
Submitted:
10 April 2024
Posted:
11 April 2024
You are already at the latest version
Abstract
Bergamot flavonoids have been shown to prevent metabolic syndrome, non-alcoholic fatty liver disease (NAFLD) and stimulate autophagy in animal models and patients. To investigate further the mechanism of polyphenol-dependent effects, we performed RT2-PCR array analysis on 168 metabolism, transport and autophagy-related genes expressed in rat livers exposed for 14 weeks to different diets: standard, cafeteria (CAF) and CAF diet supplemented with 50 mg/kg of Bergamot Polyphenol Fraction (BPF). CAF diet caused a strong upregulation of the gluconeogenesis pathway (Gck, Pck2) and a moderate (>1.7 fold) induction of genes regulating lipogenesis (Srebf1, Pparg, Xbp1), lipid and cholesterol transport or lipolysis (Fabp3, Apoa1, Lpl) and inflammation (Il6). However, only one -oxidation gene (Cpt1a) and a few autophagy genes were modulated in the CAF group compared to controls. While most of these transcripts were significantly modulated by BPF, we observed a particularly potent effect on lipogenesis genes, like Acly, Acaca, Fasn and Scd1, which were suppressed far below the mRNA levels of control livers as confirmed by alternative primers-based RT2-PCR analysis and western blotting. These effects were accompanied by downregulation of pro-inflammatory cytokines (Il6, Tnfa, and Il10) and diabetes-related genes. Few autophagy (Map1Lc3a, Dapk) and no -oxidation gene expression changes were observed, suggesting minor transcriptional effects on autophagy. In conclusion, chronic BPF supplementation efficiently prevents NAFLD by modulating hepatic energy metabolism and inflammation gene expression programs, with no effect on -oxidation, but profound suppression of de-novo lipogenesis.
Keywords:
flavonoids
; hepatic steatosis
; lipogenesis
; Citrus Bergamia
; gene profiling
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) and its more advanced form (NASH), are the most common liver disorders in industrialized countries, caused by fat and sugar-rich diet, sedentary lifestyle and genetic predisposition [1,2,3]. To date, there are no specific drugs approved for NAFLD [4,5], but antioxidant polyphenols, especially in the form of plant-derived extracts are emerging as an important therapeutic option for the management of NAFLD and NASH [6,7] in addition to dietary measures and physical activity. In this scenario, Bergamot Polyphenol Fraction (BPF) appears as a particularly promising food supplement. Several preclinical studies documented the efficacy of BPF against hepatic steatosis in rodent models of diet-induced NAFLD and NASH as both a preventive measure or post-induction treatment and clinical studies BPF® obtained from juice and peels of Bergamot (Citrus Bergamia Risso et Poiteau) fruits exceptionally rich in flavonoids, is characterized by a unique profile of flavonoids such as naringin, brutieridin and melitidin and many other flavonoid and non-flavonoid compounds with lipid-lowering, anti-inflammatory, proautophagic and detoxifying activity [8,9,10,11]. The chemical composition and beneficial effects of BPF antioxidants have been documented in numerous analytical [8,9,12], preclinical [10,13,14,15] and clinical studies [16,17,18,19,20], but little is known about molecular mechanisms underlying these effects.
Hepatic lipid accumulation is a product of an imbalance between fatty acid (FA) synthesis and FA oxidation, but defective lipolysis and lipophagy, esterification of plasma-free FA or increased dietary fat intake also play a role in steatosis pathophysiology [21]. FA synthesis is the first step of de novo lipogenesis (DNL), by which lipids are endogenously synthesized from acetyl-CoA. The first reaction of endogenous FA synthesis is the conversion of citrate or acetate to acetyl-CoA by the action of citrate lyase (ACLY) or hepatic ACSS2, respectively [22]. Acetyl-CoA is then converted to malonyl-CoA by acetyl-CoA carboxylase (ACACA). FA synthase (FAS) sequentially utilizes malonyl-CoA to extend the growing fatty acyl chains, which can be elongated and desaturated by stearoyl-CoA desaturase (SCD1). The esterification of FA produces triglycerides and other lipids and its final step is catalyzed by diacylglycerol acyltransferase (DGAT) among other enzymes. Most lipogenic enzymes are upregulated to a different extent in high-fat diet [23,24,25], fructose or sucrose [22,26,27] or Western diet-induced [28] animal models of NAFLD and NASH, but their differential expression has never been investigated in CAF diet compared to normal diet livers.
The induction of DNL and their catabolism are tightly regulated at the transcription level [29]. Proteins coded by Sterol regulatory element-binding factor (SREBF1), such as SREB protein 1A (SREB1A) and 1C (SREBP1C), liver X receptors (LXRs), carbohydrate-responsive element-binding protein (ChREBP) and X-box binding protein 1 (XBP-1), which also regulates endoplasmic reticulum (ER) stress, play critical roles in this process. Other transcriptional factors involved in FA synthesis are the peroxisome proliferator-activated receptors (PPARs) that can either enhance genes involved in β-oxidation/translocation of FA (PPAR-α) or promote lipogenesis and glucose uptake and storage (PPAR-γ). SREBP-2 activates genes involved in sterol metabolism and it is the predominant transcriptional factor affecting cholesterol synthesis [30]. The gene expression of the above transcriptional modulators is also altered in patients with NAFLD/NASH and in animal models of the disease [29]. Excessive lipogenesis and β-oxidation of FA are opposing processes that strongly contribute to the oxidative imbalance in hepatocytes and other cells [31].
The vast scientific literature suggests that phenolic phytochemicals may (1) interact with specific proteins in signaling pathways and modulate the activity and/or expression of key antioxidant proteins; (2) regulate the epigenetic mechanisms of gene expression; and (3) modulate the gut microbiota profile and metabolites. For example, flavone- and tyrosol derivatives are implicated in the activation of AMP kinase (AMPK) and as a consequence in the regulation of several metabolic enzymes that play a role in NAFLD [32,33]. In addition, AMPK stays at the crossroads of autophagy and energy (lipid and carbohydrate) metabolism and AMPK activation should explain both hypolipemic and proautophagic proprieties of these compounds in hepatic steatosis [34,35]. AMPK activation has been shown to be mediated by the direct interaction of certain polyphenols with phosphodiesterases (PDEs) [36], but it can be also modulated by NRH-quinone oxidoreductase 2 (NQO2) [32] and possibly by other enzymes among more than 5000 proteins predicted as direct targets of these compounds [37].
The interaction of flavonoids with their protein targets leads to short- and long-term gene expression changes. The latter are mediated by epigenetic mechanisms such as DNA methylation, histone acetylation and deacetylation, causing spatial reorganization of the chromatin [38,39]. In fact, flavones and other dietary polyphenols have been shown to exhibit epigenetic modulatory effects on several gene targets [40,41], which are epigenetically altered in NAFLD [42,43].
Autophagy is a catabolic pathway that contributes to liver homeostasis through its role in cell quality control, by removing misfolded proteins, damaged organelles and lipid droplets [44]. Autophagy and energy metabolism gene expression programs are interconnected [45,46]. Lipid metabolism and autophagy are orchestrated by several common transcription factors, such as transcriptional factor EB (TFEB), a master regulator of lysosomal biogenesis, autophagy and lipid catabolism. Specifically, TFEB is activated in mouse livers and C. elegans upon starvation, while it is downregulated after the administration of a high-fat diet (HFD) to mice, likely modulating the expression of autophagy-related genes [47,48]. Accordingly, autophagy is considered to be an important therapeutic target in the management of NAFLD and NASH [34,44,49]. It is well-known that flavonoids and their phytocomplexes induce autophagy in many cell types including in vivo models of NAFLD [50,51]. It is more difficult to study autophagy in vivo, but there is well-documented evidence that NAFLD and NASH-inducing diets suppress hepatic autophagy levels [48,52]. In addition, our group has demonstrated that Citrus Bergamia flavonoids induce evident bulk and lipid autophagy (lipophagy), detectable after three months of chronic supplementation of BPF, while efficiently preventing NAFLD in rats treated with CAF diet [10].
Here, we evaluated the impact of chronic supplementation of BPF in CAF-treated rats on mRNA levels of 84 genes representing different energy metabolism pathways, often altered in hepatic steatosis and 84 genes representing autophagy machinery. Our data clearly show that suppression of BPF hepatic, de-novo lipogenesis at the level gene expression of and is the most important target of bergamot polyphenols, while no b-oxidation most of autophagy genes is not subjected to transcriptional regulation by polyphenols.
2. Materials and Methods
2.1. Animal Procedures and Experimental Design
Male 5-week-old Rcc: Han WISTAR rats (Harlan Laboratories, Indianapolis, IN, USA) were housed two rats/cage in the animal house facility had access to water and standard chow (SC) diet 2016 (“SC,” Harlan Teklad) or SC and CAF diet ad libitum and maintained in standard conditions as previously described [10]. CAF diet included different sweet or briny foods and condensed milk. The exact composition and feeding protocol has been previously described [10]. All animal studies were approved by the Italian Health Ministry and by the local ethics committee. At 8 weeks of age, the rats were weighed, marked on the tail for recognition, and randomly assigned to two basic experimental groups: CAF diet group (CAF, n = 10 rats) or SC diet group (SC, n = 5 rats). CAF group was subsequently subdivided into two subgroups, of which one received BPF extract (~50 mg/kg body weight/day) as a supplement in drinking water (CAF+BPF, n = 5) and the other received drinking water without BPF (CAF, n = 5). After a week of adaptation to the new cage mate, the administration of CAF diet started (day “0”) and lasted 91–94 days until the day of sacrifice. Food consumption and body weight gain were monitored weekly for 14 weeks. The animals were sacrificed under Zoletil (80mg/kg) and Dormitor anesthesia for tissue collection.
The blood was collected by cardiac puncture as previously described [10] and cholesterol and triglycerides analyses were performed using commercial reagents on a Dimension EXL analyzer (Siemens Healthcare Diagnostics s.r.l.,Milano, Italy).
2.2. Gene Expression Analysis
3 representing rats were chosen for each experimental group (SC, CAF, CAF+BPF). Small pieces (0.4 - 1 g) of the central part of the main lobe of rat livers were shock-frozen in liquid nitrogen and stored until needed at -80°C. Frozen tissue was further fragmented and 50-100 mg samples were homogenized with a glass bouncer on ice with 1mL of TRIzol Reagent (Cat. No. 15596026, Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Total RNA (totRNA) was extracted using the TRIzol Reagent method followed by DNase treatment (Cat. No. 79254, Qiagen Gmbh, Hilden, Germany). Total RNA was carefully quantified, and its integrity was verified on 0.8% denaturing agarose gel. Equal amounts of totRNA from 3 rats of the same experimental group were pooled and cDNA was synthesized from 500 ng of pooled RNA with RT2 PreAMP cDNA Synthesis Kit (Cat. No. 330451; Qiagen Gmbh) according to manufacturer instructions. The relative gene expression was assayed on 96-well format arrays: Rat Fatty liver (PARN-157Z) and Rat autophagy (PARN-084Z) RT2 Profiler PCR arrays (SABiosciences, Qiagen Gmbh), each containing 84 key genes, likely involved in the pathogenesis of hepatic steatosis and the regulation of autophagy, respectively. Each plate included also six housekeeping genes and control wells for genomic DNA (gDNA) contamination and RT-PCR efficiencies. RT-PCR was performed on an iQ5 real-time PCR (Bio-Rad Laboratoires, Inc., Hercules, CA, USA) using SYBR Green (universal cycling conditions: 95°C, 10 min; 95°C, 15 s; and 60°C, 1 min; repeated for 40 cycles) and subsequent analyses were carried out according to the manufacturer’s recommended protocol (SABioscience, Qiagen Gmbh). Melt curve analysis confirmed the amplification of a single product. Each pooled cDNA was analysed twice on independent plates to calculate the mean. This and further analysis was performed by using a dedicated software available at the Gene Globe Data Analysis Center (http://www.qiagen.com/it/shop/genes-and-pathways/data-analysis-center-overview-page/). Briefly, two housekeeping (hk) control genes—hypoxanthine phosphoribosyl transferase (Hprt1) and B2-microglobulin (B2m) were selected for normalization. Each replicate cycle threshold (CT) was normalized to the average CT of hk controls on a per-plate basis. The comparative CT method was used to calculate the relative quantification of gene expression. The following formula was used to calculate the relative amount of the transcripts in the treated samples (CAF or CAF+BPF) and the control samples (SC or CAF), both of which were normalized to hk controls: ΔΔCT =ΔCT (tt) –ΔCT (con). ΔCT is the log2 difference in CT between the target gene and hk controls by subtracting the average CT of controls from each replicate. The fold change for each treated sample relative to the control sample = 2–ΔΔCT.
