenza Virus. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, Q.; Chen, S.; Gao, S.; Song, L.; Liu, P.; Huang, W. Influenza A Virus NS1 Induces G0/G1 Cell Cycle Arrest by Inhibiting the Expression and Activity of RhoA Protein. J Virol 2013, 87, 3039–3052. [Google Scholar] [CrossRef]
Figure 1.
Comparative analysis of upregulated (a) and downregulated (b) proteins expression changes at different time intervals in duck lung tissues.
Figure 1.
Comparative analysis of upregulated (a) and downregulated (b) proteins expression changes at different time intervals in duck lung tissues.
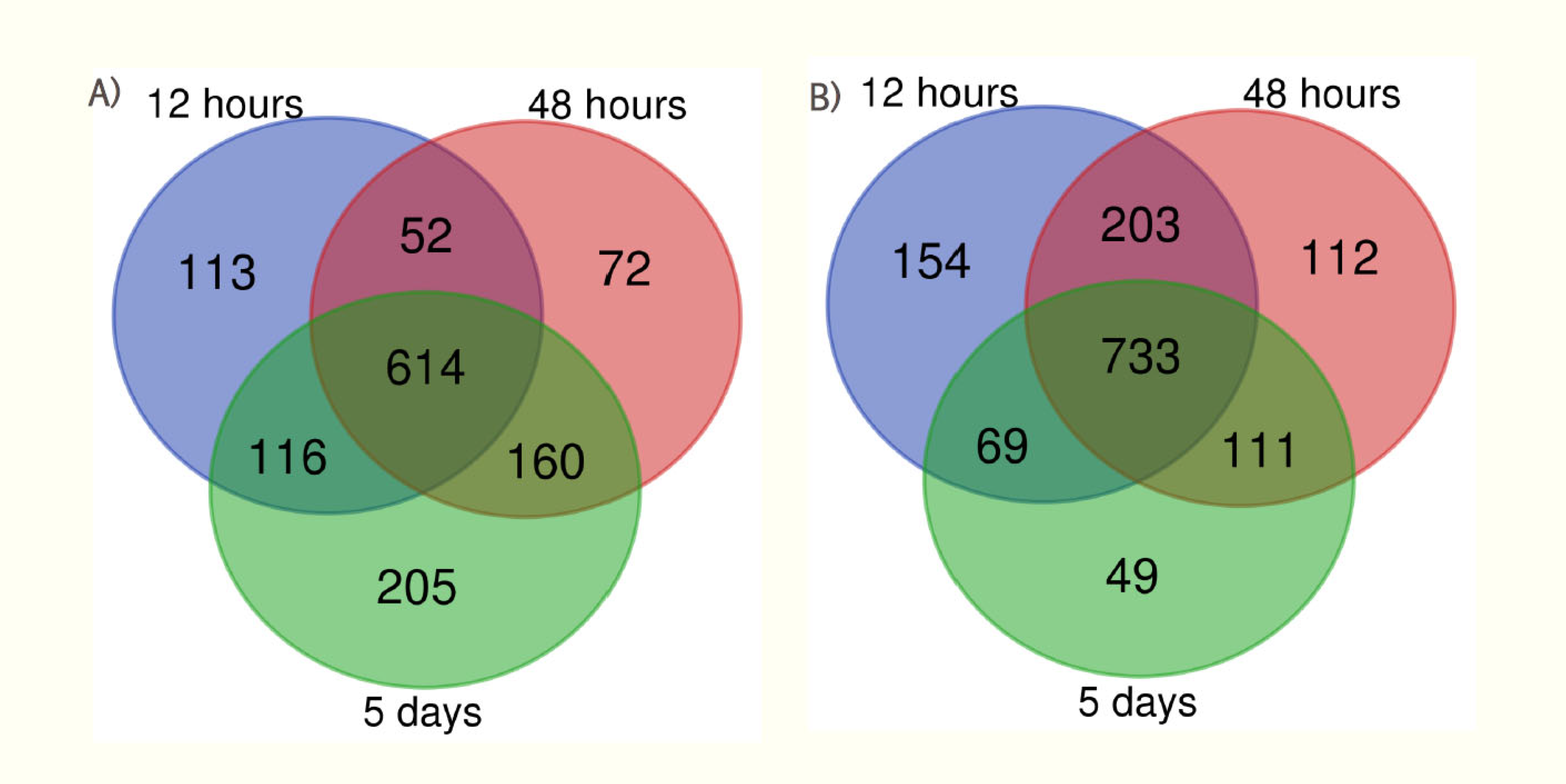
Figure 2.
