
Submitted:
16 April 2024
Posted:
17 April 2024
You are already at the latest version
Abstract
Faba bean is an important legume crop wordwide, grown both for food and feed. As for many legume crops, postemergence weed control is difficult. The aim of this work was to assess potential of intercropping on weed management by assessing the effect of mixing faba bean with different crops (wheat, barley, and pea) on weeds in the South of Spain. Four field trials were conducted over four consecutive seasons, and results found that addition alternate intercropping of faba bean and barley achieved high levels of weed control, both for coverage and biomass (92.7% and 76.6%, respectively). Intercropping with wheat did not reduce weed coverage but did decrease weed biomass by 46.1% as compared to faba bean monocrop. Intercropping with pea did not provide significant reductions for weed coverage neither for weed biomass. The diversity of the weed community was not altered by the different crop combinations tested. Additional experiments under controlled conditions were carried out to determine a possible role of barley allelopathy in weed suppression. A strong allelopathic effect entailing a reduction in the number of plants and biomass was detected for four weeds: Sinapis arvensis, Polypogon monspeliensis, Matricaria camomilla, and Medicago trucatula. It can be concluded that intercropping faba bean with barley is an excellent strategy for weed control in the Southern Spain.
Keywords:
allelopathy
; faba bean
; intercropping
; weeds
1. Introduction
The Crop legumes are an essential component of sustainable agriculture, given the ecological services they provide, especially their ability to fix atmospheric nitrogen [1]. However, legume cultivation is hampered by biotic and abiotic stresses, which can seriously affect their yields [2]. One of these is the competition of weeds, which is a serious challenge for managing these crops. Not only may weeds reduce crop yields, but they also may affect the quality of the grain [3], serve as alternative hosts for diseases and pests [4,5], or even hinder harvesting. In general, weed control methods must be implemented for legume crops since these are inefficient competitors [6]. Herbicides are the most widely used weed management method, but these present several drawbacks, such as high costs for the farmer, environmental damage, and the eventual appearance of resistance to the active ingredients in weeds. These resistances may be especially problematic, as no herbicides with new modes of action have appeared in recent years [7]. Other strategies are mechanical control and management of the spatial arrangement of crops [8,9].
Faba bean (Vicia faba L.) stands out as one of the most cultivated legumes worldwide for its use for both animal feed and human consumption [10]. Weeds are one of the major biological constraints for faba bean, reducing yields by up to 50%, although they appear more competitive with weeds than other legumes [11].
Intercropping is the practice of growing two or more crop species simultaneously in one field. It may help in the control of weeds in many crops [12], complementing herbicides if necessary [13]. In the case of legumes, weed reduction has been reported when combined with diverse crops, such as corn, wheat, oat, barley or rape [14,15,16,17,18,19]. There are different types of intercropping, depending on the arrangement and sowing densities of the combined crops; mixed intercropping, when plants are completely mixed; and alternate intercropping, when rows of each crop alternate regularly [20]; in addition intercropping all crops are sown at their normal densities, while in replacement intercropping densities are adjusted for the presence of the other(s) crop(s) [21].
Different mechanisms explain the smothering of weeds in intercropping systems. The most common is the competition of crops and weeds for natural resources such as light, water or nitrogen [18]. Another one is allelopathy, which is a characteristic of plants that may be exploited for weed control [22]. Allelopathy is defined as the capability of plants to exert positive or negative influence in the surrounding area by releasing chemicals [23]. It has been known for a long time that several crops present allelopathic activity, such as oat, wheat, barley, and sorghum [24].
The main objective of this work was to establish if it is possible to control weeds in faba bean by intercropping, determining the best crop combination if so. To this effect, field trials were performed across different years. Additionally, experiments under controlled conditions were carried out to investigate the possible role of allelopathy in weed control in intercrops of faba bean.
2. Materials and Methods
2.1. Field trials
Five field trials were carried out from 2014 to 2018 in Córdoba (South of Spain) to assess the effect of different crop combinations on the control of weeds in faba bean (Table 1). The combinations tested were faba bean/wheat, faba bean/barley, and faba bean/pea as well as the monocrops of the four crops, and a monocrop of faba bean at 50% of sowing density (doubling the distance between rows) to check the effect of density on weed infestation. The cultivars used were 'Muchamiel' (faba bean), 'Califa' (wheat), 'Henley' (barley), and 'Messire' (pea). Sowing densities were 80 seeds/m2 for legumes and 200 seeds/m2 for cereals. Two different intercropping systems were evaluated: alternate with replacement at 50%, where rows of each crop are alternated, giving a final sowing rate of 50/50, i.e., half of the normal density of each crop; and alternate with addition, where rows are also alternated, but to a final rate of 100/100, i.e., the normal density of each crop. Replacement intercropping was evaluated in trials Córdoba-15-r and Córdoba-16-r, and addition intercropping was evaluated in trials Córdoba-16-a, Córdoba-17-a, and Córdoba-18-a. The experimental plots had a length of 3 m and comprised eight rows at 35 cm distance between them in the case of monocrops and replacement intercropping and 16 rows at 17.5 cm in the case of addition intercropping; the monocrop of faba bean at 50% had four rows at 70 cm per plot (Figure 1). For each trial the experiment was designed as a randomized complete block with four replications.
Two areas of 0.7 m2 were randomly evaluated in the central part of each plot at the faba bean maturity stage to assess weed infestation. The weeds were counted and classified by botanic families. Two diversification indices were calculated in trials Córdoba-16-a, Córdoba-17-a, and Córdoba-18-a:
Richness (R) and Relative density (D):
R = nFi/nFT
R: Richness; nFi: number of weed families present in a given plot (i); nFT: total number of weed families found in the whole trial
Dji = (Nji/NTi)×100
Dij: Relative density of weed family j in plot i; Nji: number of plants of weed family j present in plot i; NTi: total number of weed plants in plot i.
