
Submitted:
14 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
Thromboembolic conditions, such as stroke and myocardial infarction [MI], are the primary causes of mortality globally. High efficiency was demonstrated by fibrinolytic enzymes isolated from microbial sources in dissolving various blood clots. The study aimed to investigate and produce bacterial fibrinolytic enzymes from various Egyptian soil conditions using recombinant DNA technology. Fifty soil samples were examined to determine the production of fibrinolytic enzymes. The plasmin plate technique assessed the thrombolytic activities of the strong fibrinolytic enzyme-producing bacterial isolates. In addition to determining the molecular mass using a mass spectrometer and the western blot method, the activity and thermostability of the enzymes were characterized. An affinity chromatography approach was used to purify the fibrinolytic enzymes. However, bacterial recombinant DNA technology produced the pure bacterial fibrinolytic enzyme. Several dosage formulations for the formulation of fibrinolytic enzymes were tested. By using affinity chromatography and the SDS-PAGE test, the isolated bacterial fibrinolytic enzyme in this investigation was almost 83% pure. Bacillus cereus ATCC 4342 was identified as the predominant bacterial isolate secreting fibrinolytic enzymes. It was discovered that the fibrinolytic enzyme activity of fifty soil samples ranged from 0.167 U/ml to 1.853 U/ml. The extracellular enzyme extracted from Bacillus cereus culture supernatant demonstrated a relatively stable fibrinolytic activity over 18 hours at a pH range of 7.4–10, as well as stability with maximum productivity at 43℃ for over 60 minutes. In this investigation, the enzyme was referred to as Fibrin-proteinase[ FP]. The fibrin clots were directly thrombolyzed by the FP. The calculated values for the specific activity and molecular mass of the FP were approximately 40.71 units/mg of protein and approximately 30 KDa, respectively. It was observed that the presence of Fe+3 marginally boosted protease activity, but the presence of EDTA decreased it. On the other hand, adding monovalent cations like K+1 and Na+1 significantly increased the fibrinolytic activity of FP. The enzyme exhibited activity throughout the temperature range of 20 to 65℃, peaking at 55℃. The ideal range for the generation of protease was 35–43℃ and 8.0–10. The thermostable fibrinolytic enzyme FP, which was isolated from Bacillus cereus ATCC 4342 in various soil environments in Egypt during the current study, was a promising fibrinolytic enzyme because it demonstrated a more extensive and effective fibrinolytic activity and fewer side effects than the fibrinolytic enzymes that are currently available worldwide. It is advised that further research be done on optimizing the right formulation of bacterial fibrinolytic enzymes in the future to help dissolve different types of blood clots and, as a result, lower the high death rate that exists worldwide.
Keywords:
fibrinolytic
; enzyme
; thrombosis
; screening
; myocardial infarction
Introduction
Thromboembolic illnesses are life-threatening conditions that need the administration of fibrinolytic drugs. Cardiovascular diseases are now the leading cause of mortality worldwide, with low- and middle-income countries suffering the most from them. Thromboembolic diseases are estimated to be the cause of around one in every four deaths globally. [1] Thrombosis, which accounts for 31% of all deaths worldwide, is one of the top causes of mortality, according to the WHO in 2017. [2] Second, the process by which the fibrin-specific protease plasmin breaks down fibrin is known as thrombolysis. [3] Plasminogen, the precursor form of plasmin, is an inactive form that circulates throughout the fibrinolytic system. [4] In the event of tissue injury, plasminogen is converted to plasmin by plasminogen activator. [5] Plasmin dissolves and breaks down fibrin clots. [6] Thrombosis, which is brought on by fibrin accumulation in blood vessels, can lead to myocardial infarction or other cardiovascular issues. [7] Typical drugs such as fibrinolytic proteins, enzymes, anticoagulants, and antiplatelets might result in allergic responses, bleeding problems, and hemorrhaging. [8] Alteplase and Tencteplase are examples of tissue plasminogen activators that are expensive. [9] It became necessary to find new, low-cost fibrinolytic enzymes to overcome the limits of the already existing fibrinolytic drugs. [10] Stroke is the second most prevalent cause of mortality globally, accounting for an estimated 5.5 million fatalities annually. [11] About 85% of stroke patients have an ischemic stroke, which means that fibrinolytic drugs are required. [12] Proactivator plasminogen and the extracellular protein streptokinase, produced by streptococci, combine to form a complex. [13] This combination of enzymes catalyses the conversion of inactive plasminogen into active plasmin. [14] Urokinase is produced by the kidney. [15] It transforms plasminogen into plasmin that is active. [16] Owing to their non-synthesis-ability for use in many oral administration modalities and non-thermostability, current fibrinolytic drugs are unsuitable for storage at room temperature or outside of a refrigerator. [17] The goal of the current study was to find new sources of microbial fibrinolytic enzymes from different soil conditions in Egypt in order to reduce the negative side effects of the fibrinolytic enzymes that are currently available on the global market.
Patients and Methos
Ethical statement:
Prioritized in the current study were all pertinent institutional, national, and/or international standards concerning the use and care of humans and animals. All study techniques involving humans and animals were approved by the Cairo University Ethics Committee for Human and Animal Handling (ECAHCU), located at the Faculty of Pharmacy, Cairo University, Egypt, in compliance with the Weatherall report's recommendations. This committee's approval number is P-9-12-2022. Every effort was made to minimize the number of participants and the suffering of the animals utilized in the research.
Type of the study:
Screening experimental study.
Place and date of the study:
This study was conducted from September 2022 to November 2023 at Cairo University's pharmacy faculty in Egypt.
Source of animal models:
The pharmacology and toxicology department of Cairo University's pharmacy faculty in Egypt procured and authorized animal models.
Inclusion criteria for animal models:
Fibrin blood clots are introduced into an adult male rabbit model that is overweight and weighs around 2 kg. The rabbits were given a week of adjustment before the testing. with a 12-hour light-dark cycle, 50% humidity, and a regulated temperature of 25°C. The rabbits had access to fresh grass.
Exclusion criteria for animal models:
Young and female rabbits; Non-obese rabbits weighing less than 2 kg.
Material
The cell lines were purchased from the American company Accegen Biological. The reference microbe utilized in this work to standardize it was Bacillus subtilis strain W23. Each reagent used was of the analytical grade.
Identification and isolation media:
Mineral Fibrinogen Agar(MFA):
A focused medium for the separation and detection of microbes capable of growing only on fibrinogen as a source of carbon and nitrogen. The following components( g/L) made up this medium:
After combining the components, they were dissolved in boiling water and constantly stirred until the mixture was fully dissolved. Subsequently, the medium was cooled in a water bath to a temperature of 37 ℃ and a pH of 7.3. The plates were immediately put into dishes with a 90 mm diameter, filling them to a capacity of 15-20 ml after the ingredients had been mixed.Every ingredient was purchased from Cairo, Egypt's Algomhoria Company for Chemicals. [18] The mineral fibrinogen agar plate composition is shown in Table 1.
Blood agar:
In addition to the gram stain, the biochemical tests, and the colony morphology, it was an enrichment medium for the discovery of beta-haemolysis of the positive isolates grown on MFA to help identify fibrin-protease generating bacterial species. The contents of the blood agar plate are listed in Table 2.
At 25 ℃, the final pH was 7.3. After the mixture was mixed and autoclaved for 15 minutes at 121℃, 5% sheep blood was added before the mixture was poured onto the plates. Every component was purchased from Cairo, Egypt's Algomhuria Company for Chemicals. [19]
Equipment
Table 3 refers to the list of instruments used in the present study.
Methods
Sample collection:
The samples were grassland soils, obtained at a depth of 11 to 20 cm, and randomly picked from different government sites. Samples were stored in sterile containers at 4℃ until processing.
Isolation of fibrinolytic producing bacterial isolates:
Each soil sample weighed one gram. Ninety nine milliliters of sterile distilled water were added to 250 millilitre Erlenmeyer flasks. The mixture was shaken for two minutes at 400 rpm using a gyrator shaker. In order to plate the dilutions from 10-1 to 10-6 on mineral Fibrinogen agar( MFA) medium, which contains 0.5 g KCl, 0.5 g Mgso4, 1 g KH2PO4, 0.1 g FeSO4, 0.1 g Znso4, 2 g Fibrinogen, and 2% Agar, the soil suspensions were serially diluted in sterile distilled water. At 37 ℃, the medium's pH was adjusted to 7.3. For 48 hours, the plates were incubated at 37 ℃. On the MFA medium, only microbes capable of using Fibrinogen as their only source of carbon and nitrogen could proliferate. Following two rounds of purification using the streak plate technique, colonies exhibiting growth were placed on nutrient agar slants and maintained at 4 ℃. It was possible to subculture the positive isolated colonies on sheep blood agar and determine whether or not beta-haemolysis was present. Every four weeks, the chosen isolate was routinely sub-cultured, and the slant culture was kept refrigerated.
Inoculum preparation:
The inoculum of the studied bacterial isolate was created in 250 ml Erlenmeyer flasks with 50 ml of pH 7 nutrient broth liquid. The medium was autoclaved at 121℃ for 15 min and then infected with a loopful of culture from a nutritional agar slant that had been left overnight. The inoculum was the inoculated flasks, which were shaken for a whole day at 150 rpm. [20]
Identification of fibrinolytic enzymes producing bacterial isolates was performed afterwards through Gram staining, biochemical reactions and 16S rRNA sequencing technique.
Gram stain:
It divided bacteria into two groups according to the composition of their cell walls. On a microscope slide, the bacteria were treated with a crystal violet solution and then iodine, causing the cells to become purple. Gram-positive bacteria retained the stain when coloured cells were treated with a solvent like acetone or alcohol, whereas gram-negative organisms lost the stain and became colourless. When the gram-negative, transparent bacteria were mixed with the counter-stain safranin, they became pink.
Spore shape:
The spore staining technique was used to find this. The slide was cleaned with a Kim-wipe and alcohol to remove any fingerprints. Two circles were drawn with a Sharpie on the bottom of the slide. Using an inoculation loop, two small droplets of water were added to each circle. Using an aseptic technique, a very little number of germs were removed from the culture tube. Microorganisms were present in the droplet of water on the slide. Air dried the slide fully. The slide was heat-fixed by passing it through the flame three or four times with the smear side up. The slide took a long time to cool entirely. The stains were covered up using a piece of paper towel that was positioned inside the slide's border. Above the slide was a beaker filled with heated water. The paper towel was drenched in a malachite green liquid and the slide was left to steam for three to five minutes. The stained paper towel was taken out and disposed of. Water was used to carefully wipe the slide in order to remove any loose paper towel particles.Safranin was used as the counter-stain for a minute. The slide's bottom was dried before it was placed on the microscope's stage and examined using the oil immersion lens.
Spore site:
During the Gram stain test, the spore location was discovered.
Cell shape:
During the Gram stain test, the cell shape was determined.
Blood haemolysis:
On blood agar media, the microorganism's capacity to haemolyze the blood was tried.
Motility test:
It was able to distinguish between germs that moved and those that did not.
The medium was punctured with a sterile needle up to 1 centimetre from the tube's bottom in order to identify a motile colony and assess its isolation. Without a doubt, the needle was held in place both throughout its insertion and withdrawal from the medium. It needed to be incubated for eighteen hours at 35°C, or until there was observable growth. In order to help identify the bacterial isolates that produced the fibrinolytic enzyme, biochemical experiments were carried out later.
Lecithinase test:
Several species were isolated and their differentiation was hypothesized based on lecithinase activity using the differentiated and enriched medium called egg yolk agar. One prevalent ingredient in egg yolks was lecithin. The enzyme lecithinase had the ability to break down lecithovitellin, a lipoprotein component of egg yolk agar, into phosphorylcholine and an insoluble diglyceride, which resulted in the formation of a precipitate in the medium. Lecithin was broken down by microorganisms that possessed the enzyme lecithinase into phosphorylcholine and insoluble diglyceride, which formed a white, opaque precipitation zone that extended outside the colony's boundaries. An opaque halo surrounded a colony that grew on egg yolk agar media, indicating a positive lecithinase activity test result. The test organism was streaked on the plate and then removed in a loop. The plate was checked for an opalescent halo surrounding the inoculates after 24 hours at 35–37°C.
Methyl red test:
Before adding the methyl red pH indicator to the MR broth contaminated tube, the Methyl Red test was conducted. When an organism employed the mixed acid fermentation pathway and generated stable acidic end products, the acids in the medium overcame the buffers, creating an acidic environment.
Catalase test:
To investigate if it might make catalase, a small inoculum of a particular bacterial strain was added to a 3% Hydrogen Peroxide solution. The quick development of oxygen bubbles was noticed.
Oxidase test:
A small piece of filter paper was treated with 1% Kovács oxidase reagent and let to air dry. Using a sterile loop, a well-isolated colony was removed from a newly cultured( 18–24 hours) bacterial plate and rubbed onto prepared filter paper. Changes in colour were observed.
Citrate utilization:
After autoclaving the Simmon Koser's citrate medium at 15 pounds for 15 minutes, five millilitres were extracted.The test tube holding the melted citrate medium was tilted to provide a distinct slant and butt. The designated microbe samples were injected on the inclined medium using labelled tubes and sterile wire. The tubes were incubated at 37°C for a whole day. The colour change of the medium was observed.
Starch hydrolysis:
The bacterial plates were injected for 48 hours at 37°C. The plates were sprayed with an iodine solution using a dropper for 30 seconds following incubation. After that, extra iodine was gushed out. The region around the bacterial growth line was examined.
Gelatin hydrolysis:
Agar medium was made using 1% gelatin. Using an inoculating loop, the provided microbe was applied to the gelatin agar plates, resulting in a single centre streak. The plates were incubated at 37 °C for a whole day. The plates were covered with a HgCl2 solution. The plates were inspected after a little period of time. Test result was positive; there was a noticeable halo around the injected area that indicated gelatin hydrolysis.
Growth at 45 0C:
On nutrient agar media, growth was observed to be possible at 45°C.
Indol test:
Five drops of the Kovács reagent were added straight to the test tube holding the inoculation microbe. An Indol test that was positive was indicated by the reagent layer turning pink to red( a cherry-red ring) a few seconds after the reagent was added to the medium.
Tolerance salinity:
Its capacity to develop on nutrient agar while being responsive to 5% and 7% NaCl was examined.
Voges-Proskauer(VP) test:
For the test, Voges-Proskauer broth, a glucose-phosphate broth loaded with microorganisms, was added to alpha-naphthol and Potassium hydroxide. A successful outcome was indicated by a cherry red tint, whereas an unfortunate outcome was indicated by a yellow-brown color.
Triple sugar Iorn test( TSI):
The triple-sugar iron agar( TSI agar) method was used to assess the fermentation of glucose, lactose, and sucrose, either with or without gas production. It also looked at whether hydrogen sulphide might be produced by amino acids. Phenol red served as the pH indicator in this test medium. The result was a TSIA slant. Using an inoculating needle, the indicated organism was injected into the butt of the TSIA slant. After the inoculating needle was taken out, TSIA zigzag-shaped lifted the surface of the slanted section of the tube. We let the slant incubate for a whole day. Following the incubation period, any alterations in the tube were recorded.
The lactose and/or sucrose fermentation was shown by the slant color. The butt color suggested that glucose was fermenting. Agar bubbles, a sign of gas production, were seen. The medium became dark, indicating the production of H2S.
Glucose fermentation test:
The fermentation reactions of glucose were investigated using glucose purple broth. Peptone and the PH indicator bromcresol purple made up the purple broth. A 1% concentration of glucose was added. Isolated colonies from a 24-hour pure culture of microorganisms were added to the glucose purple broth as an inoculant. Parallel to the inoculation of the glucose-based medium, a control tube of purple broth base was used. The inoculated medium was incubated aerobically for 3 days at a temperature of 35–37 ℃. The medium began to become yellow, which was a sign of a successful outcome. A poor carbohydrate fermentation response was indicated by the lack of yellow color development.
Fructose fermentation test:
The inoculum of a pure culture was aseptically added to a sterile tube with phenol red fructose broth. The infected tube was incubated at 35–37 ℃ for 18–24 hours. An acidic pH change, shown by a colour shift from red to yellow, was a positive reaction.
Maltose fermentation test:
Aseptic transfer of a pure culture inoculum was performed to a sterile tube filled with phenol red maltose broth. The infected tube was incubated at 35–37 ℃ for 18–24 hours. An acidic pH change, shown by a colour shift from red to yellow, was a positive reaction.
Sucrose fermentation test:
The inoculum of a pure culture was aseptically moved to a sterile tube filled with phenol red sucrose broth. The infected tube was incubated at 35–37 ℃ for 24 hours. An acidic pH change, shown by a hue shift from red to yellow, was a positive reaction.
By cultivating multiple soil samples previously obtained on MFA, the ideal environmental and physiological factors affecting the growth of some selected bacterial isolates producing fibrinolytic enzymes, including metal ions such as Fe+2, Mg+2, Co+2, Mn+2, Ba+2, K+1, Na+1, and Ni+2 at a concentration of 10 mM, were determined. By measuring the fibrinolytic activity using the plasmin[ fibrin] plate test, the effects of different pH values[ 4–12], incubation temperatures[ 20–50 ℃], and inoculum quantities[ 1–10%] were also examined.
Optimization of soil bacterial fibrinolytic enzymes production:
The metal ions, nitrogen and carbon sources were obtained from Alnasr pharmaceutical chemical company, Abo-zabal, Alkhanka, Egypt.
