
Submitted:
12 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
The legume-rhizobia symbiosis is one of the most important plant-microbe interactions in sustainable agriculture due to its ability to provide much needed N to cropping systems. This interaction is mediated by the mutual recognition of signaling molecules from the two partners, namely legumes and rhizobia. With legumes, these molecules are in the form of flavonoids and anthocyanins, which are responsible for the pigmentation of plants parts, such as seeds, flowers, fruits and even leaves. Seed coat pigmentation in legumes is a dominant factor influencing gene expression relating to N2 fixation, and may be responsible for the different N2-fixing abilities observed among legume genotypes under field conditions in African soils. Cowpea, Kersting’s bean and Bambara groundnut landraces with black seed coat color are reported to release higher concentrations of nod-gene-inducing flavonoids and anthocyanins when compared to the Red and Cream landraces, hence the black seed coat pigmentation is considered a biomarker for enhanced nodulation and N2 fixation. Cowpea, Bambara groundnut and Kersting’s bean with differing seed coat colors are known to attract different native soil rhizobia, confirmable by PCR-RFLP analysis of bacterial DNA from root nodules of these legumes. Even when seeds of the same legume with diverse seed coat colors were planted together in one hole, the nodulating bradyrhizobia clustered differently in the PCR-RFLP dendrograms. In one study, Kersting’s groundnut, Bambara groundnut and cowpea with differing seed coat colors were selectively nodulated by different bradyrhizobial species. Multilocus sequence analysis showed that different Bradyrhizobium species nodulated the Kersting's bean based on seed coat color. Phylogenetic analysis also placed the bradyrhizobial isolates in close proximity to different Bradyrhizobium species such as B. vignae 7-2T, B. subterraneum 58 2-1T, B. kavangense 14-3T, B. liaoningense 2281 (USDA 3622)T, B. yuanmingense LMG 21827T, B. huanghuaihaiense CCBAU 23303T, B. pachyrhizi PAC48T, and a reference type strain of B. elkanii according to seed coat color. Using 16S rDNA amplicon sequencing, we also found significant selective influences of seed coat pigmentation on microbial community structure in the rhizosphere of five Kersting’s groundnut landraces. For example, the rhizosphere of Belane Mottled landrace was dominated by Proteobacteria, while Bacteroidetes dominated the rhizospheres of the other landraces. With legumes, seed coat pigmentation therefore plays a dominant role in the selection of the bacterial symbiotic partner.
Keywords:
flavonoids
; anthocyanin
; pigmentation
; soil microbes
; disease resistance
; nodulation and nitrogen fixation
1. Introduction
Plants and microorganisms have coexisted for millions of years. At the root-soil interface, plants and microorganisms engage in complex signaling to establish beneficial associations known as symbiosis[1]. These interactions are crucial for nutrient acquisition and fitness improvement, and serve as the foundation for symbiotic relationships, with the most ancient being those formed with nitrogen-fixing bacteria. A key alternative to soil N improvement via fertilization in cropping systems is biological N2 fixation (BNF) [2], a process that has been recognized as the most smart environmentally-friendly and economically-sustainable alternative for overcoming soil N infertility [3]. BNF contribution to the N economy of terrestrial ecosystems can be symbiotic, associative, or by free-living microorganisms [4]. However, symbiotic N2 fixation in legumes has contributed the most N to global agriculture due to its ability to fix large amounts of biological N in cropping systems for use by the legume and succeeding crops [5].
Within the broader context of the legume-rhizobia symbiosis, seed coat pigmentation has emerged as a significant factor influencing the establishment of this plant-bacterial association [6]. Though often overlooked, seed coat pigmentation plays a crucial role in mediating partner selection during the early stages of the legume host/strain symbiosis.
Effective nodulation and N2 fixation depend on the presence of host plants and compatible nodulating soil bacteria in adequate numbers. Once these conditions are met, legumes offer a great opportunity for improving the N economy of nutrient-poor soils through their ability to establish effective N2-fixing symbiosis with compatible soil bacteria that can contribute symbiotic N to cropping systems [7,8]. While N2-fixing bacteria can contribute significantly to this BNF process, the interplay between seed coat pigmentation and N2 fixation remains relatively unexplored and untapped for sustainable development. The aim of this review is to highlight the role of seed coat pigmentation in partnership development and nitrogen fixation in the legume/rhizobia symbiosis.
2. A Historical Perspective on Legume Seed Coat Pigmentation
Grain legumes have played an important role in the development of Neolithic agriculture some 12,000 years ago [9] The evolution of seed coat pigmentation spans millions of years, reflecting both dynamic environmental pressures and intricate adaptations [10]. Diversity in seed coat pigmentation is evident across the plant kingdom, and reflects the unique ecological niches and adaptive strategies adopted by different plant species. A wide range of seed coat colors exist within and among legume species [9] (see Figure 1).
The color of legume seed coats is a trait that is influenced by polyphenolic substances, and holds significant evolutionary and agricultural implications. However, crop domestication processes have led to the loss of seed coat pigmentation, possibly as a trade-off for protection from foragers [11,12]. Wild chickpea (Cicer arietinum L.) progenitors, for example, are known to exhibit seed coat pigmentation that mimicks soil color, thus emphasizing the evolutionary role of seed coat color [13]. Pigmentation loss has however been linked to the level of bitter-tasting compounds, which suggest a potential relationship between seed coat color and metabolite concentration [14].
Legume genotypes have been shown to fix varying amounts of symbiotic N. Although the effect of seed coat color in legume nodulation has been established [6,15,16,17,18], little is known about its influence on the choice of rhizobia by legumes for symbiotic partnership, root nodulation and N contribution. The importance of seed coat color in legume nodulation has been properly documented [15,17,19,20,21], and darker seed coat color is reported to contain greater levels of flavonoids, anthocyanins, and anthocyanidins that promote superior symbiotic functioning [17,22,23]. Hungria and Phillips [17] and dos Santos Sousa [24] have, in fact, shown that common bean (Phaseolus vulgaris L.) seedlings generated from a black-seeded genotype formed more root nodules compared to its white isogenic counterpart.
Though known for their aesthetics in Africa, legume seed coat coloration goes beyond mere aesthetics as it serves as an adaptive trait with functional significance. The ability of seed coats to exhibit diverse colors is often linked to ecological adaptations such as UV-B protection, signaling mechanisms, and interactions with symbiotic microorganisms. An understanding of the adaptive significance of seed coat color is likely to unveil its role as a dynamic and strategic trait in the evolutionary arms race between plants and microbes. In the African marketplace, legume accessions are often given local names based on the color of the testa or seed coat [23]. From the primitive seed coat color to the sophisticated pigmentations observed in legumes today seems to suggest a historical perspective on the fascinating journey of seed coat pigmentation.
