
Submitted:
15 April 2024
Posted:
16 April 2024
You are already at the latest version
Abstract
It has been proposed that procedures which up-regulate mitochondrial biogenesis and autophagy by replacing damaged mitochondria with healthy ones, may prevent the development of several heart diseases. A member of serine and threonine kinases, AMPK, could play essential roles in the autophagy and/or mitophagy. AMPK is widely distributed in various cells, which might play diverse regulatory roles in different tissues and/or organs. In fact, changes in the kinase function of AMPK due to alteration of activity have been linked with diverse pathologies including cardiac disorders. AMPK can regulate mitochondrial biogenesis via PGC1α signaling and also improve oxidative mitochondrial metabolism through inhibition of mTOR pathway, which may also modulate the autophagy/mitophagy through ULK1 and/or TGF-β signaling. Therefore, the modulation of AMPK in autophagy/mitophagy pathway might be considered as a therapeutic tactic for several cardiac disorders. As kinases are amongst the most controllable proteins, in general, the design of small molecules targeting kinases might be an eye-catching avenue to modulate the cardiac function. Some analyses of the molecular biology underlying mitophagy suggests that nutraceuticals and/or drugs including specific AMPK modulator as well as physical exercise and/or dietary restriction that could modulate AMPK may be useful against several heart diseases. In particular, some nutraceutical regimens may have encouraging potential for controlling some of cardiac disorders.
Keywords:
autophagy
; mitophagy
; AMPK
; cardiomyocyte
; cardiac arrhythmia
; cardiovascular disease
; cardiac failure
1. Introduction
Autophagy is a conserved lysosome-dependent pathway, which is also an intracellular self-protective process that can cut down damaged molecules and/or organelles [1]. In conditions of mild stress, autophagy maintains cellular energy homeostasis by recycling damaged organelles. Under severe stress conditions, however, too much increased activation of autophagy may lead to non-selective degradation of related substances and molecules/organelles, thereby exacerbating mitochondrial damage and triggering energy imbalance and/or cellular apoptosis [2]. Therefore, the autophagy may be an adaptive cellular response to oxidative stress, inflammation, aging, hypoxia, starvation, metabolic abnormalities and other external stimuli [3]. Autophagy can exert significant influence on various cardiac pathologies including cardiac arrhythmias, cardiovascular diseases, and/or heart failure [4]. In general, autophagy is also involved in the maintenance of intracellular homeostasis in most types of cells including cardiomyocytes, vascular smooth muscle cells, fibroblasts, macrophages, and endothelial cells [5, 6]. Cardiomyocytes are terminally differentiated cells, which might considerably rely on autophagy for the elimination of abnormal substances [7]. Therefore, the regulation of autophagy is crucial for the safeguarding of heart homeostasis.
Mitophagy is a selective autophagy in mitochondrial damages [8]. The common feature of mitophagy is the form of autophagic vacuole enclosing damaged mitochondria, which is also well-defined as a mitophagosome [9]. Therefore, mitophagy is a kind of autophagic response that explicitly targets injured and/or cytotoxic mitochondria. Interestingly, imperfections in autophagy or mitophagy have been identified to exacerbate the character to instinctively develop cardiac disorders. (Figure 1) For example, the depletion of a key mediator of mitophagy may cause left ventricular dysfunction and pathological cardiac hypertrophy in mice [10]. Mitophagy in heart can be induced by hypoxia and/or excessive production of reactive oxygen species (ROS) [11]. Aberrant mitophagy may induce the accumulation of damaged mitochondria, decrease of cardiomyocytes, and dysfunction of heart contraction. In addition, the abnormalities in mitochondrial dynamics and mitophagy are directly related to the defective clearance of damaged mitochondria and the activation of inflammatory responses, which eventually contribute to heart failure [12]. Following acute myocardial infarction, for instance, serious impairment of cardiac function may occur, leading to complications such as cardiac arrhythmia and/or severe heart failure. These cardiac disorders with mitochondrial dysfunction being an important factor in their development are the leading reason for death globally. Therefore, protecting the heart and/or improving cardiac function are important therapeutic goals after acute myocardial infarction. Given the abundance of mitochondria in cardiomyocytes, it is expected that dysfunction of mitochondria could impact cellular activity, and may eventually promote the development of those serious heart diseases.
Mitochondria continuously adjust their shape through fission and fusion in response to alterations in adenosine triphosphate (ATP) supply and demand. Interestingly, compared to those in healthy cardiomyocytes, diabetic cardiomyocytes have a reduced mitochondrial size and enlarged spatial density, which may increase the ATP supply of mitochondrial oxidative phosphorylation [13]. In this regards, the role of adenosine monophosphate (AMP)-activated protein kinase (AMPK) in mitochondrial homeostasis has been well-established. The AMPK has been shown to be involved in mitochondrial biogenesis, and promotion of autophagy. For example, resveratrol could function as an antioxidant through the stimulation of AMPK in mitochondria [14]. In addition, the AMPK is considered as an important protein kinase working in the signaling pathway that has been shown to exert prominent cardioprotective effects on the pathophysiological mechanisms of various heart diseases. In addition, the modification of AMPK activity could be a significant dietary approach for preventing and/or treating several heart diseases [15]. Western-style diet often leads to food overconsumption, which triggers the development of comorbidities such as obesity, hypercholesterolemia, cardiovascular disease, and chronic heart failure. Several studies suggest that intermittent fasting may also protect against the development of those morbidities [16]. Interestingly, it has been shown that fasting may be related to the modulation of autophagy and/or mitophagy [17]. Dietary interventions involving caloric restriction could induce autophagy/mitophagy in the heart [18]. Based on the current evidence, this review may present the potential molecular mechanisms of the beneficial effects by which some diets could play important roles against several heart diseases. Additional nutraceuticals which might have potential modulating effects on mitophagy with heart protective outcomes are also discussed.
2. Relationship between Autophagy/Mitophagy and Several Diseases Including Cardiac Discorders
Cardiomyocytes are predominantly occupied by numerous mitochondria, which provide sufficient but tremendous adenosine triphosphate (ATP) required for cardiac function [19]. In addition, the cardiac mitochondria have a critical role in the modulation of calcium storage and/or cell apoptosis [20]. Remarkably, some quota of the Ca2+ released from sarcoplasmic reticulum may be taken up by mitochondria through the interaction between mitochondria and sarcoplasmic reticulum, which can trigger the production of ATP [21]. Abnormal mitochondrial function would result in inadequate energy supply, thereby increasing the generation of ROS [22]. In general, the ROS accumulation may lead to injure to mitochondrial lipids, proteins, and DNAs involved in the production of ATP, thereby contributing to severe oxidative damage and/or cell death [23]. Such damage may be participated in different diseases as shown by neurodegenerative and/or cardiovascular diseases or in different terminal organs [24].
As a result, the function of the heart is strongly influenced by the condition of mitochondria, which are often also damaged in various cardiac disorders [25]. Considering the critical role of mitophagy in the removal of damaged mitochondria, therefore, accumulating evidence supposes that alterations in mitophagy could contribute to the progression of cardiac disorders [26]. For example, mitophagy may slow down the progression of cardiac arrhythmia by eliminating damaged mitochondria and/or modulating ROS levels [27]. In addition, the dysfunction of mitochondria might affect the ATP production and/or cardiac electrical transmission, leading to altered K+ fluxes in sarcolemma via the ATP-dependent potassium channels [27]. Mitochondrial dysfunction may also indorse to decrease ATP levels and/or increases ROS levels, which might provoke the heterogeneity of cardiac potential activity [28]. Subsequently, mitochondrial dysfunction might be worsened associated with reduced ATP production. This rubbish cycle might contribute to electrophysiological modifications and/or cardiac arrhythmia. Therefore, suitable mitophagy is indispensable for the inhibition of cardiac arrhythmia. Alteration of ATP and/or ROS levels may play an imperative role in arrhythmia [29]. The development of arrhythmia and inappropriate energy metabolism with the alteration of mitophagy in cardiomyocytes might be closely connected [30]. Mitophagy may protect cardiac cells from myocardial damage in ischemic heart disease that is the consequence of insufficient coronary blood flow [31]. However, several myocardial damages in cardiac ischemia could impede the mitophagy, which rather induces apoptosis in myocardial cells. Similarly, insufficient mitophagy may further exacerbate heart injury. Decreased levels of mitophagy are frequently detected in patients with severe heart failure that is categorized by mitochondrial dysfunction [32]. The vital role of mitophagy in the progression of heart failure has been also suggested by patients with mild heart failure [33].
