
Submitted:
17 April 2024
Posted:
18 April 2024
You are already at the latest version
Abstract
Enterococcus faecalis is a growing cause of nosocomial and antibiotic-resistant infections. Treating drug-resistant E. faecalis requires novel approaches. The use of bacteriophages (phages) against multidrug-resistant (MDR) bacteria has recently garnered global attention. Biofilms play a vital role in E. faecalis pathogenesis as they enhance antibiotic resistance. Phages eliminate biofilms by producing lytic enzymes, including depolymerases. In this study, Enterococcus phage vB_Efs8_KEN04 (ФKEN04), isolated from a sewage treatment plant in Nairobi, Kenya, was tested against clinical strains of MDR Enterococcus faecalis. This phage had a broad host range against 100% (26/26) of MDR E. faecalis clinical isolates and cross-species activity against Enterococcus faecium. It was able to withstand acidic and alkaline conditions, from pH 3 to 11, as well as temperatures between -80ᵒC and 37ᵒC. It could inhibit and disrupt the biofilms of MDR E. faecalis. Its linear double-stranded DNA genome of 142,402 bp contains 238 coding sequences with a G+C content and coding gene density of 36.01% and 91.46%, respectively. Genomic analyses showed that ФKEN04 belongs to the genus Kochikohdavirus in the family Herelleviridae. It lacked antimicrobial resistance, virulence, and lysogeny genes, and its stability, broad host range, and cross-species lysis indicate strong potential for the treatment of Enterococcus infections.
Keywords:
bacteriophage
; biofilm
; Enterococcus faecalis
; genome
; multidrug resistance
; phage therapy
1. Introduction
Enterococcus faecalis is a gram-positive, facultative anaerobic cocci that causes difficult-to-treat infections in the nosocomial setting [1]. It is commonly found in nature and is a part of the human intestinal microbiota, comprising less than 1% of the microbiome [2,3]. Early in its evolution, Enterococcus faecalis acquired traits that enabled it to become an effective nosocomial pathogen, resistant to several drugs and causing severe infections in humans. It causes many human infections, including bacteremia, soft tissue and wound infections, pneumonia, endocarditis, and urinary tract infections [4,5,6]. It can persist for extended periods on medical equipment, and because of its high tolerance and genetic adaptability, E. faecalis is a significant contaminant in the hospital environment [7]. The ability of Enterococcus faecalis to form biofilms is particularly concerning in clinical settings, as its biofilms form on medical equipment such as catheters and prosthetic heart valves, leading to persistent infections that exhibit increased resistance to antibiotics within the biofilm structure [8]. Biofilms are organized communities of microorganisms that attach to surfaces and are embedded in self-produced extracellular polymeric substances (EPS) consisting of proteins, extracellular DNA, and polysaccharides [9]. Bacterial biofilms enhance pathogenicity; for example, they contribute significantly to persistent chronic urinary tract infections (UTIs), including recurrences and relapses [10]. Existing antibiotics have limited efficacy in eliminating biofilms and are less effective in treating the growing number of MDR infections [11], prompting the exploration of phage-based therapies as promising alternatives for eradicating biofilms and treating MDR pathogens. The dramatic increase in the frequency of antibiotic therapy failures due to resistance has prompted scientists to search for novel solutions.
Bacteriophages, viruses that infect bacteria, have been investigated for the development of highly effective antimicrobials with low toxicity and minor environmental impact. Bacteriophages, known for their narrow host range, are the most represented biological entities on Earth, and their number in ecosystems is estimated to exceed 1031 [12]. Phages can eliminate biofilms by producing enzymes that prevent biofilm formation and disrupt existing biofilms [13]. Depolymerases and lysins are bacteriophage enzymes that selectively degrade biofilms’ extracellular polymeric substance matrix components, enhancing the phages’ access to bacterial biofilm [14].
Phages with narrow host ranges are highly specific for specific bacterial strains or species. This specificity can be advantageous when precise targeting is needed, such as treating specific bacterial infections [15]. However, this makes them less valuable when targeting a wide range of bacteria, such as when treating polymicrobial infections or during the emergence of phage resistance [16]. In this regard, a phage with a broad host range is particularly advantageous because it can target more than one bacterial strain, presumably leading to fewer treatment failures [15].
Many Enterococcus faecalis phages have been identified to date [17,18,19,20] and have been shown to inhibit and disrupt the biofilms of their host bacteria [21,22,23]. For instance, studies have shown the ability of phage EFDG1 to reduce two-week-old biofilms of E. faecalis V583 [24]. Additionally, a genetically engineered orthocluster VIII phage phiEf11 reduced the established biofilm of E. faecalis strains JH2-2 and V583, which had formed on coverslips [25]. After 24 and 48 h of incubation, a significant decrease of 10–100-fold in viable cells was observed [26]. Despite the promising nature of phage therapy in the fight against antimicrobial-resistant bacteria, a few infrequent case studies have identified certain limitations. As an illustration, a patient suffering from a Pseudomonas aeruginosa multidrug-resistant prosthetic vascular graft infection was treated using a cocktail of phages (PT07, 14/01, and PNM) in combination with ceftazidime-avibactam. The outcome, nonetheless, did not meet expectations. After phage treatment and without antimicrobial therapy, a new bloodstream infection, increased biofilm production, and the emergence of phage-resistant mutants in the bacterial isolate occurred, highlighting the challenges and potential risks associated with phage therapy in complex infections [27].
This study presents the genomic characterization and antibiofilm activity of Enterococcus faecalis phage vB_Efs8_KEN04 (ФKEN04), isolated from community wastewater in Nairobi, Kenya. This phage exhibits a relatively broad host range against clinical MDR E. faecalis isolates and a potent capacity to disrupt (eliminate already formed) and inhibit (prevent biofilm initiation) E. faecalis biofilms under laboratory conditions. It also evaluates the stability of lytic ФKEN04 in vitro under different temperatures and pH ranges. The discovery of ФKEN04 offers a promising phage-based therapy to effectively combat multidrug-resistant enterococcal infections and their biofilms.
2. Materials and Methods
2.1. Strains and Cultural Conditions
Archived clinical isolates of multidrug-resistant Enterococcus faecalis and Enterococcus faecium (37 in total, 26 MDR E. faecalis and 11 MDR E. faecium) from patients in different hospitals around Kenya were obtained from an ongoing surveillance study (protocol WRAIR 209/KEMRI 2767) in the Department of Emerging Infectious Diseases, Walter Reed Army Institute of Research-Africa (WRAIR-A) in Kenya. Bacterial identity and antimicrobial susceptibility testing profiles were first confirmed using the Vitek 2 version 9.02 automated platform (bioMérieux, Marcy-l'Étoile, France), and they were cultured in tryptic soy broth (Oxoid Ltd., Basingstoke, Hampshire, England) under aerobic conditions with agitation at 37°C and a speed of 200 rpm for phage isolation.
2.2. Bacteriophage Isolation, Purification, and Propagation
Raw sewage water samples were collected from a sewage treatment plant in Nairobi East. It treats domestic and industrial wastewater, handling approximately 80% of the wastewater generated in Nairobi city daily. It is, therefore, a significant source of bacteria in the environment [28]. Enterococcus phage vB_Efs8_KEN04 was isolated through an enrichment method using E. faecalis isolate EFS8 as a host according to the method described by D’Souza et al. with slight modifications (D’Souza et al., 2020). Briefly, 50 mL of environmental wastewater was centrifuged at 12,000 × g for 10 min (Thermo Fisher Scientific, Waltham, Massachusetts, United States). Eight milliliters of water sterilized by filtration through a 0.22 µm membrane was mixed with 2 ml of 5x Tryptic Soy Broth (TSB; Oxoid Ltd., Basingstoke, Hampshire, England) and 50 µL of bacterial culture grown in 1x TSB for 16-24 h at 37°C with agitation at 200 rpm. The mixture was incubated for 24 h at 37°C with agitation at 200 rpm. Bacterial debris was eliminated by centrifugation, and the supernatant was filter-sterilized. Serial 10-fold dilutions of ФKEN04 in sodium chloride-magnesium sulfate (SM) buffer were spotted onto double-layer (0.7% top/1.5% bottom) Tryptic Soy Agar (TSA) agar overlaid with 100 µL of a culture of E. faecalis isolate EFS8 in the semisolid top layer. The next day, a well-isolated phage plaque was suspended in SM buffer and filter sterilized. Phage KEN04 was purified by three rounds of single-plaque isolation through plaque assays and propagated to reach a high titer. Briefly, the following components were mixed in a 50 ml falcon tube to amplify the phage: 20 ml of TSB, 10 µL of 1 M CaCl2, 40 µL of 1M MgCl2, 200 µL of 10% glucose, and 400 µL of the overnight host bacteria and incubated at 37ᵒC with shaking at 200 rpm for 1−2 h to reach the mid-log phase. When the host bacterium reached the exponential growth phase, 250 µL of a single pure phage suspension was added, followed by incubation at 37ᵒC, 200 rpm until lysis occurred. Bacterial debris was removed by a 10-minute centrifugation at 12,000 x g, after which the supernatant was filtered through a sterile 0.22 µm filter. The filtrate was centrifuged for 16−18h at 10,000 × g to pellet the phages. The supernatant was discarded after ultracentrifugation, leaving approximately 2 ml of supernatant to resuspend the pellet. The phage titers were determined using a spot assay [29].
2.3. Phage Stability
2.3.1. Thermal Stability
Thermal stability was determined by dispensing 20 µL of the propagated phage suspension with a titer of 2 × 109 PFU/ml into 0.2 ml PCR tubes and incubating at different temperatures (−80ᵒC, −20°C, +4°C, 20°C, 22−30ᵒC, 37°C, 40°C, 45°C, 50°C, and 60°C) for 1 h. After incubation, the phage lysate was diluted in SM buffer using a 10-fold serial dilution technique in 96-well round-bottom (U) microplates (Thermo Scientific, Danmark), and the phage titer was then evaluated using a spot assay as described elsewhere [29]. The experiment was performed in triplicate, and the phage lysate stored at +4ᵒC was used as the reference titer.
