
Submitted:
16 April 2024
Posted:
17 April 2024
You are already at the latest version
Abstract
Candidiasis is considered an emerging public health concern because of the occurrence of drug-resistant Candida strains and the lack of available structurally diverse antifungal drug armamentariums. The indole alkaloid globospiramine from the anticandidal Philippine medicinal plant Voacanga globosa exhibits a variety of biological activities; however, its antifungal properties remain to be explored. In this study, we report the in vitro antifungal activities of globospiramine against Candida albicans and Candida tropicalis and explore its possible target proteins using in silico methods. The colony-forming unit (CFU) viability assay revealed time- and concentration-dependent anticandidal effects of the alkaloid by decreasing almost 50% of the viable CFUs 60 min after treatment. Results of the MIC and MFC assays indicated inhibitory and fungicidal effects of globospiramine against C. albicans (MIC = 8 µg/mL; MFC = 4 µg/mL) and potential fungistatic effects against C. tropicalis at lower concentrations (MIC = 4 µg/mL; MFC > 64 µg/mL). The FAM-FLICA poly-caspase assay showed metacaspase activation in C. albicans cells at concentrations of 16 and 8 µg/mL, which agreed with the MIC and MFC values. Molecular docking and molecular dynamics simulation experiments indicated stable, strong binding of globospiramine with the target proteins 1,3-β-glucan synthase and Als3 adhesin enzymes, which are indirectly involved in apoptotic-driven candidal inhibition.
Keywords:
globospiramine
; bisindole alkaloid
; Voacanga globosa
; Candida albicans
; Candida tropicalis
; molecular docking
; molecular dynamics
; antifungal
; apoptosis
1. Introduction
The increased occurrence and severity of fungal infections have greatly contributed to the escalation of disease-associated morbidity and mortality rates , with approximately 1.5 million deaths annually [1,2,3]. Treatment failures are mostly attributed to the emergence and re-emergence of resistant strains, which is among the consequences of the irresponsible use of available antifungal drugs and innate evolutionary mechanisms of the causative agents against therapeutic pressures [4,5].
Fungal infections are caused by different species, mostly belonging to the genera Aspergillus, Pneumocystis, Candida, and Cryptococcus. Among these, Candida spp. are the most frequently reported pathogen [6]. Candida spp. transition into opportunistic pathogens in immunologically weak and immunocompromised patients, leading to local and systemic infections [7]. However, in the last few decades, there has been an increased incidence of deep fungal infections or chronic candidiasis (long-term infection caused by Candida species) even in healthy individuals [8,9]. In addition to C. albicans, C. tropicalis, C. glabrata, C. parapsilosis, and C. krusei infections have been reported in clinical settings. These Candida cells have developed mechanisms to ensure efficient colonization and infection of their hosts despite therapeutic pressures. Therefore, this necessitates the discovery of new antifungal agents with novel and/or multitargeting mechanisms [10,11].
Current clinically approved antifungals belong to the following structural classes: polyenes, azoles, and echinocandins [12]. Polyenes like amphotericin B target ergosterol, which is selectively found in fungal cell membranes. Azoles inhibit fungal cytochrome P450-dependent enzymes, resulting in impaired ergosterol synthesis. Echinocandins block the 1,3-β-D-glucan synthase, disrupting fungal cell wall biosynthesis [13]. The fact that most chemotherapeutic-based treatment armamentarium against candidiasis is limited to these three classes calls for new and diverse drug congeners from different structural classes.
Relevant to our study, indole alkaloids have long been explored for their biological properties. The indole moiety contributes to the efficient binding of compounds to a number of disease targets, thus conferring favorable biological activities. In the context of anticandidal drug discovery, some reported indole alkaloids include kopsifolines A, G, H, I, J, and K from Kopsia fruticosa [14], ibogaine [15], and chetomin and chaetoglobinol A from Chaetomium globosusm [16]. Moreover, six out of 27 isolated monoterpene indole alkaloids from the plant Rhazya strictra demonstrated inhibitory properties against six Candida strains [17]. Other anticandidal indole alkaloids derived from marine organisms include indolepyrazines A and B from Acinetobacter sp. ZZ1275 [18]. Thus, indole alkaloids and their derivatives have great potential for development as anticandidal agents. Among the prolific producers of biologically active indole alkaloids is the genus Voacanga of the family Apocynaceae. In the Philippines, the medicinal plant Voacanga globosa (Blanco) Merr. has numerous biological activities [19,20,21], including C. albicans inhibition [22]. As part of our research effort to explore antimicrobial agents from Philippine medicinal plants, we hereby disclose the antifungal activities of the spirobisindole alkaloid globospiramine (Figure 1) previously obtained from Voacanga globosa [19] against C. albicans and C. tropicalis using in vitro and computational approaches (molecular docking and molecular dynamics simulations).
2. Materials and Methods
2.1. Test Compound
Globospiramine was isolated from Voacanga globosa as previously reported [19]. The test concentrations of globospiramine were prepared by dilution with dimethyl sulfoxide (DMSO).
2.2. Colony-Forming Unit Viability Assay
A colony-forming unit (CFU) viability assay was performed following previously reported methods [23]. Briefly, C. albicans ATCC 90028 and C. tropicalis ATCC 750 cells were inoculated in Sabouraud dextrose broth (SDB) and incubated overnight. The resulting culture must reach OD600 = 0.3 to 0.8 prior to serial dilution until approximately 500 CFUs/mL were attained. The test compound globospiramine in varying concentrations (2.5, 50, and 100 µg/mL) was added to the diluted suspension and 50 µL were aliquoted from the diluted suspension and spread plated on Sabouraud dextrose agar (SDA) plates after 0, 30, 60, and 90 min. After 48 h of incubation, percentage CFU viability was measured using the following equation:
where is the number of surviving colonies at each specific time point (x = 30, 60, or 90 minutes) for each test concentration of globospiramine while is the number of colonies at the starting point (time = 0). DMSO (negative/vehicle) and amphotericin B B were utilized as controls. The experiment was performed in triplicate.
2.3. MIC and MFC Determination
The minimum inhibitory concentration (MIC) was assessed based on CLSI M27-A3 2008 guidelines on microdilution for yeast species. Fresh yeast suspension (OD600 = 0.3 to 0.8) in SDB was diluted to yield 5.0 x 102 to 2.5 x 103 cells/mL. Two-fold serial dilutions from 256 µg/mL to 1 µg/mL were prepared in a 96-well microplate for globospiramine. For the positive control amphotericin B, the concentrations used were 0.125 to 16 µg/mL. Each well contained 100 µL of test concentrations, 5 µL standardized yeast suspension in SDB, and 100 µL RPMI 1640 broth medium. Absorbance was recorded using the Glomax Discover Microplate Reader (Promega) after 24 h of incubation. MIC was recorded as the lowest concentration to inhibit increase in OD600. Triplicates were performed. For the minimum fungicidal concentration (MFC) determination assay, 10 µL from the well containing the MIC and two concentrations higher were spread plated on SDA. MFC was recorded as the lowest concentration where no colony growth was observed in the three independent plates post 48 h of incubation.
2.4. FAM-FLICA Poly-Caspase Assay
The FAM-FLICA poly-caspase assay was performed according to the manufacturer’s instructions provided in the kit (ImmunoChemistry Technologies, Bloomington, MN) with modifications. The FAM-FLICA reagent has been used in studies to elucidate yeast metacaspase activities [24,25]. For the treatment concentrations, the MIC, 2 x MIC and 4 x MIC of globospiramine against C. albicans and C. tropicalis were used. In the 96-well microplate, Candida cells were exposed to the treatment groups for 24 h. Each wells contained approximately 1 x 106 candidal cells/mL. The FAM-FLICA reagent was then added. After 50 min of incubation at 30 °C, fluorescence readouts were obtained using the Glomax Microplate Discover Reader (Promega) (excitation: 490 nm; emission: 530 nm). Three independent experiments were conducted. For statistical analysis, one-way ANOVA was performed followed by pairwise analysis with DMSO (negative/vehicle control) as the reference (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001).
2.5. Molecular Docking against C. albicans Proteins
2.5.1. Ligand and Protein Preparation
Globospiramine was considered as the ligand, whereas the C. albicans target proteins as receptors and/or molecular targets. The following PDB IDs were used: 1EQP (1,3-β-glucan synthase), 4QUV (δ-14-sterol reductase), 5TZ1 (lanosterol 14-alpha demethylase or CYP51), 5UIV (thymidylate kinase), 4LEB (Als3 adhesin), and 2Y7L (Als9-2). These PDB IDs have already served as key targets in other molecular docking studies and/or are considered pharmaceutical targets of current anticandidal drugs [26,27,28]. To prepare the ligand, its structure was drawn in ChemDraw (18.1), optimized in Avogadro (1.2.0), and saved as .mol2 file. For the protein preparation, non-standard residues were removed in UCSF Chimera (1.17.3), followed by minimization using the steepest descent and conjugate gradient methods. The output file was saved as .pdb [29].