Heatmap of differentially expressed proteins of HPAIV infected lung tissues. The expression levels are visualized using a gradient colour scheme.
Figure 2.
Heatmap of differentially expressed proteins of HPAIV infected lung tissues. The expression levels are visualized using a gradient colour scheme.
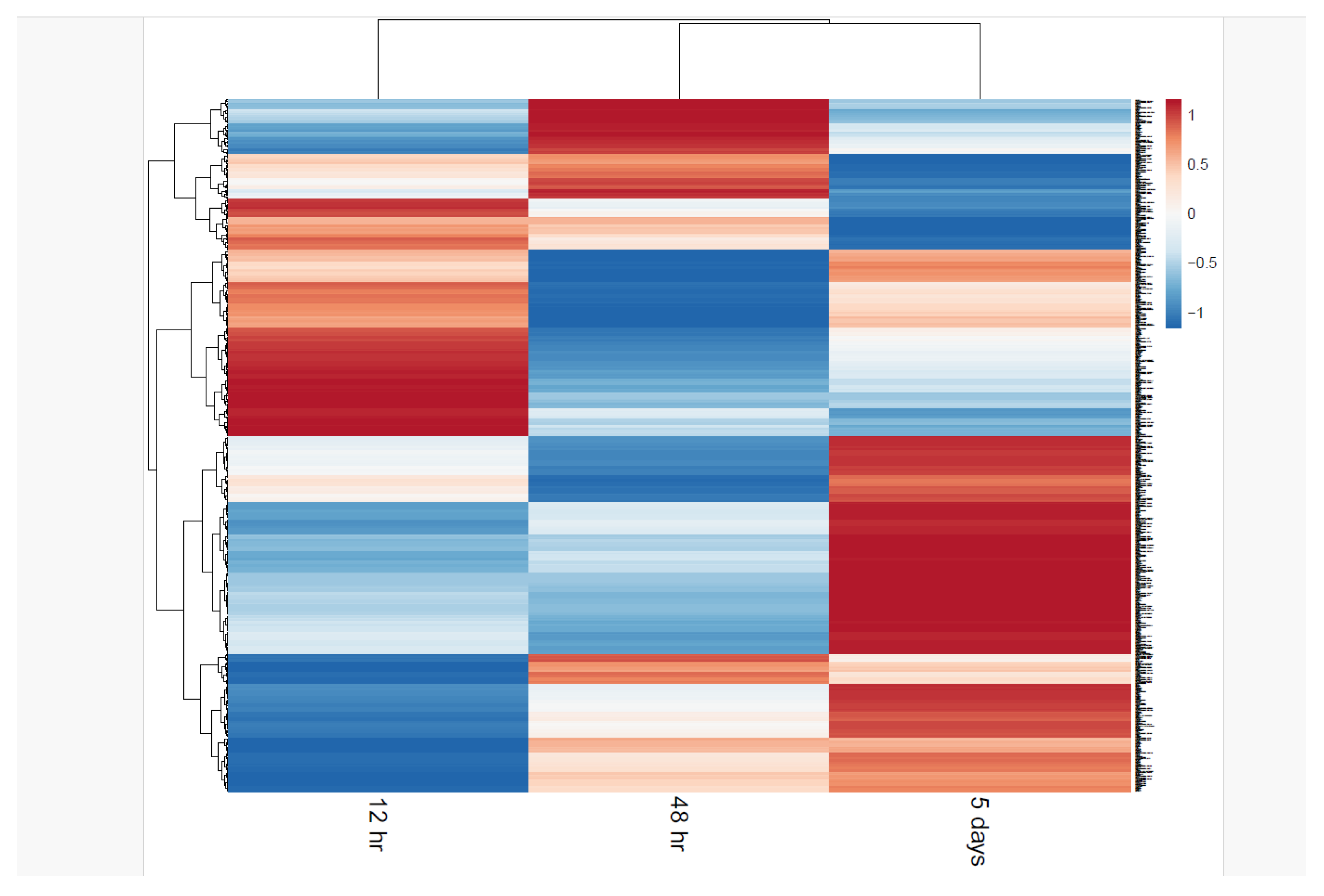
Figure 3.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 12 hr post infection condition. GO terms selected based on cut off P value > 0.05.
