
Submitted:
14 April 2024
Posted:
15 April 2024
You are already at the latest version
Abstract
(1) Background: Grapes and their associated by-products (as grape pomace, GP) stand out for their polyphenol content, which makes them a source of bioactive compounds with antioxidant capacity. The aim of this research was to determine if the inclusion of 50 g/kg of GP in the diet of hens could enrich egg with antioxidants and to study its effect on internal and external egg quality parameters. (2) Methods: A trial was conducted with two genetic lines of hens, which were fed either a control diet or a diet containing 50 g/kg of GP. Performance, internal and external egg quality, and egg yolk content of vitamins E and A, and gallic acid were determined; (3) Results: In eggs laid by hens fed GP diet, Haugh units and yolk colour score were enhanced, and eggshell became thinner, but without affecting the breaking strength. No dietary effect was observed on vitamin contents in yolk. Higher gallic acid content was observed in yolk of eggs laid by hens fed the GP diet, suggesting that some dietary phenolic compounds could be transferred to the eggs. Hen genetics influenced egg weight, albumen Haugh units, shell thickness and α- and γ-tocopherol concentration in yolks; (4) Conclusions: Dietary inclusion of GP improved the internal quality of eggs, enriching yolks with a phenolic compound, but reduced shell thickness.
Keywords:
grape byproducts
; polyphenols
; antioxidants
; egg quality
; gallic acid
; vitamins
; poultry
; laying hen
1. Introduction
The increase in food demand driven by population growth, coupled with the reduction in the supply of raw materials owing to factors such as climate change, poses a significant challenge for the agricultural and livestock sectors. Among the different processes associated with egg production, the feeding of hens represents the largest source of environmental impact [1]. Nonetheless, the revaluation of agri-food by-products through their incorporation into animal diets represents a promising strategy in order to reduce the environmental impact derived from waste generation and disposal, as well as a way to contribute to ensuring sustainable consumption and production patterns and to reducing nutrient loss, as outlined in some of the Sustainable Development Goals (specifically, targets 2.4 and 12.5) of the 2030 Agenda. Certain by-products have a high content in antioxidant compounds and could contribute to improving animal health and the quality of animal products. Among these, by-products originating from the winemaking process, such as grape pomace (GP), are a rich source of natural antioxidants like polyphenols. The GP consists of a mixture of stems, pulp and seeds in varying proportions. Spain holds a significant position in the international wine economy, leading to a substantial availability of these grape by-products, which have the potential to be used as ingredients in animal nutrition [2]. In recent years, some authors have demonstrated the antioxidant capacity of these by-products in poultry [3,4], their prebiotic effect [5], as well as their potential to improve meat quality [6,7]. Furthermore, the identification of phenolic metabolites in the intestinal content and plasma of chickens that had consumed grape by-products [8,9] suggests that these compounds could be transferred to tissues where they may exert their antioxidant effect. Although less research has been conducted with laying hens, it has also been shown [10] that the inclusion of GP in the diet of hens can modify egg composition (e.g. yolk fatty acid profile, by reducing the proportion of saturated fatty acids while increasing that of polyunsaturated ones) and can improve the oxidative stability of yolk lipids during refrigerated storage. Moreover, recent research works have demonstrated that it is possible to reduce oxidation and to increase egg fertility by incorporating grape polyphenols into the diet of hens [11,12]. Additionally, phenolic metabolites have been identified in the plasma of hens after feeding the latter with diets containing grape seed extract [13]. These results suggest that certain grape polyphenols could also transfer to the egg and exert their antioxidant action there. However, there is no information demonstrating whether dietary grape polyphenols and/or their metabolites can be deposited in the egg after being metabolized by laying hens and therefore, whether it is possible to enrich eggs with these antioxidant compounds. Due to the efficiency of laying hens in depositing nutrients from the diet into the egg, incorporating bioactive compounds into laying hens’ diets could be a successful strategy to enhance the nutraceutical value of eggs. Actually, it has been proven that when antioxidants are included in the feed of laying hens, the metabolism of hens allocate these dietary antioxidants preferentially in the egg instead of in the body [14].
On the other hand, several authors have highlighted the ability of dietary grape polyphenols to increase the content of vitamin E in plasma and tissues in birds [4,6,7,9,15]. Polyphenols, similarly to vitamin C, can interact with radicals generated during the metabolism of vitamin E, thereby recycling alpha-tocopherol in plasma and tissues. Furthermore, it has been demonstrated that it is possible to increase the content of vitamin E in eggs by means of hens’ diet [16,17], making it interesting to study whether the dietary inclusion of GP would also allow an increase in the deposition of vitamin E in egg yolk. Likewise, considering the potential interaction in intestinal absorption between fat-soluble vitamins [18], modifications in the deposition of vitamin E could affect the content of vitamin A in egg yolk.
Thus, the aim of this study was to determine if the inclusion of 50 g/kg of GP in hens’ diet might produce enrichment in phenolic compounds in the egg and to study the effect of this dietary inclusion on internal and external egg quality parameters and on α- and γ-tocopherol and retinol contents in egg yolk.
2. Materials and Methods
2.1. Standards and the Tested Product
All chemical used were HPLC analytical grade and the water ultrapure. Gallic acid, catechin, cyanidin 3-O-glucoside and quercetin 3-O-glucoside, α-tocopherol, γ-tocopherol and retinol were purchased from Sigma Chemical Co (St. Louis, MO). Methanol, ethanol and hexane were obtained from Panreac (Castellar del Vallés, Barcelona, Spain).
The grape by-product (whole pomace), consisting of a mixture of seeds and skins, was obtained fresh, immediately after racking (following 15 days of fermentation) and mechanical separation of stems. Grapes originated from September 2021 harvest season and were obtained at Roquesán Cooperative Winery (Quemada, Burgos, Spain). The GP was dried in an oven at 50 °C for 4 days and subsequently stored in darkness. Thereafter, GP was ground with a 1-mm screen and incorporated to the experimental diet. The nutritional composition and the phenolic profile of GP are reported in Table 1 and Table 2, respectively.
2.2. Birds and Diets
This trial was conducted at the experimental facilities of the Faculty of Veterinary Science of the Complutense University of Madrid (Spain). Thirty ISA White and thirty ISA Brown laying hens were used. All hens were 22 weeks old at the beginning of the experiment. Hens of each genetic line were allocated randomly to 6 wire cages (5 hens per cage) in a completely enclosed fan-ventilated and environmentally controlled building with a daily light programme of 16 h (photoperiod from 06:00 am to 10:00 pm). Cages used in this research work were enriched cages fulfilling the requirements set by the Council Directive 1999/74/EC (750 cm2 of cage area per hen, more than 0.20 m of height at any point of the cage, at least 0.15 m of perch per hen, at least 0.12 m of feeder length per hen, a nest within the cage, etc.). Experimental procedures were approved by the Complutense University of Madrid Animal Care and Ethics Committee (protocol code NP0926052022-2022) in compliance with the guidelines for the Care and Use of Animals for Scientific Purposes of the Ministry of Agriculture, Fishery and Food.
Hens of each genetic line were fed either a control diet or a diet including GP at 50 g/kg (3 replicate cages of each genetic line per diet). Ingredients and nutrient composition of the two experimental diets are shown in Table 3. Diets were formulated to be isocaloric, isonitrogenous and isofibrous and to contain the same amount of calcium and phosphorus. To make diets be isofibrous, straw was included in the control diet. Diets were exempt from colourants and antioxidants. Feed, offered in mash form, and water were provided ad libitum throughout the whole experiment.
Thus, the present trial considered the following four treatments arranged in a factorial 2 x 2 design: (1) ISA White hens – control diet; (2) ISA White hens – GP 50 g/kg diet; (3) ISA Brown hens – control diet; (4) ISA Brown hens – GP 50 g/kg diet.
2.3. Hen Performance Assessment and Egg Collection
After a two-week adaptation period, laying hen performance was assessed for four weeks (from week 24 to week 28 of age). Feed intake by laying hens was recorded weekly and then divided by the number of birds per cage and by seven in order to determine the daily feed intake per hen. All laid eggs were collected daily and the number of eggs obtained in each cage was written down. All eggs were then weighed. Daily egg mass was calculated per cage by multiplying daily egg production by average egg weight divided by 100. Feed conversion ratio was calculated by dividing feed intake by egg mass.
On the first and second day of week four of the trial (week 27 of age of hens), 9 freshly laid eggs were collected daily per treatment (daily, 3 eggs per replicate of each of the four treatments), individually weighed and used for determination of shell proportion, shell thickness, albumen Haugh units and egg yolk colour.
On the third and fourth day of week four of the trial (week 27 of age of hens), 12 freshly laid eggs were collected daily per treatment (daily, 4 eggs per replicate of each of the four treatments). These eggs were broken and for each one of them the yolk was separated from the egg white. The 24 yolks thereby obtained per treatment were frozen at -20 °C, freeze-dried and later on used for the determination of α- and γ-tocopherol and retinol content.