Additionally, two evaluators visually estimated the weed coverage as a percentage of the area covered by weeds. At the end of the experiment, in trials Córdoba-16-a and Córdoba-17-a, the weeds in a central area of 2 m2 in each plot were harvested, and their biomass determined by drying them in an oven at 60 ºC for three days and subsequently weighing them.
2.2. Controlled-Condition Experiments
Two experiments were carried out under controlled conditions to investigate a possible allelopathic effect of barley. Thus, four weeds that had grown in the field trials were selected for this: Polypogon monspeliensis (L.) Desf., Matricaria camomilla L., Sinapis arvensis L., and Medicago truncatula Gaertm, belonging to the botanical families Poaceae, Asteraceae, Cruciferae and Fabaceae, respectively. In the first one, 10 seeds of barley were sown on plastic pots (5× 5 × 10 cm) filled with a mixture of sand and peat at 1:3 rate (v:v) and grown in a growth chamber with a photoperiod of 12 h of visible light (150 μmol m−2 s−1 photon flux density) at 25 ºC, and 12 h of darkness at 20 ºC. Two weeks later, the barley plants were removed, and 20 seeds of the same weed species were sown in the same pots maintaining the substrate. Weed plants were counted every two weeks for 42 days. Then, weed plants were removed, dried in an oven at 60ºC for three days, and weighed to obtain their biomass. A treatment control was used, in which weeds were sown in pots where barley has not been previously grown. All this was done in separate pots for each one of the four weeds. The second experiment was carried out in the same way as in the first one, but barley plants remained in the pots when weeds were sown, and they grew together till the end of the experiment. The design in the two cases was randomized with seven replications. Each of these experiments was performed twice.
2.3. Statistical Analysis
Considering the type of data, generalized linear models (GLMs) were performed to study the influence of the treatment on weed coverage and biomass in field trials, following the next model:
Y = βo+β1X1+β2X2+β3X1×X2+Ɛ
Where Y is the dependent variable, βo, β1 and β3 denote the estimated parameters (weights), and Ɛ is the residual. Model fits were evaluated through the residual plots. Subsequently, we assessed the contribution of the treatment, year, and treatment×year according to Cohen’s f factor [25].
Given that the interactions treatment×year were significant (Cohen’s f > 0.4) for both coverage and biomass, and since the study's main objective was to compare the intercropping treatments independently of the year×crop interaction, a Fridman nonparametric two-way analysis of variance were used. Subsequently, Dunn’s test were performed to compare the treatment mean ranks at P ≤ 0.05. Additional ANOVA analyses were carried out to study the effect of the treatment on the weed richness and weed relative density in each experiment.
In the experiments under controlled conditions, the Poisson regression procedure was applied using the maximum likelihood estimation method to study how the plant counts depended on the presence or absence of barley plants. Thus, data were fitted to the following equation:
Y = Exp(βo+β1X1+β2X2)
Where Y is the number of plants, βo is the independent term, β1 and β2 are the estimated associated with the treatment (presence or not of barley). Difference between treatments was evaluated according to the p-value of the Poisson regression. Additionally, a t-test was performed to calculate the barley presence's effect on the weed species' biomass and compare the proportion of reduction of weed plants and weed biomass between the two types of experiments. Finally, the proportion of weed plants and biomass reduction between weed species was assessed by ANOVA analysis.
Data were analyzed using Statistix software (Version 10; Statistix, Tallahassee, FL) and the R statistical software package.
3. Results
3.1. Field trials
A high pressure of weed infestation was present in all field trials.
3.1.1. Field Trials – Replacement System
In the experiments where the replacement intercropping system was used there was no difference in weed coverage between any intercropping treatment and faba bean monocrop (Supplementary Data Table S1). In these trials, faba bean at 50% density presented the highest level of weed infestation (not statistically different from wheat and pea monocrops in the case of Córdoba-16-r). That is, intercropping was ineffective in controlling weeds in faba bean when rows of faba bean plants were replaced by rows of other crops (pea, wheat, or barley).
3.1.2. Field Trials – Addition System
Clear differences were found among treatments according to weed infestation values for the addition system. In general, pea and faba bean at 50% density presented the highest levels of weed infestation in all trials, both for weed coverage and weed biomass. The combined analysis of the experiments found the lowest weed coverage for the combination of faba bean with barley, with a reduction of 92.7% about the faba bean monocrop (Figure 2). For weed biomass, the combinations of faba bean with barley and faba bean with wheat showed the lowest values, not significantly different from those of barley as a sole crop, getting decreases in comparison with monocropped faba bean of 76.6% and 46.1%, respectively (Figure 3).
The weeds found were classified into 10 different botanical families along the trials Córdoba-16, Córdoba-17, and Córdoba-18 (Table 2). No difference was found for Richness (R) between faba bean monocrop and any of the intercropping mixtures in any of the three trials where it was calculated (Table 3). Equally, no significant differences for Relative density (D) of any of the 10 botanical families of weeds were detected for the different treatments tested (p > 0.05, ANOVA test; Supplementary Data Table S2).
Controlled-Condition Experiments
The number of emerged weed plants was significantly lower (p < 0.05) in the pots where barley had grown previously (either removed or maintained when weed seeds were sown) than in the pots used as control. The number of plants did not vary as the experiments progressed, so the counts of the three-time points were very similar (data not shown). The results for the final count of plants are shown in Table 4, both for the experiment where barley was removed before sowing the seeds of weeds and for the experiment where barley remained in the pots after the seeds of weeds were sown. The biomass of weeds at the end of the experiments was significantly lower in the pots with barley than in the control ones (Table 5).