Figure 3.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 12 hr post infection condition. GO terms selected based on cut off P value > 0.05.
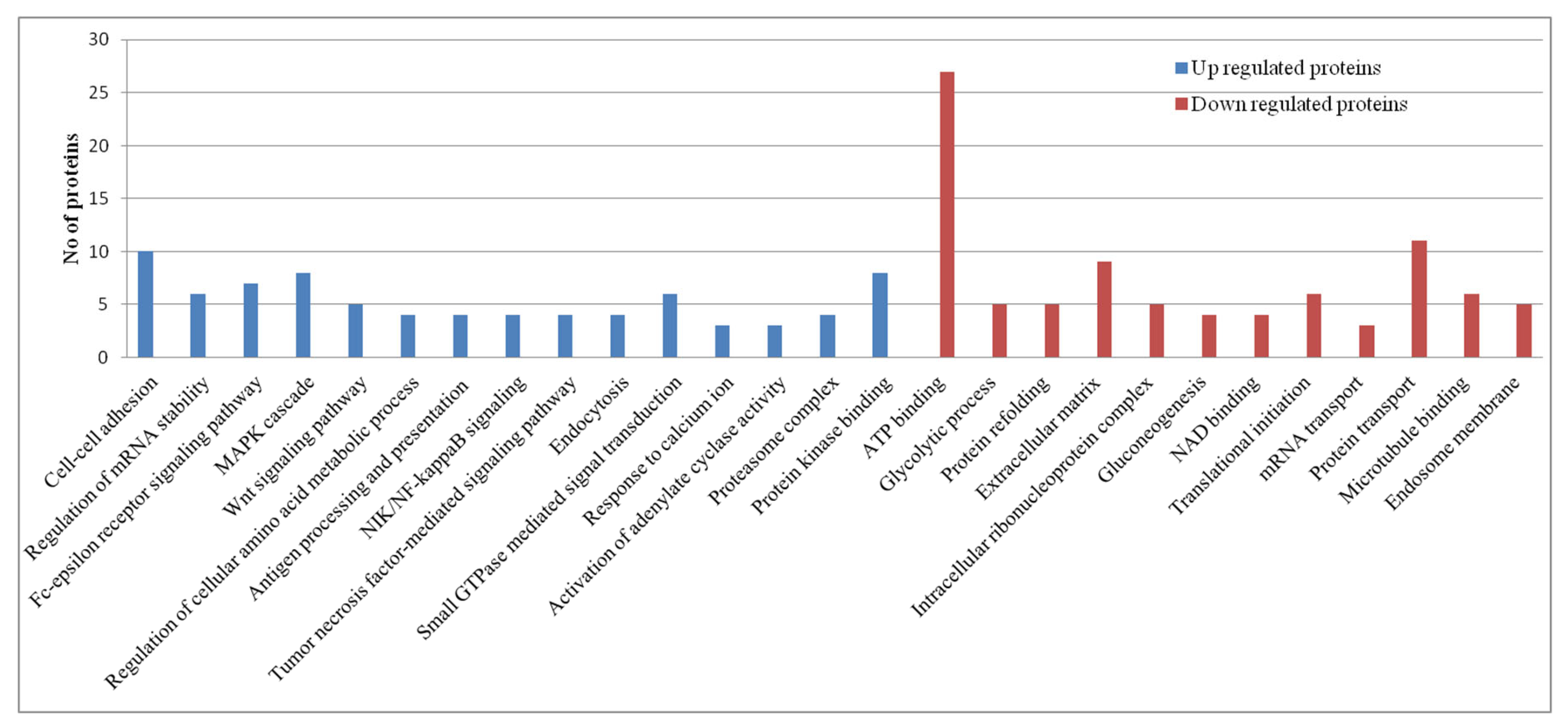
Figure 4.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 48 hr post infection condition. GO terms selected based on cut off P value > 0.05.
Figure 4.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 48 hr post infection condition. GO terms selected based on cut off P value > 0.05.