2.3. RT-qPCR Analysis of Gene Expression
For standard RT-qPCR analysis, totRNA was isolated as described above from 5 rats for each experimental group, but RNA from each rat was processed separately. cDNA was synthesized from 5 mg of totRNA with TransScript® II First-Strand cDNA Synthesis SuperMix (Cat. No. AH301-02) according to manufacturer instructions (TransGen Biotech Co., Ltd, Haidian District, Beijing, China). RT-qPCR was performed on QuantStudio 3 Real-Time PCR Detection System (Applied Biosystems Europe, Monza (MI), Italy) as previously described [15]. The list of used primers is shown in Table 1.
The applied cycling conditions were: 95 °C, 10 min; 95 °C, 15 s; and 61 °C, 1 min; repeated for 40 cycles. Samples were analyzed in triplicate with hypoxanthine phosphoribosyl transferase 1 (Hrpt1) as a HK control. Only results with the amplification of a single product, as verified by melting curve analysis, were considered. Relative gene expression was calculated according to the 2ΔΔCT method using the SC liver tissue cDNA (15 weeks on SC diet) as a starting point control.
2.4. Liver Histology and Lipid Droplets (LDs) Staining and Analysis
10 μm thick frozen sections of the perfused liver from the central portion of the main lobe were prepared as previously described [10]. For lipid droplets (LDs) staining, the liver sections mounted on slides were rinsed with distilled water and immersed in 60% isopropanol (Sigma-Aldrich, St. Louis, MO, USA) and then LDs were stained with freshly prepared Oil Red O (ORO, Sigma-Aldrich) working solution for 15 min. The sections were then rinsed with 60% isopropanol and washed with de-ionized water three times for 30 sec. Then, half of the sections were counterstained with Mayer’s hematoxylin (05-06002/L; Bio-Optica Milano Spa, Milano, Italy) for 60 sec to visualize the nuclei, rinsed with running tap water for 10 min and then covered with a film of embedding medium SlowFade Gold (Cat. No. S36936; Life Technologies, Thermo Fischer Scientific, Road Grand Island, NY, USA) and a coverslip. Equivalent ORO and ORO plus hematoxylin-stained liver sections of each experimental group were examined in bright-field with Leica Microscope DM4000B (Leica Microsystems GmbH, Wetzlar, Germany) equipped with 10×, 40×, and 100× objective lenses. For quantitative LDs accumulation analysis, digital images of ORO-stained sections were acquired by Sp2 confocal microscopy (Leica Microsystems GmbH) by using a TRITC wide emission filter (540–580 nm) and accumulation settings. For each liver section, at least five independent images from equivalent central lobe areas were captured at 40× magnification. Within each area, four different regions were chosen randomly and acquired with zoom at 4.65. These images were processed and analyzed using a semiautomatic procedure implemented in Image J2x software.
2.5. Tissue Homogenization and Western Blotting (WB)
Liver fragments were homogenized on Bullet Blender Storm 24 tissue homogenizer (Next Advance, Inc., Averill Park, NY, USA) according to manufacturer indications, using 1 mm diameter Zirconium Oxide Beads (Next Advance) at 4°C in RIPA lysis buffer [Tris-HCl 20 mM (pH 7,5) NaCl 150 mM, Igepal 1%, EDTA 1 mM, SDS 0,1%] supplemented with protease inhibitors (cOmplete Mini, EDTA free, REF 11836170001; Roche Diagnostics Gmbh, Mannheim, Germany), NaF 2 mM and sodium orthovanadate 2 mM. Protein concentration was evaluated and then 40 µg was subjected to gradient 4-12% Bis-Tris electrophoresis NuPAGE (#NP0335BOX) according to manufacturer’s instructions (Invitrogen, Thermo Fisher Scientific) or 12% SDS- PAGE (for anti-LC3 and ATG16 WB). WB was performed on polyacrylamide gel and the primary antibody was usually incubated overnight at 4°C, followed by 1-h incubation with a secondary antibody at RT. Blots were developed with ImmunoBlot ECL reagents (Cat. # 170-5061; Bio-Rad Laboratories, Inc.).
2.6. Antibodies
The antibodies used for WB were as follows: rabbit polyclonal (rp) anti-GCK (H-88) (sc-7908; Santa Cruz Biotechnology Inc., Dallas, TX, USA; 1:1000); rp anti-PCK2/PEPCK (sc-32879; Santa Cruz Biotech. Inc.; 1:400); rp anti-ACLY (Cat. No. 15421-1-AP; Proteintech Group, Inc. Rosemont, IL 60018, USA; 1:1000); mouse monoclonal (mm) anti-ACACA/ACC (Cat. No: 67373-1-Ig, Proteintech Group, Inc.; 1:1000); rp anti-ATG16 (Code No. PM040Y; MBL International., Woburn, MA, USA; 1:1000); rp anti-ADRP/Perilipin 2 (Cat. No. 15294-1-AP, Proteintech Group, Inc.); mm anti-α-tubulin (T6074, Sigma Aldrich, Darmstadt, Germany; 1:1000); rp anti-LC3 (Code No. M186-3 MBL International; 1:1000); rp anti-GAPDH (sc-87752, 1:500; Santa Cruz Biotech. Inc.; 1:500) were used as primary antibodies.
2.7. Data Analysis and Statistical Procedures
WB optical density was analyzed as previously described [53]. Each liver lysate (from 1 rat) was analyzed at least twice by WB, and the results were expressed as the mean ± standard error (SEM). The data were evaluated using ordinary one-way ANOVA, followed by Tukey’s post-test and occasionally by uncorrected Fisher’s Least Significant Difference (LSD) test as indicated in the legends, for normal distribution of data and Mann-Whitney test when the data were not normally distributed. Data from the experiments are expressed as the mean ± SEM or mean ± standard deviation (SD) for the number n of independent experiments. The between-group differences were considered significant at p < 0.05.
3. Results
3.1. Bergamot Polyphenols Efficiently Prevent CAF Diet-Induced Hepatic Steatosis in Rats
CAF diet supplementation for 15 weeks according to Figure 1 induced typical NAFLD-related disorders in Wistar male rats. CAF diet-fed rats showed considerably higher body weight (Figure 2A), higher blood triglycerides (TGL) (Figure 2B), liver fat accumulation (Figure 2D) and a grade of micro and macro-steatosis in the liver (Figure 3). The undesired effects of CAF diet were mitigated when it was supplemented with BPF (50 mg/kg/rat daily). Although the bioactive compounds did not exert a significant effect on final body weight (Figure 2A), they considerably reduced TGL and cholesterol levels in the blood (Figure 2B,C), as well as liver fat accumulation (Figure 2D). A potent reduction of hepatic steatosis by BPF was confirmed by a strong suppression of an LD-coating protein ADRP/Perilipin 2 in liver specimens (Figure 2E,F) as well as by oil red staining which allows sensitive detection of intracellular TG and cholesterol esters.
This technique revealed the accumulation of numerous, considerably larger LDs in hepatocytes of CAF-fed rats (Figure 3B) compared to SC-fed rats (Figure 3A). BPF supplementation strongly attenuated hepatic LDs accumulation (Figure 3C). These observations were confirmed by the analysis of numbers and size of lipid droplets (LD) in oil-red stained liver sections (Figure 3G). Hematoxylin staining revealed important differences in liver histology: in SC and groups, liver parenchyma appeared homogeneous, and the hepatocytes showed uniform size with large rounded nuclei usually located in the center of the cells and cytoplasmic glycogen granules, with few LDs (Figure 3D). In contrast, the cytoplasm of CAF hepatocytes appeared highly vacuolated, rich in both glycogen granules and LDs (Figure 3E). Importantly, CAF+BPF livers resembled SC livers with moderately increased oil red staining (Figure 3F).
3.2. Bergamot Polyphenols Strongly Suppress Lipogenesis-Related Genes in the Liver
To investigate the transcriptional changes induced by CAF diet and the chronic effects of BPF, we performed RNA profiling of key genes that might play a role in the pathogenesis of liver steatosis. The quantitative (q) PCR-array analysis included a total of 84 genes (Figure 4C) belonging to the pathways of insulin and adipokine signaling, b-oxidation, cholesterol and lipid metabolism and transport, carbohydrate metabolism, inflammatory response and apoptosis. Compared to the control SC diet, the expression of 15 genes was considerably changed in the liver of rats fed with CAF diet (Figure 4A).
As expected for a diet rich in simple sugars, CAF diet upregulated genes involved in glucose utilization (Gck and Pck2) and its conversion into lipids, although lipogenesis genes (Acly, Acaca and Fasn) were only moderately induced in RT2 array experiment. In CAF vs SC group (Figure 4B) 10 genes related to Lipid Transport (Fabp3, Lpl), Carbohydrate Metabolism (Gck, Pck2), Endoplasmic Reticulum (ER) stress (Xbp1) Inflammatory Response (Il6) cholesterol metabolism and transport (Apoa1, Lepr, Pparg, Srebf1) were upregulated and 5 genes related to Adipokine signaling (Cd36 and Serpine1), Beta Oxidation (Cpt1a), Insulin signaling (Insr) and Lipid Metabolism (Hnf4a) were downregulated. Interestingly in CAF+BPF vs CAF experimental groups (Figure 4D) 23 genes related to Cholesterol and Lipid Metabolism (Abcg1, Apoa1, Ldlr, Lepr, Nr1h3, Srebf2, Acaca, Fabp5, Fasn, Lpl, Scd), Insulin and Adipokine Signaling (Akt1, Slc2a1, Slc2a4, Ppargc1a), Carbohydrate Metabolism (Acly, G6pd, Gck, Pck2), Inflammatory Response (Il10, Il6, Tnf) were downregulated and surprisingly Cyp2e1, presumably related to fatty liver phenotype, was upregulated. For the gene expression analysis between CAF vs SC (Figure 4B) and CAF vs CAF+BPF (Figure 4E) groups, a gene induction higher than 1.7 folds and a reduction of expression to at least -1.7 were defined as cut-off values. The majority of genes were regulated below the threshold and they were equally distributed between positive and negative fold regulation (Figure 4C, F). The data presented in the scatter plots (Figure 4A,D) were also normalized to the expression levels in SC group (Figure 6).
3.3. BPF Supplementation Has a Minor Effect on Liver Autophagy Gene Expression
Next, mRNA levels of autophagy-related genes were assessed in the same samples of cDNA, but on autophagy-specific arrays. A scatter plot of 84 autophagy-related genes, showed that the expression of 5 genes (Tnf, Ins2, Atg16I2, Map and Hsp90) were significantly modulated in CAF vs SC group (Figure 5A, B) while comparison between CAF+BPF and CAF group showed changes in only 3 genes (Ins2, Map1lc3a and Dapk1) (Figure 5C, D). The in CAF and CAF+BPF were also compared to the expression levels in SC group (Figure 6). Thus, compared to the genes responsible for the development of fatty liver, autophagy-related gene expression is much less affected by bergamot polyphenols.
Figure 5.
Regulation of autophagy-related transcripts in steatotic livers exposed to CAF diet for 14 weeks as compared to control SC diet (A, B) and the effects of chronic supplementation of bergamot polyphenols to CAF diet (C-D). (A) CAF vs SC and (C) CAF+BPF vs CAF scatter plots for differential analysis of 84 genes coding for proteins regulating autophagy. Below are the lists of genes modulated more than 1.7-fold when CAF are compared to SC livers (B) and CAF+BPF are compared to CAF livers (D). Bergamot polyphenols have a slight effect on the expression of autophagy-related genes in the liver in contrast to lipid and glucose metabolism genes that are strongly modulated.
Figure 5.
Regulation of autophagy-related transcripts in steatotic livers exposed to CAF diet for 14 weeks as compared to control SC diet (A, B) and the effects of chronic supplementation of bergamot polyphenols to CAF diet (C-D). (A) CAF vs SC and (C) CAF+BPF vs CAF scatter plots for differential analysis of 84 genes coding for proteins regulating autophagy. Below are the lists of genes modulated more than 1.7-fold when CAF are compared to SC livers (B) and CAF+BPF are compared to CAF livers (D). Bergamot polyphenols have a slight effect on the expression of autophagy-related genes in the liver in contrast to lipid and glucose metabolism genes that are strongly modulated.