2.5.2. Molecular Docking and Visualization of Interactions
The prepared ligands and protein targets were combined using UCSF Chimera (1.17.3). Actual docking simulation experiments were carried out using the flexible ligand in a flexible active site protocol based on the BFGS algorithm coupled with AutoDock Vina. Grids were generated to encompass the target binding domains. To visualize the interactions, the output files were processed using BIOVIA Discovery (4.1). [30]
2.6. Molecular Dynamics Simulations
MD simulation experiments were conducted using the Desmond package (Desmond Molecular Dynamics System 6.4 academic version, D.E. Shaw Research (“DESRES”), New York, NY, USA, 2020. Maestro-Desmond Interoperability Tools, Schrödinger, New York, NY, USA, 2020). The three-dimensional ligand/protein complexes (globospiramine within 1,3-β-glucan synthase (PDB ID: 1EQP) and Als3 adhesin (PDB ID: 4LEB)) obtained by molecular docking studies were prepared using the system builder tool, available in Desmond software, to produce suitable complexes for MD simulation studies. Accordingly, the ligand/protein complexes were placed into an orthorhombic box and solvated by water molecules (TIP3P water model) [31,32]. A physiological concentration of monovalent ions (0.15 M) was used by adding to the biological systems Na+ and Cl− ions. MD simulation studies were conducted using the OPLS3 force field [33], and calculations were performed utilizing the CUDA API technology on two NVIDIA graphics processing units (GPUs). A constant number of particles, constant temperature (300 K by Nosé–Hoover thermostat method [34], and pressure (1.01325 bar by Martyna–Tobias–Klein method [35] were considered using the NPT ensemble class. To assess the motion for bonded and non-bonded interactions within the short-range cutoff, the RESPA integrator was adopted (inner time step of 2.0 fs) [36]. To calculate long-range electrostatic interactions (short-range electrostatic interactions were fixed at 9.0 Å), the particle mesh Ewald method (PME) was employed [37]. To equilibrate the biological systems, the default protocol available in Desmond was used. The protocol consists of several constrained minimizations and MD simulations that were applied to each biological system to progressively relax and bring them to equilibrium. The Desmond application’s simulation event analysis tools were utilized to examine the MD results produced throughout the MD simulation calculations, as previously reported [38].
3. Results
3.1. Effects of Globospiramine on C. albicans and C. tropicalis CFU Viability
Screening for inhibitory activities of globospiramine against the pathogenic yeast species C. albicans and C. tropicalis, CFUs, were investigated in vitro using a CFU viability assay. Globospiramine exhibited time- and concentration-dependent activities against C. albicans and C. tropicalis by significantly decreasing the percentage of viable CFUs. After 60 min exposure to globospiramine at 2.5 µg/mL, the CFU count for both Candida species was reduced to approximately 50% of the original count (Figure 2).
3.2. MIC and MFC of Globospiramine versus C. albicans and C. tropicalis
The MIC and MFC were determined for both Candida species to support the CFU viability data using microdilution and spread plate techniques. The lowest concentration that significantly prevented increased OD600 was considered as MIC, whereas MFC was the lowest concentration that completely inhibited the growth of yeast colonies. In agreement with the results of the CFU viability assay, globospiramine showed moderately strong MIC (8 ug/mL) and MFC (8 µg/mL) values against C. albicans. (Table 1) compared with the positive standard drug control, amphotericin B. Interestingly, a lower concentration of globospiramine was effective against C. tropicalis (MIC = 4 µg/mL); however, such activity might be limited to growth inhibition, or a much higher concentration (MFC > 64 µg/mL) is necessary to promote fungicidal effects (Table 1).
3.3. Apoptosis-Inducing Activities of Globospiramine vs C. albicans and C. tropicalis
The metacaspase-activating activity of globospiramine vs C. albicans and C. tropicalis cells was also investigated to determine its possible mechanism of action. Globospiramine induced a significant increase in relative fluorescence units (RFUs) compared with the vehicle control DMSO – triggering metacaspase activation responses in C. albicans cells at 16 (p < 0.01) and 8 µg/mL (p < 0.05) test concentrations (Figure 3). These concentrations corroborated to the MIC and MFC values shown in Table 1. Meanwhile, globospiramine did not induce apoptosis in C. tropicalis cells. This is to be expected since the MFC value against C. tropicalis was noted > 64 µg/mL.
3.3. Molecular Docking against C. albicans Targets
Probing the putative molecular targets of globospiramine is the next step after demonstrating its in vitro anti-C. albicans properties. Six protein targets previously reported to play significant roles in the pathogenesis of candidiasis caused by C. albicans were selected for molecular docking experiments, namely 1EQP (1,3-β-glucan synthase), 4QUV (δ-14-sterol reductase), 5TZ1 (lanosterol 14-alpha demethylase or CYP51), 5UIV (thymidylate kinase), 4LEB (Als3 adhesin), and 2Y7L (Als9-2). Globospiramine showed the best binding affinities to 1,3-β-glucan synthase and Als3 adhesin (Table 2, Figure 4A-4B). Compared with the positive controls caspofungin and amphotericin B, our compound showed much better binding energy against all these targets.
3.4. Molecular Dynamics Simulations
To improve the reliability of target identification and validate the docking results, we conducted MD simulation experiments on the most promising targets. In particular, the complexes 1,3-β-glucan synthase/globospiramine and Als3 adhesin/globospiramine were considered for MD simulation studies. Figure 5 shows the MD simulation output for the complex 1,3-β-glucan synthase/globospiramine. Based on 100 ns of MD simulation, we observed a general stability of the selected biological system, highlighted by the low RMSD values of the protein and the ligand and by the low RMSF value, indicating small fluctuations in the biological system. Considering the main interactions found by molecular docking studies, we observed that the H-bond established with residue His253 was maintained, although it became water-mediated. In addition, a strong H-bond network was detected with residue Asp227. Other polar contacts, mainly water-mediated with His254, Glu262, Arg265, and Asp280, were observed. The hydrophobic interactions with Phe229, Tyr255, Phe258, and Trp277 were well maintained during the simulation, with additional hydrophobic contacts with Phe232 and His252.
Regarding the Als3 adhesin/globospiramine complex, the MD simulation results are illustrated in Figure 6. In addition, in this case, we observed a general stability of the biological system with a small fluctuation of the protein, as indicated by the RMSD and RMSF values. Considering the main contacts governing the binding mode of globospiramine within the selected binding site of the Asl3 adhesin, we observed that the H-bond with Thr168 was well maintained as well as the ionic interactions with the reside Asp169. Additional polar contacts that could contribute to stabilizing the binding mode were detected with residues Ala19, Asn22, and Arg294. Hydrophobic interactions with residues Val161, Tyr166, and Leu167 were still evident at low frequencies. More favorable hydrophobic contacts were established from globospiramine with Tyr226 and Trp295.
4. Discussion
Globospiramine is a spirobisindole alkaloid from the Philippine endemic medicinal plant Voacanga globosa. It has been reported to exhibit biological activities including cholinesterase inhibitory, antiviral, and antimycobacterial properties [19,20]. Meanwhile, V. globosa extracts demonstrated anticancer and antifungal activities [21,22]. Generally, indole alkaloids are known to exert antifungal activities [14,39]. For example, decreased viability of fluconazole-resistant C. albicans was observed upon treatment with Tabernaemontana divaricata indole alkaloids [40]. Other studies have reported potential targets of these indole alkaloids, such as isocitrate lyase and other extracellular enzymes implicated in the lipolytic and proteinase activities of C. albicans [15,41].
Among the emerging targets in anticandidal drug discovery is the apoptosis pathway. Herein, we report the apoptosis-inducing potential of globospiramine against C. albicans and C. tropicalis cells using the FAM-FLICA poly-caspase assay. Yeast and mammalian apoptosis share markers such as DNA fragmentation, metacaspase activation, and reactive oxygen species (ROS) accumulation. FAM-FLICA has been reported to bind to the yeast metacaspase, thus being used to assess yeast apoptosis [24,25,42,43]. The antifungal activities of natural products have been investigated in the context of apoptosis induction as a mechanism of their fungicidal action [44,45,46]. Our results indicate that the fungicidal effect of globospiramine against C. albicans occurs through metacaspase activation leading to apoptosis, which might also explain the correlation between MIC, MFC, and metacaspase-inducing activity at the same concentration. Meanwhile, against C. tropicalis, globospiramine may be fungistatic and not fungicidal, although other possible modes of cell death like necrosis may be investigated. Therefore, further mechanistic investigations on the effect of globospiramine in promoting the growth inhibition of C. tropicalis cells are warranted. Thus, our study indicates for the first time that the V. globosa phytoconstituent globospiramine has anticandidal activity and could be responsible for the purported antifungal activity of the medicinal plant.