Once hens reached the age of 28 weeks, the trial was continued for another additional six weeks only with the ISA White laying hens. These 30 hens kept on eating the same diet they had been receiving since they were 22 weeks old (15 ISA White laying hens per diet). On weeks 31, 32 and 33 of age of laying hens, all eggs produced were collected daily, counted, marked and weighed. From the eggs obtained in weeks 31 and 32, 36 eggs per cage were set aside. Nine out these 36 eggs were used on collection day for assessment of albumen Haugh units, yolk colour score and eggshell quality parameters (shell thickness, shell breaking strength, total rupture area and shell rupture force peaks). The remaining 27 eggs per cage were stored in a refrigeration chamber at 4 °C. At 15, 21 and 31 days of storage, 9 eggs per cage were taken out of the chamber and also used for assessment of albumen Haugh units and yolk colour score. On week 32 of age of laying hens, 9 additional eggs per cage were used for measurement of shell thickness.
On the first day of week 33, 4 freshly laid eggs were taken per cage and broken. For each one of these eggs, the yolk was separated from the egg white. The four yolks originating from the same cage were pooled, hence obtaining 3 pools per dietary treatment. These 6 pools of yolks were frozen, freeze-dried and subsequently used for the determination of gallic acid concentration.
2.4. Measurements of Egg Quality Parameters
Eggshells were washed, the testaceous membranes were removed and then, the shells were dried in the air at room temperature for 48 h. Shell thickness was assessed at egg equator with a digital Mitutoyo shell thickness micrometre (Kawasaki, Japan).
Albumen height was measured with a QCH device (TSS, York, UK). Haugh units were calculated thereafter with the formula:
Haugh units = 100 × log (h − 1.7 × w0.37 + 7.57), where h = albumen height (mm) and w = egg weight (g) [19].
At week 27 of age of laying hens, colour of egg yolk was measured using a Minolta Chromameter (Model CR-400, Minolta Co., Osaka, Japan). According to the CIELAB colour space, colour values were expressed as L* (lightness), a* (redness) and b* (yellowness). White standard was used to calibrate the chromameter. In eggs laid by hens aged 31 and 32 weeks, yolk colour was assessed using the Roche Yolk Colour Fan and data were expressed in the standard DSM Roche Fan values (from 1 for light yellow to 15 for orange).
Mechanical measurements in the shells were performed using a TA-XT2i Texture Analyzer (Stable Micro Systems, Ltd., Godalming, UK) provided with Texture Exponent software (version 6.1.20.0) and equipped with a 30 kg load cell. A compression test was performed with a 45-mm diameter flat cylindrical stainless-steel probe (SMS P/45). Eggs were placed on a specific stationary platform in a horizontal position to the probe. This specific platform has three vertices and a central hole designed for the shape of the egg, collecting the white and the yolk separately. A pressure force was applied with the probe until the eggshell is completely broken. Test speed was set at 1 mm/s, considering a trigger test force of 0.001 N. Data acquisition was performed with a rate of 500 pps (points per second). From the force-time curves, the shell breaking strength (N) and the total rupture area (N.s), corresponding respectively to the maximum force and area required to crush the eggshell surface, were obtained. In addition, the number of shell rupture force peaks, calculated for a drop in force higher than 0.05 N, was derived.
2.5. Chemical Analyses
Dry matter (930.15), crude protein (976.05), crude fibre (978.10), ashes (942.05) and ether extract by Soxhlet fat analysis after acid hydrolysis (method 920.39) were analysed in accordance with the methods of the Association of Official Analytical Chemists [20]. Chemical analyses were performed in triplicate.
2.5.1. Vitamins A and E Determinations
Retinol and α- and γ-tocopherol extraction was performed following the protocol indicated by Claeys et al. [21]. In an amber glass tube with screw cap, 200 mg of egg yolk were weighed and stirred vigorously with 6 ml of ethanol with ascorbic acid (5 g/L). It was incubated in a water bath at 78 °C for 30 min, mixing the contents with a glass rod after 10 min incubation. The tube was cooled under tap water, 1 mL of distilled water was added and it was incubated in an ice bath for 15 min. It was centrifuged for 5 min at 1100 x g and the supernatant was transferred to another amber tube. Two millilitres of KCl (57.5 g/L) and 2 mL of hexane (containing 0.02 g/L BHT) were added, vigorously mixed and centrifuged for 5 min at 720 x g. The hexane phase was transferred to another amber tube using a Pasteur pipette. The hexane extraction step was repeated and the pooled hexane phase was evaporated under N2 gas. Finally, the precipitate was dissolved in 1 mL methanol, filtered through a 0.2 μm cellulose membrane (Scharlab S.L.) and collected in an amber vial.
Reversed-phase HPLC technique was used, with a Gilson 305 pump and a 5 μm (100 mm x 4.6 mm) KromaPhase C18 chromatography column. The mobile phase was methanol-water (97:3) with the following conditions: 1 mL/min flow rate, around 0.78 kpsi pressure and 25 °C. A Jasco UV-2075Plus UV detector was used, and absorbance was recorded at 292 nm for α- and γ-tocopherol and 325 nm for retinol. Quantification was carried out immediately after extraction. Standard solutions of known concentration of α-tocopherol, γ-tocopherol and retinol (Sigma-Aldrich®) were injected and peak areas were used to calculate α-tocopherol, γ-tocopherol and retinol concentrations in the samples.
2.5.2. Phenolic Compounds
For the extraction of phenolic compounds in GP and diets, 50 mg of the sample was placed in a capped centrifuge tube and suspended in 1 mL of acidified methanol 50:50 vortexed and sonicated over 15 min. Subsequently, samples were centrifuged at 10,400 x g for 10 min at 4 °C and the supernatant was collected. The resulting pellet was re-suspended in 500 μL of methanol-water (50:50) acidified with formic acid (0.1%), the extraction process being repeated twice. The obtained supernatants were combined (2 mL in total) and used to quantify the total extractable polyphenols by Folin-Ciocalteu method and for phenolic compound identification. For subsequent HPLC-QTOF-MS and HPLC-MS analysis of phenolic compounds, supernatants were filtered (0.45 µm) and placed in vials.
For egg yolk extraction, 200 mg of freeze-dried egg yolk were re-suspended in 1.5 mL of acidified methanol-water (80:20, 0.1% formic acid) and taxifolin was added as internal standard. Samples were gently mixed for 1 hour, sonicated over 15 min and centrifuged at 10,400 x g for 10 min at 4 °C. The supernatant was collected and the resulting pellet was extracted, repeating the process twice. Then, supernatants were mixed, concentrated (N atmosphere), re-suspended in 500 μL of methanol-water (50:50, 0.1% formic acid) and placed in vials for subsequent HPLC-QTOF-MS analysis of phenolic compounds.
The identification of the different compounds present GP and egg yolk was performed using HPLC coupled with a mass spectrometer (HPLCMS-QTOF). The HPLC (Agilent 1200, Agilent Technologies, Waldrom, Germany) with a quaternary pump (model G1311A) was coupled with a diode array detector (Agilent model G1315B) and an Agilent 6530 Accurate-Mass QTOF-MS with Electrospray Ionization (ESI) with Jet Stream technology (Agilent Technologies, Santa Clara, CA, USA). Separation was performed on a Phenomenex Luna C18 column (5 μm, 4.6 mm × 150 mm; Phenomenex, Alcobendas, Spain), set thermostatically at 25 °C. A gradient between solvent A (water/formic acid, 99.9:0.1, v/v) and solvent B (acetonitrile/formic acid, 99.9:0.1, v/v) was applied at a flow rate of 0.5 mL/min as follows: 10% B at 0 min, 30% B at 30 min, 35% B at 35 min, 40% B at 45 min, 10% B at 50 min, and 10% at 60 min. The electrospray ionization (ESI) parameters were as follows: drying gas flow, 8 L/min; nebulizer pressure, 45 psi; gas drying temperature, 325 °C; sheath gas temperature, 300 °C; sheath gas flow, 11 L/min; capillary voltage, 4000 kV; and fragmentator, 120 V. The ESI was operated in positive and negative mode to provide extra certainty in the determination of the molecular masses. Compounds were identified using extracted-ion chromatogram (EIC) data, by comparing with external standard and confirmed by tandem mass spectrometry fragmentation spectra (MS/MS). For mass spectrometry experiments, quite generic collision energy of 20 V was used. Spectral signal data were also acquired at 280, 320, and 520 nm. Data acquisition and processing were performed with a Masshunter Data Acquisition B.05.01 and Masshunter Qualitative Analysis B.07.00 SP2 software (both from Agilent Technologies, Santa Clara, CA, USA). Compounds were identified by comparing mass spectra and retention time with the corresponding standard if available. The quantification was performed by interpolation into the calibration curve of the standard or structurally related compound used to quantify (equivalent) and expressed as g per g of dry matter (DM) as follows: epicatechin for oligomeric procyanidins, quercetin-3-O-glucoside for flavonols, and cyanidin-3-O-glucoside for anthocyanidins.