3. Mechanisms of Partner Selection in Symbiotic Interactions
Partner selection in symbiotic interactions involves intricate genetic and molecular mechanisms that govern the recognition and compatibility between plants and microorganisms. The rhizobia-legume symbiosis is an excellent model system for studying partner selection in mutualistic relationships. Despite the differences in host specificity during symbiotic establishment, commonalities and overlaps do exist in signaling pathways that can provide insights for understanding how seed coat pigmentation contributes to partner recognition and selection during nodulation. Although seed coat pigmentation is a significant determinant in partner recognition during symbiotic establishment, our understanding of these roles is essential for unraveling the broader dynamics of symbiosis.
At the molecular level, the pathways linking seed coat pigmentation to N2 fixation remain a focal point of investigation. In the absence of effective indigenous soil bacteria, rhizobial strains must be applied as inoculants for nodulation to occur in legumes. The ability of rhizobia to successfully establish a N2-fixing relationship with a legume depends on the symbiotic signals involved in the molecular conversation between legume and rhizobia during nodule formation [25]. Usually the first signals are produced by the plant in the form of flavonoids which interact with nodD proteins of the rhizobia. The nodD-flavonoid complex then activates the transcription of nod boxes that cause the deformation of root hairs, thus allowing entry of the rhizobia into plant roots [25].
The flavonoids released by legume seed coats and young roots, act as chemo-attractants, cell growth enhancers, and nod-gene inducers in symbiotic rhizobia [16,26,27]. The same flavonoid molecule can therefore elicit different responses in different rhizobia [18]. Unraveling these pathways can provide a molecular framework for understanding the functional role of seed coat pigmentation in N2-fixing symbioses
4. Genetic Determinants of Legume Seed Coat Pigmentation
The genetic control of seed coat color involve can the influence of specific genes that have dysfunctional pathways with potential for causing variations. For instance, the stay-green type seed coat color in chickpea is caused by the loss of function in the 'CaStGR1' gene which affects chlorophyll degradation and retention [28]. In soybean, however, seed pigmentation is tied to the I locus, which affects the expression of chalcone synthase gene and subsequently flavonoid biosynthesis [29]. Similar homologous genes in other legumes such as the P locus in common bean, Mendel’s A locus in pea, B locus in chickpea, and Tan gene in lentil, play major roles in anthocyanin and flavonol biosynthesis [30,31,32,33]. Faba bean presents an intriguing case with two identified homologous genes [34,35].
Recently, studies of the genetics of seed coat color in common bean using phenotypic segregation of a 'TU' × 'Musica' cross-derived recombinant inbred line population indicated three genes that controlled seed coat color: one for white (mapped on Pv07) and two for black (mapped on Pv06 and Pv08) [36]. The mapped positions were consistent with classical studies on the V gene and C locus. Crossing three selected lines validated these regions, confirming that TU alleles resulted in a black phenotype. Furthermore, two genes involved in anthocyanin biosynthetic pathway were identified as Phvul.006G018800 encoding a flavonoid 3′5’hydroxylase and Phvul.008G038400 encoding MYB113 transcription factor [36].
Campa et al [37] also examined 308 common bean lines for 10 seed coat color traits, and discovered 31 significant SNP-trait associations (QTNs) that were distributed across 20 chromosome regions. Of these, chromosome Pv08 played a central role, as it influenced phenolic metabolites on chromosomes Pv01, Pv02, Pv04, Pv08, and Pv09 and seed coat color on chromosomes Pv01, Pv02, Pv06, Pv07, and Pv10. Chromosome Pv08 therefore plays a key role in the phenylpropanoid pathway, with significant effect on common bean seed coat pigmentation.
The genetic basis of red seed coat color in cowpea was found to involve two loci, Red-1 (R-1) on chromosome Vu03 and Red-2 (R-2) on chromosome Vu07 [38]. The candidate gene for R-1 was Vigun03g118700, encoding dihydroflavonol 4-reductase which is a cyanidin biosynthesis catalyst, while the candidate gene for R-2 was Vigun07g118500, with a nucleolar function and elevated expression in developing seeds. The red color was attributed to the accumulation of cyanidin in the seed coat [38].
5. Genetic Determinants of Host Plants in Symbiotic Interactions with Rhizobia
The genetic makeup of host plants can play a pivotal role in shaping the outcomes of symbiotic interactions. From signaling pathways to nutrient exchange, the genetic determinants of host plant can contribute significantly to the success of the symbiosis. As a heritable trait seed coat pigmentation can contribute to determining host plants in symbiotic partnership. As found with common bean, cowpea also exhibits different seed coat colors that can significantly affect the N2-fixing ability of genotypes [39,40,41]. The varying seed coat pigmentation of cowpea was due to genetic differences [11,23,38], with seed coat color being a major determinant of the differences in N2 fixation under field conditions. Many strategies have been used to select and breed for enhanced N2 fixation in legumes [42]. Despite the known importance of seed coat color in legume nodulation and N2 fixation, cowpea breeding in West Africa has generally favored genotypes with cream (or white) seed coat color [43]. Recently, Mohammed et al [44] reported a large variation in plant growth, grain yield and symbiotic functioning between and among genetically diverse cowpea accessions with different seed coat colors, which was attributed to the ability of different seed coat colors to attract different native soil rhizobia.
In another study involving Kersting's groundnut (Macrotyloma geocarpum Harms), which is a neglected, underutilized, and endangered food and medicinal legume in Africa, analysis of SSR sequences revealed significant genetic diversity among the Kersting's groundnut landraces with diverse seed coat colors [45]. Due to the unavailability of genetic information specific to Kersting's groundnut, Mohammed et al [45] employed SSR markers derived from cowpea. The successful cross-genus transferability of SSRs in that study did not only indicate a practical approach for future studies, but also suggested an evolutionary closeness between cowpea and Kersting's groundnut.
According to Puozaa et al. [6], seed coat pigmentation is responsible for the observed differences in symbiotic functioning among legumes with different seed coat colors. Furthermore, it was found that three landraces with many similar traits except for seed coat color performed differently. Despite having similar growth habits, including phenology, flowering and maturity dates, the seed coat color emerged as the dominant factor influencing gene expression relating to N2 fixation [6]. In fact, studies of seed coat colors (e.g. black, brown and yellow) in soybean found isoflavone malonyl-CoA acyltransferase GmMaT2 to be involved in the nodulation of soybean through modifying the synthesis and secretion of isoflavones by the seed coat [19].
However, the effective cross-nodulation of Kersting's groundnut plants by cowpea-nodulating strain CB756 [6], which was initially isolated from Macrotyloma africanum by Blumenthal and Staples [46] suggested potential synteny between cowpea and Kersting's groundnut. A comparison of transcriptomes of contrasting seed coat colors (black and yellow) and RNA-seq analysis of soybean also discovered the presence of 318 differentially expressed genes involved in ethylene, lipid, brassinosteroid, lignin, and sulfur amino acid biosynthesis in cultivars with black seed coat pigmentation.