There have been investigations into several process variables that increase fibrinolytic enzymes yield. Peptone, Soyabean, and ammonium nitrate at a concentration of 1% W/V were used as nitrogen sources, and the effects of adding carbon sources such as sucrose, mannitol, dextrose and soluble starch at a concentration of 2% W/ V were also studied. The experiments were run in triplicate, and the mean results were presented. To determine the effective parameters and substrates, a series of fermentations were carried out in 500 ml Erlenmeyer flask containing 100 ml of MFA medium with different carbon( 2% W/ V), nitrogen( 1% W/ V) and metal ion( 9 mM) sources as well as inoculation volumes( 1- 10%), PH( 4-12) and temperatures( 20-500C). The carbon sources used were mannitol, dextrose, lactose and sucrose. The nitrogen sources were tryptone, peptone, soyabean, sodium nitrite, ammonium sulfate and ammonium nitrate. The salts of different metal ions including CU +2, Ca +2, Mg +2, Fe +2, Mn +2, Zn +2, Co +2, Ni +2 and Ba +2 were utilized in the culture medium. [21]
Molecular detection:
The predominant bacterial isolate with high fibrinolytic activity was identified using 16S rRNA sequencing and other biochemical tests. Nucleic acid was extracted from a swab by bead-beating in a buffered solution containing Phenol, Chloroform and Isoamyl alcohol. Variable region of 16S rRNA gene was then amplified from the resulting nucleic acid using PCR. The genomic DNA was extracted from 18 hours cultured cells using a DNA purification kit[ PurreLinkTM Genomic DNA Mini Kit with Catalog number: K182002 was purchased from Invitrogen, USA] according to the protocol provided by the manufacturer of DNA purification kit. The 16S rRNA gene was amplified by PCR[ PCR SuperMix kit was purchased from Invitrogen,USA] using forward[ 5-AGAGTTTGATCCTGGCTCAG-3-] and reverse[ 5-GGTTACCTTGTTACGACTT-3-] primers. PCR amplicons from up to hundreds of samples were then combined and sequenced on a single run. The resulting sequences were matched to a reference database to determine relative bacterial abundances. Polymerase Chain Reaction( PCR) is a powerful method for amplifying particular segments of DNA. PCR uses the enzyme PlatinumTM Taq DNA polymerase with catalog number 10966018[ purchased from Invitrogen, USA] that directs the synthesis of DNA from deoxynucleotide substrates on a single-stranded DNA template. DNA polymerase adds nucleotides to the 3` end of a custom-designed oligonucleotide when it is annealed to a longer template DNA. Thus, if a synthetic oligonucleotide is annealed to a single-stranded template that contains a region complementary to the oligonucleotide, DNA polymerase can use the oligonucleotide as a primer and elongate its 3` end to generate an extended region of double stranded DNA. Denaturation is the initial PCR cycle stage The DNA template is heated to 94° C. This breaks the weak hydrogen bonds that hold DNA strands together in a helix, allowing the strands to separate creating single stranded DNA. Annealing is the second PCR cycle. The mixture is cooled to anywhere from 50-70° C. This allows the primers to bind (anneal) to their complementary sequence in the template DNA. Extension is the final step of PCR cycle. The reaction is then heated to 72° C, the optimal temperature for DNA polymerase to act. DNA polymerase extends the primers, adding nucleotides onto the primer in a sequential manner, using the target DNA as a template. With one cycle, a single segment of double-stranded DNA template is amplified into two separate pieces of double-stranded DNA. These two pieces are then available for amplification in the next cycle. As the cycles are repeated, more and more copies are generated and the number of copies of the template is increased exponentially. The amplified PCR product was sequenced using a genetic analyzer 3130XL[ purchased from Applied biosystems, USA]. DNA sequence homology search analysis of the predominant bacterial isolate was achieved using Blast n algorithm at NCBI website. [22]
Upstream process:
Bacterial recombinant DNA production of fibrinolytic enzymes:
Primer for expression of FP,
Forward primer: TTCCAACAAAATGGACGTGA
Reverse primer: CAGGATAGCCGATTGTGCTT.
The expression vector was pET-14b( purchased from Novagene in the United States). The expression vector was designed via GenSmart™ Design sortware. The promotor was T7 Lac, and the tagged protein was 6x histidine linked to the C terminus of an molecule. Escherichia coli BL21[ DE3] polys S served as the expression host. All of them were used in the manufacture of Fibrin-proteinase fibrinolytic enzyme using bacterial recombinant DNA technology. IPTG was the transcription process's inducer. The principal host for the plasmid synthesis and replication was Escherichia coli DH5 [obtained from Stratagene corporation, USA]. Fibrin-proteinase genomic DNA was isolated from Bacillus cereus ATCC 14579 discovered in several Egypt soil conditions using restriction endonuclease type II enzymes[ DNA cutting enzymes] Taq I and Hae III. These cutting enzymes were bought from the German business Sigma-Aldrich. Furthermore, genomic DNA was amplified using the polymerase chain reaction method before being sub-cloned to the prokaryotic expression vectorPET-14b using the same restriction endonuclease type II enzymes used to extract genomic DNA from Bacillus cereus ATCC 14579. This was followed by the transformation of pET-14b into the polys S expression host Escherichia coli BL21[ DE3]. The addition of IPTG stimulated transcription at the promotor site T7 Lac, hence initiating gene expression. Luria agar and broth[ LA, LB] were used for routine bacterial culture for 24 hours at 37 0C incubation temperature. Ampicillin and/or Kanamycin were added to the culture medium according to the manufacturer's guidelines. [23]
Downstream process:
Purification was carried out using 500 ml of crude enzyme extract. For recombinant expression yield, a centrifuge tube was spun for 5 minutes at 4000 rpm. The expressed protein was extracted from the supernatant through salting out using 70% Ammonium sulfate.
Procedure of salting out using 70% Ammonium sulfate:
38 grams of ammonium sulfate were dissolved in 100 ml distilled water at 25 ℃ to prepare 70% saturated solution. The retentate was subjected to 70% W/ V Ammonium sulfate precipitation via stirring using a magnetic stirrer at 4 ℃ for over night; moreover the precipitate formed was centrifuged at 10000 rpm for 30 min in a centrifuge 4K15( purchased from Sigma-Aldrich, German) and the precipitate obtained was dissolved in a minimal quantity of 60 mM Tris-HCl buffer, pH 7.2. Furthermore, the expressed protein was purified using Ni-NTA Agarose Column affinity chromatography technique[ Ni-NTA Purification System with catalog number K95001 was purchased from Invitrogen, USA]. [24]
Procedure of Ni-NTA Agarose Column affinity chromatography:
8 ml of cell lysate was added to a prepared purification column. To keep the resin suspended in the lysate solution, it was bound for 15–30 minutes at room temperature using gentle agitation on a rotating wheel. The resin was settled by low speed centrifugation at 500 rpm and carefully the supernatant was aspirated. The column was washed with 5 ml denaturing binding buffer by resuspending the resin and rocking for 3 minutes. The column was washed with 4 ml denaturing wash buffer( pH 6.0) by resuspending the resin and rocking for two minutes; then the column was washed with 10 ml native wash buffer by resuspending the resin and rocking for 3 minutes. The column was clamped in a vertical position and the cap was snapped off on the lower end. The protein of interest was eluted with 9 ml native elution buffer. An 1 ml of the supernatants after each wash were kept at 4º C for SDS-PAGE analysis. [25]
SDS-PAGE procedure:
SDS-PAGE Kit utilized during the present study was HiPer® SDS-PAGE Kit with Catalog number: HTP001 purchased from Hi-Media Laboratories, India. Procedure of SDS-PAGE was performed according to the manufacturer's instructions. The electrophoresis unit was assembled such that the glass plates were clamped to the unit along the spaces placed in-between them at two vertical edges. 1% Agarose( 0.5 g in 5 ml of distilled water) was prepared. The agarose was boiled to dissolve then a thin horizontal layer was poured the lower edge of the plates to seal the assembly. The agarose was allowed to solidify via allowing it to cool down for 5- 10 minutes. The preparation of 12% separating gel was carried out through the addition of the following:
The gel was poured in-between the plates and was allowed to solidify for an hour. Immediately after the gel was poured, the distilled water was added to level the gel. After one hour the water was poured off by inverting the casting assembly. 5% stacking gel was prepared through the addition of the following components:
After the addition of TEMED, the components were mixed gently by swirling the beaker. The stacking gel was poured was poured on the top of the separating gel and immediately the comb avoiding air bubbles was placed; as well as the gel was allowed to solidify for 30 minutes. 1X Tris-Glycine-SDS gel running buffer was poured in the unit such that the buffer connects the two electrodes, and hence completed the flow of current. The comb was removed from the stacking gel carefully. 5 µl of prestained protein ladder and 20 µl of samples immediately were loaded after the heat treatment in the wells created by the comb in the stacking gel. The power cord was connected to the electrophoertic power supply according to the conventions: red Anode and black Cathode. It was allowed to electrophorese at 100 volts and 10 mA until dye front reached 0.5 cm above the sealing gel. The gel was carefully removed from in-between the plates using spatula into the plastic tray containing distilled water. The gel was washed for 1 minute. The water was discarded and staining destaining procedure was proceeded. Staining and destaining of the gel included the addition of 50 ml of staining solution in the tray containing gel, till the bands were visible. The gel; afterwards was removed the staining solution. The staining solution could be reused 2-3 times; then the gel was washed by rinsing in distilled water till a considerable amount of stain was leached out from the gel. Changing the distilled water was kept for 3-4 times. 50 ml of destaining solution was added to the gel with constant moderate shaking. The destaining was continued till clear and distinct bands were observed. In the end of the process, the gel was removed from the destaining solution. After staining and destaining of the gel the molecular weights of samples were compared with that of the protein markers. Molecular weights were confirmed via a mass spectrometer. [26] Table 4a refers to the composition of 12 SDS-PAGE gel; while Table 4b showed the components of 5% stacking gel.

Table 4a.
It demonstrates the components of 12% SDS-PAGE gel.
| Component | Volume |
|---|---|
| 30% Acrylamide-bis-acrylamide solution | 6 ml |
| Distilled water | 3 ml |
| 2.5 X Tris-SDS Buffer( PH 8.8) | 6ml |
| 10% APS solution | 125 µl |
| TEMED | 18 µl |
Table 4b.
It refers to the components of 5% stacking gel.
| Component | Volume |
|---|---|
| 30% Acrylamide-bis-acrylamide solution | 1.3 ml |
| Distilled water | 5.1 ml |
| 5X Tris-SDS Buffer( PH 6.8) | 1.6 ml |
| 10% APS solution | 75 µl |
| TEMED | 10 µl |
Quantitative purified fibrinolytic protein estimation by lowry method:
The determination of of the unknown concentration of purified fibrinolytic enzyme in a sample was carried out using standard curve obtained through Lowry assay using bovine serum albumin as the standard, and the values were expressed as mg/ ml. [27]
Determination of cellular position of fibrinolytic enzyme:
This was determined through spectrophotometric analysis of thrombolytic activity( SATA) assay, standardized by tissue plasminogen activator( tPA), which could quantitatively measure in vitro thrombolytic activity. Blood clots were formed, uniformly, by mixing citrated whole blood with partial thromboplastin time( PTT) reagent, together with calcium chloride. Then, designated concentrations of tPA as a standard fibrinolytic drug, supernatants and cell lysates of different centrifuged soil samples containing test fibrinolytic enzymes at 500 rpm were added. The released red blood cells from each clot were quantified using spectrophotometry( λmax=405nm) as an indicator of thrombolytic activity. [28]
Assessment of secretion of antibodies to bacterial fibrinolytic enzyme:
The quantity of IgG antibodies to the enzyme of interest in mouse serum was dictated via ELISA. 100 µl of an emulsion containing 50 µg of FP and an equal volume of complete Freund adjuvant[ purchased from Sigma Aldrich, USA) at 3 week intervals to 10 mice.Sera separated and used as primary antibodies. [29]
The kinetic parameters Km and Vmax determination:
The parameters of kinetic, Michaelis–Menten constant( Km) and maximum velocity ( Vmax) of purified fibrinolytic enzymes were ascertained with suited concentrations of fibrinogen( 2–11 mM) as the substrate. Data was connected to the nonlinear exponential stage union curve of regression. The outcome of fibrinolytic enzymes were computed aside measurement of the hydrolysis rate of fibrin clots below standard laboratory circumstances victimizing the equation of Michaelis-Menten. [30]
Formulation of bacterial fibrinolytic enzymes:
The secreted fibrinolytic enzymes from the predominant bacterial isolate grown on SFA medium was formulated as a sterile solution intended for the utilization via parenteral routes of administration either IV, IM or SC injections. Lyophphilized powder of recombinant FP was obtained for preparation of a solution for injection of 50 units and 100 units in vials. The preparation was administered intravenously in the initial dose of 200 IU in 60 ml of isotonic solution of sodium chloride during 30 minutes( 25 drops per minute). Later, the administration of FP in the dose of 150 IU per hour was followed. Table 5 demonstrates the composition of parenteral dosage form of FP.
Compatibility studies of fibrinolytic enzyme and polymer:
These were performed via FTIR spectroscopy and DSC. Recombinant FP and different excipients utilized in the preparation of intravenous and subcutaneous parenteral formulations were characterized by FT-IR( Perkins-Elmer 1600 FTIR spectrophotometer) spectroscopy and DSC( Shimadzu-DSC 50) to find the compatibility. The optimized formulations were blended with 100 mg KBr; then compressed into discs which were scanned at 5mm/ sec with a resolution of 1 cm-1 at a range of 4000-300 cm-1. Experiments of thermal analysis were carried out utilizing various scanning calorimeter( DSC). The samples of the optimized formulation were heated in hermetically sealed Aluminium pans at a temperature range of 0-4000 0C at a constant rate of 105 0C/minute under a purge of nitrogen( 40 ml/ min).
Testing the fibrinolytic activity of recombinant fibrinolytic enzymes:
This was achieved through the following calibration assays:
Fibrin plate method:
Sigma-Aldrich in the United States provided the human fibrinogen, plasmin, and thrombin. Using a fibrin plate test, fibrinolytic activity was ascertained. In a petri dish, agarose solution( 6 ml, 2% w/v, in 50 mM pH 7.4 sodium phosphate buffer), human fibrinogen (7 ml, 0.5% w/v, in 60 mM pH 7.4 sodium phosphate buffer), and 0.1 ml of Thrombin( 5 NIH U/ml in 60 mM pH 7.4 sodium phosphate buffer) were combined. A fibrin clot layer formed after the solution was kept at room temperature for two hours. A 10 mm diameter well-punched fibrin plate was filled with 50 µl of the fibrinolytic enzyme-laden supernatant from various samples collected throughout the experiments. The plate was then incubated at 37°C for 24 hours. The lytic area was measured in order to quantify the enzyme activity. The quantity of enzyme in 30 µl of enzyme solution that created a clear zone measuring 1 mm2 at pH 7.4 and 37°C for 18 hours is known as one unit of fibrinolytic enzyme activity. The FEA of the samples was determined by creating a calibration plot with plasmin( 1000 U/ml) as the reference standard. [31]
Casein digestion method:
Using the casein digestion method—which measures the amount of tyrosine produced from digested casein—fibrinolytic enzyme activity was ascertained. Utilizing the modified Sutar et al. technique, activity was confirmed to be 100% victimized (1986). 10 mg casein, 50 mM Tris-HCl, pH 9.0, and 0.1 ml( or the suitable diluent) of supernatant were added to the reaction mixture( 3 ml). After 20 minutes at 37° C, the reactions were halted by adding 0.3 ml of 4.4 M HCl and 3.5 ml of 6% w/v trichloroacetic acid( TCA). After that, the reaction was placed on ice for thirty minutes, and then Whatman #1 filter paper was used for filtering. At 280 nm, the TCA soluble fraction's absorbance was discussed. The standard plasmin curve was used to calculate the units of enzymatic activity. [32]
Effect of plasminogen addition on fibrinolytic enzyme activity:
200 µl of 2% fibrin( pH 8), 200 µl of 0.2 M Tris-HCl( 20 mM CaCl2, pH 7.8), 100 µl of the enzyme solution, and 100 µl of plasminogen( 0.730 U) were all included in the incubation mixture. By employing the fibrin plate test, fibrinolytic activity was quantified and contrasted with the enzyme's activity in the absence of plasminogen. Five millilitres of 2.1% fibrin (pH 8) were added to four millilitres of 35 mM Tris-HCl( containing 0.80% agar and 0.20 M NaCl, pH 7.9) at 45°C to create the fibrin plate. [33]
Effects of pH and temperature on enzyme activity:
The reaction mixture was incubated for two hours at temperatures ranging from 25 to 95 degrees Celsius in 605 millimolar phosphate buffer (pH 7.0) in order to establish the ideal temperature for the enzyme activity of extracellular fibrinolytic enzymes. The fibrin plate assay was used to measure enzyme activity. The pH range that was shown to be ideal for enzyme activity was pH 4.0 to pH 11.0. The enzyme was combined with several buffers in 65 mM sodium phosphate buffer (pH 7.0). The 0.2 M citrate-phosphate buffer( pH 2.0-5.0), 65 mM sodium phosphate buffer (pH 6.0-7.0), Tris-Cl buffer( pH 8.0-9.0), and glycine-NaOH buffer( pH 10.0-12.0) were the buffers that were employed. After the enzyme was incubated in 65 mM sodium phosphate buffer( pH 6.5), the amount of activity that persisted was used to measure the thermostability of the enzyme activity. [34]
Effect of metallic ions on enzyme fibrinolytic activity:
The effect of the enzyme was investigated using MgSO4, CoCl2, MnCl2, ZnCl2, FeSO4, CuSO4, and CaCl2. The partial purified protein was incubated in both the absence and the presence of metal ions including Mg2+, Co2+, Mn2+, Zn2+, Fe2+, Cu2+ and Ca2+ with a final concentration of 10 mM in Tris buffer( pH7.3) for 2 hrs at 37o C and the fibrinolytic activity was assayed using fibrin plate assay. [35]
Experimental venous thrombosis Technique:
Biological activity of the test fibrinolytic enzyme was compared to standard activity in a rabbit model of experimental venous thrombosis. 300 male rabbit models weighing nearly 2 kg were collected and divided into three groups (test, standard, and control). Each group consisted of 100 of rabbit models. In test and control groups, thrombosis was induced by jugular vein stasis and injection of 10 mg/kg thromboplastin into the ear vein of each rabbit model. Animals were randomized to receive IV test fibrinolytic enzyme 0.2, 2.0, 4.0, or 9.0 mg/ kg or vehicle control and compared to standard intravenous plasmin and vehicle control. On the other hand, activated partial thromboplastin time( APTT) test was performed to detect the anticoagulant effect of fibrinolytic enzymes; as well as the clotting time( CT), thrombin time( TT), Euglobulin lysis time( ELT), fibrinogen( FIG) and hemorheology conditions were determined. [36]
Determination of phamacokinetic profile:
In vitro fibrinolytic enzyme release and in vivo bioavailability study of optimized formulations were determined according to British pharmacopoeia 2023. A 0.5- 1 ml rabbit animal model blood samples were withdrawn before dosing and immediately after dosing at 30, 60, 90, 120, 240 and 360 minutes; afterwards blood samples were refrigerated within 1 hour of sampling and centrifuged at 40C. Fibrin-proteinase concentrations were measured using HPLC. HPLC analysis was performed on a reverse- phase column using phosphate buffer[ PH 4.3] and acteonitrile[ 750: 250, V/V] as a mobile phase with a flow rate of 1 ml / min. The limit of the determination with UV detection was at 225 nm. [37]
Statistical Analysis
Triplets were used for all cultural experiments. They used a means and standard deviation display. Both statistical analysis based on Excel spreadsheet software and one-way analysis of variance( p value≤0.05) were employed as methods. In the current investigation, the F test was applied.