2.6. Statistical Analysis
Data of variables were subjected to a one-way analysis of variance (ANOVA) with laying hen genetics, diet and their interaction as the main sources of variation by using the general linear model procedure (Version 9.4, SAS Institute Inc., Cary, NC, USA). When the effect was declared significant (P < 0.05), comparisons among all the treatment means were made using a t-test.
The cage with 5 hens represented the experimental unit for performance parameters, whereas the egg was the experimental unit for external and internal quality parameters and for α- and γ-tocopherol and retinol concentrations. A pool of four egg yolks from the same dietary treatment constituted the experimental unit for gallic acid concentration.
3. Results and Discussion
3.1. Productive Performance of Laying Hens
Genetics of laying hens and the diet consumed are the main sources of variation affecting egg production and quality [22,23]. Conventionally, two types of laying hens are used worldwide in laying hen husbandry: light hens (hens laying white eggs) and semi-heavy hens (hens laying brown eggs). Hence, in the present study, a strain of light hens (ISA White) and a strain of semi-heavy hens (ISA Brown) were used and compared.
Table 4 shows the effect of hen genetics and dietary treatment on productive performance of hens in the present research work. In keeping with productive results published in ISA Poultry performance guides, daily egg mass was lower for ISA White than for ISA Brown hens (55.2 vs 58.0 g/d, P = 0.004). However, since feed intake of ISA White hens was also lower than that of ISA Brown hens (111 vs 123 g/d, P < 0.001), ISA White hens turned out to be more efficient in egg production (lower feed conversion ratio, 2.02 vs 2.13, P = 0.012). Both strains of hens showed very high daily egg production (97.1%, on average), as it should have been expected for selected hens of this age (at 24-26 weeks of age, these laying hens reach peak egg production). In the current research work, the dietary inclusion of GP at 50 g/kg had no effect on egg weight. On the contrary, in a previous study [10] the inclusion of GP at 30 or 60 g/kg in the diet of laying hens reduced the weight of eggs laid. The different effect observed between these two works could be ascribable to the different grape polyphenol concentration in the experimental diets.
3.2. Egg Internal Quality
The effect of genetics and diet on various parameters determining the internal quality of freshly laid eggs is shown in Table 5. The Haugh units (HU) are an important indicator used to assess albumen quality, based on variables such as egg white height and egg weight [24]. In this context, a higher value of HU is associated with better protein quality in the egg, in accordance with the height of the albumen at its point of maximum viscosity and thickness. Indeed, high values of HU are largely considered indicators of egg freshness [25]. As it can be observed in Table 5, both strains of hens exhibited high HU values and, consequently, excellent quality of albumen, with no significant differences due to dietary inclusion of GP being detected. These results are in disagreement with those obtained by Romero et al. [10], who reported an increase in the HU of light laying hens (Hy-Line strain) fed diets including GP (either at 30 or 60 g/kg), as compared with the control treatment. Nevertheless, it should be noted that HU values of the present work are 22.5% higher, on average, than those of Romero et al. [10] and hence, it could be surmised that, since the HU values obtained in the current study in hens fed the control diet were already very high, very little further improvement could be achieved with the dietary inclusion of GP. Other authors, such as Zhu et al. [25], found a decrease in HU values in commercial Lohmann pink-shell laying hens when feeding them with different tea polyphenols in comparison with the control group. These differences could be explained by the different nature and content of the polyphenols present in tea and GP. Additionally, the use of different breeds could also contribute to explain the different response.
Concerning the yolk colour, when determined according to the CIELAB colour space, results (Table 5) showed that lightness (L*) was not affected either by diet or hen genetics. However, the parameter a* (redness tendency) was affected by both treatments, with a significant decrease in a* observed in eggs from hens that had consumed GP (2.73 vs 3.53, P < 0.001) and with higher values being found in semi-heavy hens (3.33 vs 2.93, P = 0.008). Likewise, the parameter b* (yellowness tendency) tended (P = 0.07) to decrease (25.9 vs 27.5) when diet of hens included GP. Therefore, it seemed that eggs from hens fed the diet including GP exhibited rather orange-coloured yolks, which may be more attractive from a sensory perspective for consumers [26].
For none of the internal quality parameters assessed in freshly laid eggs, the interaction between hen genetics and dietary inclusion of GP was found to be significant. Hence, the effect of diet and that of storage time (15, 21 and 31 days) of eggs on HU and yolk colour score, now measured using the Roche scale, was only evaluated with one strain of hens (light laying hens). These results are provided in Table 6. In keeping with the decreasing effect on HU due to storage duration observed by Grashorn et al. [27], a significant reduction (P < 0.001) over time of HU was detected in the present study (96.7, 93.4, 87.4 and 69.6 at 0, 15, 21 and 31 days of storage, respectively). On the contrary, the dietary inclusion of GP tended (P = 0.06) to increase the HU (87.8 vs 85.8), with the interaction between diet and storage time not being significant. The positive effect on HU resulting from the dietary inclusion of GP had not been detected previously when hens were 24 to 28 weeks old. Nonetheless, at that age, the diet containing GP led to HU values 1.45% higher than those of the control group but this difference did not reach significance level. Now, in hens aged 31 to 34 weeks, the effect of dietary GP could have become significant because a higher number of eggs per treatment were evaluated. Similarly, dietary inclusion of GP (either at 30 or 60 g/kg) also increased egg HU in a previous work [10]. As aforementioned, HU are related with the quality of albumen. The changes that this portion of the egg undergoes during storage have been well-studied for decades. Reduced HU values observed throughout storage indicate a decline in the functional properties of albumen. The consistency of the albumen was compromised when the HU score fell below 70 during the storage period [28]. Shan et al. [29] concluded that the highly glycosylated ovomucin in the egg white undergoes degradation as its glycan chain hydrolyses, accompanied by a thinning of the egg white. Another study [30] has suggested that ovalbumin gradually converts to S-ovalbumin during storage, with the content of S-ovalbumin being negatively correlated with HU.
As regards yolk colour, assessed according to the Roche scale, no effect due to storage time of eggs was detected (Table 6), whereas the presence of GP in the diet at 50 g/kg increased (P < 0.001) the yolk colour score by 9.36%. Again, as previously observed from 24 to 28 weeks of age of hens, the diet including GP made egg yolks look more orange. In previous research [10], the dietary inclusion of GP at 60 g/kg increased the yolk colour score by 12.8%. While this index does not imply nutritional value, it is crucial for consumer acceptance of eggs and it has been largely proven that egg yolk colour can be influenced by the diet fed to the hens [31,32]. Actually, it has been observed that European consumers prefer yolk colouration between 9 and 14, with differences between northern and southern countries. Southern countries tend to prefer yolks with more intense colours (11-14), while northern countries prefer paler yolks (9-10) [33]. In this study, means lower than 11 (ranging from 9.29 to 10.7) were obtained, which meets the quality standards of only northern European countries. This could be explained by the absence of colourants in both feeds. In a previous study [34], aimed at finding natural pigments to replace synthetic ones in the diets of laying hens, it was concluded that, among the studied ingredients (dandelion, marigold and basil), only eggs from diets supplemented with marigold flowers, a compound with a high content of xanthophylls, showed acceptable yolk colour values for southern European countries. Tufarelli et al. [35] also indicated that hens consuming 150 g/kg of dehydrated tomato pomace and 50 g/kg of dehydrated GP showed significantly higher yolk colour than those fed the diet containing flaxseed meal. In the current research, the interaction between diet and storage time was not found to be significant but Grčević et al. [36] reported that, while storage duration did not affect the yolk colour value of control eggs, eggs from experimental diets supplemented with lutein (in the form of powdered marigold extract) showed significantly higher yolk colour values in eggs stored for 20 days.
3.3. Egg External Quality
The external quality of eggs was evaluated on the basis of shell thickness and mechanical parameters derived from force-time curves (shell breaking strength, total rupture area and shell rupture force peaks). When hens were aged 24 to 28 weeks, no effect of dietary inclusion of GP was observed on shell thickness (Table 5), whereas in 31- to 34-week old laying hens (Table 7), the diet containing GP resulted in an impairment of shell thickness (361 vs 424 μm, P < 0.001). Polyphenols are known to have the ability to chelate calcium ions [37], which could have reduced the intestinal absorption of calcium in hens fed the GP diet and thereby, could have hindered the correct formation of shell in the uterus. This negative effect of dietary inclusion of flavonoids on shell thickness of eggs was also observed by Zhu et al. [27]. These authors showed that the addition of six tea by-products with different caffeine content to the diet of laying hens caused a marked reduction in shell thickness and weight. They reported that high quantities of caffeine could eventually bring about shell fracture, caused by calcium loss and availability. Finally, as commonly observed in laying hen husbandry [23], eggshell was thinner for light hens than for semi-heavy ones (370 vs 386 μm, P = 0.038).