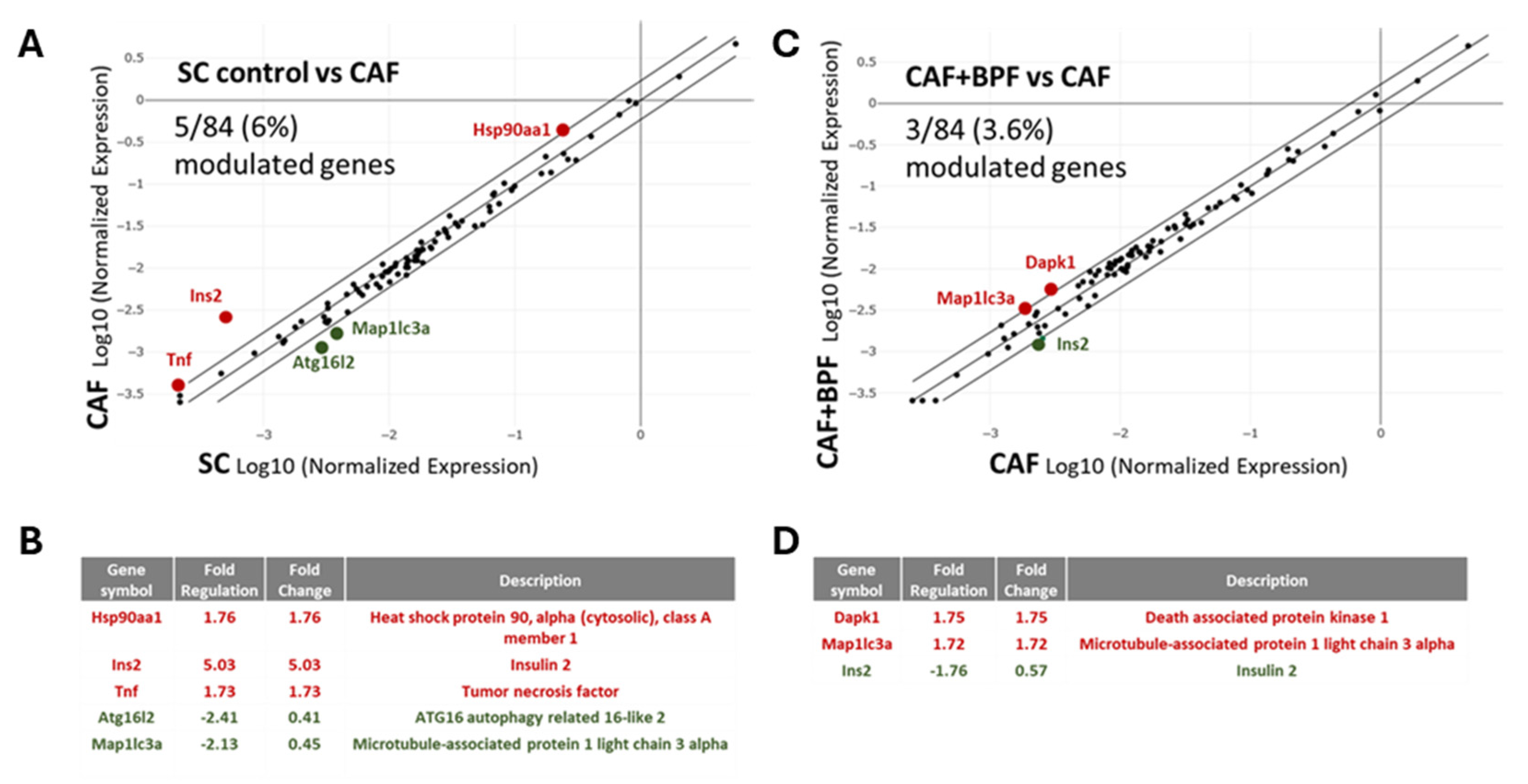
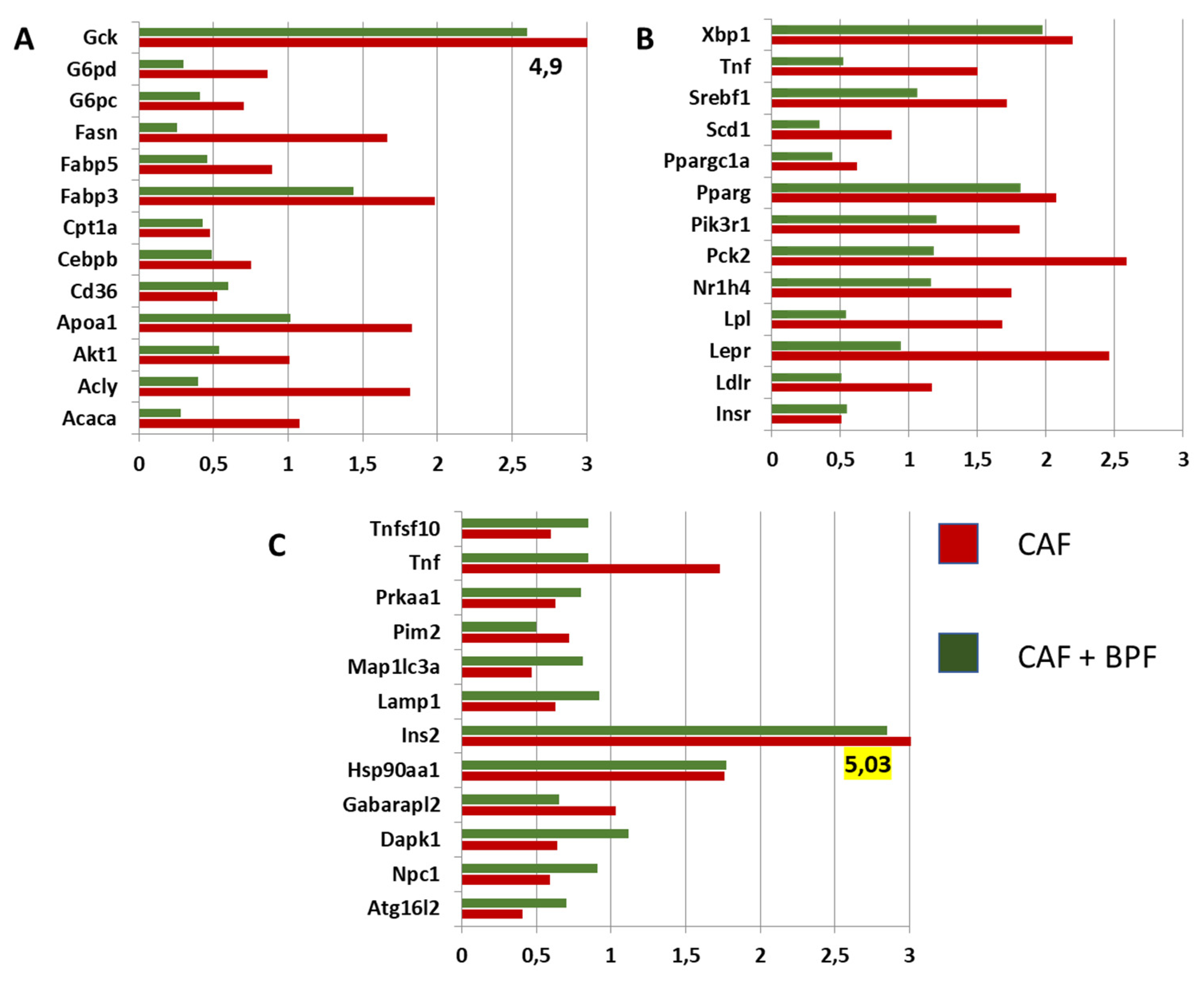
Figure 6.
Hepatic genes modulated in CAF and CAF+BPF groups when compared to control SC livers. (A-B). Fold change expression in metabolic genes associated with lipid- and glucose-metabolism significantly modulated by CAF and CAF+BPF treatments. Only genes with fold change > 1.7 or < 0.5 in at least one group are reported. (C) Gene expression changes as in A and B for autophagy-related transcripts. Only genes with fold change > 1.7 or < 0.65 in at least one group are reported. All the fold change values are relative to the expression levels of individual transcripts in SC livers normalized to 1.
Figure 6.
Hepatic genes modulated in CAF and CAF+BPF groups when compared to control SC livers. (A-B). Fold change expression in metabolic genes associated with lipid- and glucose-metabolism significantly modulated by CAF and CAF+BPF treatments. Only genes with fold change > 1.7 or < 0.5 in at least one group are reported. (C) Gene expression changes as in A and B for autophagy-related transcripts. Only genes with fold change > 1.7 or < 0.65 in at least one group are reported. All the fold change values are relative to the expression levels of individual transcripts in SC livers normalized to 1.
3.4. Standard qRT-PCR Confirms RT2-PCR Array Data on BPF-Induced Gene Modulation
Changes in the expression of a subset of the candidate regulated genes identified by the array approach were confirmed by individual quantitative real-time PCR (qRT-PCR) assays, later referred to as “single qRT-PCR”. The induction of lipogenesis genes, Fasn, Acly and Acaca, was not statistically significant when mRNA from 5 individual animals was separately analyzed (Figure 7). Our results, in line with the array data, showed a more marked downregulation produced by BPF treatment in some genes Acaca, Fasn, Pck2 and Ins, compared to both CAF and SC groups (Figure 7). Interestingly in the BPF group the following genes Fasn, Acaca, Acly, Pck2 and Ins were suppressed far below the mRNA levels of the control livers. In addition, CAF diet caused a dramatic upregulation in Gck, Pck2 and Ins genes compared to the SC group without affecting Srebp1 and Pparg genes, which showed a similar trend observed in array analysis (Figure 7).
3.5. Most BPF-Induced Effects on Gene Expression Can Be Replicated at the Protein Level
To verify the BPF-induced effects on gene expression, we performed Western blot analysis of liver lysates for the most representative gene products. The increased ACACA and PCK2 protein levels in rats fed with CAF diet were reversed by BPF supplementation and in the case of ACACA significantly reduced when compared with the SC group (Figure 8A,B). CAF diet was not able to modulate the protein levels of ACLY and GCK enzymes in comparison to the SC diet, but BPF treatment significantly decreased it (Figure 8A,B). This finding contrasts with the highly elevated Gck mRNA in CAF livers (Figure 4B,C, Figure 6A and Figure 7), suggesting that not all significant transcriptional effects are mirrored by protein levels. Concerning the expression level of autophagy-related proteins, both LC3I and LC3II forms of LC3 were significantly reduced under CAF treatment when compared to the SC group (Figure 8C,D) and only autophagosome marker LC3II was significantly upregulated in CAF+BPF group with respect to CAF group. However, in contrast to RNA expression data in Figure 5, ATG16L was found slightly upregulated in CAF group but not modulated in CAF+ BPF group (Figure 8C,D).
4. Discussion
Citrus Bergamia flavonoids efficiently prevent NAFLD, systemic redox imbalance and other features of metabolic syndrome in rats fed with CAF diet [10,14]. These original findings have been subsequently confirmed in other models and extended to NASH [15,54,55,56] and NAFLD patients with metabolic syndrome patients in subsequent studies [17,18,56]. Accordingly, the rats used in the present study responded to BPF supplementation with a strong reduction of lipid accumulation in CAF-fed livers characterized by a 60% and 80% decrease in fat content and numbers of big lipid droplets, respectively, confirmed by histopathological analysis of liver sections. BPF largely and significantly reduced diabetes and hypertriglyceridemia in CAF rats, while it had no significative impact on the body mass in this experiment; this confirms that BPF is hepatoprotective but has no or little effect on obesity [10,15]. In this article, we evaluated the impact of chronic supplementation of BPF on mRNA levels of a battery of 168 genes to address possible mechanisms behind the widely demonstrated efficacy of bergamot polyphenols against NAFLD.