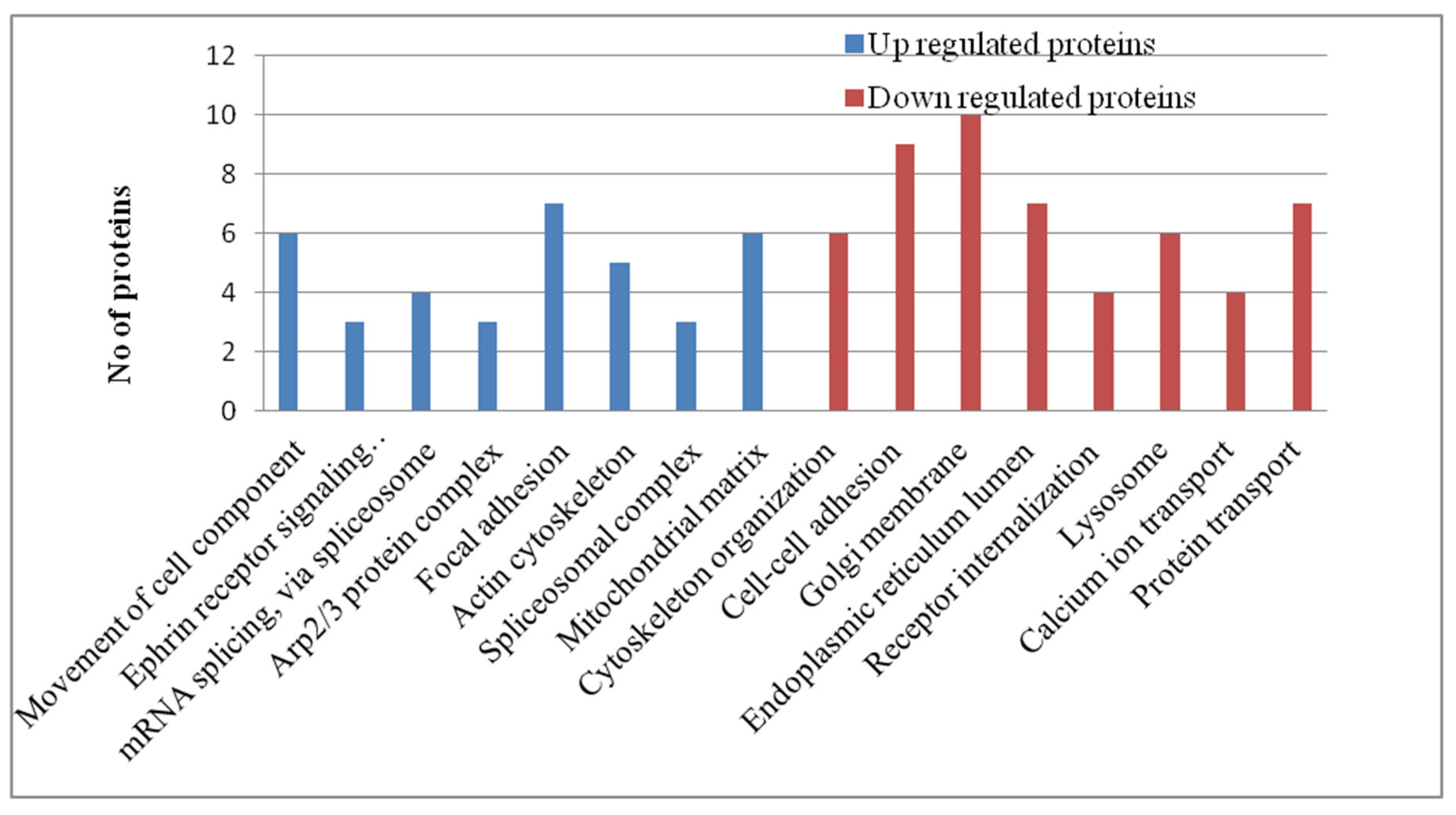
Figure 5.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 5 days post infection condition. GO terms selected based on cut off P value > 0.05.
Figure 5.
Gene ontology term analysis of exclusively upregulated and downregulated proteins in duck lung tissues infected with HPAI H5N1 virus at 5 days post infection condition. GO terms selected based on cut off P value > 0.05.