Figure 1 is an example of the shell breaking force-time curves of samples from the control (Figure 1a) and GP diet (Figure 1b) groups. Eggs were compressed in their equatorial position until they were completely broken, obtaining very similar force-time curves. In eggs from both dietary treatments, a noticeable maximum peak of force was observed at around 20 s, corresponding with the shell breaking strength, followed by numerous smaller fracturability force peaks until the total rupture of the eggs ends, which happened at around 80 s. The mechanical parameters derived from these force-time curves are also shown in Table 7. Unlike what was detected for shell thickness, no negative effect of dietary inclusion of GP was found on any of the mechanical parameters, for which no significant differences were observed between the dietary treatments.
On the other hand, it is important to indicate that the number of eggs per treatment in which the shell breaking test was carried out was quite lower (n = 27) than that of eggs used to determine shell thickness (n = 54), so it would be advisable to increase the number of samples in future studies and thereby, confirm that this reduction in shell thickness does not have a negative effect on shell strength.
3.4. Bioactive Compounds Measurements
3.4.1. Vitamins A and E Determinations
The effect of diet and genetics on the composition of vitamin E (α- and γ-tocopherol) as well as vitamin A (retinol) in egg yolk is presented in Table 8. No effect on vitamin E and A concentration was observed when diets were compared. Previous research in chicken has shown an increase in α- and γ-tocopherol concentration in plasma and meat after feeding chickens with a diet including GP [6,7]. As far as seen in literature, this is the first time in which the amount of vitamins A and E is studied in the egg yolk after inclusion of GP in the diet of laying hens. The increase of vitamin E in plasma obtained by Romero et al. [6] may not be translated into the egg yolk, as in our case, due to the regulatory role of the liver in the incorporation of certain products into the egg yolk. Thus, the liver may need to be saturated before certain substances are incorporated into the egg yolk and this could be the case of the studied vitamins [38]. A study performed in laying hens with a hempseed supplemented diet observed an increase in α- and γ-tocopherol in egg yolks [17], but there are some important methodological differences (duration of the treatment, strains raised, degree of supplementation...) between their study and ours that could account for the absence of effect of the diet in our research work. It is also noteworthy to mention that our GP diet contained a higher percentage of sunflower oil than the control diet, in order to balance the energetic input between diets. Sunflower oil is rich in polyunsaturated fatty acids that may make the eggs more prone to oxidation [39]. Despite this, we have not found lower levels of α- and γ-tocopherol in the egg yolks from hens fed with the GP diet, which may indicate indeed a positive effect of dietary GP on the amount of vitamin E in the egg yolk. We hypothesize that polyphenols present in GP could stimulate the turnover from α-tocopheroxil to α-tocopherol, reducing the last one that is oxidised under oxidative stress and contributing to regenerate the reduced α-tocopherol pool. Another explanation could be that the polyphenols get oxidised preventing the antioxidants present in the egg yolk from being consumed. Concentration of retinol in egg yolks was not different in hens fed the GP diet than in hens fed with the control diet. The amount of retinol in eggs seems to be closely related to its presence in diet, either in form of retinol or its precursors, carotenoids [40,41]. In spite of the high number of carotenoids present in the grapes, these are usually degraded during the alcoholic fermentation [42] and we believe that the amount of carotenoids remaining in the GP incorporated to the diet may not have been enough to increase retinol content in the egg.
Regarding the influence of genetics on the amount of vitamins A and E in egg yolks, we observed a higher amount of α-tocopherol in ISA Brown egg yolks than in eggs laid by ISA White hens. However, the opposite was observed when γ-tocopherol was analysed, being higher in ISA White egg yolks than in ISA Brown. Differences between the strains used have also been shown in other parameters of this study, indicating the importance of genetics on the egg quality. In agreement with this, other studies have also described an effect of genetics on the amount of vitamins [41] and quality parameters [43]. In line with our results, a higher concentration of α-tocopherol was found in eggs laid by Rhode Island hens, which are semi-heavy hens just like the ISA Brown hens of the present study, in comparison with eggs of Leghorn hens, light hens as those of ISA White strain [41]. When analysing retinol concentration, we found no influence of genetics on the amount of this vitamin in the egg yolk. Remarkably, as shown in Table 6, we did not find any interaction between diet and genetics, indicating hence that the lack of effect of GP diet was consistent independently of genetics.
3.4.2. Phenolic Compounds
Laying hens have the capacity to transfer health-promoting components, such as vitamins and minerals, from the diet into eggs. Thus, incorporating bioactive compounds into laying hens’ diets could be a successful strategy to enhance the nutraceutical value of eggs. Among them, α-tocopherol, the most common antioxidant utilised in animal nutrition, is easily deposited in egg yolk from the diet [16,44]. Additionally, eggs can be enriched with other antioxidants such as carotenoids (mainly xanthophylls) or selenium [44,45]. Regarding the transfer of polyphenols into eggs, some few works have been carried out, in which the transference of phenolic compounds present in soybean [46] and in corn/wheat [47] was studied. Saitoh et al. [46,48] studied the biotransformation of soy isoflavone-glycosides in laying hens and detected preferential accumulation of equol, a metabolite of daidzein, into the egg yolk. However, so far, no further studies have been conducted regarding the transfer of phenolic compounds/metabolites into the egg. On the other hand, Nimalaratne et al. [47], after investigating the potential transfer of polyphenols from cereals (corn and wheat-based diets) to egg yolk, concluded that only trace amounts of ferulic acid were detected. Additionally, these authors pointed out that free aromatic amino acids, such as tryptophan and tyrosine, were the major contributors to the total antioxidant capacity of egg yolk extracts. Our results (Figure 2 and Figure 3) showed the presence of gallic acid in egg yolk from laying hens fed both control and GP diets. This could be explained by the fact that gallate esters are commonly used as antioxidants in both food and animal feed. Gallate compounds possess antioxidant capacity in both aqueous and lipid medium scavenging peroxy radicals [49]. Interestingly, we also observed that eggs yolks from hens fed the GP diet presented a gallic acid content higher than that of eggs obtained from hens fed the control diet, suggesting hence the potential transfer of some dietary grape polyphenols to the eggs. Previous works indicated a relatively high bioavailability of gallic acid, proved by the presence in different tissues of its metabolites (like 3-O-methylgallic, 4-O-methylgallic and 3-4-O-dymethylgallic), together with the parent compounds, after 1-2 hours of ingestion [50]. However, in the present study we have only detected the presence of gallic acid in the egg yolk and none of these metabolites. To the best of our knowledge, the presence of gallic acid in eggs had not been previously reported. An increased transfer of gallic acid to the eggs could contribute to improve their antioxidant capacity. As a consequence, the antioxidant capacity in both aqueous and lipid medium might protect egg white and egg yolk alike and could be associated with the effect observed in HU as well as in yolk colour, which were both improved in hens fed the diet containing GP. As aforementioned, high HU value is an indicator of albumen quality and egg freshness. It has been demonstrated that the incorporation of different vegetable sources in hen diets can be used as a nutritional strategy to increase eggs self-life and to improve the preservation of their nutritional value [51].
4. Conclusions
The inclusion of grape pomace in the diet of laying hens enabled an enrichment of egg yolks in gallic acid and enhanced egg quality parameters like albumen Haugh units and egg yolk color score. No dietary effect was observed on the content of α- and γ-tocopherol nor on the retinol content of the yolks. On the other hand, the dietary inclusion of grape pomace made eggshell be thinner, but this was not reflected in mechanical parameters. The influence of hen genetics was confirmed in parameters such as egg weight, shell thickness and vitamins concentration in the yolk.
Author Contributions
Conceptualization, S.C.; Formal analysis, M.L.T., I.S.R., B.H., M.D.A., C.R. and S.C.; Funding Acquisition, S.C. and C.R.; Investigation, I.A., A.V., B.H., M.D.A., S.P.T., I.S.R., M.L.T., and S.C.; Writing—original draft preparation, B.H. C.R., M.L.T., I.S.R. and S.C.; Writing—review and editing, B.H., C.R. and S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been partially supported by the “Instituto de Estudios del Huevo” (Spanish Institute for Egg Studies).
Data Availability Statement
Data reported in this work are available on request from the corresponding author. They are not publicly available because of privacy restrictions.