The gene expression analysis of livers after long-term 14 weeks of CAF diet feeding revealed that CAF diet produced both expected and few unexpected effects on the transcriptional profiles of hepatic genes compared to other hypercaloric diets. There is a pattern of insulin and leptin resistance characterized by: upregulation of insulin mRNA 5 to 10 times, downregulation of Insr mRNA but upregulation of leptin receptor Lepr transcript 2 and 2.3-fold, respectively. The key transcription factors of lipid and cholesterol synthesis Srebf1, Xbp1 and Pparg, were also upregulated around 2-fold, which is common to many rodent models of NAFLD and found in patients with histologically diagnosed NAFLD
However, there was only a weak or not statistically significant increase in transcripts coding for lipogenesis enzymes (Acly, Acaca, Fasn, Scd-1) in CAF livers. This is in contrast to fructose-induced rat and murine models of NAFLD, where the main targets of SREBP-1c mediated transactivation Acaca and Fasn, were upregulated 6 to 16-fold compared to control animal [22,57]. Yet, such a strong difference between those models might be explained by overnight fasting, but continued fructose supply before sacrifice [58], while in our study all the animals were deprived for 5 h of all energy sources for blood analysis. Indeed, a study reported that in High-fat diet (HFD)-fed mice Acaca was more expressed in HFD than in the control group and decreased to baseline levels in fasted control animals. In the same experiment, Fasn upregulation by 3 to 4-fold in HFD mice did not seem to depend on fasting [23]. Accordingly, we found consistent, but modest upregulation of Fasn mRNA in the CAF group by around 1.6, but less consistent data for Acaca and Acly in both array pool (Figure 4) and individual assays (Figure 7), suggesting that lipogenesis enzymes expression is flexible and quickly responds to nutrient status. In contrast, the upstream transcription factors, such as Srebf1 and Pparg maintained the stable 2-fold upregulation, regardless of short-term fasting [23]. Interestingly, Acly was potently induced at the protein level.
According to the phenotypic changes in rat livers, BPF was able to suppress several NAFLD-related genes. Among them, mRNA levels of Acly, Acaca, Fasn, and Scd1 were strongly suppressed in BPF-treated livers far below the mRNA levels of CAF and control livers, as confirmed also by an alternative primer-pair RT2-PCR analysis. For some genes, we could also observe that their mRNA downregulation (Acaca and Acly) led to a marked protein decrease in BPF-treated livers. BPF also considerably downregulated liver expression of Apoa1 in comparison to CAF diet. APOA1 is the major protein component of HDL particles in plasma and it is involved in lipid metabolism and lipid transport from the liver [59]. It has been proposed that the upregulation of PPARs might promote the expression and molecular transportation of APOA1 to form a PPAR-APOA1 signaling pathway regulating the pathogenesis of NAFLD [60]. In our study liver expression of PPARgc1a was considerably reduced by supplementation with BPF suggesting that the pathogenic pathway related to APOA1 might be attenuated.
The efficacy of different polyphenols and plant extracts, including Citrus fruit extracts, have been tested in several NAFLD and NASH rodent models, but none of these studies reported such a dramatic transcriptional suppression of lipogenesis as bergamot polyphenols in CAF-induced NAFLD/NASH in rats. This is true for both purified compounds as well as for complex plant extracts [51,58,61,62,63,64,65]. In another study, the same array of 84 mouse genes related to NAFLD was used to characterize the antisteatotic effect of a complex mixture of natural extracts containing silymarin, curcumin and chlorogenic acid. Yet, even though this extract almost fully reverted NAFLD induced by HFD in mice, only Scd-1 and Fabp5 were found downregulated after 16 weeks of treatment [62].
Another important finding in this work is no regulation of fatty acid oxidation genes, such as Acadl, Acox1, Cpt1a, Cpt2, Fabp1, Irs1, mTOR, and Ppara by long-term BPF supplementation. The only effect was lower Carnitine palmitoyltransferase 1a (Cpt1a) expression in CAF and CAF+BPF livers with respect to SC group (Figure 7A). CPT1A is the key enzyme in the carnitine-dependent transport of fatty acids across the external mitochondrial membrane. Its lower expression should limit fatty acids uptake by mitochondria and thus reduce b-oxidation, as an adaptative process to mitochondria-derived oxidative stress, found in fructose- [26,27], but not in high-fat diet-induced NAFLD models [27,66]. In fact, both mitochondrial as well as peroxisomal fatty acid oxidation are ROS-generating processes and their excessive stimulation is deleterious for liver tissue [31]. Lack of positive stimulation of fatty-acid oxidation gene expression in BPF-treated livers is in line with a clinical study in which a multifactorial diet (rich in polyphenols and polyunsaturated fatty acids) downregulated lipogenesis, but did not regulate plasma levels of b-hydroxybutyrate and thus b-oxidation [67]. Remarkably, the stimulation of hepatic b-oxidation is a typical response to different phytochemicals in vitro in cellular models of NAFLD [68,69,70,71], but it is only occasionally reported in rodents treated with purified phenolic compounds [72,73]. In conclusion, our data demonstrate that transcriptional suppression of lipogenesis, and not stimulation of b-oxidation, is the main mechanism of BPF counteracting fat accumulation in NAFLD and NASH in vivo as depicted in Figure 9. This finding is consistent with an antioxidant function of flavonoids, mediated by the regulation of expression of genes contributing to healthy redox balance rather than through direct ROS-scavenging.
An unexpected effect of BPF was the induction of Cytochrome 2e1 (CYP2E1) by BPF, a phase I drug metabolism enzyme, but not by CAF diet feeding without BPF. At first glance, it seems surprising since CYP2E1 activity and expression were found elevated in human steatohepatitis and rodent alcohol and methionine-induced liver steatosis [74]. However, In line with our findings, CYP2E1 expression has not been found increased in most rodent NAFLD models [71], while it might play a role in polyphenol metabolism and its induction is a hepatic response to flavonoids and related metabolites. In fact, mice treated with flavone-8-acetic acid showed a substantial induction of hepatic CYP2E1 [75].
Modulation of autophagy to degrade LDs and relieve hepatic inflammation is a potential therapeutic target for NAFLD. Since common transcription factors are believed to regulate lipid metabolism and autophagy, we expected huge transcriptional changes in hepatic autophagy genes, induced by both CAF diet and bergamot polyphenols. Surprisingly, we found that both CAF diet and BPF had limited effects on autophagy-related gene expression. Indeed, it slightly upregulated Hsp90aa1 and downregulated Atg16l2, and Maplc3a, while BPF upregulated only Maplc3a and Dapk1. The other two genes Ins2 and Tnf, upregulated by CAF diet and downregulated by BPF (Tnf not significantly) are not typical autophagy-related genes and they play more important roles in diabetes and inflammation. At the protein level, we were able to confirm the downregulation of LC3I and LC3II levels in CAF livers and its upregulation by BPF diet, but not in the case of ATG16L, suggesting that there is a partial correlation between mRNA levels and protein expression. This indicates that polyphenols regulate autophagy mainly by post-translational mechanisms. In fact, Beclin-1 and p62/SQSTM1 were previously found to be regulated by chronic supplementation of BPF in rats [10], but they are not regulated at mRNA levels in this study. To our knowledge, the data set presented here is the first characterization of autophagy-related gene expression in response to the CAF diet and polyphenols.
5. Conclusions
In conclusion, among many pleiotropic effects of polyphenols, the main mechanisms underlying the anti-steatotic effect of BPF supplementation appear to be transcriptional and include a potent suppression of lipogenesis and an effective reduction of gluconeogenesis. Interestingly, b-oxidation is not induced transcriptionally, while autophagy-related genes are only marginally modulated by chronic treatment with BPF, indicating that autophagy is regulated mainly by posttranslational mechanisms.
Considering that oxidation of fatty acids is the main source of oxidative stress in NAFLD, no induction of b-oxidation with an extremely potent suppression of lipogenesis might be an efficient antioxidant mechanism of bergamot flavonoids in vivo. This mechanism provides a convincing explanation of the high efficacy of BPF against NAFLD far beyond its anti-inflammatory and proautophagic effects.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Original Western blots Janda 2024.
Author Contributions
Conceptualization, E.J.; methodology, M.P., D.L., B.F., E.J.; validation, A.L., F.C., and C.R.; formal analysis, E.J., M.P., F.C. and C.R.; investigation, D.L., M.P., A.L and E.J.; resources, E.J.; data curation, E.J., M.P., F.C.; writing—original draft preparation, E.J.; writing—review and editing, E.J. and B.F.; visualization, F.C., C.R and E.J.; supervision, E.J.; funding acquisition, E.J. and V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NUTRAMED Consortium (PON03PE 00078) and received a financial support from Herbal and Antioxidant Derivatives (H&AD) S.r.l., Bianco (RC), Italy, awarded to V.M. and E.J., respectively.
Institutional Review Board Statement
This animal study was approved by a local animal welfare committee and by the Italian Ministry of Health (project code: 0124/09/2013), according to Legislative Decree 116/1992, which was in force when the study was proposed. All surgery was performed under anesthesia and all efforts were made to minimize animal suffering.
Data Availability Statement
The data is contained within the article and Supplementary material. Other original data supporting reported results are available upon request.
Acknowledgments
The authors wish to thank H&AD srl for the kind gift of BPF and Daniele Vumbaca from Department of Health Sciences, UMG, Catanzaro, Italy for excellent technical assistance. We also thank Dr. Antonio Pisano from Experimental Medicine Department, UMG, Catanzaro, Italy, for critical reading and useful suggestions to improve the manuscript.
Conflicts of Interest
E.J. received a small financial support from Herbal and Antioxidant Derivatives (H&AD) S.r.l., Bianco (RC), Italy for research on BPF™. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Sookoian, S., et al., Genetic Pathways in Nonalcoholic Fatty Liver Disease: Insights From Systems Biology. Hepatology, 2020. 72(1): p. 330-346. [CrossRef]
- Bence, K.K. and M.J. Birnbaum, Metabolic drivers of non-alcoholic fatty liver disease. Mol Metab, 2021. 50: p. 101143. [CrossRef]
- Yaqub, S., et al., Decoding the Pathophysiology of Non-alcoholic Fatty Liver Disease Progressing to Non-alcoholic Steatohepatitis: A Systematic Review. Cureus, 2021. 13(9): p. e18201. [CrossRef]
- Majumdar, A., J. Verbeek, and E.A. Tsochatzis, Non-alcoholic fatty liver disease: Current therapeutic options. Curr Opin Pharmacol, 2021. 61: p. 98-105. [CrossRef]
- Tokushige, K., et al., Evidence-based clinical practice guidelines for nonalcoholic fatty liver disease/nonalcoholic steatohepatitis 2020. J Gastroenterol, 2021. 56(11): p. 951-963. [CrossRef]
- Son, C.G., et al., Medicinal Herbs and Their Active Compounds for Fatty Liver Diseases. Evid Based Complement Alternat Med, 2017. 2017: p. 3612478. [CrossRef]
- Bagherniya, M., et al., Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol Res, 2018. 130: p. 213-240. [CrossRef]
- Baron, G., et al., Analytical Profile and Antioxidant and Anti-Inflammatory Activities of the Enriched Polyphenol Fractions Isolated from Bergamot Fruit and Leave. Antioxidants (Basel), 2021. 10(2). [CrossRef]
- Janda, E., et al., Qualitative and quantitative analysis of the proautophagic activity of Citrus flavonoids from Bergamot Polyphenol Fraction. Data Brief, 2018. 19: p. 1327-1334. [CrossRef]
- Parafati, M., et al., Bergamot polyphenol fraction prevents nonalcoholic fatty liver disease via stimulation of lipophagy in cafeteria diet-induced rat model of metabolic syndrome. J Nutr Biochem, 2015. 26(9): p. 938-48. [CrossRef]
- Walker, R., E. Janda, and V. Mollace, Chapter 84 - The Use of Bergamot-Derived Polyphenol Fraction in Cardiometabolic Risk Prevention and its Possible Mechanisms of Action, in Polyphenols in Human Health and Disease, R.R. Watson, V.R. Preedy, and S. Zibadi, Editors. 2014, Academic Press: San Diego. p. 1087-1105.