6. Seed Coat Pigmentation and N2 Fixation in Legumes
The importance of seed coat pigmentation in the nodulation of legumes has been reported before [6,15,17,21,47,48] (see Figure 1), and apparently the variation in seed coat color may be responsible for the different N2-fixing abilities observed among legume genotypes under field conditions in African soils [6,45,47]. The efficiency of N2 fixation in plants could be intricately linked to seed coat pigmentation. In fact, Puozaa et al. [6] recently reported that seed coat color is a biomarker for increased nodulation, N2 fixation, and plant growth. Studies of focused on Bambara groundnut (Vigna subterranea L. Verdc) landraces in Ghana, using 15N natural abundance and ureide analysis of xylem sap and petiole extracts consistently revealed a correlation between ureide levels and shoot δ15N values, as well as between ureide levels and %N derived from fixation. Where shoot δ15N values were low and %N derived from fixation was high, ureide concentrations in xylem sap and petiole extracts were also high, indicating a robust precision in the estimation of N2 fixation in diverse seedcoat-colored Bambara groundnut landraces.
Those studies by Puozaa et al. [6] found that Black Bambara groundnut landraces consistently exhibited greater nodule number and nodule fresh weight per plant across all sites. Although the Black and Red landraces generally displayed lower shoot δ15N values, derived a higher proportion of their N from fixation, and contributed more symbiotic N, the Black landrace surpassed the Red. This superior symbiotic performance of the Black Bambara groundnut landrace was attributed to its ability to produce and release higher concentrations of nod-gene-inducing anthocyanins and flavonoids when compared to the Red and Cream landraces [49,50,51]. An earlier study by Hungria and Phillips [17] had also demonstrated that a black-seeded common bean genotype that released higher concentrations of nod-gene-inducing flavonoids similarly elicited greater nodulation than its isogenic cream counterpart with reduced flavonoid biosynthesis.
Furthermore, Mbah and Dakora [52] also found that root nodulation, N2 fixation and shoot micronutrient accumulation differed significantly among different grain legumes (Bambara groundnut, soybean, and Kersting’s groundnut) with diverse seed coat colors in South Africa. However, supplying 5 mM NO3− to inoculated seedlings of Bambara groundnut, soybean, and Kersting’s bean led to reduced plant growth, nodule formation, and nodule dry matter across all seed coat colors. Furthermore, seedlings of Bambara groundnut with diverse seed coat colors revealed varied nodulation and plant growth under different field conditions in African soils in Cameroon, Burkina Faso and Nigeria [53,54,55].
7. Effect of Seed Coat Pigmentation on Microbial Colonization
Besides N2-fixing efficiency, seed coat pigmentation can also influence root colonization by both rhizobia and plant growth-promoting microbes [56,57] (Figure 1). Puozaa et al [58,59] found that PCR-RFLP analysis of bacterial DNA from root nodules of cowpea and Bambara groundnut landraces (namely, Black, Red, Cream, Red mottled, Black mottled and Blackeye) that were grown in Ghanaian and South African soils revealed differences in nodule occupancy based on seed coat color. In other words, seeds differing in seed coat color attracted different rhizobial strains. Even when planted together in one hole, the nodulating bradyrhizobia clustered differently in the PCR-RFLP dandrograms [58]. This suggested that the different Bambara groundnut seed coat colors (e.g. Black, Black mottled, and Cream) probably contributed to a soil environment that was conducive to rhizobial richness. In fact, Mohammed et al [57](2019), Dlamini et al. [60], and Puozaa et al. [6] found that Kersting’s groundnut, Bambara groundnut and cowpea with differing seed coat colors were selectively nodulated by different rhizobial species.
This was further confirmed by multilocus sequence analysis performed on rhizobia nodulaing Kersting’s groundnut. The results revealed different Bradyrhizobium species responsible for nodulating Kersting's groundnut with differing seed coat colors [57]. Phylogenetic analysis also placed the test rhizobial isolates in close proximity with different Bradyrhizobium species such as B. vignae 7-2T, B. subterraneum 58 2-1T, B. kavangense 14-3T, B. liaoningense 2281 (USDA 3622)T, B. yuanmingense LMG 21827T, B. huanghuaihaiense CCBAU 23303T, B. pachyrhizi PAC48T, and the type strain of B. elkanii based mostly according to seed coat color [57]. However, in some instances, a significant divergence was found between test isolates and the reference type strains, suggesting those isolates could be novel Bradyrhizobium species [57].
The higher genetic diversity observed among the nodule occupants of the dark-seeded Kersting's groundnut landraces in this study could be linked directly or indirectly to the greater phenolic compounds produced by the Black landrace compared to the white-seeded Boli landrace [57,61]. Earlier studies have, however, reported higher concentrations of phenolic compounds exuded by seeds with dark seed coat pigmentation when compared to those with lighter seed coat color [49,50], and these molecules are known to play a role in signal exchange during the legume-rhizobia symbiosis.
The effect of different seedcoat colors on microbial community structure in the rhizosphere of five Kersting’s groundnut landraces (namely, Belane Mottled, Boli, Funsi, Puffeun, and Heng Red Mottled) was investigated by Jaiswal et al. [56] using 16S rDNA amplicon sequencing. The results revealed significant selective influences of the landraces on rhizosphere bacteria. The microbial composition and abundance differed significantly among the landraces, with a major landrace effect on some phyla. For example, the rhizosphere of landrace Belane Mottled was dominated by Proteobacteria, while Bacteroidetes dominated the rhizospheres of the other landraces. Furthermore, except for Puffeun (with Black seed coat), whose rhizosphere was dominated by Mesorhizobium, only Bradyrhizobium and Rhizobium species of alpha-Proteobacteria were present in the rhizosphere of all the other landraces, though indole-3-acetic–acid-producing Sphingomonas and cellulose-degrading Fibrobacteres were also abundant in the rhizosphere of all landraces [56].
8. Evolutionary Dynamics of the Legume-Rhizobia Partnership
Symbiotic relationships between plants and microbes are THE outcome of intricate co-evolutionary processes. Most grain legumes, including cowpea, soybean, groundnut, and Bambara groundnut, meet a significant proportion of their N requirements from atmospheric N2 fixation. Like all grain legumes, seed coat color variation is strong among Bambara groundnut landraces, and can range from black to mottled to lighter cream color. This diversity is primarily attributed to different types of flavonoid compounds and their concentrations present in the seed coat [62]. Seed coat colors are also reported to correlate with distinct N2-fixing abilities under different agro-ecologies, thus underscoring the role of seed coat compounds in selecting microsymbiont partners [58]. Independent of the cropping system, seed coat pigmentation is reported to significantly influence the diversity of microsymbionts residing in Bambara groundnut root nodules [58]. Even when co-planted in the same hole, landraces with different seed coat colors formed nodules with bacterial partners that exhibited divergent clustering at the 16S-23S rDNA-RFLP level. This suggests a link between seed coat pigmentation and the specificity of the symbiosis associated with each landrace [58].
The flavonoid profile of leguminous seeds is often dictated by seed coat color, which strongly contributes to the selection of rhizobial partners in symbiotic relationships. For example, Bambara groundnut landraces with distinct seed coat pigmentation were found to contain varying levels of flavonoids and anthocyanins in their seed exudates [6,62]. These phenolic compounds are abundantly present in the seed coat, and often act as chemo-attractants for rhizobia, bacterial cell growth promoters, and inducers of nod-genes in symbiotic legumes [16,17,25,63].