It has been shown that activation of a serine-threonine kinase, AMPK, signaling pathway can promote mitophagy, which may also decrease oxidative stress in myocardial cells and improves heart function [34]. Therefore, the myocardial remodeling and/or the dysregulation of mitophagy with the alteration of AMPK signaling may be considerable pathological processes in the development of heart failure characterized by myocardial hypertrophy and/or apoptosis of myocardial cells [35]. In fact, the maintenance of normal function in myocardial cells may deeply depend on basal levels of autophagy with proper AMPK signaling, and impaired autophagy with inappropriate AMPK signaling might lead to myocardial cell hypertrophy [36]. It has been revealed that insufficient mitophagy during heart failure is associated with the alteration of PI3K/AKT/ mechanistic/mammalian target of rapamycin (mTOR) signaling pathway as well as the impaired AMPK signaling. (Figure 2) The upregulation of AMPK along with inhibition of the mTOR pathway may stimulate autophagy, which might lessen myocardial hypertrophy and improves heart failure [37]. AMPK can control other cellular important functions such as energy metabolism, apoptosis, oxidative stress, and inflammation as well as autophagy and mitophagy [38]. Therefore, the activity of AMPK might have a significant role in various heart diseases that could cause cardiac arrhythmias by changing arrhythmogenic circumstances, downregulating ion channels and triggering physiological dysfunction [39]. In addition, activation of AMPK is involved in the reduction of ROS levels [40]. This effect may be related to the improvement of inflammatory responses, which can further reduce the incidence of cardiac arrhythmias [41]. In fact, AMPK -/- mice may exhibit more extensive cardiac inflammation [41]. In addition, the underlying mechanism of several heart diseases may suggest that AMPK can decrease acute oxidative stress-induced cardiac inflammation [42].
3. Characterization of AMPK and Connection between AMPK and Autophagy
AMPK is a heterotrimeric complex made up of the three subunits, which exists one catalytic α-subunit and two β- and γ- regulatory subunits. The β subunit has an N-terminus region that can be myristoylated participating in the regulation of the AMPK recruitment to mitochondria. While each α and β subunits have two isoforms (α1, α2 and β1, β2), the γ subunit has three isoforms (γ1, γ2, and γ3) [43]. As a result, AMPK is an intricate heterotrimer that widely exists in eukaryotes. The combination of different isoform subunits may construct different AMPK functions that are differentially expressed in cells, tissues and/or organs. In addition, the structure of AMPK includes a carbohydrate binding segment that permits the binding of AMPK to glycogen, impeding AMPK activity by this means [44]. An increase in AMP triggers AMPK γ subunit to bind AMP, enhancing the catalytic activity of the complex and increasing the phosphorylation of AMPK [45]. Therefore, AMPK is a kind of sensor of intracellular AMP levels. The carboxy (C)-terminal domain of the β subunit may associate with the α- and γ- subunits, then works as a scaffold protein. The γ subunit enables AMPK to respond to alterations in the AMP/ATP ratio, hence acting as an exact energy sensor [46].
Continuing suitable levels of mitophagy may be essential in the myocardium. AMPK can respond to heart failure by promoting the creation of autophagosomes and by limiting oxidative stress and/or cardiac apoptosis [47]. Consequently, AMPK might be dynamic in a coordinated intracellular network adjusting cellular homeostasis. [48]. (Figure 2) Too much increased cardiac ATP request and/or severe oxidative damages could intensely exacerbate imbalances of ATP production [49]. AMPK can improve energy status by promoting the ATP production pathway and/or by inhibiting the ATP consumption pathway, when the cellular situation would be extremely stirred by external factors [50]. In addition, AMPK can control mitochondrial biogenesis through the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α) signaling pathway, which could boost mitochondrial oxidative metabolism in ischemic conditions [51]. AMPK activation directs to significant inhibition of the mTOR signaling pathway, which successfully decreases the apoptosis of cells [52]. Additionally, AMPK can also arise levels of autophagy via the autophagy activating kinase 1 (ULK1) and partially by inhibiting transforming growth factor beta (TGF-β) signaling [53]. As a key regulator of cellular metabolism, AMPK may keep energy homeostasis at both cellular and physical levels during metabolic and/or oxidative stresses [54]. As mentioned above, AMPK is generally recognized as an accurate energy sensor due to its critical role in regulating the pathways of energy production and consumption in cells and/or body [55]. In addition, AMPK could also control various other cellular processes including the inhibition of protein synthesis, cell proliferation, and DNA repair [56]. AMPK might exert several protective effects on the heart by controlling energy homeostasis [57]. Also, AMPK could perform as an imperative endogenous protection character that can punctually respond to damaging stimuli such as cardiac ischemia [58]. Remarkably, AMPK can promote glucose and/or fat oxidation, thereby, which could powerfully improve energy metabolism [59]. Therefore, these distinctive regulatory roles of AMPK in diverse tissues and/or organs may be essential for sustaining normal physiological function [60]. Unquestionably, AMPK activation should be assumed to play a substantial cardioprotective role against cardiotoxicity, which is faithfully associated with the cardiac remodeling process via the mitophagy and/or autophagy.
4. Some Molecules Involved in the Regulation of AMPK
Metformin upregulates AMPK to induce autophagy and thus improves severe complications of diabetes, including cardiac remodeling and/or heart failure [61]. Metformin can inhibit complex I of the electron transport chain to increase the cellular level of AMP, increasing the AMP/ATP ratio, which may modulate AMPK exerting physiological effects [62]. Zinc can also modulate mitophagy to suppress the production of superoxide in mitochondria to alleviate cardiac injury by upregulating PINK1 and Beclin1 [63]. However, zinc overload may increase ROS levels, leading to cell death and impaired mitochondrial function and dynamics [64]. Statins, the most effective lipid/cholesterol lowering drugs, are commonly used in the treatment of hyperlipidemia. Statins can also activate AMPK, thereby affecting cell autophagy through the AMPK/mTOR signaling pathway [65]. Osmotin, a protein present in tobacco, is structurally and functionally similar to adiponectin, which can also activate AMPK. Interestingly, osmotin can protect mice from Aβ-induced memory impairment and neurodegeneration by reducing cholesterol biosynthesis and ameliorating the pathology of Alzheimer’s disease [66]. Ghrelin, a gastric hormone released during fasting, targets the hypothalamus to encourage starvation. The ghrelin can reduce triglyceride production and controls the AMPK and the mTOR signaling pathways to induce autophagy by repairing AMPK phosphorylation and/or preventing mTOR phosphorylation [67]. The ghrelin can activate the AMPK in hypothalamus of brain, which subsequently stimulate glucose metabolism for energy production, thereby preventing the decline of memory function in rats [68]. Resveratrol can also activate the AMPK by increasing intracellular calcium levels and promotes CaMKKβ-mediated AMPK phosphorylation, which might inhibit the mTOR-induced autophagy to reduce Aβ levels in the brain of Alzheimer’s disease. Similarly, resveratrol analogs could also activate the AMPK by promoting CaMKKβ-dependent manner, promoting autophagy to reduce Aβ levels in the brain of Alzheimer’s disease[69]. Polygalae radix, a well-known traditional Chinese medicine, can likewise induce autophagy by activating the AMPK/mTOR pathway, thereby reducing Aβ levels of Alzheimer’s disease [70]. Instead of exerting a helpful effect, however, the mitophagy activation may also be harmful to disease progression. For example, peroxynitrite treatment may worsen cerebral injury in stroke through the induction of mitophagy [71]. Estrogen could activate the phosphorylation of AMPK and PGC1α via the function of estrogen receptor, which can amend mitochondrial respiration in skeletal muscle cells in vitro [72]. Salsalate is a prodrug of salicylate, which is similar to acetylsalicylic acid, aspirin. Salicylate can stimulate the AMPK through the direct interaction with a serine residue of AMPK subunit [73]. It has been revealed that cellular effects of Rosmarinus officinalis (rosemary) extract are associated with an escalation in levels of AMPK phosphorylation in lung cancer cells [74]. In addition, honokiol treatment could activate the AMPK in kidney tubular cells [75]. A small molecule 2,6-disubstituted purine, reversine, can activate the autophagy via upregulation of AMPK in senescent myoblast cells [76]. Leptin is a peptide hormone secreted from fat cells, which can selectively phosphorylate the AMPK in skeletal muscle cells [77]. Adiponectin can also activate the AMPK signaling pathway through various mechanisms including by the activation of CaMKKβ [78]. Additionally, adiponectin could bind to α subunit of AMPK to stimulate AMPK heteromerization [79]. Adiponectin can inhibit fatty acid synthesis in obese cells reducing ATP consumption, which may activate the AMPK [80]. Moreover, adiponectin could increase levels of intracellular NAD+, which can indirectly activate the AMPK [81]. Furthermore, adiponectin could increase liver glycogenase activity, which also enhances NADH production via glycolysis, indirectly activating the AMPK[82]. As a final point, intermittent fasting and ketogenic diets may also activate the AMPK and/or inhibit the mTOR pathway, contributing the suppression of oxidative stress and/or inflammation levels through boosting mitochondrial plasticity [83].