The proportion of reduction of the number of weed plants and weed biomass per pot compared to the control (considered as the average of all the control pots for that weed) was calculated in both experiments (removing barley before sowing the weeds and not removing it). There were no significant differences in the proportion of the number of plants for the two types of experiments. On the contrary, there were differences in weed biomass per pot for Matricaria chamomilla and Sinapis arvensis: final weed biomass was lower when barley remained in the pots till the end of the experiment (Table 6).
Additionally, the proportion of reduction of the number of weed plants and weed biomass per pot as compared to the control was compared between the four weed species in the experiment where barley was removed, to investigate if any one of them was more affected by barley than the others and no significant differences were found between them (ANOVA, p > 0.05).
4. Discussion
In this work, we have evaluated intercropping as a tool to control weeds in faba bean in the South of Spain. The level of control attained was very high, reaching reductions of 92.7% in weed coverage when combined with barley and 76.6% and 46.1% in weed biomass when mixed with barley and wheat, respectively. The mixture of faba bean and barley, then, might be effective enough to dispense with the application of herbicides in this agroecosystem.
Previously only one work had studied the effect of the combination of faba bean and barley on weeds: Dhima et al. [26] reported that alternate replacement intercropping achieved levels of reduction of corn poppy (Papaver rhoeas L.) of around 90%. As for other cereals, the faba bean and wheat mixture has proven to reduce weed biomass by 60% in alternate and mixed replacement intercropping [27]. Equally, Boutagayout et al. [28] found that an alternate intercrop of faba bean with wheat and oat decreased the weed biomass by around 50%. As far as we know, our work is the first to achieve such a level of weed supression in faba bean, validated across three field experiments. Equally, barley has proved to be successful in reducing broomrape (Orobanche crenata Forsk.) infestation in faba bean [29], and also to be effective in controlling rust disease in faba bean [30]. In these cases, the intercropping systems were different from those assessed in this work, so it would be necessary to integrate them in the best way to maximize the benefits of faba bean / barley intercropping. Furthermore, it is remarkable that the combination of faba bean and wheat reduced 64% weed biomass compared to the wheat monocrop, making this mixture an exciting option for farmers that can be combined with some herbicide applications. Pea, however, has proved to be a poor competitor against weeds, both as monocrop and in mixture with faba bean, which is in line with the fact that the combination of grain legumes alone is not a good strategy to control weeds. Actually, these combinations are not found in the literature dealing with weed supression by intercropping [31].
We did not find differences in weed diversity among all the tested treatments: no weeds belonging to a particular botanic family were more affected by any crop combination or monocrops than others. This is in contrast with what has been found in other intercropping systems [32,33], although there are also situations where weed diversity has not been influenced by intercropping [33]. In our case, the weed community has proved to be relatively stable regardless of the crop or crops present.
Of the two types of intercropping we tested, alternate with replacement and alternate with addition, only the latter was adequate for controlling weeds. The relationship between crops and weeds is based on the competition for available resources such as water, nutrients or light [34,35]. Plant density is one key factor in the improvement of crop competitiveness against weeds [36], and that is precisely the difference between both intercropping systems. In addition, intercropping plant density is doubled in comparison with alternate intercropping. The high weed pressure levels that we observed in the plots with faba bean sown at half density confirm the importance of plant density.
Plant density, however, is not the only mechanism that explains weed suppression in intercropping. If that were the case, we would have observed similar results with barley, wheat, or pea. Weed pressure levels for them as sole crops illustrate that not all crops have the same competitive ability against weeds: barley presents very low weed infestation compared to the other two, with pea ranking the highest. Other factors influence the performance in the presence of weeds, such as plant architecture, vigor, or allelopathy [37,38].
Barley has been described as one of the most competitive crops against weeds by different authors [39,40]. The rapid biomass accumulation and high growth rates that barley shows at the beginning of its cycle are some of the reasons for this [40]. Another reason is the efficiency of barley in taking nitrogen: it has been reported to be more competitive for nitrogen than pea in intercrops, so depriving weeds of this nutrient. Beyond that, barley is considered a crop with high levels of allelopathy [41], and as many as 44 potential allelochemicals have been identified so far [42]. The two more important are the alkaloids Gramine and Hordeine, which appear in barley plants' leaves, roots, and roots exudates [23,42]. All this has made barley a common partner in crop diversification for weed control [31].
The experiments under controlled conditions aimed at evaluating the role that barley allelopathy might have on our results. Different types of bioassays under controlled conditions may be used to assess the allelopathic ability of a plant species, such as testing extracts from the allelopathic plant [43,44], agar bioassays [39] or pots screenings [45], where plants are grown together in Petri dishes or pots with soil, respectively. We opted for pot screening because it may better reflect the conditions under which allelopathy operates. Besides, our design, in which removing barley plants is compared with the effect of not removing them, allows for discriminating allelopathy from competition effects.
The results from the pots where barley was removed before sowing the weeds show a substantial allelopathic effect against them. All four weed species presented a decrease in plant emergence and biomass that can only be explained by the presence in the soil of chemical compounds previously released by the barley plants. These results also suggest that the main allelopathic effect is related to the first stages of seed germination and seedling development. Remarkably, there were no significant differences between the final number of weed plants in the pots where barley had been removed and in those where it remained till the end. As for weed biomass, however, in the case of two species (Sinapis arvensis and Matricaria camomilla) the decrease was higher when barley remained than when it was removed. This is probably due to additional allelopathic effects, although it is more difficult to separate them from competition effects in this case. Barley allelopathy had been previously tested on Sinapis arvensis [46,47]. Still, as far as we know, this is the first time it has been assessed on the other three weeds.