- Formisano, C., et al., Detailed Phytochemical Characterization of Bergamot Polyphenolic Fraction (BPF) by UPLC-DAD-MS and LC-NMR. J Agric Food Chem, 2019. 67(11): p. 3159-3167. [CrossRef]
- Mollace, R., et al., Comparative Effect of Bergamot Polyphenolic Fraction and Red Yeast Rice Extract in Rats Fed a Hyperlipidemic Diet: Role of Antioxidant Properties and PCSK9 Expression. Nutrients, 2022. 14(3). [CrossRef]
- La Russa, D., et al., Oxidative Imbalance and Kidney Damage in Cafeteria Diet-Induced Rat Model of Metabolic Syndrome: Effect of Bergamot Polyphenolic Fraction. Antioxidants (Basel), 2019. 8(3). [CrossRef]
- Parafati, M., et al., Bergamot Polyphenols Boost Therapeutic Effects of the Diet on Non-Alcoholic Steatohepatitis (NASH) Induced by "Junk Food": Evidence for Anti-Inflammatory Activity. Nutrients, 2018. 10(11). [CrossRef]
- Mare, R., et al., A new breakfast brioche containing bergamot fiber prevents insulin and glucose increase in healthy volunteers: a pilot study. Minerva Endocrinol (Torino), 2021. 46(2): p. 214-225. [CrossRef]
- Ferro, Y., et al., Randomized Clinical Trial: Bergamot Citrus and Wild Cardoon Reduce Liver Steatosis and Body Weight in Non-diabetic Individuals Aged Over 50 Years. Front Endocrinol (Lausanne), 2020. 11: p. 494. [CrossRef]
- Capomolla, A.S., et al., Atherogenic Index Reduction and Weight Loss in Metabolic Syndrome Patients Treated with A Novel Pectin-Enriched Formulation of Bergamot Polyphenols. Nutrients, 2019. 11(6). [CrossRef]
- Gliozzi, M., et al., Bergamot polyphenolic fraction enhances rosuvastatin-induced effect on LDL-cholesterol, LOX-1 expression and protein kinase B phosphorylation in patients with hyperlipidemia. Int J Cardiol, 2013. 170(2): p. 140-5. [CrossRef]
- Musolino, V., et al., The synergistic effect of Citrus bergamia and Cynara cardunculus extracts on vascular inflammation and oxidative stress in non-alcoholic fatty liver disease. J Tradit Complement Med, 2020. 10(3): p. 268-274. [CrossRef]
- Softic, S., D.E. Cohen, and C.R. Kahn, Role of Dietary Fructose and Hepatic De Novo Lipogenesis in Fatty Liver Disease. Dig Dis Sci, 2016. 61(5): p. 1282-93. [CrossRef]
- Zhao, S., et al., Dietary fructose feeds hepatic lipogenesis via microbiota-derived acetate. Nature, 2020. 579(7800): p. 586-591. [CrossRef]
- Chattopadhyay, M., et al., Ablation of PI3K p110-alpha prevents high-fat diet-induced liver steatosis. Diabetes, 2011. 60(5): p. 1483-92. [CrossRef]
- Porras, D., et al., Protective effect of quercetin on high-fat diet-induced non-alcoholic fatty liver disease in mice is mediated by modulating intestinal microbiota imbalance and related gut-liver axis activation. Free Radic Biol Med, 2017. 102: p. 188-202. [CrossRef]
- Luna-Vital, D., et al., Maize extract rich in ferulic acid and anthocyanins prevents high-fat-induced obesity in mice by modulating SIRT1, AMPK and IL-6 associated metabolic and inflammatory pathways. J Nutr Biochem, 2020. 79: p. 108343. [CrossRef]
- Mizuno, G., et al., High-fructose corn syrup intake has stronger effects on the transcription level of hepatic lipid metabolism-related genes, via DNA methylation modification, in childhood and adolescence than in other generations. Life Sci, 2022. 301: p. 120638. [CrossRef]
- Softic, S., et al., Dietary Sugars Alter Hepatic Fatty Acid Oxidation via Transcriptional and Post-translational Modifications of Mitochondrial Proteins. Cell Metab, 2019. 30(4): p. 735-753 e4. [CrossRef]
- Wisniewska, A., et al., Inhibition of Atherosclerosis and Liver Steatosis by Agmatine in Western Diet-Fed apoE-Knockout Mice Is Associated with Decrease in Hepatic De Novo Lipogenesis and Reduction in Plasma Triglyceride/High-Density Lipoprotein Cholesterol Ratio. Int J Mol Sci, 2021. 22(19). [CrossRef]
- Steensels, S., J. Qiao, and B.A. Ersoy, Transcriptional Regulation in Non-Alcoholic Fatty Liver Disease. Metabolites, 2020. 10(7). [CrossRef]
- Shi, Q., et al., Intracellular Cholesterol Synthesis and Transport. Front Cell Dev Biol, 2022. 10: p. 819281. [CrossRef]
- Chen, Z., et al., Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic Biol Med, 2020. 152: p. 116-141. [CrossRef]
- Janda, E., et al., Apigenin and Luteolin Regulate Autophagy by Targeting NRH-Quinone Oxidoreductase 2 in Liver Cells. Antioxidants (Basel), 2021. 10(5). [CrossRef]
- Hu, M., et al., Salidroside Activates the AMP-Activated Protein Kinase Pathway to Suppress Nonalcoholic Steatohepatitis in Mice. Hepatology, 2021. 74(6): p. 3056-3073. [CrossRef]
- Garcia-Aguilar, A., et al., Dietary Polyphenols in Metabolic and Neurodegenerative Diseases: Molecular Targets in Autophagy and Biological Effects. Antioxidants (Basel), 2021. 10(2). [CrossRef]
- Janda, E., et al., Molecular mechanisms of lipid- and glucose-lowering activities of bergamot flavonoids. PharmaNutrition, 2016. 4: p. S8-S18. [CrossRef]
- Park, S.J., et al., Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell, 2012. 148(3): p. 421-33. [CrossRef]
- Lacroix, S., et al., A computationally driven analysis of the polyphenol-protein interactome. Sci Rep, 2018. 8(1): p. 2232. [CrossRef]
- Yessenkyzy, A., et al., Polyphenols as Caloric-Restriction Mimetics and Autophagy Inducers in Aging Research. Nutrients, 2020. 12(5). [CrossRef]
- Dincer, Y. and S. Yuksel, Antiobesity effects of phytochemicals from an epigenetic perspective. Nutrition, 2021. 84: p. 111119. [CrossRef]
- Jiang, Y., et al., Epigenetic mechanisms underlying the benefits of flavonoids in cardiovascular health and diseases: are long non-coding RNAs rising stars? Crit Rev Food Sci Nutr, 2022. 62(14): p. 3855-3872. [CrossRef]
- Russo, G.L., et al., Dietary polyphenols and chromatin remodeling. Critical Reviews in Food Science and Nutrition, 2017. 57(12): p. 2589-2599. [CrossRef]
- Pirola, C.J. and S. Sookoian, Metabolic dysfunction-associated fatty liver disease: advances in genetic and epigenetic implications. Curr Opin Lipidol, 2022. 33(2): p. 95-102. [CrossRef]
- Theys, C., et al., PPARalpha in the Epigenetic Driver Seat of NAFLD: New Therapeutic Opportunities for Epigenetic Drugs? Biomedicines, 2022. 10(12). [CrossRef]
- Allaire, M., et al., Autophagy in liver diseases: Time for translation? J Hepatol, 2019. 70(5): p. 985-998. [CrossRef]
- Feng, Y., et al., Interplay of energy metabolism and autophagy. Autophagy, 2023: p. 1-11. [CrossRef]
- Xie, Y., et al., Interplay Between Lipid Metabolism and Autophagy. Front Cell Dev Biol, 2020. 8: p. 431. [CrossRef]
- Settembre, C., et al., TFEB controls cellular lipid metabolism through a starvation-induced autoregulatory loop. Nat Cell Biol, 2013. 15(6): p. 647-58. [CrossRef]
- Zhang, H., et al., Dynamic MTORC1-TFEB feedback signaling regulates hepatic autophagy, steatosis and liver injury in long-term nutrient oversupply. Autophagy, 2018. 14(10): p. 1779-1795. [CrossRef]
- Filali-Mouncef, Y., et al., The menage a trois of autophagy, lipid droplets and liver disease. Autophagy, 2021: p. 1-24. [CrossRef]
- Scafuri, B., et al., Molecular Docking Simulations on Histone Deacetylases (HDAC)-1 and -2 to Investigate the Flavone Binding. Biomedicines, 2020. 8(12). [CrossRef]
- Terzo, S., et al., Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice. Nutrients, 2018. 10(12). [CrossRef]
- Wu, X. and L.E. Nagy, MLKL contributes to Western diet-induced liver injury through inhibiting autophagy. Autophagy, 2020. 16(7): p. 1351-1352. [CrossRef]
- Janda, E., et al., Autophagy and neuroprotection in astrocytes exposed to 6-hydroxydopamine is negatively regulated by NQO2: a potential novel target in Parkinson’s disease. Scientific Reports, 2023. Preprint(In revision). [CrossRef]
- Nucera, S., et al., MAFLD progression contributes to altered thalamus metabolism and brain structure. Sci Rep, 2022. 12(1): p. 1207. [CrossRef]
- Siqueira, J.S., et al., Bergamot leaf extract treats cardiorenal metabolic syndrome and associated pathophysiological factors in rats fed with a high sugar fat diet. Mol Cell Endocrinol, 2022. 556: p. 111721. [CrossRef]
- Musolino, V., et al., The effect of bergamot polyphenolic fraction on lipid transfer protein system and vascular oxidative stress in a rat model of hyperlipemia. Lipids Health Dis, 2019. 18(1): p. 115. [CrossRef]
- Liao, C.C., et al., Prevention of diet-induced hyperlipidemia and obesity by caffeic acid in C57BL/6 mice through regulation of hepatic lipogenesis gene expression. J Agric Food Chem, 2013. 61(46): p. 11082-8. [CrossRef]
- Liu, L., et al., Modulation of hepatic sterol regulatory element-binding protein-1c-mediated gene expression contributes to Salacia oblonga root-elicited improvement of fructose-induced fatty liver in rats. J Ethnopharmacol, 2013. 150(3): p. 1045-52. [CrossRef]
- van der Vorst, E.P.C., High-Density Lipoproteins and Apolipoprotein A1. In: Hoeger, U., Harris, J. (eds) Vertebrate and Invertebrate Respiratory Proteins, Lipoproteins and other Body Fluid Proteins. Subcellular Biochemistry 2020. 94. [CrossRef]
- Chen, C., Li, H., Song, J., Zhang, C., Li, M., Mao, Y., Liu, A., & Du, J. (2022). Role of Apolipoprotein A1 in PPAR Signaling Pathway for Nonalcoholic Fatty Liver Disease. PPAR research, 2022, 4709300., Role of Apolipoprotein A1 in PPAR Signaling Pathway for Nonalcoholic Fatty Liver Disease. PPAR research 2022. [CrossRef]
- Wang, Y., et al., Prevention and treatment effect of total flavonoids in Stellera chamaejasme L. on nonalcoholic fatty liver in rats. Lipids Health Dis, 2015. 14: p. 85. [CrossRef]
- Amato, A., et al., NAFLD and Atherosclerosis Are Prevented by a Natural Dietary Supplement Containing Curcumin, Silymarin, Guggul, Chlorogenic Acid and Inulin in Mice Fed a High-Fat Diet. Nutrients, 2017. 9(5). [CrossRef]
- Chen, Q., et al., Effects of Natural Products on Fructose-Induced Nonalcoholic Fatty Liver Disease (NAFLD). Nutrients, 2017. 9(2). [CrossRef]
- Marine-Casado, R., et al., Cherry consumption out of season alters lipid and glucose homeostasis in normoweight and cafeteria-fed obese Fischer 344 rats. J Nutr Biochem, 2019. 63: p. 72-86. [CrossRef]
- Lee, G.H., et al., Citrus Peel Extract Ameliorates High-Fat Diet-Induced NAFLD via Activation of AMPK Signaling. Nutrients, 2020. 12(3). [CrossRef]
- Moody, L., et al., Epigenetic regulation of carnitine palmitoyltransferase 1 (Cpt1a) by high fat diet. Biochim Biophys Acta Gene Regul Mech, 2019. 1862(2): p. 141-152. [CrossRef]
- Costabile, G., et al., Reduction of De Novo Lipogenesis Mediates Beneficial Effects of Isoenergetic Diets on Fatty Liver: Mechanistic Insights from the MEDEA Randomized Clinical Trial. Nutrients, 2022. 14(10). [CrossRef]
- Badolati, N., et al., The Hepatoprotective Effect of Taurisolo, a Nutraceutical Enriched in Resveratrol and Polyphenols, Involves Activation of Mitochondrial Metabolism in Mice Liver. Antioxidants (Basel), 2020. 9(5). [CrossRef]
- Mirarchi, A., et al., Bergamot Polyphenol Extract Reduces Hepatocyte Neutral Fat by Increasing Beta-Oxidation. Nutrients, 2022. 14(16). [CrossRef]
- Rafiei, H., K. Omidian, and B. Bandy, Dietary Polyphenols Protect Against Oleic Acid-Induced Steatosis in an in Vitro Model of NAFLD by Modulating Lipid Metabolism and Improving Mitochondrial Function. Nutrients, 2019. 11(3). [CrossRef]
- Rebollo-Hernanz, M., et al., Phytochemicals from the Cocoa Shell Modulate Mitochondrial Function, Lipid and Glucose Metabolism in Hepatocytes via Activation of FGF21/ERK, AKT, and mTOR Pathways. Antioxidants (Basel), 2022. 11(1). [CrossRef]
- Alberdi, G., et al., Hepatic lipid metabolic pathways modified by resveratrol in rats fed an obesogenic diet. Nutrition, 2013. 29(3): p. 562-7. [CrossRef]
- Chao, J., et al., Gallic Acid Ameliorated Impaired Lipid Homeostasis in a Mouse Model of High-Fat Diet-and Streptozotocin-Induced NAFLD and Diabetes through Improvement of beta-oxidation and Ketogenesis. Front Pharmacol, 2020. 11: p. 606759. [CrossRef]
- Harjumaki, R., C.S. Pridgeon, and M. Ingelman-Sundberg, CYP2E1 in Alcoholic and Non-Alcoholic Liver Injury. Roles of ROS, Reactive Intermediates and Lipid Overload. Int J Mol Sci, 2021. 22(15). [CrossRef]
- Pham, M.H., et al., Identification and induction of cytochrome P450s involved in the metabolism of flavone-8-acetic acid in mice. Drug Metab Lett, 2011. 5(2): p. 73-84. [CrossRef]
Figure 1.