Results
The present study revealed the possibility of the production Fibrin proteinase secreted from Bacillus cereus ATCC 4342 as a fibrinolytic agent via bacterial recombinant DNA technology.
Table 6 displays the positive bacterial isolates distribution on MFA. Table 7 displays distribution of the main producing fibrinolytic bacterial bacilli on MFA.
Isolation of the bacterial fibrinolytic enzymes fromdifferent soil environments in Egypt:
During screening of fibrinolytic enzyme producing bacterial isolates, a total of 31 Bacillus SPP were isolated from different soil environments in Egypt. These isolates were subjected to primary screening via MFA plates. Only the isolates which were able to utilize fibrinogen as a sole metabolic carbon and nitrogen source were able to grow on MFA plates. All 31 isolates gave relatively large clear zones on fibrin plate assay. The fibrinolytic enzyme activities of 31 Bacillus isolates were determined between 478- 623 U/ ml. In secondary screening on blood agar plates 17 isolates out of 31 were detected with considerable high fibrinolytic activities through different fibrinolytic assays such as casein digestion technique and fibrin plate assay. Only these 17 isolates were tested for their thermostability. The morphological, biochemical reactions, Gram staining and molecular detection using 16S rRNA gene analysis revealed that Bacillus cereus ATCC 4342 and Bacillus subtilis DC 27 were the main positive bacterial isolates producing fibrinolytic enzymes in the present study. Bacillus cereus ATCC 4342 represented 17 out of positive bacterial isolates; while Bacillus subtilis DC 27 represented 14 out of the positive bacterial Bacilli producing fibrinolytic enzymes. The isolate Bacillus cereus ATCC 4342 was found to have the highest level of fibrinolytic activity together with the highest thermostability; thus it was selected for further investigations.
Bacillus cereus ATCC 4342 was noticed to be a Gram positive Bacillus shaped, endospore forming bacterium. The cells were 2- 3 µm long and 0.8- 1 µm wide. Its spores were oval in form and 0.7- 0.8 µm wide with 1- 1.4 µm long. During 16S rRNA gene analysis, both major bacterial isolates were identified based on 16S rRNA gene sequence using Blast analysis which showed high degree of similarity with these both types of Bacilli. Molecular mass of fibrinolytic enzyme Fibrin-proteinase[ FP] produced by Bacillus cereus ATCC 4342 was approximately 30 KDa as determined via mass spectrometer and confirmed with SDS-PAGE technique.
Purification of the test fibrinolytic enzymes:
The yielded fibrinolytic enzyme after the purification via immobilized Nickel affinity chromatography ranged from 0.184- 1.926 U/ ml. The productivity reached a maximum at 24 hours after which it decreased due to sporulation occurrence during culturing the collected soil samples from different environments in Egypt on MFA plates. The degree of purity of fibrinolytic enzymes reached nearly 83% as determined by SDS-PAGE chromatography and western blot technique. The fibrinolytic enzyme specific activity reached 40.71 U/ mg estimated via lowry assay. The average total protein extracted from the supernatant of culture plates reached nearly 0.228 mg. The total enzyme activity was detected to be 9.281 U. The specific activity was calculated via dividing total activity on total protein content extracted from the supernatant of the culture through Lowry assay.
Optimization of the growth media included different carbon and nitrogen sources. Optimization of various PH, Temperature, inoculum volumes and metal ions was performed in an attempt to determine the optimum physiological and environmental factors affecting the growth of the predominating bacterial isolates producing fibrinolytic enzymes.
Effect of different carbon sources on fibrinolytic enzyme productivity:
A series of experiments with different carbon sources: starch, lactose, sucrose, mannitol, dextrose were performed to determine the best carbon source of fibrinolytic enzyme productivity of Bacillus SPP. The concentration of carbon source was maintained constant at 2 % W/ V in all experiments. Mannitol was found to be the optimum carbon source as it yielded 3.15 U/ ml of fibrinolytic enzymes.
Table 8 confirm the outcomes of different carbon sources on productivity of fibrinolytic enzymes.
Effect of different inoculum volumes on FP productivity:
The inoculum volume was optimized via comparing the productivity resulting with 1- 7 % V/ V inoculum volumes. The highest level of productivity was detected at 2% inoculum volume( 1.89 U/ ml).
Table 9 demonstrates the influence of different inoculum volumes on the production of fibrinolytic enzymes at incubation conditions PH 7.4, temperature 37 0C and 18 hours incubation period.
Effect of different Nitrogen sources on FP productivity:
A series of experiments were performed with different nitrogen sources. Peptone was noticed to be the best organic nitrogen source due to increase in productivity( 2.013 U/ml) as compared with the control.
Table 10 shows the effects of different nitrogen sources on FP productivity.
Effect of different metal ions on fibrinolytic enzyme productivity:
Zn+2 and Hg+2 metal ions showed negligible effects on the production of fibrinolytic enzyme Fibrin-proteinase secreted from Bacillus cereus ATCC4342; while Ca+2 and Mn+2 demonstrated the optimum conditions on the growth of Bacillus SPP producing fibrinolytic enzymes during the present study.
Table 11 demonstrates the effects of different metal ions on the fibrinolytic enzyme productivity.
Determination of fibrinolytic enzyme cellular position:
The fibrinolytic enzymes were extracellular with 91%; while 9 % were showed as intracellular.
Table 12 shows the cellular position of the fibrinolytic enzymes via the spectrophotometric assay.
Effect of different PH on FP productivity:
The PH range 8- 10 was optimal for optimization of the growth of bacterial isolates producing fibrinolytic enzymes on MFA plates with the highest productivity was noticed at PH 8. negligible growth was detected at acidic PH degrees. The fibrinolytic enzyme FP purified from the culture supernatant of Bacillus cereus ATCC 4342 was relatively was relatively stable at PH range 7.4- 10 for 18 hours.
Effect of different incubation temperatures on FP productivity:
The fibrinolytic enzyme FP purified from the culture supernatant of Bacillus cereus ATCC 4342 was relatively stable with the highest productivity at temperature 43 ℃ for nearly 60 minutes. The optimum temperature of protease production was at 35-43 ℃ at incubation time ranged from 18-24 hours.
Table 14 demonstrates the effects of different temperatures degrees on FP production.
Effect of different PH degrees on fibrinolytic enzyme activity:
FP retained the fibrinolytic activity and stability at PH range 4-12. The activity was detected at alkaline PH range 8-10 with the highest enzyme action at PH 8.
Effect of different temperatures on fibrinolytic enzyme activity:
Effect of different Temperatures on fibrinolytic enzyme activity:
The purified enzyme was stable and retained 100 % of its activity at a temperature ranged from 20 to 65 0C with the highest stability and activity was detected at 55 0C( 727 U/ ml) for 1 hour. On the other hand, Tm of refined enzyme was recorded at 58 0C.
Effect of different metal ions and EDTA on fibrinolytic enzyme activity:
EDTA inhibited the proteolytic enzyme activity; while Fe+3 metal ion showed negligible effects on enzyme action[ 23 U/ ml]. On the other hand, the fibrinolytic activity reached the highest activity with Na+1[ 681 U/ ml] and K+1[ 653 U/ ml] monovalent cations respectively. Mn+2, K+1, Na+1, Co+2, Ni+2, Ni+2, Ba+2, Fe+2 metal ions were found to be enzymatic activators that increased FP activity by 29 %, 31 %, 35 %, 19 %, 17 %, 10 %, 21 %, 16 % respectively.
Effect of plasminogen addition on fibrinolytic activity:
The mixture of plasminogen and the extracted fibrinolytic enzymes addition did not give a higher fibrinolytic activity through fibrin plate assay than the activity of fibrinolytic enzymes alone because a similar result was obtained from fibrin plate analysis; thus this suggested the test fibrinolytic enzymes degraded fibrin directly and it was not the activity of plasminogen activator which degraded fibrin via activating plasminogen.
Production of FP via bacterial recombinant DNA technology:
The gene of interest encoding FP was isolated from Bacillus cereus ATCC 4342 using restriction endonuclease type II cutting enzymes which cut DNA at specific locations; Then joined to the plasmid expression vector pET-14b through ligase enzyme and the homo-polymer tailing. The recombinant plasmid was then introduced into the bacterial expression host Escherichia coli BL 21(DE3) Ploys S through transformation process. On the other hand the expression host was cloned as well as induced for expression of recombinant protein through addition of IPTG. The produced recombinant( transformed) FP fibrinolytic enzyme was screened for its production through the selection and the screening of the transformed cells.
Purification of recombinant FP fibrinolytic enzyme:
The crude enzyme yielded after salting out with 70% ammonium sulfate suggested that SDS-PAGE could retain the specific molecular mass proteins. The fibrinolytic enzyme produced from the predominating bacterial isolate on MFA was called Fibrin-proteinase( FP). Its productivity increased via bacterial recombinant DNA technology 15folds because it reached to 28.5 U/ ml. The specific activity reached nearly 600 U/ mg protein. The total protein reached approximately 3.5 mg. FP was purified from gel filtration during SDS-PAGE procedure. A single band having a molecular mass of nearly 30 KDa was observed on SDS-PAGE. FP showed purification fold 3.2± 2.7 with final enzyme recovery 56± 3.14 %.
Kinetic parameters Km,Vmax, Kcat of purified fibrinolytic enzyme were 5.78 mM, 100 µmol/ mg/ min and 2.41× 10( 2) M( -1) S( -1) respectively. Kinetic parameters values during the present study were determined through Michaelis- Menten plot. These parameters values reflected the presence of high affinity of FP towards Fibrin substrate. The isoelectric point of purified enzyme reached 8.3.
Table 15 demonstrates Michaelis Menten Plot.
Table 17 represents fibrinolytic activity of test thermostable FP by Casein digestion assay.
Table 18 represents Fibrin-proteinase release profile.
Table 20 represents the effects of various temperatures on the fibrinolytic activity of Fibrin-proteinase.
Table 21 represents the effects of different metal ions and inhibitors on fibrinolytic activity of Fibrin-proteinase.
Determination of phamacokinetic profile of FP:
Table 22 displays the pharmacokinetics profile of Fibrin-proteinase.
Like other proteins, FP was noticed to be metabolized proteolytically in the liver and eliminated via the kidneys. A little amount of the FP enzyme was detected to be excreted unchanged in bile. The pharmacokinetic profile of FP is represented in Table 21. The test enzyme reached the peak concentration within one hour after parenteral SC administration. Repeat usage after 5 days and for 12 months was not recommended due to the development of mild neutralizing antibodies because of its prokaryotic origin. It was noticed that the usage of moderately high loading dose up on repeating the dose overcome the neutralizing antibodies; thus giving excellent efficacy as a fibrinolytic enzyme. The drug efficacy diminished after 5-6 hours and it was inefficacious after 12 hours. The loading dose was noticed to neutralize any anti- FP antibodies which appeared up on repeated doses. The invitro release of FP parenteral SC formulation was up to 15 minutes. The percentage relative bioavailability of FP from optimized SC parenteral formulation reached 76 %.
Determination of fibrinolytic activity of FP:
This was determined through fibrin plate assay, the spectrophotometric method and Casein digestion test. Fibrin plate assay revealed that the fibrinolytic enzymatic activity ranged from 372± 19.04- 529± 30. 86 U/ ml. FP was detected to possess casein-lytic and proteolytic activities similar to serine proteinases. FP showed proteolytic trypsin like activity; while it did not demonstrate any chymotrpsin proteolytic like actions. The effect of plasminogen addition revealed that this enzyme was a plasminogen independent type and degraded the fibrin clot directly which makes the enzyme a potent secure candidate for oral administration. The purified FP was highly specific against N-Suc-Ala-Ala-Pro-Phe-pNA( 190/ mmol/ min/ ml) and clot lytic activity reached 19± 2.3 % within 1 hour invitro. The purified fibrinolytic enzyme showed little erythrocytic lysis activity; thus confirmed safety to prevent various health risks, including hemolytic anemia. Based on this study, administration of fibrinolytic enzyme produced from Bacillus cereus ATCC 4342 was safe for clinical applications. FP prolonged CT, PT, APTT and TT. FP improved the hemorheology conditions in the rabbit models. Activated partial thromboplastin time increased by approximately 50 %, fibrinogen content decreased by 63 %. Fibrin degradation also increased obviously, but the plasminogen content did not change obviously. Thrombogenesis inhibition rate was noticed to be approximately 65 %. The changes of these biochemical data revealed the thrombolytic properties of FP in vivo. The thrombolytic effect of FP[ 3841 U.kg-1 on venous thrombus in rabbit animal models was most remarkable versus that of control[ P< 0.001], but was not divergent versus that of standard plasmin drug at the same dose[ P< 0.05]. The results showed that FP is a plasmin like protein. The protease fully hydrolyzed fibrin clots in 2-3 hours. The fibrinogen degradation pattern generated by FP as a function of time was similar to that obtained with plasmin. The results demonstrated FP was a serine protease analog to the Substilin family consisting of 275 amino acid residues. N-terminal aminoacid sequence was detected to be QATVPYEQPHCSL which was different from that of other known fibrinolytic enzymes. Results of the fibrinolysis pattern showed that FP rapidly hydrolyzed the fibrin alpha and gamma chains of fibrin clots; while it slowly hydrolyzed the beta chains of fibrin clots. The enzyme showed anticoagulant activity measured with activated with partial thrombin time and pro-thrombin time tests. FP slightly decreased ELT.
Bioinformatic analysis and molecular docking of FP:
Further bioinformatic analysis using Swiss model and Clustal omega soft-wares demonstrated that FP had a PI of 5.8, low instability index, high aliphatic index and low GRAVY value. Molecular docking analysis using Autodock Vina software indicated that there were favorable interactions between FP and the fibrin substrate, as indicated by a high binding affinity( ∆G:- 20.7 Kcal/ mol) and low Kd value( 5.1E- 14 M). BLASTn analysis of nucleotide sequence encoding FP revealed approximately 70 % homology with the gene encoding the fibrinolytic enzyme Subtilisin DFE.
Determination of compatibility studies:
No drug polymer interaction or incompatibility was detected via FTIR and DSC studies.
Determination of Immunogenicity of FP:
This was determined via ELISA which showed no detectable antibodies after the administration of the initial dose. On the other hand, moderate antibodies were produced up on repeated doses. It was noticed that the utilization of loading dose of FP could lessen immunogenicity up on repeating doses for a maximum 4 days. FP lost more than 70 % its fibrinolytic activity after its administration more than 5 days due to evoking graded titers of neutralizing antibodies.
Table 23 informs Molecular 16 S rRNA detection of the predominating bacterial isolates producing FP.
Table 23.
It indicates Molecular 16 S rRNA detection of the predominating bacterial isolates producing FP.
Table 23.