5. Possible Treatment Tactics with Certain Small Molecules against Several Heart Diseases
Given the critical role of mitophagy in the pathogenesis of heart disease, mitophagy acts as a promising therapeutic target. The promotion or inhibition of mitophagy could be used to treat the development of heart diseases. In other words, several inhibitors such as metformin could target AMPK pathway, thereby mediating the modulation of mitophagy for the treatment of heart diseases. Beneficial effects on the heart diseases might also be related to the regulation of the cellular antioxidant system as well as mitophagy. Remarkably, modulation of mitophagy may be advantageous both in improving cardiac function and/or in treating several cardiomyopathy. For example, recent studies in animal models of an aging heart have shown that fasting-induced autophagy may improve the cardiac function and/or longevity, which may also be related to autophagic clearance of impaired cellular components [84]. Energy deprivation activates the AMPK as cellular energy sensor, which may successively induce the autophagy. Fasting can also release the ghrelin which can activate the AMPK, as mentioned before. Additionally, the metformin administration could prevent cardiomyopathy by activating AMPK and mitophagy in diabetic mice [85]. Treatment with the metformin could also decrease cardiovascular inflammation via increasing AMPK and/or reducing ROS [86]. Some statins have been recognized to make available a protective effect of heart, which could also reduce the severity of myocardial injury [87]. In addition, arachidonic acid could protect against ischemic myocardial damage by endorsing mitochondrial autophagy and/or mitochondrial renewal [88]. The polyamine spermidine can stimulate both mitophagy and mitochondrial biogenesis, and these are associated with improved cardiac function and prolonged lifespan of individuals. Spermidine treatment can offer a beneficial effect on cardiac mitochondrial alterations associated with aging [89]. Polyamines, as spermidine and spermine, are aliphatic cations necessary for cell proliferation and/or differentiation, with anti-inflammatory and/or anti-apoptotic effects. They are now evolving as natural autophagy regulators with anti-aging properties. Interestingly, polyamine levels were considerably changed in the skeletal muscles of aged animals [90]. Furthermore, spermine could prevent hypoxia-induced ROS production and the reduction of mitophagy in cardiomyocytes [91]. Berberine has a therapeutic effect on cardiovascular health [92]. Berberine can protect against myocardial injury by inducing the mitophagy. Therefore, berberine may serve as a novel therapeutic drug for myocardial injury [93]. In addition, berberine can diminish mitochondrial dysfunction by inducing autophagy, which may contribute to the key cardioprotective mechanisms of berberine [94]. Similarly, treatment of honokiol could improve myocardial injury, which may be significant for the cardioprotective role of honokiol by enhancing autophagy [95]. Carnosic acid, the principal constituent of rosemary, has various biological activities, including anti-inflammatory and antioxidant, which could be possible against cardiotoxicity via the modulation of autophagy and/or apoptosis [96]. Elevation in butyrate levels might enhance mitochondrial biogenesis and oxygen consumption and may increase resistance to oxidative/redox stress, hence resulting in improvement of cardiac function [97]. The protective effect of resveratrol on hypertension-induced cardiac dysfunction has been reported by modulating cellular stress responses and/or mitophagy [98]. In addition, resveratrol could improve cardiomyopathy by promoting autophagy/mitophagy in the heart of Duchenne muscular dystrophy (DMD) model mouse [99]. Resveratrol can protect mitochondria by modulating mitophagy through the alteration of AMPK signaling pathway in myocardial cells [100]. Furthermore, resveratrol can inhibit hyperglycemia-induced cardiomyocyte hypertrophy by reducing oxidative stress via stimulating mitophagy to keep mitochondrial homeostasis [101]. Interestingly, resveratrol can dilate retinal arterioles and can protect against retinal neurodegeneration via the AMPK-dependent signaling pathway [102]. (Figure 3)
6. Future Perspectives
The abnormal cardiac function due to the damages of mitochondria may remain the main pathogenesis in several heart diseases. Therefore, mitochondrial autophagy/mitophagy might be a hot subject among the molecular mechanisms of organelle-specific autophagy. The induction of mitophagy can strongly alleviate damaged mitochondria and keep suitable mitochondria for normal cellular homeostasis. The mechanisms for the mitophagy are complicated and firmly regulated by multiple interlinked factors. These factors may function during important biological processes, including cell growth, cell differentiation, cell senescence, and cell death. Aberrant regulation of mitophagy may affect normal cellular function and also influence several heart diseases, thereby providing mitophagy a promising therapeutic target for those heart diseases.
Interestingly, it has been shown that AMPK is required for cofilin activation. pyridoxal phosphatase (PDXP) is known to directly dephosphorylate the cofilin, which may be involved in mitochondrial fission and/or mitophagy [103]. Cofilin signaling pathway induces the activation of TGFβ and/or PDGF pathway for the vascularization in pluripotent stem cell-derived cardiac organoids [104]. Some researchers have revealed that stem cells can repair damaged myocardium and/or improve heart function [105]. Therefore, AMPK and PDXP might be a key regulator for the regeneration of vascularization and/or cardiomyocytes.
Continual progress has been made in identifying the constituents of natural plant products that are cardioprotective by modulating AMPK signaling pathway. However, clinical and/or preclinical trials would be mandatory to confirm the safety and/or efficacy of the plant metabolites for the treatment and/or prevention of several heart diseases. Whether the induction or inhibition of autophagy/mitophagy are helpful for the pathogenesis of heart disease remains to be more precisely understood. It is possible that mitophagy may be contributed by the distinctive characteristics of altered molecules. Future studies are also indispensable for the explanation of various effects of various molecules involved in the induction of mitophagy in several heart diseases. Further comprehension into the molecular mechanisms of mitophagy/autophagy in heart could facilitate in understanding how to prevent cardiac disorders from fundamental causing.
7. Conclusion
The damaged mitochondria may play an important role in maintaining the normal function of cardiovascular cells. An in-depth knowledge of the role of AMPK and autophagy/mitophagy in heart diseases may be valuable for developing new clinical treatment. This review may provide insight to support future research into understanding this AMPK signaling axis in cardiac disorders.