Based on the molecular docking and MD simulation experimental data, globospiramine potentially exerts its antifungal activity by targeting 1,3-β-glucan synthase and Als3 adhesin. The first putative target, 1,3-β-glucan synthase, is an enzyme important for fungal cell wall synthesis. This enzyme facilitates the creation of β(1→3) glycosidic bonds within 1,3-β-glucan molecules, utilizing uridine diphosphate–activated glucose (UDP-Glc) as the source of sugar and transporting the resulting glucan across the membrane [47]. In general, echinocandins like amphotericin B are known to elicit inhibitory effects on this enzyme in the plasma membrane. Unstable and impaired fungal cell walls then result in morphogenic and intracellular changes that cause cell death [48,49]. In addition, the fact that fungal cell walls and the enzyme itself are absent in human cells make it more ideal as a therapeutic target. Recently, this enzyme has served as a key target among newly discovered compounds and other antifungals [50,51]. Drug-induced damage to the C. albicans cell membrane and cell wall upon inhibition of this enzyme was also reported to result in cellular stress, subsequent apoptosis, and G0/G1 cell cycle arrest [52]. However, there are a myriad of reports on phenotypic changes in the enzyme caused by gene mutations in the coding region of FKS1, a gene involved in biosynthesis of 1,3-β-glucan synthase. As a result, such mutations promoted echinocandin resistance [53,54]. Therefore, the discovery of multitargeting agents against 1,3-β-glucan synthase and other virulence factors and/or molecular entities is deemed a logical strategy.
The second putative target is Als3 adhesin. The ALS gene family encodes cell surface proteins in C. albicans. These proteins function in adhesion to host cells and various surfaces and are thus implicated in biofilm formation. Biofilms are important determinants in yeast infections and are directly associated with drug failure and resistance. Interestingly, ALS proteins, including Als3 adhesin, are not expressed in human cells [55,56,57]. Therefore, Als3 is considered an emerging, pathogenetically relevant target for new generation antifungals against Candida species [58]. It is noticeable that globospiramine showed almost similar binding propensity and stability to this protein target compared with that of 1,3-β-glucan synthase. This strengthens the multitargeting nature of our compound against these two proteins. The fact that few studies on Als3 as a therapeutic target have been reported provides another opportunity for globospiramine to be considered as a promising alkaloidal template against novel molecular targets in C. albicans.
This study reports the possible multitargeting antagonistic effects of globospiramine on C. albicans. Two scenarios are possible: (1) these effects are independent from each other, such as the case of the anticandidal drug caspofungin, which exhibits concentration-dependent mechanisms against C. albicans, including inhibition of 1,3-β-glucan synthesis, promotion of apoptosis, and induction of necrosis [59], or (2) the inhibition of these proteins may directly or indirectly result in apoptosis and/or necrosis. It is important to note that previous studies have indicated the role of these enzymes in apoptosis. For example, 1,3-β-glucan synthase is involved in cell wall synthesis. Globospiramine effectively bound to 1,3-β-glucan synthase may potentially interrupt the normal biosynthetic pathway to polymerize the candidal cell wall leading to ROS hyperaccumulation – a hallmark of apoptosis [60]. Rapid killing of C. albicans by certain set of antimicrobial peptides may be due to apoptosis preceded by cell wall disruption and ROS accumulation [61]. A lipopeptide has also been reported to inhibit C. albicans growth by directly impairing the fungal cell wall, leading to an increase in ROS levels and mitochondrial dysfunction, which can activate apoptotic pathways [62]. Inhibition of Als3 on the other hand triggers stress signals and responses due to loss of adhesion-dependent survival signals in C. albicans. These stress signals can activate the apoptotic machinery via oxidative stress, DNA damage, or mitochondrial impairment. Additionally, biofilm formation may be disrupted upon inhibition of this protein. In fact, apoptosis has been reported in Candida cells under the therapeutic pressure of amphotericin B [46,63].
5. Conclusions
Overall, our study reports the fungicidal potential of the spirobisindole alkaloid globospiramine from the Philippine medicinal plant Voacanga globosa, particularly against Candida albicans via apoptotic-aided mechanisms, along with potential dual-inhibitory activity against the disease-implicated targets 1,3-β-glucan synthase and Als3 adhesin. Our in vitro findings also demonstrated the fungistatic effects of globospiramine against C. tropicalis. Accordingly, globospiramine could represent a good biomolecular candidate for exploring and discovering anticandidal drugs with improved therapeutic effects.
Author Contributions
Conceptualization, J.A.H.M., A.P.G.M. and N.A.; methodology, J.A.H.M., S.B. and V.C.; software, J.A.H.M., and S.B.; validation, J.A.H.M., S.B., V.C., A.P.G.M. and N.A.; formal analysis, J.A.H.M., S.B., V.C., A.P.G.M. and N.A.; investigation, , J.A.H.M., S.B., V.C., A.P.G.M. and N.A.; resources, S.B., V.C., A.P.G.M. and N.A.; data curation, J.A.H.M. and S.B.; writing—original draft preparation, J.A.H.M.; writing—review and editing, J.A.H.M., S.B., V.C., A.P.G.M. and N.A.; visualization, J.A.H.M. and S.B.; supervision, S.B., A.P.G.M. and N.A.; project administration, S.B., V.C., A.P.G.M. and N.A.; funding acquisition, J.A.H.M., A.P.G.M. and N.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the Department of Science and Technology – Science Education Institute (DOST-SEI) of the Republic of the Philippines under the Accelerated Science and Technology Human Research Development Program (ASTHRDP) provided to J.A.H.M. The Large Equipment Grant by the Alexander von Humboldt Foundation (3.4-8151 / 20 006) to APGM.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author/s.
Acknowledgments
J.A.H.M. acknowledges research assistants Jonah Ray Hernandez, Gabriel Mendoza, and Marc Martin Sagolili for their help in some aspects of the experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vandeputte, P.; Ferrari, S.; Coste, A.T. Antifungal resistance and new strategies to control fungal infections. International Journal of Microbiology 2012, 1–26. [Google Scholar] [CrossRef]
- Chang, Y.-L.; Yu, S.-J.; Heitman, J.; Wellington, M.; Chen, Y.-L. New facets of antifungal therapy. Virulence 2016, 8, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Campoy, S.; Adrio, J.L. Antifungals. Biochemical Pharmacology 2017, 133, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A. Antifungal drug resistance: Mechanisms, epidemiology, and consequences for treatment. The American Journal of Medicine 2012, 125. [Google Scholar] [CrossRef] [PubMed]
- Tobudic, S.; Kratzer, C.; Presterl, E. Azole-resistant Candida spp. – emerging pathogens? Mycoses 2012, 55, 24–32. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Science Translational Medicine 2012, 4. [Google Scholar] [CrossRef]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. The Lancet Infectious Diseases 2011, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Eggimann, P.; Garbino, J.; Pittet, D. Epidemiology of Candida species infections in critically ill non-immunosuppressed patients. The Lancet Infectious Diseases 2003, 3, 685–702. [Google Scholar] [CrossRef]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: Dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Martins, N.; Ferreira, I.C.; Barros, L.; Silva, S.; Henriques, M. Candidiasis: Predisposing factors, prevention, diagnosis and alternative treatment. Mycopathologia 2014, 177, 223–240. [Google Scholar] [CrossRef]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Letters in Applied Microbiology 2018, 66, 2–13. [Google Scholar] [CrossRef] [PubMed]
- 8. Heard, S.C.; Wu, G.; Winter, J.M. Antifungal natural products. Current Opinion in Biotechnology 2021, 69, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Onishi, J.; Meinz, M.; Thompson, J.; Curotto, J.; Dreikorn, S.; Rosenbach, M.; Douglas, C.; Abruzzo, G.; Flattery, A.; Kong, L.; Cabello, A.; Vicente, F.; Pelaez, F.; Diez, M.T.; Martin, I.; Bills, G.; Giacobbe, R.; Dombrowski, A.; Schwartz, R.; Morris, S.; Harris, G.; Tsipouras, A.; Wilson, K.; Kurtz, M.B. Discovery of novel antifungal (1,3)-β-glucan synthase inhibitors. Antimicrobial Agents and Chemotherapy 2000, 44, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Long, S.-Y.; Li, C.-L.; Hu, J.; Zhao, Q.-J.; Chen, D. Indole alkaloids from the aerial parts of Kopsia fruticosa and their cytotoxic, antimicrobial and antifungal activities. Fitoterapia 2018, 129, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, M.; Dimitrova, P.; Patkar, S.; Saso, L.; Ivanovska, N. Inhibition of Candida albicans extracellular enzyme activity by selected natural substances and their application in Candida infection. Canadian Journal of Microbiology 2008, 54, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.-B.; He, G.; Bai, H.-H.; Yang, T.; Zhang, G.-L.; Wu, L.-W.; Li, G.-Y. Indole alkaloids from Chaetomium globosum. Journal of Natural Products 2015, 78, 1479–1485. [Google Scholar] [CrossRef]
- Ahmed, A.; Li, W.; Chen, F.-F.; Zhang, J.-S.; Tang, Y.-Q.; Chen, L.; Tang, G.-H.; Yin, S. Monoterpene indole alkaloids from Rhazya stricta. Fitoterapia 2018, 128, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Anjum, K.; Kaleem, S.; Yi, W.; Zheng, G.; Lian, X.; Zhang, Z. Novel antimicrobial indolepyrazines A and B from the marine-associated Acinetobacter sp.. ZZ1275. Marine Drugs 2019, 17, 89. [Google Scholar] [CrossRef]
- Macabeo, A.P.; Vidar, W.S.; Chen, X.; Decker, M.; Heilmann, J.; Wan, B.; Franzblau, S.G.; Galvez, E.V.; Aguinaldo, Ma. A.; Cordell, G.A. Mycobacterium tuberculosis and cholinesterase inhibitors from Voacanga globosa. European Journal of Medicinal Chemistry 2011, 46, 3118–3123. [Google Scholar] [CrossRef]