It indicates Molecular 16 S rRNA detection of the predominating bacterial isolates producing FP.
| Description | Scientific Name | Max Score | Total Score | Query Cover | E value | Per. ident |
|---|---|---|---|---|---|---|
| Bacillus cereus ATCC 4342, complete genome | Bacillus cereus ATCC 4342 | 1423 | 1423 | 100% | 0 | 100 |
| Bacillus cereus strain FT9, complete genome | Bacillus cereus | 1395 | 1395 | 100% | 0 | 99.35 |
| Bacillus cereus strain BC-AK genomic sequence | Bacillus cereus | 1373 | 1373 | 100% | 0 | 98.83 |
| Bacillus cereus G9241, complete genome | Bacillus cereus G9241 | 1373 | 1373 | 100% | 0 | 98.83 |
| Bacillus cereus strain FM1, complete genome | Bacillus cereus | 1373 | 1373 | 100% | 0 | 98.83 |
| Bacillus cereus strain 03BB87, complete genome | Bacillus cereus | 1373 | 1373 | 100% | 0 | 98.83 |
| Bacillus cereus strain BHU2 chromosome | Bacillus cereus | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus MRY14-0075 DNA, complete genome | Bacillus cereus | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus strain EFR-1 chromosome | Bacillus cereus | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus NC7401 genomic DNA, complete genome | Bacillus cereus NC7401 | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus Q1, complete genome | Bacillus cereus Q1 | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus AH187, complete genome | Bacillus cereus AH187 | 1367 | 1367 | 100% | 0 | 98.7 |
| Bacillus cereus strain AR156, complete genome | Bacillus cereus | 1362 | 1640 | 100% | 0 | 98.57 |
| Bacillus cereus strain CC-1 chromosome, complete genome | Bacillus cereus | 1351 | 1351 | 100% | 0 | 98.31 |
| Bacillus cereus 30090 DNA, complete genome | Bacillus cereus | 1351 | 1351 | 100% | 0 | 98.31 |
| Bacillus cereus ATCC 10987 chromosome | Bacillus cereus ATCC 10987 | 1267 | 1267 | 100% | 0 | 96.36 |
| Bacillus cereus strain G9241 chromosome | Bacillus cereus | 1267 | 1267 | 100% | 0 | 96.36 |
| Bacillus cereus FRI-35, complete genome | Bacillus cereus FRI-35 | 1267 | 1267 | 100% | 0 | 96.36 |
| Bacillus cereus ATCC 10987, complete genome | Bacillus cereus ATCC 10987 | 1267 | 1267 | 100% | 0 | 96.36 |
| Methyl red | + |
| Voges-proskauer | + |
| Citrate utilization | + |
| Starch hydrolysis | + |
| Gelatin hydrolysis | + |
| Growth at 45 0C | + |
| TSI | + |
| Tolerance salinity | |
| 5% NaCl | + |
| 7% NaCl | + |
| Saccharide fermentation | |
| Glucose | + |
| Fructose | + |
| Maltose | + |
| Sucrose | + |
Table 24 shows the biochemical reactions of the predominating bacterial isolates producing FP.
Discussion
Thromboembolic diseases rank among the world's major causes of mortality. The few fibrinolytic medications on the market today, such as urokinase, streptokinase, alteplase, and tencteplase, have significant drawbacks such as haemorrhage propensity, low specificity, and high production costs. To get around these problems, it is necessary to look for new sources of fibrinolytic enzymes.
In the current study, a new fibrinolytic enzyme( FP) has been identified. It is secreted by the predominant bacterial isolates that were obtained from several environmental soil samples in Egypt. Based on morphology, biochemical responses, and molecular detection, the predominant bacterial isolate generating FP fibrinolytic enzyme in this investigation was Bacillus cereus ATCC 4342, as determined by high-throughput sequencing of 16S rRNA. Every carbon source that might potentially provide fibrinolytic enzymes was selected. It was interesting to note that mannitol showed relatively the highest productivity( 3.15 U/ ml) during the present study; while, Narasimhan M study, 2015 stated that Corn flour was ideal agent as a carbon source for the production of fibrinolytic enzymes and soyabean powder was the optimum Nitrogen source. [38] In the present study, Nitrogen source was found to play a major role in enhancing the fibrinolytic productivity in the media. The optimum Nitrogen source in the present study was noticed to be peptone( 2.013 U/ ml). Fibrin-proteinase obtained from Bacillus cereus ATCC 4342 demonstrated higher productivity and activity at basic PH. The optimum inoculum volume for the production of fibrinolytic enzymes from different soil environments was consummated at 2% V/ V. Syahbanu F et al study, 2020 confirmed that 1% V/V inoculum volume was the optimal one for the growth of the predominating bacterial isolates producing the fibrinolytic enzymes in different culturing media. [39] At inoculum volumes higher than 7% V/ V, there was a decline in the productivity. Seon Ah Kom et al study, 2011 stated that maximum fibrinolytic enzyme production was obtained at 35- 45 0C. [40] The experimental results in the current study demonstrated comparable outcomes where the productivity of fibrinolytic enzyme from Bacillus cereus ATCC 43 42 was high at a range of temperature 35- 43 0C. Divalent cations such as Ca+2, Cu+2 and Mn+2 were found to be optimum cofactors for the productivity of FP during the present study. Hg+2, Zn+2 and Fe+3 were detected to inhibit the production of fibrinolytic enzymes in the current work. In the the present investigation the production of FP was detected to be high at PH 8- 10. The optimization media composition resulted in nearly 3 folds increase( 3.15 U/ ml) in fibrinolytic productivity when compared to the original media composition. The fibrinolytic enzyme yield obtained during the present study was higher than the results reported on fibrinolytic enzyme production via Bacillus vallisomritis ACE 02( in the previous study of Kim et al, 2007. [41] FP produced via bacterial recombinant DNA technology demonstrated more significant productivity and yield( nearly 15 folds increase) than the original initial non hybrid yield at near physiological temperature 37 0C and PH 7.4. The fibrinolytic enzyme secreted from Bacillus cereus ATCC 43 42 was shown to be extracellular in nature as noticed via the spectrophotometric method, so its purification and yield would be economically feasible. Recombinant FP was more active and stable in basic PH environments( PH 8-10). Similar results were found that Subtilisin DFE from Bacillus amyloliquefaciens DC-4 was active and stable at PH( 6-10). [42] The effect of plasminogen addition revealed that the test fibrinolytic enzyme was a plasminogen independent type and degraded the fibrin clot directly suggesting that recombinant FP a potent secure candidate for parenteral and oral administration.FP showed anticoagulant activity as demonstrated through APTT, TT, ELT and pro-thrombin time tests. The molecular mass of FP was estimated through mass spectrometer and SDS-PAGE gel filtration to be approximately 30 KDa which was higher than that of Nattokinase( 27.7 KDa) and Subtilisin CK ( 28.2 KDa) but smaller that that of Streptokinase( 47 KDa). [43] The molecular homology detection via Clustal Omega software and BLASTn analysis at NCBI website demonstrated that FP was approximately 70% similar to Subtilisin family especially Subtilisin DEF. The specific activity of recombinant FP was nearly 40 U/ mg; while it was 45 U/ mg for Subtilisin CK. [44] FP maintained the highest thermal stability at 55 0C for 1 hour. On the other hand, Tm of the fibrinolytic enzyme was observed at 58 0C. in another previous study Subtilisin DJ-4 showed thermal stability up to 60 0C for 60 minutes. Kinetic parameters Km,V max, Kcat of purified fibrinolytic enzyme FP in the present study were 5.78 mM, 100 µmol/ mg/ min and 2.41× 10( 2) M( -1) S( -1) respectively; while Km, Vmax and K cat of Subtilisin DJ-4 were 0.130mM, 1.89 U/ mg/ min and 3.87× 10( 4) M( -1) S( -1) respectively. [45] These data reflected that test FP possessed high affinity and catalytic activity for fibrin substrate. The in vitro and in vivo assays revealed that the enzyme could catalyze blood clot lysis effectively indicating that this enzyme could be useful as a fibrinolytic agent. The invitro release of FP parenteral SC formulation was up to 15 minutes. The percentage relative bioavailability of FP from optimized SC parenteral formulation reached 76 %. The biological half life of test enzyme was noticed to be about 90 minutes and its action lasted for nearly 6 hours. On the other hand the biological half life of Streptokinase[ indirect fibrinolytic drug which catalyzes the conversion of plasminogen into plasmin dissolving fibrin clots was about 80 minutes and its action lasts for approximately 5 hours. [46] The fibrinolytic enzyme activity of test recombinant FP was activated by monovalent cations such as Na+1 and K+1, while Fe+3 showed negligible effects on enzyme activity. On the other hand, EDTA inhibited FP fibrinolytic activity which was confirmed via Autodock Vina molecular docking software. Molecular modeling analysis using Swiss model software suggested that the 163 threonine residue of FP was located near the cation binding site; consequently tight binding of monovalent cations such as Na+1 and K+1 with FP occurred which probably increased thermostability and action of the test fibrinolytic enzyme. As well as, Molecular docking revealed that FP was analog to serine proteinase family type. BLASTn analysis of nucleotide sequence encoding FP revealed approximately 70 % homology with the gene encoding the fibrinolytic enzyme Subtilisin DFE. To avoid bleeding tendency risk, FP was not suggested to be administered with blood thinners such as heparin anticoagulants during the present study. I.M parenteral formulation was noticed to cause hematoma to rabbit animal models at the site of injection suggesting its exclusion in the usual dosage regimens. FP showed both anticoagulant and fibrinolytic activities during the present work suggesting the necessity to avoid its administration with heparin anticoagulant drug to minimize the risk of heavy bleeding. FP was detected in the current study to enhance bleeding tendency when was utilized with other blood thinner such as aspirin and warfarin. On the other hand, FP was observed to be metabolized proteolytically in the liver; however little amount was metabolized in the bile. FP was eliminated through the kidney after the termination of its fibrinolytic action within 300-360 minutes. In the present study, in vivo efficacy during randomized preclinical trials phase reached approximately 63% as demonstrated experimentally on rabbit animal models diseased previously with venous thrombi. The optimal dosage form for FP was found to be intermittent intravenous injection or deep subcutaneous injection. Figure 20 represents Phylogenetic tree of FP as a fibrinolytic agent generated via Clustal Omega multiple sequence alignment software. Molecular homology revealed that FP possesses more than 70 % similarity with Subtilisin DFE fibrinolytic agent. It consists of 275 amino-acids. Figure 18 displays the percentage of fibrinolysis of Fibrin-proteinase during a randomized preclinical trials stage. Figure 2 shows Positive Gram staining of bacterial Bacillus subtilis and cereus isolates producing thermostable fibrinolytic enzymes. Figure 1 shows fibrinolytic activity of the bacterial thrombolytic enzymes secreted from Bacillus SPP on nutrient agar plates inoculated with 3% fibrinogen solution. Figure 3 represents 3D structure of Fibrin-proteinase excreted from Bacillus cereus. The molecular mass reached nearly 30 KDa as determined via mass spectrometer. Figure 8 demonstrates the effects of various incubation temperatures on fibrinolytic enzyme productivity. Figure 4 demonstrates the SDS-PAGE purification of Fibrin-proteinase secreted from Bacillus cereus ATCC 43 42. The degree of purity reached approximately 80%. Figure 19 shows compatibility studies of FP and its excipients. There were no drug polymer interactions or incompatibilities. Figure 7 demonstrates the effects of different PH values on fibrinolytic enzymes productivity. Figure 5 shows growth of Bacillus cereus and Bacillus subtilis bacterial isolates on MFA. Figure 6 shows the effect of different inoculum volumes on FP productivity at PH 7.4, temperature 37℃ and 18 hours incubation time. Figure 12 demonstrates effect of temperature on fibrinolytic enzyme activity. Figure 9 represents effect of different carbon sources on the productivity of fibrinolytic enzymes. Figure 11 shows Michaelis Menten Plot. Figure 10 shows effect of different nitrogen sources on the productivity of fibrinolytic enzymes. Figure 13 demonstrates the effects of different PH values on the fibrinolytic enzyme action. Figure 17 represents fibrinolytic and proteolytic activities of test FP by casein digestion assay. Graph 15 represents a recombinant Fibrin-proteinase release profile. Graph 16 represents AUC of recombinant Fibrin- proteinase given through SC route of administration. Figure 14 demonstrates the effects of different metal ions and EDTA on fibrinolytic enzyme activity. The enzyme fibrinolytic action was inhibited via EDTA.
Figure 1.
It shows fibrinolytic activity of the bacterial thrombolytic enzymes secreted from Bacillus SPP on nutrient agar plates inoculated with 3% fibrinogen solution.
Figure 1.
It shows fibrinolytic activity of the bacterial thrombolytic enzymes secreted from Bacillus SPP on nutrient agar plates inoculated with 3% fibrinogen solution.

Figure 2.
It shows Positive Gram staining of bacterial Bacillus subtilis and cereus isolates producing thermostable fibrinolytic enzymes.
Figure 2.
It shows Positive Gram staining of bacterial Bacillus subtilis and cereus isolates producing thermostable fibrinolytic enzymes.
Figure 3.
It represents 3D structure of Fibrin-proteinase excreted from Bacillus cereus. The molecular mass reached nearly 30 KDa as determined via mass spectrometer.
Figure 3.
It represents 3D structure of Fibrin-proteinase excreted from Bacillus cereus. The molecular mass reached nearly 30 KDa as determined via mass spectrometer.
Figure 4.
It demonstrates the SDS-PAGE purification of Fibrin-proteinase secreted from Bacillus cereus ATCC 43 42. The degree of purity reached approximately 80%.
Figure 4.
It demonstrates the SDS-PAGE purification of Fibrin-proteinase secreted from Bacillus cereus ATCC 43 42. The degree of purity reached approximately 80%.
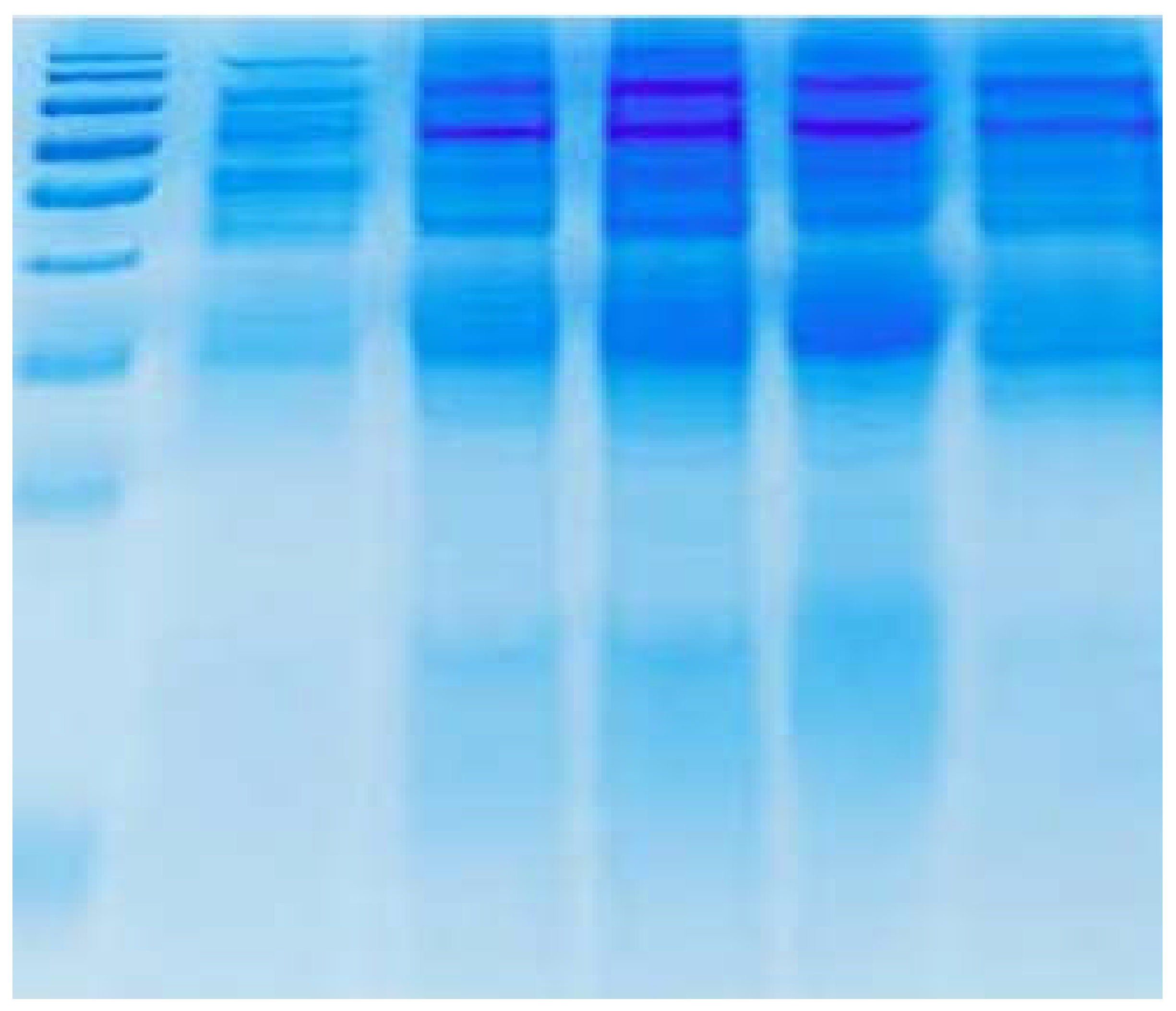
Figure 5.
It shows growth of Bacillus cereus and Bacillus subtilis bacterial isolates on MFA.
Figure 6.
It shows the effect of different inoculum volumes on FP productivity at PH 7.4, temperature 37℃ and 18 hours incubation time.
Figure 6.
It shows the effect of different inoculum volumes on FP productivity at PH 7.4, temperature 37℃ and 18 hours incubation time.
