
Submitted:
12 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
Slimy sculpin (Uranidea cognata) inhabitat coldwater streams in southeastern Minnesota, USA, many of which were subjected to probable 2000-year flood events in August 2007. Floods scoured streambeds, created new stream channels, and greatly reduced benthic invertebrate communities that serve as the primary food resource for sculpin. Diets and Fulton condition of sculpin in Gilmore Creek (with moderate flooding) and Garvin Brook (with very severe flooding) had been examined just prior to flooding, and were re-examined two weeks after flooding to assess possible diet and condition changes. Diets, condition, and reproductive fitness of sculpin were examined seven months post-flood in these same two streams, plus nearby Trout run (also experienced very severe flooding). Sculpin condition declined significantly post-flood in Garvin Brook, but improved in Gilmore Creek. Prior to spring spawning, condition of Garvin Brook sculpin had improved, but Gilmore Creek fish condition worsened. Sculpin diets were more diverse before and after flooding in Gilmore Creek than in Garvin Brook, although diets of fish from both streams were dominated (>55%) by midge (Diptera: Chironomidae) larvae. Diets differed significantly before versus after flooding in the more severely flooded Garvin Brook, but not in Gilmore Creek. Prey number per sculpin stomach declined post-flood in Gilmore Creek, but not in Garvin Brook, although dry mass of prey/fish wet mass declined post-flood in both streams. Pre-spawn sculpin displayed no patterns in reproductive fitness (gonadosomatic index, hepatosomatic index, oocyte number) among the three streams that may have been related to flooding severity the previous summer. Although sculpin diets and condition were altered significantly immediately post-flood in the more severely flooded stream, food resource recovery apparently was rapid enough to prevent longer-term impacts on sculpin condition and reproductive fitness.
Keywords:
flooding
; benthic macroinvertebrates
; sculpin
; Fulton condition
1. Introduction
River or stream flooding (i.e., out-of-banks flow) is a common phenomenon worldwide, brought on naturally by heavy rainfall or extensive snowmelt [1], or artificially by managed releases from reservoirs [2]. Flooding can occur in a wide range of systems, from tropical rivers [3] to desert streams [4], from high mountain streams [5] to lowland coastal rivers [3,6,7], and from high-gradient low-order headwaters [5,8,9] to high-order wide-floodplain systems [10,11]. Some rivers and streams are prone to frequent, recurring flooding [5,8], whereas others rarely experience floods. Floods also exhibit extreme variability, in both magnitude (total discharge and/or elevation change relative to baseflow) and duration (time spent out-of-banks) [1,12].
Floods can dramatically alter the environments of both rivers [13] and their riparia [14], impacting both physical and biological components of aquatic habitats and their adjoining terrestrial corridors [3,8,9,15]. These effects can be either negative or positive, depending on flood variables, location within the stream or river system [11,16], and the physical or biological stream/riparian characteristic being examined [12,17,18]. From a human perspective, small floods may have neutral to positive effects on many of the ecosystem services (the benefits humans obtain from the natural functioning of the system) provided by a river or stream, but extreme floods cause losses in virtually every ecosystem service [19].
Extreme (also termed catastrophic or historic) floods can have truly devastating impacts on river and stream environments [1,5,9,16]. These once-in-100-years to once-in-2000-years events wreak havoc on instream and riparian physical features [3,9] and may have long-lasting, negative effects on aquatic biota [1,5,9]. However, some biota may be resistant and/or resilient to even these drastic floods, demonstrating amazing abilities to survive scouring flows and returning to pre-flood abundances only weeks or months after floodwaters subside [5,8,9].
Although many aquatic organisms can recover quickly from extreme floods, others may not. For example, species that are poor recolonizers, those that inhabit vulnerable habitats, or those that are in more sensitive life stages at the time of flooding are likely to be severely impacted by extreme floods [8]. Such species may include benthic fishes like sculpins (Cottidae) and longnose dace (Rhinichthys cataractae), which are considered poor recolonizers [8,20] and may be vulnerable to crushing by bedload movement of their cobble/boulder habitats during catastrophic flooding [8,21]. Young-of-year or juvenile life stages of these and other species also may be at greater risk than larger or adult fish during flood events, due to both small size and poor swimming ability [22].
Sculpin are common fish of marine and freshwater systems, especially in circumpolar regions of the Northern Hemisphere [23,24]. Many species inhabit cold and coolwater rivers and streams, where they can numerically dominate fish communities [20,25]. The slimy sculpin (Uranidea cognatus), with the widest geographical distribution of any sculpin species in North America [26,27], has been suggested as an excellent species for environmental effects monitoring due to its small size, limited mobility, early maturity, fast growth rate, high reproductive output, and high abundance [20,28,29].
A once-in-2000-years flood impacted many coldwater streams and rivers across southeastern Minnesota in August 2007 [9,30], scouring stream habitats and reducing aquatic macroinvertebrate densities by up to 95% [9]. Post-flood surveys on several regional streams indicated that salmonids (brook trout, Salvelinus fontinalis and brown trout, Salmo trutta) and slimy sculpin had survived the floods, although numbers were greatly reduced (N. Mundahl, unpublished data). However, there was concern that reduced aquatic prey resources might result in poorer fish condition, which ultimately could impact subsequent spawning success. Consequently, this study was undertaken to assess the impact of flooding on autumn diets and condition of slimy sculpin in two area streams that differed in severity of flooding and where, fortuitously, sculpin had been collected one to eight weeks prior to flooding for examination of fish condition and diets. It was hypothesized that sculpin would exhibit poorer condition and reduced diets (fewer prey and reduced prey biomass) post- versus pre-flood in both streams, with reductions greater in the stream with more severe flooding. In addition, sculpin in these two streams, plus one additional stream that also had experienced catastrophic flooding, were examined the following spring (seven months post-flood) just prior to spawning to assess diets, fish condition, and reproductive fitness. It was expected that sculpin in the two streams that had the worst flooding the previous summer would display poorer diets and condition and reduced reproductive fitness relative to sculpin in the stream that had less severe flooding.