The Experimental Design and screening for genes regulated by BPF in NAFLD rat model in this work: Rat division for dietetic treatment and BPF administration for 15 weeks. On the right side, flow description from the liver sampling to RT2 profiling.
Figure 1.
The Experimental Design and screening for genes regulated by BPF in NAFLD rat model in this work: Rat division for dietetic treatment and BPF administration for 15 weeks. On the right side, flow description from the liver sampling to RT2 profiling.
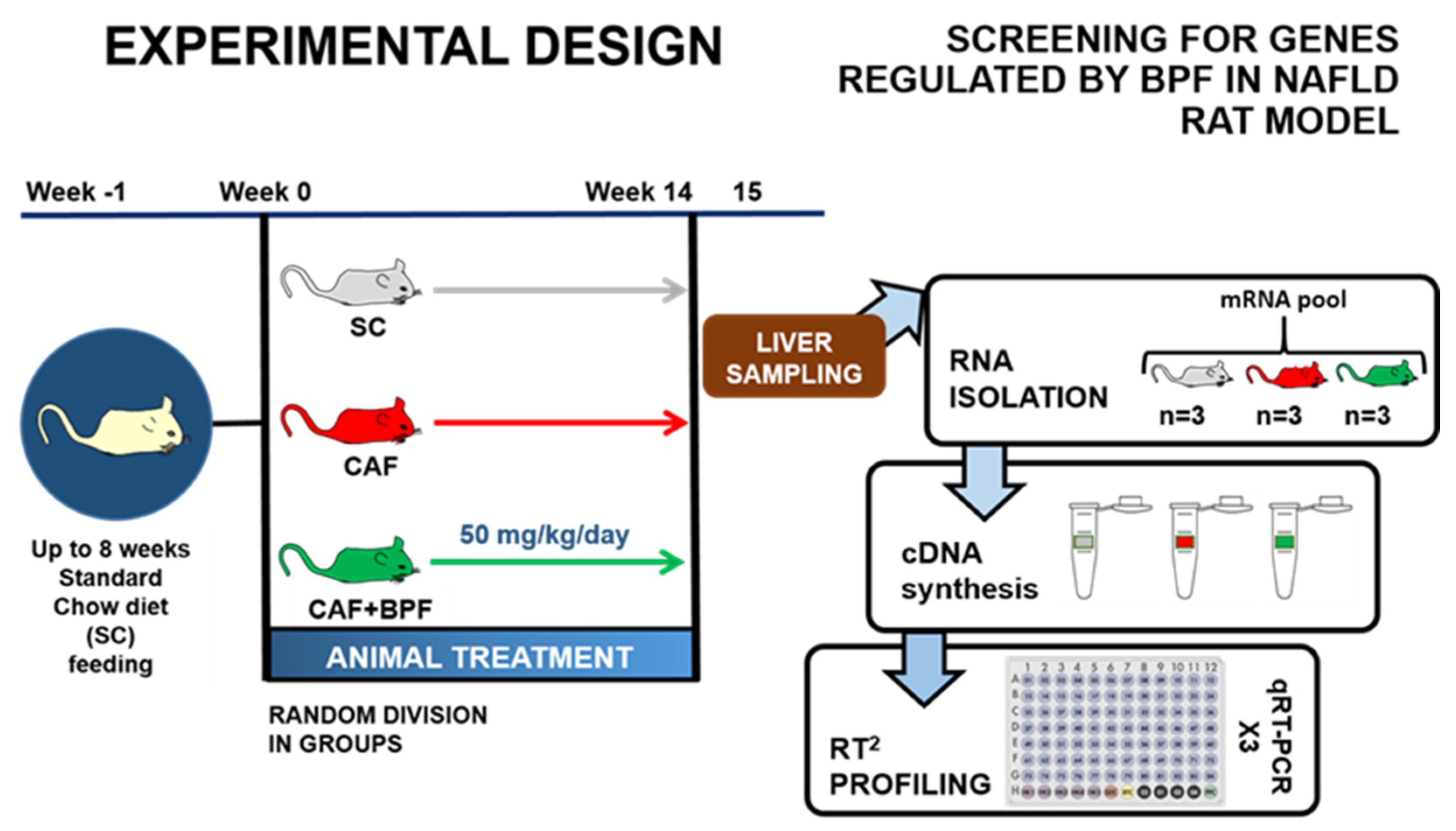
Figure 2.
Bergamot polyphenols prevent CAF diet-induced obesity, hypertriglyceridemia and intracellular fat accumulation in Wistar rats. (A) Final body weight, (B) Blood triglycerides, (TGL) (C) Blood total cholesterol. Data are presented as the mean ± SD of n = 5 rats. Each dot represents a rat. (D) The total lipid content in 400 mg of liver tissue was determined by Folch’s method. Data are presented as the mean ± SD of n = 4 livers for each group. (E) Representative blots for ADRP/Perilipin 2 and GAPDH as a loading control, showing liver lysates from 3 different rats for each group. (F) OD ratio of ADRP to GAPDH expression levels. Data are expressed as the mean ± SEM of n = 6 rat liver lysates for each group. Statistical analysis in A-D and F: One-way ANOVA with Tukey’s post-test. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, ns – not significant change.
Figure 2.
Bergamot polyphenols prevent CAF diet-induced obesity, hypertriglyceridemia and intracellular fat accumulation in Wistar rats. (A) Final body weight, (B) Blood triglycerides, (TGL) (C) Blood total cholesterol. Data are presented as the mean ± SD of n = 5 rats. Each dot represents a rat. (D) The total lipid content in 400 mg of liver tissue was determined by Folch’s method. Data are presented as the mean ± SD of n = 4 livers for each group. (E) Representative blots for ADRP/Perilipin 2 and GAPDH as a loading control, showing liver lysates from 3 different rats for each group. (F) OD ratio of ADRP to GAPDH expression levels. Data are expressed as the mean ± SEM of n = 6 rat liver lysates for each group. Statistical analysis in A-D and F: One-way ANOVA with Tukey’s post-test. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, ns – not significant change.
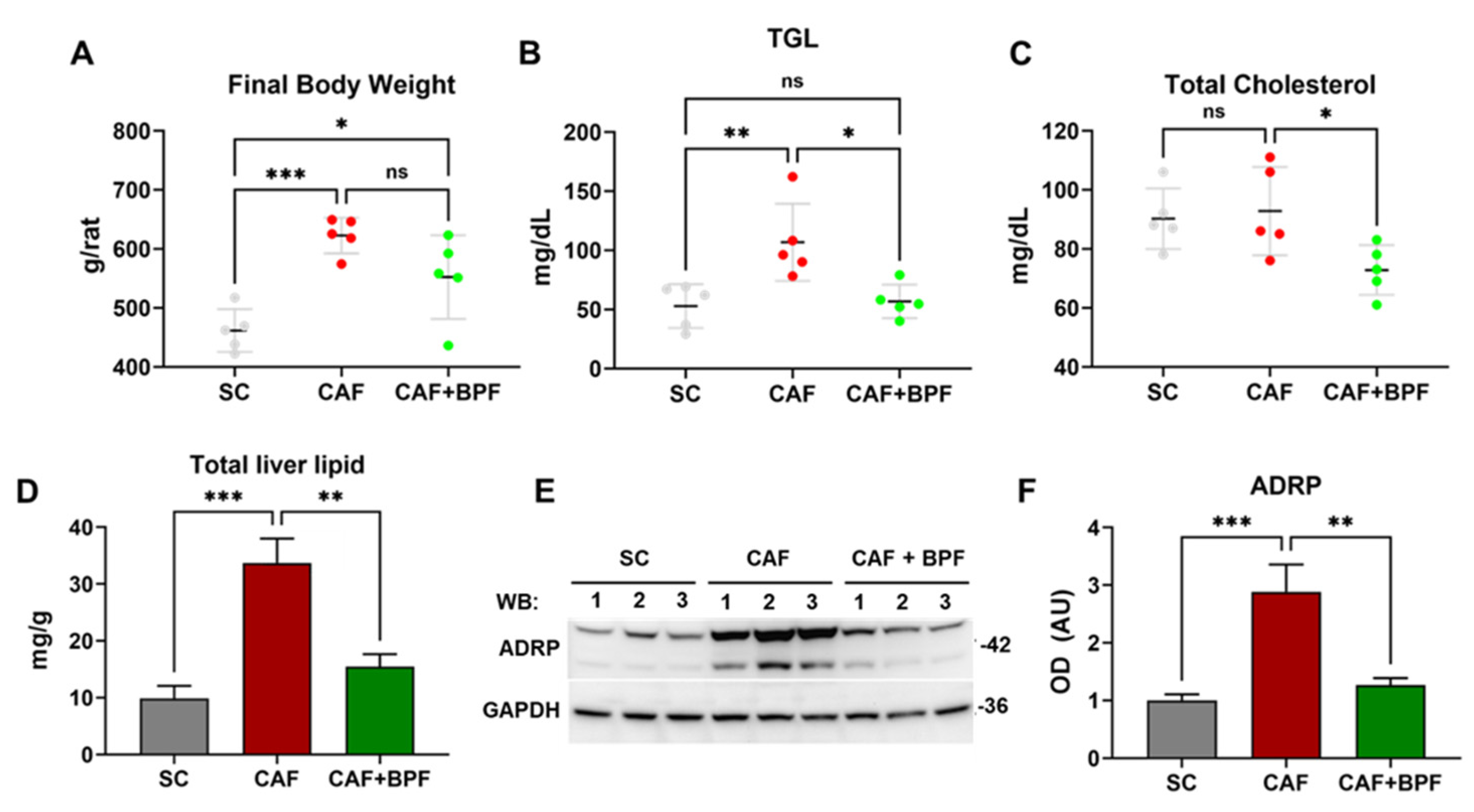
Figure 3.
BPF prevents CAF diet-induced hepatic steatosis. Histopathological changes of rat liver tissues between different dietary groups. (A-C) Representative HE and oil-red (ORO) stained liver sections were visualized at high magnification x40, by confocal microscopy bar = 25 μm; (D-F) and by bright-field ×40, bar = 40 μm; Images in the third row show magnified regions of D to F images in indicated by blue boxes, bar = 20 μm to visualize micro- and macro-vesicular steatosis. (G) LDs size and number quantification on confocal sections (as in A-C). LDs sized between 1-3 µm indicate microsteatosis and 4-7 µm macrosteatosis, respectively. Data are presented as the mean ± SEM (n = 3 livers and 9 images for each group). for LDs sized between 1-3 µm and 4-7 µm, respectively. Statistical analysis: Mann-Whitney test. Significant difference in CAF vs SC livers: * p ≤ 0.05, *** p ≤ 0.001. Significant change in CAF+BPF vs CAF group: # p ≤ 0.05.
Figure 3.