2.3.2. pH Stability
The effect of pH 1, 3, 5, 7, 9, 11, and 13 on phage titer and viability was studied for 1 h in TSA plates using the spot test method, as described elsewhere [29]. The pH of the SM buffer was adjusted to the desired value using 1M NaOH and 1M HCl. The pH of SM was determined using a pH meter (Thermo Scientific, Danmark). After incubation, the phage titer was evaluated. The experiment was performed in triplicate, and the phage lysate stored at pH 7.5 was used as the reference titer.
2.4. Host Range Analysis
To investigate the activity of ФKEN04 against other endemic bacterial strains, its host range was determined using a spot test [29] against a panel of 27 clinical isolates of MDR enterococci, and the efficiency of plating (EOP) was determined using a double-layer agar plate method, following a previously described protocol [30,31]. The bacterial strains used for this study were associated with skin and soft tissue infection, urinary tract infection, surgical site infection, and blood infection and were widely spread across Kenyan regions. All the tested strains were cultured in broth overnight at 37ᵒC. Briefly, 2 µL of an individual phage stock was spotted on a TSA plate with a lawn of 100 µL of host bacteria cultured overnight in soft agar, which was examined for bacterial lysis after 18–24 h. Host range tests were performed in duplicates. A phage was termed ‘potent’ upon infecting and lysing bacterial strain in the host range panel [32]. The EOP was calculated by dividing the average plaque-forming units (PFU) of the test bacteria by the average PFU of the host bacteria. Phages were categorized as high production (EOP ≥ 0.5) when the productive infection on the test bacteria resulted in at least 50% of the PFU found for the primary host; medium production (0.1 ≤ EOP < 0.5); low production efficiency (0.001 <EOP< 0.1); inefficient (EOP ≤ 0.001), and reference (EOP = 1) [33,34].
2.5. Biofilm Formation Assay
The ability of enterococci to form biofilms was assessed using a crystal violet biomass assay [35]. Briefly, the bacterial isolates were grown overnight at 37ᵒC, 200 rpm in tryptic soy broth. The enterococcal cultures were diluted 1:100 in fresh TSB containing 2% glucose monohydrate (Oxoid Ltd., Basingstoke, Hampshire, England), and 100 µL of the diluted solution was dispensed into the wells of 96-well round-bottom (U) microplates (Thermo Scientific, Danmark) and incubated under static conditions at 37ᵒC, 5% CO2 for 72 h without changing the medium. Wells with sterile TSB containing 2% glucose were used as controls for contamination. Enterococcus faecalis strain ATCC 29212 was used as a positive biofilm control, whereas Enterococcus faecalis isolate EFS4 (ST947), an in-house isolate, was used as the negative control. The experiments were performed in triplicate. After incubation, planktonic bacteria were pipetted off, the wells were washed three times with distilled water, and the plates were allowed to air-dry for 15 min. Adhesive bacteria were fixed at 60ᵒC for 1 h and stained with 100 µL of 1% crystal violet for 20 min. This was followed by three washes with 100 µL of sterile distilled water to remove the excess dye. The microplates were air-dried for 15 min. Then, 100 µL of 33% glacial acetic acid was added to each well, followed by pipetting to release the bound crystal violet dye from the biofilm [36]. The stained adherent cells’ optical density (OD) was quantified at 630 nm using a microtiter plate reader (BioTek Instruments, Gen5TM version 3.10, USA). The strains were divided into groups based on the OD values of the bacterial biofilms. Bacterial strains were classified as follows: OD values ≤0.0551 as non-biofilm formers, weak biofilm-producing isolates (0.0551<OD<0.102), moderate biofilm formers (0.102<OD<0.204), and those with OD > 0.204 were classified as strong biofilm-producing bacterial strains [37,38].
2.6. Biofilm Inhibition Assay by Phage
The anti-biofilm effect of the phage was evaluated as described by Goodarzi et al., with some modifications [39]. To investigate the inhibitory effect of phages on biofilm formation, a 3-day-old biofilm was formed in the presence of phages. Briefly, single colonies of E. faecalis strains were cultured in TSB at 37°C, 200 rpm for 24 h. After incubation, the bacterial culture was diluted 1:100 in fresh TSB, supplemented with 2% glucose. Diluted bacterial culture (100 µL) was dispensed into the wells, and 2 µL of phage lysate (titer 9 × 109 PFU/ml) was added. The plates were incubated under static conditions at 37ᵒC, 5% CO2 for 72 h without changing the medium. Wells with sterile TSB containing 2% glucose were used as controls for contamination. Enterococcus faecalis strain ATCC 29212 was used as a positive biofilm control, whereas Enterococcus faecalis isolate EFS4 (ST947) was used as the negative control. Biofilm formation was performed in triplicate for treated and untreated samples. After incubation for 72 h, the suspension was drained from the wells and rinsed with sterile distilled water three times, and biofilm fixation, staining, and OD measurements were performed as described in Section 2.5.
2.7. Biofilm Disruption Assay by Phage
A 2-day-old biofilm was formed in the absence of phages and then treated. Briefly, single colonies of E. faecalis strains were cultured in TSB at 37°C, 200 rpm for 24 h. After incubation, the bacterial culture was diluted 1:100 in fresh TSB, supplemented with 2% glucose. The diluted bacterial culture (100 µL) was dispensed into the wells, followed by incubation under static conditions at 37ᵒC, 5% CO2 for 48 h without changing the media. After 48 h of incubation, the plates were removed from the incubator, the planktonic bacteria were pipetted off, the wells were washed twice to remove all planktonic cells, and 100 µL of the phage lysate was added. For untreated wells, the medium was replaced with TSB supplemented with 2% glucose. The plates were then placed back in the incubator for 24 h. Biofilm formation was performed in triplicate for treated and untreated samples. After 24 h of treatment, the suspension was drained from the wells and rinsed three times with sterile distilled water. Biofilm fixation, staining, and OD measurements were performed as described in Section 2.5.
2.8. Genomic DNA Extraction
Before DNA extraction, the pure phage suspension (2 × 109 PFU/ml) was propagated to reach a titer of 3.5 x 1011 PFU/ml. The phage was concentrated by centrifugation for 18 h at 10,000 x g. The supernatant was then discarded. One milliliter of the propagated phage suspension was treated with RNase A (Thermo Fisher Scientific, USA) and DNase I (ThermoFisher Scientific, USA) to remove host RNA and DNA, respectively. Deproteinization was achieved by adding Proteinase K and incubating at 56°C for 1 h 30 min [40]. Phage DNA was isolated and purified using the Norgen phage DNA isolation kit (Norgen Biotek Corporation, Thorold, ON, Canada), following the manufacturer’s instructions. The quality and quantity of the extracted DNA were determined using a Nanodrop One spectrophotometer and a Qubit4 fluorometer (Fisher Scientific, Waltham, MA, USA), respectively.
2.9. Genome Sequencing and Bioinformatic Analysis of Sequencing Data
The genome was sequenced on the Illumina MiSeq platform (Illumina, Inc., USA). The quality of the raw reads was assessed using FastQC v0.12.1 [41], trimmed with fastp v0.23.4 [42], and assembled using shovill v1.1.0 (https://github.com/tseemann/shovill). Genome annotation was performed using pharokka 1.5.1 [43]. The complete phage genome was further queried against CRISPR-Cas Finder (https://proksee.ca/, accessed on December 11, 2023), PhageLead [44] (https://phageleads.dk/, accessed on 11 December 2023), PhageTerm platforms [45] (https://cpt.tamu.edu/galaxy-pub, accessed on 27 November 2023) to determine CRISPR-like systems, lysogeny genes, and termini in the phage genome, respectively. ARAGORN v1.2.41 [46] and tRNAscan-SE v2.0.12 [47] were used to predict the tRNA and tmRNA genes. Nucleic acid sequence similarity searches were performed using default parameters in BLASTn [48]. The identification of antimicrobial resistance and virulence genes was conducted by scanning the assembled nucleotide sequence using Abricate version 1.0.1 [49] at https://github.com/tseemann/abricate, accessed on November 27, 2023, with the following datasets: NCBI AMRFinderPlus [50], Comprehensive Antibiotic Resistance Database (CARD) [51], Virulence FactorDatabase (VFDB) [52] and ResFinder [53].
2.10. Phylogenetic Tree and Comparative Genomics of Phage Genomes
To examine the genetic relationships between ФKEN04 and other Enterococcus phages, a phylogenetic tree was generated using the entire genome sequences of 38 phages retrieved from the NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 23 December 2023), including ФKEN04. The phages included in the phylogenetic tree were selected according to the following criteria: i) should be a complete genome sequence [54]; ii) should exhibit a high similarity of >70% to ФKEN04 [55]; and iii) should have a genome size similar to that of ФKEN04 [56]. The analysis was conducted using the Virus Classification and Tree Building Online Resource (VICTOR), a method for the genome-based phylogeny and classification of prokaryotic viruses [54] (https://victor.dsmz.de, accessed on 23 December 2023), enabling the evaluation of similarities and differences in genetic characteristics. The nucleotide sequences were compared using the genome explosion distance phylogeny (GBDP) method in the settings recommended for prokaryotic viruses [57], and the branch length was magnified using the distance formula d0, according to GBDP. In addition, the intergenomic similarities between ФKEN04 and the 20 closest related Enterococcus phages were determined using a virus intergenomic distance calculator (VIRIDIC) [58] to further our understanding of their interactions. The nucleotide identity of the complete genome length cut-off for genera (>70%) and species (>95%) was used [55].
2.11. Statistical Analysis
Statistical analysis of the biofilm results was conducted using GraphPad Prism 8.4.0 (GraphPad Software, Inc., San Diego, CA, USA), and a Student’s t-test was employed to determine significance. Statistical significance was set at p < 0.05.
3. Results
3.1. Bacteriophage Isolation and Purification
Phage vB_Efs8_KEN04 isolated from a wastewater plant using the Enterococcus faecalis isolate EFS8 as the host strain. ФKEN04 forms clear plaques on a double-layered agar plate, as shown in Figure 1.