Acknowledgments
Authors are grateful to Bodegas Roquesán for providing the grape by-product.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dao, H.T.; Sharma, N.K.; Swick, R.A.; Moss, A.F. Feeding recycled food waste improved feed efficiency in laying hens from 24 to 43 weeks of age. Sci. Rep. 2023, 13, 8261–8273. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Goñi, I.; Centeno, C.; Saura-Calixto, F.; Arija, I. Effect of grape seed extract on growth performance, protein and polyphenol digestibilities and antioxidant activity in chickens. Span. J. Agric. Res. 2010, 8, 326–333. [Google Scholar] [CrossRef]
- Chamorro, S.; Viveros, A.; Rebolé, A.; Arija, I.; Romero, C.; Álvarez, I.; Rey, A.; Brenes, A. Addition of exogenous enzymes to diets containing grape pomace: Effects on intestinal utilization of catechins and antioxidant status of chickens. Food Res. Int. 2017, 96, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Nardoia, M.; Arija, I.; Viveros, A.; Rey, A.I.; Prodanov, M.; Chamorro, S. Feeding broiler chickens with grape seed and skin meals to enhance α- and γ-tocopherol content and meat oxidative stability. Antioxid. 2021, 10, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Nardoia, M.; Brenes, A.; Arija, I.; Viveros, A.; Chamorro, S. Combining grape byproducts to maximise biological activity of polyphenols in chickens. Animals 2021, 11, 3111–3122. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, S.; Romero, C.; Brenes, A.; Sánchez-Patán, F.; Bartolomé, B.; Viveros, A.; Arija, I. Impact of a sustained consumption of grape extract on digestion, gut microbial metabolism and intestinal barrier in broiler chickens. Food Funct. 2019, 10, 1444–1454. [Google Scholar] [CrossRef]
- Muñoz-Gonzalez, I.; Chamorro, S.; Pérez-Jiménez, J.; López-Andrés, P.; Álvarez. Acero, I.; Herrero, A.M.; Nardoia, M.; Brenes, A.; Viveros, A.; Arija, I.; Rey, A.; Ruiz-Capillas, C. Phenolic metabolites in plasma and thigh meat of chickens supplemented with grape byproducts. J. Agric. Food Chem. 2019, 67, 4463–4471. [Google Scholar] [CrossRef]
- Romero, C.; Arija, I.; Viveros, A.; Chamorro, S. Productive performance, egg quality and yolk lipid oxidation in laying hens fed diets including grape pomace or grape extract. Animals 2022, 12, 1076–1091. [Google Scholar] [CrossRef]
- Kara, K.; Kocaoğlu Güçlü, B.; Baytok, E.; Şentürk, M. Effects of grape pomace supplementation to laying hen diet on performance, egg quality, egg lipid peroxidation and some biochemical parameters. J. Appl. Anim. Res. 2016, 44, 303–310. [Google Scholar] [CrossRef]
- Barbe, A.; Mellouk, N.; Ramé, C.; Grandhaye, J.; Staub, C.; Venturi, E.; Cirot, M.; Petit, A.; Anger, K.; Chahnamian, M.; Ganier, P.; Callut, O.; Cailleau-Audouin, E.; Metayer-Coustard, S.; Riva, A.; Froment, P.; Dupont, J. A grape seed extract maternal dietary supplementation in reproductive hens reduces oxidative stress associated to modulation of plasma and tissue adipokines expression and improves viability of offsprings. Plos One 2020, 15, e0231131–e0231154. [Google Scholar] [CrossRef] [PubMed]
- Grandhaye, J.; Douard, V.; Rodriguez-Mateos, A.; Xu, Y.; Cheok, A.; Riva, A.; Guabiraba, R.; Zemb, O.; Philippe, C.; Monnoye, M.; Staub, C.; Venturi, E.; Barbe, A.; Ramé, C.; Dupont, J.; Froment, P. Microbiota changes due to grape seed extract diet improved intestinal homeostasis and decreased fatness in parental broiler hens. Microorganisms 2020, 8, 1141–1159. [Google Scholar] [CrossRef] [PubMed]
- Loetscher, Y.; Kreuzer, M.; Messikommer, R.E. Late laying hens deposit dietary antioxidants preferentially in the egg and not in the body. J. Appl. Poult. Res. 2014, 23, 647–660. [Google Scholar] [CrossRef]
- Goñi, I.; Brenes, A.; Centeno, C.; Viveros, A.; Saura-Calixto, F.; Rebolé, A.; Arija, I.; Esteve, R. Effect of dietary grape pomace and vitamin E on growth performance, nutrient digestibility and susceptibility to meat lipid oxidation in chickens. Poult. Sci. 2007, 86, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Galobart, J.; Barroeta, A.C.; Cortinas, L.; Baucells, M.D.; Codony, R. Accumulation of alpha-tocopherol in eggs enriched with omega3 and omega6 polyunsaturated fatty acids. Poult. Sci. 2002, 81, 1873–1876. [Google Scholar] [CrossRef]
- Skřivan, M.; Englmaierová, M.; Vít, T.; Skřivanová, E. Hempseed increases gamma-tocopherol in egg yolks and the breaking strength of tibias in laying hens. Plos One 2019, 14, e0217509–e0217517. [Google Scholar] [CrossRef] [PubMed]
- Guilland, J.C. Les interactions entre les vitamines A, D, E et K : synergie et/ou compétition. OCL 2011, 18, 59–67. [Google Scholar] [CrossRef]
- Eisen, E.J.; Bohren, B.B.; Mckean, H.E. The Haugh unit as a measure of egg albumen quality. Poult. Sci. 1962, 41, 1461–1468. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists International: Arlington, VA, USA, 1995. [Google Scholar]
- Claeys, E.; Vossen, E.; de Smet, S. Determination of α-tocopherol by reversed-phase HPLC in feed and animal-derived foods without saponification. J. Sci. Food Agr. 2015, 96, 522–529. [Google Scholar] [CrossRef]
- Kim, C.H.; Kang, H.K. Effects of energy and protein levels on laying performance, egg quality, blood parameters, blood biochemistry, and apparent total tract digestibility on laying hens in an aviary system. Animals 2022, 12, 3513–3522. [Google Scholar] [CrossRef] [PubMed]
- Calik, J.; Obrzut, J. Influence of genotype on productivity and egg quality of three hen strains included in a biodiversity program. Animals 2023, 13, 1848–1861. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Qiu, N.; Ma, M.H.; Jin, Y.G.; Yang, H.; Geng, F.; Sun, S.H. Estimation of egg freshness using S-ovalbumin as an indicator. Poult. Sci. 2012, 91, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.F.; Wang, J.P.; Ding, X.M.; Bai, S.P.; Qi, S.R.N.; Zeng, Q.F.; Xuan, Y.; Zhang, K.Y. Effect of different tea polyphenol products on egg production performance, egg quality and antioxidative status of laying hens. Anim. Feed Sci. Tech. 2020, 267, 114544–114551. [Google Scholar] [CrossRef]
- Ortiz, D.; Lawson, T.; Jarrett, R.; Ring, A.; Scoles, K.L.; Hoverman, L.; Rocheford, E.; Karcher, D.M.; Rocheford, T. The impact of orange corn in laying hen diets on yolk pigmentation and xanthophyll carotenoid concentrations on a percent inclusion rate basis. J. Appl. Poult. Res. 2022, 31, 100218–100225. [Google Scholar] [CrossRef]
- Grashorn, M.; Juergens, J.; Bessei, W. Effects of storage conditions on egg quality. Lohmann Information 2016, 50, 22–27. [Google Scholar]