2. Study Sites and Flooding
Gilmore Creek, Garvin Brook, and Trout Run are 1st- to 2nd-order coldwater trout streams in southeastern Minnesota, USA (Figure 1). They lie in adjacent, but separate, watersheds, each draining to the Mississippi River. All three streams contain similar fish communities comprised of native brook trout and slimy sculpin and non-native brown trout.
During a 48-hour period in August 2007, the southeastern Minnesota region experienced an historic precipitation event that produced 30 to 43 cm of rainfall across a wide area. This rain event produced variable but significant flooding on many regional streams and rivers, resulting in loss of human life and widespread damage to homes, businesses, roadways, bridges, farmlands, and parks. Numerous hillslope rockslides delivered materials to valley floors, and floodwaters cut new stream channels and filled old channels with rocks and debris. Whitewater State Park (containing Trout Run) and Farmers Community Park (Garvin Brook) were closed to the public for many months to repair damaged and destroyed bridges and other infrastructure. Both Garvin Brook and Trout Run exhibited severe channel scour, new channel formation, and large instream and riparian deposits of cobbles and boulders. In contrast, flooding in Gilmore Creek was less severe, with much less channel scour, no new channels, and no major damage to bridges or roadways. Gilmore Creek flows returned within banks only a few hours after cresting >3 m above normal water level [31]. None of these streams have gauging stations to monitor stream elevations or discharge.
3. Methods and Materials
3.1. Field Work
One to eight weeks prior to and two weeks after flooding, slimy sculpin were collected from Gilmore Creek and Garvin Brook with a backpack electrofisher. After assessing that sculpin populations were adequate to support collections, a small, representative sample of fish from each stream was anesthetized and euthanized in a solution of tricaine methanesulfonate (MS-222; 300–400 ppm), fixed in 7% formalin, and preserved in 70% ethanol. These fish were used to assess general fish condition and the composition of their diets. Seven months post-flood (during early March 2008), just prior to the spring spawning season of slimy sculpin in the region, addition sculpin were collected from Gilmore Creek and Garvin Brook, and also from nearby Trout Run (after first assessing for adequate abundance). Fish were again euthanized and preserved as described previously. As with earlier collections, these fish were used to assess sculpin condition and diets, as well as to determine their reproductive fitness.
3.2. Laboratory Work
In the laboratory, sculpin were processed to gather data on size, condition, diet, and reproductive condition. Individual fish were first towel-dried, then weighed (nearest 0.01 g wet mass) and measured (mm total length [TL]). Masses and TLs were used to calculate a Fulton condition factor (K, where K = [mass/TL3] X 100,000) for each fish [32,33]. Fish were dissected and the entire stomach was removed, cut open, and the contents washed into a watch glass. Prey items were identified (primarily to family or order) and counted with the aid of a dissecting microscope (8 to 50 X magnification). Following this, all prey items were placed into a pre-weighed aluminum pan, dried at room temperature under a constant-flow fume hood, then weighed (nearest 0.0001 g) to determine the total dry mass of all prey consumed. This mass was divided by the fish’s total wet mass to calculate a standardized prey mass (mg dry mass of prey/fish total wet mass).
Fish collected during spring (March) were examined further to assess reproductive fitness immediately prior to spawning. During dissections, the entire liver was removed and weighed (nearest 0.0001 g wet mass), and a hepatosomatic index (HSI, where HSI = [mg liver/mg total fish mass] X 100) was calculated as an assessment of fish health and reproductive status [33]. In addition, both gonads (either ovaries or testes) were removed and weighed (nearest 0.0001 g wet mass), and a gonadosomatic index (GSI, where GSI = [mg gonads/mg total fish mass] X 100) was calculated to assess reproductive condition [34]. After weighing ovaries, the total oocytes in one ovary were counted by placing the ovary in a watch glass, splitting open the ovary membrane, and counting oocytes with the aid of a dissecting microscope. Average oocyte mass was calculated by dividing the combined weights of both ovaries by the estimated total oocyte count (count in one ovary X 2).
3.3. Data Analyses
Diets of slimy sculpin were compared before and after flooding using several different variables. First, the composition of diets before and after flooding were compared within each stream. The diets of all fish from a specific stream and date were combined to form a composite diet sample, then a Bray-Curtis similarity index (IBC) [35] was calculated separately for each stream by comparing fish diets before and after flooding. A Bray-Curtis index value >0.6 was used to indicate significant diet similarity, whereas a value <0.6 indicated significantly different diets [35]. Diets were not compared between streams because sculpin diets typically vary among streams within this region under normal conditions [36]. Diets also were compared among dates and streams using two-factor analysis of variance (ANOVA) with replication for both numbers of prey per fish stomach and standardized prey weights.
Both general fish condition and reproductive fitness of slimy sculpin were examined relative to flooding. Fulton condition of fish in Gilmore Creek and Garvin Brook were assessed separately across the three sampling periods with single-factor ANOVAs to determine if condition changed relative to flooding. During spring, a series of single-factor ANOVAs were used to compare fish lengths, Fulton condition, GSIs, HSIs, oocyte numbers, and oocyte wet mass among fish from the three different streams. Separate comparisons were made for male and female fish.
4. Results
4.1. Sculpin Size and Condition
Slimy sculpin collected from three coldwater trout streams in Minnesota ranged in size from 25 to 101 mm TL and 0.1 to 18.0 g wet mass (Table 1). Neither length nor mass differed among the three streams. There were significant differences in sculpin Fulton condition before versus after flooding within both Gilmore Creek (ANOVA F2,63 = 11.67, p < 0.0001) and Garvin Brook (ANOVA F2,54 = 9.29, p = 0.0004), but the pattern of change differed between streams (Figure 2). In Gilmore Creek, sculpin were in better condition immediately after flooding compared to before flooding, or prior to spring spawning. In contrast, sculpin in Garvin Brook were in better condition before flooding and prior to spawning than they were immediately after flooding. Condition of sculpin in Trout Run prior to spring spawning was intermediate between those of fish from the other two streams (Figure 2).
4.2. Sculpin Diets
In total, 4069 individual prey organisms representing 19 different taxa were consumed by slimy sculpin examined from the three streams (Table 2). Midge larvae (Diptera: Chironomidae) dominated diets in all three streams (Figure 3), comprising 55 to 99% of individuals consumed in the different streams. Overall, diets were most diverse in Gilmore Creek and least diverse in Garvin Brook (Figure 3).