3.2. Host Range Analysis
The host range and efficiency of plating studies were conducted on 37 multidrug-resistant (MDR) Enterococci isolates, with 26 being E. faecalis isolates and the remaining eleven being E. faecium isolates. These bacteria were isolated from urinary tract infections, skin and soft tissue infections, surgical site infections, and blood infections in humans. Enterococcus faecalis phage vB_Efs8_KEN04 was active against all 26 MDR E. faecalis and 1/11 of the E. faecium in the spot assay (Table 1). The EOP was greater than 0.5 for 13/27 (48.15%) isolates, indicating high production of the phage and an EOP≥0.5 is considered good for therapy [34].
3.3. Phage Stability
The stability of vB_Efs8_KEN04 was evaluated at different temperatures and pH values. The results revealed that ФKEN04 was stable from − 80°C to 37°C (Figure 2A, Table S1). Phage titer declined at temperatures of 40°C and above. Similarly, the stability rate of ФKEN04 was high at pH 5–11 (slightly acidic to strongly basic) but low at pH 3 (strongly acidic). No phage activity was observed at pH 1 or 13 (Figure 2B, Table S2). These findings suggest that phage vB_Efs8_KEN04 can withstand moderate acidic and alkaline conditions and a wide range of temperature conditions between − 80°C and 37°C.
3.4. Biofilm Formation of Enterococcus faecalis
Of the 26 clinical MDR E. faecalis isolates that were examined for biofilm formation, 22 isolates (84.62%) were strong biofilm formers, one isolate (3.85%) was identified as a weak biofilm former, one (3.85%) isolate as a moderate biofilm former, and two isolates (7.69%) as non-biofilm formers (Table S3). A total of 24 MDR E. faecalis isolates (92.31%) showed the ability to produce biofilms. Enterococcus faecalis isolate EFS8, the phage host, was a strong biofilm former (Figure 3).
3.5. Biofilm Inhibition and Disruption by Phage vB_Efs8_KEN04.
The effects of phage vB_Efs8_KEN04 treatment on the inhibition and disruption of biofilms of multidrug-resistant E. faecalis isolates are shown in Figure 4. For the inhibition of biofilm formation (Figure 4A, Table S4), phage vB_Efs8_KEN04 treatment for 72 h at 37ᵒC reduced the bacterial population significantly (**, p <0.05) for some susceptible bacteria. However, it could not prevent other bacteria (EFS5, EFS6, EFS15, EFS17, EFS21, EFS22, EFS26, EFS27, EFS30, EFS31, EFS31, EFS32, and EFS33) from forming biofilms (p>0.05, highlighted by ns). For the disruption of already formed biofilms (Figure 4B, Table S5), phage vB_Efs8_KEN04 treatment for 24 h significantly reduced the bacterial population (**, p <0.05) for all the bacteria, including its host bacteria EFS8, except for the isolate EFS18 (ns, p>0.05).
3.6. Genome Characteristics of Enterococcus faecalis Phage vB_Efs8_KEN04
The genome structure of phage vB_Efs8_KEN04, a newly isolated E. faecalis phage in Kenya, was investigated in this study. The phage genome contained 8 tRNA genes and was shown to be a linear double-stranded DNA with a length of 142,402 base pairs and a G+C content of 36.01% (Figure 5). It belongs to the genus Kochikohdavirus of the family Herelleviridae. The genome contained 238 coding sequences (CDS) with a coding gene density of 91.46%. Seventy CDSs (29.41%) were predicted to encode functional proteins, and the remaining 168 (70.59%) were annotated as hypothetical proteins (Table S6). The functional proteins were divided into the following categories:
- i)
- DNA replication, transcription, translation, and nucleotide metabolism: A total of 25 CDSs were predicted to encode for DNA replication, transcription regulation, translation, and metabolism-related proteins, such as HNH homing endonuclease, DNA helicase, exonucleases, transcriptional repressor, DNA helicase, DNA primase, and a transcriptional regulator, RNA polymerase beta subunit, and thymidylate synthase.
- ii)
- Structural and packaging proteins: 27 CDS were predicted to encode for tail, head, and packaging proteins such as portal proteins, head proteins, tail fiber proteins, head maturation proteases, virion structural proteins, tail proteins, tail assembly chaperones, minor and major head proteins, and terminase large and small subunits.
- iii)
- Host lysis and adhesion proteins: Two CDS were predicted to encode holin and endolysin proteins. BLASTP analysis of the Enterococcus phage vB_Efs8_KEN04 genome revealed no similarities to the genes encoding integrase or excisionase. The genome of phage vB_Efs8_KEN04 lacks genes encoding toxins, virulence factors, antibiotic resistance genes, and CRISPR. These data indicate that phage vB_Efs8_KEN04 is a strictly lytic phage that can be used to treat E. faecalis infection.
- iv)
- Sixteen CDS encode for moron, auxiliary metabolic genes, and host takeover.
Analysis of phage vB_Efs8_KEN04 DNA termini and phage packaging mechanisms revealed long Direct Terminal Repeats (DTR) of 2,849 bp with a specific packaging site called the cos site, which serves as a recognition signal for the packaging machinery. This DTR is comparable to that of bacteriophage T5 [59].
3.7. Phylogenetic Analysis
To gain a deeper understanding of the evolution and relationship between phage vB_Efs8_KEN04 and other Enterococcus phages, the genome of phage vB_Efs8_KEN04 was compared with that of 37 Enterococcus phages. These phage sequences were obtained from the National Center for Biotechnology Information (NCBI) database, and all had homology ranging from 78.06 to 99.29% with phage vB_Efs8_KEN04 (Table S7).
The phylogenetic tree, generated with the whole genome sequences, indicated that phage vB_Efs8_KEN04 had the highest similarity to Enterococcus phage PBEF129 (GenBank accession number MN854830.2), Enterococcus phage phiM1EF22 (GenBank accession number AP018715.1), Enterococcus phage ECP3 (GenBank accession number NC_027335.2), and Enterococcus phage vB_EfaM_Ef2.3 (GenBank accession number MK721192.1) (Figure 6A, Table S7). Subsequently, we employed VIRIDIC to compute the inter-genomic similarities, revealing the degree of similarity between phage vB_Efs8_KEN04 and the top 20 phages most closely linked to it (Figure 6B). This indicated that the similarity of phage vB_Efs8_KEN04 and the other Enterococcus phages' complete genome was significantly greater than the genus threshold of 70% and lower than the species threshold of 95% [55], suggesting that they belong to the same genus but distinct species. The intergenomic similarities between phage vB_Efs8_KEN04 and the four most closely related Enterococcus phages were as follows: Enterococcus phage PBEF129 (94.6%), Enterococcus phage phiM1EF22 (93.3%), Enterococcus phage ECP3 (94.5%), and Enterococcus phage vB_EfaM_Ef2.3 (93.5%) (Figure 6B).
4. Discussion
The use of bacteriophages, also known as phages, in treating bacterial infections, including E. faecalis, has gained particular attention. This is due to the increase in antibiotic resistance and the phages’ ability to infect and eliminate bacteria. Phages have the potential to serve as a natural, safe, and efficient method for preventing and controlling multidrug-resistant (MDR) organisms [60]. Studies have also shown that phages can be used to control biofilms formed by Enterococcus faecalis [23,61].
This study characterized a highly lytic and broad-spectrum E. faecalis phage vB_Efs8_KEN04 isolated from environmental wastewater in Nairobi, Kenya. It is a dsDNA phage belonging to group I of Baltimore’s classification of viruses. The genome of this phage did not encode any lysogenic, antibiotic resistance, or virulence and CRISPR-cas genes. Thus, it is an effective and safe candidate for phage therapy. Based on genome annotation, phage vB_Efs8_KEN04 was classified as a member of the genus Kochikohdavirus in the family Herelleviridae. Its genome contains eight genes encoding for transfer RNAs (tRNAs). The presence of tRNAs in bacteriophage genomes is widespread, especially among virulent phages [62]. However, their precise role has remained ambiguous for almost five decades as phages utilize the host’s transcriptional machinery to control the expression of their own genes after initial infection [63]. Several hypotheses have been proposed for the role of these phage-encoded tRNAs. The most established is codon compensation, where codons rarely used by the host but necessary to the phage are supplemented by the tRNAs encoded by the phage [62]. Recently, a study proposed a new hypothesis that phage-encoded tRNAs counteract the tRNA-depleting strategies of the host using enzymes such as VapC, PrrC, Colicin D, and Colicin E5 to defend from viral infection, and they have evolved to be insensitive to host anticodon nucleases [64].
Phage vB_Efs8_KEN04 exhibited a wide host range by displaying lysis activity against all 26 clinical MDR E. faecalis isolates tested. These isolates belonged to various sequence types (ST6, ST44, ST28, ST59, ST368, ST947, ST1903, and ST1904) and were associated with different infection types such as urinary tract infection, skin and soft tissue infection, surgical site infection, and blood infection. It also has an EOP≥0.5 for 13 isolates, indicating high progeny production. In addition, it showed a cross-species activity against a clinical isolate of multidrug-resistant Enterococcus faecium. The phage’s ability to target its host bacteria is due to its host receptors involved in recognition, interaction and adsorption during phage attachment [65]. Additionally, the receptors are recognized by the ends of the virion’s long tail fibers of the phage towards the host bacteria [66]. When subjected to different temperatures and pH conditions, phage vB_Efs8_KEN04 showed the ability to withstand moderate acidic and alkaline conditions from pH 3-11 and a wide temperature range from – 80°C to 37°C. Many external physical and chemical factors, including but not limited to temperature, acidity, salinity, and ions, determine bacteriophages occurrence, viability, and storage. These factors can inactivate the phage by damaging its structural components (head, tail, envelope), lipid depletion, and/or DNA structural changes [67]. The studied phage exhibited remarkable stability throughout a broad range of temperatures and pH levels, making it advantageous for formulation into a suitable pharmaceutical form and therapeutic applications. Furthermore, the phage's stability across acidic and alkaline environments (pH 3–11) enables it to be administered orally without compromising its viability in the gastrointestinal tract [23].