Author Contributions
Conceptualization, MN, NS, and SM; original draft preparation and editing, MN, NS, and SM; visualization, NS and SM; supervision, SM. Each author (MN, NS, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
| AMP | adenosine monophosphate |
| ATP | adenosine triphosphate |
| AMPK | adenosine monophosphate-activated protein kinase |
| FMT | fecal microbiota transplantation |
| mTOR | mechanistic/mammalian target of rapamycin |
| PDXP | pyridoxal phosphatase |
| PGC1α | peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| QOL | quality of life |
| ROS | reactive oxygen species |
| TGF-β | transforming growth factor beta |
| ULK1 | autophagy activating kinase 1 |
References
- Zhang, Z.; Yang, X.; Song, Y.Q.; Tu, J. Autophagy in Alzheimer's disease pathogenesis: therapeutic potential and future perspectives. Ageing Res. Rev. 2021, 72, 101464. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Hariharan, N.; Monden, Y.; Zablocki, D.; Sadoshima, J. Is autophagy in response to ischemia and reperfusion protective or detrimental for the heart? Pediatr. Cardiol. 2011, 32, 275–281. [Google Scholar]
- Aventaggiato, M.; Vernucci, E.; Barreca, F.; Russo, M.A.; Tafani, M. Sirtuins’ control of autophagy and mitophagy in cancer. Pharmacol. Ther. 2021, 221, 107748. [Google Scholar] [CrossRef] [PubMed]
- Hebah, H.A.; Kamel, H.M.; Bastawy, I.M.; Ahmed, F.A. Association between serum beclin 1 level and cardiac valvular calcification in hemodialysis patients. Curr Probl Cardiol. 2024, 49, 102519. [Google Scholar] [CrossRef]
- Nodaand, N.N. , Inagaki, F. Mechanisms of autophagy. Annual Review of Biophysics. 2015, 44, 101–122. [Google Scholar]
- Sharma, V.; Verma, S.; Seranova, E.; Sarkar, S.; Kumar, D. Selective Autophagy and Xenophagy in Infection and Disease. Front. Cell Dev. Biol. 2018, 6, 147. [Google Scholar] [CrossRef]
- Bravo-San, Pedro, J. M.; Kroemer, G.; Galluzzi, L. Autophagy and Mitophagy in Cardiovascular Disease. Circ. Res. 2017, 120, 1812–1824. [Google Scholar] [CrossRef]
- Kim, I.; Lemasters, J.J. Mitophagy selectively degrades individual damaged mitochondria after photoirradiation. Antioxidants & Redox Signaling. 2011, 14, 1919–1928. [Google Scholar]
- Doblado, L.; Lueck, C.; Rey, C.; Samhan-Arias, A.K.; Prieto, I.; Stacchiotti, A.; Monsalve, M. Mitophagy in human diseases. International Journal of Molecular Sciences. 2021, 22, 3903. [Google Scholar] [CrossRef] [PubMed]
- Billia, F.; Hauck, L.; Konecny, F.; Rao, V.; Shen, J.; Mak, T.W. PTEN-inducible kinase 1 (PINK1)/Park6 is indispensable for normal heart function. Proceedings of the National Academy of Sciences of the United States of America. 2011, 108, 9572–9577. [Google Scholar] [CrossRef]
- De, Gaetano, A. ; Gibellini, L.; Zanini, G.; Nasi, M.; Cossarizza, A.; Pinti, M. Mitophagy and oxidative stress: the role of aging. Antioxidants. 2021, 10, 794.
- Vasquez-Trincado, C.; Garcia-Carvajal, I.; Pennanen, C.; Parra, V.; Hill, J.A.; Rothermel, B.A.; Lavandero, S. Mitochondrial dynamics, mitophagy and cardiovascular disease. The Journal of Physiology. 2016, 594, 509–525. [Google Scholar] [CrossRef]
- Jarosz, J.; Ghosh, S.; Delbridge, L.M.D.; Petzer, A.; Hickey, A.J.R.; Crampin, E.J.; Hanssen, E.; Rajagopal, V. Changes in mitochondrial morphology and organization can enhance energy supply from mitochondrial oxidative phosphorylation in diabetic cardiomyopathy. Am J Physiol-Cell Physiol. 2017, 312, C190–C197. [Google Scholar] [CrossRef] [PubMed]
- Yi, D.; Yu, H.; Lu, K.; Ruan, C.; Ding, C.; Tong, L.; Zhao, X.; Chen, D. AMPK Signaling in Energy Control, Cartilage Biology, and Osteoarthritis. Front Cell Dev Biol. 2021, 9, 696602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, Y.; Guo, D.; Luo, M.; Zhang, Q.; Zhang, L.; Zhang, D. Exercise Improves Heart Function after Myocardial Infarction: The Merits of AMPK. Cardiovasc Drugs Ther. 2024. [CrossRef] [PubMed]
- Garza-González, S.; Nieblas, B.; Solbes-Gochicoa, M.M.; Altamirano, J.; García, N. Intermittent Fasting as Possible Treatment for Heart Failure. Curr Vasc Pharmacol. 2022, 20, 260–271. [Google Scholar] [CrossRef]
- Parvaresh, H.; Paczek, K.; Al-Bari, M.A.A.; Eid, N. Mechanistic insights into fasting-induced autophagy in the aging heart. World J Cardiol. 2024, 16, 109–117. [Google Scholar] [CrossRef]
- Wohlgemuth, S.E.; Julian, D.; Akin, D.E.; Fried, J.; Toscano, K.; Leeuwenburgh, C.; Dunn, W.A. Jr. Autophagy in the heart and liver during normal aging and calorie restriction. Rejuvenation Res. 2007, 10, 281–292. [Google Scholar] [CrossRef]
- Nunes, J.P.S.; Andrieux, P.; Brochet, P.; Almeida, R.R.; Kitano, E.; Honda, A.K.; Iwai, L.K.; Andrade-Silva, D.; et al. Co-exposure of cardiomyocytes to IFN-γ and TNF-α induces mitochondrial dysfunction and nitro-oxidative stress: implications for the pathogenesis of chronic chagas disease cardiomyopathy. Front Immunol. 2021, 12, 755862. [Google Scholar] [CrossRef]
- Morales, P.E.; Arias-Duran, C.; Avalos-Guajardo, Y.; Aedo, G.; Verdejo, H.E.; Parra, V.; Lavandero, S. Emerging role of mitophagy in cardiovascular physiology and pathology. Molecular Aspects of Medicine. 2020, 71, 100822. [Google Scholar] [CrossRef]
- Seidlmayer, L.K.; Kuhn, J.; Berbner, A.; Arias-Loza, P.A.; Williams, T.; Kaspar, M.; Czolbe, M.; Kwong, J.Q.; et al. Inositol 1,4,5-trisphosphate-mediated sarcoplasmic reticulum-mitochondrial crosstalk influences adenosine triphosphate production via mitochondrial Ca2+ uptake through the mitochondrial ryanodine receptor in cardiac myocytes. Cardiovasc Res. 2016, 112, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Zweier, J.L. Cardiac mitochondria and reactive oxygen species generation. Circulation Research. 2014, 114, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, H.; Dudley, S.C. Jr. Reactive oxygen species originating from mitochondria regulate the cardiac sodium channel. Circulation Research. 2010, 107, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: impact on human health. Pharmacognosy Reviews. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Geng, L.; Ying, L.; Shu, L.; Ye, K.; Yang, R.; Liu, Y.; Wang, Y.; et al. FGF21-Sirtuin 3 axis confers the protective effects of exercise against diabetic cardiomyopathy by governing mitochondrial integrity. Circulation. 2022, 146, 1537–1557. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Krigman, J.; Luo, H.; Ozgen, S.; Yang, M.; Sun, N. Mitophagy in cardiovascular homeostasis. Mechanisms of Ageing and Development. 2020, 188, 111245. [Google Scholar] [CrossRef] [PubMed]
- Akar, F.G.; O'Rourke, B. Mitochondria are sources of metabolic sink and arrhythmias. Pharmacology & Therapeutics. 2011, 131, 287–294. [Google Scholar]