Before and immediately after the flood, sculpin diets in Gilmore Creek contained 13 total taxa, whereas diets of Garvin Brook sculpin contained only eight total taxa. Seven taxa in Gilmore Creek but only one taxon in Garvin Brook were consumed by sculpin both before and after flooding. Diets of Gilmore Creek sculpin before and after flooding were significantly similar (Bray-Curtis IBC = 0.606), whereas those of Garvin Brook sculpin differed significantly (Bray-Curtis IBC = 0.225) before versus after flooding.
The numbers and dry mass of prey contained in sculpin stomachs both displayed significant variation (Figure 4). Prey numbers varied both between streams (two-factor ANOVA with replication: between streams F1,118 = 21.23, p < 0.0001) and among sampling dates (among dates F2,118 = 4.05, p = 0.0020), although the pattern of change varied between streams (stream*date interaction F2,118 = 12.43, p < 0.0001). Numbers of prey in stomachs decreased immediately after flooding for Gilmore Creek sculpin before increasing again in spring, but prey numbers increased after flooding for Garvin Brook sculpin, before decreasing again in spring (Figure 4a). Dry masses of prey in stomachs did not differ between streams (two-factor ANOVA with replication: between streams F1,118 = 0.34, p = 0.561), but displayed significant declines post-flood (among dates F2,118 = 17.62, p < 0.0001) (Figure 4b).
4.3. Sculpin Size, Condition, and Reproductive Variables during Spring Pre-Spawn
Sizes of reproductive-age slimy sculpin did not differ among the three streams examined just prior to spring spawning, but Fulton condition of fish from Gilmore Creek was lower than that of fish in one or both of the other two streams (Table 3). HSIs, GSIs, and oocyte numbers did not differ significantly among the streams. Mean oocyte wet masses were similar for sculpin in Gilmore Creek and Garvin Brook, whereas they were significantly larger in Trout Run (Table 3).
5. Discussion
Flooding during August 2007 had severe effects on streams, rivers, and the surrounding landscape throughout southeastern Minnesota and west-central Wisconsin. Damage to human structures was extensive ($272 million US in losses) across a broad geographic area, and flooding resulted in 10 human fatalities. Streams and rivers crested many meters above flood stage [31], flowing with enough power to destroy bridges and roadways, lift houses off their foundations, sweep cars off roads, and obliterate agricultural crops [37]. Rock and mud landslides blocked some streams, forcing the creation of new channels. In the aftermath of such devastation, concern eventually shifted from anthropogenic losses to the status of fish that had inhabited these streams prior to flooding [30].
The impacts of extreme flooding on lotic fish communities can be highly variable and complicated to understand or explain [1]. Although juvenile or small-bodied fishes may be highly susceptible to heavy losses during extreme floods in some systems [1], in other systems this same size group of fish may be unaffected or actually increase in abundance post-flood [16]. Habitat complexity may play a crucial role in reducing flood impacts on fish [8]. In addition, many fish species display remarkable resistance and resilience to even extreme flooding [2,5,8,16], in fact even depending on both regular seasonal and periodic catastrophic floods to clean spawning substrates, deepen channels and pools, reduce non-native fish abundance, and reset food webs [1,5,8,16,38]. Long-term exposure of many fish species to the variable abiotic environments of lotic systems has allowed them to evolve and/or adapt unique sets of characteristics (e.g., morphological, physiological, behavioral) that allow them to better cope with flood conditions [1].
Despite the catastrophic flooding, surveys after the flooding in southeastern Minnesota indicated that many native brook trout and introduced brown trout had survived [30]. Most juvenile trout (5–8 cm total length) that had hatched out five months prior to the floods were lost, but all sizes of older fish remained in the streams, although in reduced numbers [30]. The resilience of those survivors was impressive, as they spawned two to three months after flooding and produced a record hatch of young trout during spring 2008 [30]. The same positive, post-flood response to late summer flooding has been observed previously for brown trout, attributed to improved fall spawning conditions (larger and higher-quality gravel beds) and enhanced environments for incubating trout eggs and, later, emerging fry (due to reduced fine sediments within spawning gravels) [5].
The presence of slimy sculpin in all streams after catastrophic flooding was encouraging, especially since small benthic species like sculpins are considered highly vulnerable to such destructive events [8,21,22]. Powerful flooding in Garvin Brook and Trout Run produced severe channel scouring and displacement of boulders >40 cm in diameter, simultaneously destroying sculpin habitat and food resources. The presence of sculpin in Gilmore Creek and Garvin Brook only two weeks post-flood suggest that either fish were able to find refuges [5,8] that enabled them to survive mass bedload rearrangement, or they were able to quickly recolonize stream reaches from nearby, less-impacted systems [39,40]. Although slimy sculpin often are considered as sedentary and unlikely to exhibit more than small-scale movements [29], there is increasing evidence that they are capable of both rapid and long-range movements, especially when recolonizing defaunated stream reaches [39,40,41]. An apparent lack of less-impacted reaches in Garvin Brook (severe flooding affected the entire stream, extending from headwater springs downstream to the confluence with the Mississippi River) suggests that survival in refuges was most likely, especially since even young-of-year sculpin were large enough by August to significantly reduce their vulnerability to elevated current velocities [5,22]. Slimy sculpin also have the ability to squeeze into extremely tight spaces by compressing their skull widths by up to 20% [42], which would permit them to retreat into very small openings beneath large boulders or into cracks between layers of sedimentary limestone bedrock that are common in regional streams [43], allowing them to wait out catastrophic flows in protected spaces. A study conducted in Gilmore Creek reported that slimy sculpin were more abundant a short time after the flood than they were prior to flooding [31].
Based on examination of slimy sculpin condition and diets before and after catastrophic flooding, it was apparent that differing severities of flooding did not impact these variables exactly as expected. Pre- and post-flood diets differed in Garvin Brook where flooding was most severe, but not in Gilmore Creek where flooding was less intense. However, sculpin in both streams consumed lower prey biomass after the flood, even though fish in Garvin Brook significantly increased the numbers of prey consumed. Condition of sculpin declined post-flood in Garvin Brook as expected, but sculpin condition improved after flooding in Gilmore Creek. Before spring spawning, sculpin condition had improved in Garvin Brook, but declined in Gilmore Creek. Just prior to spawning, sculpin from the two most severely flood-impacted streams were in better condition than those in the less-impacted stream, but no measure of reproductive fitness other than oocyte size differed among streams.