In this study, we also investigated the effect of phage vB_Efs8_KEN04 on the biomass reduction of E. faecalis biofilm by inhibition and disruption experiments. Biofilms are communities of bacteria that can be highly resistant to antibiotics and contribute to persistent infections [24,68]. Several factors contribute to the enhanced antimicrobial resistance of microorganisms in a biofilm. These include the physical barrier created by the extracellular matrix, which hinders the diffusion of antimicrobial agents [69]. Additionally, nutrient and oxygen depletion within the biofilm can cause certain bacteria to enter a stationary state, making them less susceptible to microbial killing [70].
Furthermore, a subpopulation of bacteria might differentiate into a phenotypically resistant state, and some bacteria within the biofilm have been found to express specific antimicrobial resistance genes unique to biofilms [71]. Recent studies have demonstrated that extracellular DNA (eDNA) in the biofilm matrix protects microbial cells against various antimicrobial agents [72]. The biofilm formation phenotype of multidrug-resistant Enterococcus faecalis was investigated, and the results revealed that out of 26 MDR tested for this purpose, 92.31% showed the ability to form biofilm. To date, several investigations have been performed to test bacteriophages’ ability to inhibit and destroy Enterococcus faecalis biofilms [17,22,73,74], but the mechanisms of phage-biofilm interaction are not well understood [75]. As indicated by the crystal violet biomass assay, the inhibition assay indicated that the isolated phage vB_Efs8_KEN04 significantly reduced biofilm biomass (p-value <0.05) compared to the control for most of the biofilm-forming E. faecalis isolates. Based on genome and structural proteome analysis, this can be explained by the endolysins encoded by phage vB_Efs8_KEN04, which may destroy the EPS component. Furthermore, phage endolysin lyses some bacteria at the edge of the EPSs [76]. Phage vB_Efs8_KEN04 then induces the production of depolymerase, such as endolysins [77] (CDS121) (Table S6) since they can penetrate the inner layers of the biofilm by degrading structural components of the established biofilm exopolymeric matrix, allowing them to break it down or disrupt its integrity [78]. The reduction of bacteria on the biofilm causes the reduction of EPS material; thus, the biofilm is completely eliminated [79]. Similarly, phage vB_Efs8_KEN04 showed the ability to significantly decrease biofilm biomass when compared with an untreated biofilm of an already formed biofilm. Therefore, phage vB_Efs8_KEN04 has the potential to be successfully used as a biofilm eradication agent. In future research, the phage-derived enzymes may be studied as biological antibacterial agents to control Enterococcus and its biofilm.
5. Conclusions
Phage vB_Efs8_KEN04 is a lytic phage belonging to the genus Kochikohdavirus in the family Herelleviridae. It was isolated from a municipal sewage treatment plant located in Nairobi East. Phage vB_Efs8_KEN04 exhibits efficacy against all the clinical multidrug-resistant strains of Enterococcus faecalis tested and one E. faecium isolates, including the ability to destroy bacterial biofilms. The genome analysis revealed that the phage lacks genes of concern, including virulence, antibiotic resistance, lysogeny genes, and the CRISPR or CRISPR-like system. The phage vB_Efs8_KEN04 has great potential as a candidate for phage therapy against enterococci infections and for controlling biofilms.
6. Limitations
A limitation of this study is the lack of animal models as representative models of enterococci infection in humans, as the experimental models can mimic the pathogenesis of natural disease.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Temperature stability test (Mean ± SD); Table S2: pH stability test (Mean ± SD); Table S3: Biofilm formation profile of MDR Enterococcus faecalis isolates; Table S4: Biofilm inhibition of phage vB_Efs8_KEN04; Table S5: Biofilm disruption of phage vB_Efs8_KEN04; Table S6: Predicted molecular function for gene products of phage vB_Efs8_KEN04; Table S7: Summary of similar genomic sequence with phage vB_Efs8_KEN04.
Author Contributions
Conceptualization, O.S., A.N. and L.M.; methodology, O.S.; software, O.S. and C.K.; validation, C.K., A.N. and L.M.; formal analysis, O.S.; investigation, O.S.; resources, O.S., C.K., A.N. and L.M.; data curation, O.S.; writing—original draft preparation, O.S.; writing—review and editing, C.K., A.N. and L.M.; visualization, O.S.; supervision, A.N. and L.M.; project administration, O.S.; funding acquisition, O.S., A.N. and L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the African Union Commission, grant number PAU/ADM/PAUSTI/9/2022, and the AFRICA-ai-JAPAN Project Innovation Research Funds, grant number JKU/ADM/10B.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material. The raw data are available in Sequence Read Archive under the bioSample accession number SAMN40604471 and the complete genome of Enterococcus phage vB_Efs8_KEN04 is available in GenBank under the accession number PP582180.
Acknowledgments
We acknowledge the Africa CDC for the sequencing support. We appreciate the project manager, Erick Odoyo, and Allan Wataka for administrative support and the lab personnel Martin Georges and Moses Gachoya for their help in environmental samples collection. James Muturi and Vanessa Onyoni are also appreciated for their support in phage genome sequencing.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Said, M.S.; Tirthani, E.; Lesho, E. Enterococcus Infections. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2023. [Google Scholar]
- Raza, T.; Ullah, S.R.; Mehmood, K.; Andleeb, S. Vancomycin Resistant Enterococci: A Brief Review. JPMA J. Pak. Med. Assoc. 2018, 68, 768–772. [Google Scholar]
- Franz, C.M.A.P.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as Probiotics and Their Implications in Food Safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Beganovic, M.; Luther, M.K.; Rice, L.B.; Arias, C.A.; Rybak, M.J.; LaPlante, K.L. A Review of Combination Antimicrobial Therapy for Enterococcus Faecalis Bloodstream Infections and Infective Endocarditis. Clin. Infect. Dis. 2018, 67, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Rajkumari, N.; Mathur, P.; Misra, M.C. Soft Tissue and Wound Infections Due to Enterococcus Spp. Among Hospitalized Trauma Patients in a Developing Country. J. Glob. Infect. Dis. 2014, 6, 189–193. [Google Scholar] [CrossRef]
- Shiadeh, S.M.J.; Pormohammad, A.; Hashemi, A.; Lak, P. Global Prevalence of Antibiotic Resistance in Blood-Isolated Enterococcus Faecalis and Enterococcus Faecium: A Systematic Review and Meta-Analysis. Infect. Drug Resist. 2019, 12, 2713–2725. [Google Scholar] [CrossRef]
- Tagliaferri, T.L.; Jansen, M.; Horz, H.-P. Fighting Pathogenic Bacteria on Two Fronts: Phages and Antibiotics as Combined Strategy. Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Dale, J.L.; Nilson, J.L.; Barnes, A.M.T.; Dunny, G.M. Restructuring of Enterococcus Faecalis Biofilm Architecture in Response to Antibiotic-Induced Stress. Npj Biofilms Microbiomes 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Blackledge, M.S.; Worthington, R.J.; Melander, C. Biologically Inspired Strategies for Combating Bacterial Biofilms. Curr. Opin. Pharmacol. 2013, 13, 699–706. [Google Scholar] [CrossRef]
- Chaudhary, N.; Maurya, R.; Singh, D.; Mohan, B.; Taneja, N. Genome Analysis and Antibiofilm Activity of Phage 590B against Multidrug-Resistant and Extensively Drug-Resistant Uropathogenic Escherichia Coli Isolates, India. Pathogens 2022, 11, 1448. [Google Scholar] [CrossRef]
- Shrestha, L.; Fan, H.-M.; Tao, H.-R.; Huang, J.-D. Recent Strategies to Combat Biofilms Using Antimicrobial Agents and Therapeutic Approaches. Pathogens 2022, 11, 292. [Google Scholar] [CrossRef]
- Brives, C.; Pourraz, J. Phage Therapy as a Potential Solution in the Fight against AMR: Obstacles and Possible Futures. Palgrave Commun. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- Ferriol-González, C.; Domingo-Calap, P. Phages for Biofilm Removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T. Bacteriophages and Their Enzymes in Biofilm Control. Curr. Pharm. Des. 2015, 21, 85–99. [Google Scholar] [CrossRef]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef]
- Principi, N.; Silvestri, E.; Esposito, S. Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections. Front. Pharmacol. 2019, 10, 513. [Google Scholar] [CrossRef]
- Song, M.; Wu, D.; Hu, Y.; Luo, H.; Li, G. Characterization of an Enterococcus Faecalis Bacteriophage vB_EfaM_LG1 and Its Synergistic Effect With Antibiotic. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Di Lallo, G.; Falconi, M.; Iacovelli, F.; Frezza, D.; D’Addabbo, P. Analysis of Four New Enterococcus Faecalis Phages and Modeling of a Hyaluronidase Catalytic Domain from Saphexavirus. PHAGE New Rochelle N 2021, 2, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, J.; Lin, Y.; Yuan, L.; El-Telbany, M.; Maung, A.; Abdelaziz, M.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Isolation, Characterization of Enterococcus Phages and Their Application in Control of E. Faecalis in Milk. J. Appl. Microbiol. 2023, 134. [Google Scholar] [CrossRef]
- Zhang, W.; Mi, Z.; Yin, X.; Fan, H.; An, X.; Zhang, Z.; Chen, J.; Tong, Y. Characterization of Enterococcus Faecalis Phage IME-EF1 and Its Endolysin. PLoS ONE 2013, 8, e80435. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, F.; Hallajzadeh, M.; Sholeh, M.; Talebi, M.; Pirhajati Mahabadi, V.; Amirmozafari, N. Anti-Biofilm Activity of a Lytic Phage Against Vancomycin-Resistant Enterococcus Faecalis. Iran. J. Pathol. 2022, 17, 285–293. [Google Scholar] [CrossRef]
- El-Telbany, M.; Lin, C.-Y.; Abdelaziz, M.; Maung, A.; El-Shibiny, A.; Noor Mohammadi, T.; Zayda, M.; Wang, C.; Lwin, S.; Zhao, J.; et al. Potential Application of Phage vB_EfKS5 to Control Enterococcus Faecalis and Its Biofilm in Food. AMB Express 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- El-Atrees, D.M.; El-Kased, R.F.; Abbas, A.M.; Yassien, M.A. Characterization and Anti-Biofilm Activity of Bacteriophages against Urinary Tract Enterococcus Faecalis Isolates. Sci. Rep. 2022, 12, 13048. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.-A.; Beyth, N.; Hazan, R. Targeting Enterococcus Faecalis Biofilms with Phage Therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef]
- Zhang, H.; Fouts, D.E.; DePew, J.; Stevens, R.H. Genetic Modifications to Temperate Enterococcus Faecalis Phage Ef11 That Abolish the Establishment of Lysogeny and Sensitivity to Repressor, and Increase Host Range and Productivity of Lytic Infection. Microbiol. Read. Engl. 2013, 159, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, J.M.; Buttaro, B.; Zhang, H.; Liss, N.; Sassone, L.; Stevens, R. Effect of a Genetically Engineered Bacteriophage on Enterococcus Faecalis Biofilms. Arch. Oral Biol. 2016, 71, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Blasco, L.; López-Hernández, I.; Rodríguez-Fernández, M.; Pérez-Florido, J.; Casimiro-Soriguer, C.S.; Djebara, S.; Merabishvili, M.; Pirnay, J.-P.; Rodríguez-Baño, J.; Tomás, M.; et al. Case Report: Analysis of Phage Therapy Failure in a Patient with a Pseudomonas Aeruginosa Prosthetic Vascular Graft Infection. Front. Med. 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Song’oro, E.; Nyerere, A.; Magoma, G.; Gunturu, R. Occurrence of Highly Resistant Microorganisms in Ruai Wastewater Treatment Plant and Dandora Dumpsite in Nairobi County, Kenya. Adv. Microbiol. 2019, 9, 479–494. [Google Scholar] [CrossRef]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions. Available online: https://link.springer.com/book/10.1007/978-1-60327-164-6 (accessed on 3 March 2024).