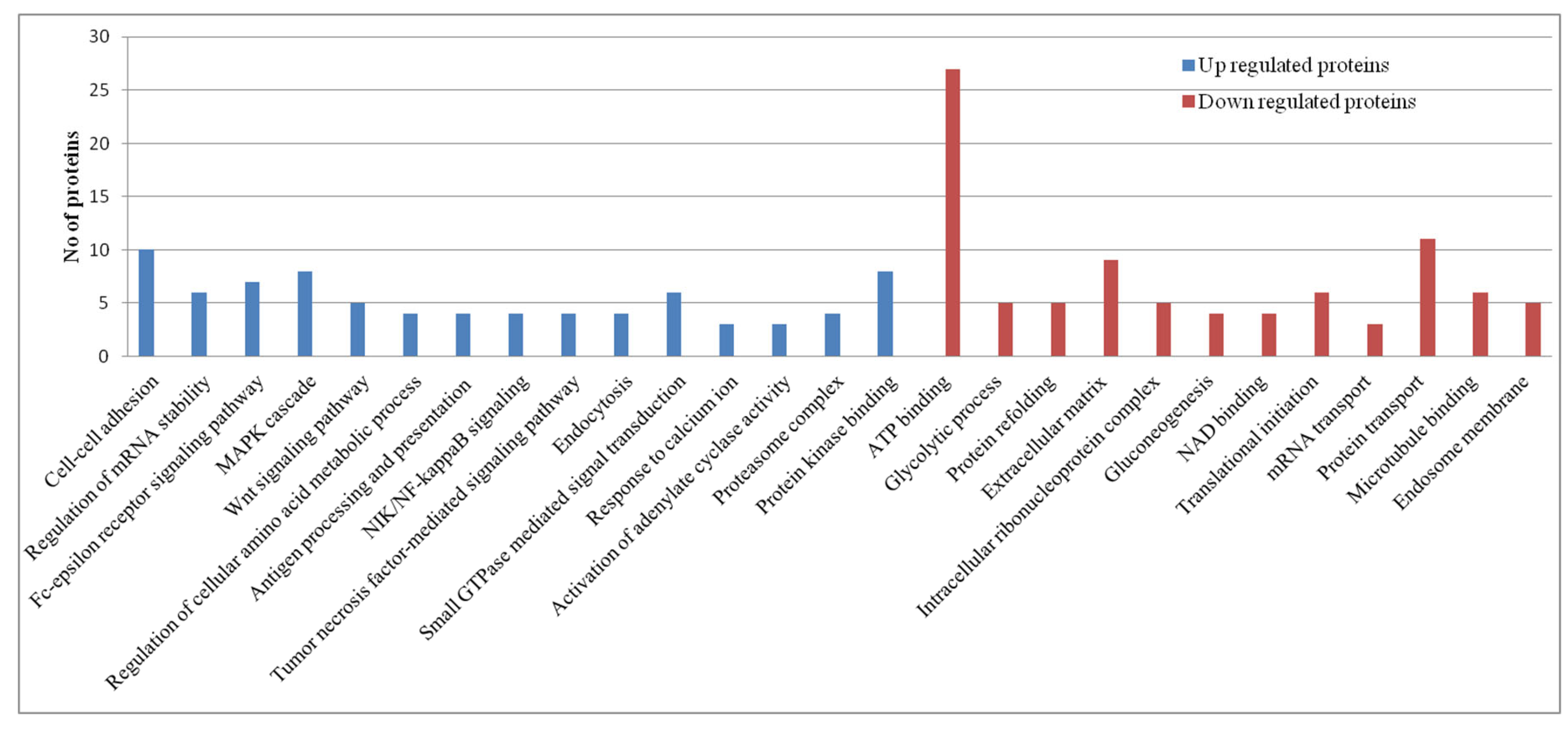
Figure 6.
Expression level of proteins involved in RIG-I-like receptor and Jak-STAT signaling pathways in duck lung tissues infected with HPAIVs.
Figure 6.
Expression level of proteins involved in RIG-I-like receptor and Jak-STAT signaling pathways in duck lung tissues infected with HPAIVs.