BPF prevents CAF diet-induced hepatic steatosis. Histopathological changes of rat liver tissues between different dietary groups. (A-C) Representative HE and oil-red (ORO) stained liver sections were visualized at high magnification x40, by confocal microscopy bar = 25 μm; (D-F) and by bright-field ×40, bar = 40 μm; Images in the third row show magnified regions of D to F images in indicated by blue boxes, bar = 20 μm to visualize micro- and macro-vesicular steatosis. (G) LDs size and number quantification on confocal sections (as in A-C). LDs sized between 1-3 µm indicate microsteatosis and 4-7 µm macrosteatosis, respectively. Data are presented as the mean ± SEM (n = 3 livers and 9 images for each group). for LDs sized between 1-3 µm and 4-7 µm, respectively. Statistical analysis: Mann-Whitney test. Significant difference in CAF vs SC livers: * p ≤ 0.05, *** p ≤ 0.001. Significant change in CAF+BPF vs CAF group: # p ≤ 0.05.
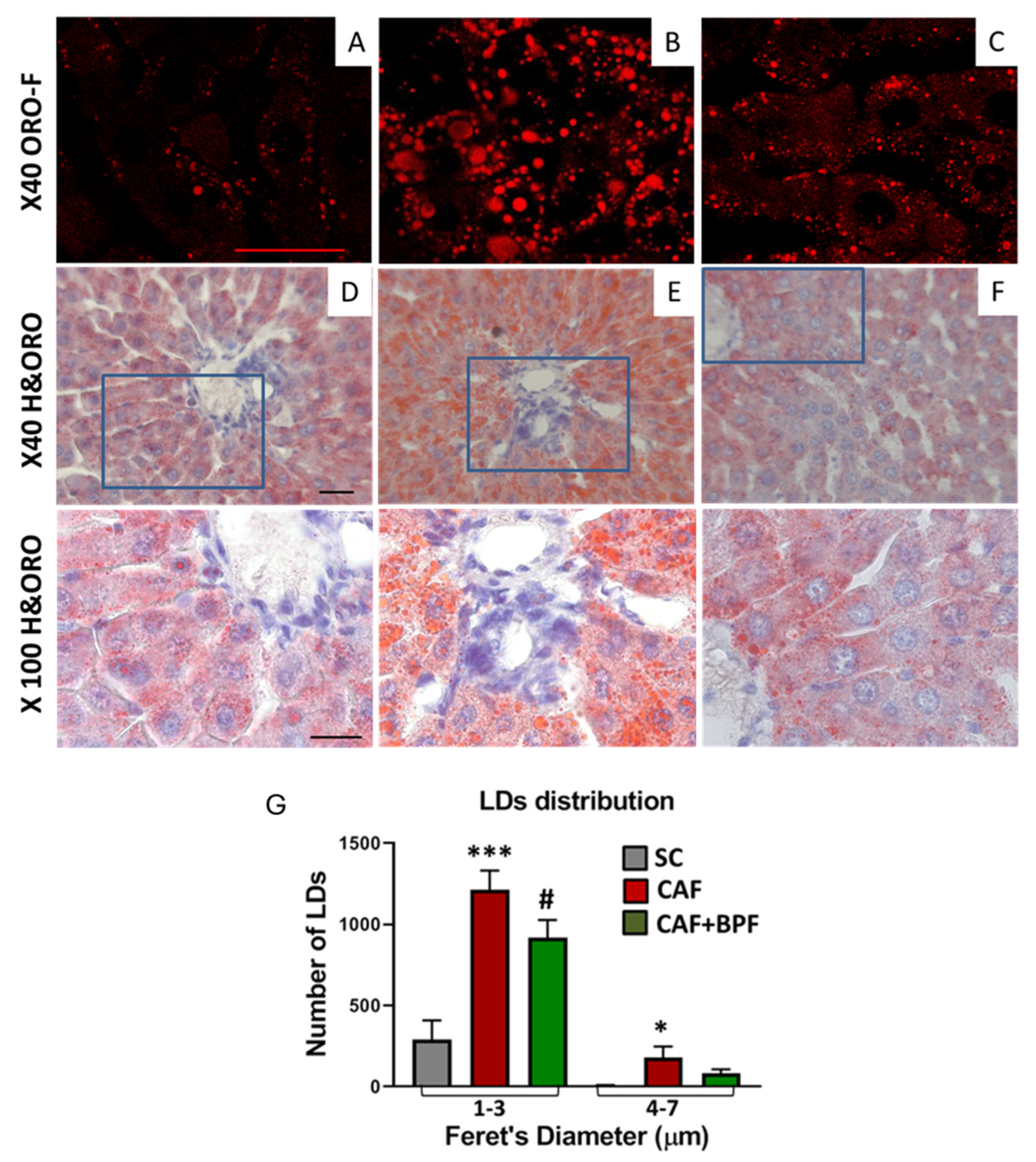
Figure 4.
Gene expression changes in steatotic livers exposed to CAF diet for 14 weeks as compared to control SC diet (A, B, C) and the effects of a chronic supplementation of bergamot polyphenols to CAF diet (D, E, F). (A) CAF vs SC and (D) CAF+BPF vs CAF scatter plots for differential RT2 analysis of 84 genes typically modulated in liver steatosis. Below, the lists of genes modulated more than 1.7 fold when CAF are compared to SC livers (B) and CAF+BPF (E) are compared to CAF livers. (B,D) The arrays of all fatty liver-associated genes with respective fold regulation values when (C) CAF vs SC and (F) CAF+BPF vs CAF livers are compared. Note, that bergamot polyphenols strongly suppress lipogenesis-related genes in the liver.
Figure 4.
Gene expression changes in steatotic livers exposed to CAF diet for 14 weeks as compared to control SC diet (A, B, C) and the effects of a chronic supplementation of bergamot polyphenols to CAF diet (D, E, F). (A) CAF vs SC and (D) CAF+BPF vs CAF scatter plots for differential RT2 analysis of 84 genes typically modulated in liver steatosis. Below, the lists of genes modulated more than 1.7 fold when CAF are compared to SC livers (B) and CAF+BPF (E) are compared to CAF livers. (B,D) The arrays of all fatty liver-associated genes with respective fold regulation values when (C) CAF vs SC and (F) CAF+BPF vs CAF livers are compared. Note, that bergamot polyphenols strongly suppress lipogenesis-related genes in the liver.

Figure 7.
The expression level of selected genes was analyzed by qRT-PCR with an independent set of primers in each animal separately. The bar represents the mean ± SD of expression data from n = 4 to 5 rat livers relative to the house-keeping control (Hrpt1). *, ** Statistically significant difference compared to the control SC group at p < 0.5 or p < 0.01 respectively. #, ##, ### - Statistically significant difference compared to the control CAF group at p < 0.5, < 0.01 or < 0.001, respectively. Red numbers indicate fold change in CAF compared to SC group. Green and black numbers indicate the fold change in CAF+BPF group compared to CAF and SC groups, respectively.
Figure 7.
The expression level of selected genes was analyzed by qRT-PCR with an independent set of primers in each animal separately. The bar represents the mean ± SD of expression data from n = 4 to 5 rat livers relative to the house-keeping control (Hrpt1). *, ** Statistically significant difference compared to the control SC group at p < 0.5 or p < 0.01 respectively. #, ##, ### - Statistically significant difference compared to the control CAF group at p < 0.5, < 0.01 or < 0.001, respectively. Red numbers indicate fold change in CAF compared to SC group. Green and black numbers indicate the fold change in CAF+BPF group compared to CAF and SC groups, respectively.
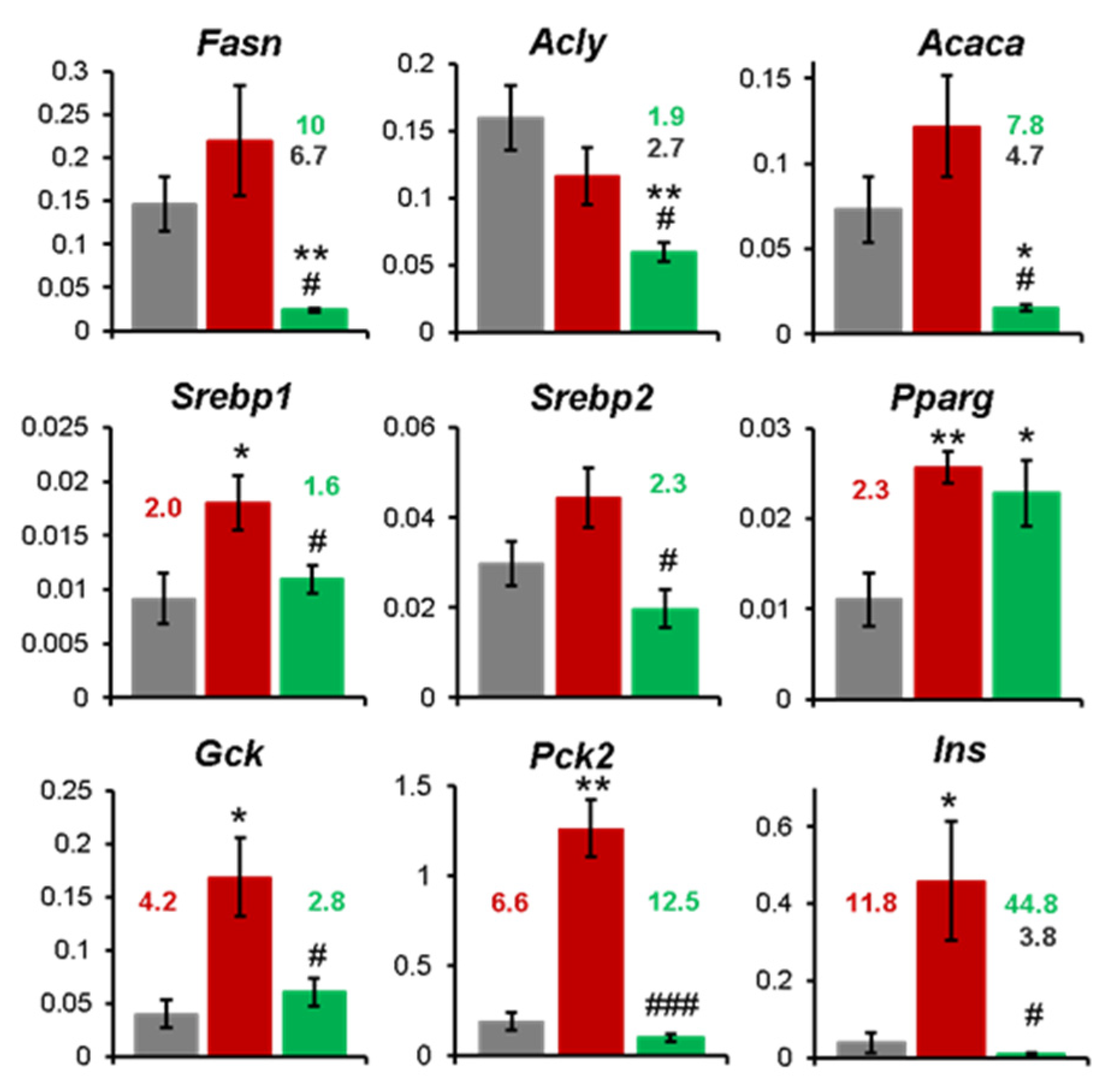
Figure 8.
Western blot analysis of protein products of selected genes modulated by CAF diet and BPF. (A) Representative blots showing 3 rat livers, for each group, for ACACA, ACLY, PCK2 and GCK. ADRP/Perilipin 2 has been shown as a marker for steatosis or lipid content and GAPDH as a loading control. (B) OD analysis of expression levels of proteins as in A compared to GAPDH n = 5 to 6 rat livers for each group. (C) Representative blots showing protein lysates from 3 rat livers for each group for autophagy proteins LC3B and ATG16. alpha-tubulin (TUBA) was used as loading control. (D) OD ratio of proteins as in C compared to TUBA in n = 5 to 6 rat livers for each group. Bars show the mean OD ratio ± SEM, normalized to the mean of SC group. Statistical analysis: One-way ANOVA with Tukey’s post-test or with LSD Fisher test for GCK OD analysis. *, **, ***, **** significant difference compared with control SC group at p < 0.5, p < 0.01, p < 0.001 or p < 0.0001, respectively. #, ##, ###, #### significant difference compared with CAF group at p < 0.5, p < 0.01, p < 0.001 or p < 0.0001, respectively. Numbers on the right of blots indicate the approximate position of molecular weights expressed in kDa.
Figure 8.