- de Jesus, Ma.; Macabeo, A.; Ramos, J.; de Leon, V.; Asamitsu, K.; Okamoto, T. Voacanga globosa spirobisindole alkaloids exert antiviral activity in HIV latently infected cell lines by targeting the NF-ΚB CASCADE: In vitro and in silico investigations. Molecules 2022, 27, 1078. [Google Scholar] [CrossRef]
- Acebedo, A.R.; Amor, E.C.; Jacinto, S.D. Apoptosis-inducing activity of HPLC fraction from Voacanga globosa (Blanco) Merr. on the human colon carcinoma cell line, HCT116. Asian Pacific Journal of Cancer Prevention 2014, 15, 617–622. [Google Scholar] [CrossRef]
- Vital, P.G.; Rivera, W.L. Antimicrobial activity, cytotoxicity, and phytochemical screening of Voacanga globosa (Blanco) Merr. leaf extract (Apocynaceae). Asian Pacific Journal of Tropical Medicine 2011, 4, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Cascio, V.; Gittings, D.; Merloni, K.; Hurton, M.; Laprade, D.; Austriaco, N. S-adenosyl-L-methionine protects the probiotic yeast, Saccharomyces boulardii, from acid-induced cell death. BMC Microbiology 2013, 13. [Google Scholar] [CrossRef]
- Gardner, J. The Effect of Acute Heavy Metal (Cu And Cd) Toxicity on ROS Generation, Apoptosis, and Intracellular Glutathione Levels in Saccharomyces cerevisiae. Master’s Thesis, Georgia State University, Atlanta, Georgia, 2019. [Google Scholar]
- Poling, B.M. Differential Effects of Acute Cadmium, Copper, and Chromium Assault on Glutathione and Transcription Profiles in Saccharomyces cerevisiae. Master’s Thesis, Georgia State University, Atlanta, Georgia, 2021. [Google Scholar]
- da Nóbrega Alves, D.; Monteiro, A.F.; Andrade, P.N.; Lazarini, J.G.; Abílio, G.M.; Guerra, F.Q.; Scotti, M.T.; Scotti, L.; Rosalen, P.L.; Castro, R.D. Docking Prediction, antifungal activity, anti-biofilm effects on Candida spp., and toxicity against human cells of cinnamaldehyde. Molecules 2020, 25, 5969. [Google Scholar] [CrossRef] [PubMed]
- Gurgel do Amaral Valente Sá, L.; da Silva, C.R.; Neto, J.B.; do Nascimento, F.B.; Barroso, F.D.; da Silva, L.J.; Cabral, V.P.; Barbosa, A.D.; Silva, J.; Marinho, E.S.; de Moraes, M.O.; Rios, M.E.; Cavalcanti, B.C.; Lima, I.S.; Júnior, H.V. Antifungal activity of etomidate against growing biofilms of fluconazole-resistant Candida spp. strains, binding to mannoproteins and molecular docking with the ALS3 protein. Journal of Medical Microbiology 2020, 69, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Bouamrane, S.; Khaldan, A.; Hajji, H.; El-mernissi, R.; Alaqarbeh, M.; Alsakhen, N.; Maghat, H.; Ajana, M.A.; Sbai, A.; Bouachrine, M.; Lakhlifi, T. In silico identification of 1,2,4-triazoles as potential Candida albicans inhibitors using 3D-QSAR, molecular docking, molecular dynamics simulations, and ADMET profiling. Molecular Diversity 2022, 27, 2111–2132. [Google Scholar]
- Manzano, J.A.; Cruz, C.L.; Quimque, M.T.; Macabeo, A.P. In silico potentials of Alpinia galanga constituents against human placental aromatase vital in postmenopausal estrogen-dependent breast cancer pathogenesis. Philippine Journal of Science 2022, 151. [Google Scholar] [CrossRef]
- Manzano, J.A.; Llames, L.C.; Macabeo, A.P. Tetrahydrobisbenzylisoquinoline alkaloids from Phaeanthus ophthalmicus inhibit target enzymes associated with type 2 diabetes and obesity. Journal of Applied Pharmaceutical Science 2023. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. The Journal of Chemical Physics 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Brogi, S.; Rossi, S.; Ibba, R.; Butini, S.; Calderone, V.; Campiani, G.; Gemma, S. In silico analysis of peptide-based derivatives containing bifunctional warheads engaging prime and non-prime subsites to covalent binding SARS-COV-2 main protease (mpro). Computation 2022, 10, 69. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. Journal of the American Chemical Society 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Physical Review A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [PubMed]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. The Journal of Chemical Physics 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Humphreys, D.D.; Friesner, R.A.; Berne, B.J. A multiple-time-step molecular dynamics algorithm for Macromolecules. The Journal of Physical Chemistry 1994, 98, 6885–6892. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. The Journal of Chemical Physics 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- da Silva, E.R.; Brogi, S.; Lucon-Júnior, J.F.; Campiani, G.; Gemma, S.; Maquiaveli, C. Dietary polyphenols rutin, taxifolin and quercetin related compounds target Leishmania amazonensis arginase. Food & Function 2019, 10, 3172–3180. [Google Scholar]
- Peng, F.; Hou, S.-Y.; Zhang, T.-Y.; Wu, Y.-Y.; Zhang, M.-Y.; Yan, X.-M.; Xia, M.-Y.; Zhang, Y.-X. Cytotoxic and antimicrobial indole alkaloids from an endophytic fungus Chaetomium sp. SYP-F7950 of Panax notoginseng. RSC Advances 2019, 9, 28754–28763. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, X.; Yuwen, H.-S.; Guo, L.-L.; Liu, J.-W.; Hao, X.-J. Alkaloids from Tabernaemontana divaricata combined with fluconazole to overcome fluconazole resistance in Candida albicans. Bioorganic Chemistry 2021, 107, 104515. [Google Scholar] [CrossRef]
- Lee, H.-S.; Yoon, K.-M.; Han, Y.-R.; Lee, K.J.; Chung, S.-C.; Kim, T.-I.; Lee, S.-H.; Shin, J.; Oh, K.-B. 5-hydroxyindole-type alkaloids, as Candida albicans isocitrate lyase inhibitors, from the tropical sponge Hyrtios sp. Bioorganic & Medicinal Chemistry Letters 2009, 19, 1051–1053. [Google Scholar]
- Leadsham, J.E.; Kotiadis, V.N.; Tarrant, D.J.; Gourlay, C.W. Apoptosis and the yeast actin cytoskeleton. Cell Death & Differentiation 2009, 17, 754–762. [Google Scholar]
- Al-Dhaheri, R.S.; Douglas, L.J. Apoptosis in Candida biofilms exposed to amphotericin B. Journal of Medical Microbiology 2010, 59, 149–157. [Google Scholar] [CrossRef]
- da Silva, C.R.; de Andrade Neto, J.B.; de Sousa Campos, R.; Figueiredo, N.S.; Sampaio, L.S.; Magalhães, H.I.; Cavalcanti, B.C.; Gaspar, D.M.; de Andrade, G.M.; Lima, I.S.; de Barros Viana, G.S.; de Moraes, M.O.; Lobo, M.D.; Grangeiro, T.B.; Nobre Júnior, H.V. Synergistic effect of the flavonoid catechin, quercetin, or epigallocatechin gallate with fluconazole induces apoptosis in Candida tropicalis resistant to fluconazole. Antimicrobial Agents and Chemotherapy 2014, 58, 1468–1478. [Google Scholar] [CrossRef]
- Soliman, S.; Alnajdy, D.; El-Keblawy, A.; Mosa, K.; Khoder, G.; Noreddin, A. Plants’ natural products as alternative promising anti-candida drugs. Pharmacognosy Reviews 2017, 11, 104. [Google Scholar] [CrossRef]
- Jia, C.; Zhang, J.; Yu, L.; Wang, C.; Yang, Y.; Rong, X.; Xu, K.; Chu, M. Antifungal activity of coumarin against Candida albicans is related to apoptosis. Frontiers in Cellular and Infection Microbiology 2019, 8. [Google Scholar] [CrossRef]
- Zhao, C.-R.; You, Z.-L.; Chen, D.-D.; Hang, J.; Wang, Z.-B.; Ji, M.; Wang, L.-X.; Zhao, P.; Qiao, J.; Yun, C.-H.; Bai, L. Structure of a fungal 1,3-β-glucan synthase. Science Advances 2023, 9. [Google Scholar] [CrossRef]
- Denning, D.W. Echinocandin antifungal drugs. The Lancet 2003, 362, 1142–1151. [Google Scholar]
- Odds, F.C.; Brown, A.J. P.; Gow, N.A. R. Antifungal agents: Mechanisms of action. Trends in Microbiology 2003, 11, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.; Latge, J.-P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiology Spectrum 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal cell wall: Emerging antifungals and drug resistance. Frontiers in Microbiology 2019, 10. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, Y. Antifungal activity of Salvia miltiorrhiza against Candida albicans is associated with the alteration of membrane permeability and (1,3)-β-D-glucan synthase activity. Journal of Microbiology and Biotechnology 2016, 26, 610–617. [Google Scholar] [CrossRef]
- Perlin, D.S. Resistance to echinocandin-class antifungal drugs. Drug Resistance Updates 2007, 10, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Edlind, T.D. Topological and mutational analysis of Saccharomyces cerevisiae FKS1. Eukaryotic Cell 2012, 11, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Filler, S.G. Candida albicans ALS3, a multifunctional adhesin and Invasin. Eukaryotic Cell 2011, 10, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Cota, E. Candida albicans agglutinin-like sequence (ALS) Family Vignettes: A review of ALS protein structure and function. Frontiers in Microbiology 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kioshima, E.S.; Shinobu-Mesquita, C.S.; Abadio, A.K.; Felipe, M.S.; Svidzinski, T.I.; Maigret, B. Selection of potential anti-adhesion drugs by in silico approaches targeted to ALS3 from Candida albicans. Biotechnology Letters 2019, 41, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.R.; Sardi, J. de; Freires, I.A.; Silva, A.C.; Rosalen, P.L. In silico approaches for screening molecular targets in Candida albicans: A proteomic insight into drug discovery and development. European Journal of Pharmacology 2019, 842, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Cheng, S.; Clancy, C.J.; Nguyen, M.H. Caspofungin kills Candida albicans by causing both cellular apoptosis and necrosis. Antimicrobial Agents and Chemotherapy 2013, 57, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhang, B.; Li, J.; Zhang, B.; Wang, H.; Li, M. Endoplasmic reticulum-derived reactive oxygen species (ROS) is involved in toxicity of cell wall stress to Candida albicans. Free Radical Biology and Medicine 2016, 99, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Maurya, I.K.; Pathak, S.; Sharma, M.; Sanwal, H.; Chaudhary, P.; Tupe, S.; Deshpande, M.; Chauhan, V.S.; Prasad, R. Antifungal activity of novel synthetic peptides by accumulation of reactive oxygen species (ROS) and disruption of cell wall against Candida albicans. Peptides 2011, 32, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, J.; Sun, J.; Zhu, X.; Zhou, L.; Lu, Z.; Lu, Y. C16-Fengycin a affect the growth of Candida albicans by destroying its cell wall and accumulating reactive oxygen species. Applied Microbiology and Biotechnology 2019, 103, 8963–8975. [Google Scholar] [CrossRef]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proceedings of the National Academy of Sciences 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of globospiramine.