- de Araújo Soares, R.; Vilela Borges, S.; Vilela Dias, M.; Hilsdorf Piccoli, R.; Fassani, E.J.; Cunha da Silva, E.M. Impact of whey protein isolate/sodium montmorillonite/sodium metabisulfite coating on the shelf life of fresh eggs during storage. LWT 2021, 139, 110611. [Google Scholar] [CrossRef]
- Shan, Y.; Tang, D.; Wang, R.; Tu, A.; Yi, Y.; Wang, X.; Liu, B.; Zhou, Y.; Huang, Q.; Lü, X. Rheological and structural properties of ovomucin from chicken eggs with different interior quality. Food Hydrocolloid. 2020, 100, 105393. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Huang, Q.; Cheng, L.; Gan, R.; Liu, L.; Wu, D.; Li, H.; Peng, L.; Geng, F. Underlying mechanism for the differences in heat-induced gel properties between thick egg whites and thin egg whites: Gel properties, structure and quantitative proteome analysis. Food Hydrocolloid. 2020, 106, 105873. [Google Scholar] [CrossRef]
- Nelson, T.S.; Baptist, J.N. Feed pigments: 2. The influence of feeding single and combined sources of red and yellow pigments on egg yolk color. Poult. Sci. 1968, 47, 924–931. [Google Scholar] [CrossRef]
- Wen, C.; Xu, X.; Zhou, D.; Yu, Q.; Wang, T.; Zhou, Y. The effects of canthaxanthin microencapsulation on yolk color and canthaxanthin deposition in egg yolk of laying hens. Poult. Sci. 2022, 101, 101889–101893. [Google Scholar] [CrossRef] [PubMed]
- Grashorn, M. Feed additives for influencing chicken meat and egg yolk color. In Handbook on Natural Pigments in Food and Beverages; Carle, R., Schweiggert, R., Eds.; Publisher: Woodhead Publishing, Cambridge, UK, 2016; pp. 283–302. [Google Scholar]
- Kljak, K.; Carović-Stanko, K.; Kos, I.; Janječić, Z.; Kiš, G.; Duvnjak, M.; Safner, T.; Bedeković, D. Plant carotenoids as pigment sources in laying hen diets: Effect on yolk color, carotenoid content, oxidative stability and sensory properties of eggs. Foods 2021, 10, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Tufarelli, V.; Baghban-Kanani, P.; Azimi-Youvalari, S.; Hosseintabar-Ghasemabad, B.; Slozhenkina, M.; Gorlov, I.; Viktoronova, F.M.; Seidavi, A.; Laudadio, V. Effect of dietary flaxseed meal supplemented with dried tomato and grape pomace on performance traits and antioxidant status of laying hens. Anim. Biotechnol. 2022, 33, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Grčević, M.; Kralik, Z.; Kralik, G.; Galović, D.; Radišić, Z.; Hanžek, D. Quality and oxidative stability of eggs laid by hens fed marigold extract supplemented diet. Poult. Sci. 2019, 98, 3338–3344. [Google Scholar] [CrossRef] [PubMed]
- O'Connell, J.E.; Fox, P.F. Effect of extracts of oak (Quercus petraea) bark, oak leaves, aloe vera (Curacao aloe), coconut shell and wine on the colloidal stability of milk and concentrated milk. Food Chem. 1999, 66, 93–96. [Google Scholar] [CrossRef]
- Squires, M.W.; Naber, E.C. Vitamin profiles of eggs as indicators of nutritional status in the laying hen: vitamin A study. Poult. Sci. 1993, 72, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Cherian, G.; Wolfe, F.W.; Sim, J.S. Dietary oils with added tocopherols: effects on egg or tissue tocopherols, fatty acids, and oxidative stability. Poult. Sci. 1996, 75, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Naber, E.C. Modifying vitamin composition of eggs: a review. J. Appl. Poult. Res. 1993, 2, 385–393. [Google Scholar] [CrossRef]
- Rey, A.I.; de-Cara, A.; Rebolé, A.; Arija, I. Short-term spirulina (Spirulina platensis) supplementation and laying hen strain effects on eggs' lipid profile and stability. Animals 2021, 11, 1944–1959. [Google Scholar] [CrossRef]
- Guedes de Pinho, P.; Silva Ferreira, A.C.; Mendes Pinto, M.; Benitez, J.G.; Hogg, T.A. Determination of carotenoid profiles in grapes, musts, and fortified wines from Douro varieties of Vitis vinifera. J. Agr. Food Chem. 2001, 49, 5484–5488. [Google Scholar] [CrossRef]
- Sharma, M.K.; McDaniel, C.D.; Kiess, A.S.; Loar, R.E.; Adhikari, P. Effect of housing environment and hen strain on egg production and egg quality as well as cloacal end eggshell microbiology in laying hens. Poult. Sci. 2022, 101, 101595–101612. [Google Scholar] [CrossRef] [PubMed]
- Skřivan, M.; Englmaierová, M. The deposition of carotenoids and α-tocopherol in hen eggs produced under a combination of sequential feeding and grazing. Anim. Feed Sci. Tech. 2014, 190, 79–86. [Google Scholar] [CrossRef]
- Bennett, D.C.; Cheng, K.M. Selenium enrichment of table eggs. Poult. Sci. 2010, 89, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, S.; Sato, T.; Harada, H.; Takita, T. Transfer of soy isoflavone into the egg yolk of chickens. Biosci. Biotechnol. Biochem. 2001, 65, 2220–2225. [Google Scholar] [CrossRef] [PubMed]
- Nimalaratne, C.; Lopes-Lutz, D.; Schieber, A.; Wu, J. Free aromatic amino acids in egg yolk show antioxidant properties. Food Chem. 2011, 129, 155–161. [Google Scholar] [CrossRef]
- Saitoh, S.; Sato, T.; Harada, H.; Matsuda, T. Biotransformation of soy isoflavone-glycosides in laying hens: intestinal absorption and preferential accumulation into egg yolk of equol, a more estrogenic metabolite of daidzein. Biochim. Biophys. Acta 2004, 1674, 122–130. [Google Scholar] [CrossRef]
- Medina, M.; Iuga, C.; Alvarez-Idaboy, J.R. Antioxidant activity of propyl gallate in aqueous and lipid media: a theoretical study. Phys. Chem. Chem. Phys. 2013, 31, 13137–13146. [Google Scholar] [CrossRef]
- Kaliora, A.C.; Kanellos, P.T.; Kalogeropoulos, N. Gallic acid bioavailability in humans. In Handbook on Gallic Acid; Thompson, M.A., Collins, P.B., Eds.; Publisher: Nova Science Publishers, New York, USA, 2013; pp. 301–312. [Google Scholar]
- Obianwuna, U.E.; Oleforuh-Okoleh, V.U.; Wang, J.; Zhang, H.J.; Qi, G.H.; Qiu, K.; Wu, S.G. Potential implications of natural antioxidants of plant origin on oxidative stability of chicken albumen during storage: A review, Antioxid. , 2022, 11, 630–649. [Google Scholar] [CrossRef]
Figure 1.
Examples of shell breaking force for eggs from ISA White laying hens fed control (a) and grape pomace (50 g/kg) (b) diets.
Figure 1.
Examples of shell breaking force for eggs from ISA White laying hens fed control (a) and grape pomace (50 g/kg) (b) diets.
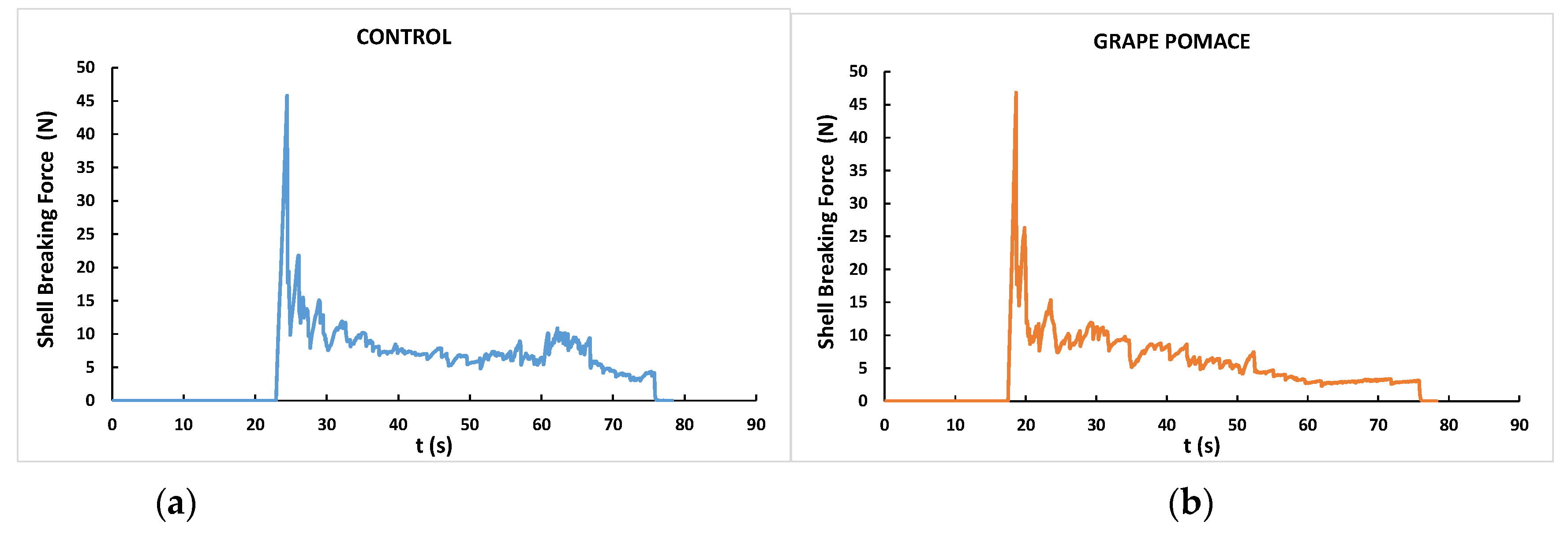
Figure 2.
Effect of dietary inclusion of grape pomace (GP) on yolk gallic acid concentration (μg/g) in eggs of 31- to 34-week old ISA White laying hens. Data are means of 3 replicates per treatment (a pool of 4 egg yolks per replicate); Figure shows means ± standard deviations. SEM = 0.104, P = 0.074.
Figure 2.
Effect of dietary inclusion of grape pomace (GP) on yolk gallic acid concentration (μg/g) in eggs of 31- to 34-week old ISA White laying hens. Data are means of 3 replicates per treatment (a pool of 4 egg yolks per replicate); Figure shows means ± standard deviations. SEM = 0.104, P = 0.074.