Western blot analysis of protein products of selected genes modulated by CAF diet and BPF. (A) Representative blots showing 3 rat livers, for each group, for ACACA, ACLY, PCK2 and GCK. ADRP/Perilipin 2 has been shown as a marker for steatosis or lipid content and GAPDH as a loading control. (B) OD analysis of expression levels of proteins as in A compared to GAPDH n = 5 to 6 rat livers for each group. (C) Representative blots showing protein lysates from 3 rat livers for each group for autophagy proteins LC3B and ATG16. alpha-tubulin (TUBA) was used as loading control. (D) OD ratio of proteins as in C compared to TUBA in n = 5 to 6 rat livers for each group. Bars show the mean OD ratio ± SEM, normalized to the mean of SC group. Statistical analysis: One-way ANOVA with Tukey’s post-test or with LSD Fisher test for GCK OD analysis. *, **, ***, **** significant difference compared with control SC group at p < 0.5, p < 0.01, p < 0.001 or p < 0.0001, respectively. #, ##, ###, #### significant difference compared with CAF group at p < 0.5, p < 0.01, p < 0.001 or p < 0.0001, respectively. Numbers on the right of blots indicate the approximate position of molecular weights expressed in kDa.
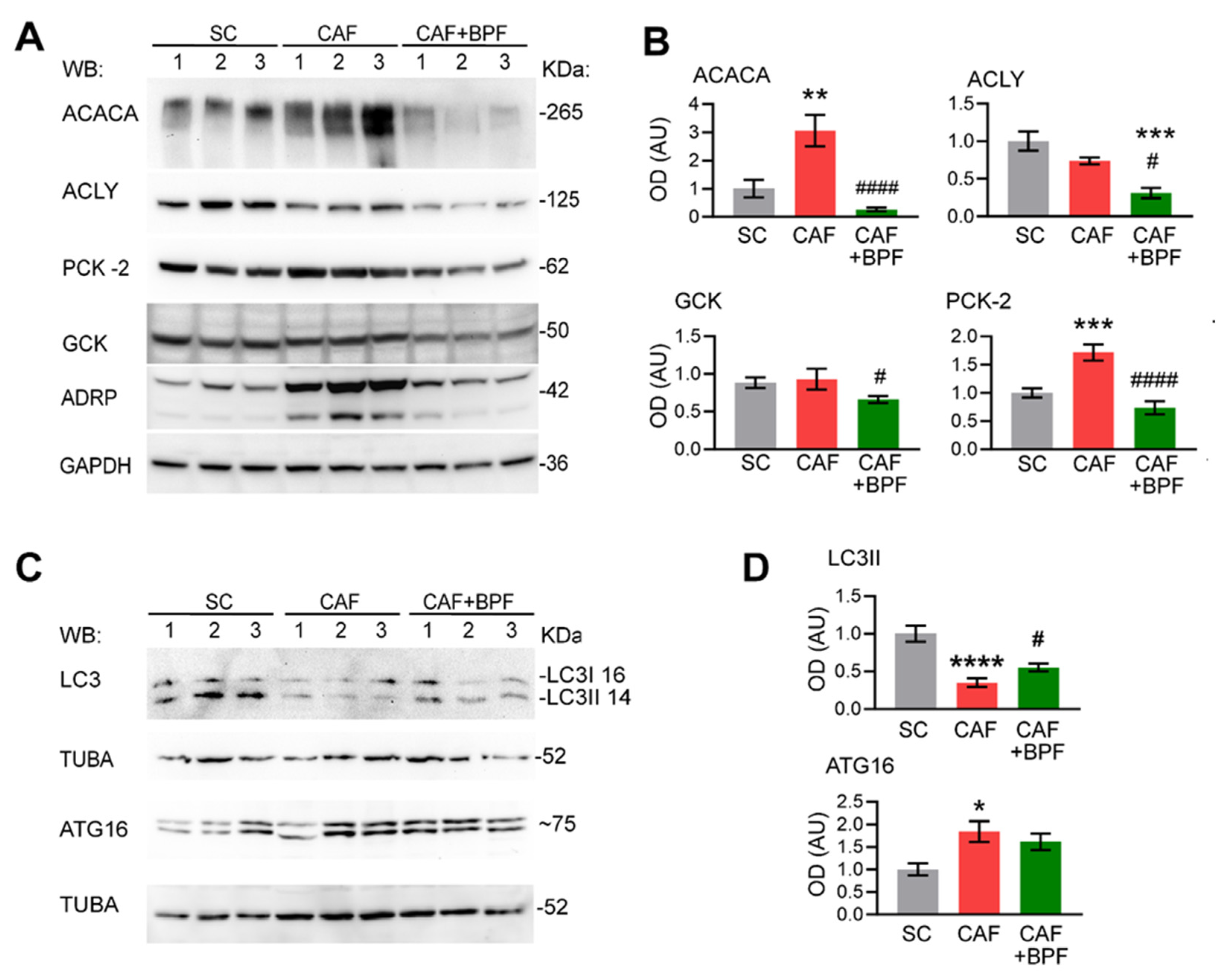
Figure 9.
Schematic representation of main lipid- and glucose-metabolism pathways and their modulation by CAF diet and BPF at mRNA level. Red and green rectangles: genes both upregulated by CAF diet and downregulated in CAF+BPF group, respectively; green rectangles: genes downregulated by BPF; violet rectangles: genes downregulated by CAF diet; light-blue rectangles and outlines: genes and proteins related to glucose metabolism; orange rectangles and outlines: genes and proteins related to lipid metabolism; blue: intermediaries in gluconeogenesis pathway.
Figure 9.
Schematic representation of main lipid- and glucose-metabolism pathways and their modulation by CAF diet and BPF at mRNA level. Red and green rectangles: genes both upregulated by CAF diet and downregulated in CAF+BPF group, respectively; green rectangles: genes downregulated by BPF; violet rectangles: genes downregulated by CAF diet; light-blue rectangles and outlines: genes and proteins related to glucose metabolism; orange rectangles and outlines: genes and proteins related to lipid metabolism; blue: intermediaries in gluconeogenesis pathway.
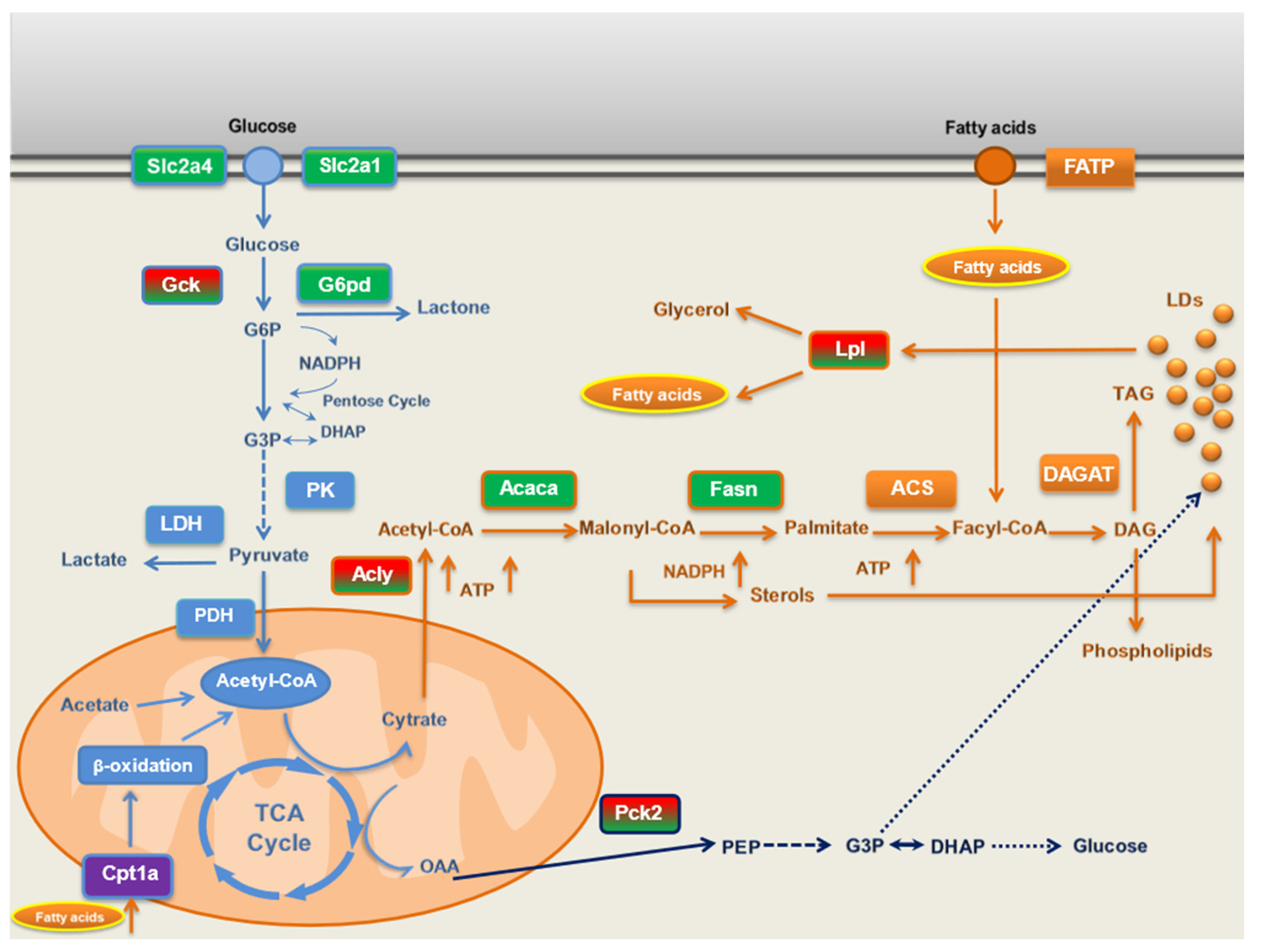

Table 1.
List of rat-specific primers for RT-qPCR analysis of gene expression. F- Forward, R- reverse.
Table 1.
List of rat-specific primers for RT-qPCR analysis of gene expression. F- Forward, R- reverse.
| ACCESSION | GENE NAME | PRIMER | SEQUENCE 5'-3' | No. BASES |
|---|---|---|---|---|
| NM_019130.2 | Ins2 | INS2-F | ATCAGCAAGCAGGTCATTGTTCCA | 24 |
| Rattus norvegicus insulin 2 | INS2-R | CTTCGCGGCGGGACATGG | 18 | |
| NM_016987.2 | Acly | ACLY-F | CGGCTCACACTGCCAACTTC | 20 |
| rat ATP citrate lyase | ACLY-R | TGGGACTGAATCTTGGGGCA | 20 | |
| NM_022193.1 | Acaca | ACC1-F | CTTCGGGGTGGTTCTTGGGT | 20 |
| acetyl-CoA carboxylase alpha | ACC1-R | TTCCAGAACGGATCCCCTGC | 20 | |
| NM_017332.1 | Fasn | FASN-F | ATTGTGGGCGGGATCAACCT | 20 |
| fatty acid synthase | FASN-R | CGGCAATACCCGTTCCCTGA | 20 | |
| NM_013124.3 | Pparg - peroxisome proliferator | PPARG-F | AGCATGGTGCCTTCGCTGAT | 20 |
| -activated receptor gamma | PPARG-R | GCCCAAACCTGATGGCATTGT | 21 | |
| NM_012565.2 | Gck | GCK-F | AGGTGTGGAGCCCAGTTGTTG | 21 |
| glucokinase | GCK-R | TCCGACTTCTGAGCCTTCTGGG | 22 | |
| NM_001108377.2 | Pck2 | PCK2-F | GGTTGAGCATGGAGGGACGA | 20 |
| phosphoenolpyruvate carboxykinase 2 | PCK2-R | CTAGCACGCGAGCGTTTTCC | 20 | |
| NM_001276707.1 | Srebf1 - sterol regulatory element | SREBF1-F | CTCTTGACCGACATCGAAGACAT | 23 |
| binding transcription factor 1 | SREBF1-R | CCCAGCATAGGGGGCATCAA | 20 | |
| NM_001033694.1 | Srebf2 - sterol regulatory element | SREBF2-F | GGCTGTCGGGTGTCATGGG | 19 |
| binding transcription factor 2 | SREBF2-R | CTGTAGCATCTCGTCGATGTCC | 22 | |
| NM_012583.2 | Hprt1 - hypoxanthine | HPRT-F | CTCATGGACTGATTATGGACAGGAC | 25 |
| phosphoribosyltransferase 1 | HPRT-R | GCAGGTCAGCAAAGAACTTATAGCC | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