The presence of a specific legume in the soil can trigger the selection of specific rhizobial groups due to rhizodeposition, consisting mainly of flavonoids released by the host plant roots. During germination, however, legume seeds release these flavonoid compounds abundantly from the seed coat [25,62,64,65].
Kersting's groundnut, is a neglected, underutilized and endangered food and medicinal grain legume native to Africa, and whose symbiosis has remained unexplored [45,56]. The multi-seedcoat colors found in Kersting's groundnut landraces are believed to be genetically influenced rather than environmentally-induced, given the observed differences in nucleotide sequences of simple sequence repeats in their genomic regions [45].
Symbiotic data from Kersting's groundnut landraces revealed marked variations in root nodulation, shoot biomass, δ15N, percent N derived from fixation, amount of N-fixed, and soil N uptake [45].
Furthermore, the Black landrace consistently outperformed others in terms of nodule number, nodule fresh weight, and N contribution from symbiosis. Phylogenetic analysis showed that Kersting's groundnut is nodulated mainly by Bradyrhizobium species as they were closely related to known strains and novel groups within the genus [57]. Because of the diverse seed coat colors of Kersting’s groundnut, the species tends to attract different microbial profiles in its rhizosphere.
9. Adapting to Environmental Changes: The Role of Seed Coat Pigmentation
Environmental changes can pose challenges to symbiotic associations that necessitate adaptive responses. Seed coat pigmentation plays a key role in the adaptive strategies employed by plants in response to changing environmental conditions. According to Puozaa et al. [59], the cultivation of cowpea with differing seed coat colors in the low rainfall site of Morwe in South Africa contributed to a higher diversity of cowpea-nodulating rhizobia. Furthermore, when cowpea seeds with Black, Red, and Cream pigmentation were planted together in one hole, they attracted diverse microsymbionts, which exhibited varied PCR-ITS-RFLP profiles and occupied distinct positions in the phylogenetic tree.
Puozaa et al. [6] also reported that the Black Bambara groundnut landrace exhibited greater plant growth (measured as shoot dry matter) at all study sites in Ghana and South Africa compared to the Red and Cream landraces. The variation in plant growth among landraces with different seed coat colors could be linked to differences in solar radiation interception efficiency, genetic variations in photosynthate production, and/or greater resistance to environmental stresses which affect crop productivity. Seedling vigor which is, crucial for early crop establishment, may also be influenced by seedcoat color, as evidenced by the differences in seedling vigor among Bambara groundnut landraces with varying seed coat pigmentation [66].
In other studies, site-specific comparisons showed that the Red landrace consistently produced higher grain yield than the Black and Cream landraces to the extent that in Cameroon and Tanzania, Red seedcoat pigmentation is associated with increased grain yield of Bambara groundnut landraces [67,68].
Although Bambara groundnut is generally considered to be drought-tolerant, little evidence current exists to support that argument, except for the report by Puozaa et al. [6] which found significant differences in shoot δ13C or water-use efficiency among the Bambara groundnut landraces tested across different study sites, with the Black landrace exhibiting higher δ13C values, and hence greater water-use efficiency than the Red and Cream landraces. The relative drought tolerance of the Black landrace can be attributed to the higher tissue concentration of flavonoids, anthocyanins and anthocyanidins, which are known to protect the photosynthetic apparatus under drought conditions [69]. Furthermore, a significant correlation was found between shoot δ13C and shoot N content, as well as between shoot δ13C and soil N uptake [6], suggesting that the enhanced N nutrition from fixation by the Black landrace contributed to the greater water-use efficiency and drought resistance relative to the Red and Cream landraces.
10. Grain legume Seed Coats Are a Natural Source of Nutraceuticals and Anthocyanins
Flavonoids secreted by legume seeds play a major role as signal molecules for attracting compatible rhizobia during symbiotic establishment. As the reproductive unit, seeds carry the genetic material of crop species over time and space, and not only does the seed coat play a vital role in protecting the embryo, and/or determining seed dormancy and germination [70,71,72], it also contains a host of novel compounds which are released following imbibition of water during germination. These molecules include flavonoids, proteins, peptides, amino acids, alkaloids and terpenoids [62,71]. Flavonoids are phenolic secondary metabolites that confer color on seeds, flowers and fruits [38]. They play many important roles in plant development, including defense against insect pests and diseases, as well as nodulation in symbiotic legumes (Table 1; Figure 1) [62,73,74,75]. Puozaa et al. [6] also found marked variations in the concentration of phenolic compounds in the seed exudate of Bambara groundnut with differing seed coat colors. In fact, seeds of the Black landrace released higher levels of flavonoids and anthocyanins, followed by the Red, with Cream being the lowest. The Black landrace also elicited significantly higher nodule number and nodule fresh weight per plant, greater shoot biomass, lower shoot δ15N, greater N derived from fixation, higher symbiotic N contribution, and increased water-use efficiency across all study sites. These findings underscore the importance of seed coat color as a biomarker for breeding Bambara groundnut varieties with enhanced nodulation, N2 fixation, and grain yield. The results of UPLC-qTOF-MS analysis by Tsamo et al. [49] confirmed earlier findings on the presence of elevated levels of phenolic compounds in darker-colored Bambara groundnut seeds, black-seeded azuki bean, and soybean. Ndakidemi and Dakora [62] also reported higher concentrations of anthocyanins and flavonoids in a black-seeded Bambara groundnut accession when compared to its Cream counterpart. Seed coat pigmentation therefore plays a crucial role in determining the profile and concentrations of flavonoids, anthocyanins, and anthocyanidins in seeds of cowpea, Bambara groundnut, and Kersting's groundnut [49,50,51]. Metabolite profiling by Tsamo et al. [51] showed that black seeds of legumes are a natural source of nutraceuticals for human nutrition/health, and a reservoir of anthocyanins that can be exploited for developing cosmetic, food, and pharmaceutical products.
However, consumer preferences for cowpea and Bambara groundnut consumption in West Africa, where most of the cowpea is produced in Africa, are influenced by seed coat pigmentation. Most consumers in West Africa prefer cowpea and Bambara groundnut with cream or white seed coat color [76]. This contrasts with Brazil, where "black beans" are highly preferred, possibly due to their high levels of total phenolics, flavonoids, anthocyanins, anthocyanidins, and antioxidant activity when compared to their counterparts with cream or white seed coat color [20,49,50,51] (Table 1). The consumption of these antioxidant compounds has implications for human nutrition and health [77,78].
11. Conclusions
Rhizobia play a pivotal role in agroecosystems, contributing significantly to enhancing overall soil health through their symbiotic relationship with legumes. Numerous reports have indicated that legumes with varied seedcoat colors exhibit diverse responses to N2 fixation and sustainable agriculture in African soils. Understanding the molecular and genetic aspects of seed coat pigmentation is crucial for unraveling symbiotic interactions. Evolutionary dynamics, illustrated by diverse seed coat colors in legumes like Bambara groundnut and Kersting's groundnut, showcase the intricate co-evolutionary processes influencing N2-fixing partnerships. The connection between seed coat pigmentation, flavonoid profiles, and rhizobial selection suggests potential applications in optimizing microsymbiont associations. Further research on neglected legumes, such as Kersting's groundnut, and the impact of seed coat colors on soybean seed quality opens avenues for improved crop productivity and sustainability.