The fact that there has been no difference in the reduction of emerged plants and biomass between four weeds belonging to such different botanical families points to a global and non-discriminatory effect of barley in our case. These weeds are a sample of the ten families found in our area, and this global effect could explain the lack of differences in the composition of weed communities between the barley intercrops and the monocrops in our field experiments.
In conclusion, this research has established that the combination of faba bean and barley has great potential for the control of weeds in the agroclimatic region of the South of Spain. Further work should focus on developing the optimal intercropping strategy to obtain the best advantages from combining these two crops. It would also be of great interest to identify the allelochemicals behind weed suppression in our experiments and determine their mechanisms of action.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1 and table S2.
Author Contributions
Conceptualization, D.R., A.V. and J.M.; methodology, D.R., A.V., J.M..; formal analysis, J.M. and A.V..; investigation, A.A., A.V. and J.M..; writing—original draft preparation, A.V..; writing—review and editing, A.V., D.R. and J.M..; supervision, D.R..; funding acquisition, D.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Spanish Ministerio de Ciencia e Innovación project PID2020-114668RB-I00, by the Horizon 2020 EU project H2020-SFS-01-2020 GA No 101000622, RADIANT and by Junta de Andalucía PAIDI 2020 project P20_00986.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ditzler, L.; van Apeldoorn, D.F.; Pellegrini, F.; Antichi, D.; Bàrberi, P.; Rossing, W.A.H. Current research on the ecosystem service potential of legume inclusive cropping systems in Europe. A review. Agronomy for Sustainable Development 2021, 41, 26. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.D.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and Challenges in Legume Breeding for Pest and Disease Resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef]
- Plaza, M.E.H.; Bastida, F.; Gibson, D.J.; Barro, F.; Giménez, M.J.; Pallavicini, Y.; Izquierdo, J.; González-Andújar, J.L. Grain Quality as Influenced by the Structural Properties of Weed Communities in Mediterranean Wheat Crops. 2022. [Google Scholar] [CrossRef]
- Kumar, S.; Bhowmick, M.; Ray, P. Weeds as alternate and alternative hosts of crop pests. INDIAN JOURNAL OF WEED SCIENCE 2021, 53, 14–29. [Google Scholar] [CrossRef]
- Dentika, P.; Ozier-Lafontaine, H.; Penet, L. Weeds as Pathogen Hosts and Disease Risk for Crops in the Wake of a Reduced Use of Herbicides: Evidence from Yam (Dioscorea alata) Fields and Colletotrichum Pathogens in the Tropics. Journal of fungi (Basel, Switzerland) 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Carton, N.; Naudin, C.; Piva, G.; Baccar, R.; Corre-Hellou, G. Differences for traits associated with early N acquisition in a grain legume and early complementarity in grain legume-triticale mixtures. Aob Plants 2018, 10. [Google Scholar] [CrossRef]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Management Science 2012, 68, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Avola, G.; Tuttobene, R.; Gresta, F.; Abbate, V. Weed control strategies for grain legumes. Agronomy for Sustainable Development 2008, 28, 389–395. [Google Scholar] [CrossRef]
- Pannacci, E.; Tei, F.; Guiducci, M. Evaluation of mechanical weed control in legume crops. Crop Protection 2018, 104, 52–59. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. 2020.
- Karkanis, A.; Ntatsi, G.; Lepse, L.; Fernandez, J.A.; Vagen, I.M.; Rewald, B.; Alsina, I.; Kronberga, A.; Balliu, A.; Olle, M.; et al. Faba Bean Cultivation - Revealing Novel Managing Practices for More Sustainable and Competitive European Cropping Systems. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef]
- Huss, C.P.; Holmes, K.D.; Blubaugh, C.K. Benefits and Risks of Intercropping for Crop Resilience and Pest Management. Journal of Economic Entomology 2022, 115, 1350–1362. [Google Scholar] [CrossRef]
- Weerarathne, L.V.Y.; Marambe, B.; Chauhan, B.S. Does intercropping play a role in alleviating weeds in cassava as a non-chemical tool of weed management? - A review. Crop Protection 2017, 95, 81–88. [Google Scholar] [CrossRef]
- Flores-Sanchez, D.; Pastor, A.; Lantinga, E.A.; Rossing, W.A.H.; Kropff, M.J. Exploring Maize-Legume Intercropping Systems in Southwest Mexico. Agroecology and Sustainable Food Systems 2013, 37, 739–761. [Google Scholar] [CrossRef]
- Banik, P.; Midya, A.; Sarkar, B.K.; Ghose, S.S. Wheat and chickpea intercropping systems in an additive series experiment: Advantages and weed smothering. European Journal of Agronomy 2006, 24, 325–332. [Google Scholar] [CrossRef]
- Begna, S.H.; Fielding, D.J.; Tsegaye, T.; Van Veldhuizen, R.; Angadi, S.; Smith, D.L. Intercropping of oat and field pea in Alaska: An alternative approach to quality forage production and weed control. Acta Agriculturae Scandinavica Section B-Soil and Plant Science 2011, 61, 235–244. [Google Scholar] [CrossRef]
- Corre-Hellou, G.; Dibet, A.; Hauggaard-Nielsen, H.; Crozat, Y.; Gooding, M.; Ambus, P.; Dahlmann, C.; von Fragstein, P.; Pristeri, A.; Monti, M.; et al. The competitive ability of pea-barley intercrops against weeds and the interactions with crop productivity and soil N availability. Field Crops Research 2011, 122, 264–272. [Google Scholar] [CrossRef]
- Sharma, G.; Shrestha, S.; Kunwar, S.; Tseng, T. Crop Diversification for Improved Weed Management: A Review. Agriculture-Basel 2021, 11. [Google Scholar] [CrossRef]
- Kemper, R.; Rinke, N.; Gerhards, R.; Böhm, H. Weed suppression and crop yield performance in sole and intercrops of common vetch and spring wheat depending on seed density ratio in organic farming. Journal fur Kulturpflanzen 2020, 72, 12–24. [Google Scholar] [CrossRef]
- Lithourgidis, A.S.; Dordas, C.A.; Damalas, C.A.; Vlachostergios, D.N. Annual intercrops: an alternative pathway for sustainable agriculture. Australian Journal of Crop Science 2011, 5, 396–410. [Google Scholar]
- Boudreau, M.A. Diseases in Intercropping Systems. In Annual Review of Phytopathology, Vol 51, VanAlfen, N.K., Ed.; Annual Review of Phytopathology; 2013; Volume 51, pp. 499–519.