Reduced prey biomass in sculpin stomachs after flooding suggests that prey availability was limited [9]. The dominance of sculpin diets by Chironomidae (midge) larvae was not surprising, as these organisms were the most abundant aquatic invertebrates after the flood and comprised a large proportion of the invertebrate assemblage for many months thereafter [9]. Sculpin are opportunistic feeders, taking advantage of whatever prey are available and abundant [36], so their ability to find and consume prey after a flood should be adequate. Differing pre- versus post-flood diets in Garvin Brook, but not in Gilmore Creek, follows expectations that more severe flooding would produce greater impacts on sculpin diets, but the increased numbers of prey in Garvin Brook sculpin stomachs relative to the pre-flood survey appears counterintuitive. Reduced condition of sculpin post-flood in Garvin Brook suggest that fish may have experienced a one-to-two-week period of food deprivation during and after flooding, followed by compensatory increases in feeding rate and prey consumption [44]. However, the small size of midge larvae still resulted in reductions in ingested biomass post-flood relative to pre-flood feeding. Reduced body condition of sculpin has been reported previously when fish consumed smaller, lower-energy prey [45].
As expected, condition of sculpin in flood-damaged Garvin Brook declined significantly after the flood, likely attributable to a combination of reduced prey resources and increased stress post-flood [9,32,33]. In contrast, sculpin in Gilmore Creek exhibited improved condition post-flood, despite reductions in numbers and dry mass of prey in stomachs. Condition is a responsive indicator of exposure of sculpin to a variety of stressors [29,46], so significant reductions in condition post-flood in Garvin Brook are suggestive of stressful environmental conditions in the few weeks following floods. Environmental conditions apparently were better in less-impacted Gilmore Creek, since fish displayed improved condition after flooding. This result is difficult to explain from a diet perspective, since Gilmore Creek sculpin contained fewer prey items in their stomachs and reduced dry weight prey biomass after flooding compared to before. Perhaps the more diverse diet of Gilmore Creek sculpin, containing organisms considerably larger than midge larvae (especially caddisfly larvae, a variety of crustaceans, and small fish), may have provided nutrients sufficient to maintain and even improve overall sculpin body condition post-flood [45].
Prior to spring spawning, temperate zone fishes in general, and sculpin in particular, direct energy resources to their reproductive organs to maximize their reproductive fitness [47,48]. Prey resources often are abundant and diverse during this time period [9], although territorial behaviors among fish may result in reduced food intake in those individuals relegated to poorer habitats [47,49]. As cold-adapted species capable of feeding effectively at winter temperatures [48], sculpin should lose little of their endogenous energy reserves and maintain condition through the winter months. However, reduced food (energy) intake during winter and early spring due to reduced prey resources post-flooding [9], while reproductive organs were enlarging in preparation for spawning, could have resulted in the reduced condition factors observed for sculpin in Gilmore Creek (both males and females).
The lack of any significant differences in GSI or HSI indices, or in oocyte numbers of sculpin among the three streams examined indicates that there were no observable effects of differing flood magnitude on sculpin reproduction seven months post-flood. Although differing availabilities of prey in the various systems may have resulted in altered food intake and possible declines in overall fish condition, this was not translated into reduced reproductive fitness. Even under adverse environmental conditions, reproductive tradeoffs in fish are common [48]. Many fish species, apparently including sculpin, can allocate significant energy reserves toward maximizing their reproductive output, even if that allocation may compromise post-spawning survival of some fish [48]. Such allocations toward reproduction have been reported in other fish species deprived of food prior to spawning [48].
6. Conclusions
Slimy sculpin diets and condition changed significantly immediately post-flood in the more severely flooded stream, but post-flood prey recovery was rapid enough to prevent any extended impacts on sculpin condition or reproductive fitness. Pre-spawn slimy sculpin displayed no obvious negative impacts to reproductive fitness (GSI, HSI, oocyte number) among three streams seven months after flooding that may have been related to flooding severity the previous summer. Taken together, these observations suggest that slimy sculpin are extremely resilient to even catastrophic flooding, able to survive and reproduce after exposure to destructive flows that rearranged stream habitats, reduced prey availability, and caused numerous human fatalities and extensive loss of infrastructure.
Funding
Partial funding for this project was provided by the Minnesota Department of Natural Resources, Division of Parks and Trails.
Institutional Review Board Statement
Fish collections were carried out under special permits (Numbers 14055, 14728) from the Minnesota Department of Natural Resources, Division of Fish and Wildlife, Section of Fisheries, and were conducted with the approval of the Winona State University Animal Care and Use Committee (1310073-2). This research complied with all ethical standards.
Data Availability Statement
Data available from the author upon reasonable request.
Acknowledgments
I thank the personnel of Farmers Community Park and Whitewater State Park for granting access to the parks while they were closed to the public after flooding, and Winona State Biology students who assisted with fish collections.
Conflicts of Interest
The author declares no competing interest.
References
- Hickey, J.T.; Salas, J.D. Environmental effects of extreme floods. United States – Italy Research Workshop on the Hydrometeorology, Impacts, and Management of Extreme Floods, Perugia, Italy, November 1995. 22 p. 19 November.