- Phadwal, K.; Vrahnas, C.; Ganley, I.G.; MacRae, V.E. Mitochondrial dysfunction: cause or consequence of vascular calcification? Frontiers in Cell and Developmental Biology. 2021, 9, 611922. [Google Scholar] [CrossRef]
- Brown, D.A.; O'Rourke, B. Cardiac mitochondria and arrhythmias. Cardiovascular Research. 2010, 88, 241–249. [Google Scholar] [CrossRef]
- Wong, A.K.; Howie, J.; Petrie, J.R.; Lang, C.C. AMP-activated protein kinase pathway: a potential therapeutic target in cardiometabolic disease. Clin. Sci. 2009, 116, 607–620. [Google Scholar] [CrossRef]
- Zhang, W.; Ren, H.; Xu, C.; Zhu, C.; Wu, H.; Liu, D.; Wang, J.; Liu, L.; et al. Hypoxic mitophagy regulates mitochondrial quality and platelet activation and determines severity of I/R heart injury. eLife. 2016, 5, e21407. [Google Scholar] [CrossRef]
- Jamialahmadi, T.; Hasanpour, M.; Vakilian, F.; Penson, P.E.; Iranshahy, M.; Sahebkar, A. Evaluation of Urolithin A Efficacy in Heart Failure Patients with Reduced Ejection Fraction: A Randomized, Double-blind, Crossover, Placebo-controlled Clinical Trial. Rev Recent Clin Trials. 2024. [CrossRef] [PubMed]
- Hu, X.; Xu, X.; Lu, Z.; Zhang, P.; Fassett, J.; Zhang, Y.; Xin, Y.; Hall, J.L.; et al. AMP activated protein Kinase-α2 regulates expression of estrogen-related Receptor-α, a metabolic transcription factor related to heart failure development. Hypertension. 2011, 58, 696–703. [Google Scholar] [CrossRef]
- Li, H.; Zheng, F.; Zhang, Y.; Sun, J.; Gao, F.; Shi, G. Resveratrol, novel application by preconditioning to attenuate myocardial ischemia/reperfusion injury in mice through regulate AMPK pathway and autophagy level. J. Cell. Mol. Med. 2022, 26, 4216–4229. [Google Scholar] [CrossRef]
- Shirakabe, A.; Zhai, P.; Ikeda, Y.; Saito, T.; Maejima, Y.; Hsu, C.P.; Nomura, M.; Egashira, K.; et al. Drp1-Dependent Mitochondrial Autophagy Plays a Protective Role Against Pressure Overload-Induced Mitochondrial Dysfunction and Heart Failure. Circulation. 2016, 133, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.B.; Gao, W.C.; Xie, M.; Li, Z.; Ma, X.; Song, W.; Luo, D.; Huang, Y.; et al. Ang II Promotes Cardiac Autophagy and Hypertrophy via Orai1/STIM1. Front. Pharmacol. 2021, 12, 622774. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, K.; Gao, C.; Ma, W.; Liu, M.; Guo, X.; Bao, G.; Han, B.; et al. Activation of FMS-like tyrosine kinase 3 protects against isoprenaline-induced cardiac hypertrophy by improving autophagy and mitochondrial dynamics. FASEB J. 2022, 36, e22672. [Google Scholar] [CrossRef] [PubMed]
- Keerthana, C.K.; Rayginia, T.P.; Shifana, S.C.; Anto, N.P.; Kalimuthu, K.; Isakov, N, Anto, R. J. The role of AMPK in cancer metabolism and its impact on the immunomodulation of the tumor microenvironment. Front. Immunol. 2023, 14, 1114582. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Y.; Jiang, T.; Liu, B.; Sun, H.; Zhang, Y.; Fan, B.; Li, X.; et al. Enhancing fatty acids oxidation via L-carnitine attenuates obesity-related atrial fibrillation and structural remodeling by activating AMPK signaling and alleviating cardiac lipotoxicity. Front. Pharmacol. 2021, 12, 771940. [Google Scholar] [CrossRef]
- Lee, C.C.; Chen, W.T.; Chen, S.; Lee, T.M. Dapagliflozin attenuates arrhythmic vulnerabilities by regulating connexin43 expression via the AMPK pathway in post-infarcted rat hearts. Biochem. Pharmacol. 2021, 192, 114674. [Google Scholar] [CrossRef]
- Zhang, M.; Alemasi, A.; Zhao, M.; Xu, W.; Zhang, Y.; Gao, W.; Yu, H.; Xiao, H. Exercise Training Attenuates Acute beta-Adrenergic Receptor Activation-Induced Cardiac Inflammation via the Activation of AMP-Activated Protein Kinase. Int J Mol Sci. 2023, 24, 9263. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, H.; Yang, Y.; Jiang, Z.; Ma, H. Dehydroepiandrosterone activates the GPER-mediated AMPK signaling pathway to alleviate the oxidative stress and inflammatory response in laying hens fed with high-energy and low-protein diets. Life Sci. 2022, 308, 120926. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Shaw, R.J. AMPK: mechanisms of cellular energy sensing and restoration of metabolic balance. Mol. Cell. 2017, 66, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, G.; Kim, Y.; Kim, J.; Ha, J. AMPK activators: mechanisms of action and physiological activities. Exp. Mol. Med. 2016, 48, e224. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Muraleedharan, R.; Dasgupta, B. AMPK in the brain: its roles in glucose and neural metabolism. Febs J. 2022, 289, 2247–2262. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, C.; Dong, B.; Xing, F.; Huang, H.; Yao, F.; Ma, Y.; He, J.; et al. AMPK attenuates ventricular remodeling and dysfunction following aortic banding in mice via the Sirt3/Oxidative stress pathway. Eur. J. Pharmacol. 2017, 814, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; et al. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature. 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Pagan, L.U.; Gomes, M.J.; Gatto, M.; Mota, G.A.; Okoshi, K.; Okoshi, M.P. The role of oxidative stress in the aging heart. Antioxidants. 2022, 11, 336. [Google Scholar] [CrossRef]
- Trefts, E; Shaw, R. J. AMPK: restoring metabolic homeostasis over space and time. Mol Cell. 2021, 81, 3677–3690. [Google Scholar] [CrossRef]
- Qi, X.; Wang, J. Melatonin improves mitochondrial biogenesis through the AMPK/PGC1α pathway to attenuate ischemia/reperfusion-induced myocardial damage. Aging. 2020, 12, 7299–7312. [Google Scholar] [CrossRef]
- Li, S.X.; Li, C.; Pang, X.R.; Zhang, J.; Yu, G.C.; Yeo, A.J.; Lavin, M.F.; Shao, H.; et al. Metformin attenuates silica-induced pulmonary fibrosis by activating autophagy via the AMPK-mTOR signaling pathway. Front Pharmacol. 2021, 12, 719589. [Google Scholar] [CrossRef]
- Timm, K.N.; Tyler, D.J. The role of AMPK activation for cardioprotection in doxorubicin-induced cardiotoxicity. Cardiovasc Drugs Ther. 2020, 34, 255–269. [Google Scholar] [CrossRef]
- González, A.; Hall, M.N.; Lin, S.C.; Hardie, D.G. AMPK and TOR: the Yin and Yang of cellular nutrient sensing and growth control. Cell Metab. 2020, 31, 472–492. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Hardie, D.G. New insights into activation and function of the AMPK. Nat Rev Mol Cell Biol. 2023, 24, 255–272. [Google Scholar] [CrossRef]
- Maharajan, N.; Ganesan, C.D.; Moon, C.; Jang, C.H.; Oh, W.K.; Cho, G.W. Licochalcone D ameliorates oxidative stress-induced senescence via AMPK activation. Int J Mol Sci. 2021, 22, 7324. [Google Scholar] [CrossRef]
- Sharma, A.; Anand, S.K.; Singh, N.; Dwivedi, U.N.; Kakkar, P. AMP-activated protein kinase: an energy sensor and survival mechanism in the reinstatement of metabolic homeostasis. Exp Cell Res. 2023, 428, 113614. [Google Scholar] [CrossRef]