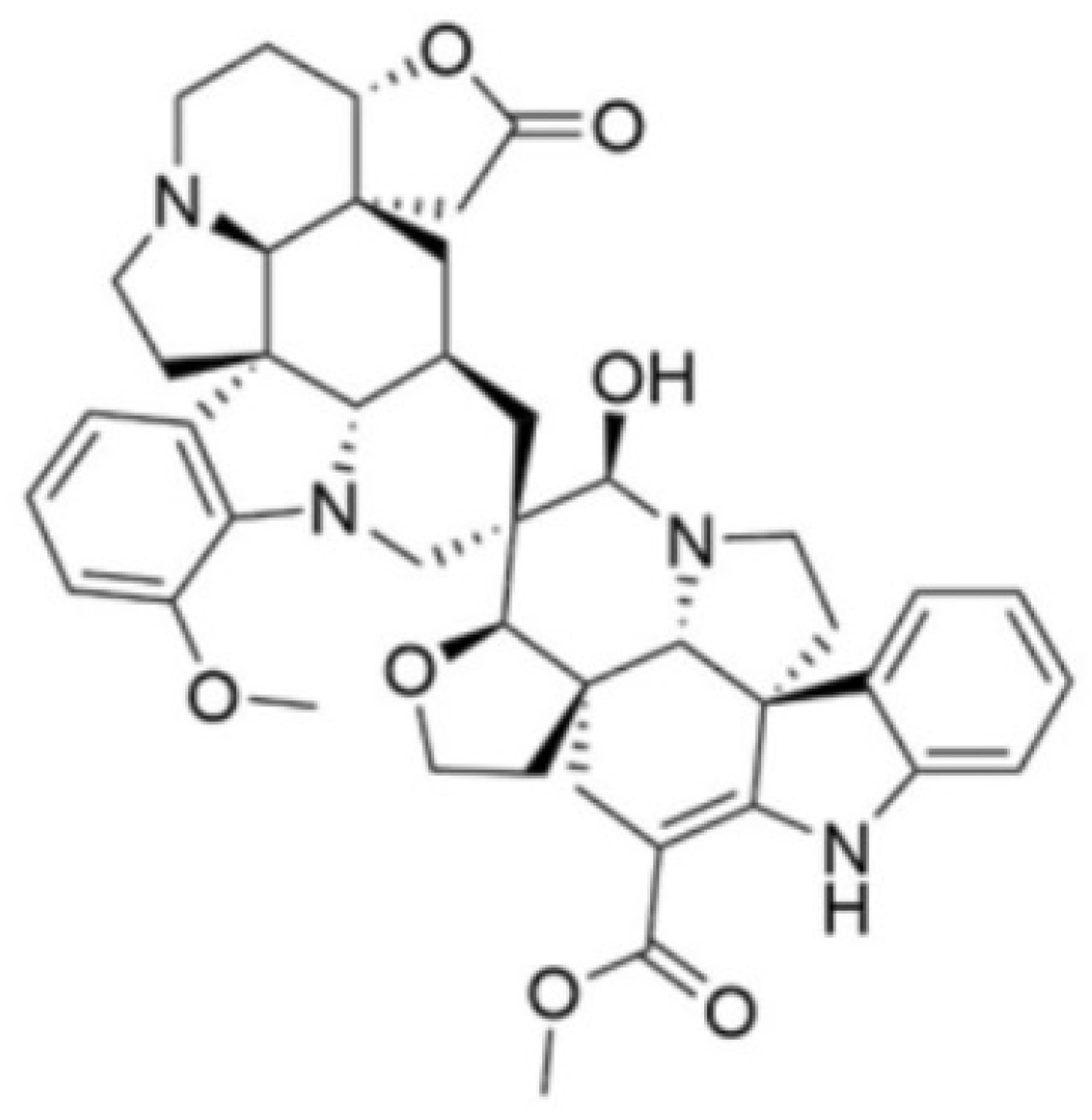
Figure 2.
Globospiramine demonstrated time- and concentration-dependent effects on the CFU viability of C. albicans and C. tropicalis.
Figure 2.
Globospiramine demonstrated time- and concentration-dependent effects on the CFU viability of C. albicans and C. tropicalis.
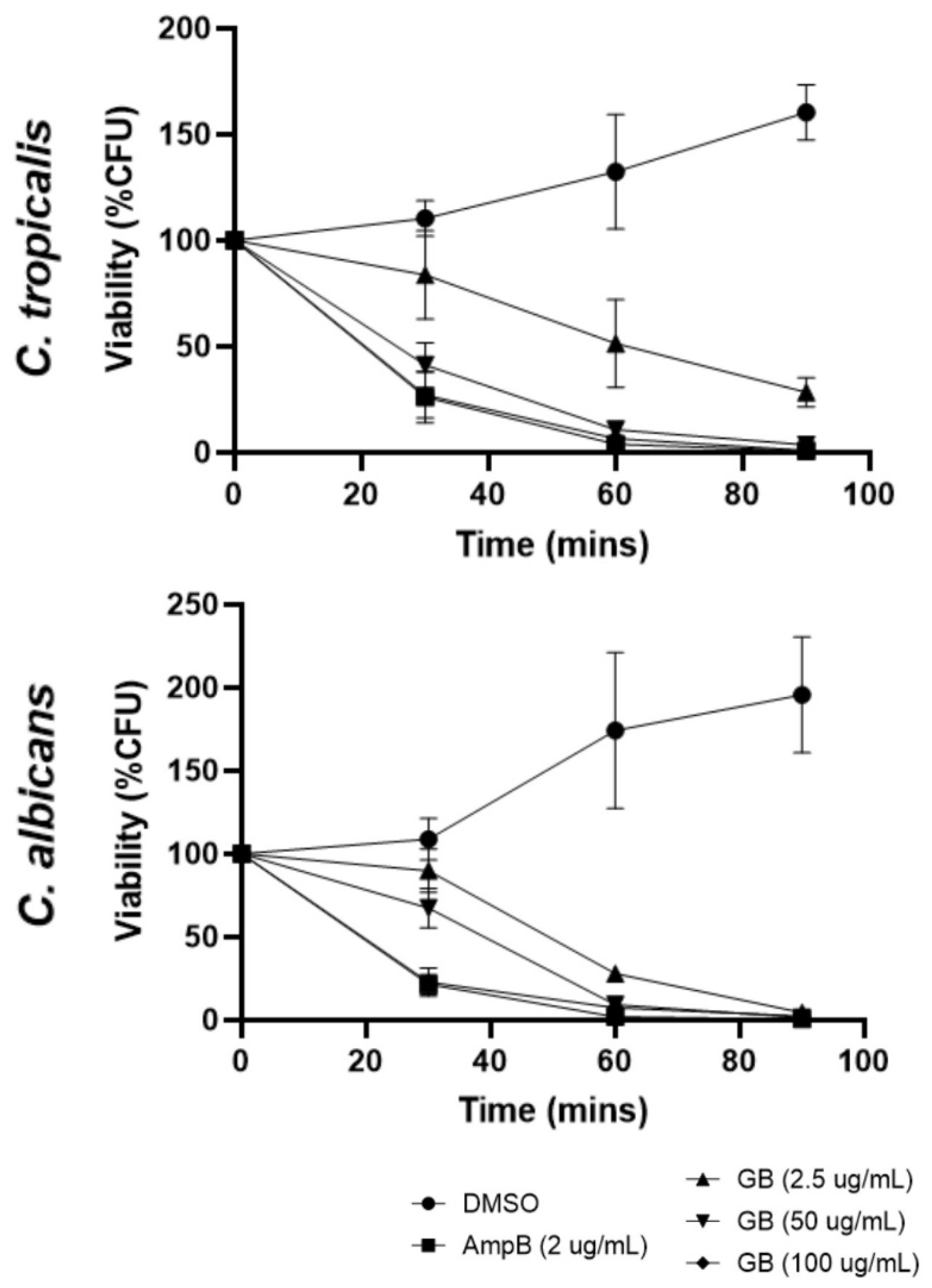
Figure 3.