- Chang, Y.; Shin, H.; Lee, J.-H.; Park, C.J.; Paik, S.-Y.; Ryu, S. Isolation and Genome Characterization of the Virulent Staphylococcus Aureus Bacteriophage SA97. Viruses 2015, 7, 5225–5242. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Correction: Isolation of Phages for Phage Therapy: A Comparison of Spot Tests and Efficiency of Plating Analyses for Determination of Host Range and Efficacy. PloS One 2015, 10, e0127606. [Google Scholar] [CrossRef]
- Mutai, I.J.; Juma, A.A.; Inyimili, M.I.; Nyachieo, A.; Nyamache, A.K. Efficacy of Diversely Isolated Lytic Phages against Multi-Drug Resistant Enterobacter Cloacae Isolates in Kenya. Afr. J. Lab. Med. 2022, 11. [Google Scholar] [CrossRef]
- Chaudhary, N.; Mohan, B.; Kaur, H.; Modgil, V.; Kant, V.; Bhatia, A.; Taneja, N. Vibrio Phage VMJ710 Can Prevent and Treat Disease Caused by Pathogenic MDR V. Cholerae O1 in an Infant Mouse Model. Antibiotics 2023, 12, 1046. [Google Scholar] [CrossRef]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Brabban, A.D.; Diez-Gonzalez, F. Isolation and Characterization of Lytic Bacteriophages against Enterohaemorrhagic Escherichia Coli. J. Appl. Microbiol. 2011, 110, 1323–1331. [Google Scholar] [CrossRef]
- Khalil, M.A.; Alorabi, J.A.; Al-Otaibi, L.M.; Ali, S.S.; Elsilk, S.E. Antibiotic Resistance and Biofilm Formation in Enterococcus Spp. Isolated from Urinary Tract Infections. Pathogens 2022, 12, 34. [Google Scholar] [CrossRef]
- Tiria, F.; Odoyo, E.; Georges, M.; Nyerere, A.; Musila, L. Molecular Detection of Key Virulence-Associated Genes and Phenotypic Analysis of Virulence Traits of Klebsiella Pneumoniae Clinical Isolates from Kenya. J. Pure Appl. Microbiol. 2023, 17. [Google Scholar] [CrossRef]
- Donelli, G.; Vuotto, C.; Cardines, R.; Mastrantonio, P. Biofilm-Growing Intestinal Anaerobic Bacteria. FEMS Immunol. Med. Microbiol. 2012, 65, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Mirzaie, A.; Ranjbar, R. Antibiotic Resistance, Virulence-Associated Genes Analysis and Molecular Typing of Klebsiella Pneumoniae Strains Recovered from Clinical Samples. AMB Express 2021, 11, 122. [Google Scholar] [CrossRef]
- Goodarzi, F.; Hallajzadeh, M.; Sholeh, M.; Talebi, M.; Pirhajati Mahabadi, V.; Amirmozafari, N. Anti-Biofilm Activity of a Lytic Phage Against Vancomycin-Resistant Enterococcus Faecalis. Iran. J. Pathol. 2022, 17, 285–293. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.; Filippov, A.A.; Sergueev, K.V.; He, Y.; Ward, A.M.; Goglin, K.; Vashee, S.; Nikolich, M.P.; Fouts, D.E. Complete Genome Sequence of Broad-Host-Range Staphylococcus Aureus Myophage ESa1. Microbiol. Resour. Announc. 2020, 9, e00730–20. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 February 2024).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinforma. Oxf. Engl. 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bouras, G.; Nepal, R.; Houtak, G.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Pharokka: A Fast Scalable Bacteriophage Annotation Tool. Bioinforma. Oxf. Engl. 2023, 39, btac776. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Rajandas, H.; Parimannan, S.; Manickam, R.; Marimuthu, K.; Petersen, B.; Clokie, M.R.J.; Millard, A.; Sicheritz-Pontén, T. PhageLeads: Rapid Assessment of Phage Therapeutic Suitability Using an Ensemble Machine Learning Approach. Viruses 2022, 14, 342. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.R.; Depardieu, F.; Fortier, L.-C.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using next-Generation Sequencing Data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canback, B. ARAGORN, a Program to Detect tRNA Genes and tmRNA Genes in Nucleotide Sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. Clifton NJ 2019, 1962, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antimicrobial and Virulence Genes. Github 2017.
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, 10.1128/aac.00483-19. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and Model-Centric Curation of the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A Comparative Pathogenomic Platform with an Interactive Web Interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-Based Phylogeny and Classification of Prokaryotic Viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef] [PubMed]
- Henz, S.R.; Huson, D.H.; Auch, A.F.; Nieselt-Struwe, K.; Schuster, S.C. Whole-Genome Prokaryotic Phylogeny. Bioinforma. Oxf. Engl. 2005, 21, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinformatics 2013, 14, 60. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC-A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.; Vincent, M.; Sun, Y.; Yu, H.; Wang, J.; Bao, Q.; Kong, H.; Hu, S. Complete Genome Sequence of Bacteriophage T5. Virology 2005, 332, 45–65. [Google Scholar] [CrossRef]
- El Haddad, L.; Harb, C.P.; Gebara, M.A.; Stibich, M.A.; Chemaly, R.F. A Systematic and Critical Review of Bacteriophage Therapy Against Multidrug-Resistant ESKAPE Organisms in Humans. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 69, 167–178. [Google Scholar] [CrossRef]
- Goodarzi, F.; Hallajzadeh, M.; Sholeh, M.; Talebi, M.; Mahabadi, V.P.; Amirmozafari, N. Biological Characteristics and Anti-Biofilm Activity of a Lytic Phage against Vancomycin-Resistant Enterococcus Faecium. Iran. J. Microbiol. 2021, 13, 691–702. [Google Scholar] [CrossRef]
- Bailly-Bechet, M.; Vergassola, M.; Rocha, E. Causes for the Intriguing Presence of tRNAs in Phages. Genome Res. 2007, 17, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ma, Y.; Wang, Y.; Yang, H.; Shen, W.; Chen, X. Transcription Regulation Mechanisms of Bacteriophages: Recent Advances and Future Prospects. Bioengineered 2014, 5, 300–304. [Google Scholar] [CrossRef]
- van den Berg, D.F.; van der Steen, B.A.; Costa, A.R.; Brouns, S.J.J. Phage tRNAs Evade tRNA-Targeting Host Defenses through Anticodon Loop Mutations. eLife 2023, 12, e85183. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host Receptors for Bacteriophage Adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed]
- Drexler, K.; Riede, I.; Montag, D.; Eschbach, M.-L.; Henning, U. Receptor Specificity of the Escherichia Coli T-Even Type Phage Ox2: Mutational Alterations in Host Range Mutants. J. Mol. Biol. 1989, 207, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.-W.; Tremblay, D.; Moineau, S. Long-Term Bacteriophage Preservation. WFCC Newsl. 2004, 38, 35–40. [Google Scholar]
- Sanz, M.; Beighton, D.; Curtis, M.A.; Cury, J.A.; Dige, I.; Dommisch, H.; Ellwood, R.; Giacaman, R.A.; Herrera, D.; Herzberg, M.C.; et al. Role of Microbial Biofilms in the Maintenance of Oral Health and in the Development of Dental Caries and Periodontal Diseases. Consensus Report of Group 1 of the Joint EFP/ORCA Workshop on the Boundaries between Caries and Periodontal Disease. J. Clin. Periodontol. 2017, 44, S5–S11. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Bamford, N.C.; MacPhee, C.E.; Stanley-Wall, N.R. Microbial Primer: An Introduction to Biofilms – What They Are, Why They Form and Their Impact on Built and Natural Environments. Microbiology 2023, 169, 001338. [Google Scholar] [CrossRef] [PubMed]
- Patel, R. Biofilms and Antimicrobial Resistance. Clin. Orthop. 2005, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Jakubovics, N.S.; Burgess, J.G. Extracellular DNA in Oral Microbial Biofilms. Microbes Infect. 2015, 17, 531–537. [Google Scholar] [CrossRef]
- Goodarzi, F.; Hallajzadeh, M.; Sholeh, M.; Talebi, M.; Mahabadi, V.P.; Amirmozafari, N. Biological Characteristics and Anti-Biofilm Activity of a Lytic Phage against Vancomycin-Resistant Enterococcus Faecium. Iran. J. Microbiol. 2021, 13, 691–702. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, Y.; Li, Y.; Lu, Y.; Xiong, K.; Zhong, Q.; Wang, J. Bacteriophage-Resistant Mutant of Enterococcus Faecalis Is Impaired in Biofilm Formation. Front. Microbiol. 2022, 13, 913023. [Google Scholar] [CrossRef] [PubMed]
- Winans, J.B.; Wucher, B.R.; Nadell, C.D. Multispecies Biofilm Architecture Determines Bacterial Exposure to Phages. PLoS Biol. 2022, 20, e3001913. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B.; Mlynek, K.D.; Hettiarachchi, H.; Alamneh, Y.A.; Biggemann, L.; Zurawski, D.V.; Black, C.C.; Bane, C.E.; Kim, R.K.; Granick, M.S. Extracellular Polymeric Substance (EPS)-Degrading Enzymes Reduce Staphylococcal Surface Attachment and Biocide Resistance on Pig Skin in Vivo. PloS One 2018, 13, e0205526. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Loessner, M.J. Bacteriophage Endolysins: Applications for Food Safety. Curr. Opin. Biotechnol. 2016, 37, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm Formation and Control Strategies of Foodborne Pathogens: Food Safety Perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm Formation in Food Industries: A Food Safety Concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
Figure 1.