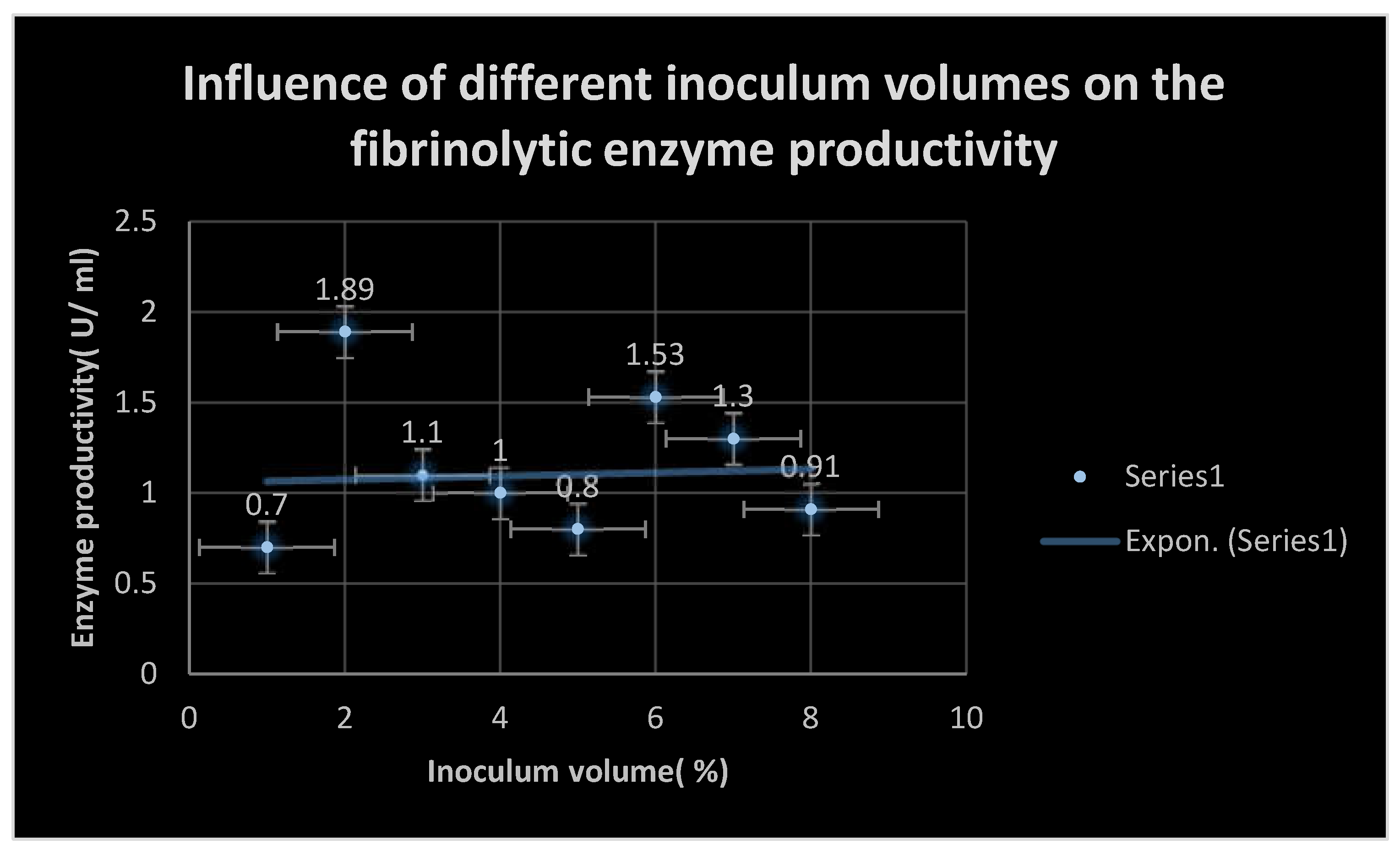
Figure 7.
It demonstrates the effects of different PH values on fibrinolytic enzymes productivity.
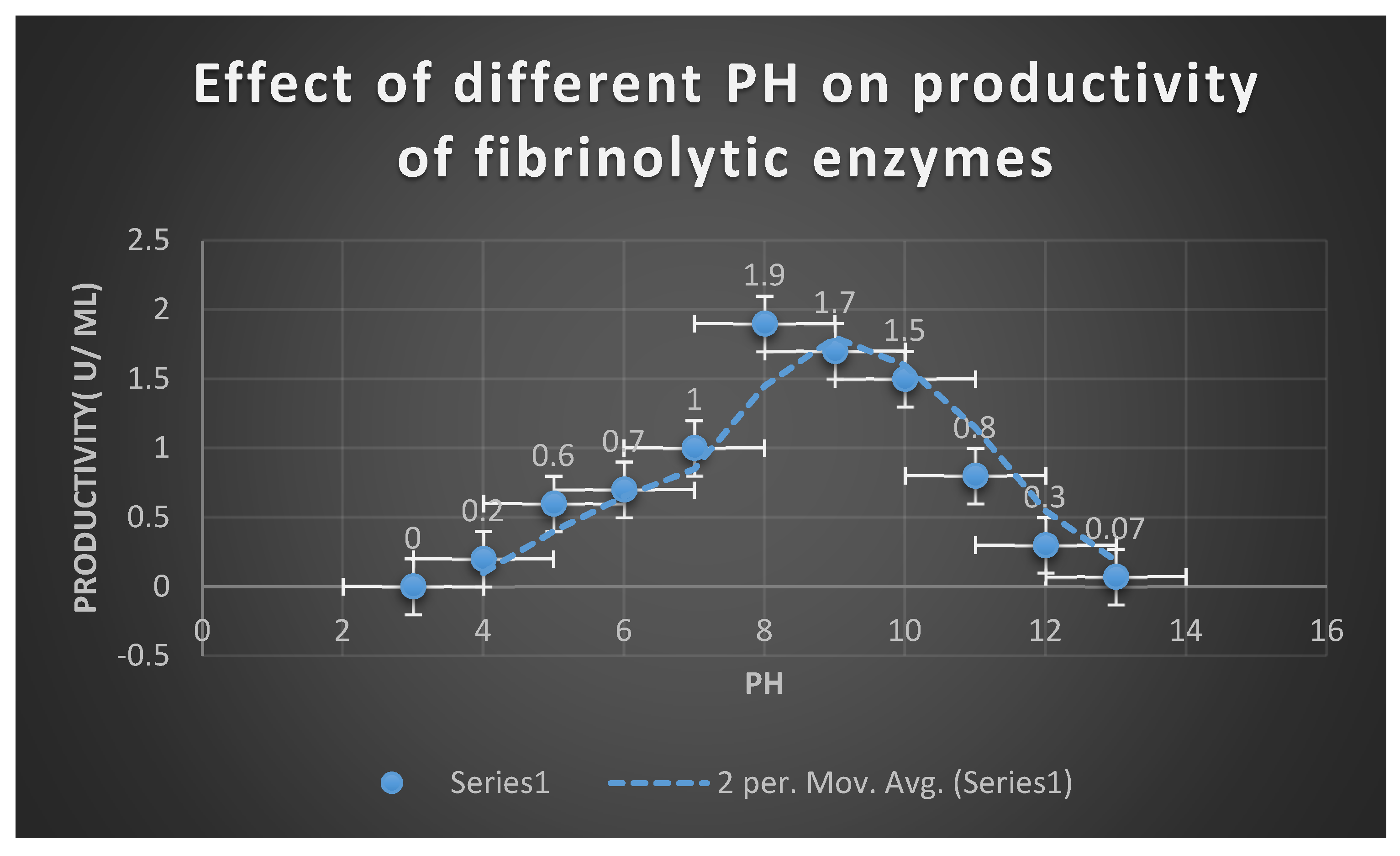
Figure 8.
It demonstrates the effects of various incubation temperatures on fibrinolytic enzyme productivity.
Figure 8.
It demonstrates the effects of various incubation temperatures on fibrinolytic enzyme productivity.
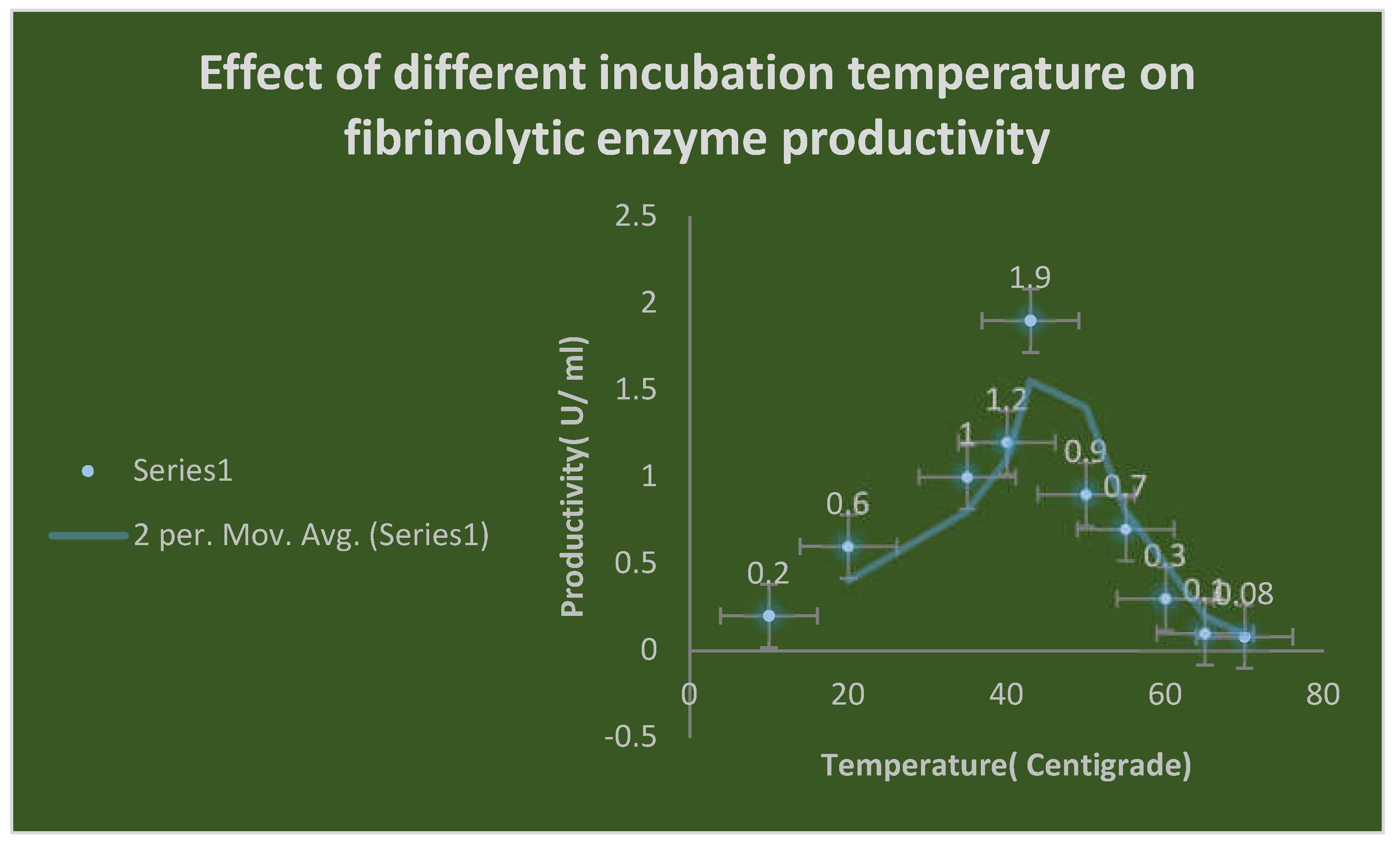
Figure 9.
Effect of different carbon sources on the productivity of fibrinolytic enzymes.
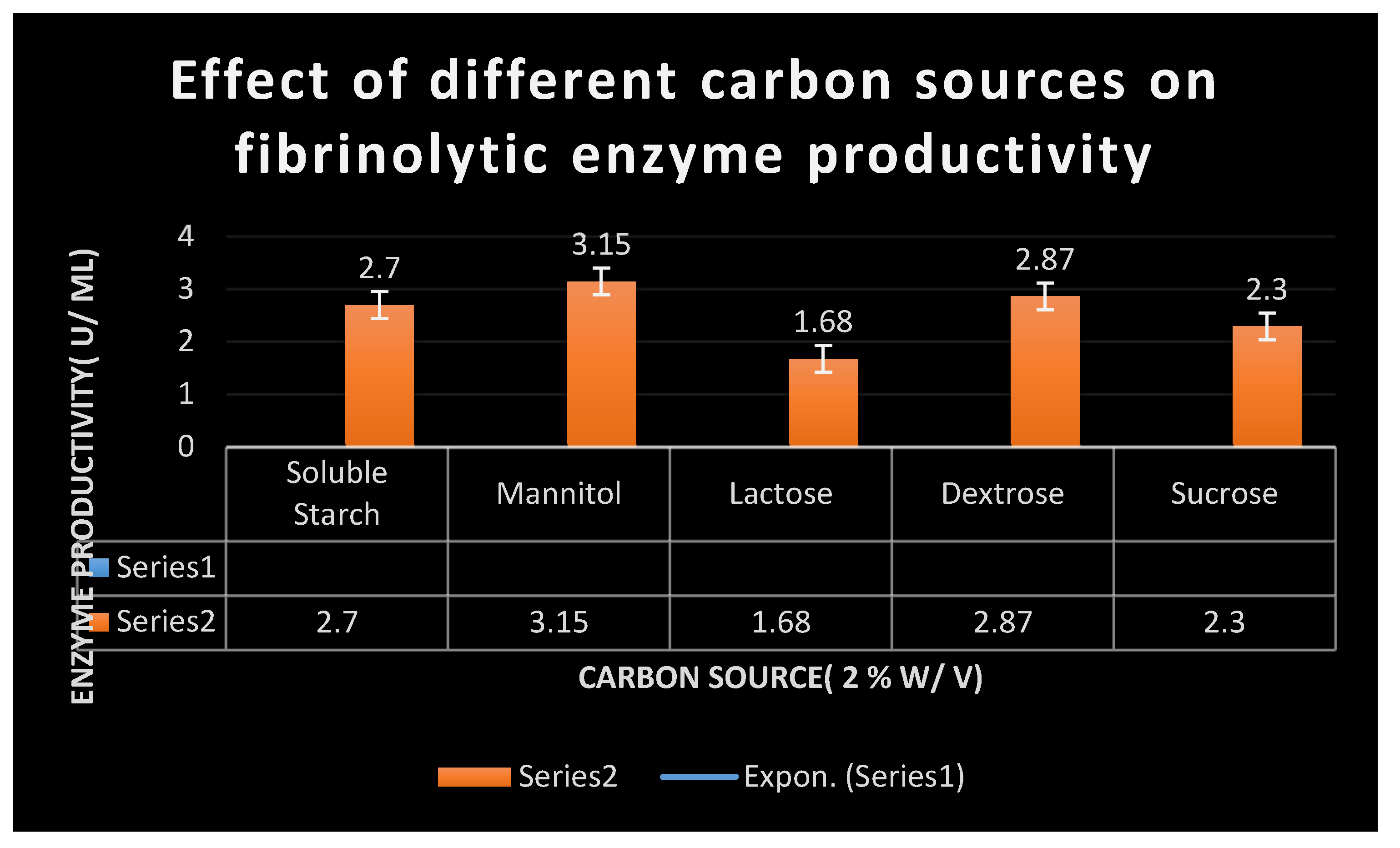
Figure 10.
Effect of different nitrogen sources on the productivity of fibrinolytic enzymes.
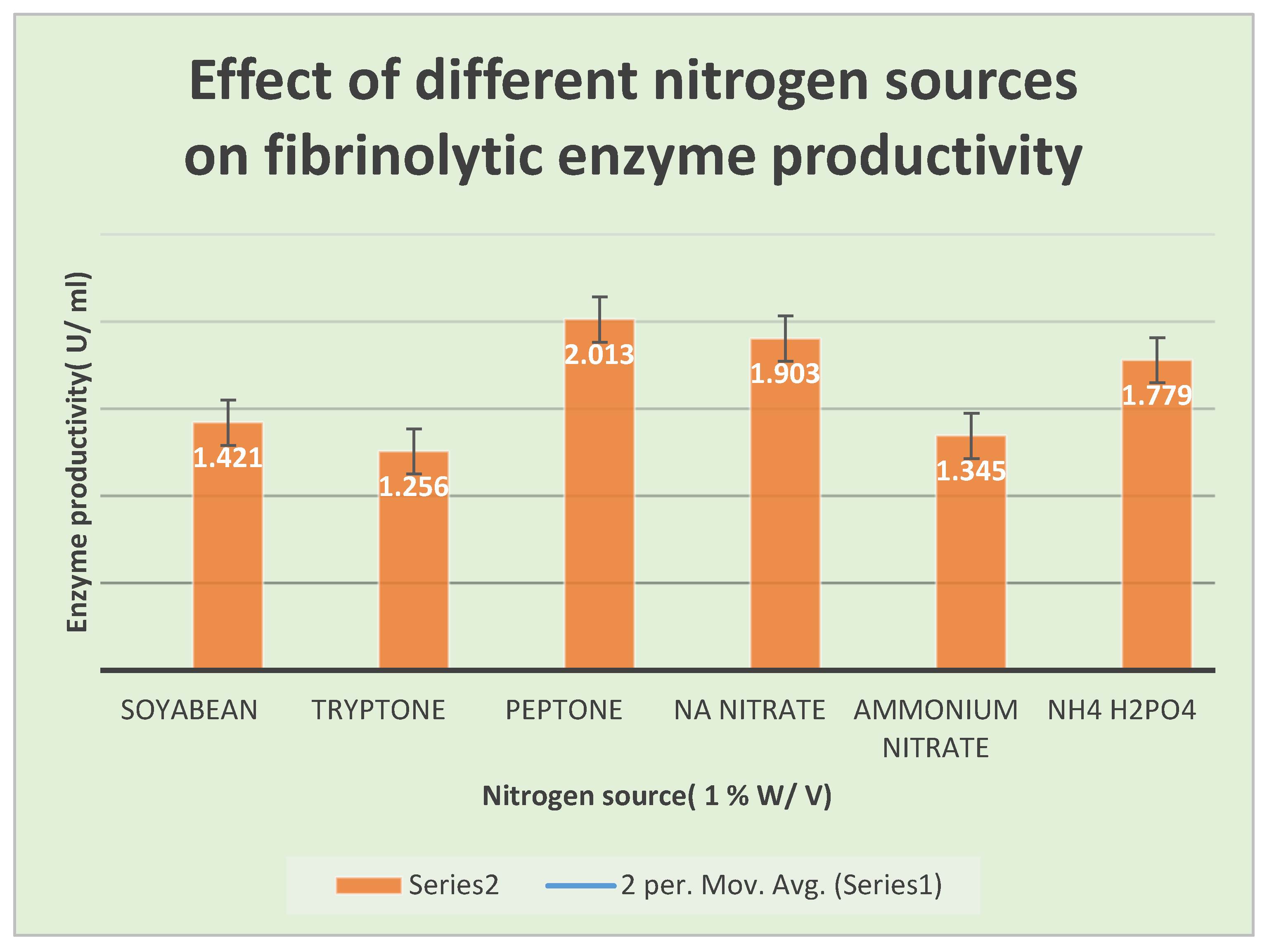
Figure 11.