- Schultz, A.A.; Maughan, O.E.; Bonar, S.A.; Matter, W.J. Effects of flooding on abundance of native and nonnative fishes downstream from a small impoundment. N. Am. J. Fish. Manag. 2003, 23, 503–511. [Google Scholar] [CrossRef]
- Rayner, T.S.; Pusey, B.J.; Pearson, R.G. Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, north-east Queensland: towards a new conceptual framework for understanding fish-habitat dynamics in small tropical rivers. Mar. Freshw. Res. 2008, 59, 97–116. [Google Scholar] [CrossRef]
- Eby, L.A.; Fagan, W.F.; Minckley, W.L. Variability and dynamics of a desert stream community. Ecol. Appl. 2003, 13, 1566–1579. [Google Scholar] [CrossRef]
- George, S.D.; Baldigo, B.P.; Smith, A.J.; Robinson, G.R. Effects of extreme floods on trout populations and fish communities in a Catskill Mountain river. Freshw. Biol. 2015, 60, 2511–2522. [Google Scholar] [CrossRef]
- Adams, S.M.; Greeley, M.S.; Law, J.M.; Noga, E.J.; Zelikoff, J.T. Application of multiple sublethal stress indicators to assess the health of fish in Pamlico Sound following extensive flooding. Estuaries 2003, 26, 1365–1382. [Google Scholar] [CrossRef]
- Blewett, D.A.; Stevens, P.W.; Carter, J. Ecological effects of river flooding on abundance and body condition of a large, euryhaline fish. Mar. Ecol. Prog. Ser. 2017, 563, 211–218. [Google Scholar] [CrossRef]
- Pearsons, T.N.; Li, H.W.; Lamberti, G.A. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Am. Fish. Soc. 1992, 121, 427–436. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Hunt, A.M. Recovery of stream invertebrates after catastrophic flooding in southeastern Minnesota, USA. J. Freshw. Ecol. 2011, 26, 445–457. [Google Scholar] [CrossRef]
- King, A.J.; Humphries, P.; Lake, P.S. Fish recruitment on floodplains: the roles of patterns of flooding and life history characteristics. Can. J. Fish. Aquat. Sci. 2003, 60, 773–786. [Google Scholar] [CrossRef]
- Pereira, L.S.; Tencatt, L.F.C.; Dias, R.M.; de Oliveira, A.G.; Agostinho, A.A. Effects of long and short flooding years on the feeding ecology of piscivorous fish in floodplain river systems. Hydrobiologia 2017, 795, 65–80. [Google Scholar] [CrossRef]
- Death, R.D. The effect of floods on aquatic invertebrate communities. In Aquatic Insects: Challenges to Populations; Lancaster, J., Briers, R.A., Eds.; Proceedings of the Royal Society’s 24th Symposium; CAB International: UK, 2008; pp. 103–121. [Google Scholar]
- Allan, J.D.; Castillo, M.M.; Cappas, K.A. Stream Ecology: Structure and Function of Running Waters, 3rd edition; Springer Nature Switzerland: Cham, Switzerland, 2021. [Google Scholar]
- Naiman, R.J.; Décamps, H.; McClain, M.E. Riparia: Ecology, Conservation, and Management of Streamside Communities; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Yin, Y.; Nelson, J.C.; Swenson, G.V.; Langrehr, H.A.; Blackburn, T.A. Tree mortality in the upper Mississippi River and floodplain following an extreme flood in 1993. In Long Term Resource Monitoring Program 1993 Flood Observations (LTRMP 94-S011); National Biological Service, Environmental Management Technical Center: Onalaska, WI, USA, 1994. [Google Scholar]
- Carlson, A.K.; Fincel, M.J.; Longhenry, C.M.; Graeb, B.D.S. Effects of flooding on fishes and aquatic habitats in a Missouri River delta. J. Freshw. Ecol. 2016, 31, 271–288. [Google Scholar] [CrossRef]
- Harrell, H.L. Response of the Devil’s River (Texas) fish community to flooding. Copeia 1978, 1978, 60–68. [Google Scholar] [CrossRef]
- Piniewski, M.; Prudhomme, C.; Acreman, M.C.; Tylec, L.; Oglęcki, P.; Okruszko, T. Responses of fish and invertebrates to floods and droughts in Europe. Ecohydrology 2017, 10, e1793. [Google Scholar] [CrossRef]
- Talbot, J.J.; Bennett, E.M.; Cassell, K.; Hanes, D.M.; Minor, E.C.; Paerl, H.; Raymond, P.A.; Vargas, R.; Vidon, P.G.; Wollheim, W.; Xenopoulos, M.A. The impact of flooding on aquatic ecosystem services. Biogeochemistry 2018, 141, 439–461. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.A.; Cunjak, R.A. Influence of water temperature and streambed stability on the abundance and distribution of slimy sculpin (Cottus cognatus). Environ. Biol. Fishes 2006, 80, 9–22. [Google Scholar] [CrossRef]
- Erman, D.C.; Andrews, E.D.; Yoder-Williams, M. Effects of winter floods on fishes in the Sierra Nevada. Can. J. Fish. Aquat. Sci. 1988, 45, 2195–2200. [Google Scholar] [CrossRef]
- Harvey, B.C. Susceptibility of young-of-the-year fishes to downstream displacement by flooding. Trans. Am. Fish. Soc. 1987, 116, 851–855. [Google Scholar] [CrossRef]
- Berra, T.M. Freshwater Fish Distribution; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Nelson, J.S. Fishes of the World, 4th edition; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Baldigo, B.P.; Lawrence, G.B. Composition of fish communities in relation to stream acidification and habitat in the Neversink River, New York. Trans. Am. Fish. Soc. 2000, 129, 60–76. [Google Scholar] [CrossRef]
- Wallace, R.L.; McAllister, D.E.; Rankin, M. Cottus cognatus Richardson, slimy sculpin. In Atlas of North American Freshwater Fishes; Lee, D.S., Gilbert, C.R., Hocutt, C.H., Jenkins, R.E., McAllister, D.E., Stauffer, J.R., Jr., Eds.; North Carolina Biological Survey Publication #1980-12; North Carolina Museum of Natural Sciences: Raleigh, NC, USA, 1980; p. 808. [Google Scholar]