Plaque morphology of phage vB_Efs8_KEN04.
Figure 2.
Phage stability test of Enterococcus phage vB_Efs8_KEN04. (A) Thermal stability test. (B) pH stability test.
Figure 2.
Phage stability test of Enterococcus phage vB_Efs8_KEN04. (A) Thermal stability test. (B) pH stability test.
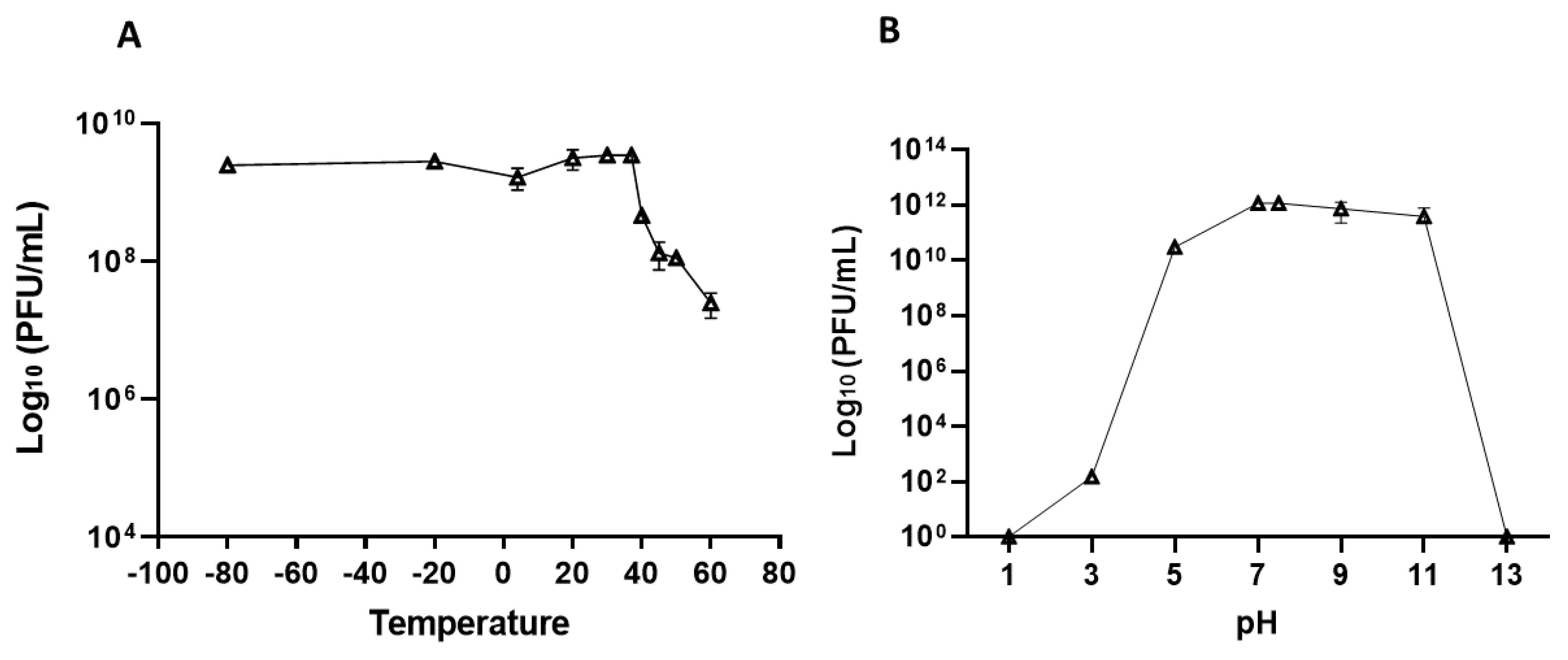
Figure 3.
Biofilm formation profile of Enterococcus faecalis isolates. The biofilm formation experiment was performed in triplicate, and the error bars represent the standard deviation. The horizontal grid line represents the OD threshold of 0.551 for biofilm-forming isolates.
Figure 3.
Biofilm formation profile of Enterococcus faecalis isolates. The biofilm formation experiment was performed in triplicate, and the error bars represent the standard deviation. The horizontal grid line represents the OD threshold of 0.551 for biofilm-forming isolates.
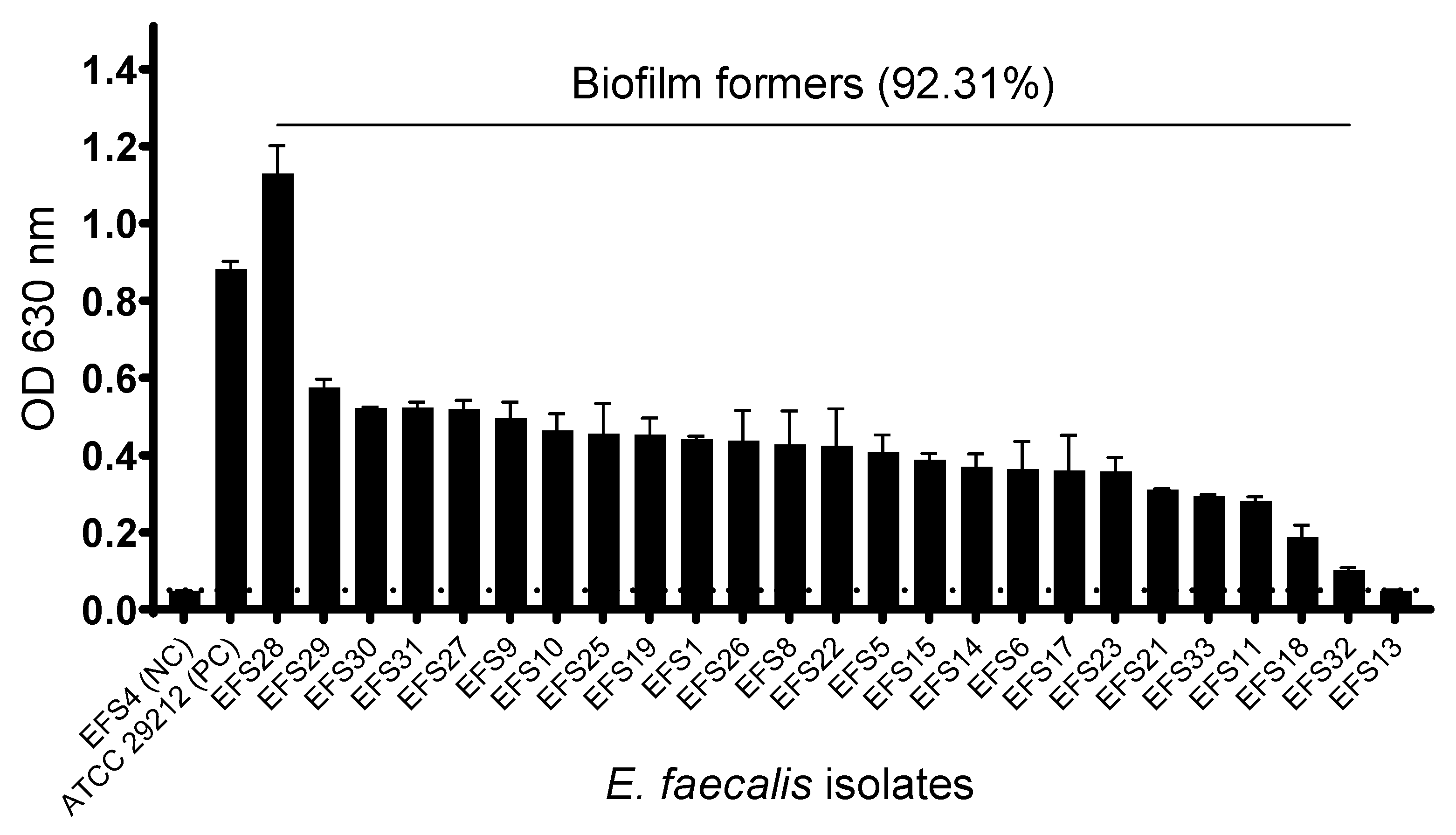
Figure 4.
(A) Inhibition of biofilm by Enterococcus phage vB_Efs8_KEN04; (B) Biofilm disruption by phage vB_Efs8_KEN04. Biofilm inhibition and disruption experiments were performed in triplicate, and the error bars represent the standard deviations. Significance level: *, p<0.05 significant; **, p<0.01 Very significant; ***, p<0.001 highly significant; ns, non-statistically significant.
Figure 4.
(A) Inhibition of biofilm by Enterococcus phage vB_Efs8_KEN04; (B) Biofilm disruption by phage vB_Efs8_KEN04. Biofilm inhibition and disruption experiments were performed in triplicate, and the error bars represent the standard deviations. Significance level: *, p<0.05 significant; **, p<0.01 Very significant; ***, p<0.001 highly significant; ns, non-statistically significant.