It shows Michaelis Menten Plot.
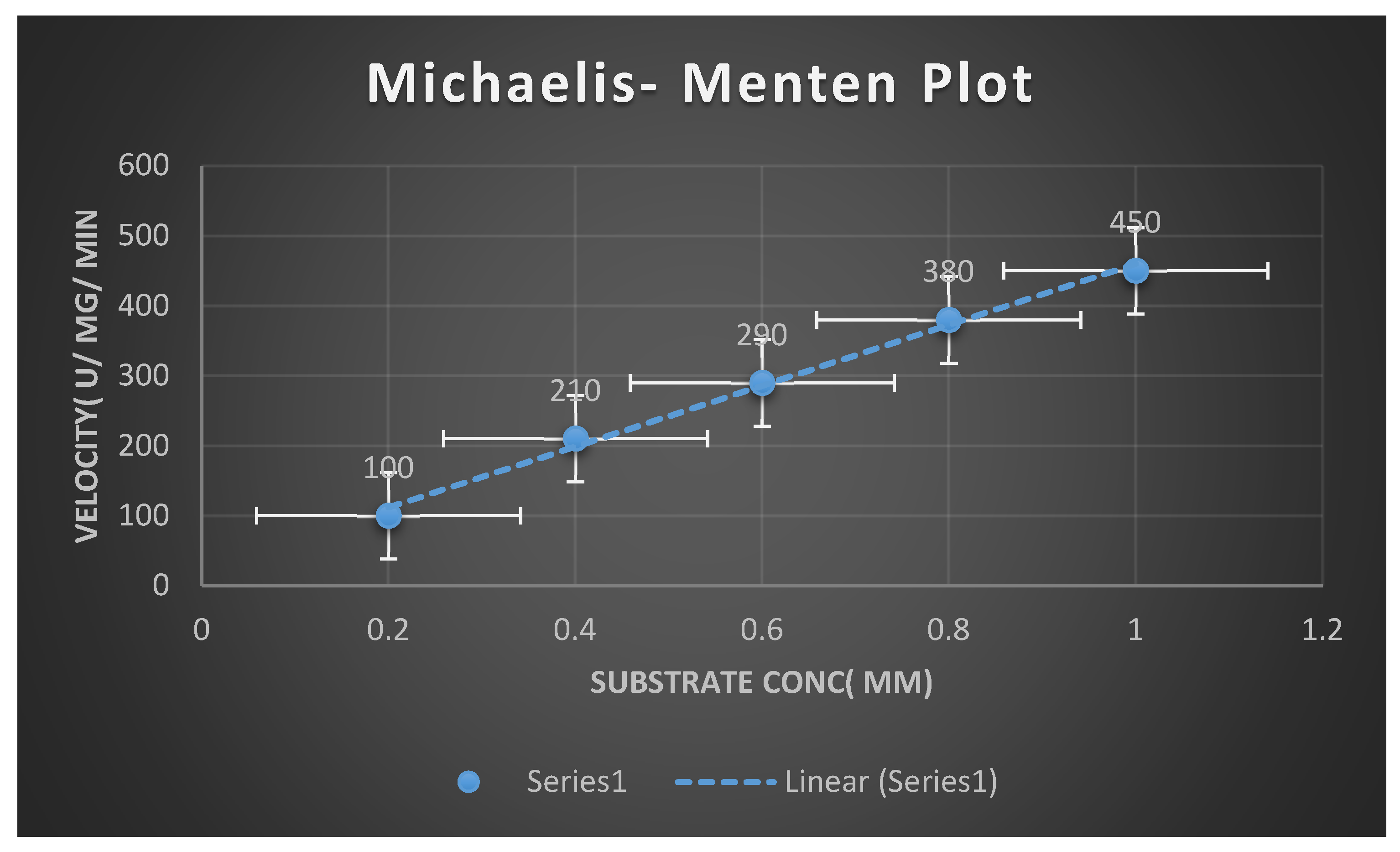
Figure 12.
Effect of temperature on fibrinolytic enzyme activity.
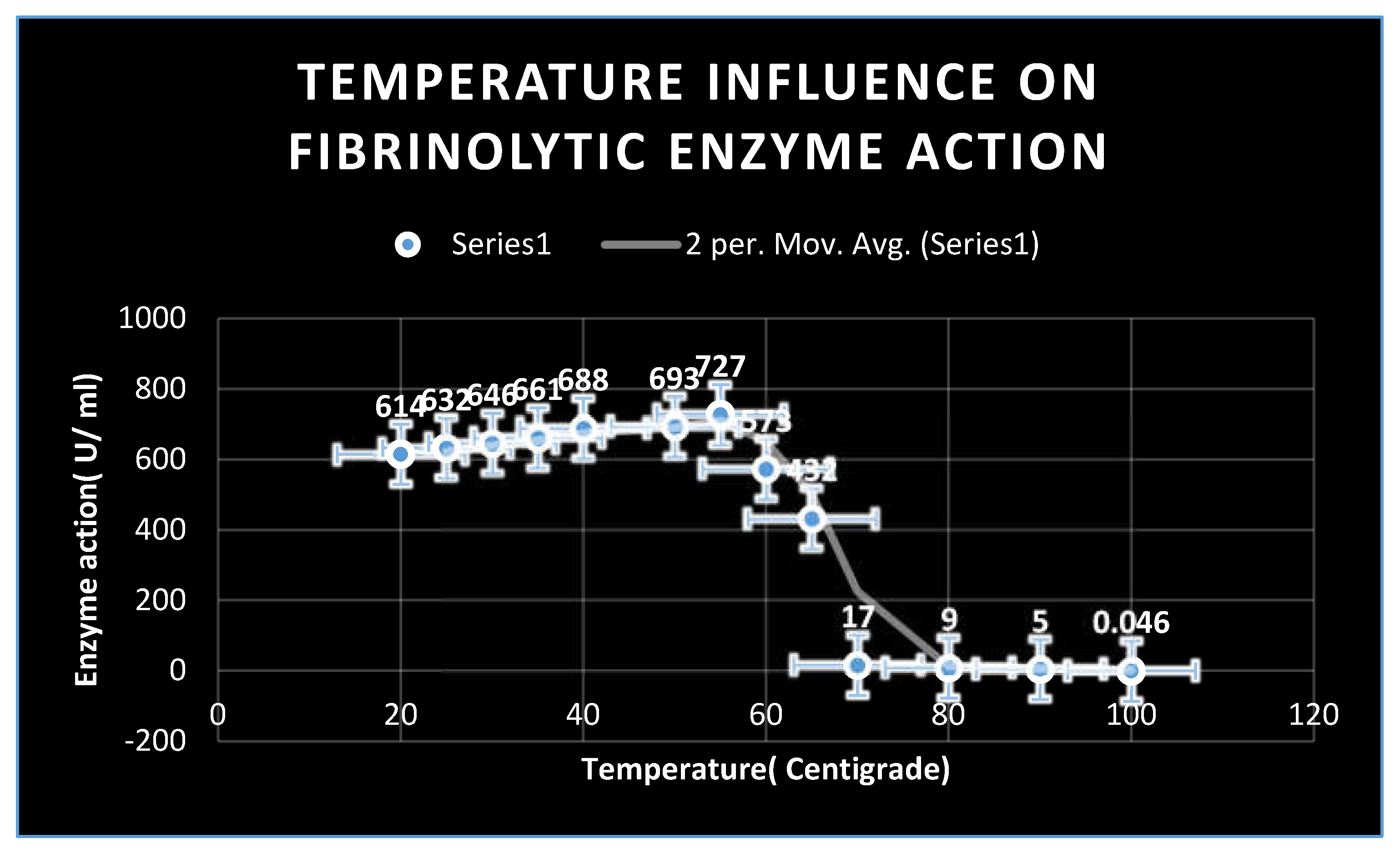
Figure 13.
It demonstrates the effects of different PH values on the fibrinolytic enzyme action.
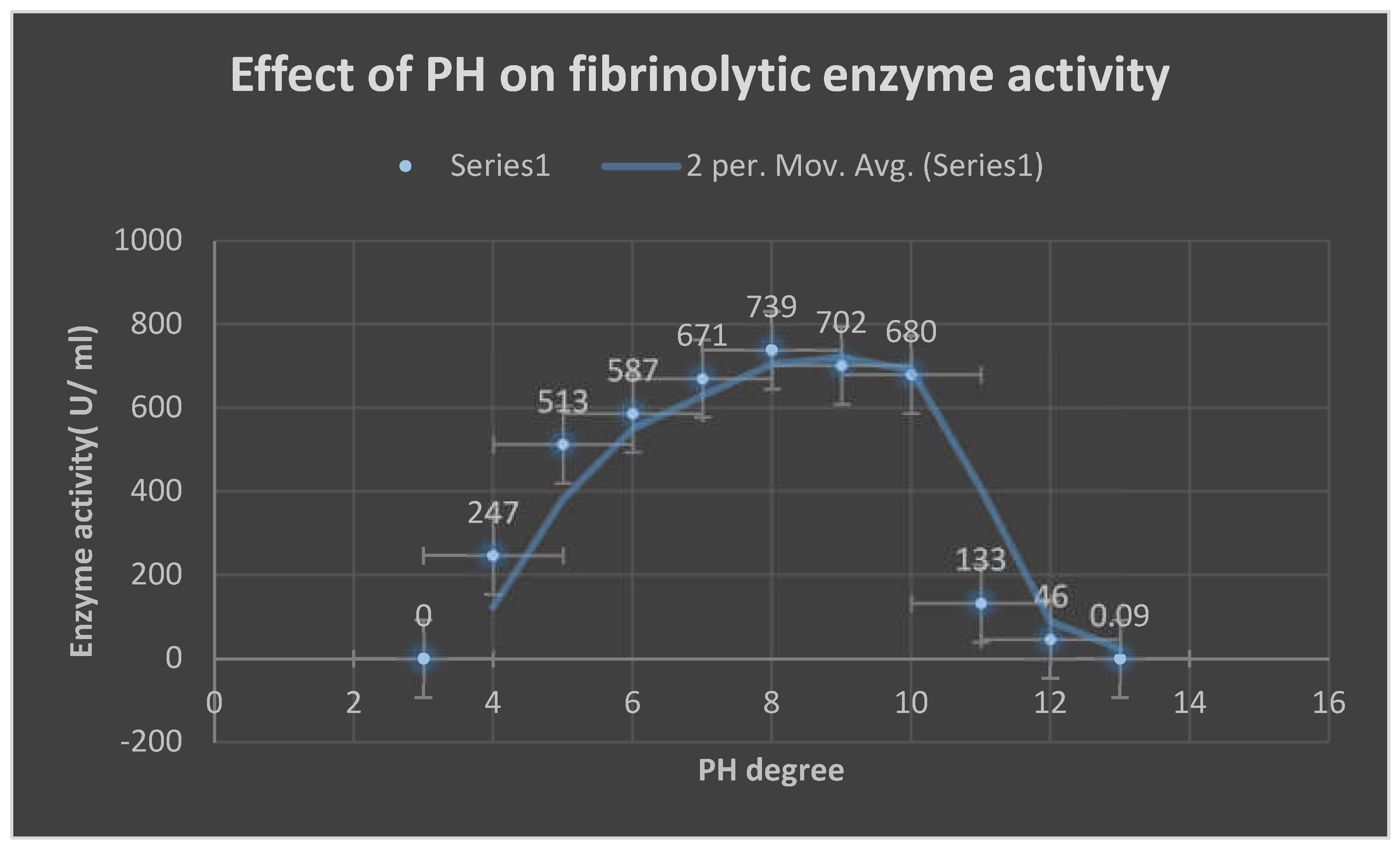
Figure 14.
It demonstrates the effects of different metal ions and EDTA on fibrinolytic enzyme activity. The enzyme fibrinolytic action was inhibited via EDTA.
Figure 14.
It demonstrates the effects of different metal ions and EDTA on fibrinolytic enzyme activity. The enzyme fibrinolytic action was inhibited via EDTA.
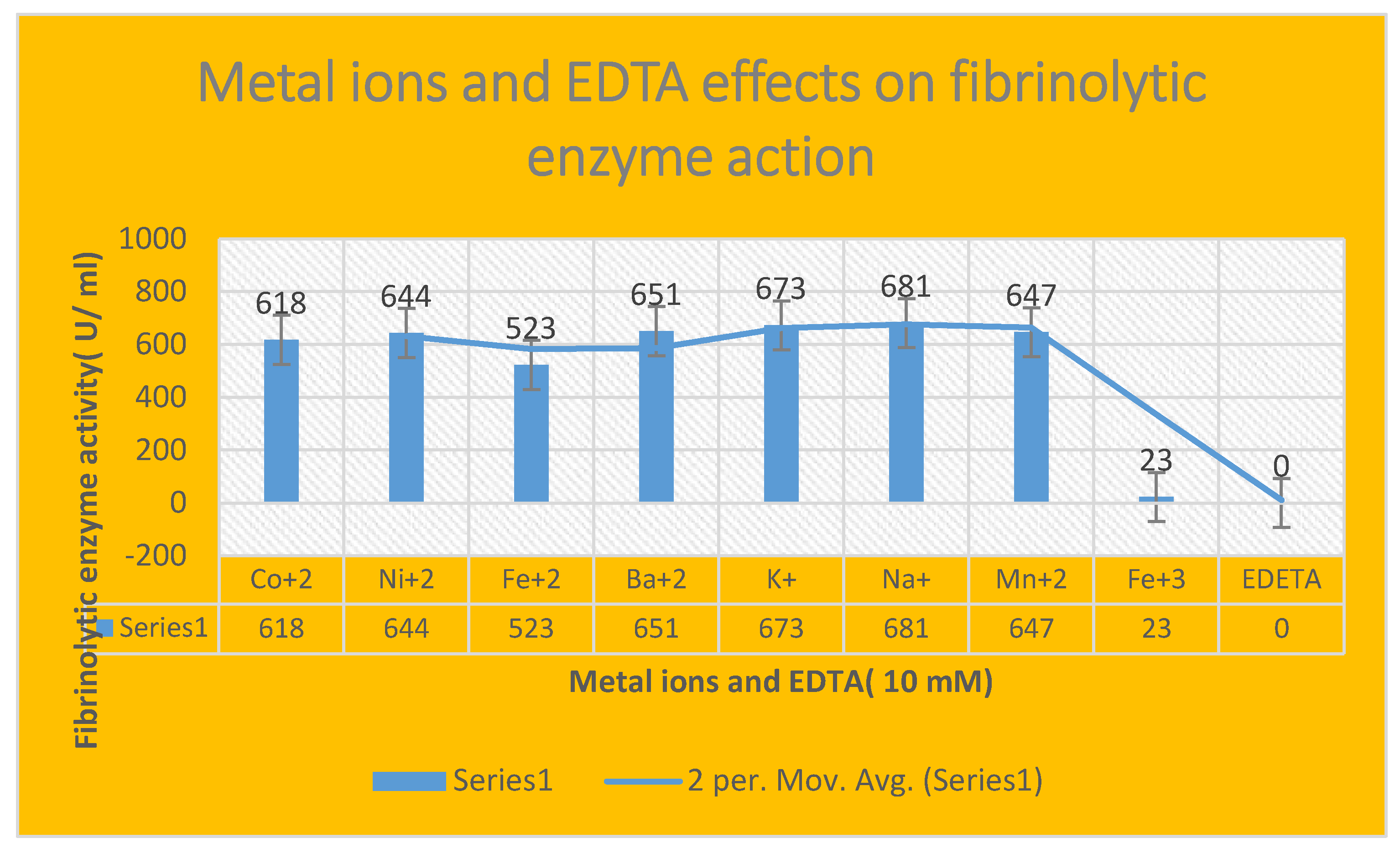
Figure 15.
It represents a recombinant Fibrin-proteinase release profile.