- Khamare, Y.; Chen, J.J.; Marble, S.C. Allelopathy and its application as a weed management tool: A review. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- Maver, M.; Miras-Moreno, B.; Lucini, L.; Trevisan, M.; Pii, Y.; Cesco, S.; Mimmo, T. New insights in the allelopathic traits of different barley genotypes: Middle Eastern and Tibetan wild-relative accessions vs. cultivated modern barley. PloS one 2020, 15, e0231976. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G. Allelopathy in crop/weed interactions - an update. Pest Management Science 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates, Inc.: New Jersey, 1988. [Google Scholar]
- Dhima, K.; Vasilakoglou, I.; Gatsis, T.; Gougoulias, N. Faba bean-barley intercrops for high productivity and corn poppy suppression. Experimental Agriculture 2016, 54, 163–180. [Google Scholar] [CrossRef]
- Eskandari, H. Intercropping of wheat (Triticum aestivum) and bean (Vicia faba): Effects of complementarity and competition of intercrop components in resource consumption on dry matter production and weed growth. African Journal of Biotechnology 2011, 10, 17755–17762. [Google Scholar] [CrossRef]
- Boutagayout, A.; Belmalha, S.; Nassiri, L.; el Alami, N.; Jiang, Y.; Lahlali, R.; Bouiamrine, E.H. Weed competition, land equivalent ratio and yield potential of faba bean (Vicia faba L.)-cereals (Triticum aestivum L. and/or Avena sativa L.) intercropping under low-input conditions in Meknes region, Morocco. Vegetos. [CrossRef]
- Fernandez-Aparicio, M.; Sillero, J.C.; Rubiales, D. Intercropping with cereals reduces infection by Orobanche crenata in legumes. Crop Protection 2007, 26, 1166–1172. [Google Scholar] [CrossRef]
- Villegas-Fernandez, A.M.; Amarna, A.A.; Moral, J.; Rubiales, D. Crop Diversification to Control Rust in Faba Bean Caused by Uromyces viciae-fabae. Journal of Fungi 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Bastiaans, L.; Anten, N.P.R.; Makowski, D.; van der Werf, W. Annual intercropping suppresses weeds: A meta-analysis. Agriculture Ecosystems & Environment 2021, 322. [Google Scholar] [CrossRef]
- Poggio, S.L.; Satorre, E.H.; de la Fuente, E.B. Structure of weed communities occurring in pea and wheat crops in the Rolling Pampa (Argentina). Agriculture, Ecosystems & Environment 2004, 103, 225–235. [Google Scholar] [CrossRef]
- Stefan, L.; Engbersen, N.; Schöb, C. Crop–weed relationships are context-dependent and cannot fully explain the positive effects of intercropping on yield. Ecological Applications 2021, 31. [Google Scholar] [CrossRef]
- Swanton, C.; Ondoua, R.; Blackshaw, R.E. Experimental methods for crop-weed competition studies. Weed Science 2015, 63, 2–11. [Google Scholar] [CrossRef]
- Singh, M.; Kukal, M.S.; Irmak, S.; Jhala, A.J. Water Use Characteristics of Weeds: A Global Review, Best Practices, and Future Directions. Frontiers in Plant Science 2022, 12. [Google Scholar] [CrossRef]
- Lemerle, D.; Cousens, R.; Gill, G.; Peltzer, S.; Moerkerk, M.; Murphy, C.; Collins, D.; Cullis, B. Reliability of higher seeding rates of wheat for increased competitiveness with weeds in low rainfall environments. The Journal of Agricultural Science 2004, 142. [Google Scholar] [CrossRef]
- Korav, S.; Dhaka, A.; Singh, R.; Reddy, G. A study on crop weed competition in field crops. 2018, 7, 3235–3240.