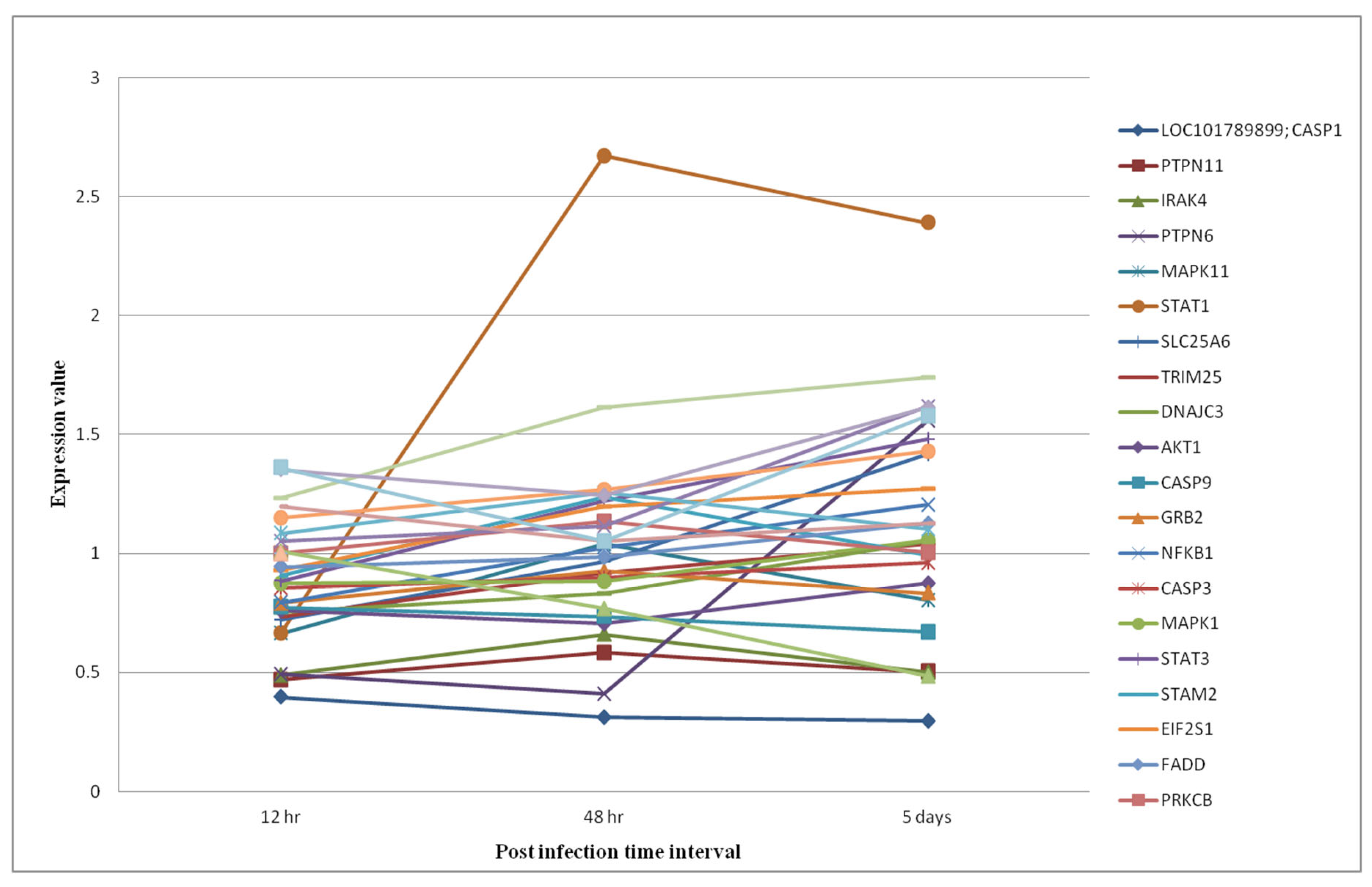
Figure 7.
The protein-protein interaction network of duck lung tissues infected with HPAI H5N1 virus. The important hub proteins involved in influenza pathogenesis were highlighted with red colour in the PPI network.
Figure 7.
The protein-protein interaction network of duck lung tissues infected with HPAI H5N1 virus. The important hub proteins involved in influenza pathogenesis were highlighted with red colour in the PPI network.