Author Contributions
SJ search literatures and drafted the manuscript. FD edited, and approved the final version of the paper. All authors contributed to the article and approved the submitted version.
Funding
FD was grateful to the NRF, Tshwane University of Technology (TUT) and the South African Research Chair in Agrochemurgy and Plant Symbiosis for continued funding of his research.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Molefe, R.R.; Amoo, A.E.; Babalola, O.O. Communication between plant roots and the soil microbiome; involvement in plant growth and development. Symbiosis 2023, 90, 231–239. [Google Scholar] [CrossRef]
- Barbieri, P.; Starck, T.; Voisin, A.-S.; Nesme, T. Biological nitrogen fixation of legumes crops under organic farming as driven by cropping management: A review. Agric. Syst. 2023, 205, 103579. [Google Scholar] [CrossRef]
- Dimkpa, C.; Adzawla, W.; Pandey, R.; Atakora, W.K.; Kouame, A.K.; Jemo, M.; Bindraban, P.S. Fertilizers for food and nutrition security in sub-Saharan Africa: an overview of soil health implications. Front. Soil Sci. 2023, 3. [Google Scholar] [CrossRef]
- Nag, P.; Shriti, S.; Das, S. Microbiological strategies for enhancing biological nitrogen fixation in nonlegumes. J. Appl. Microbiol. 2020, 129, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Ngwenya, Z.D.; Mohammed, M.; Dakora, F.D. Monocropping and Intercropping of Maize with Six Food Legumes at Malkerns in Eswatini: Their Effects on Plant Growth, Grain Yield and N2 Fixation, Measured using the 15N Natural Abundance and Ureide Techniques. Symbiosis 2024, 1–13. [Google Scholar] [CrossRef]
- Puozaa, D.K.; Jaiswal, S.K.; Dakora, F.D. Black seedcoat pigmentation is a marker for enhanced nodulation and N2 fixation in Bambara groundnut (Vigna subterranea L. Verdc.) landraces. Front. Agron. 2021, 3, 692238. [Google Scholar] [CrossRef]
- Bopape, F.L.; Beukes, C.W.; Katlego, K.; Hassen, A.I.; Steenkamp, E.T.; Gwata, E.T. Symbiotic performance and characterization of Pigeonpea (Cajanus cajan L. Millsp.) Rhizobia occurring in South African soils. Agriculture 2022, 13, 30. [Google Scholar] [CrossRef]
- Nyemba, R.C.; Dakora, F.D. Evaluating N 2 fixation by food grain legumes in farmers ’ fields in three agro-ecological zones of Zambia, using 15 N natural abundance. Biol Fertil Soils 2010, 461–470. [Google Scholar] [CrossRef]
- Bohra, A.; Tiwari, A.; Kaur, P.; Ganie, S.A.; Raza, A.; Roorkiwal, M.; Mir, R.R.; Fernie, A.R.; Sm\`ykal, P.; Varshney, R.K. The key to the future lies in the past: insights from grain legume domestication and improvement should inform future breeding strategies. Plant Cell Physiol. 2022, 63, 1554–1572. [Google Scholar] [CrossRef] [PubMed]
- Konzen, E.R.; Tsai, S.M. Genetic variation of landraces of common bean varying for seed coat glossiness and disease resistance: valuable resources for conservation and breeding. Rediscovery Landrac. as a Resour. Futur. London IntechOpen 2018, 177–193. [Google Scholar]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar] [CrossRef] [PubMed]
- Paauw, M.; Koes, R.; Quattrocchio, F.M. Alteration of flavonoid pigmentation patterns during domestication of food crops. J. Exp. Bot. 2019, 70, 3719–3735. [Google Scholar] [CrossRef] [PubMed]
- Von Wettberg, E.J.B.; Chang, P.L.; Ba\csdemir, F.; Carrasquila-Garcia, N.; Korbu, L.B.; Moenga, S.M.; Bedada, G.; Greenlon, A.; Moriuchi, K.S.; Singh, V.; et al. Ecology and genomics of an important crop wild relative as a prelude to agricultural innovation. Nat. Commun. 2018, 9, 649. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.-S.; Contador, C.A.; Ng, M.-S.; Yu, J.; Chung, G.; Lam, H.-M. The effects of domestication on secondary metabolite composition in legumes. Front. Genet. 2020, 11, 581357. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, U. a; Maxwell, C. a; Joseph, C.M.; Phillips, D. a Chrysoeriol and Luteolin Released from Alfalfa Seeds Induce nod Genes in Rhizobium meliloti. Plant Physiol. 1990, 92, 116–22. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, U. a; Joseph, C.M.; Phillips, D. a Flavonoids Released Naturally from Alfalfa Seeds Enhance Growth Rate of Rhizobium meliloti. Plant Physiol. 1991, 95, 797–803. [Google Scholar] [CrossRef]
- Hungria, M.; Phillips, D. Effects of a seed colour mutation on rhizobial nod-gene-inducing flavonoids and nodulation in common bean. Mol. Plant-Microbe Interact. 1993, 6, 418–422. [Google Scholar] [CrossRef]
- Cooper, J.E. Early interactions between legumes and rhizobia: Disclosing complexity in a molecular dialogue. J. Appl. Microbiol. 2007, 103, 1355–1365. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Zhang, Y.; Zeng, X.; Li, P.; Wang, X.; Benedito, V.A.; Zhao, J. Isoflavone malonyl-CoA acyltransferase GmMaT2 is involved in nodulation of soybean by modifying synthesis and secretion of isoflavones. J. Exp. Bot. 2021, 72, 1349–1369. [Google Scholar] [CrossRef] [PubMed]
- Hungria, M.; Joseph, C.M.; Phillips, D.A. Anthocyanidins and Flavonols, Major nod Gene Inducers from Seeds of a Black-Seeded Common Bean. 1991, 751–758. [Google Scholar]
- Subramanian, S.; Stacey, G.; Yu, O. Distinct, crucial roles of flavonoids during legume nodulation. Trends Plant Sci. 2007, 12, 282–5. [Google Scholar] [CrossRef] [PubMed]
- Mishra, G.P.; Ankita Aski, M.S.; Tontang, M.T.; Choudhary, P.; Tripathi, K.; Singh, A.; Kumar, R.R.; Thimmegowda, V.; Stobdan, T.; et al. Morphological, molecular, and biochemical characterization of a unique lentil (Lens culinaris medik.) genotype showing seed-coat color anomalies due to altered anthocyanin pathway. Plants 2022, 11, 1815. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Gillaspie, A.G.; Morris, J.B.; Pittman, R.N.; Davis, J.; Pederson, G.A. Flavonoid content in different legume germplasm seeds quantified by HPLC. Plant Genet. Resour. 2008, 6, 62–69. [Google Scholar] [CrossRef]
- dos Santos Sousa, W.; Teixeira, I.R.; Souza Campos, T.; da Silva, G.C.; da Silva, M.; Guimarães Moreira, S. Supplementary reinoculation in topdressing of Rhizobium tropici in common bean crop: effects on nodulation, morphology, and grain yield. J. Plant Nutr. 2022, 45, 3094–3108. [Google Scholar] [CrossRef]
- Shumilina, J.; Soboleva, A.; Abakumov, E.; Shtark, O.Y.; Zhukov, V.A.; Frolov, A. Signaling in Legume--Rhizobia Symbiosis. Int. J. Mol. Sci. 2023, 24, 17397. [Google Scholar] [CrossRef] [PubMed]