The spirobisindole alkaloid globospiramine significantly induced apoptosis in (A) C. albicans cells at 16 and 8 µg/mL concentrations, which agreed with its MIC and MFC. However, these effects were not observed in (B) C. tropicalis cells (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001). DMSO was used as a negative control and amphotericin B (AmpB) (0.5 µg/mL) as a positive control.
Figure 3.
The spirobisindole alkaloid globospiramine significantly induced apoptosis in (A) C. albicans cells at 16 and 8 µg/mL concentrations, which agreed with its MIC and MFC. However, these effects were not observed in (B) C. tropicalis cells (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001). DMSO was used as a negative control and amphotericin B (AmpB) (0.5 µg/mL) as a positive control.
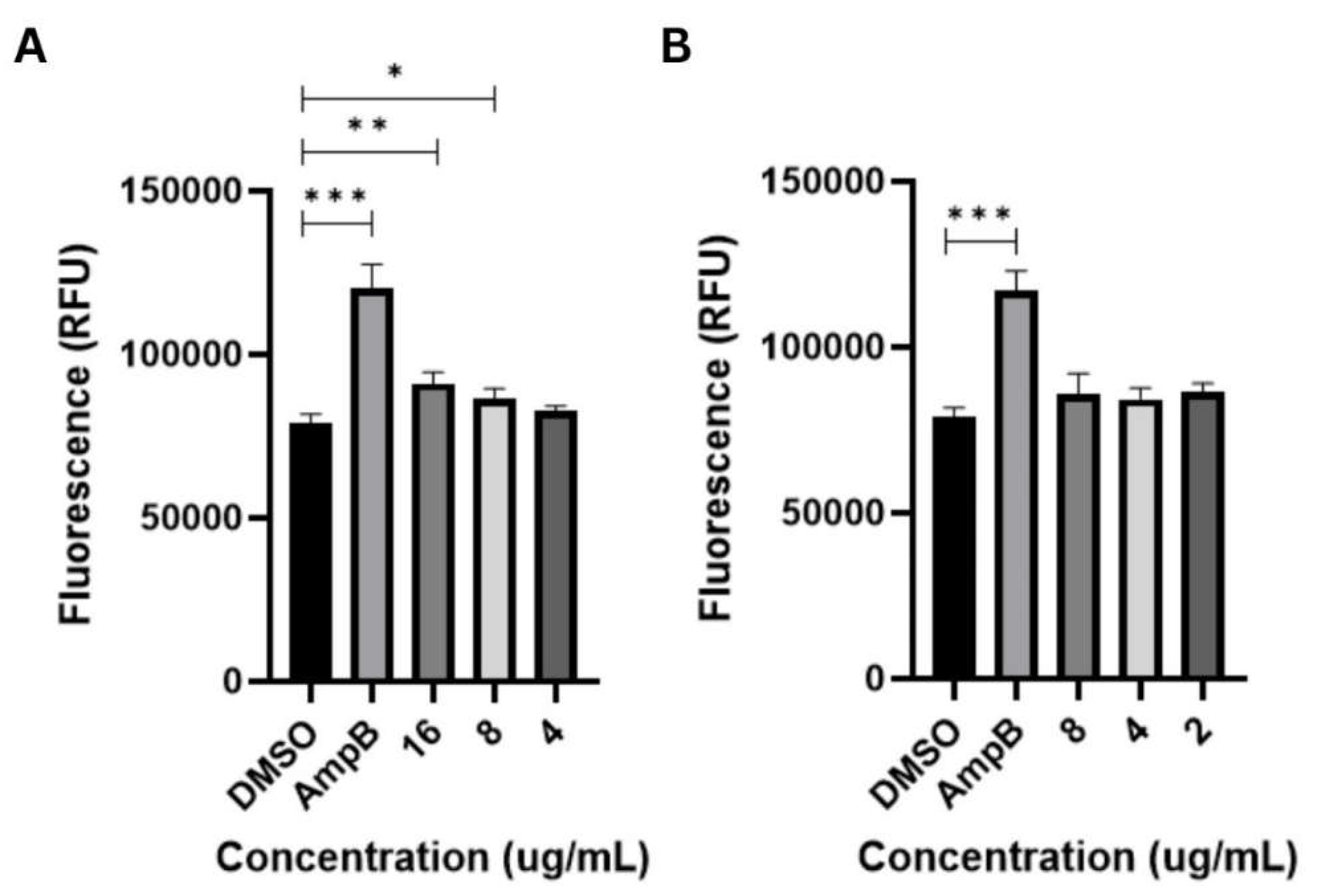
Figure 4.
Dock poses of globospiramine to (A) 1,3-β-glucan synthase and (B) Als3 adhesin.
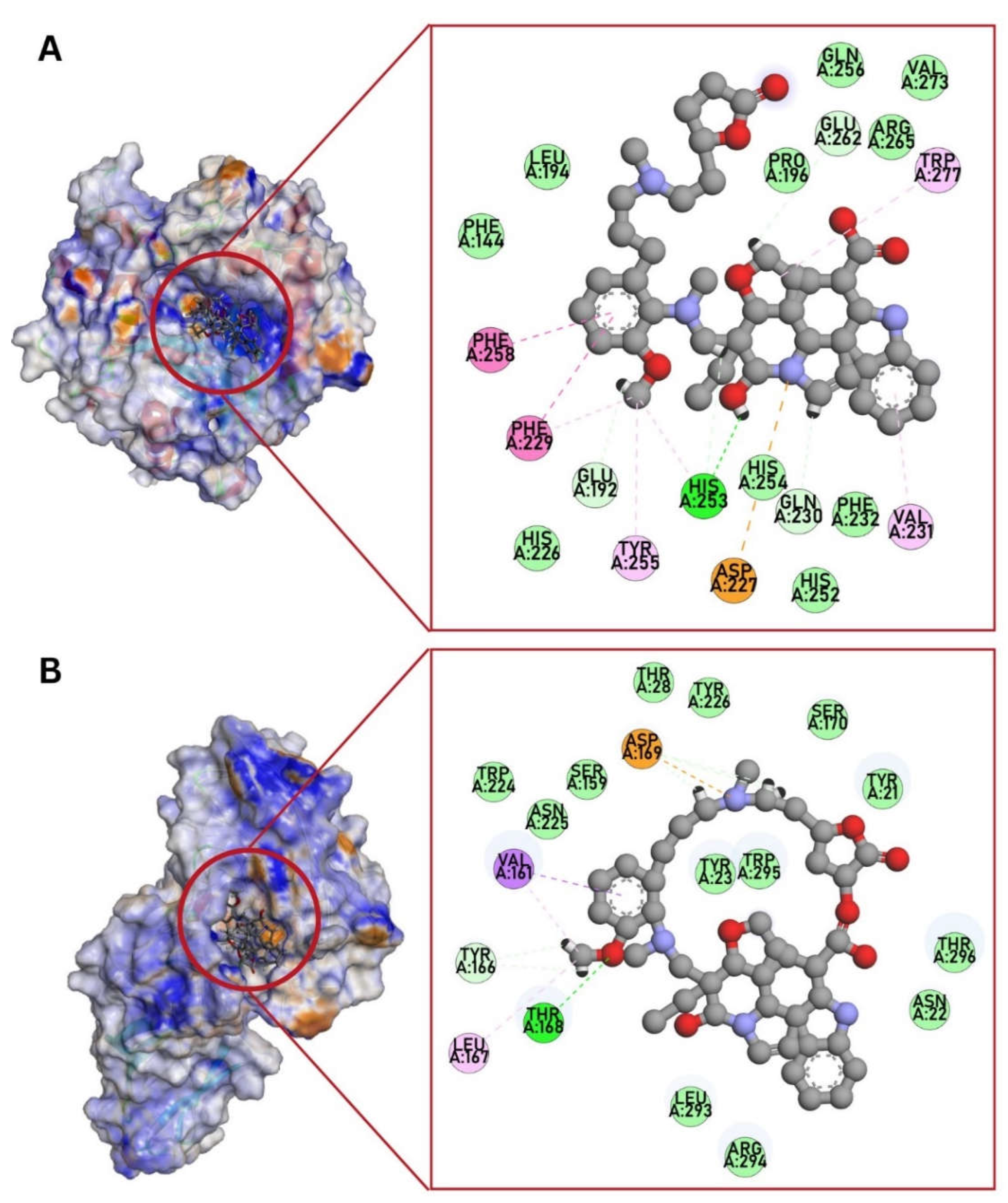
Figure 5.