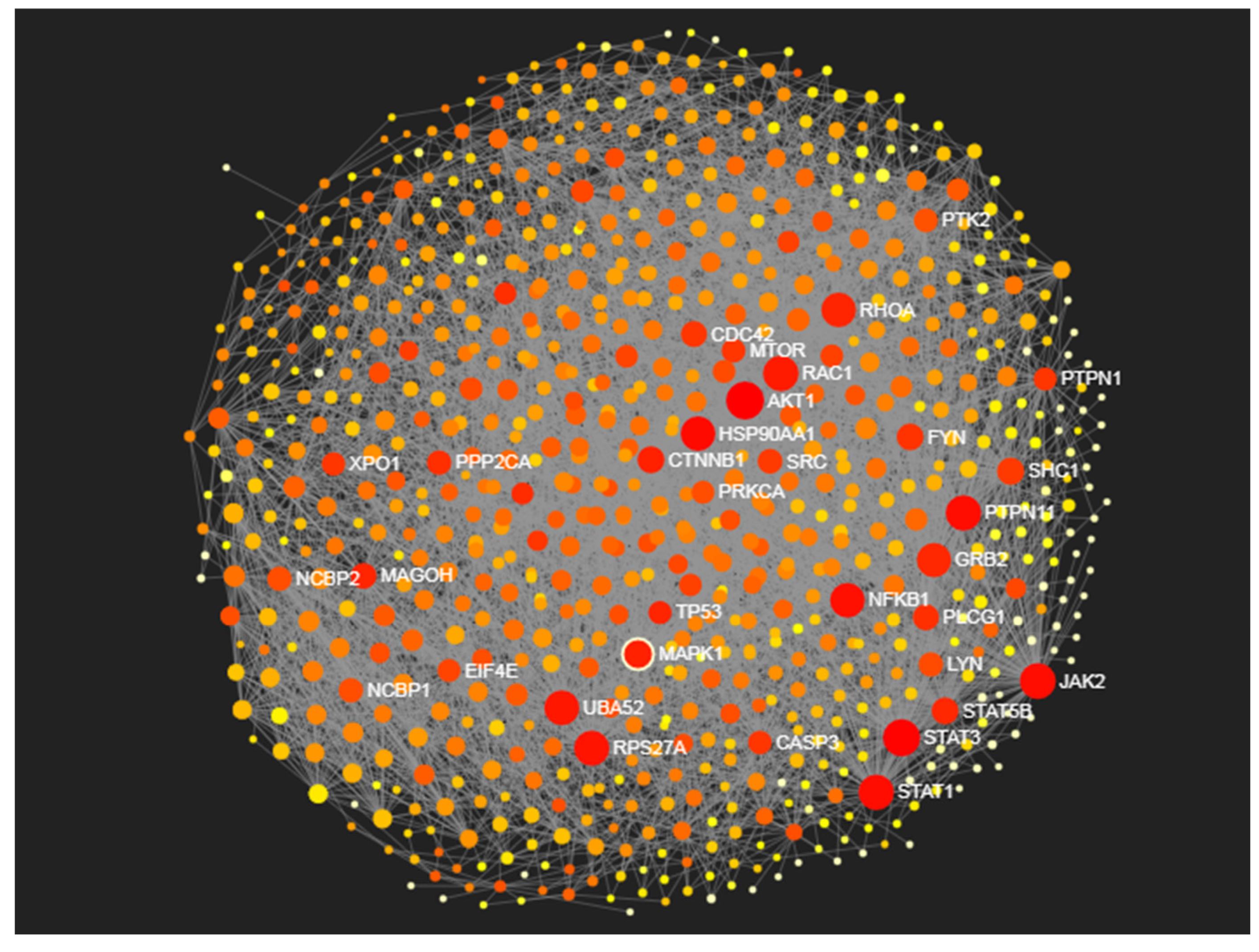

Table 1.
Differential protein expression analysis in ducks infected with HPAI H5N1 virus.
| Condition condition |
Upregulated proteins | Downregulated proteins | Upregulated proteins (>1.5 fold) | Downregulated Proteins (<1.5 fold) |
|---|---|---|---|---|
| 12 hr interval | 895 | 1159 | 128 | 225 |
| 48 hr interval | 898 | 1159 | 199 | 277 |
| 5 days interval | 1095 | 962 | 296 | 264 |
Table 2.
KEGG pathway analysis of differentially expressed proteins in ducks lung tissues infected with HPAI H5N1 virus.