- Compton, K.K.; Scharf, B.E. Rhizobial chemoattractants, the taste and preferences of legume symbionts. Front. Plant Sci. 2021, 12, 686465. [Google Scholar] [CrossRef] [PubMed]
- Samac, D.A.; Graham, M.A. Recent advances in legume-microbe interactions: recognition, defense response, and symbiosis from a genomic perspective. Plant Physiol. 2007, 144, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Sivasakthi, K.; Marques, E.; Kalungwana, N.; Carrasquilla-Garcia, N.; Chang, P.L.; Bergmann, E.M.; Bueno, E.; Cordeiro, M.; Sani, S.G.A.S.; Udupa, S.M.; et al. Functional dissection of the chickpea (Cicer arietinum L.) stay-green phenotype associated with molecular variation at an ortholog of Mendel’s I gene for cotyledon color: implications for crop production and carotenoid biofortification. Int. J. Mol. Sci. 2019, 20, 5562. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, J.H.; Zabala, G.; Varala, K.; Hudson, M.; Vodkin, L.O. Endogenous, tissue-specific short interfering RNAs silence the chalcone synthase gene family in Glycine max seed coats. Plant Cell 2009, 21, 3063–3077. [Google Scholar] [CrossRef] [PubMed]
- McClean, P.E.; Bett, K.E.; Stonehouse, R.; Lee, R.; Pflieger, S.; Moghaddam, S.M.; Geffroy, V.; Miklas, P.; Mamidi, S. White seed color in common bean (Phaseolus vulgaris) results from convergent evolution in the P (pigment) gene. New Phytol. 2018, 219, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Moreau, C.; Lin-Wang, K.; Schwinn, K.E.; Thomson, S.J.; Fiers, M.W.E.J.; Frew, T.J.; Murray, S.R.; Hofer, J.M.I.; Jacobs, J.M.E.; et al. Identification of Mendel’s white flower character. PLoS One 2010, 5, e13230. [Google Scholar] [CrossRef] [PubMed]
- Varma Penmetsa, R.; Carrasquilla-Garcia, N.; Bergmann, E.M.; Vance, L.; Castro, B.; Kassa, M.T.; Sarma, B.K.; Datta, S.; Farmer, A.D.; Baek, J.-M.; et al. Multiple post-domestication origins of kabuli chickpea through allelic variation in a diversification-associated transcription factor. New Phytol. 2016, 211, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Mirali, M.; Purves, R.W.; Stonehouse, R.; Song, R.; Bett, K.; Vandenberg, A. Genetics and biochemistry of zero-tannin lentils. PLoS One 2016, 11, e0164624. [Google Scholar] [CrossRef]
- Gutierrez, N.; Torres, A.M. Characterization and diagnostic marker for TTG1 regulating tannin and anthocyanin biosynthesis in faba bean. Sci. Rep. 2019, 9, 16174. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, N.; Avila, C.M.; Torres, A.M. The bHLH transcription factor VfTT8 underlies zt2, the locus determining zero tannin content in faba bean (Vicia faba L.). Sci. Rep. 2020, 10, 14299. [Google Scholar] [CrossRef] [PubMed]
- Garc\’\ia-Fernández, C.; Campa, A.; Ferreira, J.J. Dissecting the genetic control of seed coat color in a RIL population of common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2021, 134, 3687–3698. [Google Scholar] [CrossRef] [PubMed]
- Campa, A.; Rodr\’\iguez Madrera, R.; Jurado, M.; Garc\’\ia-Fernández, C.; Suárez Valles, B.; Ferreira, J.J. Genome-wide association study for the extractable phenolic profile and coat color of common bean seeds (Phaseolus vulgaris L.). BMC Plant Biol. 2023, 23, 158. [Google Scholar] [CrossRef] [PubMed]
- Herniter, I.A.; Munoz-Amatriain, M.; Lo, S.; Guo, Y.-N.; Lonardi, S.; Close, T.J. Identification of candidate genes controlling red seed coat color in cowpea (Vigna unguiculata [L.] Walp). Horticulturae 2024, 10, 161. [Google Scholar] [CrossRef]
- Belane, A.K.; Dakora, F.D. Measurement of N2 fixation in 30 cowpea (Vigna unguiculata L. Walp.) genotypes under field conditions in Ghana, using the15N natural abundance technique. Symbiosis 2009, 48, 47–56. [Google Scholar] [CrossRef]
- Belane, A.K.; Dakora, F.D. Symbiotic N2 fixation in 30 field-grown cowpea (Vigna unguiculata L. Walp.) genotypes in the Upper West Region of Ghana measured using 15 N natural abundance. Biol. Fertil. Soils 2010, 46, 191–198. [Google Scholar] [CrossRef]
- Pule-Meulenberg, F.; Belane, A.K.; Krasova-Wade, T.; Dakora, F.D. Symbiotic functioning and bradyrhizobial biodiversity of cowpea (Vigna unguiculata L. Walp.) in Africa. BMC Microbiol. 2010, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Herridge, D.; Rose, I. Breeding for enhanced nitrogen ® xation in crop legumes. 2000, 65. [Google Scholar]
- Egbadzor, K.F.; Yeboah, M.; Gamedoagbao, D.K.; Offei, S.K.; Danquah, E.Y.; Ofori, K. Inheritance of seed coat colour in cowpea (Vigna unguiculata (L.) Walp). 2014. [Google Scholar] [CrossRef]
- Mohammed, H.; Jaiswal, S.K.; Mohammed, M.; Mbah, G.C.; Dakora, F.D. Insights into nitrogen fixing traits and population structure analyses in cowpea (Vigna unguiculata L. Walp) accessions grown in Ghana. Physiol. Mol. Biol. Plants 2020, 26, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.; Jaiswal, S.K.; Sowley, E.N.K.; Ahiabor, B.D.K. Symbiotic N2 fixation and grain yield of endangered Kersting’s groundnut landraces in response to soil and plant associated Bradyrhizobium inoculation to promote ecological resource-use efficiency. Front. Microbiol. 2018, 9, 2105. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, M.J.; Staples, L.B. Origin, evaluation and use of Macrotyloma as forage. Trop Grassl 1993, 27, 16–29. [Google Scholar]
- Hassen, A.I.; Bopape, F.L.; van Vuuren, A.; Gerrano, A.S.; Morey, L. Symbiotic interaction of bambara groundnut (Vigna subterranea) landraces with rhizobia spp. from other legume hosts reveals promiscuous nodulation. South African J. Bot. 2023, 160, 493–503. [Google Scholar] [CrossRef]
- Wilker, J.; Navabi, A.; Rajcan, I.; Marsolais, F.; Hill, B.; Torkamaneh, D.; Pauls, K.P. Agronomic performance and nitrogen fixation of heirloom and conventional dry bean varieties under low-nitrogen field conditions. Front. Plant Sci. 2019, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Tsamo, A.T.; Ndibewu, P.P.; Dakora, F.D. Phytochemical profile of seeds from 21 Bambara groundnut landraces via UPLC-qTOF-MS. Food Res. Int. 2018, 112, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Tsamo, A.T.; Mohammed, M.; Ndibewu, P.P.; Dakora, F.D. Identification and quantification of anthocyanins in seeds of Kersting�s groundnut [Macrotyloma geocarpum (Harms) Marechal & Baudet] landraces of varying seed coat pigmentation. J. Food Meas. Charact.