- Hu, Y.; Dong, Y.D.; Wu, Y.C.; Wang, Q.X.; Nan, X.; Wang, D.L. AMPK inhibitor BML-275 induces neuroprotection through decreasing cyt c and AIF expression after transient brain ischemia. Bioorg Med Chem. 2021, 52, 116522. [Google Scholar] [CrossRef]
- Spaulding, HR.; Yan, Z. AMPK and the adaptation to exercise. Annu Rev Physiol. 2022, 84, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Li, Y.; Fu, Y.; Yang, Y. Research progress of AMP-activated protein kinase and cardiac aging. Open Life Sci. 2023, 18, 20220710. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Hao, R.; Wang, W.; Gao, H.; Wang, C. SIRT1/Atg5/autophagy are involved in the antiatherosclerosis effects of ursolic acid. Mol. Cell. Biochem. 2016, 420, 171–184. [Google Scholar] [CrossRef] [PubMed]
- LaMoia, TE.; Shulman, GI. Cellular and molecular mechanisms of metformin action. Endocr. Rev. 2021, 42, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, T.; Li, Z.; Liu, N.; Yan, Y.; Liu, B. Role of Mitophagy in cardiovascular disease. Aging and Disease. 2020, 11, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, P.; Guo, J.; Ma, T.; Hu, Y.; Huang, L.; Xing, B.; He, Y.; et al. Zinc Overload Induces Damage to H9c2 Cardiomyocyte Through Mitochondrial Dysfunction and ROS-Mediated Mitophagy. Cardiovasc Toxicol. 2023, 23, 388–405. [Google Scholar] [CrossRef]
- Sun, W.; Lee, T.S.; Zhu, M.; Gu, C.; Wang, Y.; Zhu, Y.; Shyy, J.Y. Statins activate AMP-activated protein kinase in vitro and in vivo. Circulation. 2006, 114, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Yoon, G.H.; Chung, S.S.; Abid, M.N.; Kim, T.H.; Lee, H.Y.; Kim, M.O. Novel osmotin inhibits SREBP2 via the AdipoR1/AMPK/SIRT1 pathway to improve Alzheimer's disease neuropathological deficits. Mol. Psychiatry. 2017, 22, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Cheng, J.; Yu, F.; Li, H.; Guo, C.; Fan, X. Ghrelin attenuated lipotoxicity via autophagy induction and nuclear factor-κB inhibition. Cell Physiol. Biochem 2015, 37, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Moon, N.R.; Kim, D.S.; Kim, S.H.; Park, S. Central acylated ghrelin improves memory function and hippocampal AMPK activation and partly reverses the impairment of energy and glucose metabolism in rats infused with β-amyloid. Peptides. 2015, 71, 84–93. [Google Scholar] [CrossRef]
- Vingtdeux, V.; Giliberto, L.; Zhao, H.; Chandakkar, P.; Wu, Q.; Simon, J.E.; Janle, E.M.; Lobo, J.; et al. AMP-activated protein kinase signaling activation by resveratrol modulates amyloid-beta peptide metabolism. J. Biol. Chem. 2010, 285, 9100–9113. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Z.C.; Wang, K.F.; Chen, X.Y. Aβ peptide secretion is reduced by Radix Polygalae-induced autophagy via activation of the AMPK/mTOR pathway. Mol. Med. Rep. 2015, 12, 2771–2776. [Google Scholar] [CrossRef]
- Feng, J.; Chen, X.; Guan, B.; Li, C.; Qiu, J.; Shen, J. Inhibition of peroxynitrite-induced mitophagy activation attenuates cerebral ischemia-reperfusion injury. Molecular Neurobiology. 2018, 55, 6369–6386. [Google Scholar] [CrossRef]
- Fu, Q.; Li, T.; Zhang, C.; Ma, X.; Meng, L.; Liu, L.; Shao, K.; Wu, G.; et al. Butyrate mitigates metabolic dysfunctions via the ERalpha-AMPK pathway in muscle in OVX mice with diet-induced obesity. Cell Commun Signal. 2023, 21, 95. [Google Scholar] [CrossRef] [PubMed]
- Hawley, S.A.; Fullerton, M.D.; Ross, F.A.; Schertzer, J.D.; Chevtzoff, C.; Walker, K.J.; Peggie, M.W.; Zibrova, D.; et al. The ancient drug salicylate directly activates AMP-activated protein kinase. Science. 2012, 336, 918–922. [Google Scholar] [CrossRef]
- O'Neill, E.J.; Moore, J.; Song, J.; Tsiani, E.L. Inhibition of Non-Small Cell Lung Cancer Proliferation and Survival by Rosemary Extract Is Associated with Activation of ERK and AMPK. Life (Basel). 2021, 12, 52. [Google Scholar] [CrossRef]
- Mao, R.W.; He, S.P.; Lan, J.G.; Zhu, W.Z. Honokiol ameliorates cisplatin-induced acute kidney injury via inhibition of mitochondrial fission. Br J Pharmacol. 2022, 179, 3886–3904. [Google Scholar] [CrossRef]
- Rajabian, N.; Choudhury, D.; Ikhapoh, I.; Saha, S.; Kalyankar, A.S.; Mehrotra, P.; Shahini, A.; Breed, K.; et al. Reversine ameliorates hallmarks of cellular senescence in human skeletal myoblasts via reactivation of autophagy. Aging Cell. 2023, 22, 13764. [Google Scholar] [CrossRef] [PubMed]
- Kandadi, M.R.; Roe, N.D.; Ren, J. Autophagy inhibition rescues against leptin-induced cardiac contractile dysfunction. Curr. Pharm. Des. 2014, 20, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Xu, X.; Wang, Q.; Ren, S.Y.; Dong, M.; Zhang, Y. Permissive role of AMPK and autophagy in adiponectin deficiency-accentuated myocardial injury and inflammation in endotoxemia. J. Mol. Cell Cardiol. 2016, 93, 18–31. [Google Scholar] [CrossRef]
- Fu, Z.; Liegl, R.; Wang, Z.; Gong, Y.; Liu, C.H.; Sun, Y.; Cakir, B.; Burnim, S.B.; et al. Adiponectin mediates dietary omega-3 long-chain polyunsaturated fatty acid protection against choroidal neovascularization in mice. Invest Ophthalmol. Vis. Sci. 2017, 58, 3862–3870. [Google Scholar]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Bungau, S.G.; Radu, A.F.; Batiha, G.E. The potential molecular implications of adiponectin in the evolution of SARS-CoV-2: inbuilt tendency. J. King Saud. Univ. Sci. 2022, 34, 102347. [Google Scholar] [CrossRef]
- Guo, Z.; Yan, X.; Wang, L.; Wu, J.; Jing, X.; Liu, J. Effect of Telmisartan or Insulin on the Expression of Adiponectin and its Receptors in the Testis of Streptozotocin-Induced Diabetic Rats. Horm. Metab. Res. 2016, 48, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tan, J.X.; He, Y.; Bai, F.; Li, S.W.; Hou, Y.W.; Ji, L.S.; Gao, Y.T.; et al. Atractylenolide III ameliorates Non-Alcoholic Fatty Liver Disease by activating Hepatic Adiponectin Receptor 1-Mediated AMPK Pathway. Int J. Biol. Sci. 2022, 18, 1594–1611. [Google Scholar]
- Paoli, A.; Tinsley, G.M.; Mattson, M.P.; De, Vivo, I. ; Dhawan, R.; Moro, T. Common and divergent molecular mechanisms of fasting and ketogenic diets. Trends Endocrinol Metab. 2024, 35, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Cavezzi, A.; Colucci, R.; d'Errico, G. Mitoresilience: Hormesis, Psycho-physical Resilience, Mitochondria and Heart Rate Variability as Relevant Interplaying Elements in Longevity Medicine. Curr Aging Sci. 2023, 16, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Lau, K.; Eby, B.; Lozano, P.; He, C.; Pennington, B.; Li, H.; Rathi, S.; et al. Improvement of cardiac functions by chronic metformin treatment is associated with enhanced cardiac autophagy in diabetic OVE26 mice. Diabetes. 2011, 60, 1770–1778. [Google Scholar] [CrossRef] [PubMed]