Table 2.
KEGG pathway analysis of differentially expressed proteins in ducks lung tissues infected with HPAI H5N1 virus.
| Pathway | Hits | FDR |
|---|---|---|
| Focal adhesion | 179 | 3.20E-21 |
| PI3K-Akt signaling pathway | 291 | 1.10E-20 |
| MAPK signaling pathway | 243 | 1.12E-17 |
| Cell cycle | 116 | 1.97E-17 |
| Endocytosis | 204 | 2.00E-16 |
| AMPK signaling pathway | 110 | 5.46E-15 |
| Regulation of actin cytoskeleton | 179 | 1.36E-14 |
| Phospholipase D signaling pathway | 130 | 3.91E-14 |
| T cell receptor signaling pathway | 94 | 4.54E-14 |
| Chemokine signaling pathway | 159 | 3.77E-13 |
| Apoptosis | 119 | 8.99E-13 |
| Spliceosome | 117 | 1.91E-12 |
| Platelet activation | 109 | 4.49E-12 |
| Ubiquitin mediated proteolysis | 117 | 4.00E-11 |
| mTOR signaling pathway | 128 | 8.53E-11 |
| Fc epsilon RI signaling pathway | 64 | 1.68E-10 |
| Influenza A | 137 | 2.43E-10 |
| Jak-STAT signaling pathway | 132 | 1.20E-09 |
| Protein processing in endoplasmic reticulum | 134 | 1.32E-09 |
| Proteasome | 44 | 4.35E-09 |
| Toll-like receptor signaling pathway | 88 | 2.57E-08 |
| RIG-I-like receptor signaling pathway | 62 | 1.09E-07 |
| Metabolic pathways | 926 | 1.26E-07 |
| TNF signaling pathway | 91 | 1.26E-07 |
| Th1 and Th2 cell differentiation | 77 | 4.78E-07 |
| Leukocyte transendothelial migration | 91 | 5.95E-07 |
Table 3.
KEGG pathway analysis of differentially expressed phosphoproteins in ducks lung tissues infected with HPAI H5N1 virus.
Table 3.
KEGG pathway analysis of differentially expressed phosphoproteins in ducks lung tissues infected with HPAI H5N1 virus.
| Pathway | Hits | P.Value |
|---|---|---|
| Jak-STAT signaling pathway | 5 | 0.0191 |
| NF-kappa B signaling pathway | 5 | 0.0026 |
| MAPK signaling pathway | 14 | 5.84E-07 |
| PI3K-Akt signaling pathway | 22 | 7.33E-13 |
| Focal adhesion | 34 | 2.84E-35 |
| Regulation of actin cytoskeleton | 24 | 5.77E-20 |
| ErbB signaling pathway | 14 | 3.30E-14 |
| Leukocyte transendothelial migration | 14 | 1.71E-12 |
| Ras signaling pathway | 16 | 3.65E-10 |
| Rap1 signaling pathway | 15 | 6.58E-10 |
| T cell receptor signaling pathway | 10 | 3.29E-08 |
| Chemokine signaling pathway | 12 | 1.97E-07 |
| Phospholipase D signaling pathway | 10 | 1.22E-06 |
| Tight junction | 9 | 3.13E-05 |
| Endocytosis | 8 | 0.00218 |
| Prolactin signaling pathway | 4 | 0.00439 |
Table 4.
Hub proteins identified in ducks PPI networks based on degree centrality and betweenness centrality measures.
Table 4.
Hub proteins identified in ducks PPI networks based on degree centrality and betweenness centrality measures.
| Proteins | Degree centrality | Betweenness centrality |
|---|---|---|
| AKT1 | 151 | 23549.86 |
| STAT3 | 138 | 20538.68 |
| JAK2 | 122 | 14684.79 |
| RAC1 | 117 | 9430.98 |
| STAT1 | 116 | 14916.66 |
| PTPN11 | 116 | 14221.68 |
| RPS27A | 113 | 11935.64 |
| UBA52 | 110 | 11175.27 |
| HSP90AA1 | 107 | 17096.24 |
| RHOA | 107 | 7556.94 |
| NFKB1 | 106 | 13320.84 |
| GRB2 | 103 | 6744.12 |
| MAPK1 | 100 | 8118.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