- Tsamo, A.T.; Mohammed, H.; Mohammed, M.; Papoh Ndibewu, P.; Dapare Dakora, F. Seed coat metabolite profiling of cowpea (Vigna unguiculata L. Walp.) accessions from Ghana using UPLC-PDA-QTOF-MS and chemometrics. Nat. Prod. Res. 2020, 34, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Mbah, G.C.; Dakora, F.D. Nitrate inhibition of N 2 fixation and its effect on micronutrient accumulation in shoots of soybean (Glycine max L. Merr.), Bambara groundnut (Vigna subterranea L. Vedc) and Kersting’s groundnut (Macrotyloma geocarpum Harms.). Symbiosis 2017, 1–12. [Google Scholar] [CrossRef]
- Zenabou, N.; Firmin, S.L.; Laurette, N.N.; Daniel, W.F.; Laurianne, T.N.; Martin, B.J. Nodulation Potential of Bambara Groundnut (Vigna subterranea L.) in Yaounde (Centre Region of Cameroon). Am. J. Food Nutr. 2022, 10, 34–39. [Google Scholar]
- Ouedraogo, M.; Kima, A.S.; Konaté, M.N.G.; Ouedraogo, H.M.; Ouoba, A.; Nikiema, B.; Kambou, D.J.; Zongo, K.F.; Nandkangre, H. Performance of bambara groundnut (Vigna subterranea [L.] Verdcourt) genotypes cropped on plinthite soil in the semi arid-zone, Burkina Faso. Int. J. Plant \& Soil Sci. 2022, 34, 1067–1075. [Google Scholar]
- Bitire, T.D.; Abberton, M.; Tella, E.O.; Edemodu, A.; Oyatomi, O.; Babalola, O.O. Impact of nitrogen-fixation bacteria on nitrogen-fixation efficiency of Bambara groundnut [Vigna subterranea (L) Verdc] genotypes. Front. Microbiol. 2023, 14, 1187250. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.K.; Mohammed, M.; Dakora, F.D. Microbial community structure in the rhizosphere of the orphan legume Kersting’s groundnut [Macrotyloma geocarpum (Harms) Marechal & Baudet]. Mol. Biol. Rep. 2019, 46. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.; Jaiswal, S.K.; Dakora, F.D. Insights into the phylogeny, nodule function, and biogeographic distribution of microsymbionts nodulating the orphan Kersting’s groundnut [Macrotyloma geocarpum (Harms) Marechal & Baudet] in African soils. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Puozaa, D.K.; Jaiswal, S.K.; Dakora, F.D. African origin of Bradyrhizobium populations nodulating Bambara groundnut (Vigna subterranea L. Verdc) in Ghanaian and South African soils. PLoS One 2017, 12, e0184943. [Google Scholar] [CrossRef] [PubMed]
- Puozaa, D.K.; Jaiswal, S.K.; Dakora, F.D. Phylogeny and distribution of Bradyrhizobium symbionts nodulating cowpea (Vigna unguiculata L. Walp)and their association with the physicochemical properties of acidic African soils. Syst. Appl. Microbiol. 2019, 42. [Google Scholar] [CrossRef] [PubMed]
- Dlamini, S.T.; Jaiswal, S.K.; Mohammed, M.; Dakora, F.D. Studies of Phylogeny, Symbiotic Functioning and Ecological Traits of Indigenous Microsymbionts Nodulating Bambara Groundnut (Vigna subterranea L. Verdc) in Eswatini. Microb. Ecol. 2021, 1–16. [Google Scholar] [CrossRef]
- Makoi, J.; Ndakidemi, P. Biological, ecological and agronomic significance of plant phenolic compounds in rhizosphere of the symbiotic legumes. African J. Biotechnol. 2007, 6. [Google Scholar]
- Ndakidemi, P.A.; Dakora, F.D. Review : Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Funct. Plant Bilogy 2003, 30, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.M.M.; Ashby, A.M.; Richards, A.J.M.; Loake, G.J.; Watson, M.D.; Shaw, C.H. Chemotaxis of Rhizobium leguminosarum biovar phaseoli towards Flavonoid Inducers of the Symbiotic Nodulation Genes. Microbiology 1988, 134, 2741–2746. [Google Scholar] [CrossRef]
- Begum, A.A. Specific flavonoids induced nod gene expression and pre-activated nod genes of Rhizobium leguminosarum increased pea (Pisum sativum L.) and lentil (Lens culinaris L.) nodulation in controlled growth chamber environments. J. Exp. Bot. 2001, 52, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Mathesius, U. The role of flavonoids in root-rhizosphere signalling: opportunities and challenges for improving plant-microbe interactions. J. Exp. Bot. 2012, 63, 3429–44. [Google Scholar] [CrossRef] [PubMed]
- Mabhaudhi, T.; Modi, A. Growth, phenological and yield responses of a bambara groundnut (Vigna subterranea L. Verdc) landrace to imposed water stress: II. Rain shelter conditions. Water SA 2013, 39, 191–198. [Google Scholar] [CrossRef]
- COLLINSON, S.T.; SIBUGA, K.P.; TARIMO, A.J.P.; AZAM-ALI, S.N. INFLUENCE OF SOWING DATE ON THE GROWTH AND YIELD OF BAMBARA GROUNDNUT LANDRACES IN TANZANIA. Exp. Agric. 2000, 36, 1–13. [Google Scholar] [CrossRef]
- Wamba, O.; Taffouo, V.; Youmbi, E.; Ngwene, B.; Amougou, A. Effects of organic and inorganic nutrient sources on growth, total chlorophyll and yield of three bambara groundnut landraces in the Coastal Region of Cameroom. J. Agron. 2012, 11, 31–42. [Google Scholar] [CrossRef]
- Nogués, S.; Allen, D.J.; Morison, J.I.L.; Baker, N.R. Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol. 1998, 117, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Del Bel, Z.; Andrade, A.; Lindström, L.; Alvarez, D.; Vigliocco, A.; Alemano, S. The role of the sunflower seed coat and endosperm in the control of seed dormancy and germination: phytohormone profile and their interaction with seed tissues. Plant Growth Regul. 2024, 102, 51–64. [Google Scholar] [CrossRef]
- Quilichini, T.D.; Gao, P.; Yu, B.; Bing, D.; Datla, R.; Fobert, P.; Xiang, D. The seed coat’s impact on crop performance in pea (Pisum sativum L.). Plants 2022, 11, 2056. [Google Scholar] [CrossRef] [PubMed]
- Mo\"\ise, J.A.; Han, S.; Gudynait\ke-Savitch, L.; Johnson, D.A.; Miki, B.L.A. Seed coats: structure, development, composition, and biotechnology. Vitr. Cell. \& Dev. Biol. 2005, 41, 620–644. [Google Scholar]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Mohammed, M.; Ibny, F.Y.I.; Dakora, F.D. Rhizobia as a Source of Plant Growth-Promoting Molecules: Potential Applications and Possible Operational Mechanisms. Front. Sustain. Food Syst. 2021, 4, 1–14. [Google Scholar] [CrossRef]
- Makoi, J.H.J.R.; Belane, A.K.; Chimphango, S.B.M.; Dakora, F.D. Seed flavonoids and anthocyanins as markers of enhanced plant defence in nodulated cowpea (Vigna unguiculata L. Walp.). F. Crop. Res. 2010, 118, 21–27. [Google Scholar] [CrossRef]
- Herniter, I.A.; Jia, Z.; Kusi, F.; others. Market preferences for cowpea (Vigna unguiculata [L.] Walp) dry grain in Ghana. African J. Ag. Res 2019, 14, 928–934. [Google Scholar]
- Jiang, J.; Xiong, Y.L. Natural antioxidants as food and feed additives to promote health benefits and quality of meat products: A review. Meat Sci. 2016, 120, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: an overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Seed coat colors and their roles in sustainable agriculture.