- Jha, P.; Kumar, V.; Godara, R.; Chauhan, B. Weed management using crop competition in the United States: A review. Crop Protection 2017, 95, 31–37. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Early vigour and allelopathy - Two useful traits for enhanced barley and wheat competitiveness against weeds. Weed Research 2005, 45, 94–102. [Google Scholar] [CrossRef]
- Dhima, K.; Vasilakoglou, I.; Gatsis, T.; Eleftherohorinos, I. Competitive interactions of fifty barley cultivars with Avena sterilis and Asperugo procumbens. Field Crops Research 2010, 117, 90–100. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Protection 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Kremer, R.; Ben-Hammouda, M. Allelopathic Plants. 19. Barley (Hordeum vulgare L). Allelopathy Journal 2009, 24, 225–242. [Google Scholar]
- Asghari, J.; Tewari, J.P. Allelopathic Potentials of Eight Barley Cultivars on Brassica jucea (L) Czern. and Setaria viridis (L) p. Beauv. Journal of Agricultural Science and Technology 2007, 9, 165–176. [Google Scholar]
- Copaja, S.; López, M.; Bravo, H. Dynamic of indole alkaloids in a soil and its relationships with allelopathic properties. Journal of the Chilean Chemical Society 2023, 68, 5787–5795. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T.; An, M. Screening methods for the evaluation of crop allelopathic potential. Botanical Review 2001, 67, 403–415. [Google Scholar] [CrossRef]
- Vasilakoglou, I.; Dhima, K.; Lithourgidis, A.; Eleftherohorinos, I. Allelopathic potential of 50 barley cultivars and the herbicidal effects of barley extract. Allelopathy Journal 2009, 24, 309–319. [Google Scholar]
- Modhej, A.; Rafatjoo, A.; Behdarvandi, B. Allelopathic inhibitory potential of some crop species (wheat, barley, canola, and safflower) and wild mustard (Sinapis arvensis). International Journal of Biosciences (IJB) 2013, 3, 212–220. [Google Scholar]
Figure 1.
Different cropping systems evaluated in the field trials. A: Faba bean monocrop. B: Alternate replacement intercropping. C: Alternate addition intercropping. D: Faba bean monocrop at 50% of sowing density. Row distances in A and B were 35 cm., in C were 17.5 cm. and in D 70 cm.
Figure 1.
Different cropping systems evaluated in the field trials. A: Faba bean monocrop. B: Alternate replacement intercropping. C: Alternate addition intercropping. D: Faba bean monocrop at 50% of sowing density. Row distances in A and B were 35 cm., in C were 17.5 cm. and in D 70 cm.

Figure 2.
Weed coverage in the different treatments evaluated across the three intercropping field trials where addition alternate intercropping was tested. Different letters mean significant differences according to Dunn’s test at p ≤ 0.05.
Figure 2.
Weed coverage in the different treatments evaluated across the three intercropping field trials where addition alternate intercropping was tested. Different letters mean significant differences according to Dunn’s test at p ≤ 0.05.
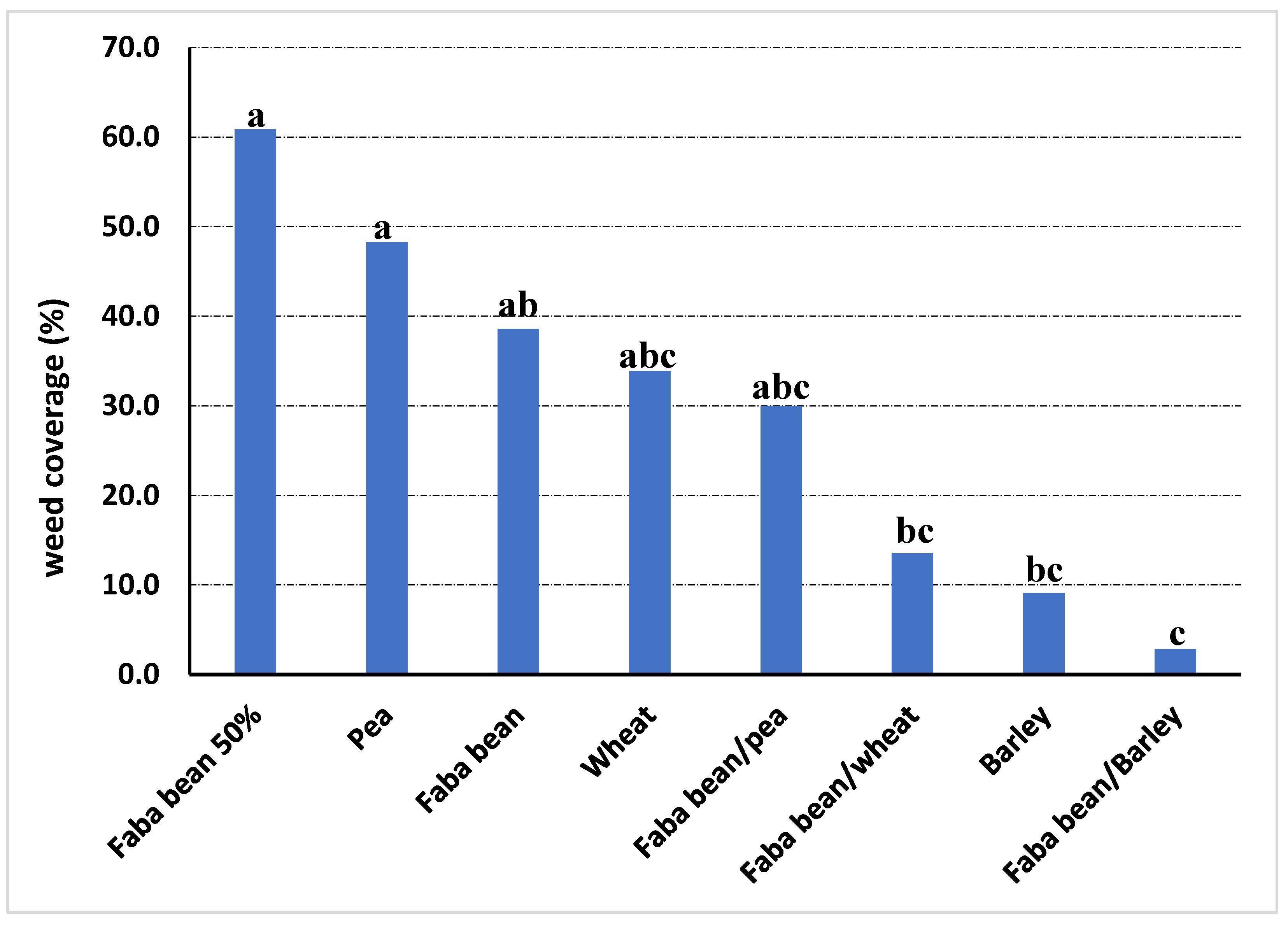
Figure 3.