- Marek-Iannucci, S.; Ozdemir, A.B.; Moreira, D.; Gomez, A.C.; Lane, M.; Porritt, R.A.; Lee, Y.; Shimada, K.; et al. Autophagy-mitophagy induction attenuates cardiovascular inflammation in a murine model of Kawasaki disease vasculitis. JCI Insight. 2021, 6, e151981. [Google Scholar] [CrossRef] [PubMed]
- Bland, A.R.; Payne, F.M.; Ashton, J.C.; Jamialahmadi, T.; Sahebkar, A. The cardioprotective actions of statins in targeting mitochondrial dysfunction associated with myocardial ischaemia-reperfusion injury. Pharmacol Res. 2022, 175, 105986. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Li, K.; Wang, S.; Wang, X.; Yue, G.; Zhang, Y.; Lv, X.; Zhao, P.; et al. Protective role of arachidonic acid against diabetic myocardial ischemic injury: a translational study of pigs, rats, and humans. Cardiovasc Diabetol. 2024, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Messerer, J.; Wrede, C.; Schipke, J.; Brandenberger, C.; Abdellatif, M.; Eisenberg, T.; Madeo, F.; Sedej, S.; et al. Spermidine supplementation influences mitochondrial number and morphology in the heart of aged mice. Mühlfeld C.J Anat. 2023, 242, 91–101. [Google Scholar] [CrossRef]
- Galasso, L.; Cappella, A.; Mulè, A.; Castelli, L.; Ciorciari, A.; Stacchiotti, A.; Montaruli, A. Polyamines and Physical Activity in Musculoskeletal Diseases: A Potential Therapeutic Challenge. Int J Mol Sci. 2023, 24, 9798. [Google Scholar] [CrossRef]
- Chai, N.; Zheng, H.; Zhang, H.; Li, L.; Yu, X.; Wang, L.; Bi, X.; Yang, L.; et al. Spermidine Alleviates Intrauterine Hypoxia-Induced Offspring Newborn Myocardial Mitochondrial Damage in Rats by Inhibiting Oxidative Stress and Regulating Mitochondrial Quality Control. Iran J Pharm Res. 2023, 21, e133776. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, T.; Mei, L.; Zhang, Y.; Liang, C.; Bai, X.; Zhang, Z.; Shi, Y.; et al. The Potential of Berberine to Target Telocytes in Rabbit Heart. Planta Med. 2024, 90, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Li, J.; Li, Y.; Zhang, Y.; Du, Q.; Hao, P.; Li, J.; Cao, X.; et al. Berberine Protects Against Simulated Ischemia/Reperfusion Injury-Induced H9C2 Cardiomyocytes Apoptosis In Vitro and Myocardial Ischemia/Reperfusion-Induced Apoptosis In Vivo by Regulating the Mitophagy-Mediated HIF-1alpha/BNIP3 Pathway. Front Pharmacol. 2020, 11, 367. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Cao, X.; Hao, P.; Zhang, Y.; Chen, Y.; Zhang, J.; Li, J.; Gao, C.; et al. Berberine attenuates mitochondrial dysfunction by inducing autophagic flux in myocardial hypoxia/reoxygenation injury. Cell Stress Chaperones. 2020, 25, 417–426. [Google Scholar] [CrossRef]
- Tan, Z.; Liu, H.; Song, X.; Ling, Y.; He, S.; Yan, Y.; Yan, J.; Wang, S.; et al. Honokiol post-treatment ameliorates myocardial ischemia/reperfusion injury by enhancing autophagic flux and reducing intracellular ROS production. Chem Biol Interact. 2019, 307, 82–90. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Yang, J.J.; Zhang, H.S. Carvedilol (CAR) combined with carnosic acid (CAA) attenuates doxorubicin-induced cardiotoxicity by suppressing excessive oxidative stress, inflammation, apoptosis and autophagy. Biomed Pharmacother. 2019, 109, 71–83. [Google Scholar] [CrossRef]
- Thai, P.N.; Miller, C.V.; King, M.T.; Schaefer, S.; Veech, R.L.; Chiamvimonvat, N.; Bers, D.M.; Dedkova, E.N. Ketone Ester D-beta-Hydroxybutyrate-(R)-1,3 Butanediol Prevents Decline in Cardiac Function in Type 2 Diabetic Mice. J Am Heart Assoc. 2021, 10, e020729. [Google Scholar] [CrossRef]
- Bal, N.B.; Bostanci, A.; Sadi, G.; Dönmez, M.O.; Uludag, M.O.; Demirel-Yilmaz, E. Resveratrol and regular exercise may attenuate hypertension-induced cardiac dysfunction through modulation of cellular stress responses. Life Sci. 2022, 296, 120424. [Google Scholar] [CrossRef]
- Kuno, A.; Hosoda, R.; Sebori, R.; Hayashi, T.; Sakuragi, H.; Tanabe, M.; Horio, Y. Resveratrol Ameliorates Mitophagy Disturbance and Improves Cardiac Pathophysiology of Dystrophin-deficient mdx Mice. Sci Rep. 2018, 8, 15555. [Google Scholar] [CrossRef]
- Wang, P.; Yang, Y.; Guo, J.; Ma, T.; Hu, Y.; Huang, L.; He, Y.; Xi, J. Resveratrol Inhibits Zinc Deficiency-Induced Mitophagy and Exerts Cardiac Cytoprotective Effects. Biol Trace Elem Res. 2024, 202, 1669–1682. [Google Scholar] [CrossRef]
- Ye, H.; Zhang, Y.; Yun, Q.; DU, R.; Li, L.; Li, Y.; Gao, Q. Resveratrol alleviates hyperglycemia-induced cardiomyocyte hypertrophy by maintaining mitochondrial homeostasis via enhancing SIRT1 expression]. Nan Fang Yi Ke Da Xue Xue Bao. 2024, 44, 45–51. [Google Scholar]
- Mori, A.; Ezawa, Y.; Asano, D.; Kanamori, T.; Morita, A.; Kashihara, T.; Sakamoto, K.; Nakahara, T. Resveratrol dilates arterioles and protects against N-methyl-d-aspartic acid-induced excitotoxicity in the rat retina. Neurosci Lett. 2023, 793, 136999. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Miura, K.; Han, R.; Seto-Tetsuo, F.; Arioka, M.; Igawa, K.; Tomooka, K.; Sasaguri, T. Differentiation-inducing factor 1 activates cofilin through pyridoxal phosphatase and AMP-activated protein kinase, resulting in mitochondrial fission. J Pharmacol Sci. 2023, 152, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.M.; Choi, S.C.; Song, M.H.; Kim, K.S.; Jun, S.; Park, J.H.; Kim, J.H.; Kim, K.; et al. The Activation of the LIMK/Cofilin Signaling Pathway via Extracellular Matrix-Integrin Interactions Is Critical for the Generation of Mature and Vascularized Cardiac Organoids. Cells. 2023, 12, 2029. [Google Scholar] [CrossRef] [PubMed]
- Huan,g F. ; Li, M.L.; Fang, Z.F.; Hu, X.Q.; Liu, Q.M.; Liu, Z.J.; Tang, L.; Zhao, Y.S.; Zhou, S.H. Overexpression of MicroRNA-1 improves the efficacy of mesenchymal stem cell transplantation after myocardial infarction. Cardiology. 2013, 125, 18–30. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the autophagy and/or mitophagy in cardiomyocytes involved in the cardiac disorders including cardiovascular disease, myocardial hypertrophy, cardiac muscular dystrophy, cardiomyopathy, and acute/chronic heart failure. Note that several important factors and/or severe inflammation with excessive ROS production triggering some critical pathways have been omitted for clarity.
Figure 1.
Schematic representation of the autophagy and/or mitophagy in cardiomyocytes involved in the cardiac disorders including cardiovascular disease, myocardial hypertrophy, cardiac muscular dystrophy, cardiomyopathy, and acute/chronic heart failure. Note that several important factors and/or severe inflammation with excessive ROS production triggering some critical pathways have been omitted for clarity.