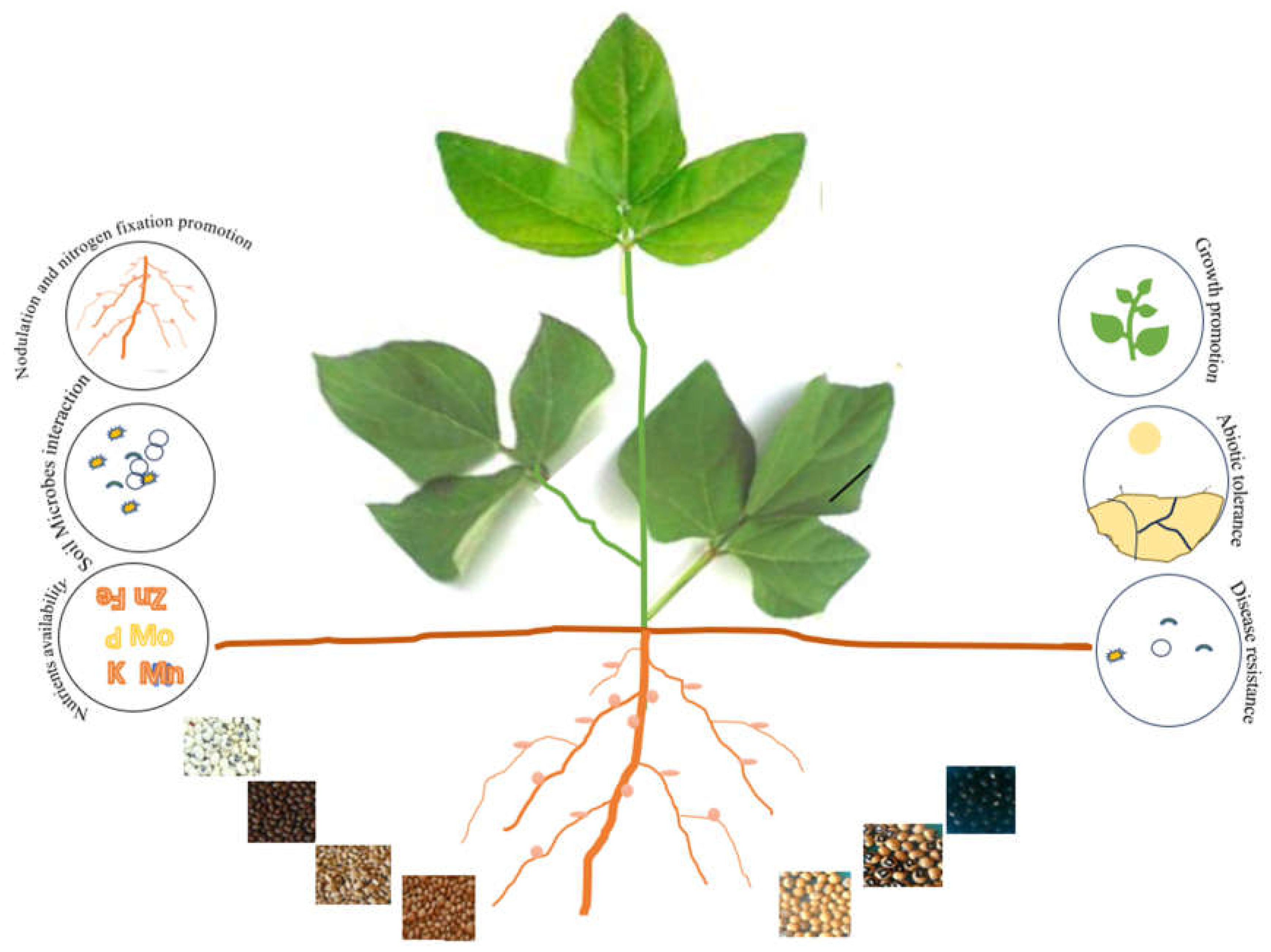

Table 1.
Legume seed coat colors and their impact on plant activities.
| Legume | Seedcoat color | Agronomical effect | References |
|---|---|---|---|
| Bambara Groundnut (Vigna Subterranea L. Verdc. | Black | Enhanced nodulation and nitrogen fixation | Puozaa et al. (2021) |
| Winged bean (Psophocarpus tetragonolobus) | Brown | Enhanced nodulation,and nitrogen fixation | Adegboyega et al. (2021) |
| Common bean (Phaseolus vulgaris L.) | light red | disease resistance and symbiotic nitrogen fixation | Wilker et al. (2020) |
| Soybean (Glycine max (L.) Merr.) | Black and Brown | Enhanced antioxidant activities and anthocyanins | Lim et al. (2021); Jung et al. (2022) |
| Soybean (Glycine max (L.) Merr.) | Yellow | Higher water absorption | Abati et al (2022) |
| Adzuki Bean (Vigna angularis L.) | Black | Higher accumulation of anthocyanins | Chu et al. (2021); Nagao et al. (2023) |
| Lentil (Lens culinaris Medik.) | Black | Higher nutraceutical values | Mishra et al. (2022) |
| Peanut (Arachis hypogaea L.) | Dark red | Higher polyphenol content | Nayak et al. 2020 |
| Kersting’s groundnut (Macrotyloma geocarpum Harms) | Black | Higher nitrogen fixation | Mohammed et al. (2018) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