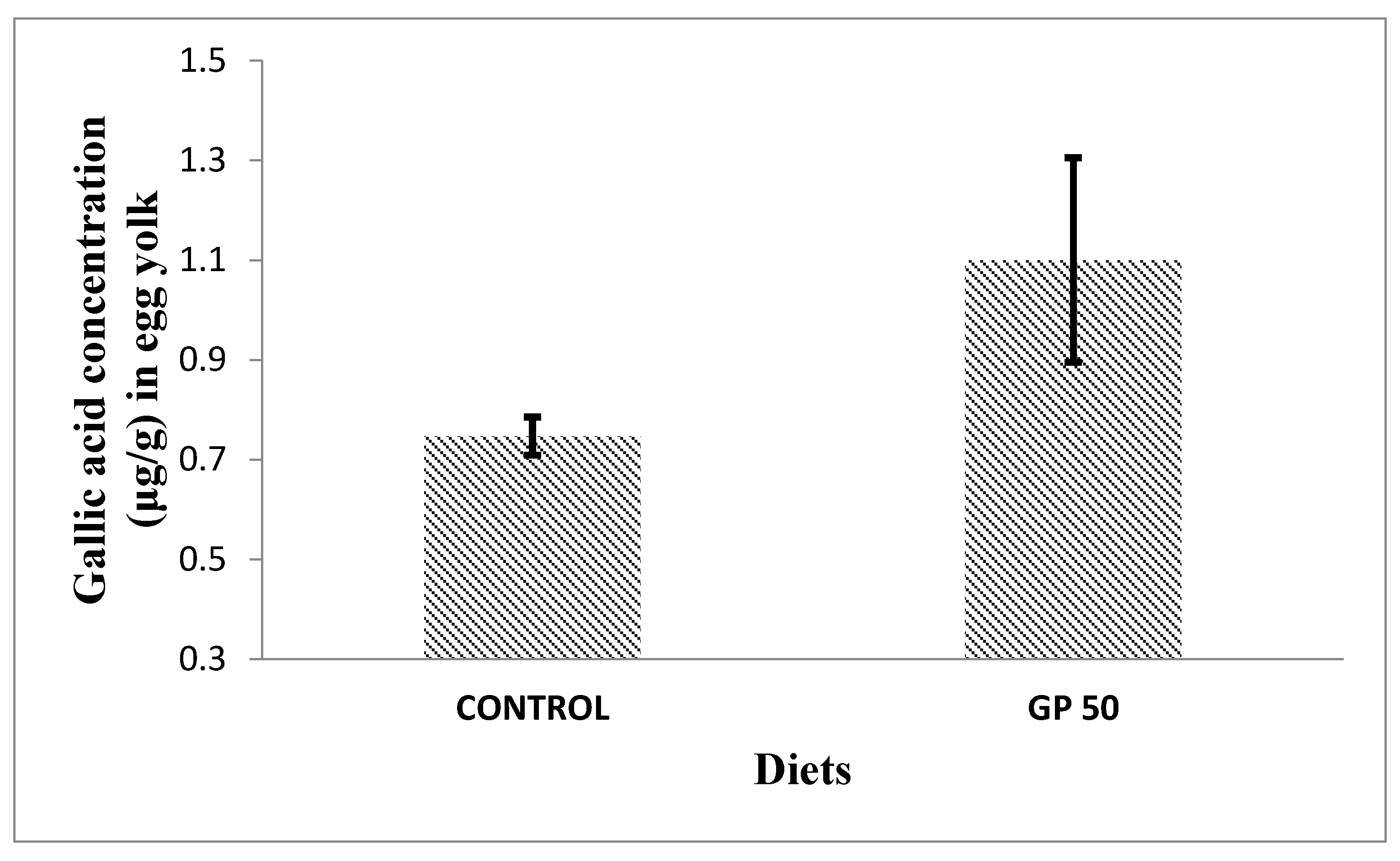
Figure 3.
Extracted ion chromatogram (EIC) of gallic acid standard and in egg yolk from hens fed grape pomace (50 g/kg) or control diet.
Figure 3.
Extracted ion chromatogram (EIC) of gallic acid standard and in egg yolk from hens fed grape pomace (50 g/kg) or control diet.
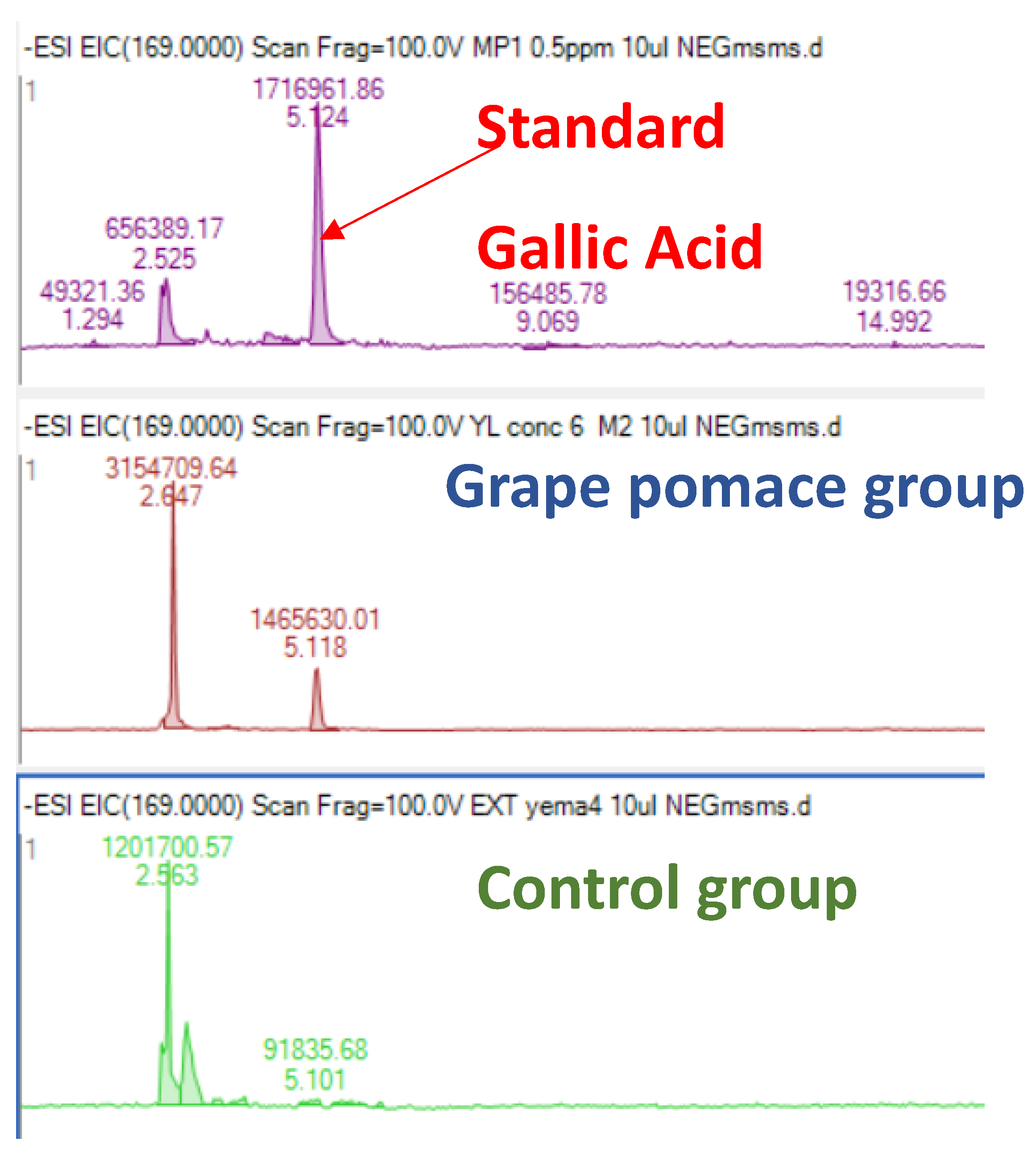

Table 1.
Proximate composition (g/100 g) of grape pomace.
| Nutrients | Grape pomace composition |
|---|---|
| Ether extract | 7.5 |
| Crude protein | 10.4 |
| Crude fibre | 25.3 |
| Ashes | 5.1 |
| Total extractable polyphenols (g gallic acid equivalents/100 g DM) |
2.10 |
Table 2.