Weed biomass (g/m2) in the different treatments evaluated across the two intercropping field trials where addition alternate intercropping was tested. Different letters mean significant differences according to Least Significant Difference (LSD) test at p ≤ 0.05.
Figure 3.
Weed biomass (g/m2) in the different treatments evaluated across the two intercropping field trials where addition alternate intercropping was tested. Different letters mean significant differences according to Least Significant Difference (LSD) test at p ≤ 0.05.
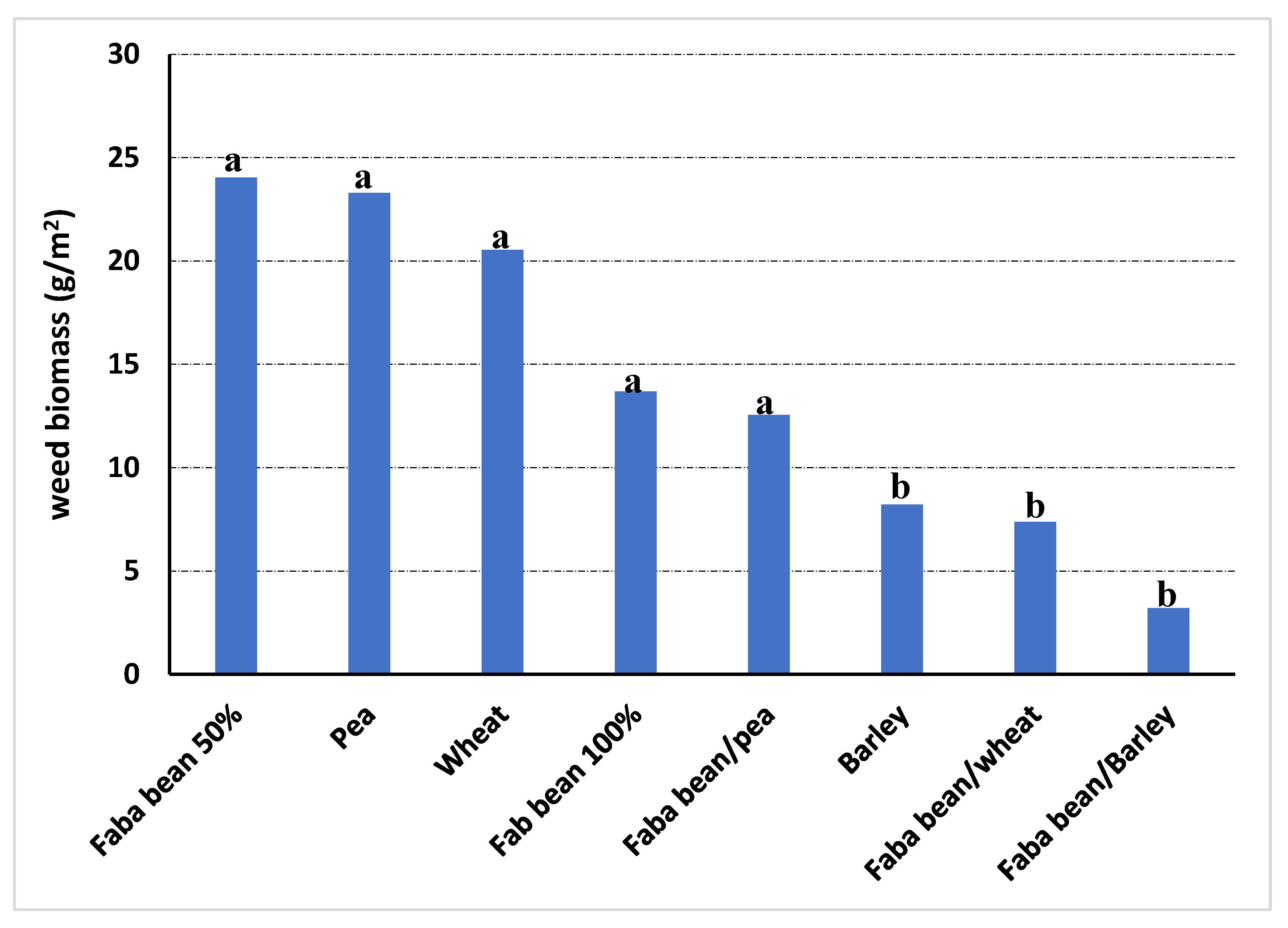

Table 1.
Field trials carried out for the study of control of weeds in faba bean by intercropping.
| Trial | Córdoba-15-r | Córdoba-16-r | Córdoba-16-a | Córdoba-17-a | Córdoba-18-a |
|---|---|---|---|---|---|
| Season | 2014/15 | 2015/16 | 2015/16 | 2016/17 | 2017/18 |
| Intercropping system | Replacement | Replacement | Addition | Addition | Addition |
| Maximum T (ºC) | 35.3 | 31.7 | 31.7 | 32.2 | 29.7 |
| Minimum T (ºC) | −3.3 | -2.3 | -2.3 | −3.4 | -3.4 |
| Mean T (ºC) | 12.3 | 12.6 | 12.6 | 14.3 | 12.0 |
| Precipitation (ml) | 150 | 336 | 336 | 143.8 | 444.6 |
Table 2.