- Page, L.M.; Burr, B.M. A Field Guide to Freshwater Fishes: North America North of Mexico; Houghton Mifflin Company: Boston, MA, USA, 1991. [Google Scholar]
- Gray, M.A.; Munkittrick, K.R. An effects-based assessment of slimy sculpin (Cottus cognatus) populations in agricultural regions of northwestern New Brunswick. Water Qual. Res. J. Canada 2005, 40, 16–27. [Google Scholar] [CrossRef]
- Gray, M.A.; Curry, R.A.; Arciszewski, T.J.; Munkittrick, K.R.; Brasfield, S.M. The biology and ecology of slimy sculpin: A recipe for effective environmental monitoring. Facets 2018, 3, 103–127. [Google Scholar] [CrossRef]
- Keillor, L. Fishing after the flood. Minn. Conserv. Vol. 2010, 73(431), 8–17. [Google Scholar]
- Cochran, P.A.; Stagg, T.W. Response of a fish assemblage to severe flooding in Gilmore Creek, a southeastern Minnesota trout stream. J. Freshw. Ecol. 2011, 26, 77–84. [Google Scholar] [CrossRef]
- Ney, J.J. Practical use of biological statistics. In Inland Fisheries Management in North America, 2nd ed.; Kohler, C.C., Hubert, W.A., Eds.; American Fisheries Society: Bethesda, MD, USA, 1999; pp. 167–191. [Google Scholar]
- Kaller, M.D.; Green, C.C.; Haukenes, A.H. Growth and development. In Methods for Fish Biology, 2nd edition; Midway, S.R., Hasler, C.T., Chakrabarty, P., Eds.; American Fisheries Society: Bethesda, MD, USA, 2022; pp. 347–398. [Google Scholar]
- Luckenbach, J.A.; Guzmán, J.M. Reproduction. In Methods for Fish Biology, 2nd edition; Midway, S.R., Hasler, C.T., Chakrabarty, P., Eds.; American Fisheries Society: Bethesda, MD, USA, 2022.; pp. 399–448. [Google Scholar]
- Brower, J.E.; Zar, J.H.; von Ende, C.N. Field and Laboratory Methods for General Ecology, 4th editionWCB McGraw-Hill: Boston, MA, USA, 1998. [Google Scholar]
- Mundahl, N.D.; Mundahl, D.E.; Merten, E.C. Success of slimy sculpin reintroductions in Minnesota trout streams: Influence of feeding and diets. Am. Midl. Nat. 2012, 168, 162–183. [Google Scholar] [CrossRef]
- Strachura, S. Six dead as flooding hits SE Minnesota. Minnesota Public Radio News. 19 August 2007. https://www.mprnews.org/story/2007/08/19/flood Accessed: 5 April 2024.
- Power, M.E.; Parker, M.S.; Dietrich, W.E. Seasonal reassembly of a river food web: floods, droughts, and impacts of fish. Ecol. Monogr. 2008, 78, 263–282. [Google Scholar] [CrossRef]
- Clarke, A.D.; Telmer, K.H.; Shrimpton, J.M. Movement patterns of fish revealed by otolith microchemistry: A comparison of putative migratory and resident species. Environ. Biol. Fish. 2015, 98, 1583–1597. [Google Scholar] [CrossRef]
- Weinstein, S.Y.; Coombs, J.A.; Nislow, K.H.; Riley, C.; Roy, A.H.; Whiteley, A.R. Evaluating the effects of barriers on slimy sculpin movement and population connectivity using novel sibship-based and traditional genetic metrics. Trans. Am. Fish. Soc. 2019, 148, 1117–1131. [Google Scholar] [CrossRef]
- Mundahl, N.D. Recovery of slimy sculpin (Uranidea cognata) after an autumn fish kill in the headwaters of a Minnesota trout stream. Water 2024, 16, 283. [Google Scholar] [CrossRef]
- Marsden, J.E.; Tobi, H. Sculpin predation on lake trout eggs in interstices: skull compression as a novel foraging mechanism. Copeia 2014, 2014, 654–658. [Google Scholar] [CrossRef]
- Waters, T. The Streams and Rivers of Minnesota; University of Minnesota Press: Minneapolis, MN, USA, 1977. [Google Scholar]
- Ali, M.; Nicieza, A.; Wootton, R.J. Compensatory growth in fishes: a response to growth depression. Fish Fish. 2003, 4, 147–190. [Google Scholar] [CrossRef]
- McGinley, E.J.; Raesley, R.L.; Seddon, W.L. The effects of embeddedness on the seasonal feeding of mottled sculpin. Am. Midl. Nat. 2013, 170, 213–228. [Google Scholar] [CrossRef]
- Hards, A.R.; Gray, M.A.; Noël, S.C.; Cunjak, R.A. Utility of condition indices as predictors of lipid content in slimy sculpin (Cottus cognatus). Diversity 2019, 11, 71. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Collins, N.C.; Cabana, G. A comparative study of sexual selection and reproductive investment in the slimy sculpin, Cottus cognatus. Oikos 1987, 51, 156–162. [Google Scholar] [CrossRef]
- Diana, J.S. Biology and Ecology of Fishes, 2nd edition; Cooper Publishing Group: Traverse City, MI, USA, 2004. [Google Scholar]
- Petty, J.T.; Grossman, G.D. Patch selection by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshw. Biol. 1996, 35, 261–276. [Google Scholar] [CrossRef]
Figure 1.
Maps indicating location of the study area within the upper midwestern USA (upper: Driftless Area Ecoregion highlighted in gray), and locations of the three study sites (lower: sites indicated by red dots) within the stream network of southeastern Minnesota (inset highlights southeastern Minnesota). Sites (left to right) were located on Trout Run, Garvin Brook, and Gilmore Creek.
Figure 1.
Maps indicating location of the study area within the upper midwestern USA (upper: Driftless Area Ecoregion highlighted in gray), and locations of the three study sites (lower: sites indicated by red dots) within the stream network of southeastern Minnesota (inset highlights southeastern Minnesota). Sites (left to right) were located on Trout Run, Garvin Brook, and Gilmore Creek.