(A) RMSD evaluation (protein: blue line; and ligand: red line). (B) RMSF assessment for the complex 1,3-β-glucan synthase (1EQP)/globospiramine, obtained by docking studies, following a 100 ns MD simulation. (C, D) globospiramine observed throughout the MD run. Four types of interactions can be distinguished: water bridges (blue), ionic (magenta), hydrophobic (grey), and H-bonds (green). Over the trajectory, the stacked bar charts are normalized. For instance, a value of 0.7 indicates that a particular contact is maintained 70% of the time during simulation. Values greater than 1.0 could occur because a protein residue could interact with the ligand more than once using the same subtype. A timeline explanation of the primary interactions is shown in the following diagram in the figure. Those residues that interact with the ligand in each trajectory frame are displayed in the output. A darker orange hue denotes several contacts that some residues have with the ligand. Maestro and Desmond software tools were utilized to generate the pictures (Maestro, Schrödinger LLC, release 2020-3).
Figure 5.
(A) RMSD evaluation (protein: blue line; and ligand: red line). (B) RMSF assessment for the complex 1,3-β-glucan synthase (1EQP)/globospiramine, obtained by docking studies, following a 100 ns MD simulation. (C, D) globospiramine observed throughout the MD run. Four types of interactions can be distinguished: water bridges (blue), ionic (magenta), hydrophobic (grey), and H-bonds (green). Over the trajectory, the stacked bar charts are normalized. For instance, a value of 0.7 indicates that a particular contact is maintained 70% of the time during simulation. Values greater than 1.0 could occur because a protein residue could interact with the ligand more than once using the same subtype. A timeline explanation of the primary interactions is shown in the following diagram in the figure. Those residues that interact with the ligand in each trajectory frame are displayed in the output. A darker orange hue denotes several contacts that some residues have with the ligand. Maestro and Desmond software tools were utilized to generate the pictures (Maestro, Schrödinger LLC, release 2020-3).
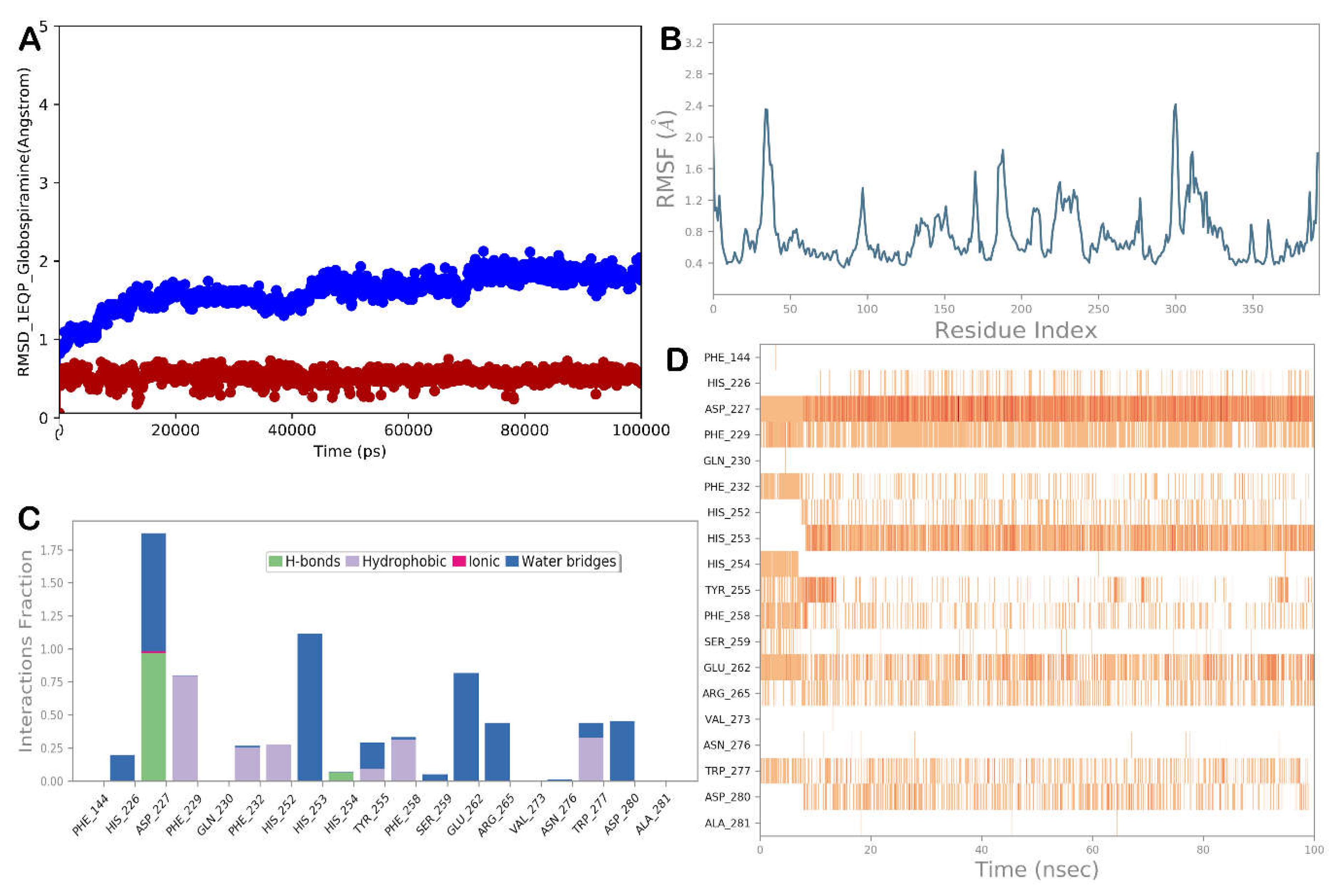
Figure 6.
(A) RMSD evaluation (protein: blue line; and ligand: red line). (B) RMSF assessment for the complex Als3 adhesin (4LEB)/globospiramine, obtained by docking studies, following a 100 ns MD simulation. (C, D) globospiramine observed throughout the MD run. Four types of interactions can be distinguished: water bridges (blue), ionic (magenta), hydrophobic (grey), and H-bonds (green). Over the trajectory, the stacked bar charts are normalized. For instance, a value of 0.7 indicates that a particular contact is maintained 70% of the time during simulation. Values greater than 1.0 could occur because a protein residue could interact with the ligand more than once using the same subtype. A timeline explanation of the primary interactions is shown in the following diagram in the figure. Those residues that interact with the ligand in each trajectory frame are displayed in the output. A darker orange hue denotes several contacts that some residues have with the ligand. Maestro and Desmond software tools were utilized to generate the pictures (Maestro, Schrödinger LLC, release 2020-3).
Figure 6.
(A) RMSD evaluation (protein: blue line; and ligand: red line). (B) RMSF assessment for the complex Als3 adhesin (4LEB)/globospiramine, obtained by docking studies, following a 100 ns MD simulation. (C, D) globospiramine observed throughout the MD run. Four types of interactions can be distinguished: water bridges (blue), ionic (magenta), hydrophobic (grey), and H-bonds (green). Over the trajectory, the stacked bar charts are normalized. For instance, a value of 0.7 indicates that a particular contact is maintained 70% of the time during simulation. Values greater than 1.0 could occur because a protein residue could interact with the ligand more than once using the same subtype. A timeline explanation of the primary interactions is shown in the following diagram in the figure. Those residues that interact with the ligand in each trajectory frame are displayed in the output. A darker orange hue denotes several contacts that some residues have with the ligand. Maestro and Desmond software tools were utilized to generate the pictures (Maestro, Schrödinger LLC, release 2020-3).