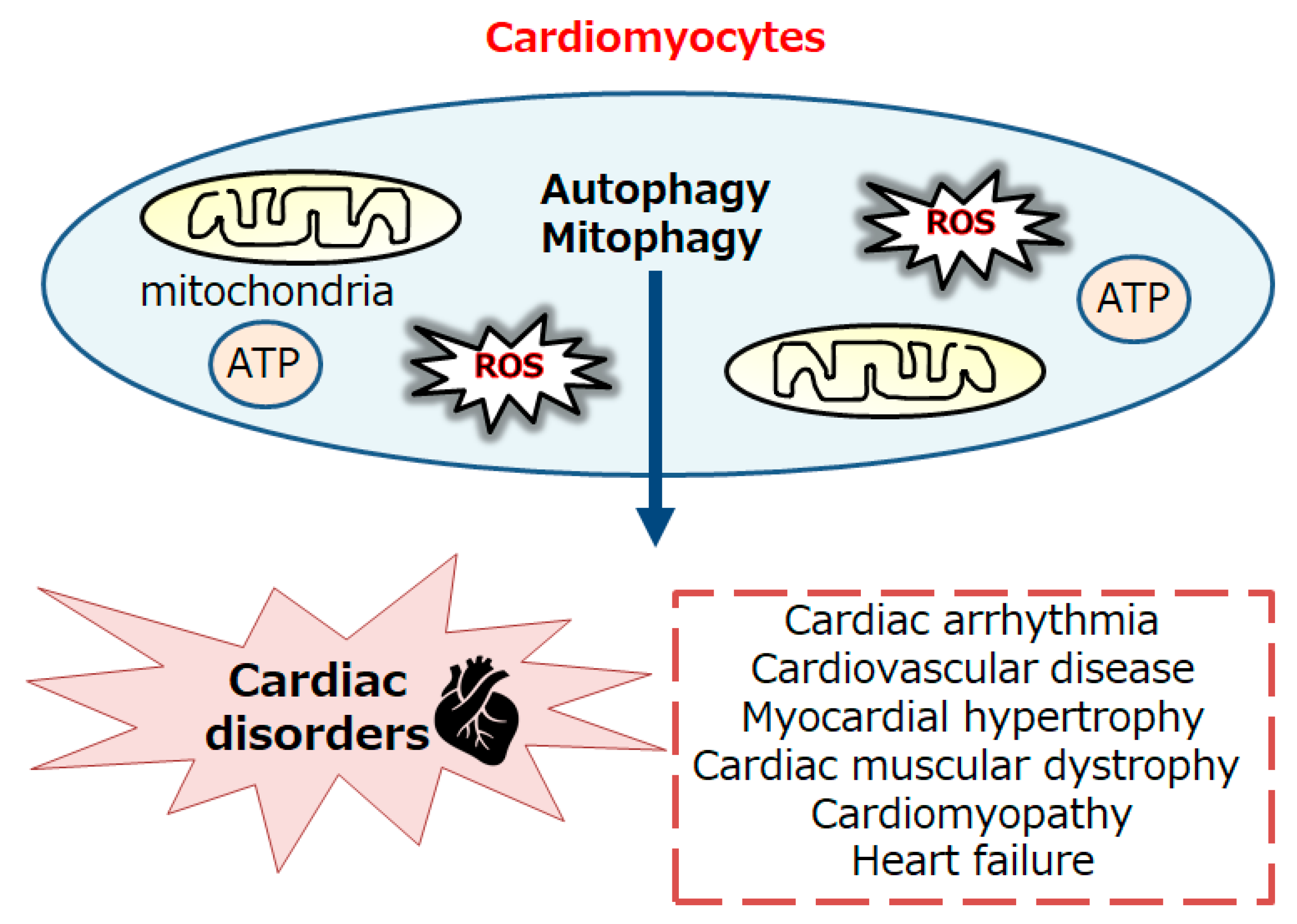
Figure 2.
An illustrative representation and overview of AMPK, mTOR, and related molecules in the regulatory pathway for the autophagy/mitophagy. Several modulator molecules linked to the AMPK/PI3K/AKT/mTOR/mTORC1 signaling are also demonstrated. Inflammation, starvation, and/or hypoxia known to act on the autophagy/mitophagy signaling are similarly shown. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several important activities such as cytokine-induction and/or intra-cellular inflammatory reactions have been omitted for clarity. Abbreviation: mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; ROS, reactive oxygen species.
Figure 2.
An illustrative representation and overview of AMPK, mTOR, and related molecules in the regulatory pathway for the autophagy/mitophagy. Several modulator molecules linked to the AMPK/PI3K/AKT/mTOR/mTORC1 signaling are also demonstrated. Inflammation, starvation, and/or hypoxia known to act on the autophagy/mitophagy signaling are similarly shown. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several important activities such as cytokine-induction and/or intra-cellular inflammatory reactions have been omitted for clarity. Abbreviation: mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; ROS, reactive oxygen species.
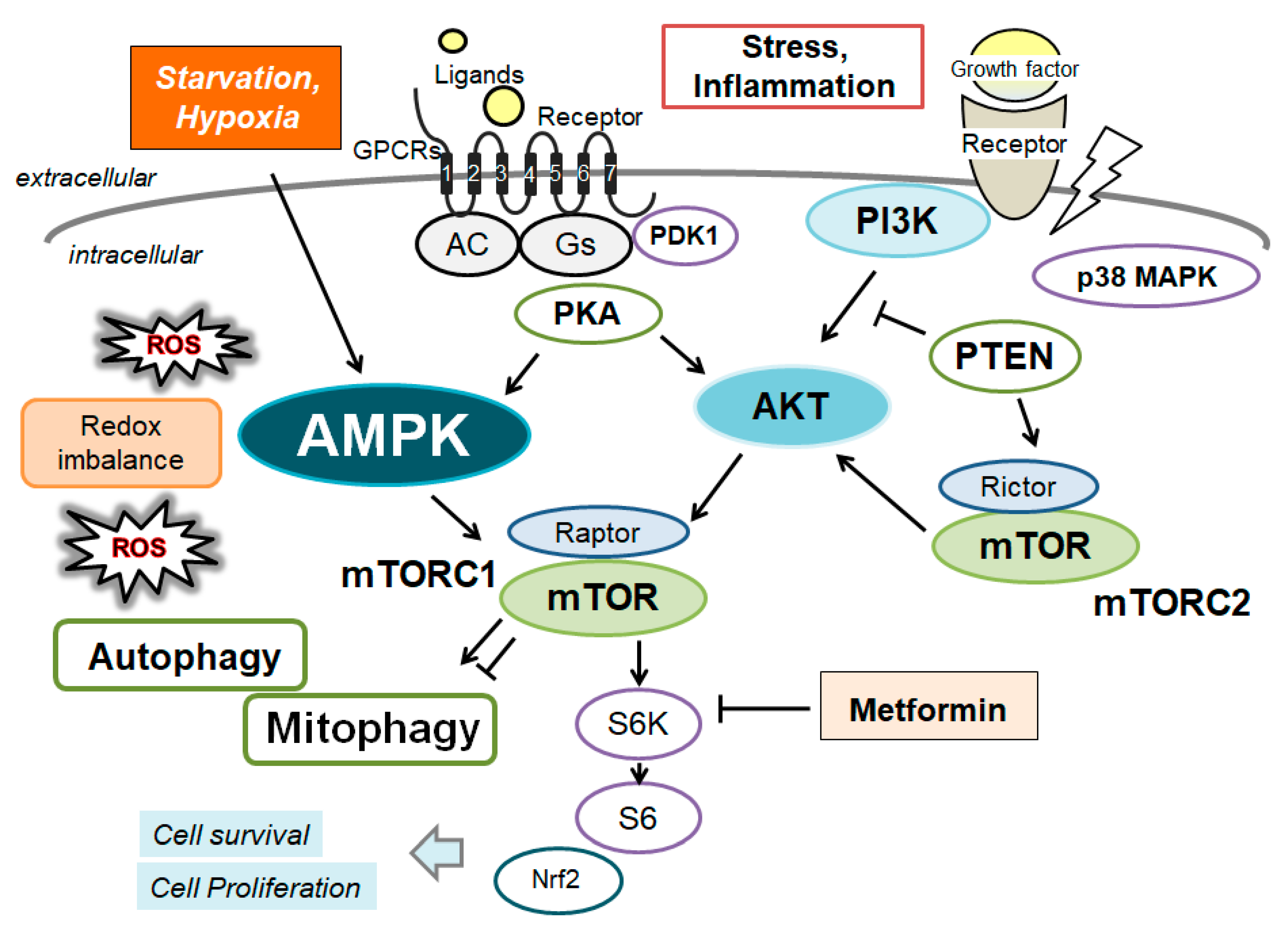
Figure 3.
Schematic demonstration of the potential inhibitory tactics against the pathogenesis of cardiac disorders. Example compounds including metformin as well as fasting, starvation, and/or exercise known to act on the mitophagy signaling are also shown. Note that some of important activities such as autophagy initiation, inflammatory reaction, and reactive oxygen species (ROS) production have been misplaced for clarity. In particular, some kinds of probiotics and/or fecal microbiota transplantation (FMT) might assist the alteration of gut microbiota for the modification of autophagy/mitophagy, which might be advantageous for the treatment of several cardiac disorders. “?” means for author speculation.
Figure 3.
Schematic demonstration of the potential inhibitory tactics against the pathogenesis of cardiac disorders. Example compounds including metformin as well as fasting, starvation, and/or exercise known to act on the mitophagy signaling are also shown. Note that some of important activities such as autophagy initiation, inflammatory reaction, and reactive oxygen species (ROS) production have been misplaced for clarity. In particular, some kinds of probiotics and/or fecal microbiota transplantation (FMT) might assist the alteration of gut microbiota for the modification of autophagy/mitophagy, which might be advantageous for the treatment of several cardiac disorders. “?” means for author speculation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