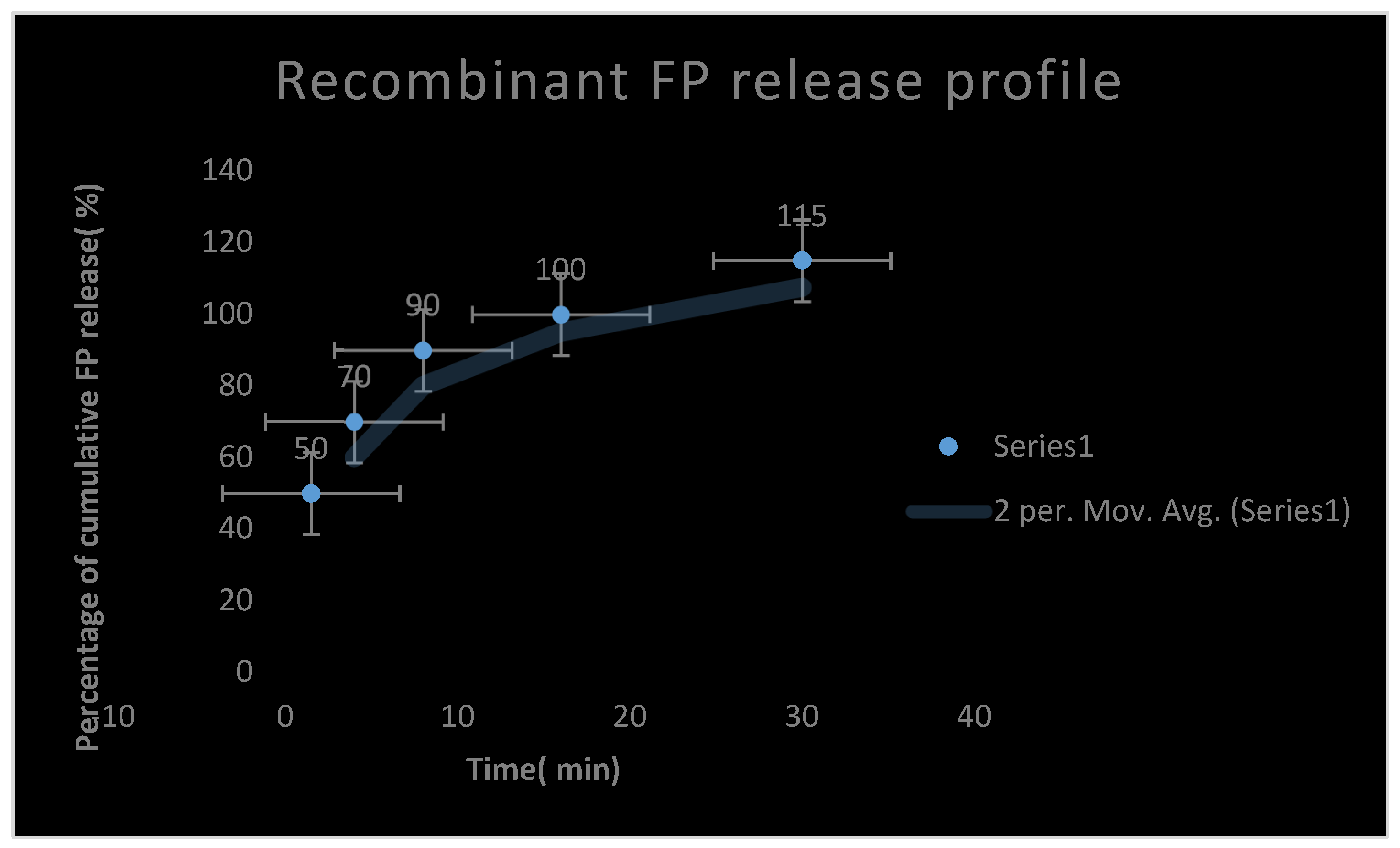
Figure 16.
It represents AUC of recombinant Fibrin- proteinase given through SC route of administration.
Figure 16.
It represents AUC of recombinant Fibrin- proteinase given through SC route of administration.
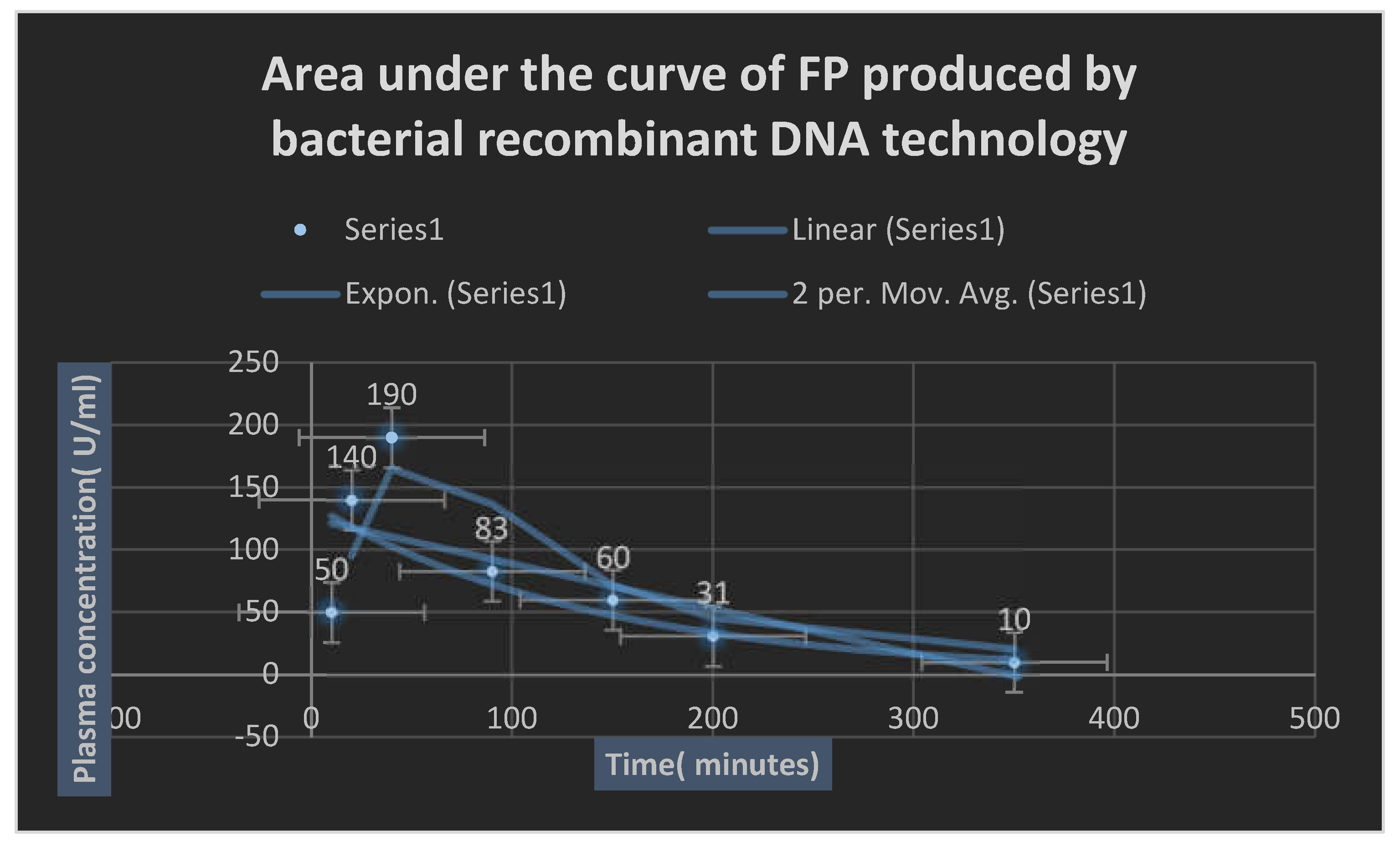
Figure 17.
It represents fibrinolytic and proteolytic activities of test FP by casein digestion assay.
Figure 17.
It represents fibrinolytic and proteolytic activities of test FP by casein digestion assay.
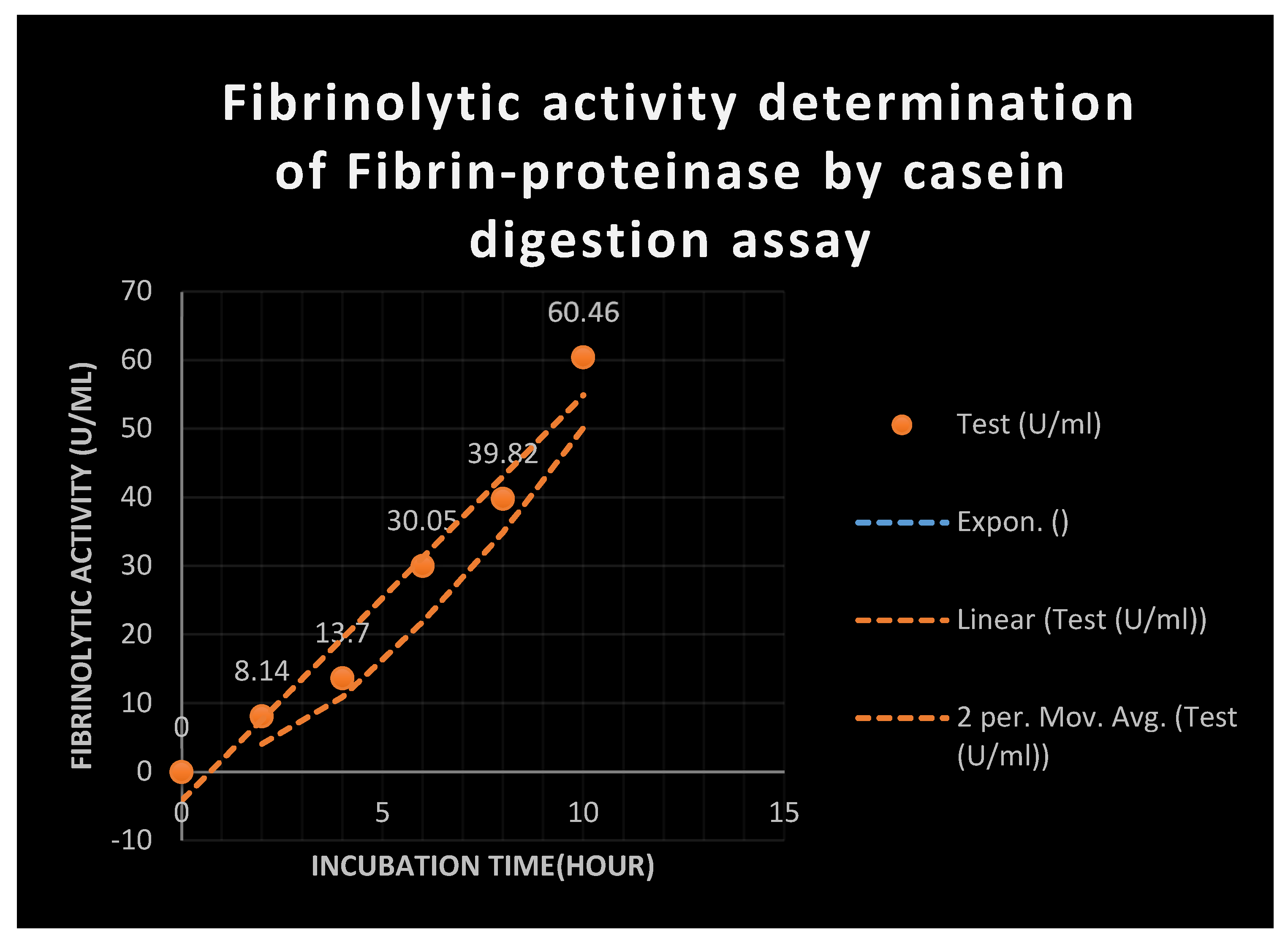
Figure 18.
Figure 18. It displays the percentage of fibrinolysis of Fibrin-proteinase during a randomized preclinical trials stage.
Figure 18.
Figure 18. It displays the percentage of fibrinolysis of Fibrin-proteinase during a randomized preclinical trials stage.
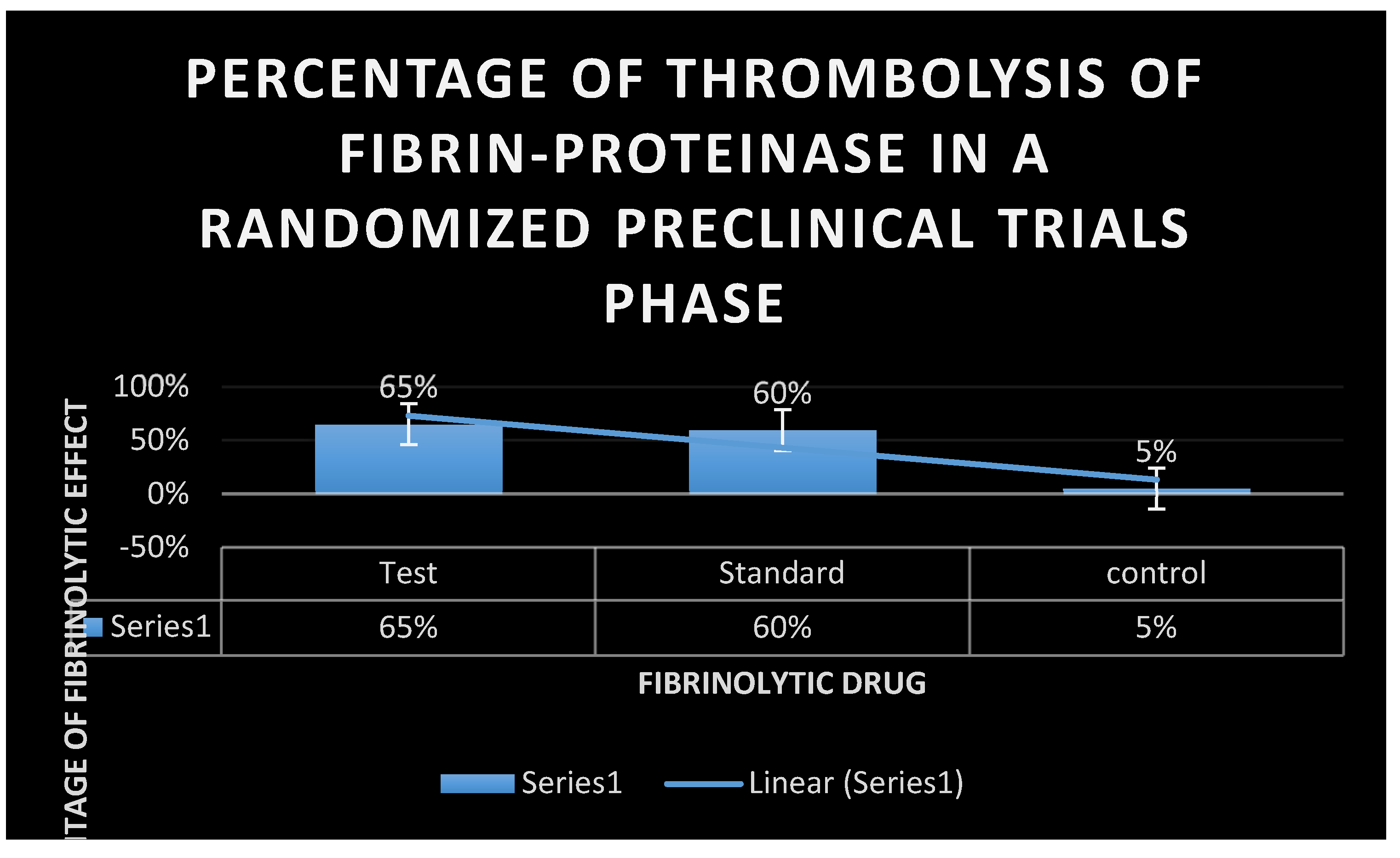
Figure 19.
It shows compatibility studies of FP and its excipients. There were no drug polymer interactions or incompatibilities.
Figure 19.
It shows compatibility studies of FP and its excipients. There were no drug polymer interactions or incompatibilities.
Figure 20.
It represents Phylogenetic tree of FP as a fibrinolytic agent generated via Clustal Omega multiple sequence alignment software. Molecular homology revealed that FP possesses more than 70 % similarity with Subtilisin DFE fibrinolytic agent. It consists of 275 amino-acids.
Figure 20.
It represents Phylogenetic tree of FP as a fibrinolytic agent generated via Clustal Omega multiple sequence alignment software. Molecular homology revealed that FP possesses more than 70 % similarity with Subtilisin DFE fibrinolytic agent. It consists of 275 amino-acids.

Conclusion
The new work is particularly promising since it uses bacterial recombinant DNA technology to manufacture FP with great fibrinolytic activity. It is suggested that additional study be conducted to ascertain the best time to use this fibrinolytic medication in order to dissolve different kinds of potentially lethal thrombi.
Funding
This study was funded by STDF.
Acknowledgement
STDF and Microbiology and Immunology department, Faculty of Pharmacy,Cairo University, Egypt are acknowledged for their support to the present study.
Conflict of interest
There is no conflict of interest.
References
- Bardal Stan,Waechter Jason, Martin Douglas( 2020). Applied pharmacology.V14: Chemotherapeutic drugs. Elsevier Edinburgh, London. 2020. 14[ 2]: 1178-1186.
- Caroline S, Zeind Michael G( 2018). Applied therapeutics, the clinical use of drugs.V23: Complications of End-Stage Liver Disease. Wolters Kluwer, London. 2018. 23[ 7]: 463-473.
- Dipro Cecily, Schwinghammer Terry, Dipro Joseph, Well Barbara ( 2021). Pharmacotherapy handbook.V13: Hepatic carcinoma. McGraw Hill Education, New York. 2021. 13( 9): 1582-1593.
- Fisher Bruce, Champe Pamela, Harvey Richard( 2021). Lippincott illustrated reviews microbiology. V6:Virology. Wolters Kluwer, London. 2021. 6( 8): 369-386.
- Golderg Stephen( 2020). Clinical physiology is made ridiculously simple.V6: Physiology of the liver. Med Master, Miami, United States of America. 2020. 6( 9): 2556-2578.
- Levinson Warren[ 2021]. Review of medical microbiology and immunology. V15: Immunology. McGraw Hill Education, New York. 2021. 15[ 13]: 937-953.
- Meeting Patricia J( 2019). Physiology.V16: Gastrointestinal physiology. McGraw Hill Education, New York. 2019. 16[ 1]: 1881-1895.
- Olson James( 2020). Clinical pharmacology made ridiculously simple. V7: Anticancer drugs. MedMaster, Miami, United States of America. 2020. 7[ 5]: 132-141.