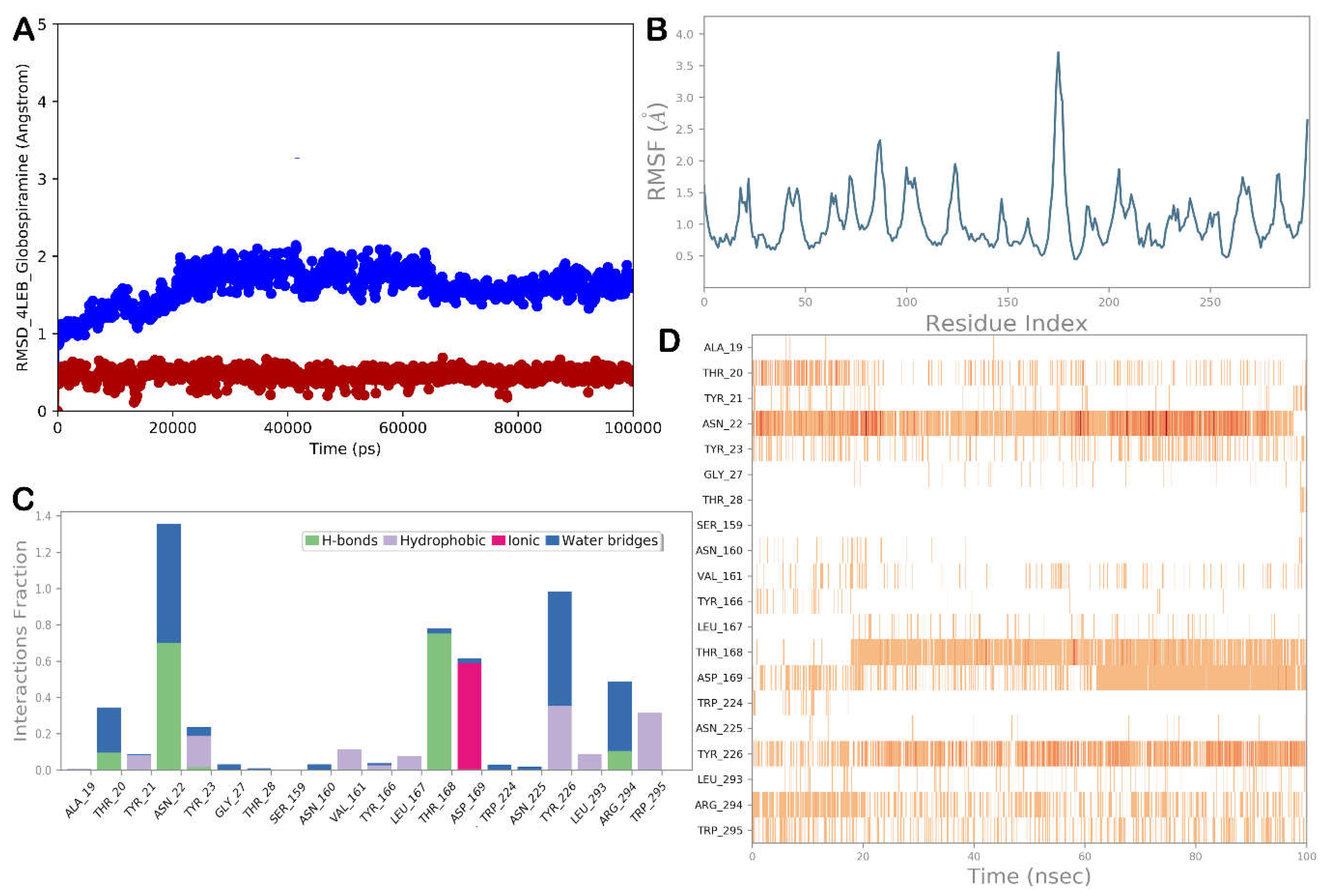

Table 1.
MIC and MFC of globospiramine and positive control amphotericin B against C. albicans and C. tropicalis.
Table 1.
MIC and MFC of globospiramine and positive control amphotericin B against C. albicans and C. tropicalis.
| Globospiramine | Amphotericin B | |
|---|---|---|
| MIC (µg/mL) | ||
| C. albicans | 8.0 | 0.50 |
| C. tropicalis | 4.0 | 0.50 |
| MFC (µg/mL) | ||
| C. albicans | 10.67 | 0.83 |
| C. tropicalis | >64.0 | 1.67 |
Table 2.
Binding energies and interactions of globospiramine against C. albicans protein targets.
| PDB IDs | Globospiramine | Positive Controls | ||||||
|---|---|---|---|---|---|---|---|---|
| Caspofungin | Amphotericin B | Co-crystallized ligand / inhibitor | ||||||
| BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | |
| 1EQP (1,3-β-glucan synthase) | -10.5 | His253 (H-bond), Phe258, Phe229 (pi-pi stacked), Trp277, Val231, Tyr255, Phe229, His253 (pi-alkyl), Glu192, His253, Gln230, Glu262 (C-H bond) | -8.0 | Trp277, Gln230 (H-bond), Asp227 (salt bridge), His254, Glu262 (C-H bond), Val273, Phe258, Phe144, Tyr255, Trp373 (alkyl, pi-alkyl), His253, Arg265 (unfavorable interaction) | -8.8 | Asn305, Asp151 (H-bond), Phe258, Phe144 (pi-alkyl), Tyr 153 (unfavorable donor-donor, C-H bond) | - | - |
| 4QUV (δ-14-sterol reductase) | -9.5 | Arg324, His320 (H-bond), Val96, His320 (pi-sigma), Leu253, Met99 (alkyl), Arg106, Arg323 (unfavorable positive-positive) | -7.2 | Arg106, Arg323, Arg324, Lys406 (H-bond), Tyr414, Trp352, Leu346, Cys403, Trp411, Lys319, Val96 (alkyl, pi-alkyl), His320 (pi-pi stacked), Gln97 (C-H bond), Arg324 (unfavorable positive-positive) | -7.5 | Gln97, Glu250, Arg323, Arg324, Gly343 (H-bond), Met99, Leu253 (alkyl), Arg106 (unfavorable positive-positive) | -9.5 | His248, Arg313, Thr254, Lys259, Lys319, Trp256, Arg395, Asn316, Thr255 (H-bond), Asp244, Asp399, Arg395 (attractive charge, pi-cation), Glu201 (C-H bond), Lys319 (unfavorable positive-positive), Tyr245 (pi-pi T-shaped), Arg398, Val252 (pi-alkyl, alkyl) |
| 5TZ1 (lanosterol 14-alpha demethylase or CYP51) | -7.4 | Arg469 (H-bond), Glu444 (attractive charge), Val452, Val454 (alkyl), Ser453, Lys451 (C-H bond) | -5.7 | Met508, Pro462, His468, Leu439, Leu471, Gly303 (H-bond), His468 (C-H bond), Ile304 (pi-sigma), Leu87, Phe233, Tyr64, Phe380, Phe228, Val509, Leu150, Ile304, Ile131, His377, Pro230, Leu88, Lys90 (pi-alkyl, alkyl), Arg381, Tyr132, Lys143 (unfavorable interactions) | -3.3 | Phe463 (H-bond), His468 (C-H bond), Tyr118 (pi-lone pair), Leu376, Ile379, Ala146, Ile304, Leu204, Phe475 (alkyl, pi-alkyl), Cys470, Ile379, Gly464, Arg381, Thr311, Phe475, Leu150, Ile471, Tyr132 (unfavorable bonds) | -10.6 | Gly303, Ile304 (C-H bond), Ser507, His377 (halogen), Tyr118, Tyr132 (pi-pi), Leu121, Phe233, Leu376, Pro230, Ile304, Ile131, Lys143 (alkyl, pi-alkyl) |
| 5UIV (thymidylate kinase) | -9.4 | Gly155, Asp91, Arg39 (H-bond), Asp13, Arg39, Glu159 (pi-cation / pi-anion / salt bridge), Glu159, Ser18 (C-H bond), Lys17 (pi-alkyl) | -8.2 | Ser18, Asp13, Asp91, Arg92, Lys17, Lys35, Arg39, Gly157 Gly155 (H-bond), Glu162, Glu159 (salt bridge, attractive charge), Asp13, Asp91, Lys35, Gly155 (C-H bond), Ile196, Arg153, Lys17, Arg39, Val199 (alkyl, pi-alkyl) | -7.7 | Arg92, Lys35, SerA (H-bond), Glu162, Gln159 (salt bridge, attractive charge), Pro37 (alkyl), Asp13 (C-H bond), Ser18 (unfavorable donor-donor) | -8.9 | Arg92, Lys17, Arg14, Ser18, Gly16 (H-bond), Glu159, Asp91, Asp13 (attractive charge, pi-anion), Lys35 (unfavorable donor-donor), Tyr100 (pi-pi), Leu51 (pi-alkyl) |
| 4LEB (Als3 adhesin) | -10.6 | Thr168 (H-bond), Asp169 (attractive charge), Asp169, Tyr166 (C-H bond), Val161 (pi-sigma), Val161, Leu167 (alkyl) | -6.5 | Thr168, Tyr226, Thr20, Asn22 (H-bond), Trp295 (pi-cation), Pro29, Arg171, Tyr21 (alkyl, pi-alkyl), Asn22 (pi-donor H-bond) | -7.7 | Asn22 (H-bond), Tyr226 (pi-alkyl), Arg294 (unfavorable positive-positive) | - | - |
| 2Y7L (Als9-2) | -8.1 | Thr293 (C-H bond), Trp294 (pi-cation), Tyr21, Pro160, Val161 (pi-alkyl) | -6.4 | Thr168 (H-bond), Arg171, Val22, Pro160, Val161, Ile167, Tyr23, Phe225, Pro29 (alkyl, pi-alkyl) | -7.1 | Glu86, Ser210, Asn213 (H-bond), Asn211 (C-H bond), Tyr261 (pi-alkyl) | - | - |
(-) not identified / no co-crystallized ligand attached.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