Phenolic profile (mg/100 g DM) of grape pomace.
| Compound | Molecular formula | Mass | mg/g |
|---|---|---|---|
| Gallic acid | C7H6O5 | (-)169,0149 | 0.12 |
| Delphinidin-3-glucoside | C21H21O12 | (+)465,1005 | 0.09 |
| Cyanidin-3-glucoside | C21H21O11 | (+)449,1099 | 0.13 |
| Procyanidin B2 | C30H26O12 | (-)577,1367 | 0.80 |
| Procyanidin B3 | C30H26O12 | (-)577,1367 | 0.21 |
| Catechin | C15H14O6 | (-)289,0733 | 0.50 |
| Peonidin-3-glucoside | C22H23O11 | (+)463,1217 | 0.90 |
| Epicatechin | C15H14O6 | (-)289,0731 | 0.52 |
| Vitisin A | C26H25O14 | (+)561,1217 | 0.04 |
| Myricetin-3-glucoside | C21H20O13 | (-)479,0848 | 0.06 |
| Malvidin-3-acetylglucoside | C25H27O13 | (+)535,1433 | 0.04 |
| Delphinidin 3-(6-coumaroylglucoside) | C30H27O14+ | (+)611,1399 | 0.03 |
| Malvidin 3-caffeoylglucoside | C32H31O15+ | (+)655,1655 | 0.05 |
| Quercetin-3-glucoside | C21H20O12 | (-)463,0892 | 0.11 |
| Cyanidin 3-(6-coumaroylglucoside) | C30H27O13 | (+)595,1440 | 0.04 |
| Quercetin-3-glucuronide | C21H18O13 | (-)477,0684 | 0.14 |
| Malvidin 3-(6-coumaroylglucoside) | C32H31O14 | (+)639,1705 | 0.08 |
| Kaempherol-3-glucoside | C21H20O11 | (-)447,0942 | 0.06 |
| Quercetin | C15H10O7 | (-)301,0340 | 0.07 |
| Kaempherol | C15H10O6 | (-)285,0442 | 0.04 |
Table 3.
Ingredients and nutrient composition of experimental diets (g/kg as fed).
| Ingredients | Experimental diets | |
|---|---|---|
| Control | GP 1 50 | |
| Corn | 500.8 | 468.6 |
| Soybean | 295.0 | 293.2 |
| Sunflower oil | 54.0 | 63.1 |
| Grape pomace | - | 50.0 |
| Straw | 25.0 | - |
| Salt | 3.0 | 3.0 |
| Monocalcium phosphate | 12.5 | 12.5 |
| Calcium carbonate | 92.5 | 92.5 |
| Vitamin-mineral premix 2 | 5.0 | 5.0 |
| DL-Methionine | 2.2 | 2.1 |
| Celite 3 | 10.0 | 10.0 |
| Analysed composition | ||
| Crude protein | 166 | 169 |
| Ether extract | 76.0 | 86.0 |
| Crude fibre | 35.0 | 35.0 |
| Total extractable polyphenols (g gallic acid equivalents/kg) |
0.644 | 0.728 |
| Calculated composition | ||
| Grape extractable polyphenols (g gallic acid equivalents/kg) 4 |
- | 1.05 |
| AME 5 (MJ/kg) | 11.4 | 11.4 |
| Calcium | 38.6 | 38.9 |
| Available P | 3.70 | 3.70 |
| Lysine | 8.85 | 8.85 |
| Meth+Cys | 7.86 | 7.74 |
1 GP = Grape pomace; 2 Vitamin-mineral mix supplied the following per kilogram of diet: vitamin A, 12,320 IU; vitamin D3, 4,620 IU; vitamin E, 15.4 IU; vitamin K, 3.08 mg; riboflavin, 6.16 mg; niacin, 46.2 mg; vitamin B12, 23.1 μg; pantothenic acid, 15.4 mg; folic acid, 0.31 mg; choline, 401 mg; Fe, as FeSO4, 50.4 mg; Zn, as ZnO, 71 mg; Mn, as MnO, 90 mg; Cu, as CuSO4, 7 mg; I, as ethylenediamine dihydroiodide, 0.7 mg; and Se, as Na2SeO3, 0.25 mg. 3 Celite Corp, Lompoc, CA; 4 Calculated on the basis of the analyses of polyphenol concentration in GP; 5 AME = apparent metabolisable energy.
Table 4.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on productive performance of 24- to 28-week old laying hens.
Table 4.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on productive performance of 24- to 28-week old laying hens.
| Genetics | ISA White | ISA Brown | SEM 1 | P | ||||
|---|---|---|---|---|---|---|---|---|
| Diet | Control | GP 50 | Control | GP 50 | Genetics | Diet | Interaction | |
| Daily egg production (%) | 97.2 | 95.7 | 97.6 | 98.1 | 0.839 | 0.10 | 0.50 | 0.20 |
| Average egg weight (g) | 57.0 | 57.3 | 59.4 | 59.1 | 0.691 | 0.002 | 0.95 | 0.65 |
| Daily egg mass (g/d) | 55.5 | 54.9 | 58.0 | 58.0 | 0.975 | 0.004 | 0.71 | 0.73 |
| Feed intake (g/d) | 111 | 111 | 124 | 123 | 1.41 | < 0.001 | 0.81 | 0.82 |
| Feed conversion ratio (g feed/ g egg mass) |
2.01 | 2.03 | 2.14 | 2.13 | 0.047 | 0.012 | 0.90 | 0.75 |
1 n = 3 replicates (5 hens per replicate).
Table 5.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on quality parameters in eggs of 24- to 28-week old laying hens.
Table 5.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on quality parameters in eggs of 24- to 28-week old laying hens.
| Genetics | ISA White | ISA Brown | SEM 1 | P | ||||
|---|---|---|---|---|---|---|---|---|
| Diet | Control | GP 50 | Control | GP 50 | Genetics | Diet | Interaction | |
| Shell (%) | 10.6 | 10.4 | 10.1 | 10.2 | 0.137 | 0.017 | 0.97 | 0.24 |
| Shell thickness (μm) | 368 | 373 | 380 | 392 | 6.0 | 0.038 | 0.21 | 0.56 |
| Haugh units | 93.8 | 96.3 | 98.6 | 99.0 | 1.30 | 0.003 | 0.23 | 0.38 |
| Egg yolk colour | ||||||||
| Lightness, L* | 43.5 | 45.6 | 43.5 | 44.5 | 1.06 | 0.61 | 0.15 | 0.58 |
| Redness, a* | 3.35 | 2.51 | 3.72 | 2.95 | 0.14 | 0.008 | < 0.001 | 0.82 |
| Yellowness, b* | 27.2 | 25.1 | 27.8 | 26.7 | 0.89 | 0.23 | 0.07 | 0.59 |
1 n = 3 replicates (6 eggs per replicate).
Table 6.
Effect of egg storage time and dietary inclusion of grape pomace (GP) on Haugh units and yolk colour score in eggs of 31- to 34-week old ISA White laying hens.
Table 6.
Effect of egg storage time and dietary inclusion of grape pomace (GP) on Haugh units and yolk colour score in eggs of 31- to 34-week old ISA White laying hens.
| Storage time | 0 days | 15 days | 21 days | 31 days | SEM 1 | P | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Control | GP 50 | Control | GP 50 | Control | GP 50 | Control | GP 50 | Storage time | Diet | Interaction | |
| Haugh units | 95.1 | 98.4 | 93.3 | 93.6 | 85.1 | 89.8 | 69.9 | 69.3 | 0.70 | < 0.001 | 0.06 | 0.22 |
| Yolk colour score | 9.62 | 10.4 | 9.56 | 10.5 | 9.29 | 10.7 | 9.59 | 10.2 | 0.217 | 0.41 | < 0.001 | 0.27 |
1 n = 3 replicates (9 eggs per replicate).
Table 7.
Effect of dietary inclusion of grape pomace (GP) on egg shell quality in eggs of 31- to 34-week old ISA White laying hens.
Table 7.
Effect of dietary inclusion of grape pomace (GP) on egg shell quality in eggs of 31- to 34-week old ISA White laying hens.
| Diet | Control | GP 50 | SEM 1 | P |
|---|---|---|---|---|
| Average egg weight (g) | 63.5 | 63.2 | 0.29 | 0.68 |
| Shell thickness (μm) | 424 | 361 | 8.6 | < 0.001 |
| Shell breaking strength (N) | 46.4 | 45.6 | 1.50 | 0.55 |
| Total rupture area (N.s) | 441 | 413 | 15.5 | 0.21 |
| Shell rupture force peaks | 92.6 | 82.6 | 3.84 | 0.06 |
1 n = 3 replicates (18 eggs per replicate for egg weight and shell thickness; 9 eggs per replicate for shell breaking strength, total rupture area and shell rupture force peaks).
Table 8.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on α- and γ-tocopherol and retinol content (μg/g) in the yolk of eggs from 24- to 28-week old laying hens.
Table 8.
Effect of laying hen genetics and dietary inclusion of grape pomace (GP) on α- and γ-tocopherol and retinol content (μg/g) in the yolk of eggs from 24- to 28-week old laying hens.
| Genetics | ISA White | ISA Brown | SEM 1 | P | ||||
|---|---|---|---|---|---|---|---|---|
| Diet | Control | GP 50 | Control | GP 50 | Genetics | Diet | Interaction | |
| α-tocopherol | 87.7 | 104 | 139 | 142 | 14.8 | 0.005 | 0.55 | 0.65 |
| γ-tocopherol | 35.3 | 41.0 | 22.1 | 22.5 | 5.03 | 0.004 | 0.56 | 0.60 |
| retinol | 42.1 | 42.0 | 57.2 | 47.5 | 6.64 | 0.14 | 0.44 | 0.48 |
1 n = 3 replicates (8 eggs per replicate).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