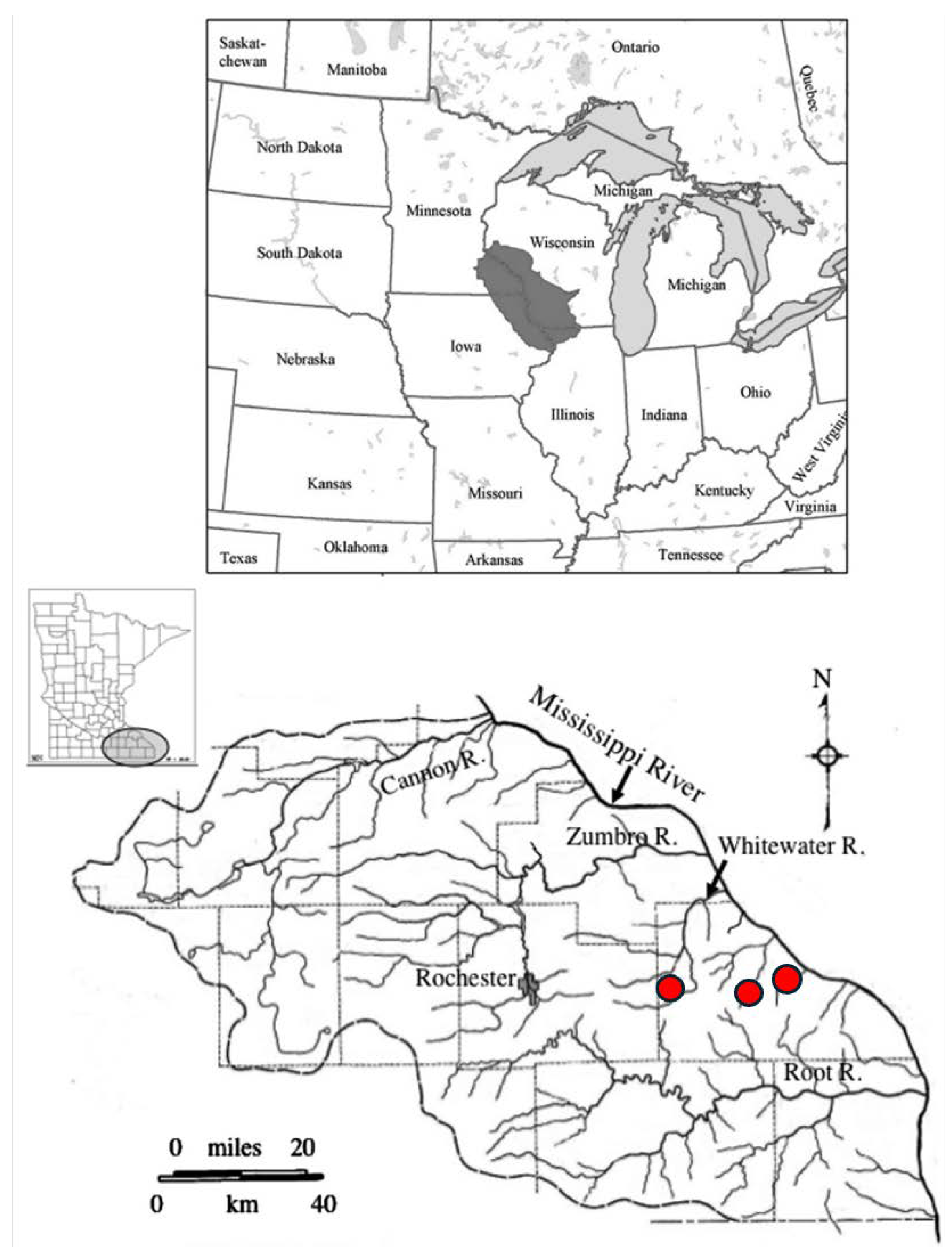
Figure 2.
Mean (+ one standard deviation) Fulton condition factors of slimy sculpin in southeastern Minnesota streams before and after August 2007 flooding, and prior to 2008 spring spawning.
Figure 2.
Mean (+ one standard deviation) Fulton condition factors of slimy sculpin in southeastern Minnesota streams before and after August 2007 flooding, and prior to 2008 spring spawning.
Figure 3.
Relative abundances of various prey items in the diets of slimy sculpin in three southeastern Minnesota streams.
Figure 3.
Relative abundances of various prey items in the diets of slimy sculpin in three southeastern Minnesota streams.

Figure 4.
Mean (+ one standard deviation) prey abundance per stomach (a) and standardized dry prey mass per fish (b) for slimy sculpin in southeastern Minnesota streams before and after August 2007 flooding, and prior to 2008 spring spawning.
Figure 4.
Mean (+ one standard deviation) prey abundance per stomach (a) and standardized dry prey mass per fish (b) for slimy sculpin in southeastern Minnesota streams before and after August 2007 flooding, and prior to 2008 spring spawning.
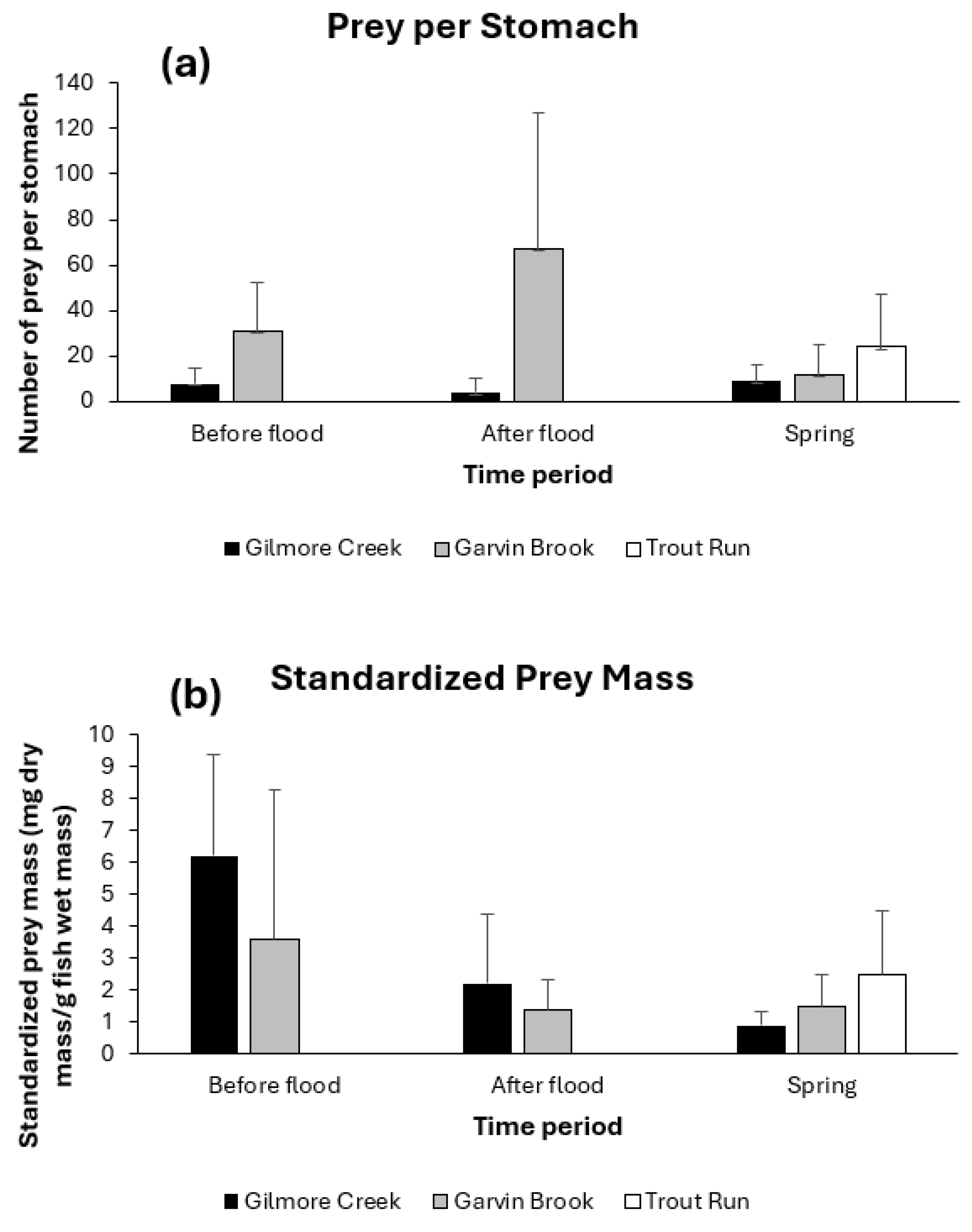

Table 1.
Size ranges of slimy sculpin collected from three streams in southeastern Minnesota before and after catastrophic flooding, 2007 and 2008. Single-factor ANOVA test statistics comparing length and mass among streams are included. n = sample size.
Table 1.
Size ranges of slimy sculpin collected from three streams in southeastern Minnesota before and after catastrophic flooding, 2007 and 2008. Single-factor ANOVA test statistics comparing length and mass among streams are included. n = sample size.
| Stream | n | Total length (mm) | Wet mass (g) |
|---|---|---|---|
| Gilmore Creek | 64 | 27–101 | 0.2–10.1 |
| Garvin Brook | 55 | 25–100 | 0.1–15.6 |
| Trout Run | 49 | 38–101 | 0.8–18.0 |
| F2,167 = 2.09 | F2,167 = 1.87 | ||
| p = 0.127 | p = 0.157 |
Table 2.
Prey taxa included in the diets of slimy sculpins from three streams in southeastern Minnesota, 2007–2008. Unid. = unidentified.
Table 2.
Prey taxa included in the diets of slimy sculpins from three streams in southeastern Minnesota, 2007–2008. Unid. = unidentified.