- Parveen Kumar( 2017). Kumar, Clark's clinical medicine.V9: Oncology. Elsevier Edinburgh London. 2017. 9[ 20]: 692-703.
- Parveen Nadaf et al( 2019). Isolation, screening and characterization of L-glutaminase producing soil bacteria. International journal of pharmaceutical sciences and research. 2019; 23( 9):1874-1887.
- Trevor Anthony, Katzung Bertram, Kruidering-Hall Marieke( 2021). Katzung Trevor pharmacology examination board review.V13: Anticancer drugs. McGraw Hill Education, New York. 2021. 13[ 7]: 912-926.
- Swanson Larry N, Souney Paul F, Muntnick Alan H, Shargel Leon ( 2019). Comprehensive Pharmacy Review for NAPLEX.V10: Oncology. Wolters Kluwer, London. 2019. 10[ 13]: 1238-1250.
- Wilson Golder N( 2019). Biochemistry and genetics. V8: Gastroenterology and nutrition. McGraw Hill Education, New York. 2019; 8( 3): 763-771.
- Agrebi R et al( 2009). BSF1 fibrinolytic enzyme from a marine bacterium Bacillus subtilis A26: purification, biochemical and molecular characterization. Journal of process biochemistry. 2009; 44: 1252-1259.
- Bryan PN et al( 2000). Protein engineering of Subtilisin. Journal of biochemistry and physics. Acta; 2000; 1543: 203-222.
- Ghasemi Y et al( 2012). Cloning of a fibrinolytic enzyme( Subtilisin) gene from Bacillus subtilis in Escherichia coli. Journal of Mol. Biotechnol. 2012; 52: 1-7.
- Jaouadi B et al( 2010). Enhancement of thermostability and the catalytic efficiency of Bacillus pumilus CBS protease by site-directed mutagenesis. Journal of biochimie. 2010; 92: 360-369.
- Jeong S-J et al( 2007). Cloning of fibrinolytic enzyme gene from Bacillus subtilis isolated from cheonggukjang and its expression in protease deficient Bacillus subtilis strain. Microbiol. Biotechnol Journal. 2007; 17: 1018-1023.
- Jeong S-J et al( 2014). Over-expression of apr E2, a fibrinolytic enzyme gene from Bacillus subtilis CH3-5 in Escherichia coli and the properties of Apr E2. Journal of Microbiol. Biotechnol. 2014; 24: 969-978.
- Jo H-D et al( 2011). Cloning and over-expression of apr E3-17 encoding the major fibrinolytic protease of Bacillus licheniformis CH 3-17. Journal of Biotechnol. Bioproc. Eng. 2011; 16: 352-359.
- Afifah, D. N. , Sulchan, M., Syah, D., Yanti Suhartono, M. T., and Kim, J. H( 2014). Purification and characterization of a fibrinolytic enzyme from Bacillus pumilus 2.g isolated from Gembus, an Indonesian fermented food. Journal of Prev. Nutr. Food Sci. 2014; 19: 213–219.
- Agrebi, R., Haddar, A., Hajji, M., Frikha, F., Manni, L. et al( 2009) Fibrinolytic enzymes from a newly isolated marine bacterium Bacillus subtilis A26: characterization and statistical media optimization. Can. J. Microbiol. 2009; 55: 1049–1061.
- Bajaj, B. K., Sharma, N., and Singh, S( 2013). Enhanced production of fibrinolytic protease from Bacillus cereus NS-2 using cotton seed cake as nitrogen source. Biocatal. Agri. Biotechnol. 2013; 2: 204–209.
- Bajaj, B. K. , Singh, S., Khullar, M., Singh, K., and Bhardwaj, S ( 2014). Optimization of fibrinolytic protease production from Bacillus subtilis I-2 using agro-residues. Journal of Braz. Arch. Biol. Technol. 2014; 57: 653– 662.
- Balaraman, K. and Prabakaran, G( 2007). Production and purification of a fibrinolytic enzyme (thrombinase) from Bacillus sphaericus. Indian J. Med. Res. 2007; 126: 459–464.
- Collen, D. and Lijnen, H. R( 2004). Tissue-type plasminogen activator: a historical perspective and personal account. J. Thromb. Haemost.2004; 2: 541–546.
- Sweta, P., Rajpal, S. K., Jayant, Y. D., Hemant, J. P., Girdhar, M. T. et al( 2006). Development of an in vitro model to study clot lysis activity of thrombolytic drugs. Thromb. J.2006; 4: 1–4.
- Yong, P., Xiaojuan, Y., and Yizheng, Z( 2005). Microbial fibrinolytic enzymes: an overview of source, production, properties, and thrombolytic activity in vivo. Appl. Microbiol. Biotechnol. 2005; 69: 126–132.
- Verhamme, I. M. and Bock, P. E( 2014). Rapid binding of plasminogen to streptokinase in a catalytic complex reveals a three-step mechanism. J. Biol. Chem. 2014; 289: 28006–28018.
- Seo, J. H. and Lee, S. P( 2004). Production of fibrinolytic enzyme from soybean grits fermented by Bacillus firmus NA-1. J. Med. Food. 2004; 7: 442–449.
- Mukherjee, A. K. and Rai, S. K( 2011). A statistical approach for the enhanced production of alkaline protease showing fibrinolytic activity from a newly isolated gram-negative Bacillus sp. strain ASS20-I. New Biotechnol. 2011; 28: 182–189.
- Mihara, H., Sumi, H., Yoneta, T., Mizumoto, H., Ikeda, R. et al( 1991). A novel fibrinolytic enzyme extracted from the earthworm, Lumbricus rubellus. Jpn. J. Physiol. 1991; 41: 461–472.
- Markland, F. S. , Jr( 1998). Snake venom fibrinogenolytic and fibrinolytic enzymes: an updated inventory. Journal of Thromb Haemost. 1998; 79: 668– 674.
- Mahajan, P. M., Nayak, S., and Lele, S. S. (2012) Fibrinolytic enzyme from newly isolated marine Bacillus subtilis ICTF-1: media optimization, purification and characterization. J. Biosci. Bioeng. 2012; 113: 307–314.
- Liu, J., Xing, J., Chang, T., Ma, Z., and Liu, H( 2005). Optimization of nutritional conditions for nattokinase production by Bacillus natto NLSSE using statistical experimental methods. Process. Biochem. 2005 40: 2757–2763.
- Kotb, E( 2013). Activity assessment of microbial fibrinolytic enzymes. Journal of Appl. Microbiol. Biotechnol. 2013; 97: 6647–6665.
- Kim, J. B., Jung, W. H., Ryu, J. M., Lee, Y. J., Jung, J. K. et al( 2007). Identification of a fibrinolytic enzyme by Bacillus vallismortis and its potential as a bacteriolytic enzyme against Streptococcus mutans. Biotechnol. Lett.2007; 29: 605–610.
- Narasimhan M et al( 2015). Fibrinolytic enzyme production by newly isolated Bacillus cereus SRM-001with enhanced in-vitro blood clot lysis potential. Journal of applied microbiology. 2015; (61): 157-164.
- Sahbanu F et al( 2020). Molecular analysis of a fibrin degrading enzyme from Bacillus subtilis K2 isolated from the Indonesian soyabean-based fermented food moromi. Mol Biol Rep. 2020 Nov; 47( 11): 8553-8563.
- Kim A et al( 2011). Characterization and production of thermostable and acid stable fibrinolytic enzymes from Cordyceps militaris. International journal of entomology. 2011, 22( 2): 83-93.
- Yeo WS et al( 2011). Biochemical analysis of a fibrinolytic enzyme purified from Bacillus subtilis A1. Journal of Microbiology. 2011; 49: 376-380.
- Yin LJ et al( 2010). Bio-properties of potent Nattokinase from Bacillus subtilis YJ1. Journal of agricultural food chemistry. 2010; 58: 5737-5742.
- Sumi H et al( 1987). A novel fibrinolytic enzyme( Nattokinase) in the vegetable cheese natto; a typical and popular soybean food in the Japanese diet. Journal of experientia. 1987; 43: 1110-1111.
- Peng Y et al( 2004). Cloning and expression of a fibrinolytic enzyme( Subtilisin DFE) gene from Bacillus amyloliquefaciens DC-4 in Bacillus subtilis. Journal of Res. Microbiol. 2004; 155: 167-173.
- Peng Y et al( 2003). Purification and characterization of a fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from douchi, a traditional Chinese soybean food. Journal of comp. Biochem. Phys. 2003; B; 134: 45-52.
- Heo K et al( 2013). Characterization of a fibrinolytic enzyme secreted by Bacillus amyloliquefaciens CB1 and its gene cloning. Journal of Microbiology and Biotechnology. 2013; 23: 974-983.
Table 1.
Ingredients of Mineral Fibrinogen Agar.
| Ingredient | Unit of measurement |
|---|---|
| Potassium chloride | 0.5 mg |
| Magnesium sulfate | 0.5 mg |
| KH2PO4 | 1.0 g |
| Ferrous sulfate | 0.1 g |
| Zinc sulfate | 0.1 g |
| Fibrinogen | 2.0 g |
| Agar | 2% |
Table 2.
list of ingredients of blood agar.
| Ingredient | Unit of measurement |
|---|---|
| peptone | 10.0 g/ L |
| tryptose | 10.0 g/ L |
| Sodium chloride | 5.0 g/ L |
| Agar | 15.0 g/ L |
| Distilled water | 960 ml |
Table 3.
List of instruments.
| Instrument | Model and manufacturer |
|---|---|
| Autoclaves | Tomy, japan |
| Aerobic incubator | Sanyo, Japan |
| Digital balance | Mettler Toledo, Switzerland |
| Oven | Binder, Germany |
| Deep freezer -70 ℃ | Artiko |
| Refrigator 5 | whirpool |
| PH meter electrode | Mettler-toledo,UK |
| Deep freezer -20 ℃ | whirlpool |
| Gyratory shaker | Corning gyratory shaker, Japan |
| 190-1100nm Ultraviolet visible spectrophotometer | UV1600PC, China |
| Light(optical) microscope | Amscope 120X-1200X, China |
Table 5.
It displays the formulation of Fibrin-proteinase as a sterile solution at PH 7.
| Ingredient | concentration |
|---|---|
| Fibrin-proteinase | 50-100 IU |
| Mono-basic sodium phosphate | USP, 2 mg + 5% |
| Di-basic sodium phosphate | USP, 3 mg + 5% |
| Sodium chloride | USP, 5 mg + 5% |
| Water for injection | Query size to 1 ml |
Table 6.
It shows the positive bacterial isolates distribution on MFA.
| No of +ve bacterial isolates | No of -ve bacterial isolates |
| 31 | 19 |
Table 7.
Distribution of the main producing fibrinolytic bacterial bacilli on MFA.
| No. of Bacillus cereus isolates | No. of Bacillus subtilis isolates |
| 17 | 14 |
Table 8.
It demonstrates the outcomes of different carbon sources on productivity of fibrinolytic enzymes.
Table 8.
It demonstrates the outcomes of different carbon sources on productivity of fibrinolytic enzymes.
| Carbon source( 2% W/ V) | Productivity( U/ml) |
|---|---|
| Soluble starch | 2.7 |
| Mannitol | 3.15 |
| Lactose | 1.68 |
| Dextrose | 2.87 |
| Sucrose | 2.3 |
Table 9.
It shows the influence of different inoculum volumes on the production of fibrinolytic enzymes at incubation conditions pH 7.4, temperature 37 ℃ and 18 hours incubation period.
Table 9.
It shows the influence of different inoculum volumes on the production of fibrinolytic enzymes at incubation conditions pH 7.4, temperature 37 ℃ and 18 hours incubation period.
| Inoculum volume( %) | Fibrinolytic enzyme productivity( U/ ml) |
|---|---|
| 1 | 0.76 |
| 2 | 1.89 |
| 3 | 1.12 |
| 4 | 1.00 |
| 5 | 0.84 |
| 6 | 1.53 |
| 7 | 1.33 |
| 8 | 0.91 |
Table 10.
It shows the effects of different nitrogen sources on FP productivity.
| Nitrogen source( 1% W/ V) | FP productivity( U/ml) |
|---|---|
| Soyabean | 1.421 |
| Tryptone | 1.256 |
| Peptone | 2.013 |
| Na Nitrate | 1.903 |
| Ammonium Nitrate | 1.345 |
| Ammonium Dihydrogen Phosphate | 1.779 |
Table 11.
It demonstrates the effects of different metal ions on the fibrinolytic enzyme productivity.
Table 11.
It demonstrates the effects of different metal ions on the fibrinolytic enzyme productivity.
| Metal ion( 10 mM) | Productivity( U/ ml) |
|---|---|
| Mg+2 | 1.8 |
| Zn+2 | 0 |
| Hg+2 | 0 |
| CU+2 | 2.1 |
| Fe+2 | 1.8 |
| Ca+2 | 2.4 |
| Mn+2 | 2.3 |
| Fe+3 | 0 |
Table 12.
It shows the cellular position of the fibrinolytic enzymes.
| Cellular position | Percentage( %) |
|---|---|
| Extracellular | 90 |
| Intracellular | 9 |
| Surface bound | 1 |
Table 13.
It demonstrates the effects of different pH degrees on FP production.
| pH | FP productivity( U/ ml) |
|---|---|
| 3 | 0 |
| 4 | 0.2 |
| 5 | 0.6 |
| 6 | 0.7 |
| 7 | 1 |
| 8 | 1.89 |
| 9 | 1.7 |
| 10 | 1.5 |
| 11 | 0.8 |
| 12 | 0.4 |
| 13 | 0.1 |
Table 14.
It demonstrates the effects of different temperatures degrees on FP production.
| T ( ℃) | FP productivity( U/ ml) |
|---|---|
| 10 | 0.2 |
| 20 | 0.6 |
| 35 | 1 |
| 40 | 1.2 |
| 43 | 1.9 |
| 50 | 0.9 |
| 55 | 0.7 |
| 60 | 0.3 |
| 65 | 0.1 |
| 70 | 0.0 |
Table 15.
It demonstrates Michaelis Menten Plot.
| Substrate concentration( mM) | Velocity( U/ mg/ min) |
|---|---|
| 0.2 | 100 |
| 0.4 | 210 |
| 0.6 | 290 |
| 0.8 | 380 |
| 1 | 450 |
Table 16.
It represents AUC of recombinant Fibrin-proteinase given through SC injection:.
| Time( minute) | plasma concentration( U/ml) |
|---|---|
| 10 | 50 |
| 20 | 140 |
| 40 | 190 |
| 90 | 83 |
| 150 | 60 |
| 200 | 31 |
| 350 | 10 |
Table 17.
It represents fibrinolytic activity of test thermostable FP by Casein digestion assay.
| Incubation time( hr) | Test( U/ml) | Standard( U/ml) |
|---|---|---|
| 0 | 0 | 0 |
| 2 | 8.14 | 7.02 |
| 4 | 13.7 | 14.07 |
| 6 | 30.05 | 28.44 |
| 8 | 39.82 | 37.29 |
| 10 | 60.46 | 58.51 |
Table 18.
It represents Fibrin-proteinase release profile.
| Time( min) | Percentage of release( %) |
|---|---|
| 1.5 | 50 |
| 4 | 70 |
| 8 | 90 |
| 16 | 100 |
| 30 | 115 |
Table 19.
It represents the effects of different pH values on FP fibrinolytic activity:.
| pH | Enzyme activity( U/ ml) |
|---|---|
| 3 | 0 |
| 4 | 247 |
| 5 | 513 |
| 6 | 587 |
| 7 | 671 |
| 8 | 739 |
| 9 | 702 |
| 10 | 680 |
| 11 | 133 |
| 12 | 46 |
| 13 | 0.09 |
Table 20.
It represents the effects of various temperatures on the fibrinolytic activity of Fibrin-proteinase.
Table 20.
It represents the effects of various temperatures on the fibrinolytic activity of Fibrin-proteinase.
| T (℃) | Enzyme activity( U/ ml) |
|---|---|
| 20 | 614 |
| 25 | 632 |
| 30 | 646 |
| 35 | 661 |
| 40 | 688 |
| 50 | 693 |
| 55 | 727 |
| 60 | 573 |
| 65 | 432 |
| 70 | 17 |
| 80 | 9 |
| 90 | 5 |
| 100 | 0.046 |
Table 21.
It represents the effects of different metal ions and inhibitors on fibrinolytic activity of Fibrin-proteinase.
Table 21.
It represents the effects of different metal ions and inhibitors on fibrinolytic activity of Fibrin-proteinase.
| Metal ion( 10mM) | Fibrinolytic enzyme activity( U/ ml) |
|---|---|
| Co+2 | 618 |
| Ni+2 | 644 |
| Fe+2 | 523 |
| Ba+2 | 651 |
| K+ | 673 |
| Na+ | 681 |
| Mn+2 | 647 |
| Fe+3 | 23 |
| EDETA | 0 |
Table 22.
It shows the pharmacokinetics profile of Fibrin-proteinase.
| Biological half life t1/2( min) | Total clearance( ml/ min) | Duration of action( min) | Onset time( min) | Apparent volume distribution( L) | Peak reach time( min) |
|---|---|---|---|---|---|
| 85-90 min | 11.6± 7.1 ml/ min | 300-360 min | 15-30 min | 2.8± 0.69 L | Within 60 minutes |
Table 24.
It demonstrates the biochemical reactions of the predominating bacterial isolates producing FP.
Table 24.
It demonstrates the biochemical reactions of the predominating bacterial isolates producing FP.
| Test | Result |
|---|---|
| Gram stain | + |
| Cell shape | Rod |
| Spore shape | Ellipsoidal |
| Spore site | Central |
| Motility | + |
| Catalase | + |
| Oxidase | + |
| Blood haemolysis | Beta haemolysis |
| Indol | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