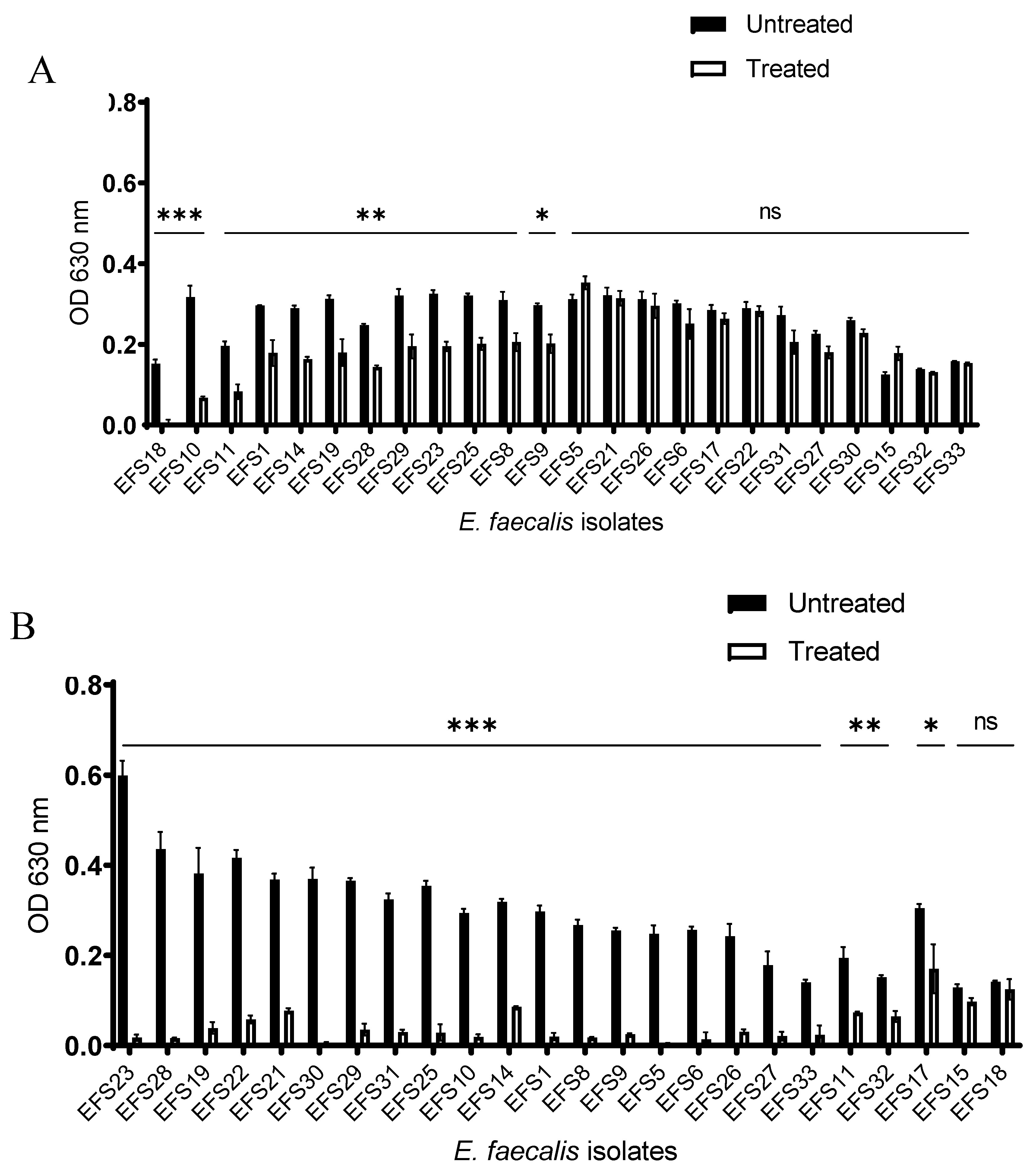
Figure 5.
Circular genome map of Enterococcus phage vB_Efs8_KEN04 constructed using CGView.
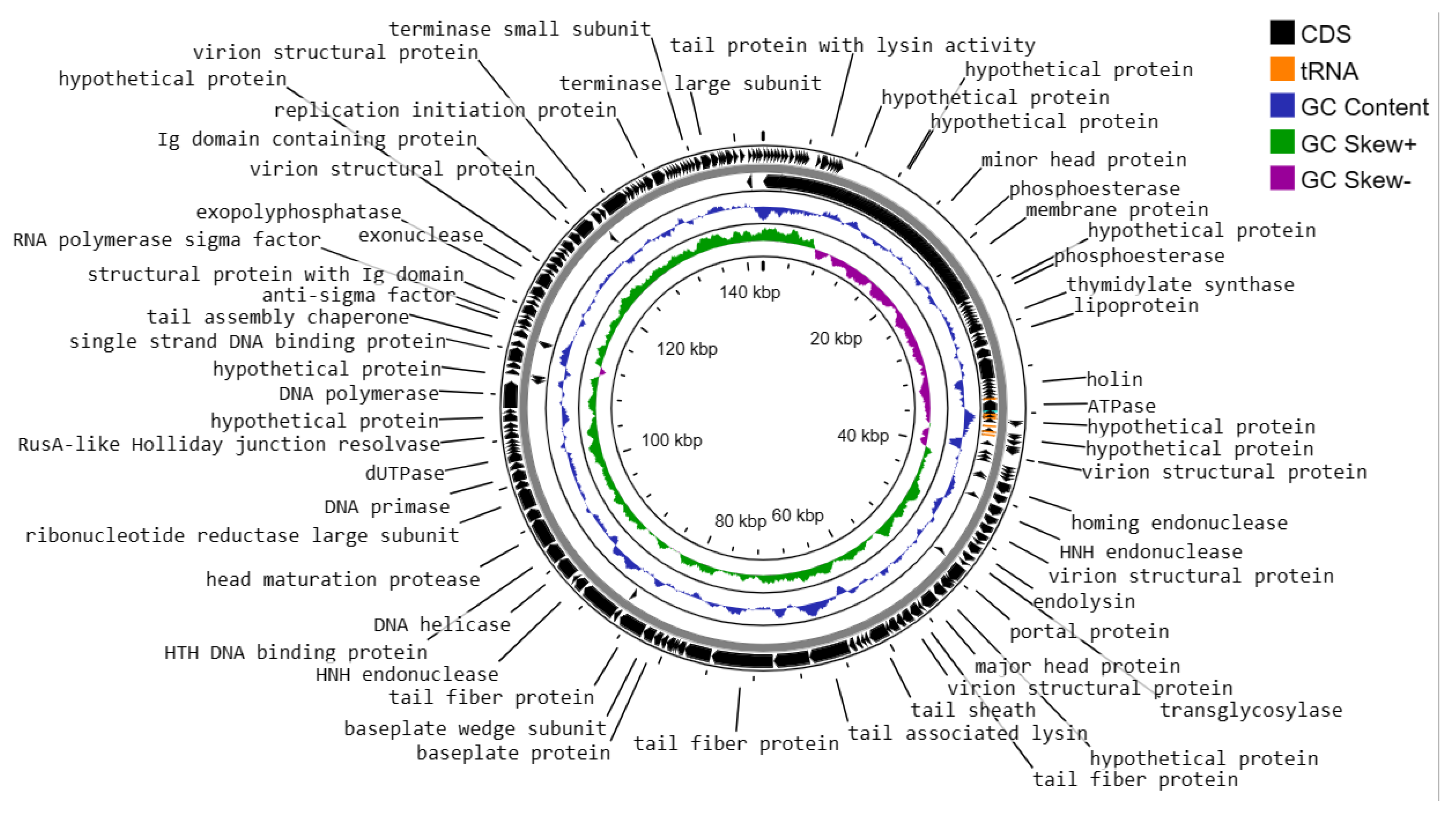
Figure 6.
(A) Phylogenetic analysis of Enterococcus phage vB_Efs8_KEN04 and other related Enterococcus bacteriophages based on the similarity of whole genome sequences. The phylogenetic tree was generated using the online VICTOR platform with the formula d0. (B) Heatmap of the average nucleotide identity values between phage vB_Efs8_KEN04 and the top 20 most similar Enterococcus bacteriophages.
Figure 6.
(A) Phylogenetic analysis of Enterococcus phage vB_Efs8_KEN04 and other related Enterococcus bacteriophages based on the similarity of whole genome sequences. The phylogenetic tree was generated using the online VICTOR platform with the formula d0. (B) Heatmap of the average nucleotide identity values between phage vB_Efs8_KEN04 and the top 20 most similar Enterococcus bacteriophages.