Presence of the different botanical families to which the weeds identified belonged in the field trials Córdoba-16, Córdoba-17 and Córdoba-18.
Table 2.
Presence of the different botanical families to which the weeds identified belonged in the field trials Córdoba-16, Córdoba-17 and Córdoba-18.
| Córdoba-16 | Córdoba-17 | Córdoba-18 | |
|---|---|---|---|
| Amarantaceae | x | x | |
| Asteraceae | x | ||
| Convolvulaceae | x | x | x |
| Cruciferae | x | x | |
| Cyperaceae | x | ||
| Fabaceae | x | x | x |
| Papaveraceae | x | x | |
| Poaceae | x | x | |
| Polygonaceae | x | x | x |
| Quenopodiaceae | x |
Table 3.
Weed richness (R) for the different treatments tested in three field trials (Córdoba-16, Córdoba-17, and Córdoba-18). Different letters mean significant differences for each trial (Tukey test, p <0.05). Richness was not measured for barley, wheat and pea monocrops in Cordoba-16.
Table 3.
Weed richness (R) for the different treatments tested in three field trials (Córdoba-16, Córdoba-17, and Córdoba-18). Different letters mean significant differences for each trial (Tukey test, p <0.05). Richness was not measured for barley, wheat and pea monocrops in Cordoba-16.
| Richness (R) | |||
|---|---|---|---|
| Córdoba-16 | Córdoba-17 | Córdoba-18 | |
| Faba bean 100% | 35.7 a | 78.6 ab | 76.2 ab |
| Faba bean / barley | 25.0 a | 50.0 ab | 42.3 a |
| Faba bean /wheat | 25.0 a | 46.4 a | 61.9 ab |
| Faba bean / pea | 50.0 a | 64.3 ab | 85.7 b |
| Faba bean 50% | 46.4 a | 75.0 ab | 85.7 b |
| Barley 100% | 83.3 b | 76.2 ab | |
| Wheat 100% | 75.0 ab | 95.0 b | |
| Pea 100% | 83.3 b | 80.9 ab | |
Table 4.
Number of weed plants per pot for each treatment in the two experiments under controlled conditions: where barley was removed before sowing the weeds and where barley was not removed after sowing the weeds. Different letters mean significant differences for each weed in each type of experiment, i.e., with barley removed or not removed. Differences were significant in all cases according to Poisson’s p-value at p ≤ 0.05.
Table 4.
Number of weed plants per pot for each treatment in the two experiments under controlled conditions: where barley was removed before sowing the weeds and where barley was not removed after sowing the weeds. Different letters mean significant differences for each weed in each type of experiment, i.e., with barley removed or not removed. Differences were significant in all cases according to Poisson’s p-value at p ≤ 0.05.
| Barley removed | Barley not removed | |||
|---|---|---|---|---|
| Control | Barley sown | Control | Barley sown | |
| Polypogon monspeliensis | 19.9 a | 5.0 b | 19.7 a | 4.6 b |
| Matricaria chamomilla | 18.9 a | 5.9 b | 18.6 a | 5.1 b |
| Sinapis arvensis | 11.1 a | 3.8 b | 11.9 a | 2.7 b |
| Medicago truncatula | 5.2 a | 1.4 b | 7.8 a | 1.4 b |
Table 5.
Biomass of weed plants per pot (mg) for each treatment in the two experiments under controlled conditions: where barley was removed before sowing the weeds and where barley was not removed after sowing the weeds. Different letters mean significant differences for each weed in each type of experiment, i.e., with barley removed or not removed, according to the t-student test at p ≤ 0.05.
Table 5.
Biomass of weed plants per pot (mg) for each treatment in the two experiments under controlled conditions: where barley was removed before sowing the weeds and where barley was not removed after sowing the weeds. Different letters mean significant differences for each weed in each type of experiment, i.e., with barley removed or not removed, according to the t-student test at p ≤ 0.05.
| Barley removed | Barley not removed | |||
|---|---|---|---|---|
| Control | Barley sown | Control | Barley sown | |
| Polypogon monspeliensis | 39.9 a | 20.2 b | 55.5 a | 24.9 b |
| Matricaria chamomilla | 115.4 a | 49.0 b | 131.6 a | 25.6 b |
| Sinapis arvensis | 70.6 a | 41.7 b | 104.6 a | 10.1 b |
| Medicago truncatula | 16.3 a | 10.6 b | 35.0 a | 4.8 b |
Table 6.
Proportion of number of weed plants and biomass per pot as compared to the control for both types of experiments under controlled conditions (removing barley and not removing it just prior to sowing the weeds). Different letters mean significant differences for each weed and parameter: number and biomass of weeds (according to the t-student test at p ≤ 0.05).
Table 6.
Proportion of number of weed plants and biomass per pot as compared to the control for both types of experiments under controlled conditions (removing barley and not removing it just prior to sowing the weeds). Different letters mean significant differences for each weed and parameter: number and biomass of weeds (according to the t-student test at p ≤ 0.05).
| No of weeds (%) | Biomass of weeds (%) | |||
|---|---|---|---|---|
| Barley removed | Barley not removed | Barley removed | Barley not removed | |
| Polypogon monspeliensis | 25.2 a | 23.6 a | 70.1 a | 61.9 a |
| Matricaria chamomilla | 31.1 a | 27.3 a | 65.5 a | 43.7 b |
| Sinapis arvensis | 32.1 a | 23.0 a | 77.0 a | 29.3 b |
| Medicago truncatula | 24.9 a | 18.4 a | 58.9 a | 31.9 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