| Taxa | Gilmore Creek | Garvin Brook | Trout Run | ||
|---|---|---|---|---|---|
| DIPTERA | Chironomidae | X | X | X | |
| Simuliidae | X | X | X | ||
| Tipula | X | ||||
| Dicranota | X | ||||
| Empididae | X | X | |||
| EPHEMEROPTERA | X | X | X | ||
| TRICHOPTERA | Brachycentrus | X | |||
| Hydropsyche | X | X | |||
| Limnephilus | X | X | |||
| Glossosoma | X | ||||
| COLEOPTERA | Agabus | X | |||
| Unid. beetle | X | ||||
| CRUSTACEA | Amphipoda | X | X | ||
| Isopoda | X | X | |||
| Ostracoda | X | ||||
| GASTROPODA | X | ||||
| OLIGOCHAETA | X | X | X | ||
| NEMATOMORPHA | X | ||||
| FISH | X | ||||
Table 3.
Size, condition, and reproductive variables of female and male slimy sculpin from three streams in southeastern Minnesota, March 2008. Values are means, with standard deviations in parentheses. ANOVA test statistics (F and p values) are included for each variable-sex combination. F = female, M = male, n = sample size, GSI = gonadosomatic index, HSI = hepatosomatic index.
Table 3.
Size, condition, and reproductive variables of female and male slimy sculpin from three streams in southeastern Minnesota, March 2008. Values are means, with standard deviations in parentheses. ANOVA test statistics (F and p values) are included for each variable-sex combination. F = female, M = male, n = sample size, GSI = gonadosomatic index, HSI = hepatosomatic index.
| Variable | Sex | Gilmore Creek | Garvin Brook | Trout Run | F | p |
|---|---|---|---|---|---|---|
| n | F | 5 | 11 | 13 | ||
| M | 6 | 7 | 8 | |||
| Total length | F | 74 (8) | 73 (7) | 77 (5) | 1.13 | 0.339 |
| (mm) | M | 84 (5) | 92 (8) | 91 (8) | 2.28 | 0.131 |
| Fulton | F | 1.36 (0.11) | 1.62 (0.11) | 1.44 (0.14) | 9.64 | <0.001 |
| condition | M | 1.39 (0.11) | 1.64 (0.11) | 1.58 (0.13) | 8.26 | 0.003 |
| GSI (%) | F | 14.74 (1.75) | 17.28 (1.63) | 15.99 (2.13) | 3.33 | 0.052 |
| M | 1.82 (0.21) | 1.93 (0.42) | 1.94 (0.57) | 0.15 | 0.864 | |
| HSI (%) | F | 2.72 (0.43) | 3.28 (0.49) | 2.95 (0.21) | 2.65 | 0.103 |
| M | 1.47 (0.18) | 1.51 (0.29) | 1.53 (0.06) | 0.11 | 0.896 | |
| Oocyte number | F | 221 (61) | 271 (61) | 223 (60) | 2.18 | 0.134 |
| Mean oocyte wet mass (mg) | F | 3.7 (0.3) | 4.1 (0.6) | 4.9 (1.0) | 4.90 | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