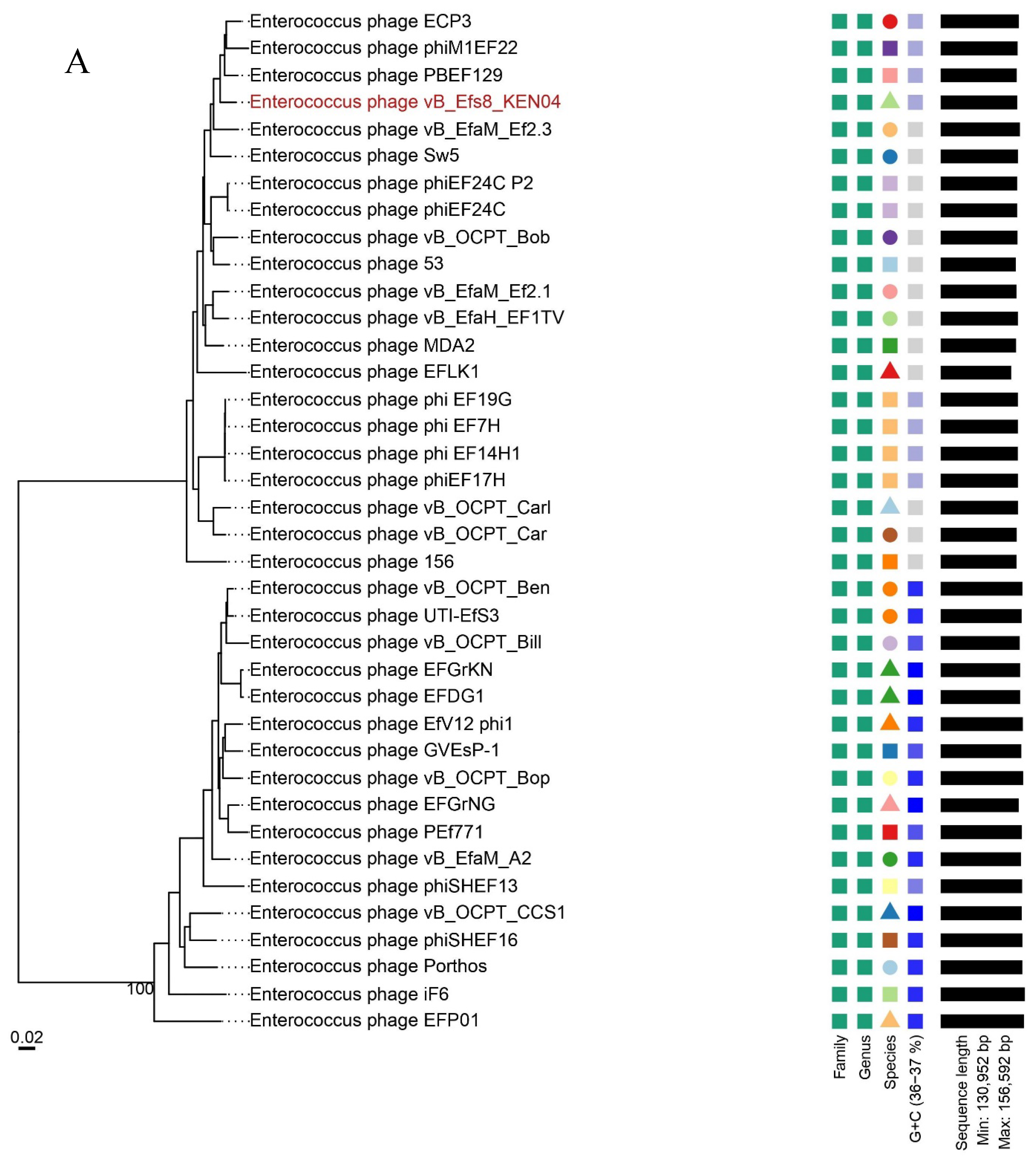
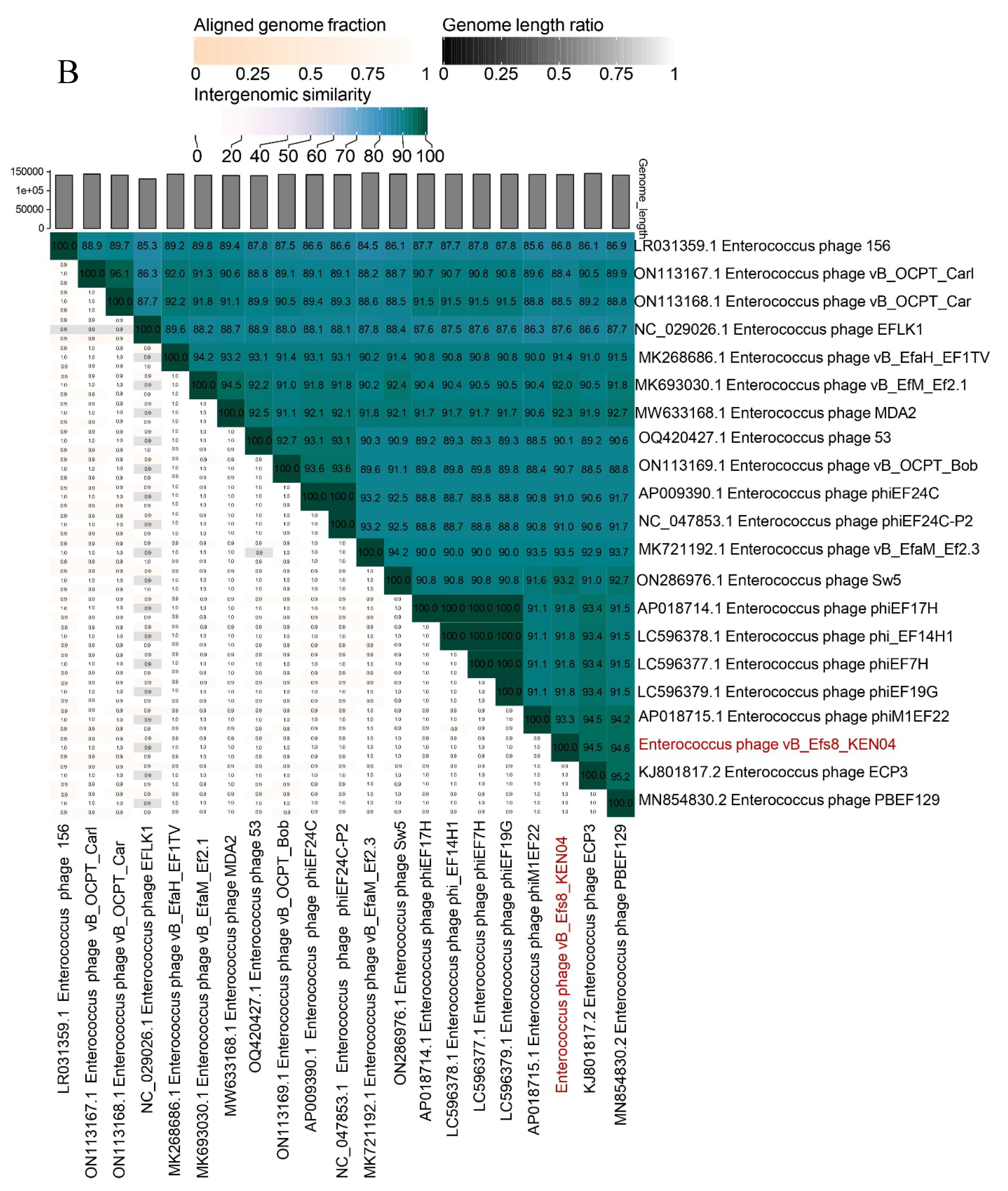

Table 1.
Host range analysis of Enterococcus phage vB_Efs8_KEN04.
| No. | Bacterial isolates | Sequence Types (ST) | Origin | Spot assay | Efficiency of Plating (EOP) |
|---|---|---|---|---|---|
| 1 | E. faecalis EFS8 * | 1904 | Urinary tract infection | ++ | 1 |
| 2 | E. faecalis EFS1 | 6 | Skin and soft tissue infection | ++ | 0.15 |
| 3 | E. faecalis EFS4 | 947 | Skin and soft tissue infection | + | <0.001 |
| 4 | E. faecalis EFS5 | 6 | Skin and soft tissue infection | ++ | 0.076 |
| 5 | E. faecalis EFS6 | 6 | Skin and soft tissue infection | ++ | 0.05 |
| 6 | E. faecalis EFS9 | 6 | Urinary tract infection | ++ | 0.01 |
| 7 | E. faecalis EFS10 | 6 | Urinary tract infection | ++ | 1.1 |
| 8 | E. faecalis EFS11 | 368 | Urinary tract infection | ++ | 0.0004 |
| 9 | E. faecalis EFS13 | 59 | Skin and soft tissue infection | ++ | 0.5 |
| 10 | E. faecalis EFS14 | 6 | Skin and soft tissue infection | ++ | 1.7 |
| 11 | E. faecalis EFS15 | Urinary tract infection | + | <0.001 | |
| 12 | E. faecalis EFS17 | 6 | Skin and soft tissue infection | + | <0.001 |
| 13 | E. faecalis EFS18 | 368 | Urinary tract infection | ++ | 0.0011 |
| 14 | E. faecalis EFS19 | Urinary tract infection | ++ | 0.12 | |
| 15 | E. faecalis EFS21 | 44 | Skin and soft tissue infection | + | <0.001 |
| 16 | E. faecalis EFS22 | Skin and soft tissue infection | + | <0.001 | |
| 17 | E. faecalis EFS23 | 6 | Urinary tract infection | ++ | 3 |
| 18 | E. faecalis EFS25 | 6 | Surgical site infection | ++ | 1.5 |
| 19 | E. faecalis EFS26 | 6 | Skin and soft tissue infection | ++ | 1.2 |
| 20 | E. faecalis EFS27 | 1903 | Urinary tract infection | ++ | 0.14 |
| 21 | E. faecalis EFS28 | 28 | Skin and soft tissue infection | ++ | 0.6 |
| 22 | E. faecalis EFS29 | 6 | Blood infection | ++ | 0.8 |
| 23 | E. faecalis EFS30 | 28 | Skin and soft tissue infection | ++ | 0.8 |
| 24 | E. faecalis EFS31 | 6 | Urinary tract infection | ++ | 1.2 |
| 25 | E. faecalis EFS32 | 1903 | Urinary tract infection | ++ | 0.9 |
| 26 | E. faecalis EFS33 | 1903 | Skin and soft tissue infection | ++ | 6 |
| 27 | E. faecium EFM5 | 80 | Urinary tract infection | + | <0.001 |
| 28 | E. faecium EFM1 | Skin and soft tissue infection | - | N/A | |
| 29 | E. faecium EFM2 | 80 | Skin and soft tissue infection | - | N/A |
| 30 | E. faecium EFM3 | Skin and soft tissue infection | - | N/A | |
| 31 | E. faecium EFM4 | Skin and soft tissue infection | - | N/A | |
| 32 | E. faecium EFM6 | 612 | Skin and soft tissue infection | - | N/A |
| 33 | E. faecium EFM7 | Skin and soft tissue infection | - | N/A | |
| 34 | E. faecium EFM8 | 80 | Urinary tract infection | - | N/A |
| 35 | E. faecium EFM9 | 80 | Skin and soft tissue infection | - | N/A |
| 36 | E. faecium EFM10 | Urinary tract infection | - | N/A | |
| 37 | E. faecium EFM11 | 761 | Surgical site infection | - | N/A |
EFS, Enterococcus faecalis; EFM, Enterococcus faecium; EOP, Efficiency of plating. The EOP was determined by dividing the mean Plaque Forming Units (PFU) of the target bacteria by the mean PFU of the host bacteria (EFS8). *Host bacteria; ++, very clear plaques; +, turbid plaques; −, no plaques; N/A, not applicable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



